
Abstract
Recent studies suggest an important role for l-homoarginine in cardiovascular, hepatic and neurological functions, as well as the regulation of glucose metabolism. However, little is known about whole-body l-homoarginine synthesis or its response to dietary l-arginine intake in animals. Four series of experiments were conducted to determine l-homoarginine synthesis and catabolism in pigs and rats. In Experiment 1, male and female pigs were fed a corn- and soybean meal-based diet supplemented with 0.0–2.42 % l-arginine-HCl. In Experiment 2, male and female rats were fed a casein-based diet, while receiving drinking water containing supplemental l-arginine-HCl to provide 0.0-3.6 g l-arginine/kg body-weight/day. In both experiments, urine collected from the animals for 24 h was analyzed for l-homoarginine and related metabolites. In Experiment 3, pigs and rats received a single oral dose of 1 or 10 mg l-homoarginine/kg body-weight, respectively, and their urine was collected for 24 h for analyses of l-homoarginine and related substances. In Experiment 4, slices of pig and rat tissues (including liver, brain, kidney, heart, and skeletal-muscle) were incubated for 1 h in Krebs-bicarbonate buffer containing 5 or 50 µM l-homoarginine. Our results indicated that: (a) animal tissues did not degrade L-homoarginine in the presence of physiological concentrations of other amino-acids; (b) 95–96 % of orally administered l-homoarginine was recovered in urine; (c) l-homoarginine was quantitatively a minor product of l-arginineg catabolism in the body; and (d) dietary l-arginine supplementation dose-dependently increased whole-body l-homoarginine synthesis. These novel findings provide a new framework for future studies of l-homoarginine metabolism and physiology in animals and humans.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
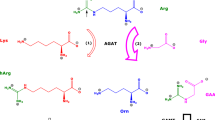
l-Homoarginine (hArg), a structural homologue of l-arginine (Arg), was discovered as a naturally occurring substance in plants (Bell 1962). Subsequently, synthesis of hArg from Arg and l-lysine in animals and humans was reported (Ryan and Wells 1964; Ryan et al. 1968, 1969). It is now known that rat liver, kidneys and the small intestine (Frenay et al. 2015; Hou et al. 2015; Pilz et al. 2015a), as well as human brain (Bernstein et al. 2015) possess the capacity for hArg synthesis. The responsible enzyme remains unknown (Tsikas and Kayacelebi 2014) and may be arginine:glycine amidinotransferase (AGAT) (Davids et al. 2012; Ryan et al. 1969).
Although recent studies suggest an important role for hArg in cardiovascular (Atzler et al. 2015; Pilz et al. 2015b), hepatic (Pilz et al. 2015a) and neurological (Bernstein et al. 2015) functions, as well as the regulation of glucose metabolism (Stockebrand et al. 2015), little is known about whole-body hArg synthesis or its responses to dietary Arg intake in animals (Tsikas and Wu 2015). The objective of this study was to determine hArg production in two animal models (pigs and rats) supplemented with various amounts of dietary Arg. Our results provide, for the first time, quantitative data on Arg metabolism via the hArg pathway and effects of Arg consumption on whole-body hArg synthesis in mammals.
Materials and methods
l-Arginine-HCl and l-alanine were products of Ajinomoto Inc. (Tokyo, Japan). Their purity was >99.9 %, as analyzed by high-performance liquid chromatography (HPLC) (Wu and Meininger 2008). hArg was purchased from Sigma Chemicals (St. Louis, MO, USA). The sources of other chemicals were the same as described previously (Dai et al. 2014b; Hou et al. 2015; Hu et al. 2015). The diets and drinking water for pigs and rats contained no hArg, nitrite, nitrate, creatine, creatinine, agmatine, or methylarginines. The experimental procedures were approved by the Institutional Agricultural Animal Care and Use Committee of Texas A&M University.
