
Abstract
Glial cells (astrocytes, oligodendrocytes and microglia) are critical for the central nervous system (CNS) in both physiological and pathological conditions. With this in mind, several studies have indicated that glial cells play key roles in the development and progression of CNS diseases. In this sense, gliotoxicity can be referred as the cellular, molecular, and neurochemical changes that can mediate toxic effects or ultimately lead to impairment of the ability of glial cells to protect neurons and/or other glial cells. On the other hand, glioprotection is associated with specific responses of glial cells, by which they can protect themselves as well as neurons, resulting in an overall improvement of the CNS functioning. In addition, gliotoxic events, including metabolic stresses, inflammation, excitotoxicity, and oxidative stress, as well as their related mechanisms, are strongly associated with the pathogenesis of neurological, psychiatric and infectious diseases. However, glioprotective molecules can prevent or improve these glial dysfunctions, representing glial cells-targeting therapies. Therefore, this review will provide a brief summary of types and functions of glial cells and point out cellular and molecular mechanisms associated with gliotoxicity and glioprotection, potential glioprotective molecules and their mechanisms, as well as gliotherapy. In summary, we expect to address the relevance of gliotoxicity and glioprotection in the CNS homeostasis and diseases.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Rudolf Virchow first described neuroglia as a connective tissue surrounding neurons in the nineteenth century [1]. Afterwards, different types of cells that constitute the neuroglia, also known as glial cells, were identified and referred as macroglia, composed by astrocytes and oligodendrocytes, and microglia. Each of these cells display several versatile functions necessary to maintain and support neuronal networks [2].
Glial cells are critical for the central nervous system (CNS) homeostasis, in both physiological and pathological conditions. In fact, it has been increasingly reported that these cells can be involved in the development and progression of neurological diseases [3]. Since each type of glial cell is able to directly affect the functionality of the others, dysfunctions in any of them can generate self-amplifying detrimental processes and synergistically impair the neuron-glia communication and/or neuronal function [4, 5]. In this sense, glial cells may represent important cellular therapeutic targets for CNS disorders. With this regard, cellular, molecular, and neurochemical changes in these cells, which enable them to mediate toxic effects or ultimately lead to an impairment of their ability to protect neurons and/or other glial cells, can be referred as gliotoxicity. On the other hand, glioprotection is associated with specific responses of glial cells, both in physiological and pathological conditions, by which they can protect themselves as well as neuronal cells, resulting in an overall improvement of the CNS functioning [6].
Considering that our research group has studied the role of glial cells on toxic and protective outcomes, this review will provide a brief summary of types and functions of glial cells and thus address molecular and cellular mechanisms associated with gliotoxicity and glioprotection, potential glioprotective molecules, and perspectives on gliotherapies (e.g., therapies for CNS pathologies focused on glial cells).
Types and Functions of Glial Cells
Astrocytes
Astrocytes are the most abundant glial cells and are recognized by a variety of homeostatic functions in the CNS. They have a refined cytoarchitecture with numerous stellate processes that allows their interaction with blood vessels, neurons, and other cell populations [7]. These morphological features of astrocytes make them important elements in the connection between peripheral and central systems, besides being associated with several of their functional roles, such as formation and maintenance of the blood–brain barrier (BBB); supply of oxygen and nutrients to the brain; regulation of synaptic transmission; and plasticity at the tripartite synapse [8,9,10]. The close contacts of astrocytes with synapses and blood vessels ensure their metabolic support to neurons through the coupling between synaptic activity and glucose utilization (neurometabolic coupling) [11].
Astrocytes participate in ionic homeostasis and metabolism of neurotransmitters, particularly of the glutamate [12]. They also regulate the diffusion and response to circulating factors, such as peripheral hormones, metabolites, and inflammatory mediators [13,14,15,16], in addition to participating in the defense against oxidative stress and in the detoxification of harmful molecules [17]. Moreover, astrocytes act as important secretory cells, releasing a wide range of signaling molecules (trophic and growth factors, gliotransmitters, peptides, and proteins) that will impact the functions of the surrounding glial, neuronal, and endothelial cells [18]. Inflammatory and immune responses are also important functional properties of astrocytes [19]. Moreover, astrocytes can respond to abnormal events in the CNS through a morphological, physiological, metabolic, biochemical, and transcriptional remodeling, which can impair their homeostatic functions and has been collectively called as reactive astrogliosis [20].
Oligodendrocytes
The major function of oligodendrocytes is synthesis of myelin sheath, a lipid-enriched specialized and compacted structure, which is extended in spirals around the axons of many neurons [21]. In an adult CNS, oligodendrocyte progenitor cells (OPCs) persist and can continuously proliferate and differentiate into mature and myelinating oligodendrocytes [22]. Both differentiation and myelination processes require a series of sequential steps and are orchestrated by several molecular and cellular signals, including those from neurons and other glial cells [23]. The rate of myelinating oligodendrocyte generation decreases throughout life, as the loss of myelin may result in cognitive disabilities and altered sensory and motor functions [21].
Recently, oligodendrocytes have been reported to provide metabolic support by transferring energy metabolites (particularly lactate and pyruvate) to neurons through monocarboxylate transporters (MCT) and cytoplasmic channels [24]. In addition, oligodendrocytes have been shown to perform immune functions by expressing both immune-related receptors and immunomodulatory molecules, which probably display pleiotropic roles in oligodendrocytes and other glial cells [25].
Microglia
Microglia are the resident immune cells of the CNS, distributed over the entire parenchyma and playing important roles to maintain brain homeostasis [26]. In response to brain damage or infections, they are usually the first cells to be activated to perform several well-established functions, among these, pathogen recognition, inflammatory responses, and phagocytosis [27, 28].
Under physiological conditions, microglia possess a specialized morphology with a small soma containing elongated, branched, and highly dynamic processes, which allows scanning the surrounding area and surveillance of the CNS [29, 30]. Upon CNS disorders, microglia quickly responds and can assume different activation patterns, which are usually associated with morphological changes. Microglial polarization may result in the neurotoxic M1-type or neuroprotective M2-type, based on the expression and release of cytokines, chemokines, and/or trophic factors [31, 32]. However, accumulating evidence has suggested that activation of microglia is heterogeneous and involves different but functionally overlapping phenotypes [33].
Besides their classical immune functions, microglia are implicated in several homeostatic processes, such as the release of trophic factors; promotion of neuronal survival, as well as the generation and maintenance of other neural cells; generation, maturation, regulation, and plasticity of synapses; synaptic pruning that redefines synapses and circuits; clearance of cells and debris; regulation of myelination; and memory formation and learning [26, 32].
Neurochemical Changes and Molecular Mechanisms Associated with Gliotoxicity
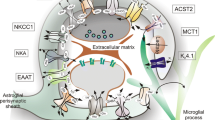
Considering the versatile and dynamic roles played by the different types of glial cells described in the previous section, it is not surprising that dysfunctions of these cells may be related to several pathological conditions. Interestingly, brain diseases may share many neurochemical, cellular, and molecular mechanisms related to gliotoxicity (Fig. 1), which is addressed in this section.

Gliotoxicity and glioprotection-associated mechanisms. Gliotoxicity may be linked to several detrimental processes, including metabolic and oxidative stresses, inflammation, and excitotoxicity. On the other hand, antioxidant defenses, metabolic and trophic support, anti-inflammatory response, and glutamate homeostasis are mechanisms associated with glioprotection. Changes in several signaling pathways in glial cells may result in both gliotoxic and glioprotective effects. The cells on the left represent reactive (dysfunctional) glial cells (astrocyte is represented in blue, microglia is represented in yellow, and oligodendrocyte is represented in purple); while the cells on the right represent functional glial cells (ramified astrocyte is represented in blue, ramified microglia is represented in yellow, and oligodendrocyte is represented in blue)
Metabolic Stresses
Glucose is the essential energy substrate for the CNS; therefore, pathological conditions associated with altered availability of glucose and/or oxygen (hypoxia/ischemia, hypoglycemia and hyperglycemia), as well as impairments in the metabolic machinery of cells, can largely impact glial functioning, inducing gliotoxicity. Hypoglycemia/glucose deprivation, hyperglycemia, and/or fluctuations in glucose concentration (hyperglycemia followed glucose deprivation) have been associated with several glial dysfunctions. In astroglial cells, metabolic stress alters glutamate metabolism, mitochondrial activity/redox balance, inflammatory response, release of trophic factors, and different signaling pathways [34,35,36]. Glucose-related metabolic stress also affects microglia and oligodendrocytes/OPCs, promoting microglial activation and contributing to inflammatory and oxidative injuries to neurons [37, 38]. Hypoglycemia inhibits oligodendrocyte development and differentiation, in addition to trigger apoptosis in OPCs [39], while glucose-oxygen deprivation causes intracellular lactate accumulation and acidosis [40]. However, hyperglycemia increased the expression of the pre-oligodendrocyte marker O4 without affecting the expression of NG2, a marker of OPC that is downregulated during the process of cell differentiation. Although hyperglycemia can improve the differentiation rate of OPCs, the mechanisms underlying this effect and its impacts are largely unknown [41].
Another important gliotoxicity condition associated with metabolic stress is caused by ammonia. Brain detoxification of ammonia occurs mainly via glutamine synthetase (GS) [42], a specific astrocytic enzyme, but hyperammonemia can exceed the metabolic capacity of cells. Ammonia-induced gliotoxicity is associated with cellular edema, energy depletion, oxidative stress, impairment in glutamate clearance and inflammatory response [43,44,45]. Moreover, ammonia can upregulate the senescence marker p21, thus potentially causing a glial-inflammaging process [14]. Although astrocytes are the primary targets of ammonia toxicity, microglia may also be affected. In a co-culture model of astrocyte and microglia, ammonia decreased cellular viability and promoted microglial activation, with an increase in the percentage of phagocytic type of microglia [46]. Additionally, ammonia induced oxidative stress and up-regulated the microglial activation marker ionized calcium-binding adaptor molecule-1 (Iba-1) in cultured microglia and in post mortem brain tissue from patients with hepatic encephalopathy, a neuropsychiatric syndrome associated with hyperammonemia [47]. Our group also have demonstrated that azide, an inhibitor of cytochrome c oxidase, induces gliotoxicity, promoting alterations in redox homeostasis, inflammatory response, and glutamate metabolism [6, 48].
