
Abstract
Of all the types of enzyme-catalyzed reactions, hydrolytic transformations involving amide and ester bonds are the easiest to perform using proteases, esterases, or lipases. The key features that have made hydrolases the favorite class of enzymes for organic chemists during the past two decades are their lack of sensitive cofactors (which otherwise would need to be recycled) and the large number of readily available enzymes possessing relaxed substrate specificities to choose from. About two-thirds of the total research in the field of biotransformations has been performed using hydrolytic enzymes of this type [1, 2].
Access provided by Autonomous University of Puebla. Download chapter PDF
2.1 Hydrolytic Reactions
Of all the types of enzyme-catalyzed reactions, hydrolytic transformations involving amide and ester bonds are the easiest to perform using proteases, esterases, or lipases. The key features that have made hydrolases the favorite class of enzymes for organic chemists during the past two decades are their lack of sensitive cofactors (which otherwise would need to be recycled) and the large number of readily available enzymes possessing relaxed substrate specificities to choose from. About two-thirds of the total research in the field of biotransformations has been performed using hydrolytic enzymes of this type [1, 2]. The reversal of the reaction, giving rise to ester or amide synthesis, has been particularly well investigated using enzymes in solvent systems of low water activity. The special methodologies involved in this latter type of reaction are described in Sect. 3.1.
Other, more complex applications of hydrolases, such as those involving the formation and/or cleavage of phosphate esters, epoxides, nitriles, and organohalides, are described elsewhere in this book. In contrast to the group of proteases, esterases and lipases, they have had less impact on organic chemistry, although their synthetic potential should not be underestimated.
2.1.1 Mechanistic and Kinetic Aspects
The mechanism of amide- and ester-hydrolyzing enzymes is very similar to that observed in the chemical hydrolysis by a base. A nucleophilic group from the active site of the enzyme attacks the carbonyl group of the substrate ester or amide. This nucleophilic ‘chemical operator’ can be either the hydroxy group of a serine (e.g., pig liver esterase, subtilisin, and the majority of microbial lipases), a carboxyl group of an aspartic acid (e.g., pepsin) [3], or the thiol functionality of cysteine (e.g., papain) [4–6].
The mechanism, which has been elucidated in greater detail, is that of the serine hydrolases [7, 8] (see Scheme 2.1): Two additional groups (Asp and His) located close to the serine residue (which is the actual reacting chemical operator at the active site) form the so-called catalytic triad [9–12].Footnote 1 The special arrangement of these three groups effects a decrease of the pK a of the serine hydroxy group thus enabling it to perform a nucleophilic attack on the carbonyl group of the substrate R1–CO–OR2 (step I). Thus, the acyl moiety of the substrate becomes covalently linked to the enzyme, forming the ‘acyl-enzyme intermediate’ by liberating the leaving group (R2–OH). Then a nucleophile (Nu), usually water, can in turn attack the acyl-enzyme intermediate, regenerating the enzyme and releasing a carboxylic acid R1–COOH (step II).

The serine hydrolase mechanism
When the enzyme is operating in an environment of low water activity – in other words, at low water concentrations – any other nucleophile can compete with the water for the acyl-enzyme intermediate, thus leading to a number of synthetically useful transformations:
-
Attack of another alcohol R4–OH leads to a different ester R1–CO–OR4. This is an interesterification reaction, called enzymatic ‘acyl transfer’ [13, 14].
-
The action of ammonia furnishes a carboxamide R1–CO–NH2 via an ammonolysis reaction [15, 16].
-
An incoming amine R3–NH2 results in the formation of an N-substituted amide R1–CO–NH–R3, yielding an enzymatic aminolysis of esters [17, 18].
-
Peracids of type R1–CO–OOH are formed when hydrogen peroxide is acting as the nucleophile [19].
-
Hydrazinolysis provides access to hydrazides [20, 21], and the action of hydroxylamine results in the formation of hydroxamic acid derivatives [22]. However, both of the latter transformations have not been used extensively.
-
Thiols (which would lead to thioesters) are unreactive [23].
During the course of all of these reactions, any type of chirality in the substrate is ‘recognized’ by the enzyme, which causes a preference for one of the two possible stereochemical pathways for a reaction. The value of this discrimination is a crucial parameter since it stands for the ‘selectivity’ of the reaction. The latter is governed by the reaction kinetics. It should be noted, that the following chapter is not an elaboration on enzyme kinetics, but rather a compilation of the most important conclusions needed for obtaining optimal results from stereoselective enzymatic transformations.
Since hydrolases nicely exemplify all different types of chiral recognition, we will discuss the underlying principles of these chiral recognition processes and the corresponding kinetic implications here [24]. Most of these types of transformations can be found within other groups of enzymes as well, and the corresponding rules can be applied accordingly.
2.1.1.1 Enantioface Differentiation
Hydrolases can distinguish between the two enantiomeric faces of achiral substrates such as enol esters possessing a plane of symmetry within the molecule [25]. The attack of the enzyme’s nucleophilic chemical operator predominantly occurs from one side, leading to an unsymmetric enolization of the unstable free enol towards one preferred side within the chiral environment of the enzyme’s active site [26]. During the course of the reaction a new center of chirality is created in the product (Scheme 2.2).

Enantioface differentiation (achiral substrates)
2.1.1.2 Enantiotopos Differentiation
If prochiral substrates possessing two chemically identical but enantiotopic reactive groups X (designated pro-R and pro-S) are subjected to an enzymatic transformation such as hydrolysis, a chiral discrimination between them occurs during the transformation of group X into Y, thus leading to a chiral product (Scheme 2.3). During the course of the reaction the plane of symmetry within the substrate is broken. The single-step asymmetric hydrolysis of a prochiral α,α-disubstituted malonic diester by pig liver esterase or α-chymotrypsin is a representative example [27]. Here, the reaction terminates after a single reaction step at the carboxylate monoester stage since highly polar compounds of such type are heavily hydrated in an aqueous medium and are therefore generally not accepted by hydrolases [28].

Enantiotopos differentiation (prochiral substrates)
On the other hand, when the substrate is a diacetate, the resulting monoester is less polar and thus usually undergoes further cleavage in a second step to yield an achiral diol [29]. However, since the second step is usually slower, the chiral monoester can be trapped in fair yield if the reaction is carefully monitored.
Similarly, the two chemically identical groups X, positioned on carbon atoms of opposite (R,S)-configuration in a meso-substrate, can react at different rates in a hydrolase-catalyzed reaction (Scheme 2.4). So, the optically inactive meso-substrate is transformed into an optically active product due to the transformation of one of the reactive groups from X into Y along with the destruction of the plane of symmetry within the substrate. Numerous open-chain or cyclic cis-meso-diesters have been transformed into chiral monoesters by this technique [30]. Again, for dicarboxylates the reaction usually stops after the first step at the carboxylate monoester stage, whereas two hydrolytic steps are usually observed with diacetoxy esters [31]. The theoretical yield of chiral product from single-step reactions based on an enantioface or enantiotopos differentiation or a desymmetrization of meso-compounds is always 100%.

Desymmetrization of meso-substrates
If required, the interconversion of a given chiral hemiester product into its mirror-image enantiomer can be achieved by a simple two-step protection–deprotection sequence. Thus, regardless of the stereopreference of the enzyme which is used to perform the desymmetrization of the bifunctional prochiral or meso-substrate, both enantiomers of the product are available and no ‘unwanted’ enantiomer is produced. This technique is often referred to as the ‘meso-trick’ [25].
Since hydrolytic reactions are performed in an aqueous environment, they are virtually completely irreversible.Footnote 2 The kinetics of all of the single-step reactions described above is very simple (Fig. 2.1): a prochiral or a meso-substrate S is transformed into two enantiomeric products P and Q at different rates, determined by the apparent first-order rate constants k 1 and k 2, respectively (Schemes 2.2–2.4). The selectivity of the reaction (α [32]) is only governed by the ratio of k 1/k 2, which is independent of the conversion and therefore remains constant throughout the reaction. Thus, the optical purity of the product (e.e.P) is not dependent on the extent of the conversion. Also, the selectivity observed in such a reaction cannot be improved by stopping the reaction at different extents of conversion, but only by changing the ‘environment’ of the system (e.g., via substrate modification, choice of another enzyme, the addition of organic cosolvents, and variations in temperature or pH). Different techniques for improving the selectivity of enzymatic reactions by variations in the ‘environment’ are presented on pp. 77–84 and 108–110.

Single-step kinetics
As mentioned above, occasionally a second successive reaction step cannot be avoided with bifunctional prochiral or meso-diesters (Schemes 2.3 and 2.4). For such types of substrates the reaction does not terminate at the chiral carboxylate monoester stage to give the desired products P and Q (step 1), but rather proceeds via a second step (usually at a slower rate) to yield an achiral product (R). Here, the reaction kinetics become more complicated.
As depicted in Fig. 2.2, the ratio of P and Q – i.e., the optical purity of the product (e.e.) – depends now on four rate constants, k 1 through k 4, since the second hydrolytic step cannot be neglected. From the fact that enzymes usually show a continuous preference for reactive groups with the same chirality,Footnote 3 one may conclude that if S is transformed more quickly into P, Q will be hydrolyzed faster into diol R than P. Thus, the rate constants governing the selectivity of the reaction are often at an order of k 1 > k 2 and k 4 > k 3. Notably, the optical purity of the product monoester (e.e.) becomes a function of the conversion of the reaction, and generally follows the curve shown in Fig. 2.2.

Double-step kinetics
Early stages of the reaction, the optical purity of the product is mainly determined by the selectivity (α) of the first reaction step, which constitutes an enantiotopos or enantioface differentiation, depending on the type of substrate.
As the reaction proceeds, the second hydrolytic step, being a kinetic resolution, starts to take place to a more significant extent due to the increased formation of monoester P/Q, and its apparent ‘opposite’ selectivity compared to that of the first step (remember that k 1 > k 2, k 4 > k 3) leads to an enhancement of optical purity of the product (e.e.P). In contrast, the product concentration [P + Q] follows a bell-shaped curve: After having reached a maximum at a certain conversion, the product concentration [P + Q] finally drops off again when most of the substrate S is consumed and the second hydrolytic step to form R at the expense of P + Q constitutes the main reaction. The same analogous considerations are pertinent for the reverse situation – an esterification reaction.
In general, it can be stated that the ratio of reaction rates of the first and the second step [(k 1 + k 2)/(k 3 + k 4)] has a major impact on the chemical yield of P + Q, whereas the symmetry of the selectivities [(k 1 > k 2, k 3 > k 4 or k 1 > k 2, k 4 > k 3)] determines the optical purity of the product. In order to obtain a high chemical yield, the first step should be considerably faster than the second to ensure that the chiral product can be accumulated, because then it is formed faster than it is further converted [(k 1 + k 2) ≫ (k 3 + k 4)]. For a high e.e.P, the selectivities of both steps should match each other (k 1 > k 2, k 4 > k 3), i.e., if P is formed predominantly in the first step from S, it should react at a slower rate than Q in the second step. Figure 2.2 shows a typical example of such a double-step process, where the first step is about ten times faster than the second, with selectivities matching (k 1 = 100, k 2 = 10, k 3 = 1, k 4 = 10).
In addition to trial-and-error experiments (i.e., by stopping such double-step reactions at various intervals and checking the yield and optical purity of the product), the e.e.-conversion dependence may also be calculated [33]. Data on the amounts of substrate S and monoester P and Q and its optical purity measured at various intervals can be used to determine the kinetic constants k 1 through k 4 for a given reaction by using the computer program ‘SeKiRe’ [34]. Thus, the enantiomeric excess of the monoester may be predicted as a function of its percentage present in the reaction mixture. The validity of this method has been verified by the desymmetrization of a prochiral meso-diacetate using pig liver esterase (PLE) and porcine pancreatic lipase (PPL) as shown in Scheme 2.5 [35].

Desymmetrization of a meso-diacetate
2.1.1.3 Enantiomer Differentiation
When a racemic substrate is subject to enzymatic hydrolysis, chiral discrimination of the enantiomers occurs [36]. It should be noted that the chirality does not necessarily have to be of a central type, but can also be axial or planar to be ‘recognized’ by enzymes (Scheme 1.2). Due to the chirality of the active site of the enzyme, one enantiomer fits better into the active site than its mirror-image counterpart and is therefore converted at a higher rate, resulting in a kinetic resolution of the racemate. The vast majority of enzymatic transformations constitute kinetic resolutions and, interestingly, this potential of hydrolytic enzymes was realized as early as 1903 [37]! It is a remarkable observation that in biotransformations, kinetic resolutions outnumber desymmetrization reactions by about 1:4, which is presumably due to the fact that there are many more racemic compounds possible as opposed to prochiral and meso-analogs [38].
The most striking difference from the above-mentioned types of desymmetrization reactions, which show a theoretical yield of 100%, is that in kinetic resolution each of the enantiomers can be obtained in only 50% yield.
In some ideal cases, the difference in the reaction rates of both enantiomers is so extreme that the ‘good’ enantiomer is transformed quickly and the other is not converted at all. Then the enzymatic reaction will cease automatically at 50% conversion when there is nothing left of the more reactive enantiomer (Scheme 2.6) [39].

Enantiomer differentiation
In practice, however, most cases of enzymatic resolution of racemic substrates are not as ideal, i.e., in which one enantiomer is rapidly converted and the other not at all. The difference in – or more precisely the ratio of – the reaction rates of the enantiomers is not infinite, but measurable. The thermodynamic reasons for this have been discussed in Chap. 1. What one observes in these cases is not a complete standstill of the reaction at 50% conversion but a marked slowdown in reaction rate at around this point. In these numerous cases one encounters some crucial dependencies:
-
The velocity of the transformation of each enantiomer varies with the degree of conversion, since the ratio of the two substrate enantiomers does not remain constant during the reaction.
-
Therefore, the optical purity of both substrate (e.e.S) and product (e.e.P) becomes a function of the conversion.
A very useful treatment of the kinetics of enzymatic resolution, describing the dependency of the conversion (c) and the enantiomeric excess of substrate (e.e.S) and product (e.e.P), was developed by C.J. Sih in 1982 [40] on a theoretical basis described by K.B. Sharpless [41] and K. Fajans [42]. The parameter describing the selectivity of a resolution was introduced as the dimensionless ‘enantiomeric ratio’ (E), which remains constant throughout the reaction and is only determined by the ‘environment’ of the system [43–46].Footnote 4 E corresponds to the ratio of the relative second-order rate constants (v A, v B) of the individual substrate enantiomers (A, B) and is related to the k cat and K m values of enantiomers A and B according to Michaelis–Menten kinetics as follows (for the thermodynamic background see Fig. 1.7):
‘Enantiomeric ratio’ is not to be confused with the term ‘enantiomer ratio’ (e.r.), which is used to quantify the enantiomeric composition of a mixture of enantiomers (e.r. = [A]/[B]) [47].
Related alternative methods for the experimental determination of E-values have been proposed [48–50].
Irreversible Reaction. Hydrolytic reactions in aqueous solution can be regarded as completely irreversible due to the high ‘concentration’ of water present (55.5 mol/L). Assuming negligible enzyme inhibition, thus both enantiomers of the substrate are competing freely for the active site of the enzyme, Michaelis–Menten kinetics effectively describe the reaction in which two enantiomeric substrates (A and B) are transformed by an enzyme (Enz) into the corresponding enantiomeric products (P and Q, Fig. 2.3).

Enzymatic kinetic resolution (irreversible reaction)
Instead of determining all individual rate constants (k cat, KM) for each of the enantiomers (a wearisome task for synthetic organic chemists, particularly when A and B are not available in enantiopure form), in order to gain access to the corresponding relative rates governing the selectivity of the reaction, the ratio of the initial reaction rates of the substrate enantiomers (E = v A/v B) can be mathematically linked to the conversion (c) of the reaction, and the optical purities of substrate (e.e.S) and product (e.e.P). In practice, these parameters are usually much easier to determine and do not require the availability of pure enantiomers.
The dependence of the selectivity and the conversion of the reaction is:
For the product | For the substrate |
|---|---|
\( E = \displaystyle\frac{{{\hbox{ln[1}} - {\hbox{c(1}} + {\hbox{e}}{\hbox{.e}}{{.}_{\hbox{P}}}{)]}}}{{{\hbox{ln[1}} - {\hbox{c(1}} - {\hbox{e}}{\hbox{.e}}{{.}_{\hbox{P}}}{)]}}} \) | \( E = \displaystyle\frac{{{\hbox{ln[(1}} - {\hbox{c)(1}} - {\hbox{e}}{\hbox{.e}}{{.}_{\hbox{S}}}{)]}}}{{{\hbox{ln[(1}} - {\hbox{c)(1}} + {\hbox{e}}{\hbox{.e}}{{.}_{\hbox{S}}}{)]}}} \) |
The above-mentioned equations give reliable results except for very low and very high levels of conversion, where accurate measurement is impeded by errors derived from sample manipulation. In such cases, the following equation is recommended instead, because here only values for the optical purities of substrate and product need to be measured. The latter are relative quantities in contrast to the conversion, which is an absolute quantity [51].
Two examples of enzymatic resolutions with selectivities of E = 5 and E = 20 are depicted in Fig. 2.4. The curves show that the product (P + Q) can be obtained in its highest optical purities before 50% conversion, where the enzyme can freely choose the ‘well-fitting’ enantiomer from the racemic mixture. So, the ‘well-fitting’ enantiomer is predominantly depleted from the reaction mixture during the course of the reaction, leaving behind the ‘poor-fitting’ counterpart. Beyond 50% conversion, the enhanced relative concentration of the ‘poor-fitting’ counterpart leads to its increased transformation by the enzyme. Thus, the e.e.P rapidly decreases beyond 50% conversion.

Dependence of optical purities (e.e.S/e.e.P) on the conversion
Analogous trends are seen for the optical purity of the residual slow-reacting enantiomer of the substrate (e.e.S, as shown in the ‘substrate’ curve). Its optical purity remains low before 40%, then climbs significantly at around 50%, and reaches its maximum beyond the 60% conversion point.
High optical purity of substrate can easily be reached by extending the reaction to the appropriate point of conversion (≥60%), but an attractive optical purity value for the product demands a very high selectivity for the enzyme-catalyzed reaction.
Using the equations discussed above, the expected optical purity of substrate and product can be calculated for a chosen point of conversion and the enantiomeric ratio (E) can be determined as a convenient conversion-independent value for the ‘enantioselectivity’ of an enzymatic resolution. Free shareware programs for the calculation of the enantiomeric ratio for irreversible reactions can be obtained from the internet [52, 53]. As a rule of thumb, enantiomeric ratios below 15 are inacceptable for practical purposes. They can be regarded as being moderate to good in the range of 15–30, and above this value they are excellent. However, values of E > 200 cannot be accurately determined due to the inaccuracies emerging from the determination of the enantiomeric excess (e.g., by NMR, HPLC, or GC), because in this range even an extremely small variation of e.e.S or e.e.P causes a significant change in the numerical value of E.
In order to obtain optimal results from resolutions of numerous racemic substrates which exhibit moderate selectivities (E values ca. 20), one can proceed as follows (see Fig. 2.5): The reaction is terminated at a conversion of 40%, where the ‘product’ curve reaches its optimum in chemical and optical yield being closest to the ‘ideal’ point X (step I). The product is isolated and the remaining substrate – showing a low optical purity at this stage of conversion – is subjected to a second hydrolytic step, until an overall conversion of about 60% is reached, where the ‘substrate’ curve is closest to X (step II). Now, the substrate is harvested with an optimal chemical and optical yield and the 20% of product from the second step is sacrificed or recycled. This two-step process [54] can be used to allow practical use of numerous enzyme-catalyzed kinetic resolutions which show incomplete selectivities.

Two-step enzymatic resolution
Reversible Reaction. The situation becomes more complicated when the reaction is reversible [55].Footnote 5 This is the case if the concentration of the nucleophile which attacks the acyl-enzyme intermediate is limited and is not in excess (like water in a hydrolytic reaction). In this situation, the equilibrium constant (K) of the reaction – neglected in the irreversible type of reaction – plays an important role and therefore has to be determined.
The equations linking the enantioselectivity of the reaction (the enantiomeric ratio E), the conversion (c), the optical purities of substrate (e.e.S) and product (e.e.P), and the equilibrium constant K are as follows:
For the product | For the substrate |
|---|---|
\( E = \displaystyle\frac{{{\hbox{ln[1}} - \hbox{(1} + {\hbox{K)c(1}} + {\hbox{e}}{.}{{\hbox{e.}}_{\hbox{P}}}{)]}}}{{{\hbox{ln[1}} - \hbox{(1} + {\hbox{K)c(1}} - {\hbox{e}}{.}{{\hbox{e.}}_{\hbox{P}}}{)]}}} \) | \( E = \displaystyle\frac{{{\hbox{ln[1}} - \hbox{(1} + {\hbox{K)(c}} + {\hbox{e}}{\hbox{.e}}{{.}_{\hbox{S}}}{\hbox{\{ 1}} - {\hbox{c\} )]}}}}{{{\hbox{ln[1}} - \hbox{(1} + {\hbox{K)(c}} - {\hbox{e}}{\hbox{.e}}{{.}_{\hbox{S}}}{\hbox{\{ 1}} - {\hbox{c\})]}}}} \) |
As shown in Fig. 2.6, the product curve of an enzymatic resolution following a reversible reaction type remains almost the same. However, a significant difference compared to the irreversible case is found in the substrate curve: particularly at higher levels of conversion (beyond 70%) the reverse reaction (i.e., esterification instead of a hydrolysis) starts to predominate. Since the main steric requirements and hence the preferred chirality of the substrate stays the same, it is clear that the same enantiomer from the substrate and the product react preferentially in both the forward and the reverse reaction. Assuming that A is the better substrate than B, accumulation of product P and unreacted B will occur. For the reverse reaction, however, P is a better substrate than Q, because it is of the same chirality as A and therefore it will be transformed back into A at a faster rate than B into Q. As a result, the optical purity of the remaining substrate is depleted as the conversion increases. In other words, the reverse reaction, predominantly taking place at higher rates of conversion, constitutes a second – and in this case an undesired – selection of chirality which causes a depletion of e.e. of the remaining substrate.

Enzymatic kinetic resolution (reversible reaction)
All attempts of improving the optical purity of substrate and product of reversible enzymatic resolutions are geared at shifting the reaction out of the equilibrium to obtain an irreversible type. The easiest way to achieve this is to use an excess of cosubstrate: in order to obtain an equilibration constant of K > 10, about 20 M equivalents of nucleophile vs. substrate are considered to be sufficient to obtain a virtually irreversible type of reaction, in most cases. Other techniques, such as using special cosubstrates which cause an irreversible type of reaction, are discussed in Sect. 3.1.1.
Sequential Biocatalytic Resolutions. For a racemic substrate bearing two chemically and stereochemically identical reactive groups, an enzymatic resolution proceeds through two consecutive steps via an intermediate monoester stage. During the course of such a reaction the substrate is forced to enter the active site of the enzyme twice – it is therefore ‘double-selected’. Since each of the selectivities of both of the sequential steps determine the final optical purity of the product, exceptionally high selectivities can be achieved by using such a ‘double-sieving’ procedure.
As depicted in Fig. 2.7, a bifunctional racemic substrate consisting of its enantiomers A and B is enzymatically resolved via a first step to give the intermediate enantiomeric products P and Q. The selectivity of this step is governed by the constants k 1 and k 3. Then, both of the intermediate monoester products (P, Q) undergo a second reaction step, the selectivity of which is determined by k 2 and k 4, to form the enantiomeric final reaction products R and S. As a result, the optical purity of the substrate (A, B), the intermediate monoester (P, Q), and the final products (R, S) are a function of the conversion of the reaction, as shown by the curve in Fig. 2.7. The selectivities of each of the steps (E 1 and E 2) can be determined experimentally and the optical purities of the three species e.e.A/B, e.e.P/Q, and e.e.R/S can be calculated [57, 58]. Free shareware computer programs for the analysis, simulation, and optimization of such processes can be obtained over the internet [59].

Sequential enzymatic kinetic resolution via hydrolysis and esterification
It has been shown that the maximum overall selectivity (E tot) of a sequential kinetic resolution can be related to the individual selectivities (E 1, E 2) of each of the steps [60]. E tot represents the enantioselectivity that a hypothetical single-step resolution would need to yield the enantiomeric purity of the two-step resolution.
This technique has been proven to be highly flexible. It was shown to work successfully not only in a hydrolytic reaction using cholesterol esterase [61] or microbial cells [62], but also in the reverse esterification direction in an organic solvent catalyzed by a Pseudomonas sp. lipase (Scheme 2.7). In a related fashion, a successful sequential resolution of a bifunctional 1,2-amine via ester aminolysis was reported [63].

Sequential enzymatic resolution via hydrolysis and esterification
A special type of sequential enzymatic resolution involving a hydrolysis-esterification [64] or an alcoholysis-esterification sequence [65] is depicted in Fig. 2.8. In view of the mechanistic symmetry of enzymatic acyl transfer reactions (Scheme 3.6), the resolution of a racemic alcohol can be effected by enantioselective hydrolysis of the corresponding ester or by esterification of the alcohol. As the biocatalyst displays the same stereochemical preference in both reactions, the desired product can be obtained with higher optical yields, if the two steps are coupled sequentially. The basis of this approach parallels that of product recycling in hydrolytic reactions. However, tedious chromatographic separation of the intermediates and the accompanying re-esterification is omitted.

Mechanism of concurrent sequential enzymatic kinetic resolution via hydrolysis-esterification in aqueous-organic solvent
As shown in Scheme 2.8, the racemic starting acetate (A/B) is hydrolyzed to give alcohols (P/Q) in an organic medium containing a minimum amount of water, which in turn, by the action of the same lipase, are re-esterified with cyclohexanoic acid present in the mixture. Thus, the alcohol moiety of the substrate has to enter the active site of the lipase twice during the course of its transformation into the final product ester (R/S). An apparent selectivity of E tot = 400 was achieved in this way, whereas the corresponding isolated single-step resolutions of this process were E 1 = 8 for the hydrolysis of acetate A/B, and E 2 = 97 for the esterification of alcohol P/Q with cyclohexanoic acid.

Sequential enzymatic kinetic resolution via hydrolysis-esterification
2.1.1.4 Deracemization
Despite its widespread use, kinetic resolution has several disadvantages for preparative applications, particularly on an industrial scale. After all, an ideal process should lead to a single enantiomeric product in 100% chemical yield. The drawbacks of kinetic resolution are as follows:
-
The theoretical yield of each enantiomer is limited to 50%. Furthermore, in general only one stereoisomer is desired and there is little or no use for the other.
-
Separation of the product from the remaining substrate may be laborious, in particular when simple extraction or distillation fails [66].
-
As explained above, the optical purity of substrate and/or product is often less than perfect for kinetic reasons.
To overcome these disadvantages by avoiding the occurrence of the undesired ‘wrong’ enantiomer, several strategies are possible [67, 68]. All of these processes which lead to the formation of a single stereoisomeric product from a racemate are called ‘deracemizations’ [69].
Repeated Resolution. In order to avoid the loss of half of the material in kinetic resolution, it has been a common practice to racemize the unwanted enantiomer after separation from the desired product and to subject it again to kinetic resolution in a subsequent cycle, until virtually all of the racemic material has been converted into a single stereoisomeric product. For obvious reasons, this laborious procedure is not justified for laboratory-scale reactions, but it is a viable option for resolutions on an industrial scale, in particular for continuous processes, where the re-racemized material is simply fed back into the subsequent batch of the resolution process. At first sight, repeated resolution appears less than ideal and it certainly lacks synthetic elegance, bearing in mind that an infinite number of cycles are theoretically required to transform all of the racemic starting material into a single stereoisomer. Upon closer examination, though, re-racemization holds certain merits: a simple calculation shows that even if only 50% of the desired enantiomer is obtained after a single cycle, the overall (theoretical) yield increases to ~94% after only four cycles [70].
In practice, however, deracemization via repeated resolution is often plagued by low overall yields due to the harsh reaction conditions required for (chemical) racemization [71]. In view of the mild reaction conditions displayed by enzymes, there is a great potential for biocatalytic racemization based on the use of racemases of EC-class 5 [72, 73].
In-Situ Inversion. The final outcome of a kinetic resolution of a racemate is a mixture of enantiomeric product and substrate. Separating them by physical or chemical means is often tedious and might pose a serious drawback to commercial applications, especially if the mixture comprises an alcohol and an ester. However, if the molecule has only a single center of chirality, the alcohol may be chemically inverted into its enantiomer before separating the products (Scheme 2.9) [74, 75]. Introduction of a good leaving group, L (e.g., tosylate, triflate, nitrate, or Mitsunobu intermediate) yields an activated ester, which can be hydrolyzed with inversion of configuration, while the stereochemistry of the remaining carboxylic acid substrate ester is retained during hydrolysis. As a result, a single enantiomer is obtained as the final product. Since the e.e.S and e.e.P are a function of the conversion, it is obvious that the point where the kinetic resolution is terminated and the in-situ inversion is performed, has to be carefully chosen in order to obtain a maximum of the final e.e.P. The optimal value for the conversion can be calculated as a function of the E value of the reaction, and it is usually at or slightly beyond a conversion of 50% [76, 77].

Kinetic resolution with in-situ inversion
Dynamic resolution is a more elegant approach [78–80]. This comprises a classic resolution with an additional feature, i.e., the resolution is carried out using conditions under which the enantiomers of the substrate are in a rapid equilibrium (racemizing). Thus, as the well-accepted substrate-enantiomer is depleted by the enzyme, the equilibrium is constantly adjusted by racemization of the poorly accepted counterpart. To indicate the nonstatic character of such processes, the term ‘dynamic resolution’ has been coined [81, 82].Footnote 6
In this case, several reactions have to occur simultaneously and their relative rates determine the stereochemical outcome of the whole process (Fig. 2.9):

Kinetic resolution with in-situ racemization
-
The enzyme should display high specificity for the enantiomeric substrates R/S (k R ≫ k S or k S ≫ k R).
-
Spontaneous hydrolysis (k spont) should be a minimum since it would yield racemic product.
-
Racemization of the substrate should occur at an equal or higher rate compared to the biocatalytic reaction in order to provide a sufficient amount of the ‘well-fitting’ substrate enantiomer from the ‘poor-fitting’ counterpart (k Subrac ≥ k R or k S, resp.).
-
Racemization of the product (k Prodrac ) should be minimal.
Although the above-mentioned criteria are difficult to meet experimentally, the benefits are impressive. Examples of this type of biotransformation have increased recently [83–89]; several examples are given in subsequent chapters.
The kinetics of a dynamic resolution is outlined in the following example [79, 90]. Figure 2.9 shows the e.e.S and e.e.P plotted for an enantiomeric ratio of E ~ 10. In a classic resolution process, the product is formed in ~83% e.e. at the very beginning of the reaction, but this value rapidly decreases when the reaction is run towards ~50% conversion as indicated by the symbol ‘*’. In a dynamic process, this depletion does not occur, because the enzyme always encounters racemic substrate throughout the reaction since the ‘well-fitting’ enantiomer is not depleted but constantly restored from the ‘poor-fitting’ counterpart via racemization. Thus, e.e.P remains constant throughout the reaction as indicated by the dashed arrow.
The e.e.P of dynamic processes is related to the enantioselectivity (E value) through the following formulas [91]:
In the case where the racemization (k Subrac ) is limited, the dynamic resolution gradually turns into a classic kinetic resolution pattern. Figure 2.9 shows the extent of the depletion of e.e.P depending on the conversion for several ratios of k Subrac /k R (E ~ 10). As can be expected, e.e.P decreases only slightly during the early stage of the reaction because the fast-reacting enantiomer is sufficiently available during this period. At higher levels of conversion, however, a serious drop in e.e.P will occur if the racemization cannot cope with the demand of the enzyme for the faster-reacting substrate enantiomer.
It is obvious that a high e.e.P for dynamic resolutions can only be achieved for reactions displaying excellent selectivities. For example, values for E ~ 19 and ~ 40 will lead to an e.e.P of 90% and 95%, respectively, but for an enantiomeric excess of 98% an enantiomeric ratio of ~100 is required.
2.1.2 Hydrolysis of the Amide Bond
The enzymatic hydrolysis of the carboxamide bond is associated to the biochemistry of amino acids and peptides [92]. The world production of enantiomerically pure amino acids accounted for more than 0.5 million tons of material and a market of ca. US $2 billion [93] per annum in 1992. The three amino acids dominating this area with respect to output and value (l-glutamic acid, l-lysine, and d,l-methionine) are produced by fermentation and by synthesis. However, a considerable number of optically pure d- and l-amino acids are prepared by using one of the enzymatic methods discussed below. l-Amino acids are increasingly used as additives for animal feed, for infusion solutions and as enantiopure starting materials for the synthesis of pharma- and agrochemicals or artificial sweeteners. Selected amino acids possessing the unnatural d-configuration have gained an increasing importance as bioactive compounds or components of such agents. For instance, d-phenylglycine and its p-hydroxy derivative are used for the synthesis of antibiotics such as ampicillin and amoxicillin, respectively, and d-valine is an essential component of the insecticidal synthetic pyrethroid fluvalinate (Table 2.1; Scheme 2.208).
Among the principal methods for the enzymatic synthesis of enantiomerically pure amino acids depicted in Scheme 2.10, the most widely applied strategy is the resolution of racemic starting material (synthetically prepared from inexpensive bulk chemicals) employing easy-to-use hydrolytic enzymes such as proteases, esterases, and lipases. In contrast, more complex procedures requiring special expertise are the (1) reductive amination of α-keto acids using α-amino acid dehydrogenases (pp. 165–166), (2) asymmetric addition of ammonia onto α,β-unsaturated carboxylic acids catalyzed by lyases (Sect. 2.5.2), and (3) amino-group transfer using transaminases (Sect. 2.6.2) [94–96].

Important enzymatic routes to enantiomerically pure α-amino acids
The generally appliable hydrolytic methods have been selected from the numerous strategies of enzymatic amino acid synthesis [92, 97–103]. They also are useful tools for the preparation of enantiomerically pure nonnatural amino acids [104], not only for industrial needs but also for research on a laboratory scale.
There is a common pattern to the majority of hydrolase reactions involving α-amino acid derivatives: the substrate enantiomer possessing the ‘natural’ l-configuration is accepted by the enzyme, while the ‘unnatural’ d-counterpart remains unchanged and thus can be recovered from the reaction medium. Different enzymes with the opposite enantiopreference are available for some selected processes such as the hydantoinase and acylase method (see below) to transform the desired enantiomer. Using strictly l-specific enzyme systems, additional synthetic protection and/or deprotection steps are required in those cases where the unnatural d-amino acid constitutes the desired product. The work-up procedure is usually easy: thus, the difference in solubility of the product and the remaining substrate at different pH in the aqueous medium facilitates their separation by conventional ion-exchange or extraction methods. Alternatively, highly insoluble Schiff bases can be prepared from aldehydes by condensation with N-unprotected amino acid derivatives in order to facilitate their isolation. The free amino acids are readily obtained from these derivatives by mild acid hydrolysis without loss of optical purity.
However, there is a limitation to the majority of these methods: the α-carbon atom bearing the amino group must not be fully substituted, since such bulky substrates are generally not accepted by hydrolases. Thus, optically pure α-methyl or α-ethyl amino acids are generally not accessible by these methods, although some exceptions are known [105, 106].
The recycling of the undesired enantiomer from the enzymatic resolution is of crucial importance particularly on an industrial scale [107]. The classical chemical method consists of the thermal racemization of an amino acid ester at about 150–170°C. Milder conditions can be employed for the racemization of the corresponding amides via intermediate formation of Schiff bases with aromatic aldehydes such as benzaldehyde or salicylaldehyde (Scheme 2.14). More recently, intense research has been devoted to the use of isomerase enzymes (such as amino acid racemases [108]) aiming at the development of dynamic resolution processes.
2.1.2.1 Esterase Method
A racemic amino acid ester can be enzymatically resolved by the action of a protease or (in selected cases) an esterase or a lipase. Remarkably, the first resolution of this type using a crude porcine pancreatic extract was reported in 1905 [109]! The catalytic activity of a protease on a carboxylic ester bond has frequently been denoted as ‘esterase activity’, although the mechanism of action involved does not differ in principle from that of an amide hydrolysis. Bearing in mind the greater stability of an amide bond as compared to that of an ester, it is reasonable that a protease, which is able to cleave a much stronger amide bond, is capable of hydrolyzing a carboxylic ester. Esterases, on the other hand, are generally unable to cleave amide bonds, although they can catalyze their formation via ester aminolysis (Sect. 3.1.3, Scheme 2.1). This does not apply to highly strained β-lactams, which can be hydrolyzed by some esterases (pig liver esterase) or lipases (Scheme 2.11) [110].

Enzymatic resolution of amino acid esters via the esterase method
The amino group of the substrate may be either free or (better) protected by an acyl functionality, preferably an acetyl-, benzoyl-, or the tert-butyloxycarbonyl-(Boc)-group in order to avoid possible side reactions such as ring-closure going in hand with the formation of diketopiperazines. The ester moiety should be a short-chain aliphatic alcohol such as methyl or ethyl to ensure a reasonable reaction rate. When carboxyl ester hydrolases such as lipases are used in this process, it is recommended to use more lipophilic alcohol residues (e.g., n-butyl, n-hexyl, n-octyl) or activated analogs bearing electron-withdrawing substituents, such as chloroethyl [111] or trifluoroethyl [112], to ensure high reaction rates.
Numerous enzymes have been used to hydrolyze N-acyl amino acid esters, the most versatile and thus very popular catalyst being α-chymotrypsin isolated from bovine pancreas (Scheme 2.12) [113–115]. Since it is one of the early examples of a pure enzyme which became available for biotransformations, its mode of action is well understood. A useful and quite reliable model of its active site has been proposed in order to rationalize the stereochemical outcome of resolutions performed with α-chymotrypsin [116, 117]. Alternatively, other proteases, such as subtilisin [118, 119], thermolysin [120], and alkaline protease [121] are also commonly used for the resolution of amino acid esters. Even whole microorganisms such as lyophilized cells of baker’s yeast, possessing unspecific proteases, can be used as biocatalysts for this type of transformation [122].

Carbonic anhydrase – an enzyme termed for its ability to catalyze the hydration of carbon dioxide forming hydrogen carbonate – can also be employed. In contrast to the above-mentioned biocatalytic systems, it exhibits the opposite enantiopreference by hydrolyzing the d-N-acylamino acid esters [123].
An efficient dynamic resolution process for α-amino acid esters using a crude industrial protease preparation from Bacillus licheniformis (‘alcalase’)Footnote 7 has been developed (Scheme 2.13) [126]. The remaining unhydrolyzed d-enantiomer of the substrate was racemized in situ, catalyzed by pyridoxal-5-phosphate (PLP, vitamin B6). Interestingly, this trick has been copied from nature, since pyridoxal-5-phosphate is an essential cofactor for biological amino-group transfer. PLP spontaneously forms a Schiff base with the amino acid ester (but not with the amino acid) which facilitates racemization through reversible proton migration. By using this method, a range of racemic amino acid esters were dynamically resolved in excellent chemical and optical yield. As a more economical substitute for pyridoxal 5-phosphate, its nonphosphorylated analog (pyridoxal) or salicylaldehyde are used in large-scale applications.

Dynamic resolution of amino acid esters
2.1.2.2 Amidase Method
α-Amino acid amides are hydrolyzed enantioselectively by amino acid amidases (occasionally also termed aminopeptidases) obtained from various sources, such as kidney and pancreas [127] and from different microorganisms, in particular Pseudomonas, Aspergillus, or Rhodococcus spp. [128]. For industrial applications, special amidases (e.g., from Mycobacterium neoaurum and Ochrobactrum anthropi) have been developed [129, 130], which, for instance, can be used to resolve α,α-disubstituted α-amino acid amides – which are otherwise not easily hydrolyzed, due to steric hindrance (Scheme 2.14) [105].

Enzymatic resolution of amino acid amides via the amidase method
Again, the l-amino acids thus formed are separated from the unreacted d-amino acid amide by the difference in solubility in various solvents at various pH. After separation, unreacted d-amino acid amides can be recycled via base-catalyzed racemization of the corresponding Schiff-base intermediates in a separate step [131]. Since amino acid amides are less susceptible to spontaneous chemical hydrolysis in the aqueous environment than the corresponding esters, the products which are obtained by this method are often of higher optical purities.
2.1.2.3 Acylase Method
Aminoacylases catalyze the hydrolysis of N-acyl amino acid derivatives, with the acyl groups preferably being acetyl, chloroacetyl, or propionyl. Alternatively, the corresponding N-carbamoyl- and N-formyl derivatives can be used [132]. Enzymes of the amino acylase type have been isolated from hog kidney, and from Aspergillus or Penicillium spp. [133–135]. The versatility of this type of enzyme has been demonstrated by the resolution of racemic N-acetyl tryptophan, -phenylalanine, and -methionine on an industrial scale using column reactors (Scheme 2.15) [136, 137].

Enzymatic resolution of N-acyl amino acids via the acylase method
On a laboratory scale, the readily available amino acylase from hog kidney is recommended [138]. It seems to be extremely substrate-tolerant, allowing variations of the alkyl- or aryl-moiety R within a wide structural range while retaining very high enantioselectivities. The latter enzyme has been frequently used as an aid in natural product synthesis (Scheme 2.17) [139, 140]. Unwanted enantiomers of N-acetyl amino acids can be conveniently racemized ex-situ by heating with acetic anhydride in a separate process. The mechanism of this racemization involves activation of the acid moiety via a mixed anhydride, which undergoes cyclization to form an oxazolinone (azlactone). The latter is subject to racemization through the intermediate achiral enol. Alternatively, the acylase method can be converted into a dynamic process if the nonreacting N-acylamino acid is racemized in-situ via an N-acylamino acid racemase [141, 142]. In contrast to the majority of amino acid racemases, which are cofactor-dependent (usually pyridoxal-5-phosphate), an enzyme which was isolated from Amycolatopsis sp. requires a divalent metal ion such as Co, Mn, or Mg for catalytic activity [143].

Resolution of cyclic N-benzyloxycarbonyl amino acids using enantiocomplementary acylases
Cyclic amino acids, such as proline and piperidine-2-carboxylic acid are valuable building blocks for the synthesis of pharmaceuticals, such as the anti-migraine-agent eleptritan or the anticancer drug incel, respectively. In order to access both enantiomers by choice of the appropriate enzyme, enantiocomplementary acylases from microbial sources were developed using classic enrichment techniques. An l-acylase from Arthrobacter sp. furnishes the free l-amino acid plus the unreacted d-N-acyl-substrate enantiomer, while opposite enantiomers were obtained using a d-specific acylase from Arthrobacter xylosoxidans (Scheme 2.16) [144, 145].
Interestingly, even N-acyl α-aminophosphonic acid derivatives have been resolved using penicillin acylase [146].

Enzymatic resolution of N-acyl amino acids for natural product synthesis
2.1.2.4 Hydantoinase Method
5-Substituted hydantoins may be obtained easily in racemic form from cheap starting materials such as aldehyde, hydrogen cyanide, and ammonium carbonate using the Bücherer–Bergs synthesis [147]. Hydantoinases from different microbial sources catalyze the hydrolytic ring-opening to form the corresponding N-carbamoyl-α-amino acids [148–150]. In nature, many (but not all) of these enzymes are responsible for the cleavage of dihydropyrimidines occurring in pyrimidine catabolism, therefore they are often also called ‘dihydro-pyrimidinases’ (Scheme 2.18) [151, 152].

Enzymatic resolution of hydantoins via the hydantoinase method
In contrast to the above-mentioned amino acid resolution methods involving amino acid esters, -amides, or N-acylamino acids where the natural l-enantiomer is preferably hydrolyzed from a racemic mixture, hydantoinases usually convert the opposite d-enantiomer [153–155], and l-hydantoinases are known to a lesser extent [156–158]. In addition, d-hydantoinases usually possess a broader substrate spectrum than their l-counterparts. The corresponding N-carbamoyl derivatives thus obtained can be transformed into the corresponding amino acids by an N-carbamoyl amino acid amidohydrolase, which is often produced by the same microbial species. Alternatively, N-carbamoyl amino acids can be chemically hydrolyzed by treatment with nitrous acid or by exposure to an acidic pH (<4). One property of 5-substituted hydantoins, which makes them particularly attractive for large-scale resolutions is their ease of racemization. When R contains an aromatic group, the enantiomers of the starting hydantoins are readily equilibrated at slightly alkaline pH (>8), which is facilitated by resonance stabilization of the corresponding enolate. In contrast, aliphatic substituted hydantoins racemize very slowly under the reaction conditions compatible with hydantoinases due to the lack of enolate stabilization. For such substrates the use of hydantoin racemases is required to render a dynamic resolution process, which ensures a theoretical yield of 100% [159, 160].
2.1.2.5 Lactamase Method
Due to their cyclic structure, cyclic amides (γ- and δ-lactams) are chemically considerably more stable and thus cannot be hydrolyzed by conventional proteases. However, they can be resolved using a special group of proteases acting on cyclic amide bonds – lactamases.
The bicyclic γ-lactam shown in Scheme 2.19 is an important starting material for the production of antiviral agents, such as Carbovir and Abacavir. It can be efficiently resolved using enantiocomplementary γ-lactamases from microbial sources: an enzyme from Rhodococcus equi produced the (S)-configurated amino acid (plus enantiomeric non-converted lactam), and another lactamase isolated from Pseudomonas solanacearum acted in an enantiocomplementary fashion by providing the corresponding mirror-image products [161].

Enzymatic resolution of bicyclic β- and γ-lactams via the lactamase method
In contrast to γ- and δ-lactams, highly strained β-lactams are more easily susceptible to enzymatic hydrolysis and thus can be (slowly) hydrolyzed by carboxyl ester hydrolyses, such as esterases [162] and lipases [163] (Scheme 2.20).

Enzymatic hydrolysis of strained β-lactams using lactamases and lipases
The bicyclic lactam shown in Scheme 2.20, which serves as starting material for the synthesis of the antifungal agent (–)-cispentacin, was efficiently resolved using Rhodococcus equi lactamase [164]. Remarkably, related benzo-fused β-lactam analogs of varying ring size were even accepted by lipase B from Candida antarctica [165]. Although the reactions were relatively slow (up to 2 days at 60°C), excellent enantioselectivities were obtained.
2.1.3 Ester Hydrolysis
2.1.3.1 Esterases and Proteases
In contrast to the large number of readily available microbial lipases, less than a dozen of true ‘esterases’ – such as pig and horse liver esterases (PLE [166,Footnote 8 167] and HLE, respectively) – have been used to perform the bulk of the large number of highly selective hydrolyses of carboxylic esters. Thus, the use of a different esterase is not easy in cases where the reaction proceeds with insufficient selectivity with a standard enzyme such as PLE.
An esterase which has been shown to catalyze the hydrolysis of nonnatural esters with exceptionally high selectivities is acetylcholine esterase (ACE). It would certainly be a valuable enzyme to add to the limited number of available esterases but it has a significant disadvantage since it is isolated from Electrophorus electricus – the electric eel. Comparing the natural abundance of this species with the occurrence of horses or pigs, its high price – which is prohibitive for most synthetic applications – is probably justified. Thus, the number of ACE applications is limited [168–171]. Additionally, cholesterol esterase is also of limited use, since it seems to work only on relatively bulky substrates which show some structural similarities to the natural substrates of cholesterol esterase, i.e., steroid esters [61, 172].
To overcome this narrow range of readily available esterases, whole microbial cells are sometimes used to perform the reactions instead of isolated enzyme preparations [173]. Although some highly selective conversions using whole-cell systems have been reported, it is clear that any optimization by controlling the reaction conditions is very complicated when viable whole cells are employed. Furthermore, for most cases the nature of the actual active enzyme system remains unknown.
More recently, novel microbial esterases [174, 175] such as carboxyl-esterase NP [176] have been identified from an extensive screening in search for biocatalysts with high specificities for certain types of substrates. Since they have been made available in generous amounts by genetic engineering [177], they are now being used more widely. Despite numerous efforts directed towards the cloning and overexpression of microbial esterases, the number of synthetically useful enzymes – possessing a relaxed substrate specificity by retaining high enantioselectivity – are limited: many novel esterases showed disappointing selectivities [178, 179].
Fortunately, as mentioned in the foregoing chapter, a large number of proteases can also selectively hydrolyze carboxylic esters and this effectively compensates for the limited number of esterases [180]. The most frequently used members of this group are α-chymotrypsin [181], subtilisin [182] and, to a somewhat lesser extent, trypsin, pepsin [183], papain [184], and penicillin acylase [185, 186]. Since many of the studies on the ester-hydrolysis catalyzed by α-chymotrypsin and subtilisin have been performed together with PLE in the same investigation, representative examples are not singled out in a separate chapter but are incorporated into the following chapter dealing with studies on PLE. A more recently established member of this group is a protease from Aspergillus oryzae which seems to be particularly useful for the selective hydrolysis of bulky esters.
As a rule of thumb, when acting on nonnatural carboxylic esters, most proteases seem to retain a preference for the hydrolysis of that enantiomer which mimics the configuration of an l-amino acid more closely [187].
The structural features of more than 90% of the substrates which have been transformed by esterases and proteases can be reduced to the general formulas given in Scheme 2.21. The following general rules can be applied to the construction of substrates for esterases and proteases:

Types of substrates for esterases and proteases
-
For both esters of the general type I and II, the center of chirality (marked by an asterisk [*]) should be located as close as possible to the site of the reaction (that is, the carbonyl group of the ester) to ensure an optimal chiral recognition. Thus, α-substituted carboxylates and esters of secondary alcohols are usually more selectively hydrolyzed than their β-substituted counterparts and esters of chiral primary alcohols, respectively.
-
Both substituents R1 and R2 can be alkyl or aryl groups, but they should differ in size and polarity to aid the chiral recognition process of the enzyme. They may also be joined together to form cyclic structures.
-
Polar or charged functional groups located at R1 and R2, such as –OH, –COOH, –CONH2, or –NH2, which are heavily hydrated in an aqueous environment should be absent, since esterases (and in particular lipases) do not accept highly polar hydrophilic substrates. If such moieties are required, they should be masked with an appropriate lipophilic protective group.
-
The alcohol moieties R3 of type-I esters should be as short as possible, preferably methyl or ethyl. If neccessary, the reaction rate of ester hydrolysis may be enhanced by linking electron-withdrawing groups to the alcohol moiety to give methoxymethyl, cyanomethyl, or 2-haloethyl esters. In contrast, carboxylates bearing long-chain alcohols are usually hydrolyzed at reduced reaction rates with esterases and proteases.
-
The same considerations are applicable to acylates of type II, where short-chain acetates or propionates are the preferred acyl moieties. Increasing the carbonyl reactivity of the substrate ester by adding electron-withdrawing substituents such as halogen (leading to α-haloacetates) is a frequently used method to enhance the reaction rate in enzyme-catalyzed ester hydrolysis [188].
-
One limitation in substrate construction is common for both types of substrates: the remaining hydrogen atom at the chiral center must not be replaced, since α,α,α-trisubstituted carboxylates and esters of tertiary alcohols are usually too bulky to be accepted by esterases and proteases, although there are some rare exceptions to this rule [189–192]. This limitation turns them into potential protective groups for carboxy- and alcoholic functionalities, such as t-butyl esters and pivalates, in case an enzymatic hydrolysis is not desired. For serine ester hydrolases, the rare ability to hydrolyze bulky esters was attributed to an atypical Gly-Gly-Gly-X-sequence motif (instead of the common Gly-X-motif) in the oxyanion cavity located within the active site, which was found in Candida rugosa and Candida antarctica lipase A [193–195].
-
It is clear that both general substrate types (which themselves would constitute racemic substrates) may be further combined into suitable prochiral or meso-substrates (Scheme 2.21).
2.1.3.2 Pig Liver Esterase and α-Chymotrypsin
Amongst all the esterases, pig liver esterase (PLE) is clearly the champion considering its general versatility. This enzyme is constitutionally complex and consists of at least five so-called isoenzymes, which are associated as trimers of three individual proteins [196]. However, for many applications this crude mixture can be regarded as a single enzyme since the isoenzyme subunits often possess similar (but not identical [197]) stereospecificities [198]. Thus, the selectivity of crude PLE may vary, depending on the source and the pretreatment of the enzyme preparation [199]. The biological role of PLE is the hydrolysis of various esters occurring in the porcine diet, which would explain its exceptionally wide substrate tolerance. For preparative reactions it is not absolutely necessary to use the expensive commercially available enzyme preparation because a crude acetone powder which can easily be prepared from pig liver is a cheap and efficient alternative [200].
It seems to generally apply that the respective esterases from related sources such as liver of chickens, hamsters, guinea pigs, or rats were found to be less versatile when compared to PLE. In certain cases, however, esterases from rabbit [201, 202] and horse liver (HLE) [203, 204] proved to be useful substitutes for PLE.
Mild Hydrolysis. Acetates of primary and secondary alcohols such as cyclopropyl acetate [205] and methyl or ethyl carboxylates (such as the labile cyclopentadiene ester [206]) can be selectively hydrolyzed under mild conditions using PLE, avoiding decomposition reactions which would occur during a chemical hydrolysis under acid or base catalysis (Scheme 2.22). For example, this strategy has been used for the final deprotection of the carboxyl moiety of prostaglandin E1 avoiding the destruction of the delicate molecule [207, 208].