Experiment 1: effects of dietary Arg supplementation on urinary excretion of hArg in pigs
Pigs used for this study were the offspring of Yorkshire × Landrace dams and Duroc × Hampshire sires, and were maintained at the Texas A&M University Swine Center. Piglets were weaned at 21 days of age to a conventional corn- and soybean meal-based diet containing 21.0 % crude protein and 1.35 % Arg, as described previously (Hu et al. 2015). At 30 days of age, 18 male or 18 female pigs with similar body weights were assigned randomly to one of three treatment groups (0, 1.21, and 2.42 % Arg-HCl) on the basis of litter origin. There were 6 males and 6 females per dietary group. The three diets were prepared by adding 0, 1.21, or 2.42 % Arg-HCl to the basal diet at the expense of cornstarch. These doses of Arg-HCl provided 0, 1, and 2 % Arg, respectively. Isonitrogenous amounts of l-alanine were added to the diets supplemented with 0 and 1.21 % Arg-HCl (Hu et al. 2015). The animals were housed individually in pens at 23–25 °C and 55–60 % relative humidity. During the entire experimental period, pigs had free access to drinking water and were offered their respective diets at 31.5 g feed per kg body weight per day (divided in two equal meals at 8:00 AM and 5:00 PM). The pigs consumed all the feed provided each day. In the last week of the study, pigs were placed individually in metabolism cages, continuously fed their respective diets, and had free access to drinking water (double-distilled and deionized water). On day 120 of age, a 24-h urine collection into a bottle (containing antibiotics) was made from each pig (Wu et al. 1999). After the completion of urine collection, jugular vein blood samples (2 ml) were collected, into EDTA-containing tubes, from each pig at 4 h after feeding. The blood samples were centrifuged immediately at 10,000 g for 1 min. The supernatant fluid (plasma) was collected and then stored at −80 °C until analyzed.
Experiment 2: effects of dietary Arg supplementation on urinary excretion of hArg in rats
Male and female Sprague–Dawley rats (5 weeks of age) were purchased from Harlan Laboratories (Indianapolis, IN). Upon arrival at the Texas A&M University Kleberg animal facilities, all rats were housed individually in carbonate cages in an animal room at 25 °C and 60 % relative humidity with a 12-h light:12-h dark cycle. During a 1-week period of adaptation, the animals were fed a regular rodent diet (Product Cat #8604, Harlan Teklad) and had free access to drinking water (double-distilled and deionized water). At 6 weeks of age, rats were assigned randomly to one of three treatment groups (0, 2.18 or 4.35 g Arg-HCl/kg body weight/day). These doses of Arg-HCl provided 0, 1.8 or 3.6 g Arg/kg body weight/day, respectively. Isonitrogenous amounts of l-alanine were added to the drinking water (Yang et al. 2015). There were 6 male and 6 female rats per dietary group. During the entire period of the experiment, rats were fed a semi-purified diet containing 4.3 % fat (Jobgen et al. 2009) and had free access to their respective drinking water (double-distilled and deionized water). This enteral diet contained 0.61 % Arg and 0.44 % l-alanine, as analyzed by HPLC (Dai et al. 2014a). Because oral arginine administration of Arg to rats via drinking water tended to increase food intake (Jobgen et al. 2009), Arg-supplemented rats were individually pair-fed with rats in the isonitrogenous control group on a per kg body weight basis to ensure similar intakes of all nutrients (except for arginine and alanine) among the three groups. Consumption of food and water by rats was determined daily. Concentrations of Arg-HCl in drinking water were adjusted daily according to the volume of water consumed by rats to provide intakes of 0, 2.18 or 4.35 g Arg-HCl/kg body weight/day, as indicated previously. In the last week of the study, rats were placed individually in metabolism cages, continuously fed their respective diets, and had free access to drinking water (double-distilled and deionized water). On day 132 of age, 24-h urine collection into a bottle (containing antibiotics) was made from each rat (Wu et al. 1999). After the completion of urine collection, rats were food deprived for 5 h. Thereafter, rats were anesthetized with CO2 and euthanized by cervical dislocation. Cardiac blood samples (2 ml) were collected into heparinized tubes and centrifuged immediately at 10,000 g for 1 min. The supernatant fluid (plasma) was collected and then stored at −80 °C until analyzed.
Experiment 3: recovery of orally administered hArg in the urine of pigs and rats
Three-month-old male (n = 6) and female (n = 6) pigs (44 and 40 kg, respectively) and 3-month-old male (n = 6) and female (n = 6) rats (365 and 240 g, respectively) were placed individually in their metabolism cages and fed their respective diets, as described above. Pigs and rats received a single oral dose of hArg-HCl (dissolved in water) at 1 or 10 mg hArg/kg body weight, respectively. The volume of water was 5 and 1 ml for pigs and rats, respectively. Urine was then collected for 24 h, and then blood samples were obtained from the animals, as described previously. The amount of hArg in the 24-h urine excreted before the day of oral hArg administration was used to correct for basal amounts of hArg excreted into urine and thus calculate the recovery of orally administered hArg in the urine of animals.