Oxidative Stress, Inflammatory Response, and Excitotoxicity
Reactive oxygen/nitrogen species (ROS/RNS) can play neuromodulatory roles at physiological concentrations, including the regulation of neuronal polarity and neurite outgrowth, differentiation, cytoskeletal changes, synaptic plasticity, and activation of a wide range of signaling pathways and gene expression [49,50,51]. ROS provide a rapid and reversible mechanism for alter protein function by modulating the redox state of amino acid residues, particularly of cysteine, thus modifying the function of signaling proteins, ion channels, transporters, and transcription factors [52, 53]. On the contrary, excessive production of ROS/RNS by activated astrocytes and microglia can induce gliotoxicity, contributing to the pathomechanisms of neuropathological conditions [54, 55]. Oxidative/nitrosative stress in the brain can also be associated with unsaturated lipid enrichment, the presence of redox active transition metals, the neurotransmitter auto-oxidation and metabolism, the excessive glutamatergic signaling, as well as the increased expression and/or activity of NADPH oxidase and inducible nitric oxide synthase (iNOS) [56].
Although brain cells are equipped with enzymatic antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPx), and glutathione reductase (GR), the non-enzymatic antioxidant defense glutathione (GSH) plays a crucial role in maintaining brain redox homeostasis [54]. Furthermore, GSH depletion in glial cells is associated with oxidative stress, and the pathophysiology of brain disorders [57] and both this depletion and oxidative stress are closely related to inflammatory response in glial cells [58, 59]. Under pathological conditions, increased ROS production triggers inflammatory responses that, in turn, exacerbate the pro-oxidant status [53], thus establishing a self-amplifying cycle that contribute to gliotoxicity. This interplay between oxidative stress and inflammation is importantly mediated by the nuclear factor kappa B (NFκB) signaling [53].
Among glial cells, inflammatory and immune responses are primarily associated with microglia and astrocytes [60], although oligodendrocytes can be also involved to a lesser extent [25]. A wide range of extracellular stimuli can elicit inflammatory responses in glial cells, including pathogen-associated molecular pattern (PAMPs) and damage-associated molecular pattern (DAMPs), cytokines, and other molecular insults [19, 27]. PAMPs are small-molecule motifs present in pathogenic bacteria, protozoa, and viruses; DAMPs are molecular motifs associated with cellular injury and tissue damage (e.g., misfolded and aggregated proteins, miss-localized nucleic acids and other alarmins originated from damaged cells). Both molecular motif types can be recognized by specific pattern recognition receptors, in particular Toll-like receptors (TLRs), which are expressed by glial cells and can trigger innate immune responses [19, 26]. Mainly under the control of NFκB, the master regulator of inflammation, microglia, and astrocytes becomes an important source of several inflammatory mediators, including tumor necrosis factor alpha (TNF-α), interleukins (IL-1β, IL-6, IL-18), chemokines, and prostaglandins, in addition to nitric oxide (NO) and ROS [32, 61]. This plethora of inflammatory mediators will impact the surrounding environment, importantly participating in the inflammatory activation of the other glial cells and in the recruitment of peripheral immune cells [32, 62]. Excessive and chronic inflammatory responses lead to neuronal death and are involved in several CNS disorders [63].
Glutamate is the predominant excitatory neurotransmitter in the CNS and can be neurotoxic when inappropriately remaining at high levels in the synaptic cleft, a phenomenon referred as excitotoxicity [64]. Glial cells, particularly astrocytes, are responsible for the rapid removal of glutamate from synaptic cleft through excitatory amino acid transporters, EAAT1 (or glutamate-aspartate transporter, GLAST), and EAAT2 (or glutamate transporter 1, GLT-1) [65]. With this regard, downregulation and/or hypofunction of glutamate transporters may be associated with pathological conditions [64, 66], and their activity can be impaired as consequence of oxidative stress and/or inflammatory responses. It is well documented that glutamate transporters are highly susceptible to oxidation, which impairs their ability to take up extracellular glutamate [67]. Moreover, TNF-α has inhibitory effect on glutamate transport [68], probably associated with TNF-α-induced oxidative stress. Therefore, excitotoxicity, oxidative stress, and inflammation are processes closely related and represent important gliotoxicity mechanisms.
Gliotoxicity and CNS Diseases
Neurochemical changes and processes related to glial cells previously discussed represent, individually or collectively, crucial points in the pathogenesis of several brain diseases, including Alzheimer’s and Parkinson’s diseases, amyotrophic lateral sclerosis (ALS), multiple sclerosis, psychiatric disorders, stroke, diabetes mellitus, hepatic encephalopathy, the aging process, and infectious diseases.
Neuropsychiatric Disorders and Aging Process
Alzheimer’s disease, Parkinson’s disease, and ALS are neurodegenerative diseases that present the accumulation and aggregation of proteins, such as β-amyloid and tau, α-synuclein, and TAR DNA-binding protein 43 (TDP-43), respectively, as a common hallmark [69]. The pathophysiology of psychiatric disorders including major depressive disorder, bipolar disorder, and schizophrenia pointed toward monoamine disturbances and glutamatergic hypothesis [70]. However, the mechanisms of cognitive dysfunctions, neuronal degeneration, onset, and progression of these neuropsychiatric disorders have not yet been clearly elucidated. Interestingly, although they affect different CNS areas, producing different outcomes and symptoms, neuropsychiatric disorders share deleterious processes that may involve glial cells, including neuroinflammation, glutamate excitotoxicity, oxidative stress, and metabolic/trophic support [71,72,73,74]. Moreover, astrocyte reactivity can contribute to the pathomechanisms of the above-referred diseases. By definition, astrocyte reactivity involves morphological, molecular, and functional changes in response to pathological situations in surrounding tissue, such as CNS disease or injury, which may be reversible or chronic. When astrocytes undergo a reactive state, loss of homeostatic functions and gain of detrimental functions may occur, including some involved in glutamate and ionic homeostasis, glucose metabolism, production of inflammatory mediators and ROS/RNS, proliferation, BBB integrity, and Ca2+ signaling [20]. These astrocyte dysfunctions potentially lead to a transition from physiologic to pathologic roles that, without being the sole or primary initiators of pathology, may affect disease outcomes/progression [20]. With this regard, glial cells are the basis of many biomarkers of CNS diseases and, consequently, have emerged as important therapeutic targets for these pathological conditions. In addition, changes in several signaling pathways in glial cells corroborate their role in the pathomechanisms of neuropsychiatric disorders and neurodegenerative diseases [75,76,77].
Aging is a complex process characterized by an intrinsic physiological and functional decline of an organism. Although brain aging increases the risk of neurodegenerative diseases, it is not pathological and may be related to adaptive mechanisms of cell physiology over time [78]. Aged human brains display only mild and heterogeneous changes in astrocyte morphology or GFAP levels [79]. However, other cellular and molecular hallmarks of aging have been studied in glial cells. Aging has been associated with decreased glucose and glycogen metabolism, as well as with decreased ATP production, decreasing astrocytic fuel provision of neurons [80,81,82]. Glutamate transporter downregulation and/or hypofunction has been also observed with aging [81, 83], which impair glutamate uptake and favor excitotoxicity. In addition, aged astrocytes and microglia have been shown to accumulate ROS and produce increased amounts of pro-inflammatory mediators, which may be exacerbated in response to harmful stimuli [84, 85]. Considering both senescence and inflammation in particular, the phenomenon of inflammaging has emerged as a mechanism shared by age-related diseases [86], notably involving astrocytes and microglia.
Neurological Dysfunctions Associated with Infectious Diseases
Despite the protective barriers, such as BBB, CNS can be directly and/or indirectly affected by bacterial and viral infections. In the context of bacterial infections, lipopolysaccharide (LPS), a toxin present in the outer membrane of gram-negative bacteria, has been widely used as a model of peripheral and central inflammatory responses and their related cognitive decline [87]. A large body of evidence has demonstrated that LPS peripherally administrated is able to induce inflammatory responses within CNS [88, 89]. Due to the position of astrocytes, they can serve as a bridge between systemic inflammation and neuroinflammation [15]. Although LPS classically activates microglia and astrocytes, oligodendroglial cells can also respond to this bacterial inflammogen [90]. Of note, glial cells can be major responsible for LPS-induced neuronal damage [91, 92]. Therefore, LPS has exhaustively used as an in vivo and in vitro experimental model of brain diseases, such as Alzheimer’s disease and schizophrenia, among others.
With regard to viral infections, glial cells can be primary targets of neurotropic viruses, such as the human immunodeficiency virus type 1 (HIV-1) and zika virus (ZIKV). Microglia and astrocytes constitute CNS reservoirs of HIV-1 [93, 94], promoting neuroinflammation, which can explain neuronal damage and neurocognitive disorders in a number of patients, considering the relative incapacity of HIV-1 to directly infect neurons [95]. Due to the presence of AXL receptor, astrocytes and microglia are potentially the primary cells targeted by ZIKV in the CNS [96]. In both astrocytes and microglia, ZIKV elicited classical inflammatory responses [96], while, for astrocytes, it induced oxidative stress, mitochondrial failure, and DNA damage in astrocytes [97]. Our group have demonstrated that an acute hippocampus exposure to ZIKV is also able to induce neuroinflammation and oxidative stress, affecting neuron-glia communication [98].
COVID-19 has been also recently associated with neurological dysfunctions, yet it is unclear whether they are consequence of direct CNS infection by SARS-CoV-2 or whether they result from peripheral cytokine storm and metabolic dysfunctions, although investigators have found that neurons and astrocytes are susceptible to SARS-CoV-2 infection [99, 100]. Anosmia and ageusia are common neurologic symptoms in COVID-19 patients, which can be associated with dysfunction in the olfactory bulb [101]. In addition, this brain structure can mediate direct viral invasion [102]. Interestingly, olfactory impairment is a common and early (preclinical) sign of neurodegenerative diseases, including Alzheimer’s and Parkinson’s diseases, in addition to be associated with depression and other neuropsychiatric disorders [103, 104]. The precise mechanisms that connect these diseases with olfactory loss are also still unclear but potentially involve neuroinflammation [103, 104]. Therefore, infectious diseases mainly target glial cells and might generate long-term consequences including cognitive deficits, neurodegenerative diseases, psychiatric disorders, and others that are currently unknown.
Mechanisms Underlying Glioprotection
Glioprotection can be achieved by endogenous homeostatic and protective functions of glial cells, which in turn may be positively modulated by a wide range of exogenous molecules, named as glioprotective molecules. They can promote protection to the CNS by improving glial functions and avoiding gliotoxicity. This section will discuss the main points associated with glioprotection (Fig. 1).