Mild ester hydrolysis by porcine liver esterase
Regio- and Diastereoselective Hydrolysis. Regiospecific hydrolysis of dimethyl malate at the 1-position could be effected with PLE as catalyst (Scheme 2.23) [209]. Similarly, hydrolysis of an exo/endo-mixture of diethyl dicarboxylates with a bicyclo[2.2.1]heptane framework occurred only on the less hindered exo-position [210] leaving the endo-ester untouched, thus allowing a facile separation of the two positional isomers in a diastereomeric mixture.

Regio- and diastereoselective ester hydrolysis by porcine liver esterase
Separation of E/Z -Isomers. With E/Z-diastereotopic diesters bearing an aromatic side chain, PLE selectively hydrolyzed the ester group in the more accessible (E)-trans-position to the phenyl ring, regardless of the p-substituent [211] (Scheme 2.24). In analogy to the hydrolysis of dicarboxylates (Scheme 2.3) the reaction stopped at the (Z)-monoester stage with no diacid being formed. Other hydrolytic enzymes (proteases and lipases) were less selective in this case.

Regioselective hydrolysis of E/Z-diastereotopic diesters by porcine liver esterase
Desymmetrization of Prochiral Diesters. PLE has been used only relatively infrequently for the resolution of racemic esters, while α-chymotrypsin has played a more important role. Instead, the ‘meso-trick’ has been thoroughly used with the former enzyme.
As depicted in Scheme 2.25, α,α-disubstituted malonic diesters can be selectively transformed by PLE or α-chymotrypsin to give the corresponding chiral monoesters [212, 213].

Desymmetrization of prochiral malonates by porcine liver esterase and α-chymotrypsin
These transformations demonstrate an illustrative example for an ‘alternative fit’ of substrates with different steric requirements. While PLE performs a selective hydrolysis of the pro-S ester group on all substrates possessing α-substituents (R) of a smaller size ranging from ethyl through n-butyl to phenyl, an increase of the steric bulkiness of R forces the substrate to enter the enzyme’s active site in an opposite (flipped) orientation. Thus, with the more bulky substituents the pro-R ester is preferentially cleaved.
The synthetic utility of the chiral monoesters was demonstrated by the stereoselective degradation of the carboxyl group in the benzyl derivative using a Curtius rearrangement via an acyl azide intermediate [27] (Scheme 2.26). Finally, hydrolysis of the remaining ester group led to an optically pure α-methyl amino acid as exemplified for (S)-α-methylphenyl alanine. As mentioned in the foregoing chapter, such sterically demanding amino acids normally cannot be obtained by resolution of the appropriate amino acid derivative.

Chemoenzymatic synthesis of α-methyl-l-amino acids
As shown in Scheme 2.27, the prochiral center may be moved from the site of the reaction into the β-position. Thus, chiral recognition by PLE [214–218] and α-chymotrypsin [219–222] is retained during the desymmetrization of prochiral 3-substituted glutaric diesters. Whole cells of Acinetobacter lowfii and Arthrobacter spp. have also been used as a source for esterase activity [223] and, once again, depending on the substitutional pattern on carbon-3, the desymmetrization can lead to both enantiomeric products.

Desymmetrization of prochiral glutarates
Acyclic meso-dicarboxylic esters with a succinic and glutaric acid backbone were also good substrates for PLE [224] and α-chymotrypsin (Scheme 2.28) [225]. Interestingly, an additional hydroxy group in the substrate led to an enhancement of the chiral recognition process in both cases.

Desymmetrization of acyclic meso-dicarboxylates by α-chymotrypsin and porcine liver esterase
The full synthetic potential of the ‘meso-trick’ has been exemplified by the desymmetrization of cyclic diesters possessing various kinds of structural patterns. A striking example of a ‘reversal of best fit’ for cyclic meso-1,2-dicarboxylates caused by variation of the ring size, is shown in Scheme 2.29 [226]: when the rings are small (n = 1, 2), the (S)-carboxyl ester is selectively cleaved, whereas the (R)-counterpart preferentially reacts when the rings are larger (n = 4). The highly flexible cyclopentane derivative of moderate ring size is somewhat in the middle of the range and its chirality is not very well recognized. The fact that the nature of the alcohol moiety of such esters can have a significant impact in both the reaction rate and stereochemical outcome of the hydrolysis was shown by the poor chiral recognition of the corresponding diethyl ester of the cyclohexane derivative, which was slowly hydrolyzed to give the monoethyl ester of poor optical purity [227].

Desymmetrization of cyclic meso-1,2-dicarboxylates by porcine liver esterase
Bulky bicyclic meso-dicarboxylates, which were extensively used as optically pure starting materials for the synthesis of bioactive products, can be accepted by PLE [228]. Some selected examples are shown in Scheme 2.30. During these studies it was shown that the exo-configurated diesters were good substrates, whereas the corresponding more sterically hindered endo-counterparts were less selectively hydrolyzed at a significantly reduced reaction rate. Again, the importance of the appropriate choice of the alcohol moiety is exemplified [216]. Thus, while the short-chain methyl and ethyl esters were hydrolyzed with high selectivities, the propyl ester was not.

Desymmetrization of polycyclic meso-1,2-dicarboxylates by porcine liver esterase
Cyclic meso-diacetates can be hydrolyzed in a similar fashion. As shown in Scheme 2.31, the cyclopentene meso-monoester [229], which constitutes one of the most important chiral synthons for prostaglandins and their derivatives [230], was obtained in an e.e. of 80–86% using crude PLE. In accordance with the above-mentioned hypotheses for the construction of esterase substrates, a significant influence of the acyl moiety of the ester was observed: the optical purity of the monoester gradually declined from 80–86% to 33% as the acyl chain of the starting substrate ester was extended from acetate to butanoate. A detailed study of the stereoselectivity of PLE isoenzymes revealed that isoenzymes PLE-1–3 gave almost identical results as the crude PLE preparation, whereas isoenzymes PLE-4 and PLE-5 showed lower stereoselectivities with a preference for the opposite enantiomer [231].

Desymmetrization of cyclic meso-diacetates by porcine liver esterase and acetylcholine esterase
In order to avoid recrystallization of the optically enriched material (80–86% e.e.), which was obtained from the hydrolysis of meso-1,3-diacetoxy-cyclopentene with crude PLE, to enantiomeric purity, a search was conducted for another esterase, which would hydrolyze this substrate with an even better stereoselectivity. Acetylcholine esterase (ACE) was shown to be the best choice [232]. It hydrolyzed the cyclopentene diester with excellent stereoselectivity but with the opposite stereopreference as with PLE. Similar results were obtained by using lipases from porcine pancreas [233] and Candida antarctica [234]. When structural analogs of larger ring size were subjected to ACE hydrolysis, a dramatic effect on the stereochemical course was observed: while the six-membered meso-diester gave the racemic product, the seven-membered analog led to optically pure monoester of opposite configuration [235].
Cyclic meso-diacetates containing nitrogen functionalities proved to be excellent substrates for PLE (Scheme 2.32). In the benzyl-protected 1,3-imidazolin-2-one system – which serves as a starting material for the synthesis of the vitamin (+)-biotin – the optical yield of PLE-catalyzed hydrolysis of the cis-diacetate [236] was much superior to that of the corresponding cis-dicarboxylate [237].

Desymmetrization of N-containing cyclic meso-diester by porcine liver esterase
Resolution of Racemic Esters. Although PLE-catalyzed resolution of racemic esters have been performed less often as compared to the desymmetrization of prochiral and meso-diesters, it can be a valuable technique.
It has been shown that chirality does not necessarily need to be located on a tetrahedral carbon atom, as in the case of the trans-epoxy dicarboxylates (Scheme 2.33) [238]. For example, the axial chirality of the racemic iron-tricarbonyl complex [239] and of the allenic carboxylic ester shown below [240], was well recognized by PLE.

Resolution of racemic carboxylic esters by porcine liver esterase
Resolution of an N-acetylaminocyclopentene carboxylate shown in Scheme 2.34 was used to access optically pure starting material for the synthesis of carbocyclic nucleoside analogs with promising antiviral activity [241]. The bulky tricyclic monoester was required for natural product synthesis [242].

Resolution of cyclic racemic carboxylic esters by porcine liver esterase
An example demonstrating the high stereospecificity of PLE is the kinetic resolution of the cyclic trans-1,2-diacetate shown in Scheme 2.35 [243]. The (R,R)-diacetate enantiomer (possessing two ester groups showing the matching (R)-configuration) was hydrolyzed from the racemic mixture via the monoester stage to yield the corresponding (R,R)-diol by leaving the (S,S)-diacetate untouched, since it possesses only nonmatching (S)-ester groups. Again, as observed in the desymmetrization of cis-meso-1,2-dicarboxylates, the enantioselectivity strongly depended on the ring size: while the four- and six-membered substrates gave excellent results with opposite enantiopreference, the five-membered substrate analog was not suitable. It should be noted that a desymmetrization of the corresponding cis-meso-1,2-diacetates is impeded a priori by nonenzymic acyl migration which leads to facile racemization of any monoester that is formed.

Resolution of a cyclic trans-1,2-diacetate by porcine liver esterase
Inspired by the broad substrate range of porcine liver esterase, cloning and overexpression of PLE isoenzymes was persued over the past few years in order to overcome imperfect stereoselectivities of crude PLE preparations and to provide a reliable enzyme source [244–246]. In addition, for industrial applications the use of enzymes from animal sources are often undesirable due to the risk of contaminations by viruses and prions and due to the fact that products derived from pigs are considered impure by several world religions.
Analysis of the amino acid sequences of PLE isoenzymes revealed that the remarkably small differences of ca. 20 amino acids are not distributed randomly but are located within distinct conserved areas. Among the different isoenzymes, PLE-1Footnote 9 and an isoenzyme termed A-PLEFootnote 10 [247] were shown to be most useful for stereoselective ester hydrolysis. The latter enzyme, which was expressed at a high level in Pichia pastoris, is remarkably stable and showed perfect enantioselectivity for the resolution of methyl (4E)-5-chloro-2-isopropyl-4-pentenoate, which is a key building block for the synthesis of the renin inhibitor aliskiren, which is used in the treatment of hypertension (Scheme 2.36) [248].

Resolution of α-chiral ester using the isoenzyme A-PLE
2.1.3.3 Microbial Esterases
Complementary to the use of isolated enzymes, whole microbial cells have also been used to catalyze esterolytic reactions. Interesting cases are reported from bacteria, yeasts, and fungi, such as Bacillus subtilis [249], Brevibacterium ammoniagenes [250], Bacillus coagulans [251], Pichia miso [26], and Rhizopus nigricans [252]. Although the reaction control becomes more complex on using whole microbial cells, the selectivities achieved are sometimes impressive. This was shown by the successful resolution of a secondary alcohol via hydrolysis of its acetate by a Bacillus sp. (Scheme 2.37), while other biocatalytic methods to obtain the desired masked chiral hydroxyaldehyde failed [253]. In order to prevent an undesired ester hydrolysis at the terminal carboxyl functionality, it was efficiently blocked as its tert-butyl ester.

Hydrolytic resolution of sec-alcohols using whole microbial cells of Bacillus sp. and baker’s yeast
To overcome the problems of reaction control arising from the metabolism of fermenting microorganisms, resting cells of lyophilized baker’s yeast have been proposed as a source of esterase activity. As shown in Scheme 2.34, 1-alkyn-3-yl acetates [254] were well resolved.
Due to the importance of α-aryl- and α-aryloxy-substituted propionic acids as antiinflammatory agents (e.g., naproxen) and agrochemicals (e.g., the herbicide diclofop), respectively, where the majority of the biological activity resides in only one enantiomer (S for α-aryl- and R for α-aryloxy derivatives,Footnote 11 a convenient way for the separation of their enantiomers was sought by biocatalytic methods. Thus, an extensive screening program carried out by the industry has led to isolation of a novel esterase from Bacillus subtilis [255] (Scheme 2.38). The enzyme, termed ‘carboxyl esterase NP’, accepts a variety of substrates esters, including naproxen [256, 257]. It exhibits highest activity and selectivity when the substrate has an aromatic side chain, as with α-aryl- and α-aryloxypropionic acids. With α-aryl derivatives the corresponding (S)-acids are obtained. α-Aryloxy analogs, however, are resolved with similar high specificities, but products have the opposite configuration (taking into account that a switch in CIP sequence priority occurs when going from aryl to aryloxy). This means that the stereochemical preference of carboxyl esterase NP is reversed when an extra oxygen atom is introduced between the chiral center and the aromatic moiety.

Resolution of α-substituted propionates by carboxylesterase NP
2.1.3.4 Esterase Activity of Proteases
Numerous highly selective ester hydrolyses catalyzed by α-chymotrypsin [115] and papain have featured in excellent reviews [258] and the examples shown above should illustrate the synthetic potential, already fully established in 1976. The major requirements for substrates of type I (see Scheme 2.21) to be selectively hydrolyzed by α-chymotrypsin are the presence of a polar and a hydrophobic group on the α-center (R1 and R2, respectively) to mimic the natural substrates – amino acids.
An interesting example describing the asymmetric synthesis of α-methyl l-amino acids is shown in Scheme 2.39. Hydrolysis of the d-α-nitro esters from the d/l-mixture leads to the formation of the corresponding labile α-nitroacids, which readily decarboxylate to yield secondary nitro compounds. The l-configured enantiomers of the unhydrolyzed esters could be recovered in high optical purity and these were further transformed into α-methyl l-amino acid derivatives [259].

Resolution of α-nitro-α-methyl carboxylates by α-chymotrypsin
Proteases such as α-chymotrypsin, papain, and subtilisin are also useful for regioselective hydrolytic transformations (Scheme 2.40). For example, while regioselective hydrolysis of a dehydroglutamate diester at the 1-position can be achieved using α-chymotrypsin, the 5-ester is attacked by the protease papain [260]. It is noteworthy that papain is one of the few enzymes used for organic synthetic transformations which originate from plant sources (papaya). Other related protease preparations are derived from fig (ficin) and pineapple stem (bromelain) [261].

Regioselective ester hydrolysis by proteases
More recently, two proteases have emerged as highly selective biocatalysts for specific purposes. Penicillin acylase is highly chemoselective for the cleavage of a phenylacetate group in its natural substrate, penicillin G. This enzyme now plays a major role in enzymatic protecting group chemistry [262, 263]. For instance, phenylacetyl groups can be removed in a highly chemoselective fashion in the presence of acetate esters (Scheme 2.41) [264, 265]. Furthermore, it can be used for the resolution of esters of primary [266] and secondary alcohols [267] as long as the acid moiety consists of a phenylacetyl group or a structurally closely related (heterocyclic) analog [268–270]. Some structural similarity of the alcohol moiety with that of the natural substrate penicillin G has been stated as being an advantage.

Chemo- and enantioselective ester hydrolyses catalyzed by penicillin acylase
The use of subtilisin (a protease which is widely used in detergent formulations) as a biocatalyst for the stereospecific hydrolysis of esters is well established [271–273]. Along the same lines, a protease derived from Aspergillus oryzae, which has hitherto mainly been used for cheese processing, has been shown to be particularly useful for the resolution of sterically hindered substrates such as α,α,α-trisubstituted carboxylates [274] (Scheme 2.42). While ‘traditional’ proteases such as subtilisin were plagued by slow reaction rates and low selectivities, the α-trifluoromethyl mandelic ester (which constitutes a precursor of a widely used chiral derivatization agent, ‘Mosher’s acid’ [275]) was successfully resolved by Aspergillus oryzae protease [276].

Resolution of bulky esters by subtilisin and Aspergillus oryzae protease
An elegant example of a protease-catalyzed hydrolysis of a carboxylic ester was demonstrated by the dynamic resolution of the antiinflammatory agent ‘ketorolac’ via hydrolysis of its ethyl ester by an alkali-stable protease derived from Streptomyces griseus (Scheme 2.43) [83]. When the hydrolysis was carried out at pH > 9, base-catalyzed in-situ racemization of the substrate ester provided more of the enzymatically hydrolyzed (S)-enantiomer from its (R)-counterpart, thus raising the theoretical yield of the resolution of this racemate to 100%.

Dynamic resolution with in-situ racemization by protease from Streptomyces griseus
2.1.3.5 Optimization of Selectivity
Many stereoselective enzymatic hydrolyses of nonnatural esters do not show a perfect selectivity, but are often in the range of 50–90% e.e., which corresponds to E values which are considered as ‘moderate’ to ‘good’ (E = 3–20). In order to avoid tedious and material-consuming processes to enhance the optical purity of the product, e.g., by crystallization techniques or via repeated kinetic resolution, several methods exist to improve the selectivity of an enzymatic transformation itself [24, 277]. Most of them can be applied to other types of enzymes.
Since every catalytic system consists of three main components – (bio)catalyst, substrate, and medium – there are three possibilities for the tuning of the selectivity:
-
The ability to choose a different biocatalyst with a superior selectivity towards a given substrate mainly depends on the number of available options within the same enzyme class. This is certainly feasible for proteases and lipases, but not within the relatively small group of esterases. Alternatively, enzyme mutants possessing altered stereospecificities may be constructed.
-
On the other hand, substrate modification (see below) is a widely employed strategy.
-
Altering the properties of the medium – pH, temperature, cosolvents – is possible within certain limits and it can be a powerful technique to enhance enzyme selectivities.
Substrate engineering is one of the most promising techniques, which is applicable to all types of enzymatic transformations. As may be concluded from some of the foregoing examples, the ability of an enzyme to ‘recognize’ the chirality of a given substrate predominantly depends on its steric shape. Electronic effects are involved but usually less important [278–281]. Thus, by variation of the substrate structure (most easily performed by variation of protective groups of different size and/or polarity) an improved fit of the substrate may be achieved, leading to an enhanced selectivity of the enzyme.
Scheme 2.44 shows the optimization of a PLE-catalyzed desymmetrization of dimethyl-3-aminoglutarate esters using the ‘substrate engineering’ approach [216]. By varying the N-protecting group in size and polarity, the optical purity of the monoester could be significantly enhanced as compared to the unprotected original substrate. In addition, a remarkable reversal in stereochemistry was achieved upon the stepwise increase of the size of the protective group X, which provides an elegant method for controlling the configuration of the product.

Optimization of porcine liver esterase-catalyzed hydrolysis by substrate modification
An interesting approach to substrate modification is based on the observation that enzyme selectivities are often enhanced with rigid substrate structures bearing π-electrons (Scheme 2.45). Thus, when a highly flexible aliphatic C-4 within a substrate (Sub) is not well recognized, it can be ‘chemically hidden’ in the corresponding thiophene derivative, which is often transformed more selectively. Then, the enantioenriched (hetero)aromatic product is desulfurized by catalytic hydrogenation using Raney-Ni to yield the saturated desired product in high e.e. [282, 283].

Optimization of selectivity via introduction of a rigid thiophene unit
Medium Engineering. Variation of the aqueous solvent system by the addition of water-miscible organic cosolvents such as methanol, tert-butanol, acetone, dioxane, acetonitrile, dimethyl formamide (DMF), and dimethyl sulfoxide (DMSO) is a promising and quite frequently used method to improve the selectivity of hydrolytic enzymes, in particular with esterases (Scheme 2.45) [284–286]. Depending on the stability of the enzyme, the concentration of cosolvent may vary from ~10 to 50% of the total volume. At higher concentrations, however, enzyme deactivation is unavoidable. Most of these studies have been performed with PLE, for which a significant selectivity enhancement has often been obtained, especially by addition of dimethyl sulfoxide or low-molecular-weight alcohols such as methanol and tert-butanol. However, the price one usually has to pay on addition of water-miscible organic cosolvents to the aqueous reaction medium is a depletion in the reaction rate. The mechanistic action of such modified solvent systems on an enzyme is only poorly understood and predictions on the outcome of such a medium engineering cannot be made; however, the selectivity-enhancing effects are often dramatic.
The selectivity enhancement of PLE-mediated hydrolyses upon the addition of methanol, tert-butanol, and dimethyl sulfoxide to the reaction medium is exemplified in Scheme 2.46. The optical purities of products were in a range of ~20–50% when a pure aqueous buffer system was used, but the addition of methanol and/or DMSO led to a significant improvement [287].

Selectivity enhancement of porcine liver esterase by addition of organic cosolvents
The ‘enantioselective inhibition’ of enzymes by the addition of chiral amines functioning as noncompetitive inhibitors has been reported for lipases [288]. This phenomenon is discussed in Sect. 2.1.3.2. For the selectivity enhancement of dehydrogenase reactions via addition of enzyme inhibitors see Sect. 2.2.3.
Variation of pH. Reactions catalyzed by hydrolases are usually performed in aqueous buffer systems with a pH closer to that of the pH optimum of the enzyme. Because the conformation of an enzyme depends on its ionization state (among others), it is reasonable to assume that a variation of the pH and the type of buffer may influence the selectivity of a given reaction. Such variations are facilitated by the fact that the pH activity profile of the more commonly used hydrolytic enzymes is rather broad and thus allows pH variations while maintaining an adequately high activity. Interestingly, the effect of pH and buffer types on the selectivity of hydrolytic enzymes have been investigated to a lesser extent [289–292].
Variation of Temperature. Enzymes, like other catalysts, generally are considered to exhibit their highest selectivity at low temperatures – as supported by several experimental observations, not only with hydrolases [293] but also with dehydrogenases. It was only recently, however, that a rational understanding of temperature effects on enzyme stereoselectivity was proposed [294, 295]. It is based on the so-called ‘racemic temperature’ (T rac) at which a given enzymatic reaction will proceed without stereochemical discrimination due to the fact that the activation energy is the same for both stereochemical forms participating in the reaction. Hence, there is no difference in free energy (ΔΔG ≠ = 0, see Figs. 1.6 and 1.7).
From the Gibb’s equation given above it follows that only the entropy term ΔΔS ≠ (but not the enthalpy ΔΔH ≠) is influenced by the temperature. Thus, the selectivity of an enzymatic reaction depends on the temperature as follows:
-
At temperatures less than T rac the contribution of entropy is minimal and the stereochemical outcome of the reaction is mainly dominated by the activation enthalpy difference (ΔΔH ≠). The optical purity of product(s) will thus decrease with increasing temperature.
-
On the other hand, at temperatures greater than T rac, the reaction is controlled mainly by the activation entropy difference (ΔΔS ≠) and enthalpy plays a minor role. Therefore, the optical purity of product(s) will increase with increasing tremperature.
However, the major product obtained at a given temperature T > T rac will be the antipode to that at T < T rac, thus a temperature-dependent reversal of stereochemistry is predicted. The validity of this rationale has been proven with the asymmetric reduction of ketones using a dehydrogenase from Thermoanaerobium brockii [296] (Sect. 2.2.2). In contrast to the above-mentioned dehydrogenases from thermophilic organisms, the majority of hydrolases used for biotransformations possess more restricted operational limits with respect to heating, thus narrowing the possibility of a significant selectivity enhancement by variation of temperature of the reaction. From the data available, it can be seen that upon lowering the temperature both an increase [297] or a decrease in the selectivity of hydrolase reactions may be observed [298]. The outcome of the latter experiments depends on whether the reaction has been performed above or below the racemic temperature of the enzyme used. The comparable low upper temperature of about 50°C for the majority of enzymes represents a serious limitation, while impressive effects have been observed upon cooling (–20 to –60°C) [299–301].
More recently, the application of microwave (MW) irradiation for the enhancement of reaction rates of organic-chemical reactions has become fashionable [302].Footnote 12 While conventional heating is due to polychromatic infrared radiation, microwaves are generated in a monochromatic manner. The benefit of MW heating has been proven in numerous types of organic reactions, but the detailed mechanistic principles of molecular action are still poorly understood and there is still much debate about the so-called hot-spot theory [303, 304]. However, for enzyme-catalyzed reactions, MW heating has been shown to be superior to conventional heating by leading to reduced enzyme deactivation and enhanced selectivities [305–307].
Monitoring of the reaction, bearing in mind the underlying kinetics (Sect. 2.1.1) can be very helpful, but only if the reaction is of a type where the extent of conversion of the reaction affects the optical purity of substrate and/or product. Re-esterification of the enantiomerically enriched (but not yet optically pure) product and subjecting the material to a repeated (second) enzymatic hydrolysis is certainly a tedious process but it may be a viable option.
Enzyme Engineering. Molecular biology has enabled the redesign of enzymes possessing improved performance in terms of enhanced stability at extreme temperatures and pH, and at high concentrations of reactants and organic (co)solvents, which is crucial for the construction of process-stable proteins for biotechnological applications. In addition to improved stability, enzymes also can be engineered for enhanced (stereo)selectivities, which represents an equivalent to ligand tuning of homogeneous catalysts. Only some key issues are discussed below since this area requires special expertise in molecular biology – not necessarily a core specialty of synthetic organic chemists. For a deeper understanding, excellent introductory chapters can be found in recent books and reviews [308–312].
There are two distinct philosophies to enzyme engineering:
-
1.
Rational protein design requires detailed knowledge of the three-dimensional structure of an enzyme, preferably from its high-resolution crystal structure or NMR measurements [313]. Alternatively, a computer-generated homology model may help, if the sequence identity is high enough.Footnote 13 In a first step, docking of the substrate to the active site allows to identify amino acid residues, which appear to interact closely with the structural features of the substrate during binding. Steric incompatibilities, such as collisional interference between residues, insufficient substrate binding in large pockets, or nonmatching polarities between hydrogen bonds or salt bridges can be identified and proposals for the replacement of (usually only few) amino acids can be made. The corresponding mutants are generated and tested for their catalytic properties. Sometimes, this rational approach yields impressive results, but (more often) mutant enzymes tell us that the rational analysis of the substrate binding based on a static (crystal) structure is insufficient to explain the dynamic process of protein (re)folding upon formation of the enzyme–substrate complex, which is a prerequisite for the successful dynamics of catalysis [314]. In addition, the tempting notion that mutations close to the active site are always better than distant ones is only a single aspect of a more complex story [315].
-
2.
Directed evolution requires the availability of the gene(s) encoding the enzyme of interest, a suitable (microbial) expression system, a method to create mutant libraries, and an effective selection system – while structural information is irrelevant here. Mutant libraries are widely available, but the crucial aspect lies in the selection problem [316]: in order to identify the one (or the few) mutant protein(s) with improved properties amongst the vast number of variants (typically 104–106), which are (more or less) randomly generated,Footnote 14 an efficient screening method is required to find the tiny needle in the very big haystack. Adequate screening methods usually rely on spectral changes during catalysis. The drawback of this first-generation screening method is the requirement for a chromogenic or fluorogenic ‘reporter group’ in the substrate, which usually consists of a large (hetero)aromatic moiety which needs (at least) 10–14 π-electrons to be ‘visible’ by UV/VIS or fluorescence spectroscopy. Classic reporter groups are (colorless) p-nitrophenyl derivatives, liberating the (yellow) p-nitrophenolate anion upon enzyme catalysis, which in turn can be spectrophotometrically monitored at 410 nm (Scheme 2.47). Unfortunately, the original substrate (e.g., a methyl ester) is modified to a structurally very different p-nitrophenyl substrate ester analog. Since the mutants are screened for optimal activity/selectivity on the surrogate substrate, their performance with the ‘real’ (methyl ester) substrate will be less efficient. In order to create ‘real’ mutant enzymes for ‘real’ substrates, more sophisticated screening methods are recommended based either on a multienzyme assay for acetate (produced during ester hydrolysis, Scheme 2.47) [317], MS analysis of (deuterated) ‘pseudo-enantiomeric’ products, or time-resolved IR thermogravimetry [318–322]. After all, you always get what you screen for.
Scheme 2.47 
Screening for Pseudomonas aeruginosa lipase mutants showing enhanced enantioselectivities using a chromogenic surrogate substrate

Scheme 2.47 shows the use of a surrogate ester substrate bearing a chromogenic (p-nitrophenyl) reporter group for the screening of Pseudomonas aeruginosa lipase mutants possessing improved enantioselectivities for the hydrolysis of an α-chiral long-chain fatty acid [323]. Pure substrate enantiomers were separately tested in 96-well microtiter plates using a plate reader for the readout of enantioselectivity. After four rounds of error-prone polymerase chain reaction (epPCR) at a low mutation rate (ensuring an average amino acid exchange rate of one per enzyme molecule), mutant A (E = 11) was obtained. The latter was improved via saturation mutagenesis through variation of all remaining 19 amino acids at position #155 yielding mutant B (E = 20). Another round of epPCR gave mutant C (E = 25), which could not be further improved. At high mutations rates, epPRC of the wild-type enzyme gave only slightly improved variants D and E (E = 3.0 and 6.5, respectively), which again could not be further improved. However, combinatorial multiple cassette mutagenesis (CMCM) of the wild-type enzyme in the ‘hot region’ of amino acids 160–163 gave mutant G (E = 30). The latter could be further improved by DNA-shuffling with mutant genes D and E to finally yield mutant J, which exhibited a top value of E > 51.
An example for the successful generation of highly enantioselective esterase mutants capable of hydrolyzing acetate esters of tert-alcohols is shown in Scheme 2.48 [324, 325]. In order to improve the modest enantioselectivity of wild-type Bacillus subtilis esterase (E = 5 and 43), a library of ca. 5,000 mutants was constructed, which encompassed 2,800 active variants. Among the latter, the G105A and E188D mutants showed significantly enhanced enantioselectivities for both substrates (E > 100). An E188W/M193C double mutant even showed inverted enantiopreference (E = 64) for the trifluoromethyl substrate [326]. In order to avoid the undesired modification of the substrate by a chromogenic reporter group, a second-generation screening method was employed based on a commercial test kit: Thus, the acetate formed during ester hydrolysis was activated into acetyl-CoA catalyzed by acetyl-CoA synthase (at the expense of ATP). In a subsequent step, the acetate unit is transferred from acetyl-CoA onto oxaloacetate yielding citrate (catalyzed by citrate synthase). The oxaloacetate required for this reaction is formed by oxidation of l-malate (catalyzed by l-malate dehydrogenase) under consumption of NAD+ yielding an equimolar amount of NADH, which can be spectrophotometrically monitored at 340 nm [327].

Enantioselectivities of wild-type Bacillus subtilis esterase and mutants acting on tert-alcohol esters using a multienzyme acetate assay
2.1.3.6 Model Concepts
Useful ‘models’ for the more commonly used enzymes have been developed to avoid trial-and-error modifications of substrate structures and to provide suitable tools for predicting the stereochemical outcome of enzymatic reactions on nonnatural substrates. These models should be able to provide the means to ‘redesign’ a substrate, when initial results are not satisfying with respect to reaction rate and/or selectivity. Since the application of such ‘models’ holds a couple of potential pitfalls, the most important principles underlying their construction are discussed here.
Molecular Modeling. The three-dimensional ‘map’ of the active site of an enzyme can be accurately determined by X-ray crystallography [328–330] or by NMR spectroscopy (provided that the enzyme is not too large). Since the tertiary structure of most enzymes is regarded as being closely related to the preferred form in a dissolved state [331], these methods allow the most accurate description of the structure of the enzyme. However, X-ray data for the prediction of the stereochemical outcome of an enzyme-catalyzed reaction can only represent a static protein structure. On the other hand, the chiral recognition process during formation of the enzyme–substrate complex is a highly complex dynamic process. Thus, any attempt of predicting the selectivity of an enzymatic reaction based on X-ray data is comparable to explaining the complex movements in a somersault from a single photographic snapshot.
Unfortunately, X-ray structures are available only for a few enzymes, such as α-chymotrypsin [116], subtilisin [181], and a number of lipases from Mucor spp. [9], Geotrichum candidum [332], Candida rugosa (formerly cylindracea) [333], Candida antarctica B [334], and Pseudomonas glumae [335] – while for a large number of synthetically useful enzymes such as pig liver esterase, relevant structural data are not available.
If the amino acid sequence of an enzyme is known either entirely or even in part, computer-assisted calculations can provide estimated three-dimensional structures of enzymes [336]. This is done by comparing the known parts of the enzyme in question with other enzymes whose amino acid sequence and three-dimensional structures are already known. Depending on the percentage of the homology, i.e., ‘overlap’, of the amino acid sequences, the results are more or less certain. In general, an overlap of about ~60% is sufficient for good results; less is considered too inaccurate.
Provided that the three-dimensional structure of an enzyme is available, several methods for predicting the selectivity and its stereochemical preference are possible:
-
The enzyme–substrate complex is constructed in its transition state for both enantiomers and the energy value for both (diastereomeric) conformations within the active site of the enzyme are calculated via molecular dynamics (MD). The difference in free energy (ΔΔG ≠) – obtained via force field calculations – yields semiquantitative results for the expected selectivity [337].
-
The difference in steric interactions during a (computer-generated) approach of two substrate enantiomers towards an acyl-enzyme intermediate can be used instead [338].
-
If the transition state is not known with some certainty, the substrate can be electronically fitted into the active site of the enzyme. The orientation of substrate enantiomers with respect to the chemical operator of the enzyme as well as possible substrate movements can be analyzed via MD [339]. This is achieved via (computer-generated) ‘heating’ of the substrate within the enzyme, followed by a slow electronic ‘cooling process’, which allows the substrate enantiomers to settle in their position representing the lowest energy minimum. Because selectivities are determined by differences in free energy of transition states, the first approach leads to the most accurate results.
Substrate Model. If neither X-ray data nor the amino acid sequence are available for an enzyme – which is unfortunately the case for many synthetically useful enzymes – one can proceed as follows: A set of artificial substrates having a broad variety of structures is subjected to an enzymatic reaction. The results, i.e., the reaction rates and enantioselectivities, then allow one to create a general structure of an imagined ‘ideal’ substrate, which an actual substrate structure should simulate as closely as possible to ensure rapid acceptance by the enzyme and a high enantioselection (Fig. 2.10). This idealized substrate structure is then called a ‘substrate model’. Such models are quite popular; inter alia they have been developed for PLE [224] and Candida rugosa lipase [340, 341].

Substrate model for porcine liver esterase
To ensure optimal selectivity by PLE, the α- and β-substituents of methyl carboxylates should be assigned according to their size (L = large, M = medium and S = small) with the preferably-accepted enantiomer being shown in Fig. 2.10.
Active Site Model. Instead of developing an ideal substrate structure one also can try to picture the structure of the (unknown) active site of the enzyme by the method described above. Thus, substrates of varying size and polarity are used as probes to measure the dimensions of the active site. Therefore this approach has been denoted as ‘substrate mapping’ [342, 343]. Such active site models are frequently employed and they usually resemble an arrangement of assumed ‘sites’ or ‘pockets’ which are usually box- or cave-shaped. A relatively reliable active-site model for PLE [344] using cubic-space descriptors was based on the evaluation of the results obtained from over 100 substrates (Fig. 2.11).

Active-site model for porcine liver esterase
The boundaries of the model represent the space available for the accommodation of the substrate. The important binding regions which determine the selectivity of the reaction are two hydrophobic pockets (HL and HS, with L = large and S = small) and two pockets of more polar character (PF and PB, with F = front and B = back). The best fit of a substrate is determined by positioning the ester group to be hydrolyzed close to the hydrolytically active serine residue and then arranging the remaining moieties in the H and P pockets.
Of course these crude models only yield reliable predictions if they are based on a substantial number of test substrates.
2.1.3.7 Lipases
Lipases are enzymes which hydrolyze triglycerides into fatty acids and glycerol [345, 346]. Apart from their biological significance, they play an important role in biotechnology, not only for food and oil processing [347–349] but also for the preparation of chiral intermediates [350, 351]. In fact, about 40% of all biotransformations reported to date have been performed with lipases. Thus, lipases constitute probably the most thoroughly investigated group of enzymes – to date numerous lipases have been cloned and more than a dozen crystal structures are available. Although they can hydrolyze and form carboxylic ester bonds like proteases and esterases, their molecular mechanism is different, which gives rise to some unique properties [352, 353].
The most important difference between lipases and esterases is the physicochemical interaction with their substrates. In contrast to esterases, which show a ‘normal’ Michaelis-Menten activity depending on the substrate concentration [S] (i.e., a higher [S] leads to an increase in activity), lipases display almost no activity as long as the substrate is in a dissolved monomeric state (Fig. 2.12). However, when the substrate concentration is gradually enhanced beyond its solubility limit by forming a second (lipophilic) phase, a sharp increase in lipase activity takes place [354, 355]. The fact that lipases do not hydrolyze substrates under a critical concentration (the ‘critical micellar concentration’, CMC), but display a high activity beyond it, has been called the ‘interfacial activation’ [356].

Esterase and lipase kinetics
The molecular rationale for this phenomenon has been shown to be a rearrangement process within the enzyme [329]. A freely dissolved lipase in the absence of an aqueous/lipid interface resides in its inactive state [Enz], because a part of the enzyme molecule covers the active site. When the enzyme contacts the interface of a biphasic water-oil system, a short α-helix – the ‘lid’ – is folded back. Thus, by opening its active site the lipase is rearranged into its active state [Enz]≠.
Lipase-catalyzed hydrolyses thus should be conducted in a biphasic medium. It is sufficient to employ the substrate alone at elevated concentrations, such that it constitutes the second organic phase, or, alternatively, it may be dissolved in a water-immiscible organic solvent such as hexane, a dialkyl ether, or an aromatic solvent. Due to the presence of an interface, physical parameters influencing the mass-transfer of substrate and product between the aqueous and organic phase such as stirring or shaking speed have a marked influence on the reaction rate of lipases. Triacylglycerols such as triolein or -butyrin are used as standard substrates for the determination of lipase activity, whereas for esterases p-nitrophenyl acetate is the classic standard. For the above-mentioned reasons it is clear that the addition of water-immiscible organic solvents to lipase-catalyzed reactions is a useful technique to improve catalytic activities. In contrast, water-soluble organic cosolvents are more often used in conjunction with esterases, which operate in a ‘true’ solution.
The fact that many lipases have the ability to hydrolyze esters other than glycerides makes them particularly useful for organic synthesis [357]. Furthermore, some lipases are also able to accept thioesters [358, 359]. In contrast to esterases, lipases have been used for the resolution of racemates much more than for effecting the ‘meso-trick’. Since the natural substrates are esters of the chiral alcohol, glycerol, with an achiral acid, it may be expected that lipases are most useful for hydrolyzing esters of chiral alcohols rather than esters of chiral acids. Although this expectation is true for the majority of substrates (see substrate type III, Scheme 2.49), a minor fraction of lipases also display high selectivity through recognizing the chirality of an acid moiety (substrate type IV).

Substrate types for lipases
Some of the general rules for substrate-construction are the same as those for esterase-substrates (Scheme 2.21), such as the preferred close location of the chirality center and the necessity of having a hydrogen atom on the carbon atom bearing the chiral or prochiral center. However, other features are different:
-
The acid moiety R3 of lipase-substrate of type III should be of a straight-chain nature possessing at least three to four carbon units to ensure a high lipophilicity of the substrate. Despite that long-chain fatty acids such as oleates would be advantageous for a fast reaction rate, they do cause operational problems such as a high boiling point of the substrate and they tend to form foams and emulsions during extractive work-up. As a compromise between the two extremes – short chains for ease of handling and long ones for a high reaction rate – n-butanoates are often the esters of choice.
-
Furthermore, the majority of lipases show the same stereochemical preference for esters of secondary alcohols (Scheme 2.49), which is known as the ‘Kazlauskas’ rule’ [341]. Assuming that the Sequence Rule order of substituents R1 and R2 is large > medium, the preferably accepted enantiomer lipase-substrate of type III possesses the (R)-configuration at the alcoholic center. It should be noted that the Kazlauskas’ rule for secondary alcohols (Type III) has an accuracy of ≥90%, whereas the predictability for the corresponding α-chiral acids (Type IV) is less reliable.
-
Many proteases (such as α-chymotrypsin and subtilisin) and pig liver esterase exhibit a stereochemical preference opposite to that of lipases. This is because the catalytic triad of lipases and proteases – where the X-ray structure is known – has been found to be arranged in a mirror-image orientation [360]. Thus, the stereochemical outcome of an asymmetric hydrolysis can often be directed by choosing a hydrolase from a different class [361–364]. Scheme 2.50 depicts the quasi-enantiomeric oxy-anion transition-state intermediates during hydrolysis of a sec-alcohol ester catalyzed by Candida rugosa lipase (PDB entry 1crl) and the protease subtilisin (PDB entry 1sbn). While the nucleophilic Ser-residues approach from the back, both His are located at the inside, with the oxy-anions pointing outside. Both active sites have limited space for one of the large and medium-sized substituents of the sec-alcohol moiety (red), but the mirror-image orientation of the catalytic center favors opposite enantiomers, which is exemplified by the hydrolytic kinetic resolution of an α-chiral indolpropionic ester using the (R)-selective Mucor sp. lipase and the (S)-selective protease α-chymotrypsin (Scheme 2.50) [363]. The activated 2-chloroethyl ester was used to ensure enhanced reaction rates.
Scheme 2.50 
Mirror-image orientation of the catalytic machinery of Candida rugosa lipase and the protease subtilisin and enantiocomplementary ester hydrolysis using Mucor sp. lipase and α-chymotrypsin
-
Substrate-type IV represents the general structure of a smaller number of esters which were hydrolyzed by lipases. When using lipases for type-IV substrates, the alcohol moiety R3 should preferentially consist of a long straight-chain alcohol such as n-butanol. For esters of type IV the stereochemical preference is often (S) (Scheme 2.49) but the predictability is less accurate than with type-III substrates [343].

A large variety of different lipases are produced by bacteria or fungi and are excreted as extracellular enzymes, which makes their large-scale production particularly easy.Footnote 15 The majority of these enzymes are created by the organisms in two isoforms (isoenzymes), usually denoted as A and B. Both are closely related and usually show the same enantiopreference, but slight structural differences do exist, leading to certain differences in enantioselectivity. Crude technical-grade lipase preparations usually contain both isoforms; the only notable exception is Candida antarctica lipase, for which both pure isoforms A and B have been made available through genetic engineering. In contrast to esterases, only a minor fraction of lipases are isolated from mammalian sources such as porcine pancreas. Since some lipases from the same genus (for instance, from Candida or Pseudomonas sp.) are supplied by different commercial sources, one should be aware of differences in selectivity and activity among the different preparations, while these are usually not in the range of orders of magnitude [365]. The actual enzyme content of commercial lipase preparations may vary significantly, from less than 1% up to ~70% – and the selectivity of a lipase preparation from the same microbial source does not necessarily increase with its price! Among the ever increasing number of commercially available lipases only those which have been shown to be of a general applicability are discussed below. For a list of the more commonly used lipases see the Appendix (Chap. 5).
As a rule of thumb, the most widely used lipases may be characterized according to the steric requirements of their preferred substrate esters (Fig. 2.13). Whereas Aspergillus sp. lipases are capable of accepting relatively bulky substrates and therefore exhibit low selectivities on ‘narrow’ ones, Candida sp. lipases are more versatile in this regard. Both the Pseudomonas and Mucor sp. lipases have been found to be highly selective on substrates with limited steric requirements and hence are often unable to accept bulky compounds. Thus, substrates which are recognized with moderate selectivities by a Candida lipase, are usually more selectively hydrolyzed by a Pseudomonas type. Porcine pancreatic lipase represents a crude mixture of different hydrolytic enzymes and is therefore difficult to predict. However, pure PPL prefers slim substrates.

Steric requirements of lipases.
Porcine Pancreatic Lipase
The cheapest and hence one of the most widely used lipases is isolated from porcine pancreas (PPL) [366–368]. The crude preparations contain a significant number of other hydrolases besides the ‘true PPL’. The crude preparation mostly used for biotransformations is called ‘pancreatin’ or ‘steapsin’ and contains less than 5% protein. The main hydrolase impurities are α-chymotrypsin, cholesterol esterase, carboxypeptidase B, phospholipases, and other unknown hydrolases. Phospholipases can usually be neglected as undesired hydrolase impurities, because they prefer negatively charged substrate esters which mimic their natural substrates – phospholipids [369, 370]. On the other hand, α-chymotrypsin and cholesterol esterase can be serious competitors in ester hydrolysis. Both of the latter proteins – and also other unknown hydrolases – can impair the selectivity of a desired PPL-catalyzed ester hydrolysis by exhibiting a reaction of lower selectivity (or even of opposite stereochemistry). Thus, any models for PPL should be applied with great caution [371, 372]. Indeed, in some cases these hydrolase impurities have been shown to be responsible for the highly selective transformation of substrates which were not accepted by purified ‘true PPL’ (the latter being available at a high price in partially purified form). Cholesterol esterase and α-chymotrypsin are likely to act on esters of primary and secondary alcohols, whereas ‘true PPL’ is a highly selective catalyst for esters of primary alcohols. Despite the possible interference of different competing hydrolytic enzymes, numerous highly selective applications have been reported with crude PPL [373–375]. Unless otherwise stated, all of the examples shown below have been performed with steapsin.
Regioselective reactions are particularly important in the synthesis of biologically interesting carbohydrates, where selective protection and deprotection of hydroxyl groups is a central problem. Selective removal of acyl groups of peracylated carbohydrates from the anomeric center [376] or from primary hydroxyl groups [377, 378], leaving the secondary acyl groups intact, can be achieved with hydrolytic enzymes or chemical methods, but the regioselective discrimination between secondary acyl groups is a complicated task [379]. PPL can selectively hydrolyze the butanoate ester on position 2 of the 1,6-anhydro-2,3,4-tri-O-butanoyl-galactopyranose derivative shown in Scheme 2.51 [380]. Only a minor fraction of the 2,4-deacylated product was formed.

Regioselective hydrolysis of carbohydrate esters by porcine panceatic lipase
A simultaneous regio- and enantioselective hydrolysis of dimethyl 2-methylsuccinate has been reported with PPL [381] with a preference for the (S)-ester and with the hydrolysis taking place at position 4 (Scheme 2.52). The residual unhydrolyzed ester was obtained with >95% e.e. but the monoacid formed (73% e.e.) had to be re-esterified and subjected to a second hydrolytic step in order to be obtained in an optically pure form. It is interesting to note that α-chymotrypsin exhibited the same enantio- but the opposite regioselectivity on this substrate, preferably hydrolyzing the ester at position 1 [382].

Regio- and enantioselective hydrolysis of dimethyl α-methylsuccinate
The asymmetric hydrolysis of cyclic meso-diacetates by PPL proved to be complementary to the PLE-catalyzed hydrolysis of the corresponding meso-1,2-dicarboxylates (compare Schemes 2.29 and 2.53). The cyclopentane derivative, which gave low e.e. using the PLE method, was now obtained with 86% e.e. [31, 383]. This selectivity was later improved by substrate modification of the cyclopentane moiety [384], giving access to a number of chiral cyclopentanoid building blocks used for the synthesis of carbacyclic prostaglandin I2 derivatives (therapeutic agents for the treatment of thrombotic diseases).

Asymmetric hydrolysis of cyclic meso-diacetates by porcine pancreatic lipase
Chiral glycerols, optically active C3-synthons, were obtained by asymmetric hydrolysis of prochiral 1,3-propanediol diesters using PPL (Scheme 2.54) [385]. A remarkable influence of a π-system located on substituents at position 2 on the optical purity of the products indicate that the selectivity of an enzyme does not depend on steric factors alone, but also on electronic issues [278, 281, 386]. Note that a rigid (E)-C=C bond or a bulky aromatic system [387] on the 2-substituent led to an enhanced selectivity of the enzyme as compared to the corresponding saturated analogs. When the configuration of the double bond was Z, a reversal in the stereochemical preference took place, associated with an overall loss of selectivity. Additionally, this study shows a positive influence of a biphasic system (using di-iso-propyl ether or toluene [388] as water-immiscible organic cosolvent) on the enantioselectivity of the enzyme.

Desymmetrization of prochiral 1,3-propanediol diesters by porcine pancreatic lipase
The last two entries in Scheme 2.54 represent a ‘chiral glycerol’ derivative where a positive influence of the chain length of the fatty acid moiety on the selectivity of the enzyme was demonstrated [389, 390].
Chiral epoxy alcohols, which are not easily available via the Sharpless procedure [391], were successfully resolved with PPL (Scheme 2.55). Interestingly, the lipase is not deactivated by a possible reaction with the epoxide moiety and it is able to accept a large variety of structures [392, 393]. The significant influence of the nature of the acyl moiety on the selectivity of the resolution – again, the long-chain fatty acid esters gave better results than the corresponding acetate – may be attributed to the presence of different hydrolytic enzymes present in the crude PPL preparation [394, 395]. In particular α-chymotrypsin and cholesterol esterase are known to hydrolyze acetates of alcohols but not their long-chain counterparts. Thus, they are more likely to be competitors of PPL on short-chain acetates. In order to improve the modest enantioselectivity of crude PPL for industrial applications, a pure hydrolase was isolated from crude pancreatin, which gave perfect enantioselectivity (E >100) in the presence of dioxane as cosolvent [396].

Resolution of epoxy esters by porcine pancreatic lipase
During a study on the resolution of the sterically demanding bicyclic acetate shown in Scheme 2.56 [397], which represents an important chiral building block for the synthesis of leukotrienes [398], it was found that crude steapsin is a highly selective catalyst for its resolution. On the other hand, pure PPL and α-chymotrypsin were unable to hydrolyze the substrate. Cholesterol esterase, another known hydrolase impurity in crude steapsin capable of accepting bulky substrates, was able to hydrolyze the ester but with low selectivity. Finally, a novel hydrolase which was isolated from crude PPL proved to be the enzyme responsible for the highly selective transformation.

Resolution of bicyclic acetate by hydrolases present in crude porcine pancreatic lipase
Certain azlactones, such as oxazolin-5-ones, represent derivatives of activated esters and thus can be hydrolyzed by proteases, esterases, and lipases (Scheme 2.57) [399]. The products obtained are N-acyl α-amino acids. When proteases are employed, only products of modest optical purity were obtained due to the fact that the enzymatic reaction rate is in the same order of magnitude as the spontaneous ring opening in the aqueous medium (k spont ≈ k R or k S).

Lipase-catalyzed dynamic resolution of oxazolin-5-ones
On the other hand, lipases were found to be more efficient catalysts [400]. Thus, when the hydrolysis was carried out under carefully controlled reaction conditions at pH 7.6, N-benzoyl amino acids of moderate to excellent optical purities were obtained depending on the substituent on C-4. Whereas PPL led to the formation of l-amino acids, the d-counterparts were obtained with a lipase from Aspergillus niger. Furthermore, the rate of equilibration of the two configurationally unstable substrate antipodes under weakly basic conditions is sufficiently rapid to provide a dynamic resolution via in-situ racemization with a theoretical yield of 100% (k Subrac ≥ k R or k S, respectively), whereas the products are configurationally stable (k Prodrac ≈ 0, compare Fig. 2.3).
Candida sp. Lipases
Several crude lipase preparations are available from the yeasts Candida lipolytica, C. antarctica (CAL), and C. rugosa (CRL, syn. C. cylindracea). The latter enzyme, the three-dimentional structure of which has been resolved by X-ray analysis [333], has been frequently used for the resolution of esters of secondary alcohols [401–406] and, to a lesser extent, for the resolution of α-substituted carboxylates [407, 408]. The CRL preparations from several commercial sources which contain up to 16% of protein [409] differ to some extent in their activity but their selectivity is very similar [410]. As CRL is able to accommodate relatively bulky esters in its active site, it is the lipase of choice for the selective hydrolysis of esters of cyclic secondary alcohols. To illustrate this point, some representative examples are given below.
The racemic cyclohexyl enol ester shown in Scheme 2.58 was enzymatically resolved by CRL to give a ketoester with an (S)-stereocenter on the α-position (77% e.e.) coupled with a diastereoselective protonation of the liberated enol, which led to an (R)-configuration on the newly generated center on the γ-carbon atom. Only a trace of the corresponding (S,S)-diastereomer was formed; the remaining (R)-enol ester was obtained in optically pure form [411]. In accordance with the concept that an acyloxy moiety is hydrolyzed preferentially to a carboxylate by ‘typical’ lipases, no significant hydrolysis on the ethyl carboxylate was observed.