Experiment 4: catabolism of hArg by animal tissues
Pigs and rats were fed as described in Exp. 1 and Exp. 2. Male (n = 6) and female (n = 6) pigs (44 and 40 kg, respectively; 121 days of age), as well as 133-day-old male (n = 6) and female (n = 6) rats (365 and 240 g, respectively) were euthanized, as we described previously (Hu et al. 2015; Yang et al. 2015). One hundred milligram slices of tissues (small intestine, large intestine liver, pancreas, spleen, heart, brain, kidney, skeletal muscle, white adipose tissue, thymus, mesenteric lymph nodes, skin, testes, or ovaries) were incubated at 37 °C for 1 h in the presence of 2 ml oxygenated Krebs bicarbonate buffer (95 % O2/5 % CO2, pH 7.4; Wu et al. 1994) containing 5 mM d-glucose, 5 or 50 µM hArg, and physiological concentrations of other AA (except for lysine) (Jobgen et al. 2009; Wu et al. 1997). Lysine was not included in the incubation medium to prevent hArg synthesis (Yang et al. 2015). At the end of the 0 and 1-h incubation period, 0.1 ml of 1.5 M HClO4 was added to the incubation medium. The acidified medium was neutralized with 0.05 ml of 2 M K2CO3. The whole tissue extract was analyzed for hArg.
Chemical analyses
Plasma and urine samples (1 ml) were deproteinized with an equal volume of 1.5 M HClO4, followed by addition of 0.5 ml 2 M K2CO3. The neutralized extracts were used for chemical analyses. Specifically, hArg, agmatine and Arg were determined by HPLC methods involving precolumn derivatization with o-phthaldialdehyde as described previously (Dai et al. 2014b; Hou et al. 2015; Wu and Meininger 2008). Creatinine was determined using a kit from Sigma Chemicals (St. Louis, MO, USA). NOx (nitrite plus nitrate) was measured by an HPLC method involving the enzymatic conversion of nitrate into nitrite and the subsequent derivatization of nitrite with 2,3-diaminonaphthalene to form 2,3-naphthotriazole (Jobgen et al. 2007). For determination of asymmetric dimethylarginine, NG-monomethyl arginine, and symmetric dimethylarginine, sample cleanup was performed using the Oasis MCX SPE column (1 ml, 30 mg) before HPLC analysis (Wu and Meininger 2008).
Statistical analysis
Results are expressed as mean ± SEM. Statistical analyses of data were performed by one-way analysis of variance using the General Linear Models procedures (Assaad et al. 2014). Differences among treatment means were determined using the Student–Newman–Keuls multiple comparison method (Assaad et al. 2014). A probability value of ≤0.05 was taken to indicate statistical significance.
Results
Concentrations of hArg and related metabolites in the plasma and urine of pigs and rats
Data on concentrations of hArg and related metabolites in the plasma and urine of pigs are summarized in Table 1. Male and female pigs responded similarly to dietary Arg supplementation. Specifically, concentrations of Arg, hArg and NOx in plasma, as well as concentrations of hArg, creatinine and NOx increased (P < 0.05) in a dose-dependent manner with dietary supplementation of 0, 315, and 630 mg Arg/kg body weight per day. Interestingly, concentrations of creatinine in plasma or concentrations of Arg in urine did not differ among treatment groups within each sex. Concentrations of methylarginines (formed by the enzymatic methylation of Arg residues in proteins and released by proteolysis) in the plasma or urine of pigs were not affected by dietary Arg supplementation (Table 2). Similar results were observed for rats supplemented with 0, 1.8 and 3.6 g Arg/kg body weight per day (Tables 3, 4).
In both pigs and rats, concentrations of hArg in plasma and urine were relatively low, compared with Arg, creatinine and NOx (Tables 1, 3). With reabsorption of water by kidney tubules, hArg was concentrated in the urine. While concentrations of creatinine and NOx in plasma were lower than those of Arg, concentrations of creatinine and NOx in urine were about 700 and 10–20 times those of Arg, respectively. Thus, reabsorption of creatinine and NOx by kidney tubules was limited in both pigs and rats.