Metabolic Support
Astrocytes are recognized as energy substrate suppliers, since they are responsible for glucose uptake and its distribution to other neural cells, besides being able to store it as glycogen [9]. Moreover, astrocytes largely metabolize glucose glycolytically to produce ATP, generating lactate, which can be later transferred to neurons to be fully oxidized under conditions of high energy demands or when glucose supply is low [9, 105]. More recently, it has been demonstrated a metabolic coupling between oligodendrocytes and neurons, in which lactate derived from the glucose metabolism of these glial cells can also be transferred to the axon, contributing to ATP synthesis in neurons [106]. Of note, besides its metabolic function, signaling roles of extracellular lactate have been also recently investigated particularly in neurons, associated with neuronal excitability, synaptic plasticity, memory consolidation, and expression of trophic factors. Such signaling effects can be mediated either by the G protein-coupled receptor GPR81, extracellular acidification, changes in redox state, or depolarization of target cells [107, 108]. In addition to lactate, astrocytes can also transfer healthy mitochondria to neurons, replacing damaged organelles of these cells and thus providing a protection against neuronal mitochondrial dysfunction [109].
Another important metabolic cooperation between astrocytes and neurons comprises the glutamate-glutamine cycle [11]. Once taken up by astrocytes, glutamate can be converted into glutamine by the enzyme GS, which participates both in glutamate metabolism and in ammonia detoxification [42]. Glutamine is then exported to neurons, allowing recycling of glutamate. Since glutamate is also the precursor of gamma aminobutyric acid (GABA), the glutamate-glutamine cycle is crucial for maintaining glutamate and GABA-based neurotransmission [110]. Moreover, de novo synthesis of glutamate in the brain occurs in astrocytes via the pyruvate carboxylase pathway and thus also depends on glucose [111].
Concerning the lipid metabolism, astrocytes are an important cholesterol source to mature neurons, since these glial cells express the enzymes for cholesterol synthesis and the apolipoproteins necessary to export it [112]. Moreover, although oligodendrocytes are able to synthesize cholesterol, a critical component of the myelin structure, they also depend on the supply from astrocytes [112]. In addition, there is a metabolic coupling between astrocytes and neurons regarding detoxification of neuronal-derived toxic fatty acids, which are transferred to astrocytes and metabolized via mitochondrial β-oxidation [113].
Trophic Support
Synthesis and release of a wide range of trophic factors by glial cells, especially by astrocytes, constitute another important mechanism of glioprotection. These trophic factors include brain-derived neurotrophic factor (BDNF), glial cell line-derived neurotrophic factor (GDNF), S100B, transforming growth factor-β (TGF-β), vascular endothelial growth factor (VEGF), insulin-like growth factor-1 (IGF-1), nerve growth factor (NGF), and neurotrophins 3 and 4. These multifunctional molecules can mostly act on nearby neurons, regulating neuronal survival, differentiation, function, plasticity and regeneration, as well as neurogenesis [32, 114, 115]. In addition, trophic factors can also target other glial and even endothelial cells, regulating several processes, such as oligodendrocyte differentiation, survival and remyelination; microglial activation; astrocyte proliferation, function and metabolism; angiogenesis; and BBB integrity [116,117,118].
Modulation of Triad Oxidative Stress, Excitotoxicity, and Inflammatory Response
Glial cells display efficient antioxidant defense mechanisms that allow their functions in the defense and repair of the brain. In particular, astrocytes are able to maintain their protective roles even after surviving intense oxidative stress, thus playing a crucial role for providing antioxidant support to neurons [119]. In light of this, neuron-astrocyte interactions mediate an essential mechanism for recycling of ascorbic acid, an important neuronal antioxidant defense [120]. In addition, GSH, a tripeptide consisting of glutamate, cysteine, and glycine that is synthesized by the enzymes γ-glutamyl cysteine ligase (GCL) and GSH synthase [121], is an important antioxidant molecule able to react with free radicals or participate in enzymatic reactions, such as those catalyzed by GPx and glutathione-S-transferase (GST). While microglia and oligodendrocytes synthetize GSH for their self-protection, astrocytes are also able to readily release it [121, 122]. This is particularly important because neurons are dependent on astrocytic GSH for providing extracellular cysteine for their synthesis of GSH, since they are less capable of importing cystine [122, 123]. Extracellular GSH, in turn, can be protected from oxidation by other “guardian” molecules, including the SOD, secreted by astrocytes [124].
GSH metabolism and glutamatergic neurotransmission/homeostasis are processes closely interconnected in several ways. Besides astrocytes, microglia and oligodendrocytes also express glutamate transporters and uptake glutamate, but they are probably associated with the GSH demands of these cells [125, 126]. Thus, glial glutamate transporters can provide intracellular glutamate for GSH synthesis, as well as for Cys-Glu exchanger (system xc−) operation. This transporter, present in glial cells, plays a crucial role for GSH synthesis, since it mediates the uptake of cystine, the bioavailable form of cysteine, in exchange for glutamate [127]. Maintenance of adequate GSH levels, therefore, is important to protect glutamate transporters from oxidation and avoid excitotoxicity. Moreover, and interestingly, it has been recently hypothesized that GSH is a relevant glutamate reservoir and could supply it for synaptic transmission when the glutamate-glutamine cycle is impaired [128].
Additionally, both microglia and astrocytes can be involved in suppression of inflammation and immune responses [129, 130]. Alternative activation patterns adopted by these glial cells are related to production and release of several anti-inflammatory molecules, such as IL-4, IL-10, IL-11, and IL-27, as well as TGF-β, that function mainly by suppressing the pro-inflammatory milieu [131, 132]. Thus, they establish a bidirectional crosstalk for a reciprocal anti-inflammatory modulation of microglia and astrocytes. To illustrate this relationship, activated M2-like microglia produce anti-inflammatory cytokine IL-10 that stimulate astrocytes to secrete TGF-β, which in turn reduces microglial pro-inflammatory activation, ultimately preserving neuronal and oligodendroglial functioning [132].
Signaling Pathways associated with Glioprotection
The nuclear factor erythroid-derived 2-like 2 (Nrf2) is a stress-responsive transcription factor that acts as a key regulator of redox, metabolic, and inflammatory homeostasis [133,134,135]. Upon activation, Nrf2 is translocated into the nucleus and controls the expression of genes that encode antioxidant enzymes, including SOD, GPx, and GST [133, 135]. It also stimulates the expression of proteins that contribute to GSH biosynthesis and homeostasis, such as system xc−, GCL, GSH synthase, and the NADPH-generating enzyme glucose-6-phosphate dehydrogenase [133]. Moreover, Nrf2 may directly or indirectly influence intermediary metabolism and mitochondrial function. It directly regulates the expression of important enzymatic steps of metabolic pathways related to synthesis of carbohydrates, nucleic acids, lipids, and amino acids. Indirectly, Nrf2 can affect its own expression [136] and the other transcription factors [e.g., peroxisome proliferator-activated receptor γ (PPARγ) and retinoid X receptor α (RXRα)] [137] that in turn regulate metabolic genes, in addition to influence the activity of metabolic enzymes that are susceptible to thiol modifications [e.g., pyruvate dehydrogenase kinase 2, pyruvate kinase, AMP-activated protein kinase (AMPK)], since Nrf2-mediated expression of antioxidant genes can prevent or reverse oxidation of cysteine residues [133].
Heme oxygenase 1 (HO-1) is one of the classical genes regulated by Nrf2, which is associated with responses against oxidative challenges. This enzyme catalyzes the degradation of heme into biliverdin, bilirubin, carbon monoxide, and free iron. Products of HO-1, in particular bilirubin and CO, mediate protective effects since they have antioxidant and anti-inflammatory properties [138]. Of note, they can inhibit iNOS activity and NFκB activation [135]; thus HO-1 is an important element in the connection between Nrf2 and NFκB signaling pathways. In fact, Nrf2 signaling negatively regulates NFκB-driven inflammatory and oxidative stress responses [135]. In the context of glioprotection, although microglia and oligodendrocytes exhibit functional Nrf2/HO-1 signaling, astrocytes may be the predominant neural cell type for activation of Nrf2 [139].
Other signaling pathways that act as key regulators of cell survival, responses to stressful conditions, and metabolic effectors can mediate glioprotective effects, including sirtuin 1 (SIRT1), AMPK, phosphoinositide3-kinase (PI3K)/Akt, and protein kinase C (PKC) [35, 140, 141].
Glioprotective Molecules
A wide range of molecules has been investigated as candidates to mediate protective effects on the CNS by targeting glial cells (Table 1). Resveratrol, a polyphenol stilbene found in grapes and wine, is one of these promising molecules. Several studies have shown that resveratrol regulates diverse astroglial functions, including antioxidant defenses, inflammatory response, trophic factor release, and glutamate homeostasis, both at basal conditions and against harmful stimuli [43, 48, 140, 142,143,144,145,146,147]. Additionally, resveratrol is able to prevent age-related functional alterations of astrocytes [148]. These effects are associated with different signaling pathways, including Nrf2/HO-1, SIRT1, PI3K/Akt, AMPK, adenosine receptors, and NFκB [48, 140, 144, 149]. Moreover, resveratrol also exhibits glioprotective effects on microglial and oligodendroglial cells [90, 145, 150] and in different in vivo experimental models [151,152,153].
Besides resveratrol, other naturally occurring molecules of plant origin can promote glioprotection, such as curcumin (polyphenolic compound found in the rhizome of Curcuma longa Linn) [154,155,156,157,158,159,160,161,162,163], isoflavones (flavonoid polyphenols present in leguminous plants) [164,165,166,167,168,169,170,171,172,173,174], and sulforaphane (isothiocyanate found in cruciferous vegetables) [175,176,177,178,179]. Endogenous mammalian compounds including lipoic acid (an essential cofactor for different mitochondrial enzymes) [141, 180,181,182,183,184,185] and guanosine (a guanine-based purine) have been also investigated as potential glioprotective agents [6, 35, 83, 186,187,188,189,190]. The mechanisms underlying the protective effects of these molecules in glial cells involve antioxidant and anti-inflammatory activities, improvement of mitochondrial function, Nrf2/HO-1 activation and NFκB inhibition, glutamate clearance and metabolism, regulation of microglial activation, survival of oligodendrocytes, and delay of demyelination (Table 1).
Perspectives on Gliotherapies
A wide variety of medications currently used to treat psychiatric disorders and neurodegenerative diseases are shown to have beneficial effects on glial cells, which may participate in their therapeutic effects. Antipsychotics, such as risperidone, haloperidol, clozapine and quetiapine are able to regulate inflammatory responses in astrocytes and/or microglia [191,192,193]. Risperidone, in particular, modulate glutamate uptake, GS activity, GSH content, and S100B release in astroglial cells [191, 194, 195]. Major antidepressants (serotonin-specific reuptake inhibitors, tricyclic antidepressants) also demonstrate anti-inflammatory properties [196], as well as improving the release of trophic factors by glial cells [196, 197]. In addition, riluzole, the only drug approved for ALS, mainly target glutamate excitotoxicity, at least in part, by improving astroglial glutamate uptake [198], and it may also increase synthesis of trophic factors and induce Nrf2/HO-1 signaling [199].