Enzymatic resolution of a cyclic enol ester by Candida rugosa lipase
Racemic 2,3-dihydroxy carboxylates, protected as their respective acetonides, were resolved by CRL [412] by using their lipophilic n-butyl esters (Scheme 2.59). It is particularly noteworthy that the bulky α-methyl derivatives could also be transformed, although compounds of this type are usually not accepted by hydrolases.

Enzymatic resolution of cyclic esters by Candida rugosa lipase
A number of cyclohexane 1,2,3-triols were obtained in optically active form via resolution of their esters using CRL as shown in Scheme 2.59 [413]. To prevent acyl migration which would lead to racemization of the product, two of the hydroxyl groups in the substrate molecule were protected as the corresponding acetal. In this case, a variation of the acyl chain from acetate to butanoate increased the reaction rate, but had no significant effect on the selectivity of the enzyme.
Typical substrates for CRL are esters of cyclic sec-alcohols, which usually give excellent enantioselectivities [397, 414–417]. In contrast, straight-chain substrates are only well resolved when sterically demanding substituents are present (Scheme 2.60) [418]. Esters of prim-alcohols usually yield modest stereoselectivities.

Typical ester substrates for Candida rugosa lipase (reacting enantiomer shown)
In order to provide a general tool which allows to predict the stereochemical outcome of CRL-catalyzed reactions, a substrate model for bicyclic sec-alcohols [54, 340, 419, 420] and an active site model [421] have been developed.
The fact that crude Candida rugosa lipase occasionally exhibits a moderate selectivity particularly on α-substituted carboxylic esters could be attributed to the presence of two isomeric forms of the enzyme present in the crude preparation [422, 423]. Both forms (denoted as fraction A and B), could be separated by Sephadex chromatography and were shown to possess a qualitatively identical (i.e., identical enantiopreference) but quantitatively different stereoselectivity (Scheme 2.61). Thus, racemic α-phenyl propionate was resolved with low selectivity (E = 10) using crude CRL, whereas enzyme-fraction A was highly selective (E > 100). The isomeric lipase fraction B showed almost the same moderate selectivity pattern as did the crude enzyme, although it possessed the same stereochemical preference. Treatment of form B with the surface-active agent deoxycholate and an organic solvent system (ethanol/ether) forced its transformation into the more stable conformer, form A, via an unfolding–refolding rearrangement. This noncovalent modification of the enzyme provided a method for the transformation of CRL of form B into its more stable and more selective isomer.

Selectivity enhancement of Candida rugosa lipase by noncovalent enzyme modification
A related selectivity-enhancing noncovalent modification of Candida rugosa lipase, which does not require tedious protein chromatography and which is therefore applicable to large-scale reactions is based on the treatment of crude CRL with 50% aqueous iso-propanol, which led (after simple centrifugation and dialysis) to a modified lipase preparation, which was not only more active, but also showed considerably enhanced enantioselectivity [424].
The most versatile ‘champion’ lipase for preparative biotransformations is obtained from the basidomycetous yeast Candida antarctica (CAL) [425]. As indicated by its name, this yeast was isolated in Antarctica with the aim of finding enzymes with extreme properties to be used in detergent formulations. Like others, the organism produces two isoenzymes A and B, which differ to a significant extent [426]: whereas lipase A (CALA) is Ca2+-dependent and more thermostable, the B-component is less thermotolerant and metal-independent. More important for preparative applications, the substrate-specificity varies a great deal, as the A-lipase is highly active in a nonspecific manner on triglycerides, showing a preference for the sn-2 ester group [427] and is not very useful for simple nonnatural esters. On the contrary, the B-component (CALB) is very active on a broad range of nonnatural esters. Both isoenzymes have been made available in pure form through cloning and overexpression in Aspergillus oryzae as the host organism [428] and various preparations of this enzyme are produced by NOVO (DK) in bulk quantities [429]. For the preparative applications discussed below, the B-component has been used more often.
CALB is an exceptionally robust protein which is deactivated only at 50–60°C,Footnote 16 and thus also shows increased resistance towards organic solvents. In contrast to many other lipases, the enzyme appears to be rather ‘rigid’ and does not show a pronounced effect of interfacial activation [430], which makes it an intermediate between an esterase and a lipase. This latter property is probably the reason why its selectivity could be predicted through computer modeling to a fair extent [431], and for the majority of substrates the Kazlauskas’ rule (Scheme 2.49) can be applied. In line with these properties of CALB, selectivity-enhancement by addition of water-miscible organic cosolvents such as t-butanol or acetone is possible – a technique which is rather common for esterases. All of these properties make CALB the most widely used lipase both in the hydrolysis [432–437] and synthesis of esters (Sect. 3.1.1).
A representative selection of ester substrates, which have been hydrolyzed in a highly selective fashion is depicted in Scheme 2.62 [234, 438–441]. The wide substrate tolerance of this enzyme is demonstrated by a variety of carboxyl esters bearing a chiral center in the alcohol- or the acid-moiety. In addition, desymmetrization of meso-forms was also achieved. In general, good substrates for CALB are somewhat smaller than those for Candida rugosa lipase and typically comprise (acetate or butyrate) esters of sec-alcohols in the (ω-1)- or (ω-2)-position with a straight-chain or monocyclic framework.

Typical ester substrates for Candida antarctica lipase (reacting enantiomer shown)
Pseudomonas sp. Lipases
Bacterial lipases isolated from Pseudomonas fluorescens, P. aeruginosa, P. cepacia, and P. glumae are highly selective catalysts [442].Footnote 17 The structures of both of the latter enzymes were elucidated by X-ray analysis [443, 444]. They seem to possess a ‘narrower’ active site than CRL, since they are often unable to accommodate bulky substrates, but they can be extremely selective on ‘slim’ counterparts [445–449]. Like the majority of the microbial lipases, the commercially available crude Pseudomonas sp. lipase preparations (PSL) all possess a stereochemical preference for the hydrolysis of the (R)-esters of secondary alcohols, but the selectivity among the different preparations may differ to some extent [450]. Various active-site models for PSL have been proposed [163, 451–453].
The exceptionally high selectivity of PSL on ‘narrow’ open-chain esters is demonstrated by the following examples (Scheme 2.63).

Desymmetrization of esters having a remote prochiral center by Pseudomonas sp. lipase
The desymmetrization of some prochiral dithioacetal esters possessing up to five bonds between the prochiral center and the ester carbonyl – the site of reaction – proceeded with high selectivity using PSL [454]. This example of a highly selective chiral recognition of a ‘remote’ chiral/prochiral center is not unusual amongst hydrolytic enzymes [455–457].
Chirality need not reside on a sp3 carbon atom to be recognized by PSL but can be located on a sulfur atom (Scheme 2.64). Thus, optically pure aryl sulfoxides were obtained by lipase-catalyzed resolution of methyl sulfinyl acetates [458] in a biphasic medium containing toluene. The latter compounds are important starting materials for the synthesis of chiral allylic alcohols via the ‘SPAC’ reaction.

Resolution of sulfoxide esters by Pseudomonas sp. lipase
Optically active α- and β-hydroxyaldehydes are useful chiral building blocks for the synthesis of bioactive natural products such as grahamimycin A1 [459] and amino sugars [460]. PSL-catalyzed resolution of the corresponding dithioacetal esters gave both enantiomers in excellent optical purity (Scheme 2.65) [461]. A significant selectivity enhancement caused by the bulky sulfur atoms was demonstrated when thioacetals were employed instead of the corresponding O-acetal esters.

Resolution of dithioacetal esters and β-acyloxynitriles by Pseudomonas sp. lipase
The selectivity of PSL-catalyzed hydrolyses may be significantly improved by substrate-modification through variation of the nonchiral acyl moiety (Scheme 2.65) [462]. Whereas alkyl- and chloroalkyl esters gave poor selectivities, again, the introduction of a sulfur atom to furnish the thioacetates proved to be advantageous. Thus, optically active β-hydroxynitriles, precursors of β-hydroxy acids and β-aminoalcohols, were conveniently resolved via the methyl- or phenyl-thioacetate derivatives.
An elegant example for a dynamic resolution of an allylic alcohol via enantioselective ester hydrolysis is depicted in Scheme 2.66 [463]. Thus, Pseudomonas sp. lipase hydrolyzed the acetate ester with high specificity, while the in-situ racemization of the substrate enantiomers was effected by a catalytic amount of PdII leading to the product alcohol in 96% e.e. and 81% yield. However, the lipase has to be chosen with great care, since other hydrolytic enzymes such as acetylcholine esterase and lipases from Penicillium roqueforti, Rhizopus niveus, and Chromobacterium viscosum were incompatible with the metal catalyst.

Dynamic resolution of an allylic alcohol ester using Pseudomonas sp. lipase and PdII catalysis
The typical substrate for Pseudomonas sp. lipases is an (ω-1)-acetate ester bearing a rather small group on one side, whereas remarkable space is available for the large group on the opposite side (Scheme 2.67) [365, 464–467]. Esters of cyclic sec-alcohols are well accepted as long as the steric requirements are not too demanding [468–470]. A special feature of PSL is its high selectivity for racemic or prochiral prim-alcohols [471, 472], where other lipases often show insufficient stereorecognition.

Typical ester substrates for Pseudomonas sp. lipase (reacting enantiomer shown)
Mucor sp. Lipases
Lipases from Mucor species (MSL) [9, 473] such as M. miehei and M. javanicus (also denoted as Rhizomucor) have frequently been used for biotransformations [401, 474]. With respect to the steric requirements of substrates they seem to be related to the Pseudomonas sp. lipases. Similar to Candida and Pseudomonas sp. lipases, the different MSL preparations are related in their hydrolytic specificity [39].
A case where only MSL showed good selectivity is shown in Scheme 2.68 [475]. The desymmetrization of meso-dibutanoates of a tetrahydrofuran-2,5-dimethanol, which constitutes the central subunit of several naturally occurring polyether antibiotics [476] and platelet-activating-factor (PAF) antagonists, was investigated using different lipases. Whereas crude PPL and CRL showed low selectivity, PSL – as may be expected from its more narrow active site – was significantly better. Mucor sp. lipase, however, was completely selective leading to optically pure monoester products. It should be noted that the analogous reaction of the 2,5-unsubstituted acetate (R = H) with PLE at low temperature resulted in the formation of the opposite enantiomer [477].

Desymmetrization of bis(acyloxymethyl)tetrahydrofurans by lipases
The majority of lipase-catalyzed transformations have been performed using PPL, CRL, CAL, PSL, and MSL – the ‘champion lipases’ – and it may be assumed that most of the typical lipase substrates may be resolved by choosing one of this group. However, the largely untapped potential of other lipases which is just being recognized is illustrated by the following examples.
Sterically demanding α-substituted β-acyloxy esters were resolved using an Aspergillus sp. lipase (see Scheme 2.69).Footnote 18 Again, introduction of a thioacetate as the acyl moiety improved the selectivity dramatically [479]. The diastereomeric syn/anti-conformation of the substrate was of critical importance due to the fact that, in contrast to the syn-substrates, only the anti-derivatives were resolved with high selectivities.

Resolution of α-substituted β-acyloxy esters by Aspergillus sp. lipase
Optically pure cyanohydrins are required for the preparation of synthetic pyrethroids, which are used as more environmentally acceptable agricultural pest-control agents in contrast to the classic highly chlorinated phenol derivatives, such as DDT. Cyanohydrins also constitute important intermediates for the synthesis of chiral α-hydroxy acids, α-hydroxyaldehydes [480] and aminoalcohols [481, 482]. They may be obtained via asymmetric hydrolysis of their respective acetates by microbial lipases (Scheme 2.70) [483]. In the ester hydrolysis mode, only the remaining unaccepted substrate enantiomer can be obtained in high optical purity, because the formed cyanohydrin is spontaneously racemized since via its equilibrium with the corresponding aldehyde, liberating hydrocyanic acid at neutral pH values. However, it has recently been shown that the racemization of the cyanohydrin can be avoided when the hydrolysis is carried out at pH 4.5 [484] or in special nonaqueous solvent systems (see Sect. 3.1).

Hydrolysis of cyanohydrin esters using microbial lipases
The resolution of the commercially important esters of (S)-α-cyano-3-phenoxybenzyl alcohol was only moderately efficient using lipases from Candida rugosa, Pseudomonas, and Alcaligenes sp. (Scheme 2.70). The best selectivities were obtained with lipases from Chromobacterium and Arthrobacter sp. [485], respectively.
The epoxy-ester shown in Scheme 2.71 is an important chiral building block for the synthesis of the Ca-channel blocker diltiazem, a potent agent for the treatment of angina pectoris, which is produced at >100 t/year worldwide. Resolution on industrial scale is performed via enantioselective hydrolysis of the corresponding methyl ester using lipases from Rhizomucor miehei (E > 100) or an extracellular lipase from Serratia marcescens (E = 135) in a membrane reactor. The (undesired) carboxylic acid enantiomer undergoes spontaneous decarboxylation yielding p-methoxyphenyl acetaldehyde, which is removed via extraction of the corresponding bisulfite adduct.

Lipase-catalyzed resolution of an epoxy-ester on industrial scale
Another less known lipase is obtained from the mold Geotrichum candidum [486, 487]. The three-dimensional structure of this enzyme has been elucidated by X-ray crystallography [332] showing it to be a serine hydrolase (like MSL), with a catalytic triad consisting of an Glu-His-Ser sequence, in contrast to the more usual Asp-His-Ser counterpart. It has a high sequence homology to CRL (~40%) and shows a similar preference for more bulky substrates like Candida rugosa lipase.
Another extracellular lipase, called ‘cutinase’, is produced by the plant-pathogenic microorganism Fusarium solani pisi for the hydrolysis of cutin – a wax ester which is excreted by plants in order to protect their leaves against microbial attack [488]. The enzyme has been purified to homogeneity [489] and has been made readily available by genetic engineering [490]. Up to now it has not been widely used for biotransformations of nonnatural esters, but it certainly has a potential as a useful pure lipase for complementary purposes [491].
2.1.3.8 Optimization of Selectivity
Most of the general techniques for an enzymatic selectivity enhancement such as adjustment of temperature [492], pH [292], and the kinetic parameters of the reaction which were described for the hydrolysis of esters using esterases and proteases, are applicable to lipase-catalyzed reactions as well. Furthermore, the switch to another enzyme to obtain a better selectivity is relatively easy due to the large number of available lipases. Substrate modification involving not only the chiral alcohol moiety of an ester but also its acyl group [493], as described above, is a valuable technique for the selectivity improvement of lipase-catalyzed transformations. Bearing in mind that lipases are rather ‘soft’ enzymes, they are subject to a strong induced-fit and pronounced interfacial activation (Fig. 2.12). Thus, medium engineering with lipases has been shown to be more effective by applying biphasic systems (aqueous buffer plus a water-immiscible organic solvent) instead of monophasic solvents (buffer plus a water-miscible organic cosolvent).
Some special optimization techniques, which were developed with lipases in particular, are described below.
Enantioselective Inhibition of Lipases. The addition of weak chiral bases such as amines or aminoalcohols has been found to have a strong influence on the selectivity of Candida rugosa [288] and Pseudomonas sp. lipase [494]. The principle of this selectivity enhancement was elaborated as early as 1930! [495]. As shown in Scheme 2.72, the resolution of 2-aryloxypropionates by CRL proceeds with low to moderate selectivity in aqueous buffer alone. The addition of chiral bases of the morphinan-type to the medium led to a significant improvement of about one order of magnitude.

Selectivity enhancement of Candida rugosa lipase by enantioselective inhibition
Kinetic inhibition experiments revealed that the molecular action of the base on the lipase is a noncompetitive inhibition – i.e., the base attaches itself to the lipase at a site other than the active site – which inhibits the transformation of one enantiomer but not that of its mirror image. Moreover, the chirality of the base has only a marginal impact on the selectivity enhancement effect. The general applicability of this method – impeded by the high cost of morphinan alkaloids and their questionable use for large-scale synthesis – has been extended by the use of more simple amines such as N, N-dimethyl-4-methoxyphenethylamine (DMPA) [235].
Alternatively, simple aminoalcohols such as methioninol (MetOH) were shown to have a similar effect on PSL-catalyzed resolutions. As depicted in Scheme 2.73, the chirality of the base had a significant influence on the selectivity of the reaction in this case.

Selectivity enhancement of Pseudomonas sp. lipase by enantioselective inhibition
Chemical Modification of Lipases. The chemical modification of enzymes involving the formation of covalent bonds are a major tool for elucidating the mechanisms of enzymatic catalysis [496–498]. These investigations were aimed primarily at defining those amino acids which participate in catalysis and those which are important in substrate binding. Furthermore, the properties of the enzyme such as solubility, pH optimum, inhibition patterns, and the relative reactivity towards different substrates – the specificity – can be varied by chemical modification. More recently, it was also shown that the enantioselectivity of a lipase may also be improved by covalent modificationFootnote 19 [499–501] (compare Scheme 2.72 and Table 2.2).
The resolution of an α-2,4-dichlorophenoxypropionic acid ester shown in Scheme 2.72 proceeded with very low selectivity when Candida rugosa lipase was used in its native form (E = 1.5). Reductive alkylation of the ε-amino groups of lysine residues using pyridoxal phosphate led to only a small improvement. However, when tyrosine residues were nitrated with tetranitromethane, the lipase proved to be highly specific (E = 33). Reduction of the modified nitro-tyrosine lipase with sodium hydrosulfite (which transforms the nitro into an amino group) slightly enhanced the selectivity even further (E = 37).
2.1.3.9 Hydrolysis of Lactones
Owing to their cyclic structure, lactones are more stable than open-chain esters and thus are generally not hydrolyzed by ‘standard’ ester hydrolases. They can be hydrolyzed by lactonases [502], which are involved in the metabolism of aldoses [503] and the deactivation of bioactive lactones, such as N-acyl homoserine lactone [504]. In the hydrolytic kinetic resolution of lactones, the separation of the formed (water-soluble) hydroxycarboxylic acid from unreacted (lipophilic) lactone is particularly easy via extraction using an aqueous-organic system.
Crude PPL has been shown to hydrolyse γ-substituted α-amino lactones with moderate to good enantioselectivity, however, the identity of the enzyme responsible remains unknown (Scheme 2.74) [505].

Enantioselective hydrolysis of γ-lactones by porcine pancreatic lipase
Well-defined lactonases were identified in bacteria [506–508] and fungi. The most prominent example for the use of a lactonase comprises the resolution of dl-pantolactone, which is required for the synthesis of calcium pantothenate (vitamin B5, Scheme 2.75). The latter is used as vitamin supplement, feed additive, and in cosmetics. An lactonase from Fusarium oxysporium cleaves the d-enantiomer from racemic pantolactone forming d-pantoate in 96% e.e. by leaving the l-enantiomer behind. After simple extractive separation, the unwanted l-lactone is thermally racemized and resubjected to the resolution process. In order to optimize the industrial-scale process, which is performed at ca. 3,500 t/year, the lactonase has been cloned and overexpressed into Aspergillus oryzae [509]. A corresponding enantiocomplementary l-specific lactonase was identified in Agrobacterium tumefaciens [510].

Resolution of pantolactone using a lactonase.
2.1.4 Hydrolysis and Formation of Phosphate Esters
The hydrolysis of phosphate esters can be equally achieved by chemical methods and by phosphatases; the application of enzymes for this reaction is only advantageous if the substrate is susceptible to decomposition. Thus, the enzymatic hydrolysis of phosphates has found only a limited number of applications. The same is true concerning the enantiospecific hydrolyses of racemic phosphates affording a kinetic resolution.
However, the formation of phosphate esters is of importance, particularly when regio- or enantioselective phosphorylation is required. Numerous bioactive agents display their highest activity only when they are transformed into phosphorylated analogs. Furthermore, a number of essential cofactors or cosubstrates for other enzyme-catalyzed reactions of significant synthetic importance involve phosphate esters. For instance, nicotinamide adenine dinucleotide phosphate (NADP+) or glucose-6-phosphate (G6P) is an essential cofactor or cosubstrate, respectively, for some dehydrogenase-catalyzed reactions (Sect. 2.2.1). Dihydroxyacetone phosphate (DHAP) is needed for enzymatic aldol reactions (Sect. 2.4.1), and adenosine triphosphate (ATP) represents the energy-rich phosphate donor for most biological phosphorylation reactions. Glycosyl phosphates are essential for glycosyl transfer reactions catalyzed by carbohydrate phosphorylases (Sect. 2.6.1).
2.1.4.1 Hydrolysis of Phosphate Esters
Chemoselective Hydrolysis of Phosphate Esters. Chemical hydrolysis of polyprenyl pyrophosphates is hampered by side reactions due to the lability of the molecule. Hydrolysis catalyzed by acid phosphatase – an enzyme named because it displays its pH-optimum in the acidic range – readily afforded the corresponding dephosphorylated products in acceptable yields [511].
Similarly, the product from a DHAP-depending aldolase-catalyzed reaction is a chemically labile 2-oxo-1,3,4-triol, which is phosphorylated at position 1. Dephosphorylation under mild conditions, without isolation of the intermediate phosphate species, by using acid phosphatase is a method frequently used to obtain the chiral polyol products [512–515]. As shown in Scheme 2.76, enzymatic dephosphorylation of the aldol product obtained from 5-substituted hexanal derivatives under mild conditions gave the sensitive chiral keto-triol in good yield. In this case the product could be transformed into (+)-exo-brevicomin, the sex pheromone of the pine bark beetle.

Chemoselective enzymatic hydrolysis of phosphate esters
Enantioselective Hydrolysis of Phosphate Esters. In comparison with the hydrolysis of carboxyl esters, enantioselective hydrolyses of phosphate esters have been seldom reported. However, acid phosphatases were applied to the kinetic resolution of serine and threonine via hydrolysis of the corresponding O-phosphate esters (Scheme 2.77) [516]. As for the resolutions of amino acid derivatives using proteases, the natural l-enantiomer was hydrolyzed in the case of threonine O-phosphate, leaving the d-counterpart behind (E > 200). After separation of the d-phosphate from l-threonine, the d-enantiomer could be dephosphorylated using an unspecific alkaline phosphatase – an enzyme with the name derived from having its pH-optimum in the alkaline region. Interestingly, the N151D mutant exhibited an opposite enantiopreference for the d-enantiomer in case of dl-serine-O-phosphate (E = 18) [517].

Resolution of rac-threonine O-phosphate using acid phosphatase
Carbocyclic nucleoside analogs with potential antiviral activity, such as aristeromycin [518] and fluorinated analogs of guanosine [519], were resolved via their 5′-phosphates using a 5′-ribonucleotide phosphohydrolase from snake venom (see Scheme 2.78). Again, the nonaccepted enantiomer, possessing a configuration opposite to that of the natural ribose moiety, was dephosphorylated by unspecific alkaline phosphatase.

Resolution of carbocyclic nucleoside analogs
2.1.4.2 Formation of Phosphate Esters
The introduction of a phosphate moiety into a polyhydroxy compound by classic chemical methods is tedious since it usually requires a number of protection and deprotection steps. Furthermore, oligophosphate esters as undesired byproducts arising from overphosphorylation are a common problem. Employing enzymes for the regioselective formation of phosphate esters can eliminate many of these disadvantages thus making these syntheses more efficient. Additionally, enantioselective transformations are also possible involving the desymmetrization of prochiral or meso-diols or the resolution of racemates.
Phosphate esters are usually synthesized by means of phosphorylating transferases called kinases, which catalyze the transfer of a phosphate moiety (or a di-Footnote 20 or triphosphate moiety) from an energy-rich phosphate donor, such as ATP. Due to the high price of these phosphate donors, they cannot be employed in stoichiometric amounts.Footnote 21 Since ATP cannot be replaced by less expensive man-made chemical equivalents, efficient in-situ regeneration (i.e., recycling) is necessary in order to reduce the cost of enzymatic phosphorylations. Fortunately, ATP recycling has become feasible on a molar scale [520, 521]. On the other hand, reversal of phosphate ester hydrolysis, i.e., the equivalent condensation reaction, has been performed in solvent systems with a reduced water content. Such systems would eliminate the use of expensive or chemically labile phosphate-donors but it is questionable if they will be of general use [522].
ATP Recycling. All phosphorylating enzymes (kinases) require nucleoside triphosphates (in most cases ATP) as a cofactor. In living organisms, these energy-rich phosphates are regenerated by metabolic processes, but for biocatalytic transformations, which are performed in vitro using purified enzymes, this does not occur. The (hypothetical) addition of stoichiometric amounts of these cofactors would not only be undesirable from a commercial standpoint but also for thermodynamic reasons. Quite often the accumulation of the inactive form of the consumed cofactor (most commonly the corresponding diphosphate, ADP) can tip the equilibrium of the reaction in the reverse direction. Thus, cofactors such as ATP are used only in catalytic amounts and are continuously regenerated during the course of the reaction by an auxiliary system which usually consists of a second enzyme and a stoichiometric quantity of an ultimate (cheap) phosphate donor (Scheme 2.79). As the nucleoside triphosphates are intrinsically unstable in solution, the XTP triphosphate species used in phosphorylation reactions is typically recycled ~100 times, with total turnover numbers (TTN) being in the range of about 106–108 mol of product per mol of enzyme.

Enzymatic recycling of ATP from ADP
The following ATP-regenerating systems have been proposed:
-
The use of the phosphoenol pyruvate (PEP)/pyruvate kinase system is probably the most useful method for the regeneration of nucleoside triphosphates [523]. PEP is not only very stable towards spontaneous hydrolysis but it is also a stronger phosphorylating agent than ATP. Furthermore, nucleosides other than adenosine phosphates are also accepted by pyruvate kinase. The drawbacks of this system are the more complex synthesis of PEP [524, 525] and the fact that pyruvate kinase is inhibited by pyruvate at higher concentrations.
-
Acetyl phosphate, which can be synthesized from acetic anhydride and phosphoric acid [526], is a commonly used regeneration system in conjunction with acetate kinase [527]. It is modestly stable in aqueous solution and while its phosphoryl donor potential is lower than that of PEP, it is considerably cheaper. As for pyruvate kinase, acetate kinase also can accept nucleoside phosphates other than adenosine, and it is inhibited by acetate.
-
Promising ATP-recycling methods for large-scale applications use cheap inorganic polyphosphate as phosphate donor and polyphosphate kinase, respectively. Polyphosphate kinase from E. coli accepts also other nucleoside diphosphates and yields up to 40 regeneration cycles [528].
-
An elegant (kinase- and XTP-independent) approach is to use of phosphatases in the reverse direction: Although these enzymes are phosphate ester hydrolases, some of them, such as the enzyme from Shigella flexeri is able to catalyse the backward (phosphorylation) reaction by accepting cheap pyrophosphate as phosphate donor [529].
-
Two further ATP-recycling systems use carbamoyl phosphate and methoxycarbonyl phosphate as nonnatural phosphate donors together with carbamate kinase and acetate kinase, respectively [530, 531]. Both systems lead to the formation of highly unstable products – carbamic acid and methyl carbonate, respectively – which readily decompose forming NH3/CO2 or MeOH/CO2 thereby driving the equilibrium towards completion. Unfortunately, both phosphate donors undergo spontaneous hydrolysis in in aqueous medium, which severely limits their applicability.
-
Regeneration of other nucleoside triphosphates (GTP, UTP, and CTP) or the corresponding 2′-deoxynucleoside triphosphates – which are important substrates for enzyme-catalyzed glycosyl transfer reactions (Sect. 2.6.1) [523, 532, 533] – can be accomplished in the same manner using the acetate or pyruvate kinase systems.
-
A number of reactions which consume ATP generate AMP rather than ADP as a product, only few produce adenosine [534]. ATP may be recycled from AMP using polyphosphate-AMP phosphotransferase and polyphosphate kinase in a tandem-process at the expense of inorganic polyphosphate as phosphate donor for both steps. Alternatively, the combination of adenosine kinase and adenylate kinase were used (Scheme 2.80) [535].
Scheme 2.80 
Enzymatic recycling of ATP from AMP via ADP

A different approach to facilitate ATP-dependent phosphorylation reactions is based on a (chemically) modified nucleoside analog [536] (Scheme 2.81). Carbocyclic surrogates are often good mimics of the corresponding natural nucleoside by displaying a better stability due to their lack of the sensitive hemiaminal bond and are thus widely used in antiviral therapy. Carbocyclic ATP analogs, such as aristeromycin-triphosphate and 6′-fluorocarbocyclic adenosine-triphosphate, were synthesized using adenylate and pyruvate kinase using phosphoenol pyruvate as the ultimate phosphate donor. Both of the carba-ATPs were well accepted by glycerol kinase and hexokinase. The multi-step syntheses required for the preparation of the ATP-mimics may be justified by their enhanced stability and by the possibility of monitoring the phosphorylation reaction using the powerful 19F-NMR in case of the fluoro-ATP analog.

Enzymatic synthesis of carbocyclic ATP-mimics
Regioselective Phosphorylation. The selective phosphorylation of hexoses (and their thia- or aza-analogs) on the primary alcohol moiety located in position 6 can be achieved by a hexokinase (Scheme 2.82) [537, 538]. The other (secondary) hydroxyl groups can be either removed or they can be exchanged for a fluorine atom. Such modified hexose analogs represent potent enzyme inhibitors and are therefore of interest as potential pharmaceuticals or pharmacological probes. The most important compound in Scheme 2.80 is glucose-6-phosphate (G6P; N, O, R = H; M, P, Q = OH; X = O), which serves as a hydride source during the recycling of NAD(P)H when using glucose-6-phosphate dehydrogenase [539, 540] (Sect. 2.2.1).

Phosphorylation of hexose derivatives by hexokinase
Another labile phosphate species, which is needed as a cosubstrate for DHAP-dependent aldolase reactions, is dihydroxyacetone phosphate (Scheme 2.83, also see Sect. 2.4.1). Its chemical synthesis using phosphorus oxychloride is hampered by moderate yields. Enzymatic phosphorylation, however, gives significantly enhanced yields of a product which is sufficiently pure so that it can be used directly in solution without isolation [541, 542].

Phosphorylation of dihydroxyacetone by glycerol kinase
5-Phospho-d-ribosyl-α-1-pyrophosphate (PRPP) serves as a key intermediate in the biosynthesis of purine, pyrimidine, and pyridine nucleotides, such as nucleotide cofactors [ATP, UTP, GTP, CTP, and NAD(P)H]. It was synthesized on a large scale from d-ribose using two consecutive phosphorylating steps [543] (Scheme 2.84). First, ribose-5-phosphate was obtained using ribokinase, subsequently a pyrophosphate moiety was transferred from ATP onto the anomeric center in the α-position by PRPP synthase. In this latter step, AMP (rather than ADP) was generated from ATP, which required adenylate kinase for ATP-recycling. Phosphoenol pyruvate (PEP) served as phosphate donor in all phosphorylation steps. The PRPP thus obtained was subsequently transformed into orotidine monophosphate (O-5-P) via enzymatic linkage of the nucleobase by orotidine-5′-pyrophosphorylase (a transferase) followed by decarboxylation of O-5-P by orotidine-5′-phosphate decarboxylase (a lyase) led to UMP in 73% overall yield.

Phosphorylation of d-ribose and enzymatic synthesis of UMP
Enantioselective Phosphorylation. Glycerol kinase [544] is not only able to accept its natural substrate, glycerol, to form sn-glycerol-3-phosphate [545], or close analogs of it such as dihydroxyacetone (see Scheme 2.83), but it is also able to transform a large variety of prochiral or racemic primary alcohols into chiral phosphates (Scheme 2.85) [546–548]. The latter compounds represent synthetic precursors to phospholipids [549] and their analogs [550].

Enantioselective phosphorylation of glycerol derivatives
As depicted in Scheme 2.85, the glycerol backbone of the substrates may be varied quite widely without affecting the high specificity of glycerol kinase. In resolutions of racemic substrates, both the phosphorylated species produced and the remaining substrate alcohols were obtained with moderate to good optical purities (88 to >94%). Interestingly, the phosphorylation of the aminoalcohol shown in the last entry occurred in an enantio- and chemoselective manner on the more nucleophilic nitrogen atom.
The evaluation of the data obtained from more than 50 substrates permitted the construction of a general model of a substrate that would be accepted by glycerol kinase (Fig. 2.14).

Substrate model for glycerol kinase
2.1.5 Hydrolysis of Epoxides
Chiral epoxides and vicinal diols (employed as their corresponding cyclic sulfate or sulfite esters as reactive intermediates) are extensively employed high-value intermediates for the synthesis of enantiomerically pure bioactive compounds due to their ability to react with a broad variety of nucleophiles [551, 552]. As a consequence, extensive efforts have been devoted to the development of catalytic methods for their production. Although several chemical strategies are known for preparing them from optically active precursors, or via asymmetric syntheses involving desymmetrization or resolution methods [553], none of them is of general applicability and each of them has its merits and limits. Thus, the Sharpless epoxidation gives excellent stereoselectivities and predictable configurations of epoxides, but it is limited to allylic alcohols [554]. On the other hand, the Jacobsen epoxidation is applicable to nonfunctionalized alkenes [555]. The latter gives high selectivities for cis-alkenes, whereas the results obtained with trans- and terminal olefins were less satisfactory. As an alternative, a number of biocatalytic processes for the preparation of enantiopure epoxides via direct or indirect methods are available [556–559]. Among them, microbial epoxidation of alkenes would be particularly attractive by providing a direct access to optically pure epoxides, but this technique requires sophisticated fermentation and process engineering (Sect. 2.3.2.3) [560]. In contrast, the use of hydrolase enzymes for this purpose would be clearly advantageous. An analogous metal-based chemocatalyst for the asymmetric hydrolysis of epoxides is available.Footnote 22
Enzymes catalyzing the regio- and enantiospecific hydrolysis of epoxides – epoxide hydrolases (EH)Footnote 23 [562] – play a key role in the metabolism of xenobiotics. In living cells, aromatics and olefins can be metabolized via two different pathways (Scheme 2.86).

Biodegradation of aromatics
In prokaryotic cells of lower organisms such as bacteria, dioxygenases catalyze the cycloaddition of molecular oxygen onto the C=C double bond forming a dioxetane (Sect. 2.3.2.7). The latter species are reductively cleaved into cis-diols. In eukaryotic cells of higher organisms such as fungi and mammals, enzymatic epoxidation mediated by monooxygenases (Sect. 2.3.2.3) is the major degradation pathway. Due to the electrophilic character of epoxides, they represent powerful alkylating agents which makes them incompatible with living cells: they are toxic, cancerogenic, and teratogenic agents. In order to eliminate them from the cell, epoxide hydrolases catalyze their degradation into biologically more innocuous trans-1,2-diols,Footnote 24 which can be further metabolized or excreted due to their enhanced water solubility. As a consequence, most of the epoxide hydrolase activity found in higher organisms is located in organs, such as the liver, which are responsible for the detoxification of xenobiotics [563, 564].
2.1.5.1 Enzyme Mechanism and Stereochemical Implications
The mechanism of epoxide hydrolase-catalyzed hydrolysis has been elucidated from microsomal epoxide hydrolase (MEH) and bacterial enzymes to involve the trans-antiperiplanar addition of water to epoxides and arene oxides to give vicinal diol products. In general, the reaction occurs with inversion of configuration at the oxirane carbon atom to which the addition takes place and involves neither cofactors nor metal ions [565]. Two types of mechanism are known (Scheme 2.87).

SN2- and borderline-SN2-type mechanism of epoxide hydrolases
S N 2-Type Mechanism. A carboxylate residue – aspartate – performs a nucleophilic attack on the (usually less hindered) epoxide carbon atom by forming a covalent glycol-monoester intermediate [566–568]. The latter species can be regarded as a ‘chemically inverted’ acyl-enzyme intermediate in serine hydrolase reactions (Sect. 2.1.1). In order to avoid the occurrence of a charged oxy-anion, a proton from an adjacent Tyr-residue is simultaneously transferred. In a second step, the ester bond of the glycol monoester intermediate is hydrolyzed by a hydroxyl ion which is provided from water with the aid of a base – histidine [569] – thereby liberating the glycol. Finally, proton-migration from His to Tyr closes the catalytic cycle. This mechanism shows striking similarities to that of haloalkane dehalogenases, where a halide is displaced by an aspartate residue in a similar manner (Sect. 2.7.2) [570, 571]. In addition, a mechanistic relationship with β-glycosidases which act via formation of a covalent glycosyl-enzyme intermediate by retaining the configuration at the anomeric center is obvious (Sect. 2.6.2) [572].
Borderline-S N 2-Type Mechanism. Some enzymes, such as limonene-1,2-epoxide hydrolase, have been shown to operate via a single-step push-pull mechanism [573]. General acid catalysis by a protonated aspartic acid weakens the oxirane to facilitate a simultaneous nucleophilic attack of hydroxyl ion, which is provided by deprotonation of H2O via an aspartate anion. Due to the borderline-SN2-character of this mechanism, the nucleophile preferentially attacks the higher substituted carbon atom bearing the more stabilized δ+-charge. After liberation of the glycol, proton-transfer between both Asp-residues closes the cycle.
The above-mentioned facts have important consequences on the stereochemical course of the kinetic resolution of nonsymmetrically substituted epoxides. In contrast to the majority of kinetic resolutions of esters (e.g., by ester hydrolysis using proteases, esterases, and lipases) where the absolute configuration of the stereogenic center always remains the same throughout the reaction, the enzymatic hydrolysis of epoxides may take place via two different pathways (Scheme 2.88).

Enzymatic hydrolysis of epoxides proceeding with retention or inversion of configuration
-
Attack of the (formal) hydroxide ion on the less hindered (unsubstituted) oxirane carbon atom causes retention of configuration and leads to a hetero-chiral product mixture of enantiomeric diol and nonreacted epoxide.
-
Attack on the stereogenic center leads to inversion and furnishes homochiral products possessing the same sense of chirality.
Although retention of configuration seems to be the more common pathway, inversion has been reported in some cases depending on the substrate structure and the type of enzyme [574, 575]. As a consequence, the absolute configuration of both the product and the substrate from a kinetic resolution of a racemic epoxide has to be determined separately in order to elucidate the stereochemical pathway. As may be deduced from Scheme 2.88, the use of the enantiomeric ratio is only appropriate to describe the enantioselectivity of an epoxide hydrolase as long as its regioselectivity is uniform, i.e., only inversion or retention is taking place, but E-values are inapplicable where mixed pathways, i.e., retention and inversion, are detected [576]. For the solution to this stereochemical problem, various methods were proposed [577].
2.1.5.2 Hepatic Epoxide Hydrolases
To date, two main types of epoxide hydrolases from liver tissue have been characterized, i.e., a microsomal (MEH) and a cytosolic enzyme (CEH), which are different in their substrate specificities. As a rule of thumb, with nonnatural epoxides, MEH has been shown to possess higher activities and selectivities when compared to its cytosolic counterpart.
Although pure MEH can be isolated from the liver of pigs, rabbits, mice, guinea pigs [578], or rats [579], a crude preparation of liver microsomes or even the 9,000 × g supernatant of homogenized liver was employed as a valuable source for EH activity with little difference from that of the purified enzyme being observed [580]. However, other enzyme-catalyzed side-reactions such as ester hydrolysis may occur with crude preparations.
Cyclic cis-meso-epoxides can be asymmetrically hydrolyzed using hepatic epoxide hydrolases to give trans-diols. In this case, the (S)-configurated oxirane carbon atom is preferentially attacked and inverted to yield an (R,R)-diol (Scheme 2.89) [581, 582]. In comparison to the microsomal epoxide hydrolase, cytosolic EH exhibited a lower stereoselectivity.

Desymmetrization of cyclic cis-meso-epoxides by hepatic epoxide hydrolases
Utilizing steroid substrates, MEH was able to hydrolyze not only epoxides, but also the corresponding heteroatom derivatives such as aziridines to form trans-1,2-aminoalcohols albeit at slower rates (Scheme 2.90) [583]. The thiirane, however, was inert towards enzymatic hydrolysis. The enzyme responsible for this activity was assumed to be the same microsomal epoxide hydrolase, but this assumption was not proven.

Enzymatic hydrolysis of steroid epoxides and aziridines by microsomal epoxide hydrolase
Although many studies have been undertaken with hepatic epoxide hydrolases, mainly aimed at the elucidation of detoxification mechanisms, it is unlikely that enzymes from these sources will be widely used as biocatalysts in preparative transformations, since they cannot be produced in reasonable amounts. In contrast, epoxide hydrolases from microbial sources have a great potential because scale-up of their production is considerably easier.
2.1.5.3 Microbial Epoxide Hydrolases
Although it was known for several years that microorganisms possess epoxide hydrolases, they were only scarcely applied to preparative organic transformations [584–588]. Thus, the hydrolysis of epoxides, which was occasionally observed during the microbial epoxidation of alkenes as an undesired side reaction causing product degradation, was usually neglected, and it was only recently that systematic studies were undertaken. It should be emphasized, that although a number of microbial epoxide hydrolases have been purified and characterized [589–593], the majority of preparative-scale reactions were performed by using whole-cell preparations or crude cell-free extracts with an unknown number of epoxide hydrolases being active.
As a result, an impressive amount of knowledge on microbial epoxide hydrolases from various sources – such as bacteria, filamentous fungi, and yeasts – has been gathered and featured in several reviews [594–598]. The data available to date indicate that the enantioselectivities of enzymes from certain microbial sources can be correlated to the substitutional pattern of various types of substrates [599]:
-
Red yeasts (e.g., Rhodotorula or Rhodosporidium sp.) give best enantioselectivities with monosubstituted oxiranes.
-
Fungal cells (e.g., Aspergillus and Beauveria sp.) are best suited for styrene-oxide-type substrates.
-
Bacterial enzymes (in particular derived from Actinomycetes such as Rhodococcus and Nocardia sp.) are the catalysts of choice for more highly substituted 2,2- and 2,3-disubstituted epoxides.
These trends are exemplified as follows (Scheme 2.91). Monosubstituted oxiranes represent highly flexible and rather ‘slim’ molecules, which make chiral recognition a difficult task [600]Footnote 25, [601–603]. Thus, the majority of attempts to achieve highly selective transformations using epoxide hydrolases from bacterial and fungal origin failed for this class of substrates. The only notable exceptions were found among red yeasts, such as Rhodotorula araucarae CBS 6031, Rhodosporidium toruloides CBS 349, Trichosporon sp. UOFS Y-1118, and Rhodotorula glutinis CIMW 147. Regardless of the enzyme source, the enantiopreference for the (R)-enantiomer was predominant and the regioselectivity prevailed for the sterically less hindered carbon atom.

Microbial resolution of monosubstituted epoxides
Styrene oxide-type epoxides have to be regarded as a special group of substrates, as they possess a benzylic carbon atom, which facilitates the formation of a carbenium ion through resonance stabilization by the adjacent aromatic moiety (Scheme 2.92). Thus, attack at this position is electronically facilitated, although it is sterically hindered, and mixed regiochemical pathways (proceeding via retention and inversion) are particularly common within this group of substrates. As a consequence, any E-values reported have to be regarded with great caution, as long as the regioselectivity has not been clearly elucidated. The biocatalysts of choice were found among the fungal epoxide hydrolases, such as Aspergillus niger LCP 521 [604], Beauveria densa CMC 3240 and Beauveria bassiana ATCC 7159. Under certain circumstances, Rhodotorula glutinis CIMW 147 might serve as well [605–607].

Microbial resolution of styrene oxide-type oxiranes
Among the sterically more demanding substrates, 2,2-disubstituted oxiranes were hydrolyzed in virtually complete enantioselectivities using enzymes from bacterial sources (E > 200), in particular Mycobacterium NCIMB 10420, Rhodococcus (NCIMB 1216, DSM 43338, IFO 3730) and closely related Nocardia spp. (Scheme 2.93) [608, 609]. All bacterial epoxide hydrolases exhibited a preference for the (S)-enantiomer. In those cases where the regioselectivity was determined, attack was found to exclusively occur at the unsubstituted oxirane carbon atom.

Enzymatic resolution of 2,2-disubstituted epoxides using microbial epoxide hydrolases
In contrast to 2,2-disubstituted epoxides, mixed regioselectivities are common for 2,3-disubstituted analogs and, as a consequence, E-values are not applicable (Table 2.3, Scheme 2.91) [610]. This is understandable, bearing in mind that the steric requirements at both oxirane positions are similar. Whereas fungal enzymes were less useful, yeast and bacterial epoxide hydrolases proved to be highly selective.
To date, only limited data are available on the enzymatic hydrolysis of trisubstituted epoxides [611–613]. For example, a racemic allylic terpene alcohol containing a cis-trisubstituted epoxide moiety was hydrolyzed by whole cells of Helminthosporium sativum to yield the (S,S)-diol with concomitant oxidation of the terminal alcoholic group (Scheme 2.94). The mirror image (R,S)-epoxide was not transformed. Both optically pure enantiomers were then chemically converted into a juvenile hormone [614].

Microbial resolution of a trisubstituted epoxide
In order to circumvent the disadvantages of kinetic resolution, several protocols were developed towards the enantioconvergent hydrolysis of epoxides, which lead to a single enantiomeric vicinal diol as the sole product from the racemate.
The first technique made use of two fungal epoxide hydrolases possessing matching opposite regio- and enantioselectivity for styrene oxide (Scheme 2.95) [615]. Resting cells of Aspergillus niger hydrolyzed the (R)-epoxide via attack at the less hindered carbon atom to yield the (R)-diol of moderate optical purity. The (S)-epoxide remained unchanged and was recovered in 96% e.e. In contrast, Beauveria bassiana exhibited the opposite enantio- and regioselectivity. It hydrolyzed the (S)-enantiomer but with an unusual inversion of configuration via attack at the more hindered benzylic position. As a result, the (R)-diol was obtained from the (S)-epoxide leaving the (R)-epoxide behind. By combining both microbes in a single reactor, an elegant deracemization technique was accomplished making use of both stereo-complementary pathways. Whereas Aspergillus hydrolyzed the (R)-epoxide with retention, Beauveria converted the (S)-counterpart with inversion. As a result, (R)-phenylethane-1,2-diol was obtained in 89% e.e. and 92% yield.

Microbial resolution and deracemization of styrene oxide
For 2,2-disubstituted oxiranes, this technique was not applicable because an enzyme to perform a highly unfavored nucleophilic attack on a fully substituted carbon atom would be required. In this case, a two-step sequence consisting of combined bio- and chemocatalysis was successful (Scheme 2.96) [616]. In the first step, 2,2-disubstituted oxiranes were kinetically resolved by using bacterial epoxide hydrolases in excellent selectivity. The biohydrolysis proceeds exclusively via attack at the unsubstituted carbon atom with complete retention at the stereogenic center. By contrast, acid-catalyzed hydrolysis of the remaining nonconverted enantiomer under carefully controlled conditions proceeds with inversion. Thus, combination of both steps in a one-pot resolution-inversion sequence yields the corresponding (S)-1,2-diols in virtually enantiopure form and in excellent yields (>90%).

Deracemization of 2,2-disubstituted oxiranes using combined bio- and chemocatalysis
An exceptional case for an enantioconvergent biocatalytic hydrolysis of a (±)-cis-2,3-epoxyalkane is shown in Scheme 2.97 [617]. Based on 18O-labeling experiments, the stereochemical pathway of this reaction was elucidated to proceed via attack of the (formal) hydroxyl ion at the (S)-configured oxirane carbon atom with concomitant inversion of configuration at both enantiomers with opposite regioselectivity. As a result, the (R,R)-diol was formed as the sole product in up to 97% e.e. in almost quantitative yield.

Deracemization of 2,3-disubstituted oxiranes via enantioconvergent enzymatic hydrolysis
As an alternative to the enzymatic hydrolysis of epoxides, nonracemic vicinal diols may be obtained from epoxides via the nucleophilic ring-opening by nitrite catalyzed by halohydrin dehalogenase (a lyase). The corresponding nitrite-monoesters are spontaneously hydrolyzed to yield diols. For the application of this technique see Sect. 2.7.2.
2.1.6 Hydrolysis of Nitriles
Organic compounds containing nitrile groups are found in the environment not only as natural products but also as a result of human activities [618]. Naturally occurring nitriles are synthesized by plants, fungi, bacteria, algae, sponges, and insects, but not by mammals. Cyanide is highly toxic to living cells and interferes with biochemical pathways by three major mechanisms:
-
Tight chelation to di- and trivalent metal atoms in metalloenzymes such as cytochromes
-
Addition onto aldehydes or ketones to form cyanohydrin derivatives
-
Reaction with Schiff-base intermediates (e.g., in transamination reactions) to form stable nitrile derivatives [619]
As shown in Scheme 2.98, natural nitriles include cyanogenic glucosides which are produced by a wide range of plants including major crops such as cassava [620] and sorghum (millet). Plants and microorganisms are also able of producing aliphatic or aromatic nitriles, such as cyanolipids, ricinine, and phenylacetonitrile [621]. These compounds can serve not only as a nitrogen storage, but also as protecting agents against attack by hungry predators, following the general philosophy that if one species has developed a defence mechanism, an invader will try to undermine it with a counterstrategy. As a consequence, it is not unexpected that there are several biochemical pathways for nitrile degradation, such as oxidation and – more important – by hydrolysis. Enzyme-catalyzed hydrolysis of nitriles may occur via two different pathways depending on steric and electronic factors of the substrate structure [622–625] (Scheme 2.99).

Naturally occurring organic nitriles

General pathways of the enzymatic hydrolysis of nitriles
-
Aliphatic nitriles are often metabolized in two stages. First they are converted to the corresponding carboxamide by a nitrile hydratase and then to the carboxylic acid by an amidase enzyme (a protease) [626].
-
Aromatic, heterocyclic, and certain unsaturated aliphatic nitriles are often directly hydrolyzed to the corresponding acids without formation of the intermediate free amide by a so-called nitrilase enzyme. The nitrile hydratase and nitrilase enzyme use distinctively different mechanisms of action.
Nitrile hydratases are known to possess a tightly bound metal atom (Co2+ or Fe3+ [627]) which is required for catalysis [628–631]. Detailed studies revealed that the central metal is octahedrally coordinated to two NH-amide groups from the backbone and three Cys–SH residues, two of which are post-translationally modified into a Cys-sulfenic (–SOH) and a Cys-sulfinic (–SO2H) moiety. This claw-like setting is required to firmly bind the non-heme iron or the non-corrinoid cobalt [632–634] (Scheme 2.100). The remaining axial ligand (X) is either a water molecule (Co2+) [635] or nitric oxide (NO) which binds to Fe3+ [636, 637]. Quite remarkably, the activity of the latter protein is regulated by light: in the dark, the enzyme is inactive, because NO occupies the binding site for the substrate. Upon irradiation with visible light, NO dissociates and activity is switched on.

Coordination sphere of the ferric ion in Brevibacterium sp. nitrile hydratase
Three proposals for the mechanism of metal-depending nitrile hydratases have been suggested, the most plausible assumes direct corrdination of the nitrile to the metal, which (by acting as Lewis-acid) increases the electrophilicity of the carbon atom to allow attack of a water-molecule. The hydroxy-imino-species thus formed tautomerizes to form the carboxamide [638–640].
On the other hand, nitrilases operate by a completely different mechanism (Scheme 2.101). They possess neither coordinated metal atoms, nor cofactors, but act through an essential nucleophilic sulfhydryl residue of a cysteine [641, 642], which is encoded in the nitrilase-sequence motif Glu–Lys–Cys [643]. The mechanism of nitrilases is similar to general base-catalyzed nitrile hydrolysis: Nucleophilic attack by the sulfhydryl residue on the nitrile carbon atom forms an enzyme-bound thioimidate intermediate, which is hydrated to give a tetrahedral intermediate. After the elimination of ammonia, an acyl-enzyme intermediate is formed, which (like in serine hydrolases) is hydrolyzed to yield a carboxylic acid [644].