Urinary excretion of hArg and related metabolites in pigs and rats
The volume of urine excreted by male and female pigs was not affected by dietary supplementation with 0, 315 and 630 mg Arg/kg body weight per day (Table 5). Likewise, urinary excretion of Arg did not differ among these three groups of pigs. In contrast, dietary supplementation with Arg dose-dependently increased (P < 0.05) urinary excretion of hArg and NOx in male and female pigs. When data from male and female pigs were combined, diertary supplementation with 0, 315 and 630 mg Arg/kg body weight/day enhanced (P < 0.05) urinary excretion of creatinine and agmatine in a dose-dependent manner (Table 6). Similar results were obtained for rats supplemented with 0, 1.8 and 3.6 g Arg/kg body weight per day (Table 7). In both pigs and rats, urinary excretion of hArg and agmatine was much lower than that of Arg, creatinine and NOx.
Recovery of orally administered hArg in the urine of pigs and rats
At 24 h after oral administration of hArg, concentrations of hArg in the plasma of pigs and rats (Table 8) were similar to those for animals not receiving an oral dose of hArg (Tables 1, 2). Approximately 96 and 95 % of orally administered hArg was recovered in the urine of pigs and rats, respectively (Table 8). The urinary recovery of hArg did not differ between males and females in either animal species. In both pigs and rats, oral administration of hArg did not affect (P > 0.05) concentrations of Arg, creatinine, agmatine, methylarginines, or NOx in plasma and urine, or urinary excretion of these metabolites.
Degradation of hArg by animal tissues
The limit of detection (defined as a signal-to-noise ratio of 3:1) for our determination of hArg in tissue extracts was 38.4 pmol/ml (Hou et al. 2015). Based on the HPLC analysis of hArg in neutralized tissue extracts (medium plus tissue) before and after a 1-h incubation period, there was no detectable disappearance of hArg from the incubation medium of any animal tissue studied, which included the small intestine, large intestine liver, pancreas, spleen, heart, brain, kidney, skeletal muscle, white adipose tissue, thymus, mesenteric lymph nodes, skin, testes, and ovaries from male and female pigs as well as male and female rats. These results indicate that pig and rat tissues do not degrade hArg in the presence of physiological concentrations of other AA.
Discussion
Arg is a substrate for the synthesis of proteins, hArg, NO, ornithine (the precursor of polyamines, proline, and glutamate), creatine, and agmatine in mammals (Brosnan and Brosnan 2007; Morris 2007; Tsikas and Wu 2015; Wu and Morris 1998). Although concentrations of hArg in tissues and physiological fluids (e.g., plasma, brain, and urine) have been reported for humans (Carmann et al. 2015; Haghikia et al. 2015; Hörster et al. 2015; May et al. 2015), little is known about hArg production in the whole body. Similarly, there is a paucity of information about effects of dietary Arg intake on hArg synthesis in animals or humans. Much-needed data on hArg synthesis and catabolism will expand our knowledge about quantitative aspects of Arg metabolism in the body.
hArg is a chemically stable metabolite in both humans and rats (Kayacelebi et al. 2015a, b; Yang et al. 2015). The major site of hArg synthesis may be the liver and kidneys (Ryan et al. 1969). hArg is not degraded by arginase and other enzymes in animal tissues (the present study), and is not likely a significant source of NO when present at physiologically low concentrations in plasma (~2 µM), as compared with 100–200 µM Arg (Wu et al. 2009). Thus, in vivo catabolism of hArg is limited in animals. Furthermore, enterocytes can absorb dietary hArg from the lumen of the small intestine via cationic AA transporters (Monné et al. 2015), but there is little uptake of hArg from arterial blood into the small intestine (Rutherfurd 2015; Yin et al. 2015). Therefore, urinary excretion of hArg can be a useful indicator of its whole-body production. This view is supported by our findings that 96 and 95 % of orally administered hArg were recovered in the urine of pigs and rats, respectively (Table 8). The unrecovered hArg may be lost through the skin and feces, or utilized by bacteria in the small intestine. In post-weaning pigs and rats, dietary Arg is extensively degraded by the gastrointestinal tract as 40 % of orally administered Arg does not enter the portal circulation (Wu 1998).