In line with this, many studies strive to develop specific gliotherapies for treatment of neurological diseases [200, 201], demonstrating that glial cells can represent a novel basis for understanding, preventing, and treating these conditions, such as Alzheimer’s disease and schizophrenia. Moreover, characterizing the role of glial cells in the pathophysiology of CNS diseases as well as identifying gliotherapeutic targets can improve future gliotherapies [202].
Concluding Remarks
The last 25 years have brought significant progress in the understanding of glial functionality, since these cells play a critical role in CNS homeostasis, as well as in pathogenesis and progression of CNS diseases. With these concept changes, it is believed that we will be able to make rapid progress in the findings, as well as in a broader and more efficient way to demonstrate that glial cells can be targets to drug development. Currently, it is well established that under oxidative and inflammatory challenges, glial cells can switch from having a protective role to a harmful phenotype. In addition, triad oxidative stress, neuroinflammation, and excitotoxicity are strongly associated with several neurological and psychiatric disorders. Considering the relevance of glial cells for physio/pathological processes, our Lab has studied these cells in different models of gliotoxicity to propose glioprotective strategies in the future, as well as to characterize the mechanisms of glioprotection. By understanding gliotoxicity, glial-based preventive/therapeutic strategies might emerge to delay and to prevent the development of CNS diseases and their consequences.
Finally, this review represents an overview of gliotoxicity and glioprotection and was wrote by researchers from the Neurotoxicity and Glioprotection Lab of Federal University of Rio Grande do Sul, as a remote activity during COVID-19 pandemic.
Data availability
Not applicable.
Code availability
Not applicable.
References
Virchow R (1858) Die Cellularpathologie in ihrer Begründung auf physiologische und pathologische Gewebelehre. Berlin, Germany(20): August Hirschwald; 1858
Verkhratsky A, Ho MS, Zorec R, Parpura V (2019) The Concept of Neuroglia. In: Verkhratsky A, Ho MS, Zorec R, Parpura V (eds) Neuroglia in Neurodegenerative Diseases. Springer Singapore, Singapore, pp 1–13
Parpura V, Heneka MT, Montana V et al (2012) Glial cells in (patho)physiology: Glial cells in (patho)physiology. J Neurochem 121:4–27. https://doi.org/10.1111/j.1471-4159.2012.07664.x
Patel DC, Tewari BP, Chaunsali L, Sontheimer H (2019) Neuron–glia interactions in the pathophysiology of epilepsy. Nat Rev Neurosci 20:282–297. https://doi.org/10.1038/s41583-019-0126-4
Bernaus A, Blanco S, Sevilla A (2020) Glia Crosstalk in Neuroinflammatory Diseases. Front Cell Neurosci 14:209. https://doi.org/10.3389/fncel.2020.00209
Quincozes-Santos A, Bobermin LD, Souza DG et al (2014) Guanosine protects C6 astroglial cells against azide-induced oxidative damage: a putative role of heme oxygenase 1. J Neurochem 130:61–74. https://doi.org/10.1111/jnc.12694
Matyash V, Kettenmann H (2010) Heterogeneity in astrocyte morphology and physiology. Brain Res Rev 63:2–10. https://doi.org/10.1016/j.brainresrev.2009.12.001
Perea G, Navarrete M, Araque A (2009) Tripartite synapses: astrocytes process and control synaptic information. Trends Neurosci 32:421–431. https://doi.org/10.1016/j.tins.2009.05.001
Barros LF, Brown A, Swanson RA (2018) Glia in brain energy metabolism: A perspective. Glia 66:1134–1137. https://doi.org/10.1002/glia.23316
Gonçalves C-A, Rodrigues L, Bobermin LD et al (2018) Glycolysis-Derived Compounds From Astrocytes That Modulate Synaptic Communication. Front Neurosci 12:1035. https://doi.org/10.3389/fnins.2018.01035
Bélanger M, Allaman I, Magistretti PJ (2011) Brain energy metabolism: focus on astrocyte-neuron metabolic cooperation. Cell Metab 14:724–738. https://doi.org/10.1016/j.cmet.2011.08.016
Benarroch EE (2016) Astrocyte signaling and synaptic homeostasis: I: Membrane channels, transporters, and receptors in astrocytes. Neurology 87:324–330. https://doi.org/10.1212/WNL.0000000000002875
Santos CL, Bobermin LD, Souza DO, Quincozes-Santos A (2018) Leptin stimulates the release of pro-inflammatory cytokines in hypothalamic astrocyte cultures from adult and aged rats. Metab Brain Dis 33:2059–2063. https://doi.org/10.1007/s11011-018-0311-6
Bobermin LD, Roppa RHA, Gonçalves C-A, Quincozes-Santos A (2020) Ammonia-Induced Glial-Inflammaging. Mol Neurobiol 57:3552–3567. https://doi.org/10.1007/s12035-020-01985-4
Bellaver B, Dos Santos JP, Leffa DT et al (2018) Systemic Inflammation as a Driver of Brain Injury: the Astrocyte as an Emerging Player. Mol Neurobiol 55:2685–2695. https://doi.org/10.1007/s12035-017-0526-2
Marina N, Turovsky E, Christie IN et al (2018) Brain metabolic sensing and metabolic signaling at the level of an astrocyte. Glia 66:1185–1199. https://doi.org/10.1002/glia.23283
Dringen R, Brandmann M, Hohnholt MC, Blumrich E-M (2015) Glutathione-Dependent Detoxification Processes in Astrocytes. Neurochem Res 40:2570–2582. https://doi.org/10.1007/s11064-014-1481-1
Vardjan N, Parpura V, Verkhratsky A, Zorec R (2019) Gliocrine System: Astroglia as Secretory Cells of the CNS. In: Verkhratsky A, Ho MS, Zorec R, Parpura V (eds) Neuroglia in Neurodegenerative Diseases. Springer Singapore, Singapore, pp 93–115
Sofroniew MV (2020) Astrocyte Reactivity: Subtypes, States, and Functions in CNS Innate Immunity. Trends Immunol 41:758–770. https://doi.org/10.1016/j.it.2020.07.004
Escartin C, Galea E, Lakatos A et al (2021) Reactive astrocyte nomenclature, definitions, and future directions. Nat Neurosci 24:312–325. https://doi.org/10.1038/s41593-020-00783-4
Stadelmann C, Timmler S, Barrantes-Freer A, Simons M (2019) Myelin in the Central Nervous System: Structure, Function, and Pathology. Physiol Rev 99:1381–1431. https://doi.org/10.1152/physrev.00031.2018
Kuhn S, Gritti L, Crooks D, Dombrowski Y (2019) Oligodendrocytes in Development. Myelin Generation and Beyond Cells 8:1424. https://doi.org/10.3390/cells8111424
Baydyuk M, Morrison VE, Gross PS, Huang JK (2020) Extrinsic Factors Driving Oligodendrocyte Lineage Cell Progression in CNS Development and Injury. Neurochem Res 45:630–642. https://doi.org/10.1007/s11064-020-02967-7
Philips T, Rothstein JD (2017) Oligodendroglia: metabolic supporters of neurons. J Clin Invest 127:3271–3280. https://doi.org/10.1172/JCI90610
Zeis T, Enz L, Schaeren-Wiemers N (2016) The immunomodulatory oligodendrocyte. Brain Res 1641:139–148. https://doi.org/10.1016/j.brainres.2015.09.021
Prinz M, Jung S, Priller J (2019) Microglia Biology: One Century of Evolving Concepts. Cell 179:292–311. https://doi.org/10.1016/j.cell.2019.08.053
Kettenmann H, Hanisch U-K, Noda M, Verkhratsky A (2011) Physiology of Microglia. Physiol Rev 91:461–553. https://doi.org/10.1152/physrev.00011.2010
Haley MJ, Brough D, Quintin J, Allan SM (2019) Microglial Priming as Trained Immunity in the Brain. Neuroscience 405:47–54. https://doi.org/10.1016/j.neuroscience.2017.12.039
Kierdorf K, Prinz M (2017) Microglia in steady state. J Clin Invest 127:3201–3209. https://doi.org/10.1172/JCI90602
Rio-Hortega P (1939) THE MICROGLIA The Lancet 233:1023–1026. https://doi.org/10.1016/S0140-6736(00)60571-8
Hu X, Leak RK, Shi Y et al (2015) Microglial and macrophage polarization—new prospects for brain repair. Nat Rev Neurol 11:56–64. https://doi.org/10.1038/nrneurol.2014.207
Colonna M, Butovsky O (2017) Microglia Function in the Central Nervous System During Health and Neurodegeneration. Annu Rev Immunol 35:441–468. https://doi.org/10.1146/annurev-immunol-051116-052358
Ransohoff RM (2016) A polarizing question: do M1 and M2 microglia exist? Nat Neurosci 19:987–991. https://doi.org/10.1038/nn.4338
Quincozes-Santos A, Bobermin LD, de Assis AM et al (2017) Fluctuations in glucose levels induce glial toxicity with glutamatergic, oxidative and inflammatory implications. Biochim Biophys Acta BBA - Mol Basis Dis 1863:1–14. https://doi.org/10.1016/j.bbadis.2016.09.013
Quincozes-Santos A, Bobermin LD, de Souza DG et al (2013) Gliopreventive effects of guanosine against glucose deprivation in vitro. Purinergic Signal 9:643–654. https://doi.org/10.1007/s11302-013-9377-0
Tramontina AC, Nardin P, Quincozes-Santos A et al (2012) High-Glucose and S100B Stimulate Glutamate Uptake in C6 Glioma Cells. Neurochem Res 37:1399–1408. https://doi.org/10.1007/s11064-012-0722-4
Rabenstein M, Vay SU, Blaschke S et al (2020) Crosstalk between stressed brain cells: direct and indirect effects of ischemia and aglycemia on microglia. J Neuroinflammation 17:33. https://doi.org/10.1186/s12974-020-1697-8