Mechanism of nitrilases
Enzymatic hydrolysis of nitriles is not only interesting from an academic standpoint, but also from a biotechnological point of view [645–652]. Cyanide represents a widely applicable C1-synthon – a ‘water-stable carbanion’ – but the conditions usually required for the chemical hydrolysis of nitriles present several disadvantages. The reactions usually require either strongly acidic or basic media incompatible with other hydrolyzable groups that may be present. Furthermore, energy consumption is high and unwanted side-products such as cyanide itself or considerable amounts of salts are formed during neutralization. Using enzymatic methods, conducted at physiological pH, most of these drawbacks can be avoided. Additionally, these transformations can often be achieved in a chemo-, regio-, and enantioselective manner. Due to the fact that isolated nitrile-hydrolyzing enzymes are often very sensitive [623], the majority of transformations have been performed using sturdy whole-cell systems.
Another important aspect is the enzymatic hydrolysis of cyanide for the detoxification of industrial effluents [653–656].
2.1.6.1 Chemoselective Hydrolysis of Nitriles
The microorganisms used as sources of nitrile-hydrolyzing enzymes usually belong to the genera Bacillus, Brevibacterium, Micrococcus, Rhodococcus , Pseudomonas, and Bacteridium and they generally show a broad metabolic diversity. Depending on the source of carbon and nitrogen – acting as ‘inducer’ – added to the culture medium, either nitrilases or nitrile hydratases are predominantly produced by the cell. Thus, the desired hydrolytic pathway leading to an amide or a carboxylic acid can often be biologically ‘switched on’ during the growth of the culture by using aliphatic or aromatic nitriles as inducers. In order to avoid substrate inhibition (which is a more common phenomenon with nitrile-hydrolyzing enzymes than product inhibition [657]) the substrates are fed continuously to the culture.
Acrylamide is one of the most important commodity chemicals for the synthesis of various polymers and is produced in an amount of about 200,000 t/year worldwide. In its conventional synthesis, the hydration of acrylonitrile is performed with copper catalysts. However, the preparative procedure for the catalyst, difficulties in its regeneration, problems associated with separation and purification of the formed acrylamide, undesired polymerization and over-hydrolysis are serious drawbacks. Using whole cells of Brevibacterium sp. [658, 659], Pseudomonas chlororapis [660, 661] or Rhodococcus rhodochrous [662] acrylonitrile can be converted into acrylamide in yields of >99%; the formation of byproducts such as acrylic acid is circumvented by blocking of the amidase activity. The scale of this biotransformation exceeds 30,000 t/year (Scheme 2.102).

Chemoselective microbial hydrolysis of acrylonitrile
Aromatic and heteroaromatic nitriles were selectively transformed into the corresponding amides by a Rhodococcus rhodochrous strain [663]; the products accumulated in the culture medium in significant amounts (Scheme 2.103). In contrast to the hydrolysis performed by chemical means, the biochemical transformations were highly selective and occurred without the formation of the corresponding carboxylic acids.

Chemoselective microbial hydrolysis of aromatic and heteroaromatic nitriles yielding carboxamides (product concentrations)
Even more important from a commercial standpoint was that o-, m-, and p-substituted cyanopyridines were accepted as substrates [664, 665] to give picolinamide (a pharmaceutical), nicotinamide (a vitamin), and isonicotinamide (a precursor for isonicotinic acid hydrazide, a tuberculostatic) (Scheme 2.103). Extremely high productivities were obtained due to the fact that the less soluble carboxamide product readily crystallized from the reaction medium in 100% purity. Nicotinamide – enzymatically produced on a scale of 6,000 t/year – is an important nutritional factor and is therefore widely used as a vitamin additive for food and feed supplies [666]. Pyrazinamide is used as a tuberculostatic.
By changing the biochemical pathway through using modified culture conditions, the enzymatic pathways of nitrile hydrolysis are switched and the corresponding carboxylic acids can be obtained (see Scheme 2.104). For instance, p-aminobenzoic acid, a member of the vitamin B group, was obtained from p-aminobenzonitrile using whole cells of Rhodococcus rhodochrous [667]. Similarly, the antimycobacterial agent pyrazinoic acid was prepared in excellent purity from cyanopyrazine [668]. Like nicotinamide, nicotinic acid is a vitamin used as an animal feed supplement, in medicine, and also as a biostimulator for the formation of activated sludge. Microbial hydrolysis of 3-cyanopyridine using Rhodococcus rhodochrous [669] or Nocardia rhodochrous [670] proceeds quantitatively, whereas chemical hydrolysis is hampered by moderate yields.

Chemoselective microbial hydrolysis of aromatic and heteroaromatic nitriles yielding carboxylic acids (product concentrations)
2.1.6.2 Regioselective Hydrolysis of Dinitriles
The selective hydrolysis of one nitrile group out of several in a molecule is generally impossible using traditional chemical catalysis and the reactions usually result in the formation of complex product mixtures. In contrast, whole microbial cells can be very efficient for this purpose [671] (Scheme 2.105).

Regioselective microbial hydrolysis of dinitriles
For instance, 1,3- and 1,4-dicyanobenzenes were selectively hydrolyzed by Rhodococcus rhodochrous to give the corresponding monoacids [672, 673]. In the aliphatic series, tranexamic acid (trans-4-aminomethyl-cyclohexane-1-carboxylic acid), which is a hemostatic agent, is synthesized from trans-1,4-dicyanocyclohexane. Complete regioselective hydrolysis was achieved by using an Acremonium sp. [674]. The outcome of regioselective nitrile hydrolysis is believed to depend on the distance of the nitrile moieties and the presence of other polar groups within the substrate [675, 676].
2.1.6.3 Enantioselective Hydrolysis of Nitriles
While most biocatalytic hydrolyses of nitriles make use of the mild reactions conditions and the chemo- and regioselectivity of nitrile-hydrolyzing enzymes, their stereoselectivity has been investigated more recently. It seems to be a common trend that both nitrilases and nitrile hydratases are often less specific with respect to the chirality of the substrate and that enantiodiscrimination often occurs during the hydrolysis of an intermediate carboxamide an amidase [677] (Scheme 2.99). As a rule, the ‘natural’ l-configured enantiomer is usually converted into the acid leaving the d-counterpart behind. This is not unexpected bearing in mind the high specificities of proteases on α-substituted carboxamides (see Sect. 2.1.2).
Desymmetrization of Prochiral Dinitriles. Prochiral α,α-disubstituted malononitriles can be hydrolyzed in an asymmetric manner by the aid of Rhodococcus rhodochrous [678] (Scheme 2.106). In accordance with the above-mentioned trend, the dinitrile was nonselectively hydrolyzed by the nitrile hydratase in the cells to give the dicarboxamide. In a second consecutive step, the latter was subsequently transformed by the amidase with high selectivity for the pro-(R) amide group to yield the (R)-amide-acid in 96% e.e. and 92% yield. This pathway was confirmed by the fact that identical results were obtained when the dicarboxamide was used as substrate. The nonracemic amide-acid product thus obtained serves as a starting material for the synthesis of nonnatural α-methyl-α-amino acids [679].

Asymmetric microbial hydrolysis of a prochiral dinitrile
In contrast, prochiral glutarodinitriles were stereoselectively hydrolyzed via two steps using whole microbial cells: in a first step, a stereoselective nitrile hydratase furnished the monoamide, which was further hydrolyzed to the corresponding carboxylic acid by an amidase [646, 680]. The cyano-acids thus obtained served as buiding blocks for the synthesis of cholesterol-lowering drugs from the statin family. An impressive example for the development of stereoselective enzymes derived from the metagenome is the discovery of >130 novel nitrilases from biotope-specific environmental DNA libraries [681, 682]. Among these enzymes, 22 nitrilases showed (S)-selectivity for the desymmetrization of the unprotected glutarodinitile (Scheme 2.106, R = H), while one produced the mirror-image (R)-enantiomer in 95–98% e.e. [683].
Kinetic and Dynamic Resolution of rac -Nitriles. α-Hydroxy and α-amino acids can be obtained from the corresponding α-hydroxynitriles (cyanohydrins) and α-aminonitriles [684], which are easily synthesized in racemic form from the corresponding aldehyde precursors by addition of hydrogen cyanide or a Strecker synthesis, respectively (Schemes 2.107 and 2.108). In aqueous systems, cyanohydrins are stereochemically labile and undergo spontaneous racemization via HCN elimination, which furnishes a dynamic resolution process. From aliphatic rac-cyanohydrins, whole cells of Torulopsis candida yielded the corresponding (S)-α-hydroxy acids [685], while (R)-mandelic acid is produced from rac-mandelonitrile on an industrial scale by employing resting cells of Alcaligenes faecalis [686] in almost >90 yield [687, 688].

Stereocomplementary enantioselective hydrolysis of α-hydroxynitriles

Enantioselective hydrolysis of α-aminonitriles
In a related fashion, α-aminonitriles are enzymatically hydrolyzed to yield α-amino acids (Scheme 2.108). Whereas the enantiorecognition in Brevibacterium imperiale or Pseudomonas putida occurs through an amidase [689, 690], Rhodococcus rhodochrous PA-43, Acinetobacter sp. APN, and Aspergillus fumigatus possess enantiocomplementary nitrilases [689, 691, 692].
Many kinetic resolutions of rac-nitriles were performed in search of a method to produce (S)-configurated α-arylpropionic acids, such as ketoprofen, ibuprofen, or naproxen, which are widely used as nonsteroidal antiinflammatory agents. Overall, enantioselectivities depended on the strain used, and whether a nitrilase- or nitrile hydratase-amidase pathway was dominant, which determines the nature of (enantiomeric) products consisting of a mixture of nitrile/carboxylic acid or amide/carboxylic acid, respectively [687, 693–696].
For organisms which express both pathways for nitrile hydrolysis, the stereochemical pathways can be very complex. The latter is illustrated by the microbial resolution of α-aryl-substituted propionitriles using a Rhodococcus butanica strain (Scheme 2.109) [697]. Formation of the ‘natural’ l-acid and the d-amide indicates the presence of an l-specific amidase and a nonspecific nitrile hydratase. However, the occurrence of the (S)-nitrile in case of Ibuprofen (R = i-Bu, e.e. 73%) proves the enantioselectivity of the nitrile hydratase [694]. In a related approach, Brevibacterium imperiale was used for the resolution of structurally related α-aryloxypropionic nitriles [698].

Enantioselective hydrolysis of α-aryl propionitriles
As a substitute for (expensive) commercial enzyme preparations for nitrile-hydrolysis, whole-cell preparations are recommended: Rhodococcus R312 [699]Footnote 26 contains both nitrile-hydrolyzing metabolic pathways, whereas Rhodococcus DSM 11397 and Pseudomonas DSM 11387 contain only nitrile hydratase (no nitrilase) and nitrilase (no nitrile hydratase) activity, respectively [700].
2.2 Reduction Reactions
The enzymes employed in redox reactions are classified into three categories: dehydrogenases, oxygenases and oxidases [701–703]. Among them, alcohol dehydrogenases – also termed carbonyl reductases – have been widely used for the reduction of carbonyl groups (aldehydes, ketones) and ene-reductases are frequently employed for the bioreduction of (electronically activated) carbon-carbon double bonds. In contrast, the asymmetric bioreduction of C=N-bonds is only feasible for special types of substrates, such as in the reductive amination of α-keto acids yielding α-amino acids, but is generally not feasible for nonactivated Schiff-base type imines.
Since reduction usually implies the transformation of a planar sp2-hybridized carbon into a tetrahedral sp3-atom, it goes in hand with the generation of a stereogenic center and represents a desymmetrization reaction (Scheme 2.110). In contrast, the corresponding reverse process (e.g., alcohol oxidation or dehydrogenation) leads to the destruction of a chiral center, which is generally of limited use.

Reduction reactions catalyzed by dehydrogenases
In contrast, oxygenases – named for using molecular oxygen as cosubstrate – have been shown to be particularly useful for oxidation reactions since they catalyze the functionalization of nonactivated C–H or C=C bonds, affording hydroxylation or epoxidation, respectively (Sect. 2.3). Oxidases, which are responsible for the transfer of electrons, have played a minor role in the biotransformation of nonnatural organic compounds, but they are increasingly used more recently (Sect. 2.3.3).
2.2.1 Recycling of Cofactors
The major and crucial distinction between redox enzymes and hydrolases described in the previous chapter, is that the former require redox cofactors, which donate or accept the chemical equivalents for reduction (or oxidation). For the majority of redox enzymes, nicotinamide adenine dinucleotide [NAD(H)] and its respective phosphate [NADP(H)] are required by about 80% and 10% of redox enzymes, respectively. Flavines (FMN, FAD) and pyrroloquinoline quinone (PQQ) are encountered more rarely. The nicotinamide cofactors – resembling ‘Nature’s complex hydrides’ – have two features in common, i.e., they are relatively unstable molecules and they are prohibitively expensive if used in stoichiometric amounts.Footnote 27 In addition, they cannot be replaced by more economical man-made substitutes. Since it is only the oxidation state of the cofactor which changes during the reaction, while the remainder of the complex structure stays intact, it may be regenerated in-situ by using a second concurrent redox-reaction to allow it to re-enter the reaction cycle. Thus, the expensive cofactor is needed only in catalytic amounts, which leads to a drastic reduction in cost. The efficiency of such a recycling process is measured by the number of cycles which can be achieved before a cofactor molecule is finally destroyed. It is expressed as the ‘total turnover number’ (TTN, Sect. 1.4.2) – which is the total number of moles of product formed per mole of cofactor during its entire lifetime. As a rule of thumb, a few thousand cycles (103–104) are sufficient for redox reactions on a laboratory scale, whereas for technical purposes, total turnover numbers of at least 105 are highly desirable. The economic barrier to large-scale reactions posed by cofactor costs has been recognized for many years and a large part of the research effort concerning dehydrogenases has been expended in order to solve the problem of cofactor recycling [521, 704–707].
Cofactor recycling is no problem when whole microbial cells are used as biocatalysts for redox reactions. In this case, inexpensive sources of redox equivalents such as carbohydrates can be used since the microorganism possesses all the enzymes and cofactors which are required for metabolism. The advantages and disadvantages of using whole-cell systems are discussed in Sect. 2.2.3.
2.2.1.1 Recycling of Reduced Nicotinamide Cofactors
The easiest and least efficient method of regenerating NADH from NAD+ is the nonenzymic reduction using a reducing agent such as sodium dithionite (Na2S2O4) [708]. Since the corresponding turnover numbers of this process are very low (TTN ≤ 100), this method has only historical interest. Similarly, electrochemical [709–711] and photochemical regeneration methods [712–715] suffer from insufficient electron transport causing side-reactions and show low to moderate turnover numbers (TTN ≤ 1,000).Footnote 28 On the other hand, enzymic methods for NADH or NADPH recycling have been shown to be much more efficient and nowadays these represent the methods of choice. They may be conveniently subdivided into coupled-substrate and coupled-enzyme types.
Coupled-Substrate Process. Aiming at keeping things as simple as possible, the cofactor required for the transformation of the main substrate is constantly regenerated by addition of a second auxiliary substrate (H-donor) which is transformed by the same enzyme, but into the opposite direction (Scheme 2.111) [716–718]. To shift the equilibrium of the reaction in the desired direction, the donor must be applied in excess [719]. In principle, this approach is applicable to both directions of redox reactions [720]. Although the use of a single enzyme simultaneously catalyzing two reactions appears elegant, some significant disadvantages are often encountered in coupled-substrate cofactor recycling:

Cofactor recycling by the coupled-substrate method
-
The overall efficiency of the process is limited since the enzyme’s activity is distributed between both the substrate and the hydrogen donor/acceptor.
-
Enzyme deactivation is frequently encountered when highly reactive carbonyl species such as acetaldehyde or cyclohexenone are involved in the recycling process.
-
Enzyme inhibition caused by the high concentrations of the auxiliary substrate – cosubstrate inhibition – is common.
-
The product has to be purified from large amounts of auxiliary substrate used in excess.
A special technique avoiding some of these drawbacks makes use of gas-membranes and is discussed in Sect. 3.2.
Coupled-Enzyme Approach. The use of two independent enzymes is more advantageous (Scheme 2.112). In this case, the two parallel redox reactions – i.e., conversion of the main substrate plus cofactor recycling – are catalyzed by two different enzymes [721]. To achieve optimal results, both of the enzymes should have sufficiently different specificities for their respective substrates whereupon the two enzymatic reactions can proceed independently from each other and, as a consequence, both the substrate and the auxiliary substrate do not have to compete for the active site of a single enzyme, but are efficiently converted by the two biocatalysts independently.

Cofactor recycling by the coupled-enzyme method
Several excellent methods, each having its own particular pros and cons, have been developed to regenerate NADH. On the other hand, NADPH may be regenerated sufficiently on a lab scale but a really inexpensive and reliable method is still needed for industrial-scale applications.
The best and most widely used method for recycling NADH uses formate dehydrogenase (FDH), which is obtained from methanol-utilizing microorganisms, to catalyze the oxidation of formate to CO2 (Scheme 2.113) [722, 723]. This method has the advantage that both the auxiliary substrate and the coproduct are innocuous to enzymes and are easily removed from the reaction, which drives the reaction out of equilibrium. FDH is commercially available, readily immobilized and reasonably stable, if protected from autooxidation [724] and trace metals. The only disadvantage of this system is the high cost of FDH and its low specific activity (3 U/mg). However, both drawbacks can be readily circumvented by using an immobilized [725] or membrane-retained FDH system [726]. Overall, the formate/FDH system is the most convenient and most economical method for regenerating NADH, particularly for large-scale and repetitious applications, with TTNs (mol product/mol cofactor) approaching 600,000. The regeneration system based on FDH from Candida boidinii used as a technical-grade biocatalyst is limited by being specific for NADH [727]. This drawback has been circumvented by application of a genetically engineered formate dehydrogenase from Pseudomonas sp., which also accepts NADPH [728–730].

Enzymatic regeneration of reduced nicotinamide cofactors
Another widely used method for recycling NAD(P)H makes use of the oxidation of glucose, catalyzed by glucose dehydrogenase (GDH, Scheme 2.113) [731, 732]. The equilibrium is shifted towards the product because the gluconolactone formed is spontaneously hydrolyzed to give gluconic acid. The glucose dehydrogenase from Bacillus cereus is highly stable [733] and accepts either NAD+ or NADP+ with high specific activity. Like FDH, however, GDH is expensive and product isolation from polar gluconate may complicate the workup. In the absence of purification problems, this method is attractive for laboratory use, and it is certainly a convenient way to regenerate NADPH.
Similarly, glucose-6-phosphate dehydrogenase (G6PDH) catalyzes the oxidation of glucose-6-phosphate (G6P) to 6-phosphoglucolactone, which spontaneously hydrolyzes to the corresponding phosphogluconate (Scheme 2.113). The enzyme from Leuconostoc mesenteroides is inexpensive, stable and accepts both NAD+ and NADP+ [540, 734], whereas yeast-G6PDH accepts only NADP+. A major disadvantage of this system is the high cost of G6P. Thus, if used on a large scale, it must be enzymatically prepared from glucose using hexokinase and this involves the regeneration of ATP using kinases (see pp. 114–116). To avoid problems arising from multienzyme systems, glucose-6-sulfate and G6PDH from Saccharomyces cerevisiae may be used to regenerate NADPH [735]. The sulfate does not act as an acid catalyst for the hydrolysis of NADPH and is more easily prepared than the corresponding phosphate [736]. The G6P/G6PDH system complements glucose/GDH as an excellent method for regenerating NADPH and is a good method for regenerating NADH.
More recently, phosphite dehydrogenase has been shown to offer a promising alternative [737, 738]: The equilibrium is extremely favorable, both phosphite and phosphate are inoccuous to enzymes and act as buffer. The wild-type enzyme from Pseudomonas stutzeri accepts ony NAD+ [739], but thermostable mutants were generated which are also able to reduce NADP+ [740–742].
Ethanol and alcohol dehydrogenase (ADH) have been used extensively to regenerate NADH and NADPH [743, 744]. The low to moderate cost of ADH and the volatility of both ethanol and acetaldehyde make this system attractive for lab-scale reactions. An alcohol dehydrogenase from yeast reduces NAD+, while an ADH from Leuconostoc mesenteroides is used to regenerate NADPH (Scheme 2.113). However, due to the low redox potential, only activated carbonyl substrates such as aldehydes and cyclic ketones are reduced in good yields. With other substrates, the equilibrium must be driven by using ethanol in excess or by removing acetaldehyde. The latter result may be achieved by sweeping with nitrogen [745] or by further oxidizing acetaldehyde to acetate [746], using aldehyde dehydrogenase thereby generating a second equivalent of reduced cofactor. All of these methods, however, give low TTNs or involve complex multi-enzyme systems. Furthermore, even low concentrations of ethanol or acetaldehyde inhibit or deactivate enzymes. Alternatively, a crude cell-free extract from baker’s yeast has been recommended as an (unspecified) enzyme source for NADPH recycling by using glucose as the ultimate reductant [747].
A particularly attractive alternative for the regeneration of NADH makes use of hydrogenase enzymes, so called because they are able to accept molecular hydrogen directly as the hydrogen donor [748, 749]. The latter is strongly reducing, innocuous to enzymes and nicotinamide cofactors, and its consumption leaves no byproduct. For organic chemists, however, this method cannot be generally recommended because hydrogenase is usually isolated from strict anaerobic organisms. Thus, the enzyme is extremely sensitive to oxidation, is not commercially available and requires sophisticated fermentation procedures for its production. Furthermore, some of the organic dyes, which serve as mediators for the transport of redox equivalents from the donor onto the cofactor are relatively toxic (see below).
2.2.1.2 Recycling of Oxidized Nicotinamide Cofactors
For oxidation, reduction reactions can be run in reverse. The best and most widely applied method for the regeneration of nicotinamide cofactors in their oxidized form involves the use of glutamate dehydrogenase (GluDH) which catalyzes the reductive amination of α-ketoglutarate to give l-glutamate (Scheme 2.114) [750, 751]. Both NADH and NADPH are accepted as cofactors. In addition, α-ketoadipate can be used instead of the corresponding glutarate [752], leading to the formation of a high-value byproduct, l-α-aminoadipate.

Enzymatic regeneration of oxidized nicotinamide cofactors
Using pyruvate together with lactate dehydrogenase (LDH) to regenerate NAD+ offers the advantage that LDH is less expensive and exhibits a higher specific activity than GluDH [753]. However, the redox potential is less favorable and LDH does not accept NADP+.
More recently, flavin-dependent nicotinamide oxidases, such as YcnD from Bacillus subtilis [754] or an enzyme from Lactobacillus sanfranciscensis [755] were employed for the oxidation of nicotinamide cofactors at the expense of molecular oxygen producing H2O2 or (more advantageous) H2O via a two- or four-electron transfer reaction, respectively [756–758]. Hydrogen peroxide can be destroyed by addition of catalase and in general, both NADH and NADPH are accepted about equally well.
Acetaldehyde and yeast-ADH have also been used to regenerate NAD+ from NADH [759]. Although reasonable total turnover numbers were achieved (103–104), the above-mentioned disadvantages of enzyme deactivation and self-condensation of acetaldehyde outweigh the merits of the low cost of yeast-ADH and the volatility of the reagents involved.
2.2.2 Reduction of Aldehydes and Ketones Using Isolated Enzymes
A broad range of ketones can be reduced stereoselectively using dehydrogenases to furnish chiral secondary alcohols [760–763]. During the course of the reaction, the enzyme delivers the hydride preferentially either from the si- or the re-side of the ketone to give (R)- or (S)-alcohols, respectively. The stereochemical course of the reaction, which is mainly dependent on the steric requirements of the substrate, may be predicted for most dehydrogenases from a simple model which is generally referred to as ‘Prelog’s rule’ (Scheme 2.115) [764].

Prelog’s rule for the asymmetric reduction of ketones
It is based on the stereochemistry of microbial reductions using Curvularia falcata cells and it states that the dehydrogenase delivers the hydride from the re-face of a prochiral ketone to furnish the corresponding (S)-configured alcohol. The majority of the commercially available dehydrogenases used for the stereospecific reduction of ketones [such as yeast alcohol dehydrogenase (YADH), horse liver alcohol dehydrogenase (HLADH)] and the majority of microorganisms (for instance, baker’s yeast) follow Prelog’s rule [765]. Thermoanaerobium brockii alcohol dehydrogenase (TBADH) also obeys this rule, yielding (S)-alcohols when large ketones are used as substrates, but the stereopreference is reversed with small substrates. Microbial dehydrogenases which lead to the formation of anti-Prelog configurated (R)-alcohols are known to a lesser extent, and even fewer are commercially available, e.g., from Lactobacillus sp. [766–768]. The substrate range of commercially available alcohol dehydrogenases has been mapped including aldehydes, (acyclic, aromatic, and unsaturated) ketones, diketones and various oxo-esters [769].
Yeast ADH has a very narrow substrate specificity and, in general, only accepts aldehydes and methyl ketones [770, 771]. Therefore, cyclic ketones and those bearing carbon chains larger than a methyl group are not accepted as substrates. Thus, YADH is only of limited use for the preparation of chiral secondary alcohols. Similarly, other ADHs from Curvularia falcata [772], Mucor javanicus and Pseudomonas sp. [773] are of limited use as long as they are not commercially available. The most commonly used dehydrogenases are shown in Fig. 2.15, with reference to their preferred size of their substrates [774].

Preferred substrate size for dehydrogenases.
Horse liver ADH is a very universal enzyme with a broad substrate specificity and excellent stereoselectivity. Historically, it is the most widely used dehydrogenase in biotransformations [775, 776].Footnote 29 The three-dimensional structure has been elucidated by X-ray diffraction [778]. Although the primary sequence is quite different, the tertiary structure of HLADH is similar to that of YADH [779]. The most useful applications of HLADH are found in the reduction of medium-ring monocyclic ketones (four- to nine-membered ring systems) and bicyclic ketones [780–782]. Sterically demanding molecules which are larger than decalines are not readily accepted and acyclic ketones are usually reduced with modest enantioselectivities [783, 784].
A considerable number of monocyclic and bicyclic racemic ketones have been resolved using HLADH with fair to excellent specificities [785–787]. Even sterically demanding cage-shaped polycyclic ketones were readily accepted [788, 789] (Scheme 2.116). For instance, rac-2-twistanone was reduced to give the exo-alcohol and the enantiomeric ketone in 90% and 68% e.e., respectively [790]. Also O- and S-heterocyclic ketones were shown to be good substrates (Scheme 2.116) [791–793]. Thus, (±)-bicyclo[4.3.0]nonan-3-ones bearing either an O or S atom in position 8 were resolved with excellent selectivities [784]. Attempted reduction of the corresponding N-heterocyclic ketones led to deactivation of the enzyme via complexation of the essential Zn2+ ion in the active site [794].

Resolution of bi- and polycyclic ketones using horse liver alcohol dehydrogenase (HLADH)
Every kinetic resolution of bi- and polycyclic ketones suffers from one particular drawback because the bridgehead carbon atoms make it impossible to recycle the undesired ‘wrong’ enantiomer via racemization. Hence the desymmetrization of prochiral diketones, making use of the enantioface- or enantiotopos-specificity of HLADH, is of advantage. For instance, both the cis- and trans-forms of the decalinediones shown in Scheme 2.117 were reduced to give (S)-alcohols with excellent optical purity. Similar results were obtained with unsaturated derivatives [743, 795].

Desymmetrization of prochiral diketones using HLADH
The wide substrate tolerance of HLADH encompassing nonnatural compounds is demonstrated by the resolution of organometallic derivatives possessing axial chirality [796]. For instance, the racemic tricarbonyl cyclopentadienyl manganese aldehyde shown in Scheme 2.118 was enantioselectively reduced to give the (R)-alcohol and the residual (S)-aldehyde with excellent optical purities [797].

Enantioselective reduction of an organometallic aldehyde using HLADH
In order to predict the stereochemical outcome of HLADH-catalyzed reductions, a number of models have been developed, each of which having its own merits. The first rationale emerged from the ‘diamond lattice model’ of V. Prelog, which was originally developed for Curvularia falcata [798]. A more recently developed cubic-space descriptor is particularly useful for ketones bearing chirality center(s) remote from the location of the carbonyl group [799]. Alternatively, a quadrant rule may be applied [800].
A useful substrate model based on a flattened cyclohexanone ring is shown in Fig. 2.16 [801]. It shows the Zn2+ in the catalytic site which coordinates to the carbonyl oxygen atom and the nucleophilic attack of the hydride occurring from the bottom face. The preferred orientation of the substrate relative to the chemical operator – the hydride ion – can be estimated by placing the substituents into the ‘allowed’ and ‘forbidden’ zones.

Substrate model for HLADH
YADH and HLADH are less useful for the asymmetric reduction of open-chain ketones, but this gap is efficiently covered by a range of alcohol dehydrogenases from mesophilic bacteria, such as Rhodococcus (ADH-A) and Lactobacillus (LBADH, LKADH), and thermophilic Thermoanaerobacter [802] and Thermoanaerobium (TBADH) strains (Scheme 2.119) [296, 803–806]. Some of these enzymes are remarkably thermostable (up to 85°C) and can tolerate the presence of organic solvents such as isopropanol, which serves as hydrogen-donor for NADP-recycling in a coupled-substrate approach [807–809].

Enantioselective reduction of ketones using Thermoanaerobium brockii alcohol dehydrogenase (TBADH)
Open-chain methyl- and ethyl-ketones are readily reduced by TBADH to furnish the corresponding secondary alcohols, generally with excellent specificities [810]. Similarly, ω-haloalkyl- [732, 811] and methyl- or trifluoromethyl ketones possessing heterocyclic substituents were converted into the corresponding secondary alcohols with excellent optical purities [812, 813]. However, α,β-unsaturated ketones and ketones where both substituents are larger than ethyl are not accepted. In general TBADH obeys Prelog’s rule with ‘normal-sized’ ketones leading to (S)-alcohols, but the stereoselectivity was found to be reversed with small substrates. In order to predict the stereochemical outcome of TBADH reductions, an active site model based on a quadrant rule was proposed [814].
One serious drawback associated with TBADH and Lactobacillus ADHs in large-scale applications is their requirement for NADP(H). In an attempt to circumvent this problem, NAD(H)-vanadate, which is spontaneously formed in solution from catalytic amounts of the cheaper NAD(H) and inorganic vanadate (HVO 2–4 ), was used as a substitute for NADPH [815]. However, undesired redox reactions associated with vanadate limited the practical applicability of this system.
The key to access both stereoisomers of a sec-alcohol via asymmetric carbonyl reduction is the availability of stereocomplementary dehydrogenases. For open-chain ketones bearing a small and large substituent at each side, this is feasible by using an appropriate enzyme showing Prelog or anti-Prelog specificity. Whereas dehydrogenases from Rhodococcus ruber, R. erythropolis , and Candida parapsilosis produce the Prelog enantiomer, Lactobacillus ADHs furnish the corresponding mirror-image product, usually with high stereoselectivity (Scheme 2.120) [816]. In an analogous fashion, α-ketocarboxylic acids were reduced to the corresponding enantiomeric α-hydroxyacids using stereocomplementary lactate dehydrogenases (LDH) [817–820], or hydroxyisocaproate dehydrogenases (HicDHs) [821, 822].

Stereocomplementary bioreduction using a Prelog and anti-Prelog dehydrogenase
Hydroxysteroid dehydrogenases (HSDH) are ideally suited enzymes for the reduction of bulky mono- [823] and bicyclic ketones (Scheme 2.121) [824]. This is not surprising if one thinks of the steric requirements of their natural substrates: steroids [825, 826].

Kinetic resolution of sterically demanding ketones using hydroxysteroid dehydrogenase (HSDH)
For instance, bicyclo[3.2.0]heptan-6-one systems were reduced with HSDH with very low selectivity when substituents in the adjacent 7-position were small (R1, R2 = H). On the other hand, TBADH showed an excellent enantioselectivity with this ‘slim’ ketone. When the steric requirements of the substrate were increased by additional methyl- or chloro-substituents adjacent to the carbonyl group, the situation changed. Then, HSDH became a very specific catalyst and TBADH (or HLADH) proved to be unable to accept the bulky substrates [827, 828]. The switch in the stereochemical preference is not surprising and can be explained by Prelog’s rule: with the unsubstituted ketone, the position 5 is ‘larger’ than position 7. However, when the hydrogen atoms on carbon atom 7 are replaced by sterically demanding chlorine or methyl groups, the situation is reversed.
The majority of synthetically useful ketones can be transformed into the corresponding chiral secondary alcohols by choosing the appropriate dehydrogenase from the above-mentioned set of enzymes (Fig. 2.15). Other enzymes, which have been shown to be useful for specific types of substrates bearing a carbonyl group, are mentioned below.
The natural role of glycerol dehydrogenase is the interconversion of glycerol and dihydroxyacetone. The enzyme is commercially available from different sources and has been used for the stereoselective reduction of α-hydroxyketones [750]. Glycerol DH has been found to tolerate some structural variation of its natural substrate – dihydroxyacetone – including cyclic derivatives. An enzyme from Geotrichum candidum was shown to reduce not only α- but also β-ketoesters with high selectivity [829].
Enzymes from thermophilic organisms (which grow in the hostile environment of hot springs with temperatures ranging from 70 to 100°C) have recently received much attention [830–833]. Thermostable enzymes are not only stable to heat but, in general, also show enhanced stability in the presence of common protein denaturants and organic solvents. Since they are not restricted to working in the narrow temperature range which is set for mesophilic, ‘normal’ enzymes (20–40°C), an influence of the temperature on the selectivity can be studied over a wider range. For instance, the diastereoselectivity of the HLADH-catalyzed reduction of 3-cyano-4,4-dimethyl-cyclohexanone is diminished at 45°C (the upper operational limit for HLADH) when compared with that observed at 5°C [834]. On the other hand, a temperature-dependent reversal of the enantiospecificity of an alcohol dehydrogenase from Thermoanaerobacter ethanolicus could be achieved when the temperature was raised to 65°C [835] (compare Sect. 2.1.3.1).
2.2.3 Reduction of Aldehydes and Ketones Using Whole Cells
Instead of isolated dehydrogenases, which require sophisticated cofactor recycling, whole microbial cells can be employed. They contain multiple dehydrogenases which are able to accept nonnatural substrates, all the necessary cofactors and the metabolic pathways for their regeneration. Thus, cofactor recycling can be omitted since it is automatically done by the living cell. Therefore, cheap carbon sources such as saccharose or glucose can be used as auxiliary substrates for asymmetric reduction reactions. Furthermore, all the enzymes and cofactors are well protected within their natural cellular environment.
However, these distinct advantages have to be taken into consideration alongside some significant drawbacks:
-
The productivity of microbial conversions is usually low since the majority of nonnatural substrates are toxic to living organisms and are therefore only tolerated at low concentrations (~0.1–0.3% per volume).
-
The large amount of biomass present in the reaction medium causes low overall yields and makes product recovery troublesome, particularly when the product is stored inside the cells and not excreted into the medium. Since only a minor fraction (typically 0.5–2%) of the auxiliary substrate is used for coenzyme recycling, the bulk of it is metabolized, forming polar byproducts which often impede product purification. Therefore, monitoring of the reaction becomes difficult.
-
Finally, different strains of a microorganism most likely possess different specificities; thus it is important to use exactly the same culture to obtain comparable results with the literature [836].
-
Stereoselectivities may vary to a great extent due to the following reasons:
On the one hand, a substrate may be reduced by a single oxidoreductase via transition states for the two enantiomers (or the enantiotopic faces) which have similar values of free energy. In other words, inefficient chiral recognition takes place allowing an alternative fit of the substrate within a single enzyme.
If two enzymes, each with high but opposite stereochemical preference, compete for the same substrate, the optical purity of the product is determined by the relative rates of the individual reactions. The latter, in turn, depend on the substrate concentration for the following reasons: At concentrations below saturation, the relative rates are determined by the ratio V max/K m for each enzyme. On the other hand, when saturation is reached using elevated substrate concentrations, the relative rates mainly depend on the ratio of k cat of the two reactions. Consequently, when two (or more) enzymes are involved in the transformation of enantiomeric substrates, the optical purity of the product becomes a function of the substrate concentration, because the values of K m and k cat for the substrate enantiomers are different for both competing enzymes. With yeasts, it is a well-known phenomenon that lower substrate concentrations often give higher e.e.ps [837].
The following general techniques can be applied to enhance the selectivity of microbial reduction reactions:
-
Substrate modification, e.g., by variation of protecting groups which can be removed after the transformation [838–840]
-
Variation of the metabolic parameters by immobilization [841–843]
-
Using cells of different age [844]
-
Screening of microorganisms to obtain strains with the optimum properties (a hard task for nonmicrobiologists) [848, 849]
-
Selective inhibition of one of the competing enzymes (see below)
2.2.3.1 Reduction of Aldehydes and Ketones by Baker’s Yeast
Asymmetric Reduction of Ketones. Baker’s yeast (Saccharomyces cerevisiae) is by far the most widely used microorganism for the asymmetric reduction of ketones [850–854]. It is ideal for nonmicrobiologists, since it is readily available at a very reasonable price. In addition, its use does not require sterile fermenters and it can therefore be handled using standard laboratory equipment. Thus, it is not surprising that yeast-catalyzed transformations of nonnatural compounds leading to chiral products have been reported from the beginning of the twentieth century [855] and the first comprehensive review which covers almost all the different strategies of yeast-reductions dates back to 1949! [856].
A wide range of functional groups within the ketone are tolerated, including heterocyclic- [857, 858], fluoro- [859–862], chloro- [863], bromo- [864], perfluoroalkyl- [865], cyano-, azido-, nitro- [866–868], hydroxyl- [869, 870], sulfur- [871–873], and dithianyl groups [874]. Even organometallic derivatives [875, 876], such as silyl- [877] and germyl groups [878] are accepted.
Simple aliphatic and aromatic ketones are reduced by fermenting yeast according to Prelog’s rule to give the corresponding (S)-alcohols in good optical purities (Scheme 2.122) [771]. Long-chain ketones such as n-propyl-n-butylketone and several bulky phenyl ketones are not accepted; however, one long alkyl chain is tolerated if the other moiety is the methyl group [879, 880]. As might be expected, best stereoselectivities were achieved with groups of greatly different size.

Reduction of aliphatic and aromatic ketones using baker’s yeast
Acyclic β-ketoesters (Scheme 2.123) are readily reduced by yeast to yield β-hydroxyesters [881, 882], which serve as chiral starting materials for the synthesis of β-lactams [883], insect pheromones [884], and carotenoids [885]. It is obvious that the enantioselectivity and the stereochemical preference for the re- or the si-side of the β-ketoester depends on the relative size of the alkoxy moiety and the ω-substituent of the ketone, which directs the nucleophilic attack of the hydride occurring according to Prelog’s rule (Scheme 2.123). Therefore, the absolute configuration of the newly generated sec-alcoholic center may be directed by substrate modification using either the corresponding short- or long-chain alkyl ester, which switches the relative size of the substituents flanking the carbonyl group [886].

Reduction of acyclic β-ketoesters using baker’s yeast
In baker’s yeast, the reason for this divergent behavior is not due to an alternative fit of the substrates in a single enzyme, but rather due to the presence of a number of different dehydrogenases, possessing opposite stereochemical preferences, which compete for the substrate [837, 887]. A d-specific enzyme – belonging to the fatty acid synthetase complex – shows a higher activity towards β-ketoesters having a short-chain alcohol moiety, such as methyl esters. By contrast, an l-enzyme is more active on long-chain counterparts, e.g., octyl esters. Therefore, the stereochemical direction of the reduction may be controlled by careful design of the substrate, or by selective inhibition of one of the competing dehydrogenases.
Inhibition of the l-enzyme (which leads to the increased formation of d-β-hydroxyesters) was accomplished by addition of unsaturated compounds such as allyl alcohol [888] or methyl vinyl ketone (Table 2.4, Scheme 2.123) [889]. The same effect was observed when the yeast cells were immobilized by entrapment into a polyurethane gel [890, 891]. As expected, l-enzyme inhibitors led to a considerable increase in the optical purity of d-β-hydroxyesters.
Various haloacetates [892], thioethers [893], and allyl bromide [894] have been found to be selective inhibitors for the d-enzyme (Table 2.5, Scheme 2.123), which leads to an increased formation of the l-enantiomer. Depending on the substrate, the effect may even be pronounced enough to effect an inversion of the configuration of the resultant β-hydroxyester. In the absence of inhibitor, the d-enantiomer was obtained in moderate e.e., but the l-counterpart was formed upon addition of allyl bromide.
Diastereoselective Reduction of Ketones by Baker’s Yeast. Asymmetric microbial reduction of α-substituted ketones leads to the formation of diastereomeric syn- and anti-products. Because the chiral center on the α-position of the ketone is stereochemically labile, rapid in-situ racemization of the substrate enantiomers occurs via enolizationFootnote 30 – leading to dynamic resolution [67, 895, 896]. Thus, the ratio between the diastereomeric syn- and anti-products is not 1:1, but is determined by the selectivities of the enzymes involved in the reduction process [897]. Under optimal conditions it can even be as high as 100:0 [898].
Diastereoselective yeast-reduction of ketones has been mainly applied to α-monosubstituted β-ketoesters leading to the formation of diastereomeric syn- and anti-β-hydroxyesters (Scheme 2.124) [899–902]. With small α-substituents, the formation of syn-diastereomers predominates, but the diaselectivity is reversed when the substituents are increased in size. In any case, the selectivity for the newly generated sec-alcohol center is always very high (indicated by the e.e.s) and its absolute configuration is determined by Prelog’s rule.

Diastereoselective reduction of α-substituted β-ketoesters using baker’s yeast
These observations have led to the construction of a simple model which allows one to predict the diastereoselectivity (i.e., the syn/anti-ratio) of yeast-catalyzed reductions of α-substituted β-ketoesters (Fig. 2.17) [903]. Thus, when α-substituents are smaller than the carboxylate moiety, they fit well into the small pocket (S), with L being the carboxylate, but substrates bearing space-filling groups on the α-carbon occupy both of the pockets in an inverted orientation (i.e., the carboxylate group then occupies the pocket S). Again, the reason for the varying selectivity of baker’s yeast reduction of α-substituted β-ketoesters is due to the presence of multiple dehydrogenases possessing different specificites for the substrate enantiomers [904].

Model for predicting the diastereoselectivity in yeast reductions
As may be predicted by this model, the yeast-reduction of cyclic ketones bearing an α-alkoxycarbonyl group exclusively leads to the corresponding syn-β-hydroxyesters (Scheme 2.125) [905–907]. The corresponding anti-diastereomers cannot be formed because rotation around the α,β-carbon–carbon bond is impossible with such cyclic structures. Furthermore, the reductions are generally more enantioselective than the corresponding acyclic substrate due to the enhanced rigidity of the system. Thus, it can be worthwhile to create a sulfur-containing ring in the substrate temporarily and to remove the heteroatom after the biotransformation to obtain the desired open-chain product (e.g., by Raney-Ni reduction) in order to benefit from enhanced selectivities (compare Scheme 2.45).

Yeast-reduction of cyclic β-ketoesters
The biocatalytic reduction of α-substituted β-ketoesters with concomitant dynamic resolution has been proven to be extremely flexible (Scheme 2.126) [908–913]. Thus, by choosing the appropriate microorganism possessing the desired enantio- and diastereoselectivity, each of the four possible diastereomeric products were obtained in excellent enantiomeric and diastereomeric purity. As expected, the corresponding Prelog-configurated products (with respect to the newly generated sec-alcohol center, pathways A, B) were obtained by using baker’s yeast and the mold Geotrichum candidum, respectively. For the diastereomers possessing the opposite configuration at the alcoholic center (anti-Prelog pathways C, D) other microorganisms had to be employed.

Stereocomplementary microbial reduction of α-substituted β-ketoesters
As long as the α-substituent consists of an alkyl- or aryl-group, dynamic resolution is readily achieved, leading to chemical yields far beyond the 50% which would be the maximum for a classic kinetic resolution. However, in-situ racemization is not possible due to electronic reasons for α-hydroxy- [914], α-alkylthio- [899], α-azido- [915], or α-acetylamino derivatives [916], which are subject to kinetic resolution. The same holds for substrates which are fully substituted at the α-position, due to the impossibility of form the corresponding enolate.
α-Ketoesters and α-ketoamides can be asymmetrically reduced to furnish the correponding α-hydroxy derivatives. Thus, following Prelog’s rule, (S)-lactate [917] and (R)-mandelate esters [897] were obtained from pyruvate and α-ketophenylacetic esters by fermenting baker’s yeast in excellent optical purity (e.e. 91–100%).
Cyclic β-diketones are selectively reduced to give β-hydroxyketones without the formation of dihydroxy products (Scheme 2.127) [918–921]. It is important, however, that the highly acidic protons on the α-carbon atom are fully replaced by substituents in order to avoid the (spontaneous) chemical condensation of the substrate with acetaldehyde, which is always present in yeast fermentationsFootnote 31 and to avoid racemization of the β-hydroxyketone formed as product. Again, with small-size rings, the corresponding syn-products are formed predominantly, usually with excellent optical purity. However, the diastereoselectivity becomes less predictable and the yields drop when the rings are enlarged. Again, the stereochemistry at the newly formed secondary alcohol center can be predicted by Prelog’s rule.

Yeast reduction of cyclic β-diketones
In contrast, the reduction of α-diketones does not stop at the keto-alcohol stage but leads to the formation of diols (Scheme 2.128). In general, the less hindered carbonyl group is quickly reduced in a first step to give the (S)-α-hydroxyketone according to Prelog’s rule, but further reduction of the (usually more sterically hindered) remaining carbonyl group yields the corresponding diols predominantly in the anti-configuration as the final product [922, 923].

Yeast reduction of α-diketones
Secondary alcohols possessing the anti-Prelog configuration can be obtained from yeast reductions via substrate modification (Scheme 2.123) or through enzyme inhibition (Scheme 2.123, Tables 2.4 and 2.5). If these techniques are unsuccessful, the use of microorganisms other than yeast [767, 773, 924–928], such as Pichia farinosa [929], Geotrichum candidum [930, 931], and Yarrowia lipolytica [932] may be of an advantage. Even plant cell cultures such as Gardenia may be employed for this purpose [933, 934]. However, in this case the help of a microbiologist is recommended for organic chemists.
The high level of technology available for the biocatalytic reduction of carbonyl compounds has allowed its implementation for the industrial-scale production of chiral building blocks containing sec-alcohol moieties. The system applied – either whole microbial (wild-type) cells, a designer bug containing a dehydrogenase plus cofactor-recycling enzyme, or the use of isolated enzymes – depends on the case and is mainly dependent on the economic and the patent situation. Representative examples are depicted in (Scheme 2.129) [935–940].

Industrial-scale bioreduction of carbonyl compounds
Deracemization via Biocatalytic Stereoinversion. Racemic secondary alcohols may be converted into a single enantiomer via stereoinversion which proceeds through a two-step redox sequence (Scheme 2.130) [38, 941, 942]: In a first step, one enantiomer from the racemic mixture is selectively oxidized to the corresponding ketone while the other enantiomer remains unaffected. Then, the ketone is reduced in a second subsequent step by another redox-enzyme displaying opposite stereochemical preference. Overall, this process constitutes a deracemization technique, which leads to the formation of a single enantiomer in 100% theoretical yield from the racemate [943]. Due to the presence of two consecutive oxidation–reduction reactions, the net redox balance of the process is zero and no external cofactor recycling is required since the redox equivalents are exchanged between both steps in a closed loop. In order to achieve a high optical purity of the product, at least one of the steps has to be irreversible for entropic reasons [944, 945].

Deracemization of sec-alcohols via microbial stereoinversion
The origin of the irreversibility of microbial/enzymatic deracemization of sec-alcohols proceeding through an oxidation–reduction sequence depends on the type of microorganism and is unknown in most cases. For instance, deracemization of various terminal (±)-1,2-diols by the yeast Candida parapsilosis has been claimed to operate via an (R)-specific NAD+-linked dehydrogenase and an irreversible (S)-specific NADPH-dependent reductase [944]. Along these lines, the enzymatic stereoinversion of (biologically inactive) d-carnitine to furnish the desired bioactive l-enantiomerFootnote 32 was accomplished by using two stereocomplementary carnitine dehydrogenases. Due to the fact that both dehydrogenase enzymes are NAD(H)-dependent, the end point of the process was close to equilibrium (64%) [950]. By contrast, the stereoinversion of β-hydroxyesters using the fungus Geotrichum candidum required molecular oxygen, which would suggest the involvement of an alcohol oxidase rather than an alcohol dehydrogenase [951]. Recently, the mechanism of enzymatic stereoinversion catalyzed by stereocomplementary dehydrogenases was elucidated to depend on the opposite cofactor-dependence for NADH and NADPH of the dehydrogenases involved [952, 953].
Microbial stereoinversion of sec-alcohols has become quite popular [954]. For instance, the deracemization of simple secondary alcohols proceeds with excellent results using the fungi Geotrichum candidum or Candida parapsilosis. In case the oxidation and reduction cannot be performed by a single species, two microorganisms may be used instead. For instance, Bacillus stearothermophilus and Yarrowia lipolytica or Pseudomonas polycolor and Micrococcus freudenreichii were coupled for the deracemization of the pheromone sulcatol and mandelic acid, respectively. In a similar fashion, (±)-pantoyl lactone – a key intermediate for the synthesis of pantothenic acid [955] – was deracemized by using washed cells of Rhodococcus erythropolis or Candida sp. (Scheme 2.131) [956, 957]. Thus, l-pantoyl lactone is oxidized to the α-ketolactone, which in turn is reduced by another dehydrogenase present in the organisms to yield the corresponding (R)-d-pantoyl lactone in 100% theoretical yield.

Microbial deracemization of pantoyl lactone
Microbial stereoinversion has been shown to be extremely flexible, as it is also applicable to sec-diols possessing two stereocenters (Scheme 2.132) [958–960]. Thus, meso- or rac-trans-cyclohexane-1,2-diol was deracemized by Corynesporium cassiicola DSM 62475 to give the (1S,2S)-enantiomer as the sole product in >99% e.e. and 83% yield. The process was shown to proceed in a stepwise fashion via the corresponding hydroxyketone as intermediate, which was detected in small amounts. More important is the deracemization of rac-trans-indane-1,2-diol, which was accomplished with excellent results in a similar fashion. The (1S,2S)-isomer is a central building block for the anti-HIV-agent indinavir [961].

Microbial stereoinversion of cyclic 1,2-diols
2.2.3.2 Reductive Amination of α-Ketocarboxylic Acids
The (reversible) transformation of an α-ketocarboxylic acid in presence of ammonia and one equivalent of NAD(P)H furnishes the corresponding α-amino acid and is catalyzed by amino acid dehydrogenases [EC 1.4.1.X] [962]. Despite major differences in its mechanism, this reaction bears a strong resemblance to carbonyl group reduction and it formally respresents a reductive amination (Scheme 2.133). As deduced for l-Leu-dehydrogenase [963], the α-ketoacid substrate is positioned in the active site between two Lys-residues. Nucleophilic attack by NH3 leads to a hemiaminal intermediate, which eliminates H2O to form an iminium species. The latter is reduced by a hydride from nicotinamide forming the l-amino acid. Since this mechanism is highly tuned for α-keto/α-amino acids, it is clear that a neutral Schiff base cannot be accepted as substrate.