Intake of protein-bound Arg by pigs was 425 mg/kg body weight/day. With a 90 % digestibility, Arg derived from dietary protein was 382 mg/kg body weight/day. In pigs supplemented with 0, 315 and 630 mg Arg/kg body weight/day, total Arg intake was 382, 697, and 1012 mg/kg body weight/day. Urinary excretion of hArg represented <0.025 % of Arg intake in pigs. Likewise, the intake of protein-bound Arg by male and female rats was 278 and 302 mg/kg body weight/day, respectively. With a 90 % digestibility, Arg derived from the dietary protein was 264 and 287 mg/kg body weight/day in male and female rats, respectively. In rats supplemented with 0, 1.8 and 3.6 g Arg/kg body weight/day, total Arg intake by male rats was 0.26, 2.06, and 3.86 g/kg body weight/day, and total Arg intake by female rats was 0.29, 2.09, and 3.89 g/kg body weight/day. Urinary excretion of hArg represented <0.045 % of Arg intake in rats. Thus, hArg was quantitatively a minor metabolite of Arg in both pigs and rats. This conclusion, however, should not be taken to indicate a minor role for hArg in animal physiology and pathophysiology (Kayacelebi et al. 2015a, b, c; Khalil et al. 2015; Langen et al. 2015; Papageorgiou et al. 2015; Sasso et al. 2015). Future studies are warranted to determine effects of hArg in nutrient absorption, blood circulation, pregnancy, lactation, and metabolic control.
A novel and important finding from the present study is that increasing dietary intake of Arg dose-dependently enhanced whole-body hArg synthesis and concentrations of hArg in plasma in both pigs and rats (Tables 1, 2). Thus, even at high intakes of Arg (e.g., up to 630 and 3600 mg/kg body weight/day, respectively, in pigs and rats), the enzyme for hArg production did not appear to be saturated in animals and renal AGAT activity was further increased (Yang et al. 2015). This provides a basis for oral administration of Arg to increase hArg availability in animals and humans. Likewise, urinary output of hArg is markedly increased in patients with hyperargininemia due to arginase I deficiency (Schlune et al. 2015). Therefore, we suggest that low extracellular concentrations of Arg can limit hArg synthesis in vivo.
As noted previously, hArg is a poor substrate of NO synthase. At high concentrations (e.g., 1 mM), hArg can even inhibit NOS (Moali 1998; Schini et al. 1991). The Km value of NOS for hArg is about 20 times greater than that for Arg (Moali 1998). Given an exceedingly low concentrations of hArg (~2 µM) in plasma (Tables 1, 2) relative to those of Arg (100–250 µM) (Wu et al. 2009), it is unlikely that hArg is a significant substrate for whole-body NO production under physiological conditions. While rat liver arginase may use hArg as a substrate, its K m value for hArg is 7.2 mM (Reczkowski and Ash 1994), which is about 70 times that for concentrations of hArg in the liver (Yang et al. 2015). The V max values of rat liver arginase for Arg and hArg are 4380 and 34 µmol/min/mg protein, respectively (Reczkowski and Ash 1994). This indicates an exceedingly low activity of purified arginase toward hArg as compared with Arg. Thus, catabolism of hArg by arginase is likely negligible in mammals. In support of this notion, results of our research indicate that a number of tissues from pigs and rats do not degrade hArg in the presence of physiological concentrations of other AA found in the plasma of animals (the current study). These tissues include the small intestine, large intestine liver, pancreas, spleen, heart, brain, kidney, skeletal muscle, white adipose tissue, thymus, mesenteric lymph nodes, skin, testes, and ovaries. Therefore, we suggest that virtually little hArg is catabolized in animals and endogenously synthesized hArg is quantitatively excreted in their urine.
In conclusion, hArg was quantitatively a minor product of Arg metabolism in pigs and rats. Approximately 95–96 % of orally administered hArg was recovered in urine, indicating limited catabolism of hArg in the body. Animal tissues do not degrade hArg in the presence of physiological concentrations of other AA, and thus its urinary excretion is a valid indicator of in vivo production of hArg. Dietary supplementation with Arg increased whole-body hArg synthesis in a dose-dependent manner. These novel results provide a new framework for future studies of hArg metabolism, as well as the roles for hArg in animal and human physiology.