Zeng X, Ren H, Zhu Y et al (2018) Gp91phox (NOX2) in Activated Microglia Exacerbates Neuronal Damage Induced by Oxygen Glucose Deprivation and Hyperglycemia in an in Vitro Model. Cell Physiol Biochem 50:783–797. https://doi.org/10.1159/000494243
Yan H, Rivkees SA (2006) Hypoglycemia influences oligodendrocyte development and myelin formation. NeuroReport 17:55–59. https://doi.org/10.1097/01.wnr.0000192733.00535.b6
Zhang N, Guan T, Shafiq K et al (2020) Compromised Lactate Efflux Renders Vulnerability of Oligodendrocyte Precursor Cells to Metabolic Stresses. ACS Chem Neurosci 11:2717–2727. https://doi.org/10.1021/acschemneuro.0c00353
da Rosa PM, Meira LAM, Souza DO et al (2019) High-glucose medium induces cellular differentiation and changes in metabolic functionality of oligodendroglia. Mol Biol Rep 46:4817–4826. https://doi.org/10.1007/s11033-019-04930-4
Brusilow SW, Koehler RC, Traystman RJ, Cooper AJL (2010) Astrocyte glutamine synthetase: Importance in hyperammonemic syndromes and potential target for therapy. Neurotherapeutics 7:452–470. https://doi.org/10.1016/j.nurt.2010.05.015
Bobermin LD, Quincozes-Santos A, Guerra MC et al (2012) Resveratrol prevents ammonia toxicity in astroglial cells. PLoS ONE 7:e52164. https://doi.org/10.1371/journal.pone.0052164
Bobermin LD, Hansel G, Scherer EBS et al (2015) Ammonia impairs glutamatergic communication in astroglial cells: protective role of resveratrol. Toxicol Vitro Int J Publ Assoc BIBRA 29:2022–2029. https://doi.org/10.1016/j.tiv.2015.08.008
Bobermin LD, Souza DO, Gonçalves C-A, Quincozes-Santos A (2018) Resveratrol prevents ammonia-induced mitochondrial dysfunction and cellular redox imbalance in C6 astroglial cells. Nutr Neurosci 21:276–285. https://doi.org/10.1080/1028415X.2017.1284375
Ismail FS, Faustmann TJ, Corvace F et al (2021) Ammonia induced microglia activation was associated with limited effects on connexin 43 and aquaporin 4 expression in an astrocyte-microglia co-culture model. BMC Neurosci 22:21. https://doi.org/10.1186/s12868-021-00628-1
Zemtsova I, Görg B, Keitel V et al (2011) Microglia activation in hepatic encephalopathy in rats and humans. Hepatology 54:204–215. https://doi.org/10.1002/hep.24326
Bellaver B, Bobermin LD, Souza DG et al (2016) Signaling mechanisms underlying the glioprotective effects of resveratrol against mitochondrial dysfunction. Biochim Biophys Acta BBA - Mol Basis Dis 1862:1827–1838. https://doi.org/10.1016/j.bbadis.2016.06.018
Oswald MCW, Garnham N, Sweeney ST, Landgraf M (2018) Regulation of neuronal development and function by ROS. FEBS Lett 592:679–691. https://doi.org/10.1002/1873-3468.12972
Calabrese V, Cornelius C, Rizzarelli E et al (2009) Nitric Oxide in Cell Survival: A Janus Molecule. Antioxid Redox Signal 11:2717–2739. https://doi.org/10.1089/ars.2009.2721
Dröge W (2002) Free Radicals in the Physiological Control of Cell Function. Physiol Rev 82:47–95. https://doi.org/10.1152/physrev.00018.2001
Holmström KM, Finkel T (2014) Cellular mechanisms and physiological consequences of redox-dependent signalling. Nat Rev Mol Cell Biol 15:411–421. https://doi.org/10.1038/nrm3801
Aguilera G, Colín-González AL, Rangel-López E et al (2018) Redox Signaling, Neuroinflammation, and Neurodegeneration. Antioxid Redox Signal 28:1626–1651. https://doi.org/10.1089/ars.2017.7099
Chen Y, Qin C, Huang J et al (2020) The role of astrocytes in oxidative stress of central nervous system: A mixed blessing. Cell Prolif 53(3):e12781. https://doi.org/10.1111/cpr.12781
Simpson DSA, Oliver PL (2020) ROS Generation in Microglia: Understanding Oxidative Stress and Inflammation in Neurodegenerative Disease. Antioxidants 9:743. https://doi.org/10.3390/antiox9080743
Cobley JN, Fiorello ML, Bailey DM (2018) 13 reasons why the brain is susceptible to oxidative stress. Redox Biol 15:490–503. https://doi.org/10.1016/j.redox.2018.01.008
Aoyama K (2021) Glutathione in the Brain. Int J Mol Sci 22:5010. https://doi.org/10.3390/ijms22095010
Lee M, Cho T, Jantaratnotai N et al (2010) Depletion of GSH in glial cells induces neurotoxicity: relevance to aging and degenerative neurological diseases. FASEB J 24:2533–2545. https://doi.org/10.1096/fj.09-149997
Arús BA, Souza DG, Bellaver B et al (2017) Resveratrol modulates GSH system in C6 astroglial cells through heme oxygenase 1 pathway. Mol Cell Biochem 428:67–77. https://doi.org/10.1007/s11010-016-2917-5
Liddelow SA, Marsh SE, Stevens B (2020) Microglia and Astrocytes in Disease: Dynamic Duo or Partners in Crime? Trends Immunol 41:820–835. https://doi.org/10.1016/j.it.2020.07.006
Jensen CJ, Massie A, De Keyser J (2013) Immune Players in the CNS: The Astrocyte. J Neuroimmune Pharmacol 8:824–839. https://doi.org/10.1007/s11481-013-9480-6
Han RT, Kim RD, Molofsky AV, Liddelow SA (2021) Astrocyte-immune cell interactions in physiology and pathology. Immunity 54:211–224. https://doi.org/10.1016/j.immuni.2021.01.013
Ransohoff RM (2016) How neuroinflammation contributes to neurodegeneration. Science 353:777–783. https://doi.org/10.1126/science.aag2590
Maragakis NJ, Rothstein JD (2004) Glutamate transporters: animal models to neurologic disease. Neurobiol Dis 15:461–473. https://doi.org/10.1016/j.nbd.2003.12.007
Rothstein JD, Dykes-Hoberg M, Pardo CA et al (1996) Knockout of Glutamate Transporters Reveals a Major Role for Astroglial Transport in Excitotoxicity and Clearance of Glutamate. Neuron 16:675–686. https://doi.org/10.1016/S0896-6273(00)80086-0
Rodríguez-Campuzano AG, Ortega A (2021) Glutamate transporters: Critical components of glutamatergic transmission. Neuropharmacology 192:108602. https://doi.org/10.1016/j.neuropharm.2021.108602
Trotti D, Danbolt NC, Volterra A (1998) Glutamate transporters are oxidant-vulnerable: a molecular link between oxidative and excitotoxic neurodegeneration? Trends Pharmacol Sci 19:328–334. https://doi.org/10.1016/S0165-6147(98)01230-9
Sitcheran R, Gupta P, Fisher PB, Baldwin AS (2005) Positive and negative regulation of EAAT2 by NF-κB: a role for N-myc in TNFα-controlled repression. EMBO J 24:510–520. https://doi.org/10.1038/sj.emboj.7600555
Soto C, Pritzkow S (2018) Protein misfolding, aggregation, and conformational strains in neurodegenerative diseases. Nat Neurosci 21:1332–1340. https://doi.org/10.1038/s41593-018-0235-9
Pinto JV, Passos IC, Librenza-Garcia D et al (2018) Neuron-glia Interaction as a Possible Pathophysiological Mechanism of Bipolar Disorder. Curr Neuropharmacol 16:519–532. https://doi.org/10.2174/1570159X15666170828170921
Mattson MP, Arumugam TV (2018) Hallmarks of Brain Aging: Adaptive and Pathological Modification by Metabolic States. Cell Metab 27:1176–1199. https://doi.org/10.1016/j.cmet.2018.05.011
Mehta A, Prabhakar M, Kumar P et al (2013) Excitotoxicity: Bridge to various triggers in neurodegenerative disorders. Eur J Pharmacol 698:6–18. https://doi.org/10.1016/j.ejphar.2012.10.032
Labzin LI, Heneka MT, Latz E (2018) Innate Immunity and Neurodegeneration. Annu Rev Med 69:437–449. https://doi.org/10.1146/annurev-med-050715-104343
Patel M (2016) Targeting Oxidative Stress in Central Nervous System Disorders. Trends Pharmacol Sci 37:768–778. https://doi.org/10.1016/j.tips.2016.06.007
Saggu R, Schumacher T, Gerich F et al (2016) Astroglial NF-kB contributes to white matter damage and cognitive impairment in a mouse model of vascular dementia. Acta Neuropathol Commun 4:76. https://doi.org/10.1186/s40478-016-0350-3
Li L, Acioglu C, Heary RF, Elkabes S (2021) Role of astroglial toll-like receptors (TLRs) in central nervous system infections, injury and neurodegenerative diseases. Brain Behav Immun 91:740–755. https://doi.org/10.1016/j.bbi.2020.10.007
Rojo AI, Innamorato NG, Martín-Moreno AM et al (2010) Nrf2 regulates microglial dynamics and neuroinflammation in experimental Parkinson’s disease: N RF 2 Regulates Microglial Dynamics. Glia 58:588–598. https://doi.org/10.1002/glia.20947
Rodríguez-Arellano JJ, Parpura V, Zorec R, Verkhratsky A (2016) Astrocytes in physiological aging and Alzheimer’s disease. Neuroscience 323:170–182. https://doi.org/10.1016/j.neuroscience.2015.01.007
Jyothi HJ, Vidyadhara DJ, Mahadevan A et al (2015) Aging causes morphological alterations in astrocytes and microglia in human substantia nigra pars compacta. Neurobiol Aging 36:3321–3333. https://doi.org/10.1016/j.neurobiolaging.2015.08.024
Souza DG, Bellaver B, Raupp GS et al (2015) Astrocytes from adult Wistar rats aged in vitro show changes in glial functions. Neurochem Int 90:93–97. https://doi.org/10.1016/j.neuint.2015.07.016
Santos CL, Roppa PHA, Truccolo P et al (2018) Age-Dependent Neurochemical Remodeling of Hypothalamic Astrocytes. Mol Neurobiol 55:5565–5579. https://doi.org/10.1007/s12035-017-0786-x
Morita M, Ikeshima-Kataoka H, Kreft M et al (2019) Metabolic Plasticity of Astrocytes and Aging of the Brain. Int J Mol Sci 20:E941. https://doi.org/10.3390/ijms20040941