Reductive amination of α-ketocarboxylic acids
Among the various amino acid dehydrogenases, Leu-DH has captured an important role for the synthesis of nonproteinogenic l-α-amino acids via asymmetric reductive amination of the corresponding α-ketoacids [964, 965]. A range of protease inhibitors used for the treatment of tumors and viral infections contain sterically hindered amino acids as key element for their biological action. The latter cannot be synthetized via the conventional (protease-dependent) methods (Sect. 2.1), but they are produced on industrial-scale making use of the relaxed substrate specificity of LeuDH in combination with NADH recycling using the formate dehydrogenase/formate system [966].
2.2.4 Reduction of C=C-Bonds
The asymmetric (bio)catalytic reduction of C=C-bonds goes in hand with the creation of (up to) two chiral centers and is thus one of the most widely employed strategies for the production of chiral compounds. Whereas cis-hydrogenation using transition-metal based homogeneous catalysts has been developed to an impressive standard,Footnote 33 stereocomplementary asymmetric trans-hydrogenation is still at the stage of development [967].
The biocatalytic counterpart for the stereoselective reduction of alkenes is catalyzed by flavin-dependent ene-reductases,Footnote 34 EC 1.3.1.31], which are members of the ‘old yellow enzyme’ family (OYE, Scheme 2.134) [968, 969], first described in the 1930s [970]. These enzymes are widely distributed in microorganisms and in plants. Some of them occur in well-defined pathways, e.g., in the biosynthesis of fatty acids and secondary metabolites, such as morphine [971] and jasmonic acid [972]. Others are involved in the detoxification of xenobiotics [973], such as nitro esters [974] and nitro-aromatics [975] like trinitrotoluene (TNT) [976].

Asymmetric bioreduction of activated alkenes using flavin-dependent ene-reductases
The catalytic mechanism of the asymmetric reduction of alkenes catalyzed by ene-reductases has been studied in great detail [977] and it has been shown that a hydride (derived from a reduced flavin cofactor) is stereoselectively transferred onto Cβ, while a Tyr-residue adds a proton (which is ultimately derived from the solvent) onto Cα from the opposite side (Scheme 2.134). As a consequence of the stereochemistry of this mechanism, the overall addition of [H2] proceeds in a trans-fashion with absolute stereospecificity [978].Footnote 35 This reaction is generally denoted as the ‘oxidative half reaction’. The catalytic cycle is completed by the so-called ‘reductive half reaction’ via reduction of the oxidized flavin cofactor at the expense of NAD(P)H, which is ultimately derived from an external H-source via another redox reaction. In contrast to alcohol dehydrogenases (carbonyl reductases), which show a rather pronounced preference for either NADH or NADPH [980], ene-reductases are more flexible in this respect: some enzymes are very specific [981], others are able to accept both cofactors equally well [982, 983]. Overall, the reaction resembles an asymmetric Michael-type addition of a chiral hydride onto an enone and, as a consequence of the mechanism, nonactivated C=C bonds are therefore completely unreactive [984]. Although the overall hydride pathway appears rather complex, practical problems are minimal since flavin cofactors are usually tighly bound to the enzyme and are thereby protected from environment.
Although the remarkable synthetic potential of enoate reductases has been recognized long ago, preparative-scale applications were severely impeded by two major problems: Simple to use whole-cell systems (most prominent baker’s yeast [985], but also fungi and yeasts, such as Geotrichum candidum, Rhodotorula rubra, Beauveria bassiana [986]), Aspergillus niger are plagued by undesired side reactions, such as carbonyl reduction (catalyzed by competing alcohol dehydrogenases/carbonyl reductases) or ester hydrolysis (mediated by carboxyl ester hydrolases) [711]. On the other hand, the first generation of isolated (cloned) enoate reductases were obtained from (strict or facultative) anaerobes, such as Clostridia [987] or methanogenic Proteus sp. [988], which were inapplicable to preparative-scale transformations due to their sensitivity towards traces of molecular oxygen. It was only recently, that this bottleneck was resolved by providing oxygen-stable OYEs from bacteria, plants, and yeasts [989–996].
The following crude guidelines for the asymmetric bioreduction of activated alkenes using ene-reductases can be delineated:
-
Only C=C-bonds which are ‘activated’ by electron-withdrawing substituents (EWG) are reduced (Scheme 2.135) [997], electronically ‘isolated’ double bonds are not accepted [998]. Acetylenic triple bonds yield the corresponding (E)-alkenes [999]. With activated, conjugated 1,3-dienes only the α,β-bond is selectively reduced, leaving the nonactivated γ,δ-bond behind (Scheme 2.135). In a similar manner, cumulated 1,2-dienes (allenes) mainly give the corresponding 2-alkenes. A rearrangement of the allene to give an acetylene may be observed occasionally [1000].
Scheme 2.135 
Asymmetric bioreduction of enals and allylic alcohols using isolated ene-reductase and baker’s yeast

The following functional groups may serve as ‘activating’ groups:
-
α,β-Unsaturated carboxaldehydes (enals) are quickly reduced in a clean fashion yielding saturated aldehydes when pure ene-reductases are used. In contrast, whole-cell reductions are heavily plagued by competing carbonyl reduction, which often outcompetes the ene-reductase to furnish the corresonding allylic alcohol (thereby depleting the substrate) and/or the saturated prim-alcohol (via over-reduction of the desired product) [1001, 1002]. These undesired side-reactions sometimes allow to use an allylic alcohol as substrates, which is transformed via the corresponding enal by whole cells [1003] (Scheme 2.135).
-
α,β-Unsaturated ketones are good substrates for ene-reductases. With whole cells, competing carbonyl-reduction is slower as compared to enals and the product distribution depends on the relative rates of competing carbonyl- and ene-reductases [1004, 1005] (Scheme 2.136).
Scheme 2.136 
Asymmetric bioreduction of α,β-unsaturated diketone
Scheme 2.137 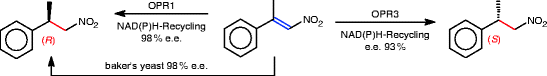
Stereocomplementary bioreduction of nitro-olefins
-
α,β-Unsaturated nitro compounds can be readily ransformed into chiral nitro-alkanes, depending on the type of OYE, reductive biodegradation may occur via the Nef-pathway [1006]. Due to the high acidity of nitroalkanes, any chiral center at Cα is racemized, wheras Cβ-analogs are perfectly stable [1007] (Scheme 2.137).
Scheme 2.138 
Stereocontrol of ene-reduction via enzyme-type or substrate-configuration
-
Cyclic imides are readily reduced without competing side reactions.
-
α,β-Unsaturated carboxylic acids or esters have to be regarded as ‘borderline’-substrates:
Simple α,β-unsaturated mono-carboxylic acids or -esters are not easily reduced by OYEs, but they are good substrates for ‘enoate-reductases’ from anaerobic organisms, which possess an additional (oxygen-sensitive) ferredoxin cofactor [1008]. However, the presence of an additional electron-withdrawing group (which alone would not be sufficient to act as activator), such as halogen, nitrile, etc., helps to overcome the low degree of activation [1009] (Scheme 2.138). Consequently, di-carboxylic acids and -esters are accepted by OYEs. Ester hydrolysis is a common side-reaction when using whole cells. Only few reports are available regarding α,β-unsaturated lactones. Due to their low carbonyl activitiy, carboxylic acids are less activated than the corresponding esters.
Scheme 2.139 
Asymmetric yeast reduction of α,β-unsaturated lactones
-
Sometimes the absolute (R/S)-configuration of the product can be controlled by starting with (E)- or (Z)-alkenes. However, this is not always the case (Scheme 2.138).
-
Steric hindrance at Cβ (where the hydride has to be delivered) seems to play an important role for OYEs. Consequently, sterically demanding substituents at the C=C-bond are more easily tolerated in the α-position.




The bioreduction of citral using the ene-reductase OPR3 (12-oxophytodienoic acid reductase) proceeds in a clean fashion yielding the fragrance compound (R)-citronellal in excellent chemical and optical yields (Scheme 2.135). In contrast, baker’s yeast reduction of a closely related enal bearing a carboxylic ester group yielded the saturated prim-alcohol as the major product cue to over-reduction of the aldehyde moiety. The less activated C=C bond adjacent to the ester remained unchanged [1010]. Instead of starting with an enal (whose aldehyde moiety would be reduced anyway) the corresponding allylic alcohol may serve as substrate in whole-cell bioreductions. Thus, geraniol gave (R)-citronellol in >97% e.e. [1011] and in a similar fashion only the α,β-bond was reduced in a conjugated 1,3-diene-1-ol [1012]. It is important to note that in whole-cell transformations the C=C-reduction of allylic alcohols always occurs at the aldehyde stage, which is reversibly formed as intermediate (Scheme 2.135).
In contrast to aldehydes, over-reduction is less pronounced on α,β-unsaturated ketones (Scheme 2.136). Nonracemic levodione, which is a precursor for the synthesis of carotenoids, such as astaxanthin and zeaxanthin, was obtained in 80% yield and >95% e.e. via yeast-mediated reduction of ketoisophorone. Two other products arising from over-reduction of the carbonyl moieties were formed in minor amounts [1013]. In contrast, no trace of carbonyl reduction was observed using ene-reductase OPR3 (Scheme 2.136).
Nitro-olefins are readily reduced by ene-reductases to form chiral nitro-alkanes (Scheme 2.137) [1014]. Using ene-reductase OPR1 or baker’s yeast, the corresponding (R)-nitroalkanes were obtained in high e.e. Surprisingly, the mirror-image product was formed by using isoenzyme OPR2, which is highly homologous to OPR3 (53%).
Simple monocarboxylic esters require the presence of an additional activating group, such as a halogen atom or a second ester, to be accepted by OYEs (Scheme 2.138). α-Substituted butenedioic esters were readily reduced by ene-reductases YqjM and OPR1 with excellent specificities, while the stereochemical outcome could be controlled by choice of the ene-reductase or by using an (E)- or (Z)-configurated substate: (R)-2-Methylsuccinate was obtained by using OPR1, regardless of the (E/Z)-configuration of the substrate. In contrast, with YqjM, the configuration of the product switched when an (E)-fumarate ester was used instead of a (Z)-maleate [1015].
A similar dependence of the configuration of the product on the (E/Z)-configuration of the substrate was observed during the yeast-mediated reduction of 2-chloro-acrylate esters [1016]. Whereas the chiral recognition of the (Z)-alkenes was perfect, the (E)-isomers gave products with lower e.e. In addition, it was shown that the microbial reduction took place on the carboxylic acid stage, which were formed enzymatically by hydrolysis of the starting esters prior to the reduction step [1017] (Scheme 2.138).
Only few reports are available on the asymmetric bioreduction of α,β-unsaturated lactones. For instance, β-substituted five-membered ring lactones were readily reduced by baker’s yeast to give the (R)-configurated saturated analogs (Scheme 2.139) [1018]. The latter constitute versatile C5-building blocks for terpenoid synthesis. The polarity of the sulfur-protecting group had a significant impact on the stereochemical course of the reaction. Whereas the thioether and the sulfoxide were readily converted with excellent selectivities, the corresponding more polar sulfone gave low chemical and optical yields.
The ultimate source of redox equivalents in microbial reduction reactions is usually a carbohydrate. Since the majority of it is metabolized by the cells and only a minor fraction (typically 0.5–2%) is used for the delivery of redox equivalents onto the substrate, the productivity of such processes is usually low and side-reactions are common. In order to avoid the undesired metabolism of the auxiliary substrate, nondegradable organic dye molecules such as viologens have been used as shuttles (‘mediators’) for the electron-transport from the donor to the oxidized cofactor [1019]. Provided that the mediators are accepted by the ene-reductases and the recycling enzymes, the productivities were improved by one to three orders of magnitude.
The reduced form of the mediator (a radical cation) can regenerate NADH. The oxidized mediator (a dication) can be enzymatically recycled using an inexpensive reducing agent, such formate in combination with formate dehydrogenase, which is often present together with the enoate reductase in the same microorganism. Alternatively, electrons from the cathode of an electrochemical cell can be used in a nonenzymatic reaction. In this case, the course of the reaction can be easily monitored by following the current in the electrochemical cell. In addition, the equilibrium of the reaction can be shifted from reduction to oxidation by choosing a mediator with the appropriate redox potential. The disadvantage of this method is the considerable toxicity of the commonly used mediators, e.g., methyl- or benzyl-viologen, which have been used for some time as total herbicides, which are banned today (Scheme 2.140).Footnote 36

Cofactor recycling via a mediator
2.3 Oxidation Reactions
Oxidation constitutes one of the key steps for the introduction of functional groups into the raw materials of organic synthesis which are almost invariably an alkane, an alkene, or an aromatic molecule.Footnote 37 Traditional methodology is plagued by several drawbacks, that is
-
Many oxidants are based on toxic metal ions such as copper, nickel, or chromium, which are environmentally incompatible.
-
Undesired side reactions are common due to a lack of chemoselective oxidation methods.
-
The most inexpensive and innocuous oxidant, molecular oxygen, cannot be used efficiently.
-
It is extremely difficult to perform oxidations in a regio- and stereoselective fashion.
Therefore, organohalogens have been widely used as intermediates for the synthesis of oxygenated compounds, which has led to severe problems in waste treatment due to recalcitrant halogenated organic compounds.
Many of the drawbacks mentioned above can be circumvented by using biological oxidation, in particular for those cases where stereoselectivity is required [1021, 1022].
The biooxidation reactions discussed in this chapter are grouped according to their requirement for the oxidant, i.e.:
-
Dehydrogenation depending on a nicotinamide cofactor [NAD(P)H] (Sect. 2.3.1)
-
Oxygenation at the expense of molecular oxygen (Sect. 2.3.2)
-
Peroxidation reactions; requiring hydrogen peroxide or a chemical derivative thereof (Sect. 2.3.3)
For a classification of biooxidation reactions see Scheme 2.144.
2.3.1 Oxidation of Alcohols and Aldehydes
Oxidations of primary and secondary alcohols catalyzed by dehydrogenases to furnish aldehydes and ketones, respectively, are common chemical reactions that rarely present insurmountable problems to the synthetic organic chemist. In contrast to the corresponding reduction reactions, oxidation reactions using isolated dehydrogenase enzymes have been rather scarcely reported [1023]. The reasons for this situation are as follows.
-
Oxidations of alcohols using NAD(P)+-dependent dehydrogenases are thermodynamically unfavorable. Thus, the recycling of the oxidized nicotinamide cofactor becomes a complicated issue (see p. 144).
-
Due to the fact that the lipophilic aldehydic or ketonic products are often more tightly bound onto the hydrophobic active site of the enzyme than the more hydrophilic substrate alcohol, product inhibition is a common phenomenon in such reactions [759].
-
Enzymatic oxidations usually work best at elevated pH (8–9) where nicotinamide cofactors and (particularly aldehydic) products are unstable.
-
Oxidation of a secondary alcohol involves the destruction of an asymmetric center (sp3 → sp2 hybrid) and is therefore of limited synthetic use.
2.3.1.1 Regioselective Oxidation of Polyols
The enzyme-catalyzed oxidation of alcohols is only of practical interest to the synthetic organic chemist if complex molecules such as polyols are involved (Scheme 2.141) [1024–1028]. Such compounds present problems of selectivity with conventional chemical oxidants. The selective oxidation of a single hydroxyl group in a polyol requires a number of protection–deprotection steps if performed by chemical means. In contrast, numerous sugars and related polyhydroxy compounds have been selectively oxidized in a single step into the corresponding keto-ols or ketoacids using a variety of microorganisms, for example Acetobacter or Pseudomonas sp. The regioselective microbial oxidation of d-sorbitol (obtained by catalytic hydrogenation of d-glucose) by Acinetobacter suboxydans yields l-sorbose, which represents the key step in the famous Reichstein–Grüssner process for the production of l-ascorbic acid (vitamin C).

Regioselective oxidation of polyols by Acetobacter suboxydans
2.3.1.2 Kinetic Resolution of Alcohols by Oxidation
The major difficulty in the resolution of alcohols via selective oxidation of one enantiomer using isolated horse liver alcohol dehydrogenase (HLADH) is the regeneration of NAD+. Besides the highly efficient enzymatic systems described in Sect. 2.2.1, a flavin mononucleotide (FMN) recycling system [1029] in which molecular oxygen is the ultimate oxidant was used to resolve numerous mono-, bi-, and polycyclic secondary alcohols by HLADH [786, 788, 1030, 1031]. To avoid enzyme deactivation, the hydrogen peroxide produced from O2 during this process was removed using catalase [1032].
Glycols having a primary hydroxyl group were enantioselectively oxidized to yield l-α-hydroxyacids using a coimmobilized alcohol and aldehyde dehydrogenase system (Scheme 2.142). In the first step, kinetic resolution of the diol furnished a mixture of l-hydroxyaldehyde and the remaining d-diol. The former was oxidized in-situ by an aldehyde dehydrogenase to yield the l-hydroxyacid in high optical purity [791, 1033].

Kinetic resolution of diols by a HLADH/aldehyde DH system
2.3.1.3 Desymmetrization of Prochiral or meso-Diols by Oxidation
In contrast to the resolution of secondary alcohols, where the more simple lipase technology is recommended instead of redox reactions, desymmetrization of primary diols of prochiral or meso-structure has been shown to be a valuable method for the synthesis of chiral lactones (Scheme 2.143) [1034].

Desymmetrization of meso-diols by HLADH
As a rule of thumb, oxidation of the (S)- or pro-(S) hydroxyl group occurs selectively with HLADH (Scheme 2.143). In the case of 1,4- and 1,5-diols, the intermediate γ- and δ-hydroxyaldehydes spontaneously cyclize to form the more stable five- and six-membered hemiacetals (lactols). The latter are further oxidized in a subsequent step by HLADH to form γ- or δ-lactones following the same (S)- or pro-(S) specificity [1035]. Both steps – desymmetrization of the prochiral or meso-diol and kinetic resolution of the intermediate lactol – are often highly selective. By using this technique, enantiopure lactones were derived from cis-meso-2,3-dimethylbutane-1,4-diol and the cyclic thia-analog [1036], similar results were obtained with sterically demanding bicyclic meso-diols [1037].
2.3.2 Oxygenation Reactions
Enzymes which catalyze the direct incorporation of molecular oxygen into an organic molecule are called ‘oxygenases’ [1038–1041]. Enzymatic oxygenation reactions are particularly intriguing since direct oxyfunctionalization of nonactivated organic compounds remains a largely unresolved challenge to synthetic chemistry. On the one hand, there are numerous (catalytic) oxidation processes developed by industry to convert simple raw materials, such as alkanes, alkenes and aromatics into more valuable intermediate products (such as aldehydes, ketones, alcohols, carboxylic acids, etc.) at the expense of O2.Footnote 38 However, the catalysts employed are highly sophisticated and thus show a very narrow substrate range, which limits their applicability to a single (or few) substrate(s) and they cannot be used for a wider range of organic compounds.Footnote 39 Unsurmountable problems persist where regio- or enantiospecificity is desired. On the other hand, highly selective oxygenation reactions may be achieved by means of biocatalysts.
Oxygen-transfer from molecular oxygen into organic acceptor molecules may proceed through three different mechanisms (Scheme 2.144).

Enzymatic oxidation reactions
-
Monooxygenases incorporate one oxygen atom from molecular oxygen into the substrate, the other is reduced at the expense of a donor (usually NADH or NADPH) to form water [1042–1044].
-
Dioxygenases simultaneously incorporate both oxygen atoms of O2 into the substrate by forming a peroxy-species, thus they are sometimes misleadingly called oxygen transferases (although they are redox enzymes belonging to EC class 1).
-
Oxidases [EC 1.1.3.X], on the other hand, mainly catalyze the electron-transfer onto molecular oxygen. This may proceed through a two- or four-electron transfer, involving either hydrogen peroxide or water as byproduct of the oxidation reaction, respectively. Oxidases include flavoprotein oxidases (amino acid oxidases, glucose oxidase, nicotinamide oxidase), metallo-flavin oxidases (aldehyde oxidase) and heme-protein oxidases (catalase, H2O2-specific peroxidases [1045]). Some of the enzymes have been found to be very useful. For instance, d-glucose oxidase is used on a large scale, in combination with catalase, as a food antioxidant [1046]; however, from a synthetic viewpoint, oxidases have not yet been utilized extensively [1047]. On the contrary, peroxidation reactions catalyzed by peroxidases at the expense of hydrogen peroxide have recently been found to be highly useful (Sect. 2.3.3).
2.3.2.1 Monooxygenases
Although the reaction mechanisms of various monooxygenases differ greatly depending on the subtype of enzyme, their mode of oxygen-activation is the same. Whereas one of the oxygen atoms from O2 is transferred onto the substrate, the other is reduced to form a water molecule. The latter requires two electrons, which are derived from a cofactor, usually NADH or NADPH, serving as ‘donor’ (Scheme 2.144).
The net reaction and a number of synthetically useful monooxygenation reactions are shown in Scheme 2.145.

Monooxygenase catalyzed reactions
The generation of the activated oxygen-transferring species is mediated either by cofactors containing a transition metal (Fe or Cu) or by a heteroaromatic system (a pteridin [1048] or flavin [1049–1051]). The catalytic cycle of the iron-depending monooxygenases, the majority of which belong to the cytochrome P-450 type (Cyt P-450) [1052–1056], has been deduced largely from studies on the camphor hydroxylase of Pseudomonas putida [1057, 1058]. A summary of the catalytic cycle is depicted in Scheme 2.146.

Catalytic cycle of cytochrome P-450-dependent monooxygenases
The iron species is coordinated equatorially by a heme moiety and axially by the sulfur atom of a cysteine residue. After binding of the substrate (Sub) by replacing a water molecule in a hydrophobic pocket adjacent to the porphine molecule [1059], the iron is reduced to the ferrous state. The single electron is delivered from NAD(P)H via another cofactor, which (depending on the enzyme) is a flavin-nucleotide, an iron-sulfur protein (ferredoxin) or a cytochrome b5. Next, molecular oxygen is bound to give a Cyt P-450 dioxygen complex. Delivery of a second electron weakens the O–O bond and allows the cleavage of the oxygen molecule. One atom is expelled as water, the other forms the ultimate oxidizing Fe4+ or Fe5+ species, which – as a strong electrophile – attacks the substrate. Expulsion of the product (SubO) reforms the iron(III)-species and closes the catalytic cycle. Despite the fact that the mechanism of Cyt P-450 enzymes has been intensively investigated over several decades, many mechanistic details are still poorly understood.
Cyt P-450 enzymes got their name from their hemoprotein character: P stands for ‘pigment’ and 450 reflects the absorption of the CO-complex at 450 nm. To date, more than 6,000 different Cyt P-450 genes have been clonedFootnote 40 and these proteins are classified into four major groups (bacterial, mitochondrial, microsomal and self-sufficient Cyt) according to the mode of the electron-transport and the interaction between the subunits. A simplified schematic organization of Cyt P-450 systems is depicted in Fig. 2.18.

Schematic organization and electron transport of cytochrome P-450 type monooxygenases
Bacterial and mitochondrial Cyt P-450 systems depend on three proteins: the P-450 monooxygenase with its heme unit, which performs the actual oxygenation of the substrate, a ferredoxin reductase, which accepts hydride equivalents from nicotinamide via an FAD cofactor and ferredoxin, which acts as electron shuttle between them using an iron-sulfur cluster as electron carrier [1060]. The microsomal system is somewhat simpler, as electron transfer occurs directly between the cytochrome P reductase (possessing an FMN and FAD cofactor) and the Cyt P-450 enzyme and thus does not require the ferredoxin. A minimal Cyt P-450 system is derived from Bacillus megaterium (BM-3) and it consists of a single (fusion) protein, which is made up of two domains, a cytochrome P reductase (containing FMN and the FeS cluster) and the P-450 enzyme. It is evident, that the latter system has been the prime target of studies directed towards the development of enzymatic oxygenation systems for preparative-scale applications [1061, 1062].
In contrast, flavin-dependent monooxygenases (see Scheme 2.147 and Table of Scheme 2.145) use a different mechanism which involves a flavin cofactor [1063–1065]. First, NADPH reduces the Enz-FAD complex thereby destroying aromaticity. The FADH2 so formed is oxidized via Michael-type addition of molecular oxygen yielding a hydroperoxide species (FAD-4a-OOH). Deprotonation of the latter affords a peroxide anion, which undergoes a nucleophilic attack on the carbonyl group of the substrate, usually an aldehyde or a ketone. The tetrahedral species thus formed (corresponding to a Criegee-intermediate) collapses via rearrangement of the carbon-framework forming the product ester or lactone, respectively. Finally, water is eliminated from the FAD-4a-OH species to reform FAD. In addition to the Baeyer-Villiger oxidation, the flavin-4a-hydroperoxy species can also mediate the hydroxylation of aromatics [1066–1070], alkene epoxidation [1071, 1072] and heteroatom-oxidation.

Catalytic cycle of flavin-dependent monooxygenases
Whereas the Cyt P-450 mechanism resembles the chemical oxidation by hypervalent transition metal oxidants through an electrophilic attack (nature’s permanganate), the FAD-dependent mechanism parallels the oxidation of organic compounds by peroxides or peracids, which act as nucleophiles (nature’s m-chloroperbenzoic acid) [1073].
Many monooxygenases are closely linked to the respiratory chain and are therefore membrane-bound, which makes their isolation and (over)expression difficult. This fact and the need for recycling of NAD(P)H makes it clear that the majority of monooxygenase catalyzed reactions have been performed by using whole microbial cells, of by employing ‘designer bug’ host-cells co-expressing the required proteins. The disadvantage of this method lies in the fact that the desired product is often obtained in low yields due to further metabolism of the oxidation product by the cell. However, selective blocking of enzymes which are responsible for the degradation of the product or using enzyme-deficient mutants has been shown to make microbial oxygenation reactions feasible on a commercial scale.
2.3.2.2 Hydroxylation of Alkanes
The hydroxylation of nonactivated centers in hydrocarbons is one of the most useful biotransformations [1040, 1074–1079] due to the fact that this process has only very few counterparts in traditional organic synthesis [1080–1082]. In general, the relative reactivity of carbon atoms in bio-hydroxylation reactions declines in the order of secondary > tertiary > primary [1083], which is in contrast to radical reactions (tertiary > secondary > primary) [1084]. There are two main groups of hydrocarbon molecules, which have been thoroughly investigated with respect to microbial hydroxylation, i.e., steroids and terpenoids. Both have in common, that they possess a large main framework, which impedes the metabolic degradation of their hydroxylated products.
Intense research on the stereoselective hydroxylation of alkanes started in the late 1940s in the steroid field, driven by the demand for pharmaceuticals [1085–1090] In the meantime, some of the hydroxylation processes, e.g., 9α- and 16α-hydroxylation of the steroid framework [1091, 1092], have been developed to the scale of industrial production. Nowadays, vitually any center in a steroid can be selectively hydroxylated by choosing the appropriate microorganism.Footnote 41 For example, hydroxylation of progesterone in the 11α-position by Rhizopus arrhizus [1094] or Aspergillus niger [1095] made roughly half of the 37 steps of the conventional chemical synthesis redundant and made 11α-hydroxyprogesterone available for hormone therapy at a reasonable cost (Scheme 2.148). A highly selective hydroxylation of lithiocholic acid in position 7β was achieved by using Fusarium equiseti [1096]. The product (ursodeoxycholic acid) is capable of dissolving cholesterol and thus can be used in the therapy of gallstones.

Regio- and stereoselective microbial hydroxylation of steroids
In a similar fashion, active pharmaceutical ingredients (APIs) are subjected to microbial hydroxylation. For instance, the regioselective allylic hydroxylation of the potent cholesterol-lowering drug simvastatin was achieved using Nocardia autotrophica to yield 6-β-hydroxy-simvastatin together with some minor side-products [1097]. An impressive amount of 15 kg of product was obtained from a 19 m3 reactor (Scheme 2.149).

Regioselective microbial hydroxylation of HMG-CoA reductase inhibitor Simvastatin
Optically active β-hydroxy-isobutyric acid has been used as a starting material for the synthesis of vitamins (α-tocopherol [1098]), fragrance components (muscone [1099]) and antibiotics (calcimycin [1100]). Both enantiomers may be obtained by asymmetric hydroxylation of isobutyric acid [1101, 1102] (Scheme 2.150). An intensive screening program using 725 strains of molds, yeasts and bacteria revealed that, depending on the microorganism, either the (R)- or the (S)-β-hydroxy-isobutyric acid was formed in varying optical purity. Best results were obtained using selected Candida and Pseudomonas strains.

Asymmetric microbial hydroxylation of isobutyric acid via the β-oxidation pathway
Although the mechanism of this reaction was initially assumed to be a ‘direct’ hydroxylation at position β, more detailed studies showed that it proceeds via the conventional β-oxidation pathway involved in fatty acid metabolism [1103]. The same sequence is followed during the transformation of 4-trimethylammonium butanoate to the β-hydroxy derivative (carnitine, see Scheme 2.212) [1104].
The production of new materials for the aroma and fragrance industry was the powerful driving force in the research on the hydroxylation of terpenes [924, 1105–1107]. For instance, 1,4-cineole, a major constituent of eucalyptus oil, was regioselectively hydroxylated by Streptomyces griseus to give 8-hydroxycineole as the major product along with minor amounts of exo- and endo-2-hydroxy derivatives, with low optical purity (Scheme 2.151) [1108]. On the other hand, when Bacillus cereus was used, (2R)-exo- and (2R)-endo-hydroxycineoles were exclusively formed in a ratio of 7:1, both in excellent enantiomeric excess [1109].

Microbial hydroxylation of 1,4-cineole
Among the many hundreds of microorganisms tested for their capability to perform hydroxylation of nonnatural aliphatic compounds, fungi have usually been more often used than bacteria. Among them, the fungus Beauveria bassiana ATCC 7159 (formerly denoted as B. sulfurescens) has been studied most thoroughly [1110–1114]. In general the presence of a polar group in the substrate such as an acetamide, benzamide or p-toluene-sulfonamide moiety seems to be advantageous in order to facilitate the orientation of the substrate in the active site [1115]. Hydroxylation occurs at a distance of 3.3–6.2 Å from the polar anchor group.Footnote 42 In cases where a competition between cycloalkane rings of different size was possible, hydroxylation preferentially occurred in the order cycloheptyl > cyclohexyl > cyclopentyl.
In the majority of cases, hydroxylation by Beauveria bassiana occurs in a regioselective manner, but high enantioselectivity is not always observed. As shown in Scheme 2.152, both enantiomers of the N-benzyl-protected bicyclic lactam are hydroxylated with high regioselectivity in position 11, but the reaction showed very low enantioselectivity. On the other hand, when the lactam moiety was replaced by a more polar benzoyl-amide, which functions as polar anchor group, high enantiodifferentiation occurred. The (1R)-enantiomer was hydroxylated at carbon 12 and the (1S)-counterpart gave the 11-hydroxylated product [1116]. A minor amount of 6-exo-alcohol was formed with low enantiomeric excess.

Regio- and enantioselective hydroxylation by Beauveria bassiana
In order to provide a tool to predict the stereochemical outcome of hydroxylations using Beauveria bassiana, an active site model was proposed [1117, 1118]. Alternatively, a substrate model containing a polar anchor group can be used [1119].
In summary, it is certainly feasible to achieve the (bio)hydroxylation of hydrocarbon compounds by using one of the many microorganisms used to date, but it is difficult to predict the likely site of oxidation for any novel substrate using monooxygenases. However, there are three strategies which can be employed to improve regio- and/or stereoselectivity in biocatalytic hydroxylation procedures:
-
Variation of the culture by stressing the metabolism of the cells
-
Broad screening of different strainsFootnote 43
-
Substrate modification, particularly aiming at the variation of the (polar) anchor group [1120–1123]
2.3.2.3 Hydroxylation of Aromatic Compounds
Regiospecific hydroxylation of aromatic compounds by purely chemical methods is notoriously difficult. There are reagents for o- and p-hydroxylation available [1124, 1125], but some of them are explosive and byproducts are usually obtained [1126]. The selective bio-hydroxylation of aromatics in the o- and p-position to existing substituents can be achieved by using monooxygenases. In contrast, m-hydroxylation is rarely observed [1127]. Mechanistically, it has been proposed that in eukaryotic cells (fungi, yeasts and higher organisms) the reaction proceeds predominantly via epoxidation of the aromatic species which leads to an unstable arene-oxide (Scheme 2.86) [1128]. Rearrangement of the latter involving the migration of a hydride anion (NIH-shift) forms the phenolic product [1129]. An alternative explanation for the catalytic mechanism of flavin-dependent oxidases [which are independent of NAD(P)H] has been proposed which involves a hydroperoxide intermediate (FAD-4a-OOH, Scheme 2.147) [1130].
Phenolic components can be selectively oxidized by polyphenol oxidase – one of the few available isolated oxygenating enzymesFootnote 44 – to give o-hydroxylated products (catechols) in high yields [1131]. Unfortunately the reaction does not stop at this point but proceeds further to form unstable o-quinones, which are prone to polymerization, particularly in water (Scheme 2.153).

o-Hydroxylation of phenols by polyphenol oxidase
Two techniques have been developed to solve the problem of o-quinone instability:
-
One way to prevent polymerization of the o-quinone is to remove it from the reaction mixture by chemical reduction (e.g., by ascorbate), which leads to catechols. Ascorbate, however, like many other reductants can act as an inhibitor of polyphenol oxidase. To circumvent the inhibition, the concentration of the reducing agent should be kept at a minimum. Furthermore, a borate buffer which leads to the formation of a complex with catechols, thus preventing their oxidation, is advantageous [1132].
-
Polymerization, which requires the presence of water, can also be avoided if the reaction is performed in a lipophilic organic solvent such as chloroform (Sect. 3.1) [1133].
The following rules for phenol hydroxylation have been deduced for polyphenol oxidase:
-
A remarkable range of simple phenols are accepted, as long as the substituent R is in the p-position; m- and o-derivatives are unreactive.
-
The reactivity decreases if the nature of the R group is changed from electron-donating to electron-withdrawing.
-
Bulky phenols (p-tert-butylphenol and 1- or 2-naphthols) are not substrates; some electron-rich nonphenolic species such as p-toluidine are accepted.
The synthetic utility of this reaction was demonstrated by the oxidation of amino acids and -alcohols containing an electron-rich p-hydroxyphenyl moiety (Scheme 2.153). Thus, l-DOPA (3,4-dihydroxyphenyl alanine) used for the treatment of Parkinson’s disease, d-3,4-dihydroxy-phenylglycine and l-epinephrine (adrenaline) were synthesized from their p-monohydroxy precursors without racemization in good yield.
Regioselective hydroxylation of aromatic compounds can also be achieved by using whole cells [1134–1138]. For instance, 6-hydroxynicotinic acid is produced from nicotinic acid by Pseudomonas acidovorans or Achromobacter xylosoxidans on a ton-scale [1139]. Racemic prenalterol, a compound with important pharmacological activity as a β-blocker, was obtained by regioselective p-hydroxylation of a simple aromatic precursor using Cunninghamella echinulata (Scheme 2.154) [1140].

Microbial hydroxylation of aromatics
2.3.2.4 Epoxidation of Alkenes
Chiral epoxides are extensively employed high-value intermediates in the synthesis of chiral compounds due to their ability to react with a broad variety of nucleophiles. In recent years a lot of research has been devoted to the development of catalytic methods for their production [551, 1141]. The Katsuki-Sharpless method for the asymmetric epoxidation of allylic alcohols [1142, 1143] and the asymmetric dihydroxylation of alkenes are now widely applied and reliable procedures. Catalysts for the epoxidation of nonfunctionalized olefins have been developed more recently [555, 1144]. Although high selectivities have been achieved for the epoxidation of cis-alkenes, the selectivities achieved with trans- and terminal olefins were less satisfactory using the latter methods.
In contrast, the strength of enzymatic epoxidation, catalyzed by monooxygenases, is in the preparation of small and nonfunctionalized epoxides, where traditional methods are limited [557, 1145]. Despite the wide distribution of monooxygenases within all types of organisms, their capability to epoxidize alkenes seems to be associated mainly with alkane- and alkene-utilizing bacteria, whereas fungi are applicable to a lesser extent [1040, 1146–1150].
Biocatalytic asymmetric epoxidation of alkenes catalyzed by monooxygenases cannot be performed on a preparative scale with isolated enzymes due to their complex nature and their dependence on a redox cofactor, such as NAD(P)H. Thus, whole microbial cells are used instead. Although the toxic effects of the epoxide formed, and its further (undesired) metabolism by the cells catalyzed by epoxide hydrolases (Sect. 2.1.5), can be reduced by employing biphasic media, this method is not trivial and requires bioengineering skills [1151]. Alternatively, the alkene itself can constitute the organic phase into which the product is removed, away from the cells. However, the bulk apolar phase tends to damage the cell membranes, which reduces and eventually abolishes all enzyme activity [1152].
Once the problems of product toxicity were surmounted by sophisticated process engineering, microbial epoxidation of alkenes became also feasible on an industrial scale [1153, 1154]. The latter was achieved by using organic-aqueous two-phase systems or by evaporation for volatile epoxides. For instance, the epoxy-phosphonic acid derivative ‘fosfomycin’ [1155], whose enantiospecific synthesis by classical methods would have been extremely difficult, was obtained by a microbial epoxidation of the corresponding olefinic substrate using Penicillium spinulosum.
The most intensively studied microbial epoxidizing agent is the ω-hydroxylase system of Pseudomonas oleovorans [1156, 1157]. It consists of three protein components: rubredoxin, NADH-dependent rubredoxin reductase and an ω-hydroxylase (a sensitive nonheme iron protein). It catalyzes not only the hydroxylation of aliphatic C–H bonds, but also the epoxidation of alkenes [1158, 1159]. The following rules can be formulated for epoxidations using Pseudomonas oleovorans (Scheme 2.155).

Microbial epoxidation of alkenes
-
Terminal, acyclic alkenes are converted into (R)-1,2-epoxides of high enantiomeric excess along with varying amounts of ω-en-1-ols or 1-als [1160], the ratio of which depends on the chain length of the substrate [1161, 1162]. Alkene-epoxidation occurs mainly with substrates of ‘moderate’ chain length, such as 1-octene. However, alkane hydroxylation predominates over epoxidation for ‘short’ substrates (propene, 1-butene) and is a major pathway for ‘long’-chain olefins.
-
α,ω-Dienes are transformed into the corresponding terminal (R,R)-bis-epoxides.
-
Cyclic, branched and internal olefins, aromatic compounds and alkene units which are conjugated to an aromatic system are not epoxidized [1163].
-
To avoid problems arising from the toxicity of the epoxide [1164] (which accumulates in the cells and reacts with cellular enzymes) a water-immiscible organic cosolvent such as hexane can be added [1165, 1166].
Besides Pseudomonas oleovorans numerous bacteria have been shown to epoxidize alkenes [1167, 1168]. As shown in Scheme 2.155, the optical purity of epoxides depends on the strain used, although the absolute configuration is usually (R) [1169]. This concept has been recently applied to the synthesis of chiral alkyl and aryl gycidyl ethers [1170, 1171]. The latter are of interest for the preparation of enantiopure 3-substituted 1-alkylamino-2-propanols, which are widely used as β-adrenergic receptor-blocking agents [1172].
More recently, the structural restrictions for substrates which were elaborated by Pseudomonas oleovorans (see above) could be lifted by using different microorganisms. As can be seen from Scheme 2.155, nonterminal alkenes can be epoxidized by a Mycobacterium or Xanthobacter sp. [1173]. On the other hand, Nocardia corallina has been reported to convert branched alkenes into the corresponding (R)-epoxides in good optical purities (Scheme 2.156). Aiming at the improvement of the efficiency of microbial epoxidation protocols, a styrene monooxygenase (StyA) and reductase StyB required for electron-transport were co-expressed into E. coli to furnish a designer-bug for the asymmetric epoxidation of styrene-type substrates [1174, 1175].

Epoxidation of styrene derivatives and branched alkenes using cloned monooxygenase and Nocardia corallina
2.3.2.5 Sulfoxidation Reactions
Chiral sulfoxides have been extensively employed as asymmetric auxiliary group that assist stereoselective reactions. The sulfoxide functional group activates adjacent carbon–hydrogen bonds to allow proton abstraction by bases, and the corresponding anions can be alkylated [1176] or acylated [1177] with high diastereoselectivity. Similarly, thermal elimination [1178] and reduction of α-keto sulfoxides [1179] can proceed with transfer of chirality from sulfur to carbon. In spite of this great potential as valuable chiral relay reagents, with rare exceptions [1180], no general method is available for the synthesis of sulfoxides possessing high enantiomeric purities.
An alternative approach involves the use of enzymatic sulfur-oxygenation reactions catalyzed by monooxygenases [1181, 1182]. The main types of enzymatic sulfur oxygenation are shown in Scheme 2.157. The direct oxidation of a thioether by means of a dioxygenase, which directly affords the corresponding sulfone, is of no synthetic use since no generation of chirality is involved. On the other hand, the stepwise oxidation involving a chiral sulfoxide, which is catalyzed by monooxygenases or oxidases,Footnote 45 offers two possible ways of obtaining chiral sulfoxides.

Enzymatic sulfur oxygenation reactions
-
The asymmetric monooxidation of a thioether leading to a chiral sulfoxide resembles a desymmetrization of a prochiral substrate and is therefore of high synthetic value.
-
The kinetic resolution of a racemic sulfoxide during which one enantiomer is oxidized to yield an achiral sulfone is feasible but it has been shown to proceed with low selectivities.
The first asymmetric sulfur oxygenation using cells of Aspergillus niger was reported in the early 1960s [1184]. Since this time it was shown that the enantiomeric excess and the absolute configuration of the sulfoxide not only depend on the species but also on the strain of microorganism used [1185]. In general, the formation of (R)-sulfoxides predominates.
Thioethers can be asymmetrically oxidized both by bacteria (e.g., Corynebacterium equi [1186], Rhodococcus equi [1187]) and fungi (e.g., Helminthosporium sp. [1188] and Mortierella isabellina [1189]). Even baker’s yeast has this capacity [1190, 1191]. As shown in Scheme 2.158, a large variety of aryl-alkyl thioethers were oxidized to yield sulfoxides with good to excellent optical purities [1192–1194]. The undesired second oxidation step was usually negligible, but with certain substrates the undesired formation of the corresponding sulfone was observed.

Microbial oxidation of aryl-alkyl thioethers
The transformation of thioacetals into mono- or bis-sulfoxides presents intriguing stereochemical possibilities. In a symmetric thioacetal of an aldehyde other than formaldehyde, the sulfur atoms are enantiotopic and each of them contains two diastereotopic nonbonded pairs of electrons (Scheme 2.159). Unfortunately, most of the products from asymmetric oxidation of thioacetals are of low to moderate optical purity [1195, 1196]. Two exceptions, however, are worth mentioning. Oxidation of 2-tert-butyl-1,3-dithiane by Helminthosporium sp. gave the (1S,2R)-monosulfoxide in 72% optical purity [1197] and formaldehyde thioacetals were oxidized by Corynebacterium equi to yield (R)-sulfoxide-sulfone products [1198] with excellent enantiomeric purity.

Microbial oxidation of dithioacetals
At present microbial sulfur oxidation reactions are certainly feasible on a preparative scale, but poor recoveries of the water-soluble products from the considerable amounts of biomass (a normal consequence of using whole cells as the biocatalyst) have to be taken into account. Whether the use of isolated oxidases will be an advantage [1199], future investigations will tell. For biocatalytic sulfur oxidation using peroxidase reactions, see Sect. 2.3.3.
2.3.2.6 Baeyer-Villiger Reactions
Oxidation of ketones by peracids – the Baeyer-Villiger reaction [1200, 1201] – is a reliable and useful method for preparing esters or lactones (Scheme 2.160). The mechanism comprises a two-step process, in which the peracid attacks the carbonyl group of the ketone to form the so-called tetrahedral ‘Criegee-intermediate’ [1202]. The fragmentation of this unstable species, which proceeds via expulsion of a carboxylate ion going in hand with migration of a carbon–carbon bond, leads to the formation of an ester or a lactone. The regiochemistry of oxygen insertion of the chemical and the enzymatic Baeyer-Villiger reaction can usually be predicted by assuming that the carbon atom best able to support a positive charge will migrate preferentially [1203].

Mechanism of the chemical and biochemical Baeyer-Villiger oxidation
All mechanistic studies on enzymatic Baeyer-Villiger reactions support the hypothesis that conventional and enzymatic reactions are closely related [1063, 1204]. The oxidized flavin cofactor (FAD-4a-OOH, see Scheme 2.147) plays the role of a nucleophile similar to the peracid. The strength of enzyme-catalyzed Baeyer–Villiger reactions resides in the recognition of chirality [1205–1207], which has been accomplished by conventional means only recently, albeit in reactions exhibiting moderate selectivities [1208].
The enzymatic Baeyer-Villiger oxidation of ketones is catalyzed by flavin-dependent monooxygenases and plays an important role in the breakdown of carbon structures containing a ketone moiety. Whereas the early studies were performed by using whole microbial cells, particularly in view of avoiding the NAD(P)H-recycling problem [1209], an impressive number of bacterial ‘Baeyer-Villigerases’ were cloned, purified and characterized more recently [1210–1213]. In order to facilitate cofactor recycling, a selfsufficient fusion protein consisting of a Baeyer-Villigerase and a phosphite dehydrogenase unit for NADH-recycling were designed [1214]. The overall performance of the fusion-protein was comparable to that of the single (non-fused) proteins.
To avoid rapid further degradation of esters and lactones in microbial Baeyer-Villiger reactions catalyzed by hydrolytic enzymes and to maximize product accumulation in the culture medium, three approaches are possible:
-
Blocking of the hydrolytic enzymes by selective hydrolase-inhibitors such as tetraethyl pyrophosphate (TEPP [1215]) or diethyl p-nitrophenylphosphate (paraoxon). However, all of these inhibitors are highly toxic and have to be handled with extreme caution.
-
Development of mutant strains lacking lactone-hydrolases or
-
Application of nonnatural ketones, whose lactone products are not substrates for the hydrolytic enzymes.
Prochiral (symmetric) ketones can be asymmetrically oxidized by a bacterial cyclohexanone monooxygenase from an Acinetobacter sp. to yield the corresponding lactones [1216, 1217]. As depicted in Scheme 2.161, oxygen insertion occurred on both sides of the ketone depending on the substituent in the 4-position. Whereas in the majority of cases products having the (S)-configuration were obtained, a switch to the (R)-lactone was observed with 4-n-butylcyclohexanone. Simple models are available, which allow the prediction of the stereochemical outcome of Baeyer-Villiger oxidations catalyzed by cyclohexanone monooxygenase of Acinetobacter and Pseudomonas sp. by determination of which group within the Criegee-intermediate is prone to migration [1218, 1219].

Desymmetrization of prochiral ketones via enzymatic Baeyer-Villiger oxidation
Racemic (nonsymmetric) ketones can be resolved via two pathways. The ‘classic’ form of a kinetic resolution involves a transformation in which one enantiomer reacts and its counterpart remains unchanged (Scheme 2.162) [1220]. For example, α-substituted cyclopentanones were stereospecifically oxidized by an Acinetobacter sp. to form the corresponding (S)-configurated δ-lactones [1221], which constitute valuable components of various fruit flavors. The nonconverted (R)-ketones accumulated in the culture medium.

Microbial Baeyer‐Villiger oxidation of monocyclic ketones involving ‘classic’ resolution
Bicyclic haloketones, which were used for the synthesis of antiviral 6′-fluoro-carbocyclic nucleoside analogs, were resolved by using the same technique [1222] (Scheme 2.163). Both enantiomers were obtained with >95% optical purity. The exquisite enantioselectivity of the microbial oxidation is due to the presence of the halogen atoms since the dehalogenated bicyclo[2.2.1]heptan-2-one was transformed with low selectivity. On the other hand, replacement of the halogens by methoxy- or hydroxy groups gave rise to compounds which were not accepted as substrates.

Microbial Baeyer-Villiger oxidation of a bicyclic ketone involving ‘classic’ resolution
The biological Baeyer-Villiger oxidation of a racemic ketone does not have to follow the ‘classic’ kinetic resolution format as described above, but can proceed via a ‘nonclassic’ route involving oxidation of both enantiomers with opposite regioselectivity. Thus, oxygen insertion occurs on the two opposite sides of the ketone at each of the enantiomers. As shown in Scheme 2.164, both enantiomers of the bicyclo[3.2.0]heptenones were microbially oxidized, but in an enantiodivergent manner [1223, 1224]. Oxygen insertion on the (5R)-ketone occurred as expected, adjacent to C7, forming the 3-oxabicyclic lactone. On the other hand, the (5S)-ketone underwent oxygen insertion in the ‘wrong sense’ towards C5, which led to the 2-oxabicyclic species. The synthetic utility of this system has been proven by the large-scale oxidation using an E. coli designer bug harboring cyclohexanone monooxygenase together with a suitabkle NADPH-recycling enzyme [1225, 1226]. In order to minimize product toxicity, in-situ substrate-feeding product removal (SFPR) was applied [1227, 1228].

Enantiodivergent microbial Baeyer-Villiger oxidation involving ‘nonclassic’ resolution
The molecular reasons of enantiodivergent Baeyer-Villiger reactions [1229, 1230] can either be the docking of the substrate in a single enzyme in two opposite modes or due to the presence of different monooxygenases present in the microbial cells [1231].
Whole-cell Baeyer–Villiger oxidations often suffer from low yields due to side reactions catalyzed by competing enzymes in the microorganisms. Furthermore, some of the most potent strains, such as Acinetobacter calcoaceticus, are potentially pathogenic and therefore have to be handled with extra care (see the Appendix, Chap. 5).Footnote 46 On the other hand, the use of isolated monooxygenases for Baeyer–Villiger oxidations is not trivial. On the one hand, there are enzymes available which show a desired opposite stereo- or enantiopreference [1232, 1233], on the contrary, the majority of these enzymes are linked to NADPH-recycling, which is notoriously difficult on a preparative scale. However, a monooxygenase is available from Pseudomonas putida which is dependent on NADH; this cofactor is much more readily recycled [1234].
In order to overcome these problems, a wide range of Baeyer-Villigerases from bacterial origin were cloned into a suitable (nonpathogenic) host, such as baker’s yeast [1235–1239] or E. coli [1240]. Investigation of the regio- and enantioselectivity of Baeyer-Villigerases from bacteria isolated from an industrial wastewater treatment plant cloned into E. coli revealed the following trends (Scheme 2.165): (a) prochiral 4-substituted cyclohexanones underwent desymmetrization yielding (R)- (e.e.max 60%) or (S)-4-alkyl-ε-caprolactones (e.e.max > 99%), depending on the enzyme used and on the size of the substituent. (b) Racemic 2-substituted cyclohexanones underwent ‘classic’ kinetic resolution with absolute regioselectivity for oxygen-insertion at the predicted side to afford enantiomeric pairs of (S)-lactone and unreacted (R)-ketone with excellent enantioselectivities (E ≥200). (c) In contrast, 3-substituted cyclohexanones furnished ‘non-classic’ kinetic resolution via oxygen-insertion at both sides with different regioselectivities to furnish regio-isomeric lactones. The enantioselectivites depended on the enzyme used and on size of the substituent [1241].

Regio- and enantioselective Baeyer-Villiger oxidation using cloned Baeyer-Villigerases exhibiting desymmetrization, and ‘classic’ and ‘nonclassic’ kinetic resolution
An alternative concept of cofactor recycling for isolated monooxygenases was developed using a coupled enzyme system (Scheme 2.166) [1242]. Thus, the substrate ketone is not used as such, but is rather produced by enzymatic oxidation of the corresponding alcohol (at the expense of NADP+ or NAD+, resp., Sect. 2.2.2) using a dehydrogenase from Thermoanaerobium brockii or from Pseudomonas sp. In a second step, the monooxygenase generates the lactone by consuming the reduced cofactor. Therefore, the NAD(P)H is concurrently recycled in a closed loop.

Closed-loop cofactor recycling via a dehydrogenase-oxygenase system
2.3.2.7 Dioxygenases
Bacterial Rieske-type iron dioxygenases are nonheme enzymes, that contain an oxygenase component, an iron-sulfur flavoprotein reductase and FeS ferredoxin [1243, 1244]. Typical dioxygenase reactions, during which two oxygen atoms are simultaneously transferred onto the substrate, are shown in Scheme 2.167. In all cases, a highly reactive and unstable peroxo species is formed, e.g., a hydro- or endoperoxide. Both intermediates are highly reactive and may be subject to further transformations such as (enzymatic or nonenzymatic) reduction or rearrangement [1245]. In general, these latter intermediates cannot be isolated but are immediately reduced to yield the more stable corresponding (di)hydroxy derivatives.