Abbreviations
- AA:
-
Amino acids
- AGAT:
-
l-Arginine:glycine amidinotransferase
- Arg:
-
l-Arginine
- hArg:
-
l-Homoarginine
- HPLC:
-
High-performance liquid chromatography
- NO:
-
Nitric oxide
- NOx:
-
Nitrite plus nitrate
References
Assaad H, Zhou L, Carroll RJ, Wu G (2014) Rapid publication-ready MS-Word tables for one-way ANOVA. SpringerPlus 3:474
Atzler D, Schwedhelm E, Choe CU (2015) l-homoarginine and cardiovascular disease. Curr Opin Clin Nutr Metab Care 18:83–88
Bell EA (1962) α, γ-Diaminobutyric acid in seeds of twelve species of Lathyrus and identification of a new natural amino-Acid, l-homoarginine, in seeds of other species toxic to man and domestic animals. Nature 193:1078–1079
Bernstein HG, Jäger K, Dobrowolny H, Steiner J, Keilhoff G, Bogerts B, Laube G (2015) Possible sources and functions of l-homoarginine in the brain: review of the literature and own findings. Amino Acids 47:1729–1740
Brosnan JT, Brosnan ME (2007) Creatine: endogenous metabolite, dietary, and therapeutic supplement. Annu Rev Nutr 27:241–261
Carmann C, Lilienthal E, Weigt-Usinger K, Schmidt-Choudhury A, Hörster I, Kayacelebi AA, Beckmann B, Chobanyan-Jurgens K, Tsikas D, Lücke T (2015) The l-arginine/NO pathway, homoarginine, and nitrite-dependent renal carbonic anhydrase activity in young people with type 1 diabetes mellitus. Amino Acids 47:1865–1874
Dai ZL, Wu ZL, Jia SC, Wu G (2014a) Analysis of amino acid composition in proteins of animal tissues and foods as pre-column o-phthaldialdehyde derivatives by HPLC with fluorescence detection. J Chromatogr B 964:116–127
Dai ZL, Wu ZL, Wang JJ, Wang XQ, Jia SC, Bazer FW, Wu G (2014b) Analysis of polyamines in biological samples by HPLC involving pre-column derivatization with o-phthalaldehyde and N-acetyl-l-cysteine. Amino Acids 46:1557–1564
Davids M, Ndika JD, Salomons GS, Blom HJ, Teerlink T (2012) Promiscuous activity of arginine:glycine amidinotransferase is responsible for the synthesis of the novel cardiovascular risk factor homoarginine. FEBS Lett 586:3653–3657
Frenay AS, Kayacelebi AA, van den Berg E, de Borst MH, Beckmann B, van Goor H, Bakker SJL, Tsikas D (2015) High urinary homoarginine excretion is associated with low rates of all-cause mortality and graft failure in renal transplant recipients. Amino Acids 47:1827–1836
Haghikia A, Kayacelebi AA, Beckmann B, Hanff E, Gold R, Haghikia A, Tsikas D (2015) Serum and cerebrospinal fluid concentrations of homoarginine, arginine, asymmetric and symmetric dimethylarginine, nitrite and nitrate in patients with multiple sclerosis and neuromyelitis optica. Amino Acids 47:1837–1845
Hörster I, Weigt-Usinger K, Carmann C, Chobanyan-Jürgens K, Köhler C, Schara U, Kayacelebi AA, Beckmann B, Tsikas D, Lücke T (2015) The l-arginine/NO pathway and homoarginine are altered in Duchenne muscular dystrophy and improved by glucocorticoids. Amino Acids 47:1853–1863
Hou YQ, Jia SC, Nawaratna G, Hu SD, Dahanayaka S, Bazer FW, Wu G (2015) Analysis of l-homoarginine in biological samples by HPLC involving pre-column derivatization with o-phthalaldehyde and N-acetyl-l-cysteine. Amino Acids 47:2005–2014
Hu SD, Li XL, Rezaei R, Meininger CJ, McNeal CJ, Wu G (2015) Safety of long-term dietary supplementation with l-arginine in pigs. Amino Acids 47:925–936
Jobgen WS, Jobgen SC, Li H, Meininger CJ, Wu G (2007) Analysis of nitrite and nitrate in biological samples using high-performance liquid chromatography. J Chromatogr B 851:71–82
Jobgen WJ, Meininger CJ, Jobgen SC, Li P, Lee MJ, Smith SB, Spencer TE, Fried SK, Wu G (2009) Dietary l-arginine supplementation reduces white-fat gain and enhances skeletal muscle and brown fat masses in diet-induced obese rats. J Nutr 139:230–237