Souza DG, Bellaver B, Bobermin LD et al (2016) Anti-aging effects of guanosine in glial cells. Purinergic Signal 12:697–706. https://doi.org/10.1007/s11302-016-9533-4
Palmer AL, Ousman SS (2018) Astrocytes and Aging Front Aging Neurosci 10:337. https://doi.org/10.3389/fnagi.2018.00337
Brawek B, Skok M, Garaschuk O (2021) Changing Functional Signatures of Microglia along the Axis of Brain Aging. Int J Mol Sci 22:1091. https://doi.org/10.3390/ijms22031091
Franceschi C, Garagnani P, Parini P et al (2018) Inflammaging: a new immune–metabolic viewpoint for age-related diseases. Nat Rev Endocrinol 14:576–590. https://doi.org/10.1038/s41574-018-0059-4
Batista CRA, Gomes GF, Candelario-Jalil E et al (2019) Lipopolysaccharide-Induced Neuroinflammation as a Bridge to Understand Neurodegeneration. Int J Mol Sci 20:2293. https://doi.org/10.3390/ijms20092293
Zhao J, Bi W, Xiao S et al (2019) Neuroinflammation induced by lipopolysaccharide causes cognitive impairment in mice. Sci Rep 9:5790. https://doi.org/10.1038/s41598-019-42286-8
Guerra M, Tortorelli LS, Galland F et al (2011) Lipopolysaccharide modulates astrocytic S100B secretion: a study in cerebrospinal fluid and astrocyte cultures from rats. J Neuroinflammation 8:128. https://doi.org/10.1186/1742-2094-8-128
Rosa PM, Martins LAM, Souza DO, Quincozes-Santos A (2018) Glioprotective Effect of Resveratrol: an Emerging Therapeutic Role for Oligodendroglial Cells. Mol Neurobiol 55:2967–2978. https://doi.org/10.1007/s12035-017-0510-x
Chien C-H, Lee M-J, Liou H-C et al (2016) Microglia-Derived Cytokines/Chemokines Are Involved in the Enhancement of LPS-Induced Loss of Nigrostriatal Dopaminergic Neurons in DJ-1 Knockout Mice. PLoS ONE 11:e0151569. https://doi.org/10.1371/journal.pone.0151569
Bronstein DM, Perez-Otano I, Sun V et al (1995) Glia-dependent neurotoxicity and neuroprotection in mesencephalic cultures. Brain Res 704:112–116. https://doi.org/10.1016/0006-8993(95)01189-7
Kramer-Hämmerle S, Rothenaigner I, Wolff H et al (2005) Cells of the central nervous system as targets and reservoirs of the human immunodeficiency virus. Virus Res 111:194–213. https://doi.org/10.1016/j.virusres.2005.04.009
Chivero ET, Guo M-L, Periyasamy P et al (2017) HIV-1 Tat Primes and Activates Microglial NLRP3 Inflammasome-Mediated Neuroinflammation. J Neurosci 37:3599–3609. https://doi.org/10.1523/JNEUROSCI.3045-16.2017
Sénécal V, Barat C, Tremblay MJ (2021) The delicate balance between neurotoxicity and neuroprotection in the context of HIV -1 infection. Glia 69:255–280. https://doi.org/10.1002/glia.23904
Meertens L, Labeau A, Dejarnac O et al (2017) Axl Mediates ZIKA Virus Entry in Human Glial Cells and Modulates Innate Immune Responses. Cell Rep 18:324–333. https://doi.org/10.1016/j.celrep.2016.12.045
Ledur PF, Karmirian K, da Pedrosa C, SG, et al (2020) Zika virus infection leads to mitochondrial failure, oxidative stress and DNA damage in human iPSC-derived astrocytes. Sci Rep 10:1218. https://doi.org/10.1038/s41598-020-57914-x
Bobermin LD, Quincozes-Santos A, Santos CL et al (2020) Zika virus exposure affects neuron-glia communication in the hippocampal slices of adult rats. Sci Rep 10:21604. https://doi.org/10.1038/s41598-020-78735-y
Song E, Zhang C, Israelow B et al (2021) Neuroinvasion of SARS-CoV-2 in human and mouse brain. J Exp Med 218:e20202135. https://doi.org/10.1084/jem.20202135
Crunfli F, Carregari VC, Veras FP, et al (2020) SARS-CoV-2 infects brain astrocytes of COVID-19 patients and impairs neuronal viability. Neurology
Srivastava DK, Bernhard SA (1987) Biophysical chemistry of metabolic reaction sequences in concentrated enzyme solution and in the cell. Annu Rev Biophys Biophys Chem 16:175–204. https://doi.org/10.1146/annurev.bb.16.060187.001135
Solomon T (2021) Neurological infection with SARS-CoV-2 - the story so far. Nat Rev Neurol 17:65–66. https://doi.org/10.1038/s41582-020-00453-w
Bathini P, Brai E, Auber LA (2019) Olfactory dysfunction in the pathophysiological continuum of dementia. Ageing Res Rev 55:100956. https://doi.org/10.1016/j.arr.2019.100956
Yuan T-F, Slotnick BM (2014) Roles of olfactory system dysfunction in depression. Prog Neuropsychopharmacol Biol Psychiatry 54:26–30. https://doi.org/10.1016/j.pnpbp.2014.05.013
Wender R, Brown AM, Fern R et al (2000) Astrocytic Glycogen Influences Axon Function and Survival during Glucose Deprivation in Central White Matter. J Neurosci 20:6804–6810. https://doi.org/10.1523/JNEUROSCI.20-18-06804.2000
Lee Y, Morrison BM, Li Y et al (2012) Oligodendroglia metabolically support axons and contribute to neurodegeneration. Nature 487:443–448. https://doi.org/10.1038/nature11314
Yang J, Ruchti E, Petit J-M et al (2014) Lactate promotes plasticity gene expression by potentiating NMDA signaling in neurons. Proc Natl Acad Sci 111:12228–12233. https://doi.org/10.1073/pnas.1322912111
Mosienko V, Teschemacher AG, Kasparov S (2015) Is L-lactate a novel signaling molecule in the brain? J Cereb Blood Flow Metab Off J Int Soc Cereb Blood Flow Metab 35:1069–1075. https://doi.org/10.1038/jcbfm.2015.77
Hayakawa K, Esposito E, Wang X et al (2016) Transfer of mitochondria from astrocytes to neurons after stroke. Nature 535:551–555. https://doi.org/10.1038/nature18928
Schousboe A, Bak LK, Waagepetersen HS (2013) Astrocytic Control of Biosynthesis and Turnover of the Neurotransmitters Glutamate and GABA. Front Endocrinol 4(4):102. https://doi.org/10.3389/fendo.2013.00102
Hertz L, Dringen R, Schousboe A, Robinson SR (1999) Astrocytes: glutamate producers for neurons. J Neurosci Res 57:417–428
Zhang J, Liu Q (2015) Cholesterol metabolism and homeostasis in the brain. Protein Cell 6:254–264. https://doi.org/10.1007/s13238-014-0131-3
Ioannou MS, Jackson J, Sheu S-H et al (2019) Neuron-Astrocyte Metabolic Coupling Protects against Activity-Induced Fatty Acid Toxicity. Cell 177:1522-1535.e14. https://doi.org/10.1016/j.cell.2019.04.001
Farina C, Aloisi F, Meinl E (2007) Astrocytes are active players in cerebral innate immunity. Trends Immunol 28:138–145. https://doi.org/10.1016/j.it.2007.01.005
Ubhi K, Rockenstein E, Mante M et al (2010) Neurodegeneration in a Transgenic Mouse Model of Multiple System Atrophy Is Associated with Altered Expression of Oligodendroglial-Derived Neurotrophic Factors. J Neurosci 30:6236–6246. https://doi.org/10.1523/JNEUROSCI.0567-10.2010
Alvarez JI, Katayama T, Prat A (2013) Glial influence on the blood brain barrier. Glia 61:1939–1958. https://doi.org/10.1002/glia.22575
Pöyhönen S, Er S, Domanskyi A, Airavaara M (2019) Effects of Neurotrophic Factors in Glial Cells in the Central Nervous System: Expression and Properties in Neurodegeneration and Injury. Front Physiol 10:486. https://doi.org/10.3389/fphys.2019.00486
Bankston AN, Mandler MD, Feng Y (2013) Oligodendroglia and neurotrophic factors in neurodegeneration. Neurosci Bull 29:216–228. https://doi.org/10.1007/s12264-013-1321-3
Bhatia TN, Pant DB, Eckhoff EA et al (2019) Astrocytes Do Not Forfeit Their Neuroprotective Roles After Surviving Intense Oxidative Stress. Front Mol Neurosci 12:87. https://doi.org/10.3389/fnmol.2019.00087
García-Krauss A, Ferrada L, Astuya A et al (2016) Dehydroascorbic Acid Promotes Cell Death in Neurons Under Oxidative Stress: a Protective Role for Astrocytes. Mol Neurobiol 53:5847–5863. https://doi.org/10.1007/s12035-015-9497-3
Dringen R, Hirrlinger J (2003) Glutathione Pathways in the Brain. Biol Chem 384(4):505–16. https://doi.org/10.1515/BC.2003.059
Dringen R, Pfeiffer B, Hamprecht B (1999) Synthesis of the Antioxidant Glutathione in Neurons: Supply by Astrocytes of CysGly as Precursor for Neuronal Glutathione. J Neurosci 19:562–569. https://doi.org/10.1523/JNEUROSCI.19-02-00562.1999
Paul BD, Sbodio JI, Snyder SH (2018) Cysteine Metabolism in Neuronal Redox Homeostasis. Trends Pharmacol Sci 39:513–524. https://doi.org/10.1016/j.tips.2018.02.007
Pope SAS, Milton R, Heales SJR (2008) Astrocytes Protect Against Copper-Catalysed Loss of Extracellular Glutathione. Neurochem Res 33:1410–1418. https://doi.org/10.1007/s11064-008-9602-3
Persson M, Rönnbäck L (2012) Microglial self-defence mediated through GLT-1 and glutathione. Amino Acids 42:207–219. https://doi.org/10.1007/s00726-011-0865-7
DeSilva TM, Kabakov AY, Goldhoff PE et al (2009) Regulation of Glutamate Transport in Developing Rat Oligodendrocytes. J Neurosci 29:7898–7908. https://doi.org/10.1523/JNEUROSCI.6129-08.2009
Ottestad-Hansen S, Hu QX, Follin-Arbelet VV et al (2018) The cystine-glutamate exchanger (xCT, Slc7a11) is expressed in significant concentrations in a subpopulation of astrocytes in the mouse brain. Glia 66:951–970. https://doi.org/10.1002/glia.23294
Sedlak TW, Paul BD, Parker GM et al (2019) The glutathione cycle shapes synaptic glutamate activity. Proc Natl Acad Sci 116:2701–2706. https://doi.org/10.1073/pnas.1817885116