Dioxygenase-catalyzed reactions
-
Alkenes may be oxidized, e.g., by a lipoxygenase, at the allylic position to furnish an allyl hydroperoxide which, upon reduction (e.g., by sodium borohydride) yields an allylic alcohol. In living systems, the formation of lipid peroxides is considered to be involved in some serious diseases and malfunctions including arteriosclerosis and cancer [1246].
-
Alternatively, an endo-peroxide may be formed, whose reduction leads to a diol. The latter reaction resembles the cycloaddition of singlet-oxygen onto an unsaturated system and predominantly occurs in the biosynthesis of prostaglandins and leukotrienes. In prokaryotic cells such as bacteria the initial step of the metabolism of aromatic compounds consists of a (formal) cycloaddition of oxygen catalyzed by a dioxygenase (Scheme 2.86) [1247, 1248]. The resulting intermediate endo-peroxide (dioxetane) is enzymatically reduced to yield synthetically useful cis-glycols (Scheme 2.167).
2.3.2.8 Formation of Peroxides
The biocatalytic formation of hydroperoxides seems to be mainly associated with dioxygenase activity found in plants, such as peas, peanuts, cucumbers, and potatoes as well as marine green algae. Thus, it is not surprising that the (nonnatural) compounds transformed so far have a strong structural resemblance to the natural substrates – fatty acids.
Allylic Hydroperoxidation. Lipoxygenase is a nonheme iron dioxygenase which catalyzes the incorporation of dioxygen into polyunsaturated fatty acids possessing a nonconjugated 1,4-diene unit by forming the corresponding conjugated allylic hydroperoxides [1249–1251]. The enzyme from soybean has received the most attention in terms of a detailed characterization because of its early discovery [1252], ease of isolation and acceptable stability [1253, 1254]. The following characteristics can be given for soybean lipoxygenase-catalyzed oxidations:
-
The specificity of this enzyme has long been thought to be restricted to an all-(Z) configurated 1,4-diene unit at an appropriate location in the carbon chain of polyunsaturated fatty acids. However, it was shown that also (E,Z)- and (Z,E)-1,4-dienes are accepted as substrates [1255], albeit at slower rates.
-
(E,E)-1,4-Dienes and conjugated 1,3-dienes are generally not oxidized.
-
The configuration at the newly formed oxygenated chiral center is predominantly (S), although not exclusively [1256, 1257].
-
In some cases, the oxidation of nonnatural substrates can be forced by increasing the oxygen pressure up to 50 bar [1258].
Oxidation of the natural substrate (Z,Z)-9,12-octadecadienoic acid (linoleic acid) proceeds highly selectively (95% e.e.) and leads to peroxide formation at carbon 13 (the ‘distal’ region) along with traces of 9-oxygenated product [1259] (the ‘proximal’ region, Scheme 2.168) [1260].

Oxidation of linoleic acid by soybean lipoxygenase
In addition, it has been shown that soybean lipoxygenase can also be used for the oxidation of nonnatural 1,4-dienes, as long as the substrate is carefully designed to effectively mimic a fatty acid [1261]. Thus, the (Z,Z)-1,4-diene moiety of several long-chain alcohols could be oxidized by attachment of a prosthetic group (PG), which served as a surrogate of the carboxylate moiety (Scheme 2.169). This group can either consist of a polar (CH2)n–CO2H or a CH2–O–(CH2)2–OH unit [1262]. The oxidation occurred with high regioselectivity at the ‘normal’ (distal) site and the optical purity of the peroxides was >97%. After chemical reduction of the hydroperoxide (e.g., by Ph3P [1263]) and removal of the prosthetic group, the corresponding secondary alcohols were obtained with retention of configuration [1264].

Oxidation of (Z,Z)-1,4-dienes by soybean lipoxygenase
In addition, the regioselectivity of the oxidation – from ‘normal’ (distal) to ‘abnormal’ (proximal) – could be inverted by changing the lipophilicity of the modifying R groups and the spacer arm linking the prosthetic group PG (see Table 2.6 and Scheme 2.169). Increasing the lipophilicity of the distal R group from n-C5 to n-C10 led to an increased reaction at the ‘abnormal’ site to form predominantly the proximal oxidation product. Consequently, when the lipophilicity of the proximal prosthetic group PG was increased by extending the spacer arm, the ‘distal’ product was formed in favor of the proximal.
α-Hydroperoxidation of Carboxylic Acids. The α-oxidation of fatty acids is known for higher plants such as pea leaves, germinating peanuts, cucumbers and potatoes as well for simple organisms such as marine green algae [1265]. The mechanism of this dioxygenase activity was elucidated in the mid-1970s (Scheme 2.170) [1266]. Occasionally, this activity has been denoted as ‘α-oxidase’, however, it should correctly be termed ‘oxygenase’, since an oxidase would produce H2O2 from O2 rather than an α-hydroperoxy fatty acid going in hand with the direct incorporation of O2 into the substrate. It is assumed that the flavoprotein-catalyzed oxidation of the fatty acid leads to an intermediary α-hydroperoxy acid, which can be further metabolized via two competing pathways: Whereas reduction furnishes an α-hydroxyacid (path A), decarboxylation leads to the corresponding aldehyde (path B). It has been speculated that the latter proceeds through an unstable intermediary α-peroxylactone. While the α-hydroxyacid is a final product and can thus be harvested, the aldehyde is further oxidized by an NAD+-dependent aldehyde dehydrogenase to the next lower homologous fatty acid, which in turn can re-enter the α-oxidation in a subsequent cycle. The fact that α-oxidases are membrane-bound has impeded their isolation in pure form, and therefore structure elucidation is still pending. However, it has recently been shown that a crude enzyme extract from germinating pea leaves can be used for the α-hydroxylation of fatty acids and derivatives thereof via path A (Scheme 2.170) [1267, 1268]. The data available so far reveal the following picture:

α-Hydroperoxidation of fatty acid derivatives
-
Substrates should be substantially lipophilic; more hydrophobic compounds, such as short-chain fatty acids and dicarboxylic acids, are not converted.
-
Saturated fatty acids having 7–16 carbon atoms are well accepted, and
-
heteroatoms such as O and S are tolerated.
-
Functional groups, such as carbon–carbon triple- and double-bonds, or heteroatoms have to be at a distance of at least three carbon atoms from the carboxylic acid moiety, otherwise α-hydroxylation does not take place.
-
The ratio between substrate degradation (i.e., decarboxylation, path B) and the desired α-hydroxylation (path A) depends on the substrate structure, but it can be shifted by using optimized reaction conditions.
-
The formation of the α-hydroperoxy acid is virtually absolute for the (R)-enantiomer.
2.3.2.9 Dihydroxylation of Aromatic Compounds
cis-Dihydroxylation by microbial dioxygenases constitutes the key step in the degradation pathway for aromatic compounds in lower organisms (Scheme 2.171) [1269, 1270], which is crucial for the removal of toxic pollutants from contaminated sites, such as oilspills. In ‘wild-type’ microorganisms, the chiral cis-glycols are rapidly further oxidized by dihydrodiol dehydrogenase(s), involving rearomatization of the diol intermediate with concomitant loss of chirality. The use of mutant strains with blocked dehydrogenase activity [1271], however, allows the chiral glycols to accumulate in the medium, from which they can be isolated in good yield [1272, 1273]. The high standard of this technology allow to perform this useful biotransformation on ton-scale [1274].

Degradation of aromatics by microbial dioxygenases
For a number of mutant strains of Pseudomonas putida, the stereospecificity is high although the substrate specificity remains low with respect to the ring substituents R1 and R2 (Scheme 2.172), which allows their use in the asymmetric dihydroxylation of substituted aromatics [1275]. An impressive number of substituted aromatic compounds have been converted into the corresponding chiral cis-glycols, with excellent optical purities, on a commercial scale [1276–1278]. Even polysubstituted benzene derivatives can be converted into cyclohexadienediols and the regioselectivity of the oxygen addition can be predicted with some accurracy using a substrate model (Scheme 2.172) [1279–1281].

Enantiocomplementary synthesis of cis-glycols
The substrates need not necessarily be monosubstituted aromatic compounds such as those shown in Scheme 2.172, but may also be extended to other species including fluoro- [1282], monocyclic- [1283], polycyclic- [1284], and heterocyclic derivatives [1285, 1286].
In order to gain access to products showing opposite configuration, a substrate modification approach using p-substituted benzene derivatives was developed. Thus, when p-iodo derivatives were used instead of the unsubstituted counterparts, the orientation of the oxygen addition was reversed, caused by the switch in relative size of substituents (I > F, I > CH3). Subsequent removal of the iodine (which served as directing group) by catalytic hydrogenation led to mirror-image products [1287].
In a useful extension of the substrate pattern, it was shown that also nonaromatic C=C bonds can be cis-dihydroxylated to yield erythro-diols, as long as they are conjugated to an aromatic system (yielding styrene-derivatives) or to (at least) one additional alkene unit [1288–1290]. In contrast, isolated olefinic bonds are more readily metabolized via an epoxidation – epoxide hydrolysis sequence furnishing the corresponding threo-diols. Thus, Pseudomonas putida harboring toluene dioxygenase or naphthalene dioxygenase was able to oxidize a range of styrene-type alkenes and conjugated di- and -trienes (Scheme 2.173). The stereoselectivities were excellent for cyclic substrates but they dropped for open-chain derivatives (e.e.max 88%) [1291]. Depeding on the substrate and the type of enzyme, hydroxylation at benzylic or allylic positions were observed as side reactions.

Dihydroxylation of conjugated alkenes using toluene dioxygenase
The synthetic potential of nonracemic cis-diols derived via microbial dihydroxylation has been exploited over recent years to synthesize a number of bioactive compounds. Cyclohexanoids have been prepared by making use of the possibility of functionalizing every carbon atom of the glycol in a stereocontrolled way. For instance, (+)-pinitol [1292] and d-myo-inositol derivatives [1293] were obtained using this approach. Cyclopentanoid synthons for the synthesis of prostaglandins and terpenes were prepared by a ring-opening/closure sequence [1294]. Rare carbohydrates such as d- and l-erythrose [1295] and l-ribonolactone [1296] were obtained from chlorobenzene as were pyrrolizidine alkaloids [1297]. Furthermore, a bio-inspired synthesis of the blue pigment indigo was developed on a commercial scale using the microbial dihydroxylation of indol [1298].
2.3.3 Peroxidation Reactions
Driven by the inability to use molecular oxygen as an oxidant efficiently for the transformation of organic compounds, chemists have used it in a partially reduced form – i.e., hydrogen peroxide [1299] or derivatives thereof.Footnote 47 H2O2 offers some significant advantages as it is cheap and environmentally benign – the only byproduct of oxidation being water. However, it is relatively stable and needs to be converted into a more active form in order to become an effective oxidant. This is generally accomplished either with organic or inorganic ‘promoters’ to furnish organic hydro- or endo-peroxides, peroxycarboxylic acids or hypervalent transition metal complexes based on V and Mo. Owing to these drawbacks, the number of industrial-scale oxidation processes using H2O2 as the oxidant is very limited.Footnote 48 On the other hand, biocatalytic activation of H2O2 by peroxidases allow a number of synthetically useful and often highly enantioselective peroxidation reactions, which offer a valuable alternative to traditional chemical methodology.
Peroxidases [EC 1.11.1.7] are a heterogeneous group of redox enzymes found ubiquitously in various sources [1300], such as plants [1301], microorganisms [1302] and animals. They are often named after their sources (e.g., horseradish peroxidase, lacto- and myeloperoxidase) or akin to their substrates (e.g., cytochrome c-, chloro- and lignin peroxidase). Although the biological role of these enzymes is quite diverse – ranging from (1) the scavenging of H2O2, (2) free radical oligomerization and polymerization of electron-rich aromatics to (3) the oxidation and halogenation of organic substrates – they have in common that they accept hydrogen peroxide or a derivative thereof (such as alkyl hydroperoxides) as oxidant. In line with these heterogeneous catalytic activities, the mechanism of action may be quite different and can involve a heme unit, selenium (glutathione peroxidase) [1303], vanadium (bromoperoxidase) [1304, 1305], manganese (manganese peroxidase) [1306] and flavin at the active site (flavoperoxidase) [1307]. The largest group of peroxidases studied so far are heme-enzymes with ferric protoporphyrin IX (protoheme) as the prosthetic group. Their catalytic cycle bears some similarities to that of heme-dependent monooxygenases (Sect. 2.3.2, Scheme 2.146), but owing to the diverse reactions they can catalyze, their pathways are more complex (Scheme 2.174). The mechanism of heme-dependent peroxidase catalysis has been largely deduced from horseradish peroxidase [1043, 1308–1310]. Its most important features are described as follows:

Catalytic cycles of heme-dependent peroxidases
In its native state, the iron-III species is coordinated equatorially by a heme unit and axially by a histidine residue and is therefore very similar to cytochrome P 450 [1311]. The first step in the reaction involves oxidation of the Fe+3 to form an iron-oxo derivative called Compound I. The latter contains a Fe+4=O moiety and a π-radical and is formally two oxidation equivalents above the Fe+3-state.Footnote 49 In a peroxidase, this oxidation is achieved in a single step at the expense of H2O2 (path 1). In the monooxygenase pathway, the Fe3+-species is oxidized by O2, which requires two additional electrons (from a nicotinamide cofactor) to cover the net redox balance. Compound I represents the central hypervalent oxidizing species, which can react along several pathways. Path 2: Abstraction of a single electron from an electron-rich substrate such as an enol or phenol (forming a substrate radical) yields an Fe+4=O species denoted Compound II (path 2a). Since the latter is still one oxidation equivalent above the Fe+3-ground state, this process can occur a second time (forming another substrate radical, path 2b) to finally re-form the enzyme in its native state. Alternatively, incorporation of an O-atom onto a substrate (going hand in hand with a two-electron transfer) can occur in a single step (path 3). In the absence of any substrate, Compound I can re-form the native enzyme via disproportionation of H2O2, denoted as ‘catalase-activity’ (path 4).
Due to the fact that – in contrast to monooxygenases – no external nicotinamide cofactor is involved in any of the peroxidase cycles, peroxidases are highly attractive for preparative biotransformations. A number of synthetically useful reactions can be achieved (Scheme 2.175) [1312–1314].

Synthetically useful peroxidase reactions
2.3.3.1 Oxidative Dehydrogenation
This type of reaction is mainly restricted to heme peroxidases and it involves one-electron transfer processes with radical cations and radicals as intermediates (path 2). As a consequence, substrates are usually electron-rich (hetero)aromatics, which upon one-electron oxidation lead to resonance-stabilized radicals, which spontaneously undergo inter- or intramolecular coupling to form dimers or oligomers. This reaction is commonly denoted as the ‘classical’ peroxidase activity, since it was the first type of peroxidase-reaction discovered. Examples of such reactions are shown in Scheme 2.176. Oxidation of phenols (e.g., guaiacol, resorcin) and anilines (e.g., aniline, o-dianisidine) leads to the formation of oligomers and polymers under mild conditions [1315–1317]. In certain cases, dimers (e.g., aldoximes [1318], biaryls [1319]) have been obtained.

Peroxidase-catalyzed oxidative dehydrogenation of aromatics
2.3.3.2 Oxidative Halogenation
A class of peroxidases specializes in the (per)oxidation of halides (Cl–, Br–, I– but not F–), thus creating reactive halogenating species (such as hypohalite), which in turn form haloorganic compounds [1320, 1321]. These reactions are described in Sect. 2.7.1.
2.3.3.3 Oxygen Transfer
From a synthetic viewpoint, selective oxygen transfer (path 3) is the most interesting peroxidation reaction. The transformations are comparable to those catalyzed by monooxygenases with one significant advantage – they are independent of redox cofactors, such as NAD(P)H.
Among the various types of reactions – C–H bond oxidation, epoxidation of alkenes and heteroatom oxidation – the most useful transformations are described below.
Hydroxylation of C–H Bonds. The only peroxidase known to effect the hydroxylation of C–H bonds is chloroperoxidase (CPO) from the marine fungus Caldariomyces fumago. Its large-scale production is facilitated by the fact that it is an extracellular enzyme, which is excreted into the fermentation medium [1322–1324].Footnote 50 In order to become susceptible towards hydroxylation by CPO, the C–H bonds have to be activated by a π-electron system. In the allylic position, hydroxylation is not very efficient and it is doubtful whether this procedure will be of practical use [1325]. By contrast, benzylic hydroxylation is readily effected and the corresponding sec-alcohols were isolated in high e.e. (Scheme 2.177) [1326, 1327]. Interestingly, in accordance with the above-described mechanism (Scheme 2.178, path 3) oxygen transfer was proven to proceed from hydrogen peroxide via Compound I onto the substrate. CPO is very sensitive with respect to the substrate structure as the stereochemistry of products was reversed from (R) to (S) when the alkyl chain was extended from ethyl to an n-propyl analog.

Products from chloroperoxidase-catalyzed benzylic and propargylic C–H hydroxylations

Asymmetric epoxidation of alkenes using chloroperoxidase
The selectivity of CPO-catalyzed propargylic hydroxylation was found to be sensitive with respect to the chain length of the alkyne substrate [1328]. Thus, whereas short alkyl groups (R1) gave rise to poor enantioselectivity, longer alkyl chains diminished the reaction rate and only medium-chain compounds gave good results. The spatial requirements adjacent to the oxidation site are very strict: whereas compounds bearing methyl groups in position R2 were oxidized with good yields, slow rates were reported for the corresponding ethyl analogs. Hydroxylation of aromatic C–H bonds seems to be possible, as long as electron-rich (hetero)aromatics, such as indol are used [1329, 1330].
Epoxidation of Alkenes. Due to the fact that the asymmetric epoxidation of alkenes using monooxygenase systems is impeded by the requirement for NADPH-recycling and the toxicity of epoxides to microbial cells, the use of H2O2-depending peroxidases represents a valuable alternative.
Unfortunately, direct epoxidation of alkenes by metal-free haloperoxidases led to racemic epoxides [1331, 1332]. Since the reaction only takes place in the presence of a short-chain carboxylic acid (e.g., acetate or propionate), it is believed to proceed via an enzymatically generated peroxycarboxylic acid, which subsequently oxidizes the alkene without the aid of the enzyme. This mechanism has a close analogy to the lipase-catalyzed epoxidation of alkenes (Sect. 3.1.5) and halogenation reactions catalyzed by haloperoxidases (Sect. 2.7.1), where enzyme catalysis is only involved in the formation of a reactive intermediate, which in turn converts the substrate in a spontaneous (nonenzymatic) followup reaction.
By contrast, chloroperoxidase-catalyzed epoxidation of alkenes proceeds with excellent enantioselectivites [1333, 1334]. For styrene oxide it was demonstrated that all the oxygen in the product is derived from hydrogen peroxide, which implies a true oxygen-transfer reaction (path 3, Scheme 2.174) [1335]. As depicted in Scheme 2.178, unfunctionalized cis-alkenes [1336] and 1,1-disubstituted olefins [1337, 1338] were epoxidized with excellent selectivities. On the other hand, aliphatic terminal and trans-1,2-disubstituted alkenes were epoxidized in low yields and moderate enantioselectivities [1339].
Sulfoxidation. Heteroatom oxidation catalyzed by (halo)peroxidases has been observed in a variety of organic compounds. N-Oxidation in amines, for instance, can lead to the formation of the corresponding aliphatic N-oxides or aromatic nitroso or nitro compounds. From a preparative standpoint, however, sulfoxidation of thioethers is of greater importance since it was shown to proceed in a highly stereo- and enantioselective fashion. Moreover, depending on the source of the haloperoxidase, chiral sulfoxides of opposite configuration could be obtained (Scheme 2.179).

Stereocomplementary oxidation of thioethers by haloperoxidases
Chloroperoxidase from Caldariomyces fumago is a selective catalyst for the oxidation of methylthioethers to furnish (R)-sulfoxides. Initial results were disappointing, as low e.e.’s were reported [1340]. The latter were caused by substantial nonenzymatic oxidation by hydrogen peroxide, which could be suppressed by optimization of the reaction conditions: whereas the use of tert-butylhydroperoxide was unsuccessful, the best results were obtained by keeping the concentration of H2O2 at a constant low level [1341, 1342].
Another vanadium-dependent haloperoxidase from the marine alga Corallina officinalis was shown to possess a matching opposite enantiopreference by forming (S)-sulfoxides [1343, 1344]. Although simple open-chain thioethers were not well transformed, cyclic analogs bearing a carboxylic acid moiety in a suitable position within the substrate were ideal candidates [1345].
Reduction of Peroxides. An intriguing alternative for the preparation of enantiopure hydroperoxides makes use of a peroxidase reaction performed in reverse, i.e., by enantioselective reduction (Scheme 2.180) [1346–1348].

Enantioselective reduction of hydroperoxides using horseradish peroxidase
Thus, when (chemically prepared) racemic hydroperoxides were subjected to the action of horseradish peroxidase, one enantiomer was reduced at the expense of o-methoxyphenol used as reductant, to give the corresponding secondary alcohol. The nonreacting enantiomer could be recovered in excellent e.e. The enzyme accepts sterically unencumbered hydroperoxides (with R1 being the ‘large’ and R2 the ‘small’ group), but those bearing sterically demanding branched substituents were unreactive. Of the various isoenzymes which are present in horseradish peroxidase, isoenzyme C is mostly responsible for the reduction [1349]. Similar activity was reported for a number of bacteria and fungi isolated from soil [1350] and for a peroxidase from the basidomycete Coprinus cinereus [1351].
Although peroxidases are more easy to use than monooxygenases, several points concerning their practical application have to be considered:
-
Given the tendency of peroxidases to undergo irreversible deactivation in the presence of substantial concentrations of H2O2 (which is common to many other enzymes) the oxidant has to be added continuously to the reaction in order to keep its concentration at a low level. This also suppresses its decomposition via the catalase activity of peroxidases (path 4, Scheme 2.174). In practice, this is conveniently achieved by using a H2O2-sensitive electrode coupled to an autotitrator which adds the oxidant in a continuous fashion (peroxy-stat) [1352]. Furthermore, this minimizes the spontaneous (background) oxidation, which leads to the formation of racemic product.
-
Depending on the enzyme–substrate combination, the replacement of hydrogen peroxide by tert-butyl hydroperoxide may be beneficial.
-
If the reaction is incompatible with a peroxo species, molecular oxygen in presence of a chemical reductant (such as ascorbic or dihydroxyfumaric acid) may be used as oxidant [1353, 1354].
2.4 Formation of Carbon–Carbon Bonds
The majority of enzymatic reactions exploited to date involve degradation processes via bond-breaking reactions. The following enzymatic systems belonging to the class of lyases, which are capable of forming carbon–carbon bonds in a highly stereoselective manner, are known and are gaining increasing attention in view of their potential in synthesis.
-
Aldol reactions catalyzed by aldolases are useful for the elongation of aldehydes by a two- or three-carbon unit.
-
A C2-fragment is transferred via transketolase reactions or via yeast-mediated acyloin or benzoin condensations.
-
For the addition of cyanide (a C1-synthon) to aldehydes by hydroxynitrile lyases see Sect. 2.5.1. For the sake of simplicity, the donor representing the umpolung reagent is drawn with bold C–C bonds throughout this chapter.
2.4.1 Aldol Reactions
Asymmetric C–C bond formation based on catalytic aldol addition reactions remains one of the most challenging subjects in synthetic organic chemistry. Although many successful nonbiological strategies have been developed [1355, 1356], most of them are not without drawbacks. They are often stoichiometric in auxiliary reagent and require the use of a metal or organocatalytic enolate complex to achieve stereoselectivity [1357–1360]. Due to the instability of such complexes in aqueous solutions, aldol reactions usually must be carried out in organic solvents at low temperature. Thus, for compounds containing polyfunctional polar groups, such as carbohydrates, the employment of conventional aldol reactions requires extensive protection protocols to be carried out in order to make them lipophilic and to avoid undesired cross-reactions. This requirement limits the application of conventional aldol reactions in aqueous solution. On the other hand, enzymatic aldol reactions catalyzed by aldolases, which are performed in aqueous solution at neutral pH, can be achieved without extensive protection methodology and have therefore attracted increasing interest [1361–1374].
Aldolases were first recognized some 70 years ago. At that time, it was believed that they form an ubiquitous class of enzymes that catalyze the interconversion of hexoses into two three-carbon subunits [1375]. It is now known that aldolases operate on a wide range of substrates including carbohydrates, amino acids and hydroxy acids. A variety of enzymes has been described that add a one-, two-, or three-carbon (donor) fragment onto a carbonyl group of an aldehyde or a ketone with high stereospecificity. Since glycolysis and glyconeogenesis are a fundamental pillar of life, almost all organisms possess aldolase enzymes. Two distinct groups of aldolases, using different mechanisms, have been recognized [1376]. Regarding the formation of the (donor) carbanion, both of the mechanisms are closely related to conventional aldol reactions, i.e., carbanion stabilization is achieved via enolate- or enamine species (Schemes 2.181 and 2.182).

Mechanism of type I aldolases

Mechanism of metal-dependent type II aldolases
Type-I aldolases, found predominantly in higher plants and animals, require no metal cofactor, and catalyze the aldol reaction through a Schiff-base intermediate, which tautomerizes to an enamine species (Scheme 2.181) [1377]. First, the donor is covalently linked to the enzyme, presumably via the ε-amino group of a lysine to form a Schiff base. Next, base-catalyzed abstraction of Hs leads to the formation of an enamine species, which performs a nucleophilic attack on the carbonyl group of the aldehydic acceptor in an asymmetric fashion. Consequently, the two new chiral centers are formed stereospecifically in a threo- or erythro-configuration depending on the enzyme. Finally, hydrolysis of the Schiff base liberates the diol and regenerates the enzyme.
Type II aldolases are found predominantly in bacteria and fungi, and are Zn2+-dependent enzymes (Scheme 2.182) [1378]. Their mechanism of action was recently affirmed to proceed through a metal-enolate [1379]: an essential Zn2+ atom in the active site (coordinated by three nitrogen atoms of histidine residues [1380]) binds the donor via the hydroxyl and carbonyl groups. This facilitates pro-(R)-proton abstraction from the donor (presumably by a glutamic acid residue acting as base), rendering an enolate, which launches a nucleophilic attack onto the aldehydic acceptor.
With few exceptions, the stereochemical outcome of the aldol reaction is controlled by the enzyme and does not depend on the substrate structure (or on its stereochemistry). Therefore, the configuration of the carbon atoms adjacent to the newly formed C–C bond is highly predictable. Furthermore, most aldolases are very restricted concerning their donor (the nucleophile), but possess relaxed substrate specificities with respect to the acceptor (the electrophile), which is the carbonyl group of an aldehyde or ketone. This is understandable, bearing in mind that the enzyme has to perform an umpolung on the donor, which is a sophisticated task in an aqueous environment!
To date more than 40 aldolases have been classified, the most useful and more readily available enzymes are described in this chapter. Bearing in mind that the natural substrates of aldolases are carbohydrates, most successful enzyme-catalyzed aldol reactions have been performed with carbohydrate-like (poly)hydroxy compounds as substrates. Depending on the donor, the carbon-chain elongation involves a two- or three-carbon unit (Scheme 2.183, donors are shown in bold).

Main groups of aldolases according to donor type
Aldolases are most conveniently classified into four groups according to their donor molecule. The best studied group I uses dihydroxyacetone or its phosphate (DHAP) as donor, resulting in the formation of a ketose 1-phosphate after reaction with an aldehyde acceptor. Within this group, enzymes capable of forming all four possible stereoisomers of the newly generated stereogenic centers in a complementary fashion are available (Scheme 2.184). Group II uses pyruvate (or phosphoenol pyruvate) as donor to yield 3-deoxy-2-keto acids as products (Scheme 2.193) [1381]. The third group consists of only one enzyme – 2-deoxyribose-5-phosphate aldolase (DERA) – which requires acetaldehyde (or close analogs) as donor to form 2-deoxy aldoses (Scheme 2.194). Finally, group IV aldolases couple glycine (as donor) with an acceptor aldehyde to yield α-amino-β-hydroxy acids (Scheme 2.196).

Stereocomplementary DHAP-dependent aldolases
2.4.1.1 Group I: Dihydroxyacetone or DHA-Phosphate-Dependent Aldolases
The development of the full synthetic potential of DHA(P)-dependent aldolases into a general and efficient methodology for asymmetric aldol additions largely depends on the availability of the complete tetrad of enzymes, which allows to create all four possible stereoisomers at will, by simply selecting the correct biocatalyst.
As shown in Scheme 2.184, all four stereocomplementary aldolases occurring in carbohydrate metabolism which generate the four possible stereoisomeric diol products emerging from the addition of DHAP onto an aldehyde have been made available by cloning and overexpression. The reaction proceeds with complete stereospecificity with respect to the configuration on carbon 3 and also (with slightly decreased specificity) on carbon 4.
Fructose-1,6-Diphosphate Aldolase. Fructose-1,6-diphosphate (FDP) aldolase from rabbit muscle, also commonly known as ‘rabbit muscle aldolase’ (RAMA), catalyzes the addition of dihydroxyacetone phosphate (DHAP) to d-glyceraldehyde-3-phosphate to form fructose-1,6-diphosphate (Scheme 2.185) [514, 1382].

Aldol reactions catalyzed by FDP aldolase from rabbit muscle
The equilibrium of the reaction is predominantly on the product side and the specificity of substituent orientation at C-3 and C-4 (adjacent to the newly formed vicinal diol bond) is absolute – always threo (Scheme 2.185). However, if the α-carbon atom in the aldehyde component is chiral (C-5 in the product), only low chiral recognition of this remote stereocenter takes place. Consequently, if an α-substituted aldehyde is employed in racemic form, a pair of diastereomeric products will be obtained.
RAMA accepts a wide range of aldehydes in place of its natural substrate, allowing the synthesis of carbohydrates [1383–1386] and analogs such as nitrogen-[513, 1387] and sulfur-containing sugars [1388], deoxysugars [1389], fluorosugars, and rare eight- and nine-carbon sugars [1390]. As depicted in Scheme 2.185, numerous aldehydes which are structurally quite unrelated to the natural acceptor substrate (d-glyceraldehyde-3-phosphate) are freely accepted [1391–1394].
For RAMA the following rules apply to the aldehyde component:
-
In general, unhindered aliphatic, α-heterosubstituted, and protected alkoxy aldehydes are accepted as substrates.
-
Sterically hindered aliphatic aldehydes such as pivaldehyde do not react with RAMA, nor do α,β-unsaturated aldehydes or compounds that can readily be eliminated to form α,β-unsaturated aldehydes.
-
Aromatic aldehydes are either poor substrates or are unreactive.
-
ω-Hydroxy acceptors that are phosphorylated at the terminal hydroxyl group are accepted at enhanced rates relative to the nonphosphorylated species.
In contrast to the relaxed specificity for the acceptor, the requirement for DHAP as the donor is much more stringent. Several analogs which are more resistant towards spontaneous hydrolysis have been successfully tested as substitutes for DHAP (Scheme 2.186) [1395], however the reaction rates were reduced by about one order of magnitude [1396–1399].

Nonnatural DHAP substitutes for fructose-1,6-diphosphate aldolase (RAMA)
Within group-I aldolases, FDP aldolase from rabbit muscle has been extensively used for the synthesis of biologically active sugar analogs on a preparative scale (Scheme 2.187). For example, nojirimycin and derivatives thereof, which have been shown to be potent anti-AIDS agents with no cytotoxicity, have been obtained by a chemoenzymatic approach using RAMA in the key step. As expected, the recognition of the α-hydroxy stereocenter in the acceptor aldehyde was low [1400, 1401].

Synthesis of aza-sugar analogs
An elegant synthesis of (+)-exo-brevicomin, the sex pheromone of the bark beetle made use of FDP-aldolase (Scheme 2.188) [1402]. RAMA-catalyzed condensation of DHAP to a δ-keto-aldehyde gave, after enzymatic dephosphorylation, a threo-keto-diol, which was cyclized to form a precursor of the pheromone. Finally, the side chain was modified in four subsequent steps to give (+)-exo-brevicomin.

Synthesis of (+)-exo-brevicomin
Despite the fact that enzymatic aldol reactions are becoming useful in synthetic carbohydrate chemistry, the preparation of aldehyde substrates containing chiral centers remains a problem. Many α-substituted aldehydes racemize in aqueous solution, which would result in the production of a diastereomeric mixture, which is not always readily separable.
The following methods have been used to avoid the (often tedious) separation of diastereomeric products [512].
-
In some cases, a stereoselective aldol reaction can be accomplished in a kinetically controlled process via kinetic resolution of the racemic α-substituted aldehyde. Thus, if the reaction is stopped before it reaches equilibrium, a single diastereomer is predominantly formed. However, as mentioned above, the selectivities of aldolases for such kinetic resolutions involving recognition of the (remote) chirality on the α-carbon atom of the aldehyde are usually low.
-
Efficient kinetic resolution of α-hydroxyaldehydes can be achieved by inserting a negative charge (such as phosphate or carboxylate) at a distance of four to five atoms from the aldehydic center in order to enhance the binding of the acceptor substrate [1403].
-
In cases wherein one diastereomer of the product is more stable than the other, one can utilize a thermodynamically controlled process (Scheme 2.189). For example, in the aldol reaction of rac-2-allyl-3-hydroxypropanal, two diastereomeric products are formed. Due to the hemiacetal ring-formation of the aldol product and because of the reversible nature of the aldol reaction, only the more stable product positioning the 5-allyl substituent in the favorable equatorial position is produced when the reaction reaches equilibrium.
Scheme 2.189 
Thermodynamic control in aldolase reactions
-
Another solution to the problem of formation of diastereomeric products is to subject the mixture to the action of glucose isomerase, whereby the d-ketose is converted into the corresponding d-aldose leaving the l-ketose component unchanged [1404].

A potential limitation on the use of FDP aldolases for the synthesis of monosaccharides is that the products are always ketoses with fixed stereochemistry at the newly generated chiral centers on C-3 and C-4. There are, however, methods for establishing this stereochemistry at other centers and for obtaining aldoses instead of ketoses. This technique makes use of a monoprotected dialdehyde as the acceptor substrate (Scheme 2.190). After the RAMA-catalyzed aldol reaction, the resulting ketone is reduced in a diastereoselective fashion with polyol dehydrogenase. The remaining masked aldehyde is then deprotected to yield a new aldose.

Synthesis of aldoses using FDP aldolase
Aldol additions catalyzed by two DHAP-dependent aldolases which exhibit a complementary stereospecificity to RAMA have been used to a lesser extent (Schemes 2.184 and 2.191) [1405, 1406]. Although the selectivity with respect to the center on carbon 3 is absolute, in some cases the corresponding C-4 diastereomer was formed in minor amounts depending on the R substitutent on the aldehyde. In those cases shown in Scheme 2.191, however, only a single diastereomer was obtained.

Aldol reactions catalyzed by fuculose- and rhamnulose-1-phosphate aldolase
One drawback common to group I aldolase reactions is that most of them them require the expensive and sensitive phosphorylated donor dihydroxyacetone phosphate. This molecule is not very stable in solution (t ~ 20 h at pH 7), and its synthesis is not trivial [1407]. DHAP may be obtained from the hemiacetal dimer of dihydroxyacetone by chemical phosphorylation with POCl3 [1408, 1409], or by enzymatic phosphorylation of dihydroxyacetone at the expense of ATP and glycerol kinase [546] (Sect. 2.1.4). Probably the most elegant and convenient method is the in situ generation of DHAP from fructose-1,6-diphosphate (FDP) using FDP aldolase, forming one molecule of DHAP as well as glyceraldehyde-3-phosphate. The latter can be rearranged by triosephosphate isomerase to give a second DHAP molecule [541]. This protocol has been further extended into a highly integrated ‘artificial metabolism’ to obtain DHAP from inexpensive feedstocks, such as glucose or fructose (yielding two equivalents of DHAP) and sucrose (four equivalents) via an enzymatic cascade consisting of up to seven enzymes [1410].
The presence of the phosphate group in the aldol adducts facilitates their purification by ion-exchange chromatography or by precipitation as the corresponding barium salts. Cleavage of phosphate esters is usually accomplished by enzymatic hydrolysis using acid or alkaline phosphatase (Sect. 2.1.4).
Attempts to use a mixture of dihydroxyacetone and a small amount of inorganic arsenate, which spontaneously forms dihydroxyacetone arsenate which is a mimic of DHAP and is accepted by FDP aldolase as a substrate, were impeded by the toxicity of arsemate [1411, 1412]. However, the use of borate ester mimics of DHAP offers a valuable nontoxic alternative for preparative-scale reactions [1413].
An elegant solution to avoid the neccessity for DHAP is the use of a recently discovered bacterial transaldolase, which catalyzes the aldol rection between glyceraldehyde-3-phosphate and (nonphosphorylated) dihydroxyacetone forming fructose-6-phosphate. Since its stereospecificity is identical to that of fructose-1,6-diphosphate aldolase, it represents a useful extension of the aldolase toolbox (Scheme 2.184) [1414–1417]. As depicted in Scheme 2.192, coupling of dihydroxyacetone with an N-protected 3-aminopropanal gave the (3S,4R)-threo-diol, which was reductively cyclized to furnish the rare aza-sugar d-fagomine, which acts as glycosidase inhibitor and shows antifungal and antibacterial activity [1418].

Aldol reaction catalyzed by dihydroxyacetone-dependent fructose-6-phosphate aldolase
2.4.1.2 Group II: Pyruvate-Dependent Aldolases
For thermodynamic reasons, pyruvate-dependent aldolases have catabolic functions in vivo, whereas their counterparts employing (energy-rich) phosphoenol pyruvate as the donor are involved in the biosynthesis of keto-acids. However, both types of enzymes can be used to synthesize α-keto-β-hydroxy acids in vitro.
Sialic Acid Aldolase. N-Acetylneuraminic acid (NeuAc, also termed sialic acid) aldolase catalyzes the reversible addition of pyruvate onto N-acetylmannosamine to form N-acetylneuraminic acid (Scheme 2.193) [1419, 1420]. Since the equilibrium for this reaction is near unity, an excess of pyruvate must be used in synthetic reactions to drive the reaction towards completion. NeuAc was previously isolated from natural sources such as cow’s milk, but increasing demand prompted the development of a two-step synthesis from N-acetylglucosamine using chemical or enzymatic epimerization to N-acetylmannosamine, followed by coupling of pyruvate catalyzed by sialic acid aldolase on a multi-ton scale [1421–1424]. Besides NeuAc, the production of structural analogs is of significance since neuraminic acid derivatives play an important role in cell adhesion and biochemical recognition processes [1425]. The cloning of the enzyme has reduced its cost [1426].

Industrial-scale synthesis of N-acetylneuraminic acid using a two-enzyme system and aldol reactions catalyzed by sialic acid aldolase
In line with the substrate requirements of FDP aldolase, the specificity of sialic acid aldolase appears to be absolute for pyruvate (the donor), but relaxed for the aldehydic acceptor. As may be seen from Scheme 2.193, a range of mannosamine derivatives have been used to synthesize derivatives of NeuAc [1427–1432]. Substitution at C-2 of N-acetylmannosamine is tolerated, and the enzyme exhibits only a slight preference for defined stereochemistry at other centers.
Other group II aldolases of preparative value are 3-deoxy-d-manno-octulosonate (KDOFootnote 51) aldolase [1433, 1434], 2-keto-3-deoxy-6-phosphogalactonate aldolase [1381, 1435] and 3-deoxy-d-arabino-heptulosonate-7-phosphate (DAHP) synthetase [1436].
2.4.1.3 Group III: Acetaldehyde-Dependent Aldolases
2-Deoxyribose-5-Phosphate Aldolase. The only aldolase known so far which accepts acetaldehyde as donor is 2-deoxyribose-5-phosphate (DER) aldolase. In vivo, DER aldolase catalyzes the reversible aldol reaction of acetaldehyde and d-glyceraldehyde-3-phosphate to form 2-deoxyribose-5-phosphate. This aldolase is unique in that it is the only aldolase that condenses two aldehydes (instead of a ketone and an aldehyde) to form aldoses (Schemes 2.183 and 2.194).Footnote 52 Interestingly, the enzyme (which has been overproduced [1438]) shows a relaxed substrate specificity not only on the acceptor side, but also on the donor side. Thus, besides acetaldehyde it accepts also acetone, fluoroacetone and propionaldehyde as donors, albeit at a much slower rate. Like other aldolases, it transforms a variety of aldehydic acceptors in addition to d-glyceraldehyde-3-phosphate.

Aldol reactions catalyzed by 2-deoxyribose-5-phosphate aldolase
An elegant method for sequential aldol reactions performed in a one-pot reaction has been discovered for 2-deoxyribose-5-phosphate aldolase (Scheme 2.195) [1439]. When a (substituted) aldehyde was used as acceptor, condensation of acetaldehyde (as donor) led to the corresponding β-hydroxy aldehyde as intermediate product. The latter, however, can undergo a second aldol reaction with another acetaldehyde donor, forming a β,δ-dihydroxy aldehyde. At this stage, this aldol cascade (which would lead to the formation of a polymeric product if uninterrupted) is terminated by the (spontaneous) formation of a stable hemiacetal. The latter does not possess a free aldehydic group and therefore cannot serve as acceptor any more.

Sequential aldol reactions catalyzed by DER aldolase
The dihydroxylactols thus obtained can be oxidized by NaOCl to the corresponding lactones, which represent the chiral side chains of several cholesterol-lowering 3-hydroxy-3-methylglutaryl-(HMG)-CoA reductase inhibitors, which are collectively denoted as ‘statins’ [1440].Footnote 53 Several derivaties thereof are produced on industrial scale using DER-aldolase mutants at product concentrations exceeding 100g/L [1441, 1442].
This concept provides rapid access to polyfunctional complex products from cheap starting materials in a one-pot reaction. It has recently been extended by combining various types of aldolases together to perform three- and four-substrate cascade reactions [1443, 1444].
2.4.1.4 Group IV: Glycine-Dependent Aldolases
One remarkable feature of group IV aldolases is their requirement for an amino acid as donor – glycine (Scheme 2.196) [1445, 1446]. Thus, α-amino-β-hydroxy acids are formed during the course of the reaction.

Aldol reactions catalyzed by l-threonine aldolase
d - and l -Threonine Aldolase. These enzymes are involved in the biosynthesis/degradation of α-amino-β-hydroxyamino acids, such as threonine and they usually show absolute specificity for the α-amino-configuration, but have only low specificities for the β-hydroxy-center, thereby leading to diastereomeric threo/erythro-mixtures of products [1447]. For biocatalytic applications, several threonine aldolases show broad substrate tolerance for various acceptor aldehydes [1448, 1449], including aromatic aldehydes; however, α,β-unsaturated aldehydes were not accepted. β-Hydroxyamino acids constitute an important class of natural products of their own right and are components of complex structures, such as the β-hydroxytyrosine moiety in vancomycin. It is thus not surprising, that threonine aldolases have been frequently used for their synthesis.
For instance, the l-enzyme from Candida humicola was used in the synthesis of multifunctional α-amino-β-hydroxy acids, which possess interesting biological properties (Scheme 2.196) [1449]. A number of benzyloxy- and alkyloxy aldehydes were found to be good acceptors. Although the stereoselectivity of the newly generated α-center was absolute (providing only l-amino acids), the selectivity for the β-position bearing the hydroxyl group was less pronounced, leading to threo- and erythro-configurated products. More recently, the use of a recombinant d-threonine aldolase was reported [1450, 1451].
Unfortunately, the position of the equilibrium does not favor synthesis, which requires to push the reaction by employing either an excess of the donor glycine (which is difficult to separate from the product) or the acceptor aldehyde (which at high concentrations may deactive the enzyme). A recently developed protocol relies on pulling of the equilibrium by (irreversible) decarboxylation of the formed α-amino-β-hydroxycarboxylic acid catalyzed by a decarboxylase to yield the corresponding aminoalcohols as final products [1452].
2.4.2 Thiamine-Dependent Acyloin and Benzoin Reactions
In the aldol reaction, C–C coupling always takes place in a head-to-tail fashion between the umpoled Cα atom of an enolate- or enamine-species (acting as donor) and the carbonyl C of an acceptor forming a β-hydroxycarbonyl product. In contrast, head-to-head coupling of two aldehydic species involving both carbonyl C atoms would lead to α-hydroxycarbonyl compounds, such as acyloins or benzoins. For this reaction, one aldehyde has to undergo umpolung at the carbonyl C, which is accomplished with the aid of an intriguing cofactor: thiamine diphosphate (ThDP, Scheme 2.197) [1453–1456]. This cofactor is an essential element for the formation/cleavage of C–C, C–N, C–O, C–P, and C–S bonds and plays a vital role as vitamin B1 [1457–1459]. A schematic representation of the mechanism of enzymatic carboligation by ThDP-dependent enzymes is depicted in Scheme 2.197. In a first step, ThDP is deprotonated at the iminium carbon, leading to a resonance-stabilized carbanion. The latter performs a nucleophilic attack on an aldehyde (R1–CH=O), which is converted into the donor by forming a covalently bound carbinol species bearing a negative charge. This umpoled species attacks the second (acceptor) aldehyde (R2–CH=O) going in hand with C–C bond formation. Tautomerization of the diolate intermediate goes in hand with release of the α-ketol (acyloin/benzoin) product and regeneration of the cofactor.

Thiamine diphosphate-dependent carboligation of aldehydes
In the enzymatic aldol reaction, the role of the donor and acceptor is strictly determined by the specificity of the enzyme and only one coupling product can be obtained. In contrast, the possible product range is more complex in acyloin and benzoin reactions: If only a single aldehyde species is used as substrate, only one product can be obtained via homocoupling; however, a pair of regioisomeric α-hydroxyketones can be obtained via heterocoupling, when two different aldehydes are used, the ratio of which is determined by the choice of substrates (e.g., benzoyl formate vs. benzaldehyde, pyruvate vs. acetaldehyde), and the specificites of enzymes, respectively (Scheme 2.198).

Regioisomeric α-hydroxyketones obtained from homo- and heterocoupling of aldehydes
Stereocontrol in mixed acyloin and benzoin reactions is high only if the carboligation encompasses at least one (large) aromatic aldehyde, whereas with two (small) aliphatic aldehydes only moderate e.e.s are generally obtained.
2.4.2.1 Acyloin and Benzoin Reactions
Historically, the biocatalytic acyloin condensation was first observed by Liebig in 1913 during studies on baker’s yeast [1460]. A few years later, Neuberg and Hirsch reported the formation of 3-hydroxy-3-phenylpropan-2-one (phenyl acetyl carbinol, PAC) from benzaldehyde by fermenting baker’s yeast [1461]. Without knowledge on the actual enzyme(s) involved, this biotransformation assumed early industrial importance when it was shown that the acyloin thus obtained could be converted into (–)-ephedrine by diastereoselective reductive amination, a process which is still utilized in almost unchanged form at a capacity of 120 t/year [1462, 1463] (Scheme 2.199). Subsequent studies revealed that this yeast-based protocol can be extended to a broad range of aldehydes [1464, 1465].

Synthesis of (–)-ephedrine via baker’s yeast catalyzed acyloin reaction and acyloin formation catalyzed by pyruvate decarboxylase
Despite its important history, it was during the early 1990s, that the reaction pathway was elucidated in detail [1466] and it turned out that the enzyme responsible for this reaction is pyruvate decarboxylase (PDC) [1467]. The C2-unit (equivalent to acetaldehyde) originates from the decarboxylation of pyruvate and is transferred to the si-face of the aldehydic substrate to form an (R)-α-hydroxyketone (acyloin) with the aid of the cofactor TDP [1468]. Since pyruvate decarboxylase accepts α-ketoacids other than pyruvate, C2- through C4-equivalents can be transferred onto a large variety of aldehydes [1469–1471]. In whole-cell (yeast) transformations, the resulting acyloin is often reduced in a subsequent step by yeast alcohol dehydrogenase to give the erythro-diol. The latter reaction is a common feature of baker’s yeast whose stereochemistry is guided by Prelog’s rule (see Sect. 2.2.3, Scheme 2.118). The optical purity of the diols is usually better than 90% [1472–1476].
It must be mentioned, however, that for baker’s yeast-catalyzed acyloin reactions the yields of chiral diols are usually in the range of 10–35%, but this is offset by the ease of the reaction and the low price of the reagents used. Depending on the substrate structure, the reduction of the aldehyde to give the corresponding primary alcohol (catalyzed by yeast alcohol dehydrogenases, see Sect. 2.2.3) and saturation of the α,β-double bond (catalyzed by ene-reductases, see Sect. 2.2.4) are the major competing reactions. To avoid low yields associated with yeast-catalyzed transformations, acyloin- [1477–1479] and benzoin-reactions [1480] are nowadays performed using isolated enzymes [1481].
2.4.2.2 Pyruvate Decarboxylase
In vivo, pyruvate decarboxylase [EC 4.1.1.1] catalyzes the nonoxidative decarboxylation of pyruvate to acetaldehyde and is thus a key enzyme in the fermentative production of ethanol. The most well-studied PDCs are obtained from baker’s yeast [1477, 1482, 1483] and from Zymomonas mobilis [1484].
From a synthetic viewpoint, however, its carboligation activity is more important [1485–1487]: All PDCs investigated so far prefer small aliphatic aldehydes as donors, used either directly or applied in the form of the respective α-ketocarboxylic acids. The latter are decarboxylated during the course of the reaction, which drives the equilibrium towards carboligation. Straight-chain α-ketoacids up to C-6 are good donors, whereas branched and aryl-aliphatic analogs are less suitable. On the acceptor side, aromatic aldehydes are preferred, leading to PAC-type regioisomeric acyloins (Schemes 2.199 and 2.200). Self-condensation of small aldehydes yielding acetoin-type products may occur.

Regiocomplementary carboligation of aldehydes catalyzed by pyruvate and benzoylformate decarboxylase
2.4.2.3 Benzoylformate Decarboxylase
BFD [EC 4.1.1.7] is derived from mandelate catabolism, where it catalyzes the nonoxidative decarboxylation of benzoyl formate to yield benzaldehyde. Again, the reverse carboligation reaction is more important [1488–1490]. As may be deduced from its natural substrate, is exhibits a strong preference for large aldehydes as donor substrates encompassing a broad range of aromatic, heteroaromatic, cyclic aliphatic and olefinic aldehydes [1480]. With acetaldehyde as acceptor, it yields the complementary regio-isomeric product to PDC (Scheme 2.200).
2.4.2.4 Benzaldehyde Lyase
Up to now, the only biochemically characterized benzaldehyde lyase (BAL) [EC 4.1.2.38] was derived from a Pseudomonas fluorescens strain, which was isolated from wood, showing the ability to grow on lignin-degradation products, such as anisoin (4,4′-dimethoxybenzoin) and benzoin. BAL cleaves these latter compounds to furnish more simple aromatic aldehydes [1491]. In contrast to PDC and BFD, BAL shows only negligible decarboxylation activity and C–C lyase- and carboligation are dominant [1492–1494]. Especially the self-ligation of benzaldehyde yields benzoin with high activity and stereoselectivity (e.e. >99), making this enzyme very interesting for industrial processes [1495]. For benzoin formation, o-, m-, and p-substituted aromatic aldehydes are widely accepted as donors [1496]. Heterocoupling of aromatic and aliphatic aldehydes (acting as acceptor) result in the formation of (R)-2-hydroxypropiophenone derivatives in analogy to BFD. On the acceptor side, formaldehyde, acetaldehyde and close derivatives, such as phenyl-, mono-, or dimethoxyacetaldehyde are tolerated.
The remarkable synthetic potential of BAL is demonstrated by the regiocomplementary benzoin condensation of α,β-unsaturated aldehydes acting as donor or acceptor, respectively. While large aldehydes acted as donors (product type A), small counterparts served as acceptors leading to isomeric olefinic acyloins B in high e.e.s [1497] (Scheme 2.201).