Kayacelebi AA, Knöfel AK, Beckmann B, Hanff E, Warnecke G, Tsikas D (2015a) Measurement of unlabeled and stable isotope-labeled homoarginine, arginine and their metabolites in biological samples by GC-MS and GC-MS/MS. Amino Acids 47:2023–2034
Kayacelebi AA, Langen J, Weigt-Usinger K, Chobanyan-Jürgens K, Mariotti F, Schneider JY, Rothmann S, Frölich JC, Atzler D, Choe CU, Schwedhelm E, Huneau JF, Lücke T, Tsikas D (2015b) Biosynthesis of homoarginine (hArg) and asymmetric dimethylarginine (ADMA) from acutely and chronically administered free l-arginine in humans. Amino Acids 47:1893–1908
Kayacelebi AA, Willers J, Pham VV, Hahn A, Schneider JY, Rothmann S, Frölich JC, Tsikas D (2015c) Plasma homoarginine, arginine, asymmetric dimethylarginine and total homocysteine interrelationships in rheumatoid arthritis, coronary artery disease and peripheral artery occlusion disease. Amino Acids 47:1885–1891
Khalil A, Hardman L, O´Brien P (2015) The role of arginine, homoarginine and nitric oxide in pregnancy. Amino Acids 47:1715–1727
Langen J, Kayacelebi AA, Beckmann B, Weigt-Usinger K, Carmann C, Hörster I, Lilienthal E, Richter-Unruh A, Tsikas D, Lücke T (2015) Homoarginine (hArg) and asymmetric dimethylarginine (ADMA) in short stature children without and with growth hormone deficiency: hArg and ADMA are involved differently in growth in the childhood. Amino Acids 47:1875–1883
May M, Kayacelebi AA, Batkai S, Jordan J, Tsikas D, Engeli S (2015) Plasma and tissue homoarginine concentrations in healthy and obese humans. Amino Acids 47:1847–1852
Moali C, Boucher J-L, Sari M-A, Steuhr DJ, Mansuy D (1998) Substrate specificity of NO synthases: detailed comparison of l-arginine, homo-l-arginine, their Nɷ-hydroxy derivatives, and Nɷ-hydroxynor-l-rginine. Biochemistry 37:10453–10460
Monné M, Miniero DV, Daddabbo L, Palmieri L, Porcelli V, Palmieri F (2015) Mitochondrial transporters for ornithine and related amino acids: a review. Amino Acids 47:1763–1777
Morris SM Jr (2007) Arginine metabolism: boundaries of our knowledge. J Nutr 137:1602S–1609S
Papageorgiou N, Androulakis E, Papaioannou S, Antoniades C, Tousoulis D (2015) Homoarginine in the shadow of asymmetric dimethylarginine: from nitric oxide to cardiovascular disease. Amino Acids 47:1741–1750
Pilz S, Putz-Bankuti C, Meinitzer A, März W, Kienreich K, Stojakovic T, Pieber TR, Stauber RE (2015a) Association of homoarginine and methylarginines with liver dysfunction and mortality in chronic liver disease. Amino Acids 47:1817–1826
Pilz S, Meinitzer A, Gaksch M, Grübler M, Verheyen N, Drechsler C, Hartaigh BÓ, Lang F, Alesutan I, Voelkl J, März W, Tomaschitz A (2015b) Homoarginine in the renal and cardiovascular systems. Amino Acids 47:1703–1713
Reczkowski RS, Ash DE (1994) Rat liver arginase: kinetic mechanism, alternate substrates, and inhibitors. Arch Biochem Biophys 312:31–37
Rutherfurd SM (2015) Use of the guanidination reaction for determining reactive lysine, bioavailable lysine and gut endogenous lysine. Amino Acids 47:1805–1815
Ryan WL, Wells IC (1964) Homocitrulline and homoarginine synthesis from lysine. Science 144:1122–1123
Ryan WL, Barak AJ, Johnson RJ (1968) Lysine, homocitrulline, and homoarginine metabolism by the isolated perfused rat liver. Arch Biochem Biophys 123:294–297
Ryan WL, Johnson RJ, Dimari S (1969) Homoarginine synthesis by rat kidney. Arch Biochem Biophys 131:521–526
Sasso S, Dalmedico L, Magro DD, Pereira EM, Wyse AT, de Lima DD (2015) Differential in vitro effects of homoarginine on oxidative stress in plasma, erythrocytes, kidney and liver of rats in the absence and in the presence α-tocopherol, ascorbic acid or L-NAME. Amino Acids 47:1931–1939
Schini VB, Vanhoutte PM (1991) l-arginine evokes both endothelium-dependent and -independent relaxations in l-arginine-depleted aortas of the rat. Circ Res 68:209–216