Shinozaki Y, Shibata K, Yoshida K et al (2017) Transformation of Astrocytes to a Neuroprotective Phenotype by Microglia via P2Y 1 Receptor Downregulation. Cell Rep 19:1151–1164. https://doi.org/10.1016/j.celrep.2017.04.047
Norden DM, Fenn AM, Dugan A, Godbout JP (2014) TGFβ produced by IL-10 redirected astrocytes attenuates microglial activation: IL-10 Redirects Immune Activated Astrocytes. Glia 62:881–895. https://doi.org/10.1002/glia.22647
Yi W, Schlüter D, Wang X (2019) Astrocytes in multiple sclerosis and experimental autoimmune encephalomyelitis: Star-shaped cells illuminating the darkness of CNS autoimmunity. Brain Behav Immun 80:10–24. https://doi.org/10.1016/j.bbi.2019.05.029
Liu L, Liu J, Bao J et al (2020) Interaction of Microglia and Astrocytes in the Neurovascular Unit. Front Immunol 11:1024. https://doi.org/10.3389/fimmu.2020.01024
Hayes JD, Dinkova-Kostova AT (2014) The Nrf2 regulatory network provides an interface between redox and intermediary metabolism. Trends Biochem Sci 39:199–218. https://doi.org/10.1016/j.tibs.2014.02.002
Ahmed SMU, Luo L, Namani A et al (2017) Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim Biophys Acta BBA - Mol Basis Dis 1863:585–597. https://doi.org/10.1016/j.bbadis.2016.11.005
Bellezza I, Giambanco I, Minelli A, Donato R (2018) Nrf2-Keap1 signaling in oxidative and reductive stress. Biochim Biophys Acta BBA - Mol Cell Res 1865:721–733. https://doi.org/10.1016/j.bbamcr.2018.02.010
Kwak M-K, Itoh K, Yamamoto M, Kensler TW (2002) Enhanced expression of the transcription factor Nrf2 by cancer chemopreventive agents: role of antioxidant response element-like sequences in the nrf2 promoter. Mol Cell Biol 22:2883–2892. https://doi.org/10.1128/MCB.22.9.2883-2892.2002
Chorley BN, Campbell MR, Wang X et al (2012) Identification of novel NRF2-regulated genes by ChIP-Seq: influence on retinoid X receptor alpha. Nucleic Acids Res 40:7416–7429. https://doi.org/10.1093/nar/gks409
Mancuso C (2004) Heme Oxygenase and Its Products in the Nervous System. Antioxid Redox Signal 6:878–887. https://doi.org/10.1089/ars.2004.6.878
Liddell J (2017) Are Astrocytes the Predominant Cell Type for Activation of Nrf2 in Aging and Neurodegeneration? Antioxidants 6:65. https://doi.org/10.3390/antiox6030065
Bobermin LD, Roppa RHA, Quincozes-Santos A (2019) Adenosine receptors as a new target for resveratrol-mediated glioprotection. Biochim Biophys Acta Mol Basis Dis 1865:634–647. https://doi.org/10.1016/j.bbadis.2019.01.004
Bobermin LD, Souza DO, Gonçalves C-A, Quincozes-Santos A (2013) Lipoic acid protects C6 cells against ammonia exposure through Na+-K+-Cl− co-transporter and PKC pathway. Toxicol In Vitro 27:2041–2048. https://doi.org/10.1016/j.tiv.2013.07.006
dos Santos AQ, Nardin P, Funchal C et al (2006) Resveratrol increases glutamate uptake and glutamine synthetase activity in C6 glioma cells. Arch Biochem Biophys 453:161–167. https://doi.org/10.1016/j.abb.2006.06.025
Bellaver B, Souza DG, Bobermin LD et al (2015) Resveratrol Protects Hippocampal Astrocytes Against LPS-Induced Neurotoxicity Through HO-1, p38 and ERK Pathways. Neurochem Res 40:1600–1608. https://doi.org/10.1007/s11064-015-1636-8
Quincozes-Santos A, Bobermin LD, Latini A et al (2013) Resveratrol Protects C6 Astrocyte Cell Line against Hydrogen Peroxide-Induced Oxidative Stress through Heme Oxygenase 1. PLoS ONE 8:e64372. https://doi.org/10.1371/journal.pone.0064372
Lu X, Ma L, Ruan L et al (2010) Resveratrol differentially modulates inflammatory responses of microglia and astrocytes. J Neuroinflammation 7:46. https://doi.org/10.1186/1742-2094-7-46
Zhang F, Lu Y-F, Wu Q et al (2012) Resveratrol promotes neurotrophic factor release from astroglia. Exp Biol Med 237:943–948. https://doi.org/10.1258/ebm.2012.012044
Daverey A, Agrawal SK (2018) Pre and post treatment with curcumin and resveratrol protects astrocytes after oxidative stress. Brain Res 1692:45–55. https://doi.org/10.1016/j.brainres.2018.05.001
Bellaver B, Souza DG, Souza DO, Quincozes-Santos A (2014) Resveratrol increases antioxidant defenses and decreases proinflammatory cytokines in hippocampal astrocyte cultures from newborn, adult and aged Wistar rats. Toxicol Vitro Int J Publ Assoc BIBRA 28:479–484. https://doi.org/10.1016/j.tiv.2014.01.006
Bastianetto S, Ménard C, Quirion R (2015) Neuroprotective action of resveratrol. Biochim Biophys Acta BBA - Mol Basis Dis 1852:1195–1201. https://doi.org/10.1016/j.bbadis.2014.09.011
Yang X, Xu S, Qian Y, Xiao Q (2017) Resveratrol regulates microglia M1/M2 polarization via PGC-1α in conditions of neuroinflammatory injury. Brain Behav Immun 64:162–172. https://doi.org/10.1016/j.bbi.2017.03.003
Ghosh AK, Rao VR, Wisniewski VJ et al (2020) Differential Activation of Glioprotective Intracellular Signaling Pathways in Primary Optic Nerve Head Astrocytes after Treatment with Different Classes of Antioxidants. Antioxidants 9:324. https://doi.org/10.3390/antiox9040324
Ghaiad HR, Nooh MM, El-Sawalhi MM, Shaheen AA (2017) Resveratrol Promotes Remyelination in Cuprizone Model of Multiple Sclerosis: Biochemical and Histological Study. Mol Neurobiol 54:3219–3229. https://doi.org/10.1007/s12035-016-9891-5
Kodali M, Parihar VK, Hattiangady B et al (2015) Resveratrol Prevents Age-Related Memory and Mood Dysfunction with Increased Hippocampal Neurogenesis and Microvasculature and Reduced Glial Activation. Sci Rep 5:8075. https://doi.org/10.1038/srep08075
Yuan J, Liu W, Zhu H et al (2017) Curcumin inhibits glial scar formation by suppressing astrocyte-induced inflammation and fibrosis in vitro and in vivo. Brain Res 1655:90–103. https://doi.org/10.1016/j.brainres.2016.11.002
Bernardo A, Plumitallo C, De Nuccio C et al (2021) Curcumin promotes oligodendrocyte differentiation and their protection against TNF-α through the activation of the nuclear receptor PPAR-γ. Sci Rep 11:4952. https://doi.org/10.1038/s41598-021-83938-y
Parada E, Buendia I, Navarro E et al (2015) Microglial HO-1 induction by curcumin provides antioxidant, antineuroinflammatory, and glioprotective effects. Mol Nutr Food Res 59:1690–1700. https://doi.org/10.1002/mnfr.201500279
Rastogi M, Ojha RP, Sagar C et al (2014) Protective effect of curcuminoids on age-related mitochondrial impairment in female Wistar rat brain. Biogerontology 15:21–31. https://doi.org/10.1007/s10522-013-9466-z
Daverey A, Agrawal SK (2016) Curcumin alleviates oxidative stress and mitochondrial dysfunction in astrocytes. Neuroscience 333:92–103. https://doi.org/10.1016/j.neuroscience.2016.07.012
Yu Y, Shen Q, Lai Y et al (2018) Anti-inflammatory Effects of Curcumin in Microglial Cells. Front Pharmacol 9:386. https://doi.org/10.3389/fphar.2018.00386
Cianciulli A, Calvello R, Porro C et al (2016) PI3k/Akt signalling pathway plays a crucial role in the anti-inflammatory effects of curcumin in LPS-activated microglia. Int Immunopharmacol 36:282–290. https://doi.org/10.1016/j.intimp.2016.05.007
Zhang J, Zheng Y, Luo Y et al (2019) Curcumin inhibits LPS-induced neuroinflammation by promoting microglial M2 polarization via TREM2/ TLR4/ NF-κB pathways in BV2 cells. Mol Immunol 116:29–37. https://doi.org/10.1016/j.molimm.2019.09.020
Eun C-S, Lim J-S, Lee J et al (2017) The protective effect of fermented Curcuma longa L. on memory dysfunction in oxidative stress-induced C6 gliomal cells, proinflammatory-activated BV2 microglial cells, and scopolamine-induced amnesia model in mice. BMC Complement Altern Med 17:367. https://doi.org/10.1186/s12906-017-1880-3
Nery-Flores SD, Ramírez-Herrera MA, Mendoza-Magaña ML et al (2019) Dietary Curcumin Prevented Astrocytosis, Microgliosis, and Apoptosis Caused by Acute and Chronic Exposure to Ozone. Molecules 24:2839. https://doi.org/10.3390/molecules24152839
Valles SL, Dolz-Gaiton P, Gambini J et al (2010) Estradiol or genistein prevent Alzheimer’s disease-associated inflammation correlating with an increase PPARγ expression in cultured astrocytes. Brain Res 1312:138–144. https://doi.org/10.1016/j.brainres.2009.11.044
Lu H, Shi J-X, Zhang D-M et al (2009) Inhibition of hemolysate-induced iNOS and COX-2 expression by genistein through suppression of NF-кB activation in primary astrocytes. J Neurol Sci 278:91–95. https://doi.org/10.1016/j.jns.2008.12.007
Du Z-R, Feng X-Q, Li N et al (2018) G protein-coupled estrogen receptor is involved in the anti-inflammatory effects of genistein in microglia. Phytomedicine 43:11–20. https://doi.org/10.1016/j.phymed.2018.03.039
Ohgomori T, Jinno S (2019) Cuprizone-induced demyelination in the mouse hippocampus is alleviated by phytoestrogen genistein. Toxicol Appl Pharmacol 363:98–110. https://doi.org/10.1016/j.taap.2018.11.009