Regiocomplementary carboligation of aldehydes catalyzed by benzaldehyde lyase
2.4.2.5 Transketolase
In the oxidative pentose phosphate pathway, ThDP-dependent transketolaseFootnote 54 catalyzes the reversible transfer of a 2-carbon hydroxyacetaldehyde-unit from a ketose phosphate (d-xylulose-5-phosphate) to an aldose (d-ribose-5-phosphate) (Scheme 2.202) [1498]. Its mechanism resembles a classical acyloin condensation mediated by ThDP. Although the substrate specificity of transketolase has not been fully explored, it appears to be a promising catalyst for use in synthesis [1499, 1500]. Fortunately, the natural phosphorylated substrate(s) can be replaced by hydroxypyruvate [1501], which is decarboxyled to furnish a hydroxyacetaldehyde unit thereby driving the reaction towards completion. The C-2 fragment is transferred onto an aldehyde acceptor yielding an acyloin possessing a threo-diol configuration. This method has allowed the synthesis of a number of monosaccharides on a preparative scale [1502–1504].

Acyloin reactions catalyzed by transketolase
Transketolases from various sources have been shown to possess a broad acceptor spectrum yielding products with complete (S)-stereospecificity for the newly formed stereocenter [1505]. Generic aldehydes are usually converted with full stereocontrol and even α,β-unsaturated aldehydes are accepted to some degree. However, hydroxylated aldehydes show enhanced rates by mimicking the natural substrate [1499]. Transketolases can be obtained from yeast [1506] and spinach [1507] and their overexpression has opened the way for large-scale production [1508, 1509].
Interestingly, transketolase recognizes chirality in the aldehydic acceptor moiety to a greater extent than the aldolases. Thus, when (stereochemically stable) racemic α-hydroxyaldehydes are employed as acceptors, an efficient kinetic resolution of the α-center is achieved (Scheme 2.202). Only the (αR)-enantiomer is transformed into the corresponding keto-triol leaving the (αS)-counterpart behind [1510]. In a related manner, when (±)-3-azido-2-hydroxypropionaldehyde was chosen as acceptor, only the d-(R)-isomer reacted and the l-(S)-enantiomer remained unchanged [1401].
The broad synthetic potential ThDP-dependent enzymes for asymmetric C–C bond formation is by far not fully exploited with the acyloin- and benzoin-condensations discussed above. On the one hand, novel branched-chain α-keto-acid decarboxylases favorably extend the limited substrate tolerance of traditional enzymes, such as PDC, by accepting sterically hindered α-ketoacids as donors [1511]. On the other hand, the acceptor range may be significantly widened by using carbonyl compounds other than aldehydes: Thus, ketones, α-ketoacids and even CO2 lead to novel types of products (Scheme 2.203).

Future potential of thiamine-dependent C–C bond formation
2.4.3 Michael-Type Additions
During an acyloin condensation mediated by baker’s yeast, a side reaction involving a Michael-type addition was observed when trifluoroethanol was added to the medium (Scheme 2.204) [1512]. Although the exact mechanism of this reaction has not yet been elucidated, it presumably proceeds via the following sequence: In a first step, trifluoroethanol is enzymatically oxidized (by yeast alcohol dehydrogenase) to trifluoroacetaldehyde, which is converted into its anion equivalent by ThDP. Instead of undergoing an acyloin reaction (resembling a 1,2-addition), the latter is added in a 1,4-addition mode across the C=C double bond of the α,β-unsaturated carbonyl compound to form trifluoromethyl ketones as intermediates. In baker’s yeast, these are stereoselectively reduced by dehydrogenase(s) to form chiral trifluoromethyl carbinols or the corresponding lactones, respectively. Absolute configurations were not specified, but are probably (R) as predicted by Prelog’s rule.

Michael addition catalyzed by baker’s yeast
Stereospecific Michael addition reactions also may be catalyzed by hydrolytic enzymes (Scheme 2.205). When α-trifluoromethyl propenoic acid was subjected to the action of various proteases, lipases and esterases in the presence of a nucleophile (NuH), such as water, amines, and thiols, chiral propanoic acids were obtained in moderate optical purity [1513]. The reaction mechanism probably involves the formation of an acyl enzyme intermediate (Sect. 2.1.1, Scheme 2.1). Being an activated derivative, the latter is more electrophilic than the ‘free’ carboxylate and undergoes an asymmetric Michael addition by the nucleophile, directed by the chiral environment of the enzyme. In contrast to these observations made with crude hydrolase preparations, the rational design of a ‘Michaelase’ from a lipase-scaffold gave disappointingly low stereoselectivities [1514–1517].

Asymmetric Michael addition catalyzed by hydrolytic enzymes
2.5 Addition and Elimination Reactions
Among the various types of transformations used in organic synthesis, addition reactions are the ‘cleanest’ since two components are combined into a single product with 100% atom efficiency [1518, 1519].
The asymmetric addition of small molecules such as hydrogen cyanide onto C=O bonds or water and ammonia across C=C bonds is typically catalyzed by lyases. During such a reaction one or (depending on the substitution pattern of the substrate) two chiral centers are created from a prochiral substrate.
2.5.1 Cyanohydrin Formation
Hydroxynitrile lyase enzymes catalyze the asymmetric addition of hydrogen cyanide onto a carbonyl group of an aldehyde or a ketone thus forming a chiral cyanohydrin [1520–1524],Footnote 55 a reaction which was used for the first time as long ago as 1908 [1525]. Cyanohydrins are rarely used as products per se, but they represent versatile starting materials for the synthesis of several types of compounds [1526]:
Chiral cyanohydrins serve as the alcohol moieties of several commercial pyrethroid insecticides (see below) [1527]. Hydrolysis or alcoholysis of the nitrile group affords chiral α-hydroxyacids or -esters and Grignard reactions provide acyloins [1528], which in turn can be reduced to give vicinal diols [1529]. Alternatively, the cyanohydrins can be subjected to reductive amination to afford chiral ethanolamines [1530]. α-Aminonitriles as well as aziridines are obtained via the corresponding α-sulfonyloxy nitriles [1531].
Since only a single enantiomer is produced during the reaction – representing a desymmetrization of a prochiral substrate – the availability of different enzymes of opposite stereochemical preference is of importance to gain access to both (R)- and (S)-cyanohydrins (Scheme 2.207). Fortunately, an impressive number of hydroxynitrile lyases of opposite stereochemical preference can be isolated from cyanogenic plants [1532–1536].
(R)-Specific enzymes are obtained predominantly from the Rosacea family (almond, plum, cherry, apricot) and they have been thoroughly investigated [1537–1540]. They contain FAD in its oxidized form as a prosthetic group located near (but not in) the active site, but this moiety does not participate in catalysis and seems to be an evolutionary relict.
On the other hand, (S)-hydroxynitrile lyases [1541–1544] were found in Sorghum bicolor [1545] (millet), Hevea brasiliensis [1546, 1547] (rubber tree), Ximenia americana [1548] (sandalwood), Sambucus niger [1549] (elder), Manihot esculenta [1543, 1550] (cassava), flax, and clover. They do not contain FAD and they exhibit a more narrow substrate tolerance, as aliphatic aldehydes are not always accepted. Furthermore, the reaction rates and optical purities are sometimes lower than those which are obtained when the (R)-enzyme is used. Based on X-ray structures [1551, 1552], the mechanism of enzymatic cyanohydrin formation has been elucidated as follows (Scheme 2.206) [1553]: The substrate is positioned in the active site with its carbonyl group bound through network of hydrogen bonds involving His/Cys/Tyr or Ser/Thr-moieties, while the lipophilic residue is accommodated in a hydrophobic pocket. Nucleophilic addition of cyanide anion occurs from opposide sides from cyanide-binding pockets, which are made of positively charged Arg/Lys- or His/Lys-residues [364].

Schematic representation of (R)- and (S)-hydroxynitrile formation by HNLs from almond and Hevea brasiliensis, respectively
The following set of rules for the substrate-acceptance of (R)-hydroxynitrile lyase was delineated [1554].
-
The best substrates are aromatic aldehydes, which may be substituted in the meta- or para-position; also heteroaromatics such as furan and thiophene derivatives are well accepted [1555–1558].
-
Straight-chain aliphatic aldehydes and α,β-unsaturated aldehydes are transformed as long as they are not longer than six carbon atoms; the α-position may be substituted with a methyl group.
-
Methyl ketones are transformed into cyanohydrins [1559], while ethyl ketones are impeded by low yields [1560].
-
For large or sterically demanding aldehydes, such as o-chlorobenzaldehyde, (R)-HNL mutants possessing a more spacious active site were constructed [1561, 1562]. The (R)-o-chloromandelonitrile thus obtained represents the chiral core of the blockbuster clopidogrel (Plavix) (Scheme 2.207).
Scheme 2.207 
Stereocomplenentary asymmetric cyanohydrin formation
Table 2.7 Synthesis of (R)-cyanohydrins from aldehydes and ketones

The (S)-hydroxynitrile lyase from Hevea brasiliensis has been made available in sufficient quantities by cloning and overexpression to allow industrial-scale applications [1563]. It should be noted that also α,β-unsaturated aliphatic aldehydes were transformed into the corresponding cyanohydrins in a clean reaction. No formation of saturated β-cyano aldehydes through Michael-type addition of hydrogen cyanide across the C=C double bond occurred. The latter is a common side reaction using traditional methodology.
Of particular interest is the industrial-scale synthesis of the (S)-configured cyanohydrin from m-phenoxybenzaldehyde (Table 2.8), which is an important intermediate for synthetic pyrethroids.
Synthetic pyrethroids comprise a class of potent insecticides with structural similarities to a number of naturally occurring chrysanthemic acid esters found in the extract of pyrethrum flowers (Chrysanthemum cinerariaefolium). These natural products constitute highly potent insecticides, but their instability (inherent to the cyclopentenone moiety) precludes their broad application in agriculture. This fact has led to the development of a range of closely related analogs, which retain the high insecticidal activity of their natural ancestors but are more stable (Scheme 2.208). All of these synthetic pyrethroids contain asymmetric carbon atoms and it is well established that their insecticidal activity resides predominantly in one particular isomer. In order to reduce the environmental burden during pest control, single isomers are marketed [1564].

Natural and synthetic pyrethroids
Two particular problems which are often encountered in hydroxynitrile lyase-catalyzed reactions are the spontaneous nonenzymatic formation of racemic cyanohydrin and racemization of the product due to equilibration of the reaction. As a result, the optical purity of the product is decreased. Bearing in mind that both the chemical formation and the racemization of cyanohydrins are pH-dependent and require water, three different techniques have been developed in order to suppress the reduction of the optical purity of the product.
-
Adjusting the pH of the medium to a value below 3.5, which is the lower operational pH-limit for most hydroxynitrile lyases.
-
Lowering the water-activity of the medium [1565] by using water-miscible organic cosolvents such as ethanol or methanol. Alternatively, the reaction can be carried out in a biphasic aqueous-organic system or in a monophasic organic solvent (e.g., ethyl acetate, di-i-propyl, or methyl t-butyl ether) which contains only traces of water to preserve the enzyme’s activity.
-
In order to avoid the use of hazardous hydrogen cyanide, trans-cyanation reactions were developed using either acetone cyanohydrin [1566] or (±)-2-pentanone cyanohydrin [1567] as donor for hydrogen cyanide. The latter are considerably more easy to handle. Using this technique, the competing chemical cyanohydrin formation is negligible due to the low concentration of free hydrogen cyanide and the use of free hydrogen cyanide is avoided.
A fascinating variant of the enzymatic cyanohydrin formation consists in the use of nitroalkanes (as nonnatural nucleophiles) instead of cyanide (Scheme 2.209) [1568, 1569]. Overall, this constitutes a biocatalytic equivalent to the Henry-reaction producing vicinal nitro-alcohols, which are valuable precursors for amino alcohols. Using (S)-HNL, the asymmetric addition of nitromethane to benzaldehyde gave the nitroalcohol in 92% e.e., while for p-nitrobenzaldehyde the stereoselectivity dropped sharply. With nitroethane, two stereocenters are created: Whereas the stereoselectivity for the alcoholic center was high (e.e. 95%), the recognition for the adjacent center bearing the nitro moiety was modest and other (dia)stereomers were formed in up to 8%.

Asymmetric Henry-reaction catalyzed by (S)-hydroxynitrile lyase
2.5.2 Addition of Water
The asymmetric addition of water or ammonia onto olefins is one of the ‘dream-reactions’ on organic synthesis and represents one of the (largely unsolved) problems of catalysis. Enzymes called hydratases or ammonia lyases can catalyze this reaction. However, they only act on activated alkenes, such as α,β-unsaturated carboxylic acids, and their substrate tolerance is rather narrow and only allow minor structural variations of their natural substrate(s), which severely limits their application in organic synthesis.
Fumarase [EC 4.2.1.2] and malease [EC 4.2.1.31] catalyze the stereospecific addition of water onto carbon–carbon double bonds conjugated with a carboxylic acid [1570]. The analogous addition of ammonia is catalyzed by aspartase [EC 4.3.1.1], 3-methylaspartase [1571], and phenylalamine ammonia lyase [EC 4.3.1.5].
Both reactions are mechanistically related and take place in a trans/anti-manner [1572, 1573], with protonation occurring from the re-side (Scheme 2.210). These close mechanistic similarities may be explained by the fact that some of these enzymes show a remarkable degree of amino acid homology [1574]. Within this group of enzymes, substrate tolerance is rather narrow, but the stereoselectivities observed are exceptionally high.

Lyase-catalyzed addition of water and ammonia onto activated C=C bonds
The addition of water onto fumaric acid leads to malic acid derivatives (Scheme 2.211). Only fumarate and chlorofumaric acid are well accepted; the corresponding (sterically more encumbered) bromo-, iodo-, and methyl derivatives are transformed at exceedingly low rates, albeit with excellent stereoselectivities. Fluoro- and 2,3-difluorofumaric acid are accepted but their transformation suffers from decomposition reactions of the first-formed fluoromalic acid. Replacement of one of the carboxylic groups or changing the stereochemistry of the double bond from (E) to (Z) is not tolerated by the enzyme [1575]. The analogous hydration of the stereoisomeric (Z)-isomer (maleic acid) produces the mirror-image (R)-malate and is catalyed by malease (maleate hydratase) [1576, 1577]. The latter enzyme also accepts 2-methylmaleate (citraconate) to form (R)-2-hydroxy-2-methylsuccinate (2-methylmaleate) [1578]. The industrial-scale production of (S)-malate, which is used as an acidulant in fruit juices, carbonated soft drinks, and candies, is performed using the above described hydratases at a capacity of up to 2,000 t/year [1579, 1580].

Fumarase-catalyzed addition of water
In contrast to the hydration of highly activated olefinic diacids, α,β-unsaturated monocarboxylic acids have to be activated via a thioester linkage onto the cofactor Coenzyme A. The latter is catalyzed by an enoyl-CoA synthetase and requires ATP as energy source. The enoyl-CoA intermediate is hydrated by an enoyl-CoA hydratase yielding the corresponding β-hydroxyacyl-CoA as product, which is finally hydrolyzed by a thioesterase to liberate the β-hydroxycarboxylic acid and CoA, which re-enters the catalytic cycle. Due to the complexity of this multienzyme-system requiring ATP and CoA, hydration of acrylic acid derivatives is always performed using whole cells [1274, 1581, 1582].
An elegant example for this biotransformation is the asymmetric hydration of crotonobetaine yielding the ‘nutraceutical’ (R)-carnitine (Scheme 2.212), which is used as an additive in baby food, geriatric nutrition and health sport. In order to avoid the undesired degradation of the product, mutant strains lacking carnitine dehydrogenase have been developed at a capacity of >100 t/year [1583–1585].

Asymmetric hydration of crotonobetaine to carnitine via a multienzyme-system
An unexpected reaction catalyzed by baker’s yeast with potential synthetic utility was observed as a side reaction during an attempt to asymmetrically reduce substituted crotonaldehyde derivatives (Scheme 2.213). Thus, a lyase-catalyzed addition of water occurred in presence of a 4-benzyloxy- (X = 2 H) or benzoyloxy substituent (X = O) in the substrate [1474, 1586].

Addition of water by fermenting baker’s yeast
The capacity of microbial cells of different origin to perform an asymmetric hydration of C=C bonds has only been poorly investigated but they show a promising synthetic potential. For instance, Fusarium solani cells are capable of hydrating the ‘inner’ (E)-double bond of terpene alcohols (e.g., nerolidol) or -ketones (e.g., geranyl acetone) in a highly selective manner [1587]. However, side reactions such as hydroxylation, ketone-reduction, or degradation of the carbon skeleton represent a major drawback. On the other hand, resting cells of Rhodococcus rhodochrous catalyzed the asymmetric addition of water onto the C=C bond of α,β-unsaturated butyrolactones with high enantioselectivity, furnishing β-hydroxylactones in moderate yields [1588].
2.5.3 Addition of Ammonia
Enzymic amination of fumaric acid using aspartase leads to the formation of l-aspartic acid, which is performed at a capacity of 1,200 t/year (Scheme 2.214) [1589–1592]. Although aspartase is one of the most specific enzymes known by accepting only its natural substrate [1593–1595], some variations concerning the nucleophile are tolerated: Hydroxylamine, hydrazine, methoxyamine, and methylamine are accepted and furnish the corresponding l-aspartate N-derivatives [1596–1598].

Amination of fumarate derivatives by aspartase and 3-methylaspartase
In contrast to aspartase, some structural variations are tolerated by the related 3-methylaspartase (Scheme 2.214). For instance, the methyl group in the natural substrate may be replaced by a chlorine atom or by small alkyl moieties [1599], but the fluoro- and the iodo-analog (X = F, I) are not good substrates. Although the bromo-derivative is accepted, it irreversibly inhibits the enzyme [1600].
In a related fashion, asymmetric amination of (E)-cinnamic acid yields l-phenylalanine using l-phenylalanine ammonia lyase [EC 4.3.1.5] at a capacity of 10,000 t/year [1274, 1601]. A fascinating variant of this biotransformation consists in the use of phenylalanine aminomutase from Taxus chinensis (yew tree), which interconverts α- to β-phenylalanine in the biochemical route leading to the side chain of taxol [1602]. In contrast to the majority of the cofactor-independent C–O and C–N lyases discussed above, its activity depends on the protein-derived internal cofactor 5-methylene-3,5-dihydroimidazol-4-one (MIO) [1603]. Since the reversible α,β-isomerization proceeds via (E)-cinnamic acid as achiral intermediate, the latter can be used as substrate for the amination reaction. Most remarkably, the ratio of α- vs. β-amino acid produced (which is 1:1 for the natural substrate, R = H) strongly depends on the type and the position of substituents on the aryl moiety: While o-substituents favor the formation of α-phenylalanine derivatives, p-substituted substrates predominantly lead to β-amino analogs. A gradual switch between both pathways occurred with m-substituted compounds. With few exceptions, the stereoselectivity remained excellent (Scheme 2.215) [1604, 1605].

Formation of α- and β-phenylalanine derivatives using phenylalanine ammonia mutase
2.6 Transfer Reactions
2.6.1 Glycosyl Transfer Reactions
Oligosaccharides and polysaccharides are important classes of naturally occurring compounds [1606]. They play a vital role in intracellular migration and secretion of glycoproteins, cell–cell interactions, oncogenesis, and interaction of cell surfaces with pathogens [1607–1609]. The building blocks are monosaccharides which (theoretically) occur in an enormous number of stereoisomers, which results in a structural diversity far greater than that possible with peptides of comparabale size [1610].Footnote 56 Fortunately, Nature is using almost exclusively pentoses and hexoses for in vivo synthesis.
The ready availability of such oligosaccharides of well-defined structure is critical for the synthesis of drug candidates. Isolation of these materials from natural sources is a complex task and is not economical on a large scale due to the low concentration of the structures of interest in the complex mixtures of carbohydrates obtained from natural sources. Chemical synthesis of complex oligosaccharides is one of the greatest challenges facing synthetic organic chemistry since it requires many protection and deprotection steps which result in low overall yields [1611]. Moreover, stereospecific chemical synthesis of oligosaccharides, in particular of the important α-sialylated structures, is difficult. In this context, biocatalysts are attractive as they allow the regio- and stereospecific synthesis of oligosaccharides with a minimum of protection and deprotection steps [1363–1365, 1612–1618]. There are two groups of enzymes which can be used for the synthesis of oligosaccharides.
Glycosyl transferases are the biocatalysts which are responsible for the biosynthesis of oligosaccharides in vivo. They require that the sugar monomer which is to be linked to the growing oligosaccharide chain is activated by phosphorylation prior to the condensation step. The activating group on the anomeric center, a leaving group (LG, Scheme 2.216), is either a nucleoside phosphate (usually a diphosphate in the Leloir pathway [1619]) or a simple phosphate (in non-Leloir pathway enzymes [1620]). Glycosyl transferases are highly specific with respect to their substrate(s) and the nature of the glycosidic bond to be formed.

Synthesis of oligosaccharides by glycosyl transferases
Glycosidases belong to the class of hydrolytic enzymes and have a catabolic function in vivo as they hydrolyze glycosidic linkages to form mono- or oligosaccharides from polysugars. Consequently, they are generally less specific when compared to glycosyl transferases. Both of these types of enzymes can be used for the synthesis of oligosaccharides and related compounds.
2.6.1.1 Glycosyl Transferases
Three fundamental steps constitute the glycoside-bond formation in the Leloir pathway: activation, transfer and modification [1621]. These steps represent biological solutions to the problems also faced by chemists, i.e., chemical activation of sugars, regio- and stereospecific formation of glycosidic linkages and final elaboration of the products. In the first step, a sugar is phosphorylated by a kinase to give a sugar-1-phosphate. This activated sugar subsequently reacts with a nucleoside triphosphate (usually uridine triphosphate, UTP) under catalysis of a nucleoside transferase and forms a chemically activated nucleoside diphosphate sugar (NDP, Scheme 2.216). These key nucleoside diphosphate sugars constitute the activated ‘donors’ in the subsequent condensation with the ‘acceptors’ (the hydroxyl group of a mono- or oligosaccharide, a protein or a lipid). The latter step is catalyzed by a glycosyl transferase. To ensure proper functioning of the cell, a large number of highly specific glycosyl transferases seem to be neccessary, since each NDP-sugar requires a distinct group of glycosyl transferases.
More than one hundred glycosyl transferases have been identified to date and each one appears to specifically catalyze the formation of a unique glycosidic linkage [1622].
Chemists apply enzymes of the Leloir pathway to the synthesis of oligosaccharides [1623–1627]. Two requirements are critical for the success of this approach, namely the availability of the sugar nucleoside phosphates at reasonable costs and the availability of the matching glycosyl transferases. Although the first issue is being resolved for most of the common NDP-sugars utilizing phosphorylating enzymes (Sect. 2.1.4), the supply of cheap glycosyl transferases remains a problem. Only a few of these enzymes are commercially available; because isolation of these membrane-bound (unstable) proteins is difficult, since they are present only in low concentrations [1628]. In this context, genetic engineering has a major impact by making glycosyl transferases more stable and readily available. Since the membrane-unbound (soluble) portion of glycosyl transferases alone (containing the catalytic domain) is stable and fully active, one promising possibility is to express only this part in engineered microorganisms [1629]. Another disadvantage associated with glycosyl transferase reactions is that the reactions often exhibit coproduct inhibition caused by the released nucleoside phosphate (LG-OH). A simple solution to these problems is to keep their concentration at a low level by in-situ regeneration of the sugar nucleotide from the released nucleoside phosphate (Scheme 2.217).

Synthesis of N-acetyllactosamine using a six-enzyme system (Leloir pathway)
Enzymatic methods [1630] of preparing the NDP-sugars have several advantages over the chemical methods [1631]. For example, the NDP-sugar may be generated in situ (e.g., by epimerization of UDP-Glc to UDP-Gal), making it possible to drive unfavorable equilibria in the required direction. Purification steps may be eliminated because the byproducts of enzyme-catalyzed methods do not interfere with further enzymatic steps.
The point of interest to synthetic chemists is the range of acceptors and donors that can be used in glycosyl transferase-catalyzed reactions. Fortunately, the specificity of glycosyl transferases is high but not absolute.
UDP-galactosyl (UDP-Gal) transferase is the best-studied transferase in terms of specificity for the acceptor sugar. It has been demonstrated that this enzyme catalyzes the transfer of UDP-Gal to a remarkable range of acceptor substrates of the carbohydrate-type [1622, 1632–1635]. Other glycosyl transferases, although less well-studied than UDP-Gal transferase, also appear to tolerate various acceptors as substrates [1636–1639] (Table 2.9).
The use of the multienzyme systems, which arise due to the need to prepare the activated UDP-sugar in situ, is exemplified with the synthesis of N-acetyllactosamine [1640] (Scheme 2.217). Glucose-6-phosphate is isomerized to its 1-phosphate by phosphoglucomutase. Transfer of the activating group (UDP) from UTP is catalyzed by UDP-glucose pyrophosphorylase liberating pyrophosphate, which is destroyed by inorganic pyrophosphatase. Then, the center at carbon 4 is epimerized by UDP-galactose epimerase in order to drive the process out of the equilibrium. Finally, using galactosyl transferase, UDP-galactose is linked to N-acetylglucosamine to yield N-acetyllactosamine.
The liberated UDP is recycled back to the respective triphosphate by pyruvate kinase at the expense of phosphoenol pyruvate. The overall yield of this sequence was in the range of 70% when performed on a scale greater than 10 g.
More recently, attention has been drawn to oligosaccharide synthesis via non-Leloir routes (Scheme 2.218). In this case, the activated donor is a more simple sugar-1-phosphate, which can be transferred by a single sugar phosphorylase. The latter catalyze the reversible cleavage/formation of a glycosyl bond using phosphate as nucleophile/leaving group, respectively [1641, 1642]. Depending on their mode of action, they belong to the group of glycosyl transferases or glycosidases [1643]. For example, trehalose, one of the major storage carbohydrates in plants, fungi, and insects, was synthesized from glucose and its 1-phosphate using trehalose phosphorylase as the catalyst [1644].

Synthesis of trehalose via the non-Leloir pathway
Glycoside synthesis becomes even more facile using sucrose phosphorylase. This bacterial transglucosidase catalyzes the cleavage of the disaccharide sucrose using phosphate as nucleophile to yield α-d-glucose-1-phosphate and d-fructose (Scheme 2.219). In the absence of phosphate, the enzyme-glucosyl intermediate can be intercepted by various nucleophiles bearing an alcoholic group to yield the corresponding α-d-glucosides in high yields [1645, 1646]. Aryl alcohols and polyhydroxylated compounds, such as sugars and sugar alcohols are often glycosylated in a highly selective fashion. The major advantage of this system is the weak hydrolase activity of sucrose phosphorylase and the high-energy content of the cheap glucosyl donor sucrose. Several of these products constitute biocompatible solutes, which regulate the water-balance of the cell, prevent protein denaturation and stabilize membranes and are thus used as natural osmolytes and moisturising agents [1647].

Synthesis of α-D-glycosides using sucrose phosphorylase
2.6.1.2 Glycosidases
In the breakdown of carbohydrates, glycosidases (also termed ‘glycohydrolases’ [1648]) cleave glycosidic bonds. Since they are true hydrolases they are independent of any cofactor. Two groups of them exist: exo-glycosidases only cleave terminal sugar residues, whereas endo-glycosidases are also able to split a carbohydrate chain in the middle. In general, glycosidases show high (but not absolute) specificity for both the glycosyl moiety and the nature of the glycosidic linkage, but little if any specificity for the aglycone component which acts as a leaving group ([LG-H], Scheme 2.220) [1649]. It has long been recognized that the nucleophile (NuH, which is water in the ‘normal’ hydrolytic pathway) can be replaced by other nucleophiles, such as another sugar or a primary or secondary (nonnatural) acceptor alcohol. This allows to turn the degradative nature of glycosyl hydrolysis towards the more useful synthetic direction [1612, 1650–1654]. Interestingly, this potential was already recognized as early as 1913! [1655].

Glycoside synthesis using glycosidases
Major advantages of glycosidase-catalyzed glycosyl transfer are that there is minimal (or zero) need for protection and that the stereochemistry at the newly formed anomeric center can be completely controlled through the choice of the appropriate enzyme, i.e., an α- or β-glucosidase. However, regiocontrol with respect to the acceptor remains a problem, particularly when mono- or oligosaccharides carrying multiple hydroxy groups are involved.
Depending on the stereochemical course of glycoside formation, i.e., whether retention or inversion of the configuration at the anomeric center is observed, glycosidases operate via two separate and distinct mechanisms (Schemes 2.221, 2.222) [1656–1660]. Examples of the retaining enzymes are β-galactosidase, invertase and lysozyme. Inverting glycosidases, such as trehalase and β-amylase, have been used for the synthesis of alkyl glycosides to a lesser extent.

Mechanism of retaining glycosidases
Although the first proposal for the mechanism of retaining glycosidases in 1953 has undergone some refinements, it is still valid in its sense (Scheme 2.221) [572, 1661, 1662]: The active site contains two glutamic acid residues (Glu1 and Glu2), which can act as an acid or a base, respectively. In the first step, Glu1 acts as an acid by protonation of the anomeric oxygen, making the (oligo)saccharide moiety [RO] a good leaving group, while the glycosyl residue is bound to the enzyme via Glu2 as oxonium ion [1663, 1664]. Then, the leaving group ROH is displaced by the incoming nucleophile NuH (usually water) via diffusion. In the second step, the nucleophile is deprotonated by Glu1 and the glycosyl-enzyme intermediate from attacks from same face from which the leaving group R-OH was expelled. Since both steps constitute an SN2-reaction, double inversion results in net retention of configuration.
In contrast, inverting glycosidases act via a single step: Direct nucleophilic displacement of the aglycone moiety (ROH) by a nucleophile (NuH) via SN2 leads to inversion of anomeric configuration (Scheme 2.222).

Mechanism of inverting glycosidases
Reverse Hydrolysis
Glycosidases can be used for the synthesis of glycosides in two modes. The thermodynamic approach is the reversal of glycoside hydrolysis by shifting the equilibrium of the reaction from hydrolysis to synthesis. This procedure uses a free (nonactivated) monosaccharide as substrate and it has been referred to as ‘direct glycosylation’ or ‘reverse hydrolysis’ (Fig. 2.19, pathway A) [1665–1668]. Since in an aqueous environment the equilibrium constant for this reaction lies strongly in favor of hydrolysis, high concentrations of both the monosaccharide and the nucleophilic component (carbohydrate or alcohol) must be used. As a consequence, yields in these reactions are generally low and reaction mixtures comprised of thick syrups up to 75% by weight are not amenable to scale-up.

Glycosylation via thermodynamic and kinetic control
Other methods to improve such procedures make use of aqueous-organic two-phase systems [1669, 1670] and polyethylene-glycol-modified glycosidases [1671]. However, the direct enzymatic synthesis of alkyl glycosides is generally hampered by the low solubility of carbohydrates in organic media. More polar solvents, such as DMF, DMSO or pyridine, would be self-defeating, considering that the products are often intended for use in food and personal care products. Alternatively, the reaction can be performed at temperatures below 0°C or the glycoside formed can be removed from the reaction medium by selective adsorption [1672]. In summary, glycoside synthesis via the reverse hydrolysis approach is less than ideal.
Transglycosylation
The second route – the kinetic approach – utilizes a preformed activated glycoside, which is coupled onto the nucleophile by an appropriate glycosidase and is referred to as ‘transglycosylation’ (Fig. 2.19, pathway B) [1673, 1674]. The enzyme-glycoside intermediate is then trapped by a nucleophile other than water to yield a new glycoside. In this case, activated glycosyl donors which possess an aglycone moiety with good leaving group properties, i.e., poor nucleophilicity, are used [1675, 1676]. Good donors are, for instance, glycosyl fluorides [537, 1677, 1678], -azides [1679, 1680], (hetero)aryl- (usually p-nitrophenyl- or nitropyridyl- [1681]), vinyl- and allyl-glycosides [1682, 1683]. Transglycosylation gives higher yields as compared to reverse hydrolysis and is generally the method of choice [1684, 1685]. Since the glycoside formed during the reaction is also a substrate for the enzyme in hydrolysis causing its degradation (Fig. 2.19, pathway C), the success of this procedure as a preparative method depends on the following crucial parameters:
-
Transglycosylation must be faster than glycoside hydrolysis
-
The rate of hydrolysis of the product being slower than that of the glycosyl donor
In practice these conditions can be attained readily. It should be emphasized that an analogous situation can be found in enzymatic peptide synthesis using proteases (Sect. 3.1.4). The primary advantages of using glycosidases in comparison to glycosyl transferases is that expensive activated sugar nucleosides are not required and glycosidases generally are more readily available than glycosyl transferases. Furthermore, there is total control over the configuration at the newly generated anomeric center.
The major drawbacks, however, are low yields and the frequent formation of product mixtures due to the limited selectivity of glycosidases with respect to the glycosidic acceptor, in particular due to the formation of undesired 1,6-linkages. The regio- and stereoselectivity of transglycosylation reactions is influenced by a number of parameters such as reaction temperature [1686], concentration of organic cosolvent, the reactivity of the activated donor [1687], the nature of the aglycone [1688, 1689], and the anomeric configuration of the acceptor glycoside [1690] (Table 2.10).
This latter fact has been used as a convenient tool to modulate the regioselectivity of glycosylation by switching the configuration at the anomeric center of the glycosidic acceptor. This technique has been commonly denoted as ‘anomeric control’ (Scheme 2.223).

Anomeric control in N-acetylglucosaminyl transfer onto α- and β-d-methylglucosides by β-galactosidase
For instance, when the α-anomer of methyl-d-glucoside was used as acceptor and p-nitrophenyl-β-N-acetyl-d-galactosaminide as donor in a trans-glycosylation reaction catalyzed by β-galactosidase from Aspergillus oryzae, two transfer products possessing a 1,4- and 1,6-linkage were formed in a ratio of ~5:1, respectively. On the other hand, when using the β-anomer of the acceptor, the corresponding 1,3- and 1,4-glucosides were formed instead (ratio ~4:1) [1691].
Besides the synthesis of natural glycosides, a considerable number of nonnatural alcohols have been employed as nucleophiles in transglycosylation reactions (Tables 2.11 and 2.12) [1692, 1693]. The types of transformation include the desymmetrization of meso-diols and the kinetic resolution of racemic primary and secondary alcohols. In discussing enantioselection towards a (chiral) nonnatural acceptor, it should be kept in mind that the donor carbohydrate moiety is chiral and, as a consequence, the glycosylation products are diastereomers rather than enantiomers. In general, the stereocontrol during desymmetrization of prochiral or the kinetic resolution of racemic alcohols by glycosidases performs much worse than, e.g., lipases and alcohol dehydrogenases.
(Bi)cyclic meso-1,2- or 1,3-diols have been transformed into the corresponding monoglycosides in moderate to good diastereoselectivity using β-galactosidase from Escherichia coli, which is readily available from the dairy industry. As may be seen from Table 2.11, the selectivity strongly depends on the structure of the aglycone component [1694].
In some cases, the kinetic resolution of racemic primary and secondary alcohols was feasible. On the one hand, the enantioselectivity of glycosidases involving the glycosylation of primary alcohol moieties in 1,2-propanediol, glycerol or glycidol was negligible [1695, 1696], however, better results were obtained for sec-alcohols (Table 2.12). This fact is understandable if one considers the rules for chiral recognition for carboxyl ester hydrolases (Sects. 2.1.3.1 and 2.1.3.2), where the distance of the center of chirality to the point of reaction should be a minimum. Since the preferentially formed (diastereomeric) product is also the better substrate for (undesired) hydrolysis, the exact determination of the enantioselection in kinetic resolutions catalyzed by glycosidases is a complex task. As may be deduced from Table 2.12, the enantioselective glycosylation of short-chain sec-alcohols gave low to moderate d.e.’s. On the other hand, better results were obtained when both of the stereogenic groups significantly differed in size [1697–1699]. In recent years, a number of thermostable glycosidases have been identified and characterized. The most remarkable among them are the β-glucosidase [1700] and the β-galactosidase from the hyperthermophilic archean Pyrococcus furiosus.
A major improvement in the use of glycosidases for glycoside synthesis was the rational re-design of the catalytic site to disable the undesired hydrolysis of the glycoside product, while maintaining glycoside synthesis activity (Scheme 2.224). Replacement of the Glu2-residue acting as base in the native enzyme by a Ser residue allowed to bind an activated glycosyl fluoride as donor. The latter is attacked by the acceptor nucleophile, which is deprotonated by Glu1, forming the glycoside product. In the native enzyme, the latter would undergo subsequent hydrolysis by a water molecule activated by Glu2 but this is impossible in the Ser-mutant. Such active-site mutants of glycosidases (aptly denoted as ‘glycosynthases’ [1701–1706]) show greatly enhanced yields of glycosides due to the elimination of their undesired hydrolysis.

Mechanistic principle of an inverting glycosynthase
2.6.2 Amino Transfer Reactions
Transaminases (also termed amino transferases [EC 2.6.1.X]) catalyze the redox-neutral amino-transfer reaction between an amine donor and a carbonyl group serving as acceptor (Scheme 2.225) [94, 1707–1712]. These enzymes require an ‘activated benzaldehyde’ (pyridoxal-5′-phosphate, PLP, vitamin B6) as cofactor, which functions as a molecular shuttle for the transfer of the NH3-moiety. In a first step, PLP forms a ketimine Schiff base with the amine-donor. Tautomerization of the C=N bond yields an aldimine, which is hydrolyzed to yield the cofactor in its aminated form (pyridoxamine, PMP). The latter reacts through the same order of events with the carbonyl group of the substrate to form the amine product and regeneration of PLP. Since the tightly bound cofactor needs no external regeneration system, asymmetric transamination has the potential for large-scale synthesis of nonracemic amines.

Transaminase-catalyzed amino-transfer
Although transaminases were discovered already half a century ago [1713, 1714], their use for the biotransformation of (nonnatural) substrates was mainly impeded by two obstacles: The majority of transaminases available were only active on α-amino/α-ketoacids as substrates and techniques to shift the equilibrium towards amine formation had to be developed. The first significant advances in transamination for organic synthesis were achieved by Celgene Co., who employed transaminases for the preparation of nonracemic amines, preferentially via (less efficient) kinetic resolution of rac-amines via enantioselective de-amination [1715, 1716]. Within the last decade, several breakthroughs with respect to the (commercial) availability of stereo-complementary transaminases possessing a broad substrate spectrum and a set of techniques to shift the equilibrium of transamination in favor of amine synthesis were accomplished, which make enzymatic transamination nowadays a reliable technique for the asymmetric synthesis of amines.
On a genomic level, transaminases are classified into (up to six) subgroups [1717, 1718], among which group II comprises the most useful transaminases, which are able to accept non-α-amino acid type substrates. Since their natural substrates typically encompass ω-aminocarboxylic acids, such as ornithine, lysine, β-alanine, and ω-aminobutyrate, they are commonly denoted as ω-transaminases (ω-TA), in contrast α-transaminases, which only act on α-amino acid-type substrates.
In view to access both stereoisomers of a chiral amine via transamination by choice of an appropriate (R)- or (S)-selective ω-TA, screening studies were undertaken which revealed an impressive number of (S)-ω-TAs and some more rarely occurring (R)-selective enzymes [1719–1722]. The most widely used enzymes are obtained from Vibrio fluvialis [1723], Chromobacterium violaceum [1724, 1725], Pseudomonas aeruginosa [1726], Bacillus megaterium [1727], and Alcaligenes denitrificans [1728]. Thermostable mutants were derived from an ω-TA from Arthrobacter citreus [1729].
Depending on the substrate preference of the employed transaminase, the following couples of sacrificial amine donor/keto acceptors were used, which are often derived from the α-aminoacid pool (such as alanine/pyruvate, phenylalanine/phenylpyruvate, glutamic acid/α-ketoglutarate, aspartic acid/α-ketosuccinate) or constitute simple amines/ketones, such as 2-propylamine/acetone and 2-butylamine/2-butanone. It should be kept in mind that the absolute configuration of a chiral amine-donor has to match the stereospecificity of the ω-TA in order to be accepted.
In transamination, equilibrium constants are close to unity at best and the amino transfer from an α-amino acid to a ketone is strongly disfavored.Footnote 57 To even worsen the situation, ω-TAs often show cosubstrate and/or coproduct inhibition at elevated concentrations, which prevents to push amine formation by employing an excess of amine donor. In order to solve this problem, several strategies have been developed (Scheme 2.226):

Shifting the equilibrium in transamination
-
The most simple approach is the physical removal of the coproduct ketone by evaporation. However, this is only feasible for simple amine-donors, such as 2-propylamine or 2-butylamine, which lead to highly volatile ketones [1730, 1731].
-
The use of an amine donor, which forms an unstable keto coproduct. For instance, cysteinesulfinic acid was used in transamination to furnish the β-sulfinic acid analog of pyruvate, which spontaneously decomposes into SO2 and pyruvate [1732]. In a related fashion, α,ω-diamino acids, such as ornithine or leucine yield amino-ketoacids, which (nonenzymatically) cyclize to the corresponding Δ2-pyrroline-5-carboxylate and Δ1-piperidine-2-carboxylate, respectively, as dead-end products [1733, 1734].
-
Removal of the coproduct via an additional enzymatic step is usually more effective: For instance, decarboxylation of an α-ketoacid (e.g., pyruvate or phenylpyruvate, formed from alanine or phenylalanine, respectively) using pyruvate or phenylpyruvate decarboxylase, yields an aldehyde and CO2 [1735, 1736]. In a similar fashion, pyruvate may be removed by condensation to acetoin going in hand with decarboxylation catalyzed by acetolactate synthase [1737].
-
Carbonyl-reduction of the keto-coproduct by a suitable dehydrogenase, such as alcohol dehydrogenase or lactate dehydrogenase, yields the corresponding alcohol or α-hydroxyacid and requires NAD(P)H-recycling [1738].
-
The most efficient approach is probably the direct recycling of alanine from pyruvate via NADH-dependent reduction in presence of ammonia catalyzed by alanine dehydrogenase. Overall, this sequence resembles a metal-free asymmetric reductive amination, which only requires ammonia and a low-cost reducing agent for NAD(P)H-recycling in molar amounts [1739].
The availability of a broad variety of ω-TAs together with efficient techniques to shift the equilibrium allows the biocatalytic synthesis of amines from the corresponding ketones via amino-group transfer. The potential of this protocol is demonstrated by a selection of amines, which can be obtained in nonracemic form by using the most prominent ω-transaminases (Scheme 2.227).

Representative substrate spectrum of selected ω-transaminases
2.7 Halogenation and Dehalogenation Reactions
Halogen-containing compounds are not only produced by man, but also by Nature [1740–1742]. A brominated indole derivative – Tyrian purple dye – was isolated from the mollusc Murex brandaris by the Phoenicians. Since that time, more than 1,000 halogenated natural products of various structural types have been isolated from sources such as bacteria, fungi, algae, higher plants, marine molluscs, insects, and mammals [1743]. Whereas fluorinated and iodinated species are rather rare, chloro and bromo derivatives are found more often. The former are predominantly produced by terrestrial species [1744] and the latter in marine organisms [1745]. For instance, about 107 tons of bromoalkanes such as bromoform and methylene bromide are released from coastal brown algae Ascophyllum nodosum into the atmosphere worldwide [1746, 1747]. Although the natural function of halogenating enzymes is not yet known, they do seem to be involved in the defence mechanism of their hosts. For instance, some algae produce halometabolites, which makes them inedible to animals [1748]. In contrast to hydrolytic or redox enzymes, which have been investigated since about a century, halogen-converting biocatalysts have been a subject of research for only the last 30 years [1749–1754].
2.7.1 Halogenation
Although an impressive number of halometabolites have been identified, only a few type of halogenating enzymes have been characterized to date [1755–1758]: Flavin-dependent halogenases [1759], α-ketoglutarate-dependent nonheme iron-halogenases [1760] and haloperoxidases. Among them, the latter group showed the broadest substrate scope and thus had a dominant impact in biotransformations [1300, 1320, 1761–1763]. This type of redox enzyme is widely distributed in nature and is capable of carrying out a multitude of halogenation reactions following the general equation shown below, where X stands for halide (Cl–, Br– and I–, but not F–). For redox reactions catalyzed by haloperoxidases which do not involve a halide (such as hydroxylation, epoxidation, or sulfoxidation) see Sect. 2.3.3.
The individual enzymes are called chloro-, bromo-, and iodoperoxidase. The name reflects the smallest halide ion that they can oxidize, in correlation to the corresponding redox potential. Bearing in mind their unique position as halogenating enzymes and the large variety of structurally different halometabolites produced by them, it is not surprising that the majority of haloperoxidases are characterized by a low product selectivity and wide substrate tolerance. As a consequence, any asymmetric induction observed in haloperoxidase-catalyzed reactions is usually low.
The most intensively studied haloperoxidases are the chloroperoxidase from the mold Caldariomyces fumago [1322] and bromoperoxidases from algae [1764] and bacteria such as Pseudomonas aureofaciens [1765], Ps. pyrrocinia [1766], and Streptomyces sp. [1767]. The only iodoperoxidase of preparative use is isolated from horseradish root [1768]. A haloperoxidase isolated from milk has been reported to be useful for the formation of halohydrins [1769].
2.7.1.1 Halogenation of Alkenes
Haloperoxidases have been shown to transform alkenes by a formal addition of hypohalous acid to produce halohydrins. The reaction mechanism of enzymatic halogenation has been debated for some time and it is now accepted that it proceeds via a halonium intermediate [1770, 1771], similar to the chemical formation of halohydrins (Scheme 2.228). The former species is derived from hypohalous acid or molecular halogen, which is in turn produced by the enzyme via oxidation of halide [1772]. In support of this, a HOCl-adduct of Fe3+-protoporphyrin IX was identified as a ‘direct’ enzyme-halogen intermediate involved in chloroperoxidase-catalyzed halogenation [1773].

Haloperoxidase-catalyzed transformation of alkenes
Functional groups present in the alkene can lead to products other than the expected halohydrin by competing with hydroxyl anion (pathway A) for the halonium intermediate. Unsaturated carboxylic acids, for instance, are transformed into the corresponding halolactones due to the nucleophilicity of the carboxylate group (pathway B) affording a halolactonization [1774, 1775]. Similarly, if the concentration of halide ion in the reaction mixture is increased, 1,2-dihalides are formed (pathway C) [1776]. Although this latter transformation may primarily be regarded as an undesired side-reaction, it offers the unique possibility of introducing fluorine, which is not oxidized by haloperoxidases, into the substrate via an enzyme-catalyzed process. Furthermore, migration of functional groups such as halogen [1777] and loss of carbon-containing units such as acetate and formaldehyde may occur, particularly when an oxygen substituent is attached to the C=C bond [1778, 1779].
All types of carbon–carbon double bonds – isolated (e.g., propene), conjugated (e.g., butadiene) and cumulative (e.g., allene) – are reactive (Scheme 2.229) [1780]. The size of the substrate seems to be of little importance since steroids [1781] and sterically demanding bicyclic alkenes [1782] are accepted equally well. Any regioselectivity observed reflects the (predominant) chemical and nonenzymatic nature of halohydrin formation.

Formation of halohydrins from alkenes
2.7.1.2 Halogenation of Alkynes
With alkyne substrates, haloperoxidase-catalyzed reactions yield α-haloketones (Scheme 2.230) [1783]. As with alkenes, the product distribution in the reaction with alkynes is dependent on the halide ion concentration. Both homogeneous and heterogeneous dihalides can be formed, dependent upon whether a single halide species or a mixture of halide ions are present.

Haloperoxidase-catalyzed reactions of alkynes
2.7.1.3 Halogenation of Aromatic Compounds
A wide range of electron-rich aromatic and heteroaromatic compounds are readily halogenated by haloperoxidases (Scheme 2.231) [1784–1786]. Bearing in mind the electrophilic character of the halogenating species, electron-rich phenols [1787, 1788] and anilines [1789] as well as their respective O- and N-alkyl derivatives are particularly well accepted. The color change of phenolic dyes such as phenol red or fluorescein upon halogenation serves as a simple assay for haloperoxidases [1790]. Since haloperoxidases are also peroxidases, they also can catalyze halide-independent peroxidation reactions of aromatics (see Sect. 2.3.3). Thus, dimerization, polymerization, oxygen insertion and de-alkylation reactions are encountered as undesired side-reactions, particularly whenever the halide ion is omitted or depleted from the reaction mixture.

Halogenation of aromatic compounds
2.7.1.4 Halogenation of C–H Groups
Similar to the chemical process, halogenation of C–H groups is only possible if they are appropriately activated by adjacent electron-withdrawing substituents, for example carbonyl groups, which facilitate enolization. Since the reactivity seems to be a function of the enol content of the substrate, simple ketones like 2-heptanone are unreactive [1791], but highly enolized 1,3-diketones are readily halogenated to give the corresponding 2-mono- or 2,2-dihalo derivatives (Scheme 2.232) [1792]. As with the formation of halohydrins from alkenes, selectivities are low and the reactivity of the substrate is independent of its size. For example, monocyclic compounds such as barbituric acid [1793] and sterically demanding polycyclic steroids are equally well accepted [1794]. β-Ketoacids are also halogenated, but the spontaneous decarboxylation of the intermediate α-halo-β-ketoacid affords the corresponding α-haloketones [1795]. The chloroperoxidase-catalyzed halogenation of oximes was shown to proceed via a two-step sequence via a halonitroso intermediate which is further oxidized to furnish a α-halonitro product [1796].

Halogenation of electronically activated C–H groups
2.7.1.5 Halogenation of N- and S-Atoms
Amines are halogenated by haloperoxidases to form unstable haloamines, which readily deaminate or decarboxylate, liberating the halogen [1797]. This pathway constitutes a part of the natural mammalian defence system against microorganisms, parasites and, perhaps, tumor cells. However, it is of no synthetic use. In an analogous fashion, thiols are oxidized to yield the corresponding sulfenyl halides. These highly reactive species are prone to undergo nucleophilic attack by hydroxyl ion or by excess thiol [1798, 1799]. As a result, sulfenic acids or disulfides are formed, respectively.
In view of the predominant chemical nature of biohalogenation, it seems that enzymatic halogenation reactions involving haloperoxidase enzymes do not show any significant advantage over the usual chemical reactions due to their lack of regio- and stereoselectivity. A benefit, however, lies in the mild reaction conditions employed.
2.7.2 Dehalogenation
The concentrations of haloorganic compounds in the ecosphere has remained reasonably constant due to the establishment of an equilibrium between biosynthesis and biodegradation. Due to man’s recent activities, a large number of halogen-containing compounds – most of which are recalcitrant – are liberated either by intent (e.g., insecticides), or because of poor practice (lead scavengers in gasoline) or through abuse (dumping of waste) into the ecosystem. These halogenated compounds would rapidly pollute the earth if there were no microbial dehalogenation pathways [1800, 1801]. Five major pathways for enzymatic degradation of halogenated compounds have been discovered (Table 2.13) [1802–1805].
Redox enzymes are responsible for the replacement of the halogen by a hydrogen atom via reductive dehalogenation [1806, 1807] and for oxidative degradation [1808]. Elimination of hydrogen halide leads to the formation of an alkene [1809], which is further degraded by oxidation. Since all of these pathways proceed either with a loss of a functional group or through removal of a chirality center, they are of little use for the biocatalytic synthesis of organic compounds. On the other hand, the enzyme-catalyzed formation of an epoxide from a halohydrin and the hydrolytic replacement of a halide by a hydroxyl functionality take place in a stereocontrolled fashion and are therefore of synthetic interest.
2.7.2.1 Dehalogenases
Hydrolytic dehalogenation catalyzed by dehalogenases (or ‘halido-hydrolases’) proceeds by formal nucleophilic substitution of the halogen atom with a hydroxyl ion [1810, 1811]. Neither cofactors nor metal ions are required for the enzymatic activity. Depending on the enzyme source, the reaction may either proceed with retention or inversion of configuration.Footnote 58 It is this stereospecificity which makes them attractive for synthetic purposes. To date, the only types of dehalogenases being of importance for preparative biotransformations due to their stereospecificity are those acting on α-haloacids (Table 2.14).
The mechanism of haloalkane dehalogenase acting with inversion of configuration was shown to have close similarities to that of epoxide hydrolases (Sect. 2.1.5) and retaining glycosidases (Sect. 2.6.2). Thus, the carboxyl moiety of an aspartate residue attacks the halide by forming an ‘alkyl-enzyme intermediate’ (Scheme 2.233). Being a carboxyl ester, the latter is hydrolyzed by a hydroxyl ion which is provided from water by the aid of a histidine [570, 1813]. On the other hand, α-haloacid dehalogenases acting with retention of configuration were shown to possess a catalytically active sulfhydryl residue.