Schlune A, vom Dahl S, Häussinger D, Ensenauer R, Mayatepek E (2015) Hyperarginemia due to arginase I deficiency: the original patients and their natural history, and a review of the literature. Amino Acids 47:1751–1762
Stockebrand M, Hornig S, Neu A, Atzler D, Cordts K, Böger RH, Isbrandt D, Schwedhelm E, Choe C (2015) Homoarginine supplementation improves blood glucose in diet-induced obese mice. Amino Acids 47:1921–1929
Tsikas D, Kayacelebi AA (2014) Do homoarginine and asymmetric dimethylarginine act antagonistically in the cardiovascular system? Circ J 78:2092–2093
Tsikas D, Wu G (2015) Homoarginine, arginine, and relatives: analysis, metabolism, transport, physiology, and pathology. Amino Acids 47:1697–1702
Wu G (1998) Intestinal mucosal amino acid catabolism. J Nutr 128:1249–1252
Wu G (2013) Amino acids: biochemistry and nutrition. CRC Press, Boca Raton
Wu G, Meininger CJ (2008) Analysis of citrulline, arginine, and methylarginines using high-performance liquid chromatography. Methods Enzymol 440:177–189
Wu G, Morris SM Jr (1998) Arginine metabolism: nitric oxide and beyond. Biochem J 336:1–17
Wu G, Davis PK, Flynn NE, Knabe DA, Davidson JT (1997) Endogenous synthesis of arginine plays an important role in maintaining arginine homeostasis in postweaning growing pigs. J Nutr 127:2342–2349
Wu G, Knabe DA, Flynn NE (1994) Synthesis of citrulline from glutamine in pig enterocytes. Biochem J 299:115–121
Wu G, Flynn NE, Flynn SP, Jolly CA, Davis PK (1999) Dietary protein or arginine deficiency impairs constitutive and inducible nitric oxide synthesis by young rats. J Nutr 129:1347–1354
Wu G, Collins JK, Perkins-Veazie P, Siddiq M, Dolan KD, Kelly KA, Heaps CL, Meininger CJ (2007) Dietary supplementation with watermelon pomace juice enhances arginine availability and ameliorates the metabolic syndrome in Zucker diabetic fatty rats. J Nutr 137:2680–2685
Wu G, Bazer FW, Davis TA, Kim SW, Li P, Rhoads JM, Satterfield MC, Smith SB, Spencer TE, Yin YL (2009) Arginine metabolism and nutrition in growth, health and disease. Amino Acids 37:153–168
Yang Y, Wu ZL, Jia SC, Dahanayaka S, Feng S, Meininger CJ, McNeal CJ, Wu G (2015) Safety of long-term dietary supplementation with l-arginine in rats. Amino Acids 47:1907–1920
Yin J, Ren W, Hou Y, Wu M, Xiao H, Duan J, Zhao Y, Li T, Yin Y, Wu G, Nyachoti CM (2015) Use of homoarginine for measuring true ileal digestibility of amino acids in food protein. Amino Acids 47:1795–1803
Acknowledgments
This research was supported by grants from Hubei Provincial Key Project for Scientific and Technical Innovation (2014ABA022), Hubei Hundred Talent program, Natural Science Foundation of Hubei Province (2013CFA097 and 2013CFB325), the International Council of Amino Acid Science (Brussels, Belgium), the Agriculture and Food Research Initiative Competitive Grant from the Animal Growth & Nutrient Utilization Program of the USDA National Institute of Food and Agriculture (2014-67015-21770), and Texas A&M AgriLife Research (H-8200). Y.Q. Hou, D.S. Che, and F.L. Wang were supported by Fellowships from the China Scholarship Council.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The use of animals for this research was approved by the Institutional Animal Care and Use Committee of Texas A&M University.
Conflict of interest
The authors declare that they have no conflict of interest.
Informed consent
This was an animal study and did not involve informed consent.
Additional information
Y. Hou and S. Hu contributed equally to this work
Rights and permissions
About this article

Cite this article
Hou, Y., Hu, S., Jia, S. et al. Whole-body synthesis of l-homoarginine in pigs and rats supplemented with l-arginine. Amino Acids 48, 993–1001 (2016). https://doi.org/10.1007/s00726-015-2145-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00726-015-2145-4