Xu SL, Bi CWC, Choi RCY et al (2013) Flavonoids Induce the Synthesis and Secretion of Neurotrophic Factors in Cultured Rat Astrocytes: A Signaling Response Mediated by Estrogen Receptor. Evid Based Complement Alternat Med 2013:1–10. https://doi.org/10.1155/2013/127075
Jeong J-W, Lee HH, Han MH et al (2014) Anti-inflammatory effects of genistein via suppression of the toll-like receptor 4-mediated signaling pathway in lipopolysaccharide-stimulated BV2 microglia. Chem Biol Interact 212:30–39. https://doi.org/10.1016/j.cbi.2014.01.012
Zhou X, Yuan L, Zhao X et al (2014) Genistein antagonizes inflammatory damage induced by β-amyloid peptide in microglia through TLR4 and NF-κB. Nutrition 30:90–95. https://doi.org/10.1016/j.nut.2013.06.006
Subedi L, Ji E, Shin D et al (2017) Equol, a Dietary Daidzein Gut Metabolite Attenuates Microglial Activation and Potentiates Neuroprotection In Vitro. Nutrients 9:207. https://doi.org/10.3390/nu9030207
Martini LH, Jung F, Soares FA et al (2007) Naturally Occurring Compounds Affect Glutamatergic Neurotransmission in Rat Brain. Neurochem Res 32:1950–1956. https://doi.org/10.1007/s11064-007-9393-y
Ariyani W, Miyazaki W, Amano I et al (2020) Soy Isoflavones Accelerate Glial Cell Migration via GPER-Mediated Signal Transduction Pathway. Front Endocrinol 11:554941. https://doi.org/10.3389/fendo.2020.554941
da Silva SI, Schaffer LF, Busanello A et al (2019) Isoflavones prevent oxidative stress and inhibit the activity of the enzyme monoamine oxidase in vitro. Mol Biol Rep 46:2285–2292. https://doi.org/10.1007/s11033-019-04684-z
Bobermin LD, Weber FB, dos Santos TM et al (2020) Sulforaphane Induces Glioprotection After LPS Challenge. Cell Mol Neurobiol. https://doi.org/10.1007/s10571-020-00981-5
Danilov CA, Chandrasekaran K, Racz J et al (2009) Sulforaphane protects astrocytes against oxidative stress and delayed death caused by oxygen and glucose deprivation. Glia 57:645–656. https://doi.org/10.1002/glia.20793
Wu Y, Gao M, Wu J et al (2019) Sulforaphane triggers a functional elongation of microglial process via the Akt signal. J Nutr Biochem 67:51–62. https://doi.org/10.1016/j.jnutbio.2019.01.019
Qin S, Yang C, Huang W et al (2018) Sulforaphane attenuates microglia-mediated neuronal necroptosis through down-regulation of MAPK/NF-κB signaling pathways in LPS-activated BV-2 microglia. Pharmacol Res 133:218–235. https://doi.org/10.1016/j.phrs.2018.01.014
Subedi L, Lee J, Yumnam S et al (2019) Anti-Inflammatory Effect of Sulforaphane on LPS-Activated Microglia Potentially through JNK/AP-1/NF-κB Inhibition and Nrf2/HO-1 Activation. Cells 8:194. https://doi.org/10.3390/cells8020194
Kleinkauf-Rocha J, Bobermin LD, de Machado P, M, et al (2013) Lipoic acid increases glutamate uptake, glutamine synthetase activity and glutathione content in C6 astrocyte cell line. Int J Dev Neurosci 31:165–170. https://doi.org/10.1016/j.ijdevneu.2012.12.006
Sanadgol N, Golab F, Askari H et al (2018) Alpha-lipoic acid mitigates toxic-induced demyelination in the corpus callosum by lessening of oxidative stress and stimulation of polydendrocytes proliferation. Metab Brain Dis 33:27–37. https://doi.org/10.1007/s11011-017-0099-9
Santos CL, Bobermin LD, Souza DG et al (2015) Lipoic acid and N-acetylcysteine prevent ammonia-induced inflammatory response in C6 astroglial cells: The putative role of ERK and HO1 signaling pathways. Toxicol In Vitro 29:1350–1357. https://doi.org/10.1016/j.tiv.2015.05.023
Scumpia PO, Kelly-Scumpia K, Stevens BR (2014) Alpha-lipoic acid effects on brain glial functions accompanying double-stranded RNA antiviral and inflammatory signaling. Neurochem Int 64:55–63. https://doi.org/10.1016/j.neuint.2013.11.006
Koriyama Y, Nakayama Y, Matsugo S et al (2013) Anti-inflammatory effects of lipoic acid through inhibition of GSK-3β in lipopolysaccharide-induced BV-2 microglial cells. Neurosci Res 77:87–96. https://doi.org/10.1016/j.neures.2013.07.001
Xiao L, Wei F, Zhou Y et al (2020) Dihydrolipoic Acid-Gold Nanoclusters Regulate Microglial Polarization and Have the Potential To Alter Neurogenesis. Nano Lett 20:478–495. https://doi.org/10.1021/acs.nanolett.9b04216
Hansel G, Ramos DB, Delgado CA et al (2014) The Potential Therapeutic Effect of Guanosine after Cortical Focal Ischemia in Rats. PLoS ONE 9:e90693. https://doi.org/10.1371/journal.pone.0090693
Jiang S, Khan MI, Lu Y et al (2003) Guanosine promotes myelination and functional recovery in chronic spinal injury. NeuroReport 14:2463–2467. https://doi.org/10.1097/00001756-200312190-00034
Bellaver B, Souza DG, Bobermin LD et al (2015) Guanosine inhibits LPS-induced pro-inflammatory response and oxidative stress in hippocampal astrocytes through the heme oxygenase-1 pathway. Purinergic Signal 11:571–580. https://doi.org/10.1007/s11302-015-9475-2
Dal-Cim T, Poluceno GG, Lanznaster D et al (2019) Guanosine prevents oxidative damage and glutamate uptake impairment induced by oxygen/glucose deprivation in cortical astrocyte cultures: involvement of A1 and A2A adenosine receptors and PI3K, MEK, and PKC pathways. Purinergic Signal 15:465–476. https://doi.org/10.1007/s11302-019-09679-w
Di Iorio P, Ballerini P, Traversa U et al (2004) The antiapoptotic effect of guanosine is mediated by the activation of the PI 3-kinase/AKT/PKB pathway in cultured rat astrocytes. Glia 46:356–368. https://doi.org/10.1002/glia.20002
Bobermin LD, Silva A, Souza DO, Quincozes-Santos A (2018) Differential effects of typical and atypical antipsychotics on astroglial cells in vitro. Int J Dev Neurosci 69:1–9. https://doi.org/10.1016/j.ijdevneu.2018.06.001
Kato T, Monji A, Hashioka S, Kanba S (2007) Risperidone significantly inhibits interferon-γ-induced microglial activation in vitro. Schizophr Res 92:108–115. https://doi.org/10.1016/j.schres.2007.01.019
Jeon S, Kim SH, Shin SY, Lee YH (2018) Clozapine reduces Toll-like receptor 4/NF-κB-mediated inflammatory responses through inhibition of calcium/calmodulin-dependent Akt activation in microglia. Prog Neuropsychopharmacol Biol Psychiatry 81:477–487. https://doi.org/10.1016/j.pnpbp.2017.04.012
Quincozes-Santos A, Bobermin LD, Tonial RPL et al (2010) Effects of atypical (risperidone) and typical (haloperidol) antipsychotic agents on astroglial functions. Eur Arch Psychiatry Clin Neurosci 260:475–481. https://doi.org/10.1007/s00406-009-0095-0
Quincozes-Santos A, Bobermin LD, Kleinkauf-Rocha J et al (2009) Atypical neuroleptic risperidone modulates glial functions in C6 astroglial cells. Prog Neuropsychopharmacol Biol Psychiatry 33:11–15. https://doi.org/10.1016/j.pnpbp.2008.08.023
Peng L, Verkhratsky A, Gu L, Li B (2015) Targeting astrocytes in major depression. Expert Rev Neurother 15:1299–1306. https://doi.org/10.1586/14737175.2015.1095094
Tramontina AC, Tramontina F, Bobermin LD et al (2008) Secretion of S100B, an astrocyte-derived neurotrophic protein, is stimulated by fluoxetine via a mechanism independent of serotonin. Prog Neuropsychopharmacol Biol Psychiatry 32:1580–1583. https://doi.org/10.1016/j.pnpbp.2008.06.001
Dall’Igna OP, Bobermin LD, Souza DO, Quincozes-Santos A, (2013) Riluzole increases glutamate uptake by cultured C6 astroglial cells. Int J Dev Neurosci 31:482–486. https://doi.org/10.1016/j.ijdevneu.2013.06.002
Daverey A, Agrawal SK (2020) Neuroprotective effects of Riluzole and Curcumin in human astrocytes and spinal cord white matter hypoxia. Neurosci Lett 738:135351. https://doi.org/10.1016/j.neulet.2020.135351
Bernstein H-G, Steiner J, Guest PC et al (2015) Glial cells as key players in schizophrenia pathology: recent insights and concepts of therapy. Schizophr Res 161:4–18. https://doi.org/10.1016/j.schres.2014.03.035
Valori CF, Guidotti G, Brambilla L, Rossi D (2019) Astrocytes: Emerging Therapeutic Targets in Neurological Disorders. Trends Mol Med 25:750–759. https://doi.org/10.1016/j.molmed.2019.04.010
Möller T, Boddeke HWGM (2016) GLIA Special Issue: Gliotherapeutics Glia 64:1608–1608. https://doi.org/10.1002/glia.23052
Acknowledgements
The authors are supported by the Universidade Federal do Rio Grande do Sul (UFRGS), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), and Fundação de Amparo à Pesquisa do Estado do Rio Grande do Sul (FAPERGS).
Author information
Authors and Affiliations
Contributions
AQS and LDB conceptualized the review. All authors written the original draft of the manuscript, revised, edited, and approved the manuscript.
Corresponding author
Ethics declarations
Ethical approval
Not applicable
Consent to participate
Not applicable
Consent for publication
Not applicable
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Quincozes-Santos, A., Santos, C.L., de Souza Almeida, R.R. et al. Gliotoxicity and Glioprotection: the Dual Role of Glial Cells. Mol Neurobiol 58, 6577–6592 (2021). https://doi.org/10.1007/s12035-021-02574-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12035-021-02574-9