Mechanism of inverting haloalkane dehalogenases
Two sub-types of α-haloacid dehalogenases have been characterized which are classified according to their preferred type of substrate: 2-Haloacid dehalogenase is able to accept all kinds of short-chain 2-haloacids [1814, 1815], while haloacetate dehalogenase exclusively acts on haloacetates [1816, 1817]. Both of them are inactive on nonactivated isolated halides. Interestingly, the reactivity of organic halides (fluoride through iodide derivatives) in dehalogenase reactions depends on the source of enzyme. In some cases it increases from iodine to fluorine derivatives, which is in sharp contrast to the corresponding chemical reactivity with nucleophiles such as hydroxyl ion [1818]. The most intriguing aspect of dehalogenases is their enantiospecificity [1819]. Depending on the growth conditions, the microbial production of (R)- or (S)-specific enzymes may be induced [1820, 1821]. This makes the stereoselective hydrolysis of α-haloacids to give the corresponding α-hydroxyacids possible [1822].
(S)-2-Chloropropionic acid is a key chiral synthon required for the synthesis of a range of important α-aryl- and α-aryloxypropionic acids used as anti-inflammatory agents and herbicides (Scheme 2.38). Several attempts to resolve racemic 2-chloropropionic acid via enzymatic methods using ‘classic’ hydrolases have been reported to proceed with varying degrees of selectivity [1823]. An elegant approach makes use of an (R)-specific dehalogenase enzyme from Pseudomonas putida NCIMB 12018 (Scheme 2.234) [1824–1826]. Thus, from a racemic mixture of α-haloacid, the (R)-enantiomer is converted into the (S)-hydroxyacid product via inversion of configuration leaving the (S)-α-haloacid behind. Some minor structural variations of the substrate are tolerated. This process has been scaled-up to industrial production at a capacity of 2,000 t/year [1827].

Resolution of 2-chloropropanoic acid derivatives by 2-haloacid dehalogenase
Two drawbacks of α-haloacid dehalogenation – i.e., the hydrolytic instability of α-bromoacids in aqueous solvent systems and the limited substrate tolerance of α-haloacid dehalogenases – can be overcome by using anhydrous organic solvents [1828]. Thus, long-chain α-haloacids (which were not accepted as substrates in water) were successfully transformed with good specificity in toluene, acetone or even in dimethyl sulfoxide.
2.7.2.2 Halohydrin Dehalogenases
The biodegradation of halohydrins proceeds through a two-step mechanism involving epoxide-formation catalysed by halohydrin dehalogenases [EC 3.8.1.5],Footnote 59 followed by epoxide hydrolase-mediated formation of vic-diols (Sect. 2.1.5), which are oxidatively degraded. A number of organisms possessing halohydrin dehalogenase and epoxide hydrolase activity, respectively, were found among bacteria (Flavo- [1830, 1831], Corynebacteria [1832], Arthrobacter erithrii [1833, 1834], Pseudomonas sp. [1835]), fungi (Caldariomyces fumago), and algae (Laurencia pacifica).
First hints on the stereoselectivity of halohydrin dehalogenases were obtained from studies on the desymmetrization of prochiral 1,3-dichloropropan-2-ol yielding epichlorohydrin using resting cells of Corynebacterium sp. (Scheme 2.235) [1836]. In two-step sequence, (R)-3-chloropropane-1,2-diol was formed in 74% e.e. via epichlorohydrin through the sequential action of an (unspecified) halohydrin dehalogenase and an epoxide hydrolase [1837]. Further studies revealed that these activities are widespread among bacteria [1838–1842].

Asymmetric microbial degradation of prochiral halohydrin by a Corynebacterium sp.
A breakthrough was achieved by cloning and overexpression of halohydrin dehalogenases from Agrobacterium radiobacter, which allowed the preparative-scale application of these enzymes under well-defined conditions [1843].
The mechanism of halohydrin dehalogenase was shown to proceed in a reversible fashion via a push-pull-like nucleophilic attack of halide (provided by a lipophilic halide binding site) with simultaneous activation of the epoxide through protonation by a Tyr residue (Scheme 2.236) [1844, 1845].

Catalytic mechanism of halohydrin dehalogenase from Agrobacterium radiobacter
Using pure halohydrin dehalogenase (HheC), competing activities observed in whole-cell preparations were eliminated and halohydrins could be resolved via enantioselective ring-closure with excellent enantioselectivities yielding enantiomeric epoxides and nonreacted halohydrins (Scheme 2.237) [1846].

Kinetic resolution of halohydrins using halohydrin dehalogenase
Subsequent studies revealed that the natural nucleophile halide (Cl, Br, I) could be replaced by nonnatural analogs, such as azide [1847], nitrite [1848], cyanide [1849], (thio)cyanate and formate by maintining the exquisite regioselectivity of nucleophilic attack at the less hindered oxirane carbon atom. Whereas the reaction rates observed with cyanide, (thio)cyanate, and formate were comparable to those using halide, azide and nitrite proved to be much better nucleophiles [1850]. Nonlinear and nonanionic nucleophiles, such as H2S, acetate, PO 3–4 , CO 2–3 , BO 3–3 , and F– were unreactive. The use of N-nucleophiles opened the way to prepare 1,2- and 1,3-aminoalcohols using azide or cyanide via the corresponding 1-azido-2-ols and 1-cyano-2-ols, respectively (Scheme 2.238).

Regio- and enantioselective ring-opening of epoxides using nonnatural nucleophiles catalysed by halohydrin dehalogenase
The mono-nitrite (or formate) esters of vic-diols obtained via enzymatic ring-opening of epoxides in presence of nitrite (or formate) are unstable and undergo spontaneous (nonenzymatic) hydrolysis to furnish the corresponding diols. This protocol offers a useful complement to the asymmetric hydrolysis of epoxides. Depending on the type of substrate and the enzymes used, enantio-complementary epoxide hydrolysis can be achieved [1851].
A one-pot two-step transformation of ethyl (S)-4-chloro-3-hydroxybutanoate (obtained via asymmetric bioreduction of the corresponding β-ketoester) via (reversible) epoxide-formation followed by ring-opening with cyanide was accomplished on a kg-scale using a halohydrin dehalogenase mutant. Ethyl (R)-4-cyano-4-butanoate was thus obtained in a highly chemoselective fashion without formation of byproducts, which plagued the chemical process. The latter product is a key intermediate for the synthesis of antihypocholesterolemic ‘statin’ agents [1852] (Scheme 2.238).
Notes
- 1.
- 2.
Since the molar concentration of water in an aqueous solution is 55.5 mol/L, the equilibrium is shifted towards the hydrolysis-side to such an extent, that the reaction can be regarded as virtually irreversible.
- 3.
These groups are called homochiral.
- 4.
The Enantiomeric Ratio (E) is a synonym for the so-called selectivity factor (s). Whereas the former term is used more often in biocatalyzed kinetic resolutions, the s-factor is more common in chemo-catalysis. In a mathematical sense, both are identical and describe the ratio of the relative (second-order) rate constants of enantiomers. For a comprehensive discussion see [47].
- 5.
For an alternative model see [56].
- 6.
- 7.
‘Alcalase’ is mainly used as additive in detergents for the degradation of proteinogenic impurities, its major enzyme component is subtilisin Carlsberg (alkaline protease A).
- 8.
It should be mentioned that some references and absolute configurations are wrong in this paper.
- 9.
PLE-1 is also named γ-PLE.
- 10.
‘Alternative pig liver esterase’, this enzyme was also termed Pharma-PLE [247].
- 11.
Be aware of the switch in the Cahn-Ingold-Prelog sequence priority.
- 12.
By definition, the range of microwave irradiation extends from 1 to 300 GHz; however, due to the resonance frequency of water (19.5 GHz), most of the applications are close to the latter range, i.e., 0.9 and 2.45 GHz.
- 13.
A sequence identity of 70% translates into a reasonably well-defined model showing a root mean square deviation of 1–2 Å, which drops to a low value of 2–4 Å for proteins having only 25% identity.
- 14.
The possible number of mutants generated from a typical protein possessing 200 amino acids exponentially increases by the number of mutations: 3,800 variants are possible for a single mutation, 7,183,900 exist for 2 mutations, and 8,429,807,368,950 are possible for only 4 mutations.
- 15.
It seems to be a common phenomenon that microbiologists keep reclassifying microbial species every once in a while. Whether this is to confuse organic chemists, or for other reasons, is often not clear. However, neither the microorganism nor the lipase are changed by a new name.
- 16.
In immobilized form, the upper operational limit increases to 60–80°C.
- 17.
Several Pseudomonas spp. were reclassified as Burkholderia spp.
- 18.
For a predictive model for Aspergillus sp. lipase see [478].
- 19.
A report on the chemical modification of crude PPL using carbamates has to be taken with some caution, since in this case the selective (chemical) deactivation of competing enzymes in the crude steapsin-mixture may likewise be the case for selectivity enhancement, see [501].
- 20.
Also termed ‘pyro-phosphates’.
- 21.
The retail price for one mole of ATP is about US $4,500, bulk prices are about one tenth of that.
- 22.
For the corresponding asymmetric hydrolysis of epoxides using a chiral Co-salen complex see [561].
- 23.
Epoxide hydrolases are occasionally also called ‘epoxide hydratases’ or ‘epoxide hydrases’.
- 24.
For an unusual cis-hydration see [587].
- 25.
It must be emphasized that the microorganism used in this study (Chryseomonas luteola) causes inner ear infections in infants and belongs to safety Class II, see [600].
- 26.
The strain was formerly denoted as Brevibacterium and is available as CBS 717.73.
- 27.
The current bulk prices for one mole are: NAD+ US $710, NADH US $3,000, NADP+ US $5,000 and NADPH US $215,000; retail prices are about ten times higher.
- 28.
For example, if the reduction of NAD(P)+ to NAD(P)H is 95% selective for hydride transfer onto the p-position of the nicotinamide ring, after 100 turnovers the residual activity of the cofactor would be 0.95100 being equivalent to only ~0.6%.
- 29.
For the catalytic mechanism see [777].
- 30.
When the chiral center is moved to the β- or γ-position, in situ racemization is impossible and, as a consequence, syn/anti-diastereomers are always obtained in a 1:1 ratio.
- 31.
Eichberger and Faber 1984, unpublished results.
- 32.
l-Carnitine is an essential factor for the transport of long-chain fatty acids across mitochondrial membranes and is used in the treatment of certain dysfunctions of skeletal muscles, acute hypoglycemia, and heart disorders.
- 33.
- 34.
Ene-reductases are also denoted as enoate reductases.
- 35.
- 36.
Salts of methyl viologen (1,1′-dimethyl-4,4′-dipyridinium dication, ‘Paraquat’) have been used as a total herbicide.
- 37.
It is an alarming fact that ≥90% of the hydrocarbons derived from drude oil are ‘wasted’ for energy-production (forming CO2), the small remainder of ≤10% is used as raw material for the chemical industry to produce of (long-lasting) products [1020].
- 38.
The most important processes with respect to scale are: p-xylene → terephthalic acid, ethylene → ethylene oxide, ethylene → acetaldehyde, ethylene/HOAc → vinyl acetate, methanol → formaldehyde, acetaldehyde → acetic acid.
- 39.
The only notable exception to this rule seems to be the Wacker-oxidation of alkenes.
- 40.
- 41.
For a complete list see [1093].
- 42.
Previously this value was believed to be 5.5 Å.
- 43.
The following strains have been used more frequently: Aspergillus niger, Cunninghamella blakesleeana, Bacillus megaterium, Bacillus cereus, Mucor plumbeus, Mortierella alpina, Curvularia lunata, Helmithosporium sativum, Pseudomonas putida, Rhizopus arrhizus, Rhizopus nigricans, Beauveria bassiana.
- 44.
Also called tyrosinase, catechol oxidase, cresolase.
- 45.
For the unusual microbial oxidation of a thioether catalyzed by a dioxygenase see [1183].
- 46.
Acinetobacter calcoaceticus NCIMB 9871 is a class-II pathogen.
- 47.
tert-Butylhydroperoxide and cumyl hydroperoxide.
- 48.
To date, the largest industrial-scale process is the oxidation of propene to propene oxide using tert-Bu–OOH.
- 49.
Compound I is comparable to the Fe+5 oxo-species in the mono-oxygenase cycle (see Scheme 2.146).
- 50.
For an X-ray structure see [1324].
- 51.
Also named 2-keto-3-deoxyoctanoate (KDO).
- 52.
For a related aldolase see [1437].
- 53.
For instance, atorvastatin (LipitorTM) or rosuvastatin (CrestorTM).
- 54.
Correctly, this enzyme has the charming name ‘d-seduheptulose-7-phosphate: d-glyceraldehyde-3-phosphate glycoaldehyde transferase’.
- 55.
Hydroxynitrile lyases were often also called ‘oxynitrilases’. However, this term should be abandoned.
- 56.
The possible number of linear and branched oligosaccharide isomers for a reducing hexasaccharide was calculated to encompass 1.05 × 1012 structures, see [1610].
- 57.
The equilibrium constant between acetophenone and alanine was reported to be 8.8 × 10–4, see [1730].
- 58.
For an example exhibiting a retention of configuration see [1812].
- 59.
Previously, halohydrin dehalogenases were also termed ‘haloalcohol dehalogenases’ or ‘halohydrin epoxidases’, see [1829].
References
References to Sect. 2.1
Bornscheuer UT, Kazlauskas RJ (2006) Hydrolases in Organic Synthesis, 2nd ed. Wiley-VCH, Weinheim
Boland W, Frößl C, Lorenz M (1991) Synthesis 1049
Lee HC, Ko YH, Baek SB, Kim DH (1998) Bioorg. Med. Chem. Lett. 8: 3379
Otto H-H, Schirmeister T (1997) Chem. Rev. 97: 133
Harrison MJ, Burton NA, Hillier IH, Gould IR (1996) Chem. Commun. 2769
Fersht A (1985) Enzyme Structure and Mechanism, 2nd edn. Freeman, New York, p 405
Kazlauskas RJ, Weber HK (1998) Curr. Opinion Chem. Biol. 2: 121
Jones JB, Beck JF (1976) Asymmetric Syntheses and Resolutions using Enzymes. In: Jones JB, Sih CJ, Perlman D (eds) Applications of Biochemical Systems in Organic Chemistry. Wiley, New York, part I, p 107
Brady L, Brzozowski AM, Derewenda ZS, Dodson E, Dodson G, Tolley S, Turkenburg JP, Christiansen L, Huge-Jensen B, Norskov L, Thim L, Menge U (1990) Nature 343: 767
Blow D (1990) Nature 343: 694
Schrag JD, Li Y, Wu S, Cygler M (1991) Nature 351: 761
Sussman JL, Harel M, Frolow F, Oefner C, Goldman A, Toker L, Silman I (1991) Science 253: 872
Kirchner G, Scollar MP, Klibanov AM (1986) J. Am. Chem. Soc. 107: 7072
Faber K, Riva S (1992) Synthesis 895
Starmans WAJ, Doppen RG, Thijs L, Zwanenburg B (1998) Tetrahedron Asymmetry 9: 429
Garcia MJ, Rebolledo F, Gotor V (1993) Tetrahedron Lett. 34: 6141
Puertas S, Brieva R, Rebolledo F, Gotor V (1993) Tetrahedron 49: 6973
Kitaguchi H, Fitzpatrick PA, Huber JE, Klibanov AM (1989) J. Am. Chem. Soc. 111: 3094
Björkling F, Frykman H, Godtfredsen SE, Kirk O (1992) Tetrahedron 48: 4587
Gotor V (1992) Enzymatic aminolysis, hydrazinolysis and oximolysis reactions. In: Servi S (ed) Microbial Reagents in Organic Synthesis, Nato ASI Series C, vol 381. Kluwer, Dordrecht, p 199
Abernethy JL, Albano E, Comyns J (1971) J. Org. Chem. 36: 1580
Silver MS (1966) J. Am. Chem. Soc. 88: 4247
Öhrner N, Orrenius C, Mattson A, Norin T, Hult K (1996) Enzyme Microb. Technol. 19: 328
Chen CS, Sih CJ (1989) Angew. Chem. Int. Ed. 28: 695
Schoffers E, Golebiowski A, Johnson CR (1996) Tetrahedron 52: 3769
Matsumoto K, Tsutsumi S, Ihori T, Ohta H (1990) J. Am. Chem. Soc. 112: 9614
Björkling F, Boutelje J, Gatenbeck S, Hult K, Norin T, Szmulik P (1985) Tetrahedron 41: 1347
Krisch K (1971) Carboxyl ester hydrolases. In: Boyer PD (ed) The Enzymes, 3rd edn, vol 5. Academic Press, New York, p 43
Ramos-Tombo GM, Schär H-P, Fernandez i Busquets X, Ghisalba O (1986) Tetrahedron Lett. 27: 5707
Gais HJ, Lukas KL (1984) Angew. Chem. Int. Ed. 23: 142
Kasel W, Hultin PG, Jones JB (1985) J. Chem. Soc., Chem. Commun. 1563
Wang YF, Chen CS, Girdaukas G, Sih CJ (1984) J. Am. Chem. Soc. 106: 3695
Kroutil W, Kleewein A, Faber K (1997) Tetrahedron Asymmetry 8: 3251
Kroutil W, Kleewein A, Faber K (1996) Free shareware programs (‘SeKiRe-Win-1.0’, ‘SeKiRe-Mac-1.0’) for the simulation, analysis and optimization of asymmetric catalytic processes proceeding through two consecutive steps running under Windows or on a Macintosh are available from the author’s website via the Internet: <http://borgc185.kfunigraz.ac.at>
Wang YF, Chen CS, Girdaukas G, Sih CJ (1985) Extending the applicability of esterases of low enantioselectivity in asymmetric synthesis. In: Porter R, Clark S (eds) Enzymes in Organic Synthesis, Ciba Foundation Symposium 111. Pitman, London, p 128
Sih CJ, Wu SH (1989) Topics Stereochem. 19: 63
Dakin HD (1903) J. Physiol. 30: 253
Gruber CC, Lavandera I, Faber K, Kroutil W (2006) Adv. Synth. Catal. 348: 1789
Klempier N, Faber K, Griengl H (1989) Synthesis 933
Chen CS, Fujimoto Y, Girdaukas G, Sih CJ (1982) J. Am. Chem. Soc. 104: 7294
Martin VS, Woodard SS, Katsuki T, Yamada Y, Ikeda M, Sharpless KB (1981) J. Am. Chem. Soc. 103: 6237
Bredig G, Fajans K (1908) Ber. dtsch. chem. Ges. 41: 752
Janes LE, Kazlauskas RJ (1997) J. Org. Chem. 62: 4560
Zmijewski MJ, Sullivan G, Persichetti R, Lalonde J (1997) Tetrahedron Asymmetry 8: 1153
Straathof AJJ, Jongejan JA (1997) Enzyme Microb. Technol. 21: 559
Gawley RE (2006) J. Org. Chem. 71: 2411
Faber K (1997) Enantiomer 2: 411
Straathof AJJ, Rakels JLL, Heijnen JJ (1992) Biocatalysis 7: 13
van Tol JBA, Jongejan JA, Geerlof A, Duine JA (1991) Recl. Trav. Chim. Pays-Bas 110: 255
Lu Y, Zhao X, Chen ZN (1995) Tetrahedron Asymmetry 6: 1093
Rakels JLL, Straathof AJJ, Heijnen JJ (1993) Enzyme Microb. Technol. 15: 1051
Faber K, Hönig H, Kleewein A (1995) Free shareware programs (‘Selectivity-Win-1.0’, ‘Selectivity-1.0’) for the calculation of the Enantiomeric Ratio for irreversible kinetic resolution running under Windows or on a Macintosh are available from the author’s website via the Internet: <http://borgc185.kfunigraz.ac.at>
Faber K, Hönig H, Kleewein A (1995) Recent developments: determination of the selectivity of biocatalytic kinetic resolution of enantiomers – the ‘enantiomeric ratio’. In: Roberts SM (ed) Preparative Biotransformations. Wiley, New York
Oberhauser T, Bodenteich M, Faber K, Penn G, Griengl H (1987) Tetrahedron 43: 3931
Chen CS, Wu, SH, Girdaukas G, Sih CJ (1987) J. Am. Chem. Soc. 109: 2812
Langrand G, Baratti J, Buono G, Triantaphylides C (1988) Biocatalysis 1: 231
Kroutil W, Kleewein A, Faber K (1997) Tetrahedron Asymmetry 8: 3263
Guo ZW, Wu SH, Chen CS, Girdaukas G, Sih CJ (1990) J. Am. Chem. Soc. 112: 4942
Kroutil W, Kleewein A, Faber K (1996) Free shareware programs (‘SeKiRe-Win-1.0’, ‘SeKiRe-Mac-1.0’) for the simulation, analysis and optimization of asymmetric catalytic processes proceeding through two consecutive kinetic resolution steps are available from the author’s website via the Internet: <http://borgc185.kfunigraz.ac.at>
Caron G, Kazlauskas R (1993) Tetrahedron Asymmetry 4: 1995
Kazlauskas RJ (1989) J. Am. Chem. Soc. 111: 4953
Wu SH, Zhang LQ, Chen CS, Girdaukas G, Sih CJ (1985) Tetrahedron Lett. 26: 4323
Alfonso I, Astorga C, Rebolledo F, Gotor V (1996) Chem. Commun. 2471
Macfarlane ELA, Roberts SM, Turner NJ (1990) J. Chem. Soc., Chem. Commun. 569
Chen CS, Liu YC (1991) J. Org. Chem. 56: 1966
Faber K (1992) Indian J. Chem. 31B: 921
Stecher H, Faber K (1997) Synthesis 1
Strauss UT, Felfer U, Faber K (1999) Tetrahedron Asymmetry 10: 107
Faber K (2001) Chem. Eur. J. 7: 5004
Guo ZW (1993) J. Org. Chem. 58: 5748
Ebbers EJ, Ariaans GJA, Houbiers JPM, Bruggink A, Zwanenburg B (1997) Tetrahedron 53: 9417
Adams E (1976) Adv. Enzymol. Relat. Areas Mol. Biol. 44: 69
Yagasaki M, Ozaki A (1997) J. Mol. Catal. B 4: 1
Danda H, Nagatomi T, Maehara A, Umemura T (1991) Tetrahedron 47: 8701
Takano S, Suzuki M, Ogasawara K (1993) Tetrahedron Asymmetry 4: 1043
Kanerva LT (1996) Acta Chem. Scand. 50: 234
Pedragosa-Moreau S, Morisseau C, Baratti J, Zylber J, Archelas A, Furstoss R (1997) Tetrahedron 53: 9707
El Gihani MT, Williams JMJ (1999) Curr. Opinion Chem. Biol. 3: 11
Kitamura M, Tokunaga M, Noyori R (1993) Tetrahedron 49: 1853
Ward RS (1995) Tetrahedron Asymmetry 6: 1475
Eliel EL, Wilen SH, Mander LN (eds) (1994) Stereochemistry of Organic Compounds. Wiley, New York, p 315
Morrisson JD (ed) (1983) Asymmetric Synthesis. Academic Press, New York, vol 1, p 1
Fülling G, Sih CJ (1987) J. Am. Chem. Soc. 109: 2845
Brinksma J, van der Deen H, van Oeveren A, Feringa BL (1998) J. Chem. Soc., Perkin Trans. 1, 4159
Um P-J, Drueckhammer DG (1998) J. Am. Chem. Soc. 120: 5605
Dinh PM, Williams JMJ, Harris W (1999) Tetrahedron Lett. 40: 749
Taniguchi T, Ogasawara K (1997) Chem. Commun. 1399
Wegman MA, Hacking MAPJ, Rops J, Pereira P, van Rantwijk F, Sheldon RA (1999) Tetrahedron Asymmetry 10: 1739
Inagaki M, Hiratake J, Nishioka T, Oda J (1992) J. Org. Chem. 57: 5643
Kitamura M, Tokunaga M, Noyori R (1993) J. Am. Chem. Soc. 115: 144
Tan DS, Günter MM, Drueckhammer DG (1995) J. Am. Chem. Soc. 117: 9093
Williams RM (1989) Synthesis of Optically Active α-Amino Acids. Pergamon Press, Oxford
Soda K, Tanaka H, Esaki N (1983) Amino acids. In: Rehm HJ, Reed G (eds) Biotechnology. Verlag Chemie, Weinheim, vol 3, p 479
Taylor PP, Pantaleone DP, Senkpeil RF, Fotheringham IG (1998) Trends Biotechnol. 16: 412
Yoshimura T, Jhee KH, Soda K (1996) Biosci. Biotechnol. Biochem. 60: 181
Ohshima T, Soda K (1989) Trends Biotechnol. 7: 210
Schmidt-Kastner G, Egerer P (1984) Amino acids and peptides. In: Rehm HJ, Reed G (eds) Biotechnology. Verlag Chemie, Weinheim, vol 6a, p 387
Kamphuis J, Boesten WHJ, Kaptein B, Hermes HFM, Sonke T, Broxterman QB, van den Tweel WJJ, Schoemaker HE (1992) The production and uses of optically pure natural and unnatural amino acids. In: Collins AN, Sheldrake GN, Crosby J (eds) Chirality in Industry. Wiley, Chichester, p 187
Enei H, Shibai H, Hirose Y (1982) Amino acids and related compounds. In: Tsao GT (ed) Annual Reports on Fermentation Processes. Academic Press, New York, vol 5, p 79
Abbott BJ (1976) Adv. Appl. Microbiol. 20: 203
Rozzell D, Wagner F (eds) (1992) Biocatalytic Production of Amino Acids and Derivatives. Hanser Publ., Munich
Yonaha K, Soda K (1986) Adv. Biochem. Eng. Biotechnol. 33: 95
Soda K, Tanaka H, Esaki N (1983) Amino acids. In: Rehm HJ, Reed G (eds) Biotechnology, vol 3. Verlag Chemie, Weinheim, p 479
Wagner I, Musso H (1983) Angew. Chem. Int. Ed. 22: 816
Kaptein B, Boesten WHJ, Broxterman QB, Peters PJH, Schoemaker HE, Kamphuis J (1993) Tetrahedron Asymmetry 4: 1113
Lu W, Rey P, Benezra A (1995) J. Chem. Soc., Perkin Trans. 1, 553
Yamada S, Hongo C, Yoshioka R, Chibata I (1983) J. Org. Chem. 48: 843
Kamphuis J, Boesten WHJ, Broxterman QB, Hermes HFM, van Balken JAM, Meijer EM, Schoemaker HE (1990) Adv. Biochem. Eng. Biotechnol. 42: 134
Warburg O (1905) Ber. dtsch. chem. Ges. 38: 187
Jones M, Page MI (1991) J. Chem. Soc. 316
Miyazawa T, Takitani T, Ueji S, Yamada T, Kuwata S (1988) J. Chem. Soc., Chem. Commun. 1214
Miyazawa T, Iwanaga H, Ueji S, Yamada T, Kuwata S (1989) Chem. Lett. 2219
Jones JB, Beck JF (1976) Applications of chymotrypsin in resolutions and asymmetric synthesis. In: Jones JB, Sih CJ, Perlman D (eds) Applications of Biochemical Systems in Organic Chemistry. Wiley, New York, part I, p 137
Dirlam NC, Moore BS, Urban FJ (1987) J. Org. Chem. 52: 3287
Berger A, Smolarsky M, Kurn N, Bosshard HR (1973) J. Org. Chem. 38: 457
Cohen SG (1969) Trans. NY Acad. Sci. 31: 705
Hess PD (1971) Chymotrypsin – chemical properties and catalysis. In: Boyer GP (ed) The Enzymes, vol 3. Academic Press, New York, p 213
Chenevert R, Letourneau M, Thiboutot S (1990) Can. J. Chem. 68: 960
Roper JM, Bauer DP (1983) Synthesis 1041
Izquierdo MC, Stein RL (1990) J. Am. Chem. Soc., 112: 6054
Chen ST, Wang KT, Wong CH (1986) J. Chem. Soc., Chem. Commun. 1514
Glänzer BI, Faber K, Griengl H (1987) Tetrahedron 43: 771
Chenevert R, Bel Rhlid R, Letourneau M, Gagnon R, D’Astous L (1993) Tetrahedron Asymmetry 4: 1137
Jones JB, Kunitake T, Niemann C, Hein GE (1965) J. Am. Chem. Soc. 87: 1777
Pattabiraman TN, Lawson WB (1972) Biochem. J. 126: 645 and 659
Chen ST, Huang WH, Wang KT (1994) J. Org. Chem. 59: 7580
Greenstein JP, Winitz M (1961) Chemistry of the Amino Acids. Wiley, New York, p 715
Boesten WHJ, Dassen BHN, Kerkhoffs PL, Roberts MJA, Cals MJH, Peters PJH, van Balken JAM, Meijer EM, Schoemaker HE (1986) Efficient enzymic production of enantiomerically pure amino acids. In: Schneider MP (ed) Enzymes as Catalysts in Organic Synthesis. Reidel, Dordrecht, p 355
Sonke T, Kaptein B, Boesten WHJ, Broxterman QB, Schoemaker HE, Kamphuis J, Formaggio F, Toniolo T, Rutjes FPJT (2000) In: Patel RN (ed) Stereoselective Biocatalysis. Marcel Dekker, New York, p 23
Eichhorn E, Roduit J-P, Shaw N, Heinzmann K, Kiener A (1997) Tetrahedron Asymmetry 8: 2533
Kamphuis J, Boesten WHJ, Broxterman QB, van Balken JAM, Meijer EM, Schoemaker HE (1990) Adv. Biochem. Eng. Biotechnol. 42: 133
Sambale C, Kula MR (1987) Biotechnol. Appl. Biochem. 9: 251
Chibata I, Tosa T, Sato T, Mori T (1976) Methods Enzymol. 44: 746
Greenstein JP (1957) Methods Enzymol. 3: 554
Chibata I, Ishikawa T, Tosa T (1970) Methods Enzymol. 19: 756
Bommarius AS, Drauz K, Klenk H, Wandrey C (1992) Ann. New York Acad. Sci. 672: 126
Tosa T, Mori T, Fuse N, Chibata I (1967) Biotechnol. Bioeng. 9: 603
Chenault HK, Dahmer J, Whitesides GM (1989) J. Am. Chem. Soc. 111: 6354
Mori K, Otsuka T (1985) Tetrahedron 41: 547
Baldwin JE, Christie MA, Haber SB, Kruse LI (1976) J. Am. Chem. Soc. 98: 3045
Verseck S, Bommarius A S, Kula MR (2001) Appl. Microbiol. Biotechnol. 55: 345
May O, Verseck S, Bommarius A S, Drauz K (2002) Org. Proc. Res. Dev. 6: 452
Tokuyama S, Hatano K (1996) Appl. Microbiol. Biotechnol. 44: 774
Shaw NM, Robins K, Kiener A (2003) Adv. Synth. Catal. 345: 425
Petersen M, Sauter M (1999) Chimia 53: 608
Solodenko VA, Kasheva TN, Kukhar VP, Kozlova EV, Mironenko DA, Svedas VK (1991) Tetrahedron 47: 3989
Bücherer HT, Steiner W (1934) J. Prakt. Chem. 140: 291
Ogawa J, Shimizu S (1997) J. Mol. Catal. B 2: 163
Yamada H, Takahashi S, Kii Y, Kumagai H (1978) J. Ferment. Technol. 56: 484
Ogawa J, Shimizu S (2000) In: Patel RN (ed) Stereoselective Bioocatalysis. Marcel Dekker, New York, p 1
Runser S, Chinski N, Ohleyer E (1990) Appl. Microbiol. BIotechnol. 33: 382
Möller, A, Syldatk C, Schulze M, Wagner F (1988) Enzyme Microb. Technol. 10: 618
Wallach DP, Grisolia S (1957) J. Biol. Chem. 226: 277
Olivieri R, Fascetti E, Angelini L (1981) Biotechnol. Bioeng. 23: 2173
Yamada H, Shimizu S, Shimada H, Tani Y, Takahashi S, Ohashi T (1980) Biochimie 62: 395
Guivarch M, Gillonnier C, Brunie JC (1980) Bull. Soc. Chim. Fr. 91
Syldatk C, Müller R, Pietzsch M, Wagner F (1992) Biocatalytic Production of Amino Acids and Derivatives. Hanser Publ., Munich, p 129
Yamada H, Takahashi S, Yoshiaki K, Kumagai H (1978) J. Ferment. Technol. 56: 484
Watabe K, Ishikawa T, Mukohara Y, Nakamura H (1992) J. Bacteriol. 174: 7989
Pietzsch M, Syldatk C, Wagner F (1992) Ann. New York Acad. Sci. 672: 478
Evans C, McCague R, Roberts SM, Sutherland AG (1991) J. Chem. Soc., Perkin Trans. 1, 656
Jones M, Page MI (1991) J. Chem. Soc., Chem. Commun. 316
Brieva R, Crich JZ, Sih CJ (1993) J. Org. Chem. 58: 1068
Evans C, McCague R, Roberts SM, Sutherland AG, Wisdom R (1991) J. Chem. Soc., Perkin Trans. 1, 2276
Forro E, Fülöp F (2006) Chem. Eur. J. 12: 2587
Zhu LM, Tedford MC (1990) Tetrahedron 46: 6587
Ohno M, Otsuka M (1989) Org. React. 37: 1
Pearson AJ, Bansal HS, Lai YS (1987) J. Chem. Soc., Chem. Commun. 519
Johnson CR, Penning TD (1986) J. Am. Chem. Soc. 108: 5655
Suemune H, Harabe T, Xie ZF, Sakai K (1988) Chem. Pharm. Bull. 36: 4337
Dropsy EP, Klibanov AM (1984) Biotechnol. Bioeng. 26: 911
Chenevert R, Martin R (1992) Tetrahedron Asymmetry 3: 199
Kotani H, Kuze Y, Uchida S, Miyabe T, Limori T, Okano K, Kobayashi S, Ohno M (1983) Agric. Biol. Chem. 47: 1363
Lambrechts C, Galzy P (1995) Biosci. Biotech. Biochem. 59: 1464
Jackson MA, Labeda DP, Becker LA (1995) Enzyme Microb. Technol. 17: 175
Quax WJ, Broekhuizen CP (1994) Appl. Microbiol. Biotechnol. 41: 425
Bornscheuer UT, Kazlauskas R J (2006) Hydrolases in Organic Synthesis, 2nd ed. Wiley-VCH, Weinheim, p 179
Zocher F, Krebsfänger N, Yoo OJ, Bornscheuer UT (1998) J. Mol. Catal. B 5: 199
Schlacher A, Stanzer T, Soelkner B, Klingsbichel E, Petersen E, Schmidt M, Klempier N, Schwab H (1997) J. Mol. Catal. B 3: 25
Jones JB (1980) In: Dunnill P, Wiseman A, Blakeborough N (eds) Enzymic and Non-enzymic Catalysis. Horwood/Wiley, New York, p 54
Schubert Wright C (1972) J. Mol. Biol. 67: 151
Philipp M, Bender ML (1983) Mol. Cell. Biochem. 51: 5
Fruton JS (1971) Pepsin. In: Boyer PD (ed) The Enzymes, vol 3. Academic Press, London, p 119
Bianchi D, Cabri W, Cesti P, Francalanci F, Ricci M (1988) J. Org. Chem. 53: 104
Fancetic O, Deretic V, Marjanovic N, Glisin V (1988) Biotechnol. Forum 5: 90
Baldaro E, Fuganti C, Servi S, Tagliani A, Terreni M (1992) The use of immobilized penicillin acylase in organic synthesis. In: Servi S (ed) Microbial Reagents in Organic Synthesis. NATO ASI Ser. C, vol 381. Kluwer, Dordrecht, p 175
Bender ML, Killheffer JV (1973) Crit. Rev. Biochem. 1: 149
Barnier JP, Blanco L, Guibe-Jampel E, Rousseau G (1989) Tetrahedron 45: 5051
Schultz M, Hermann P, Kunz H (1992) Synlett. 37
Moorlag H, Kellogg RM, Kloosterman M, Kaptein B, Kamphuis J, Schoemaker HE (1990) J. Org. Chem. 55: 5878
Kallwass HKW, Yee C, Blythe TA, McNabb TJ, Rogers EE, Shames SL (1994) Bioorg. Med. Chem. 2: 557
Chen ST, Fang JM (1997) J. Org. Chem. 62: 4349
Henke E, Bornscheuer UT, Schmid RD, Pleiss J (2003) ChemBioChem 4: 485
Henke E, Pleiss J, Bornscheuer UT (2002) Angew. Chem. Int. Ed. 41: 3211
Krishna S H, Persson M, Bornscheuer UT (2002) Tetrahedron Asymmetry 13: 2693
Heymann E, Junge W (1979) Eur. J. Biochem. 95: 509
Öhrner N, Mattson A, Norin T, Hult K (1990) Biocatalysis 4: 81
Lam LKP, Brown CM, De Jeso B, Lym L, Toone EJ, Jones JB (1988) J. Am. Chem. Soc. 110: 4409
Polla A, Frejd T (1991) Tetrahedron 47: 5883
Seebach D, Eberle M (1986) Chimia 40: 315
Reeve DC, Crout DHG, Cooper K, Fray MJ (1992) Tetrahedron Asymmetry 3: 785
Senayake CH, Bill TJ, Larsen RD, Leazer J, Reiter PJ (1992) Tetrahedron Lett. 33: 5901
De Jeso B, Belair N, Deleuze H, Rascle MC, Maillard B (1990) Tetrahedron Lett. 31: 653
Tanyeli C, Sezen B, Demir AS, Alves RB, Arseniyadis S (1999) Tetrahedron Asymmetry 10: 1129
Jongejan JA, Duine JA (1987) Tetrahedron Lett. 28: 2767
Burger U, Erne-Zellweger D, Mayerl CM (1987) Helv. Chem. Acta 70: 587
Hazato A, Tanaka T, Toru T, Okamura N, Bannai K, Sugiura S, Manabe K, Kurozumi S (1983) Nippon Kagaku Kaishi 9: 1390
Hazato A, Tanaka T, Toru T, Okamura N, Bannai K, Sugiura S, Manabe K, Kurozumi S (1984) Chem. Abstr. 100: 120720q
Papageorgiou C, Benezra C (1985) J. Org. Chem. 50: 1145
Sicsic S, Leroy J, Wakselman C (1987) Synthesis 155
Schirmeister T, Otto HH (1993) J. Org. Chem. 58: 4819
Schneider M, Engel N, Boensmann H (1984) Angew. Chem. Int. Ed. 23: 66
Luyten M, Müller S, Herzog B, Keese R (1987) Helv. Chim. Acta 70: 1250
Huang FC, Lee LFH, Mittal RSD, Ravikumar PR, Chan JA, Sih CJ, Capsi E, Eck CR (1975) J. Am. Chem. Soc. 97: 4144
Herold P, Mohr P, Tamm C (1983) Helv. Chim. Acta 76: 744
Adachi K, Kobayashi S, Ohno M (1986) Chimia 40: 311
Ohno M, Kobayashi S, Iimori T, Wang Y-F, Izawa T (1981) J. Am. Chem. Soc. 103: 2405
Mohr P, Waespe-Sarcevic, Tamm C, Gawronska K, Gawronski JK (1983) Helv. Chim. Acta 66: 2501
Cohen SG, Khedouri E (1961) J. Am. Chem. Soc. 83: 1093
Cohen SG, Khedouri E (1961) J. Am. Cherm. Soc. 83: 4228
Roy R, Rey AW (1987) Tetrahedron Lett. 28: 4935
Santaniello E, Chiari M, Ferraboschi P, Trave S (1988) J. Org. Chem. 53: 1567
Gopalan AS, Sih CJ (1984) Tetrahedron Lett. 25: 5235
Mohr P, Waespe-Sarcevic N, Tamm C, Gawronska K, Gawronski JK (1983) Helv. Chim. Acta 66: 2501
Schregenberger C, Seebach D (1986) Liebigs Ann. Chem. 2081
Sabbioni G, Jones JB (1987) J. Org. Chem. 52: 4565
Björkling F, Boutelje J, Gatenbeck S, Hult K, Norin T (1985) Appl. Microbiol. Biotechnol. 21: 16
Bloch R, Guibe-Jampel E, Girard G (1985) Tetrahedron Lett. 26: 4087
Laumen K, Schneider M (1984) Tetrahedron Lett. 25: 5875
Harre M, Raddatz P, Walenta R, Winterfeldt E (1982) Angew. Chem. Int. Ed. 21: 480
Hummel A, Brüsehaber E, Böttcher D, Trauthwein H, Doderer K, Bornscheuer UT (2007) Angew. Chem. Int. Ed. 46: 8492
Deardorff DR, Mathews AJ, McMeekin DS, Craney CL (1986) Tetrahedron Lett. 27: 1255
Laumen K, Schneider MP (1986) J. Chem. Soc., Chem. Commun. 1298
Johnson CR, Bis SJ (1992) Tetrahedron Lett. 33: 7287
Sih CJ, Gu QM, Holdgrün X, Harris K (1992) Chirality 4: 91
Wang YF, Sih CJ (1984) Tetrahedron Lett. 25: 4999
Iriuchijima S, Hasegawa K, Tsuchihashi G (1982) Agric. Biol. Chem. 46: 1907
Mohr P, Rösslein L, Tamm C (1987) Helv. Chim. Acta 70: 142
Alcock NW, Crout DHG, Henderson CM, Thomas SE (1988) J. Chem. Soc., Chem. Commun. 746
Ramaswamy S, Hui RAHF, Jones JB (1986) J. Chem. Soc., Chem. Commun. 1545
Sicsic S, Ikbal M, Le Goffic F (1987) Tetrahedron Lett. 28: 1887
Klunder AJH, Huizinga WB, Hulshof AJM, Zwanenburg B (1986) Tetrahedron Lett. 27: 2543
Crout DHG, Gaudet VSB, Laumen K, Schneider MP (1986) J. Chem. Soc., Chem. Commun. 808
Lange S, Musidlowska A, Schmidt-Dannert C, Schmitt J, Bornscheuer UT (2001) ChemBioChem 2: 576
Musidlowska A, Lange S, Bornscheuer UT (2001) Angew. Chem. Int. Ed. 40: 2851
Dominguez de Maria P, Garcia-Burgos CA, Bargeman G, van Gemert RW (2007) Synthesis 1439
Hermann M, Kietzmann MU, Ivancic M, Zenzmaier C, Luiten RGM, Skranc W, Wubbolts M, Winkler M, Birner-Gruenberger R, Pichler H, Schwab H (2008) J. Biotechnol. 133: 301
May O (2009) Green chemistry with biocatalysis for production of pharmaceuticals. In: Tao J, Lin GQ, Liese A (eds) Biocatalysis for the Pharmaceutical Industry. Wiley Asia, Singapore, p 310
Sugai T, Kuwahara S, Hishino C, Matsuo N, Mori K (1982) Agric. Biol. Chem. 46: 2579
Oritani T, Yamashita K (1980) Agric. Biol. Chem. 44: 2407
Ohta H, Miyamae Y, Kimura Y (1989) Chem. Lett. 379
Ziffer H, Kawai K, Kasai K, Imuta M, Froussios C (1983) J. Org. Chem. 48: 3017
Takaishi Y, Yang YL, DiTullio D, Sih CJ (1982) Tetrahedron Lett. 23: 5489
Glänzer BI, Faber K, Griengl H (1987) Tetrahedron 43: 5791
Mutsaers JHGM, Kooreman HJ (1991) Recl. Trav. Chim. Pays-Bas 110: 185
Smeets JWH, Kieboom APG (1992) Recl. Trav. Chim. Pays-Bas 111: 490
Stahly GP, Starrett RM (1997) Production methods for chiral non-steroidal anti-inflammatory profen drugs. In: Collins AN, Sheldrake GN, Crosby J (eds) Chirality in Industry II. Wiley, Chichester, pp 19–40
Jones JB, Beck JF (1976) Applications of chymotrypsin in resolution and asymmetric synthesis. In: Jones JB, Sih CJ, Perlman D (eds) Applications of Biochemical Systems in Organic Synthesis. Wiley, New York, p 137
Lalonde JJ, Bergbreiter DE, Wong CH (1988) J. Org. Chem. 53: 2323
Shin C, Seki M, Takahashi N (1990) Chem. Lett. 2089
Miyazawa T, Iwanaga H, Yamada T, Kuwata S (1994) Biotechnol. Lett. 16: 373
Pathak T, Waldmann H (1998) Curr. Opinion Chem. Biol. 2: 112
Waldmann H, Reidel A (1997) Angew. Chem. Int. Ed. 36: 647
Francetic O, Deretic V, Marjanovic N, Glisin V (1988) Biotech-Forum 5: 90
Waldmann H (1988) Liebigs Ann. Chem. 1175
Fuganti C, Grasselli P, Servi S, Lazzarini A, Casati P (1988) Tetrahedron 44: 2575
Waldmann H (1989) Tetrahedron Lett. 30: 3057
Fernandez-Lafuente R, Guisan JM, Pregnolato M, Terreni M (1997) Tetrahedron Lett. 38: 4693
Pohl T, Waldmann H (1995) Tetrahedron Lett. 36: 2963
Baldaro E, Faiardi D, Fuganti C, Grasselli P, Lazzarini A (1988) Tetrahedron Lett. 29: 4623
Uemura A, Nozaki K, Yamashita J, Yasumoto M (1989) Tetrahedron Lett. 30: 3819
Kvittingen L, Partali V, Braenden JU, Anthonsen T (1991) Biotechnol. Lett. 13: 13
Pugniere M, San Juan C, Previero A (1990) Tetrahedron Lett. 31: 4883–4886
Berger B, de Raadt A, Griengl H, Hayden W, Hechtberger P, Klempier N, Faber K (1992) Pure Appl. Chem. 64: 1085
Dale JA, Dull DL, Mosher HS (1969) J. Org. Chem. 34: 2543
Feichter C, Faber K, Griengl H (1991) J. Chem. Soc., Perkin Trans. 1, 653
Faber K, Ottolina G, Riva S (1993) Biocatalysis 8: 91
Kamezawa M, Raku T, Tachibana H, Ohtani T, Naoshima Y (1995) Biosci. Biotechnol. Biochem. 59: 549
Naoshima Y, Kamezawa M, Tachibana H, Munakata Y, Fujita T, Kihara K, Raku T (1993) J. Chem. Soc., Perkin Trans. 1, 557
Guanti G, Banfi L, Narisano E (1992) Asymmetrized tris(hydroxymethyl)methane and related synthons as new highly versatile chiral building blocks. In: Servi S (ed) Microbial Reagents in Organic Synthesis. Nato ASI Series C, vol 381. Kluwer, Dordrecht, pp 299–310
Morgan B, Oehlschlager AC, Stokes TM (1992) J. Org. Chem. 57: 3231
Nguyen BV, Nordin O, Vörde C, Hedenström E, Högberg HE (1997) Tetrahedron Asymmetry 8: 983
Nordin O, Hedenström E, Högberg H-E (1994) Tetrahedron Asymmetry 5: 785
Björkling F, Boutelje J, Gatenbeck S, Hult K, Norin T (1986) Bioorg. Chem. 14: 176
Guanti G, Banfi L, Narisano E, Riva R, Thea S (1986) Tetrahedron Lett. 27: 4639
Santaniello E, Ferraboschi P, Grisenti P, Aragozzini F, Maconi E (1991) J. Chem. Soc., Perkin Trans. 1, 601
Björkling F, Boutelje J, Gatenbeck, S, Hult K, Norin T (1985) Tetrahedron Lett. 26: 4957
Guo Z-W, Sih CJ (1989) J. Am. Chem. Soc. 111: 6836
Barton MJ, Hamman JP, Fichter KC, Calton GJ (1990) Enzyme Microb. Technol. 12: 577
Grandjean D, Pale P, Chuche J (1991) Tetrahedron Lett. 32: 3043
Sugai T, Hamada K, Akeboshi T, Ikeda H, Ohta H (1997) Synlett. 983
Liu YY, Xu JH, Xu QG, Hu Y (1999) Biotechnol. Lett. 21: 143
Lam LKP, Hui RAHF, Jones JB (1986) J. Org. Chem. 51: 2047
Phillips RS (1992) Enzyme Microb. Technol. 14: 417
Phillips RS (1996) Trends Biotechnol. 14: 13
Keinan E, Hafeli EK, Seth KK, Lamed R (1986) J. Am. Chem. Soc. 108: 162
Holmberg E, Hult K (1991) Biotechnol. Lett. 13: 323
Boutelje J, Hjalmarsson M, Hult K, Lindbäck M, Norin T (1988) Bioorg. Chem. 16: 364
Sakai T, Kawabata I, Kishimoto T, Ema T, Utaka M (1997) J. Org. Chem. 62: 4906
Sakai T, Kishimoto T, Tanaka Y, Ema T, Utaka M (1998) Tetrahedron Lett. 39: 7881
Sakai T, Mitsutomi H, Korenaga T, Ema T (2005) Tetrahedron Asymmetry 16: 1535
Varma RS (1999) Green Chem. 43
Obermayer D, Gutmann B, Kappe CO (2009) Angew. Chem. Int. Ed. 48: 8321
de la Hoz A, Diaz-Ortiz A, Moreno A (2005) Chem. Soc. Rev. 34: 164
Loupy A, Petit A, Hamelin J, Texier-Boullet F, Jacquault P, Mathe D (1998) Synthesis 1213
Parker MC, Besson T, Lamare S, Legoy MD (1996) Tetrahedron Lett. 37: 8383
Carrillo-Munoz JR, Bouvet D, Guibe-Jampel E, Loupy A, Petit A (1996) J. Org. Chem. 61: 7746
Bornscheuer UT, Kazlauskas RJ (2006) Hydrolases in Organic Synthesis, 2nd ed. Wiley-VCH Weinheim, p 43
Grunwald P (2009) Biocatalysis, Imperial College Press, p 777
Bommarius AS, Riebel BR (2004) Biocatalysis. Wiley-VCH, Weinheim, p 61
Penning TM, Jez JM (2001) Chem. Rev. 101: 3027
Reetz MT, Zonta A, Schimossek K, Liebeton K, Jaeger KE (1997) Angew. Chem. Int. Ed. 36: 2830
Toscano MD, Woycechowsky KJ, Hilvert D (2007) Angew. Chem. Int. Ed. 46: 3212
Gerlt JA, Babbitt PC (2009) Curr. Opinion Chem. Biol. 13: 10
Morley KL, Kazlauskas RJ (2005) Trends Biotechnol. 23: 231
Shivange AV, Marienhagen J, Mundhada H, Schenk A, Schwaneberg U (2009) Curr. Opinion Chem. Biol. 13: 19
Baumann M, Stürmer R, Bornscheuer UT (2001) Angew. Chem. Int. Ed. 40: 4201
Reetz MT, Becker MH, Kühling KM, Holzwarth A (1998) Angew. Chem. Int. Ed. 37: 2647
Turner NJ (2009) Nature Chem. Biol. 5: 567
Reetz MT (2006) Adv. Catal. 49: 1
Demirjian DC, Shah PC, Moris-Varas F (1999) Top. Curr. Chem. 200: 1
Wahler D, Reymond JL (2001) Curr. Opinion Chem. Biol. 5: 152
Reetz MT (2002) Tetrahedron 58: 6595
Kourist R, Bartsch S, Bornscheuer UT (2007) Adv. Synth. Catal. 349: 1391
Henke E, Pleiss J, Bornscheuer UT (2003) ChemBioChem 4: 485
Bartsch S, Kourist R, Bornscheuer UT (2008) Angew. Chem. Int. Ed. 47: 1508
Böttcher D, Bornscheuer UT (2006) Nature Protocols 1: 2340
Jansonius JN (1987) Enzyme mechanism: what X-ray crystallography can(not) tell us. In: Moras D, Drenth J, Strandlberg B, Suck D, Wilson K (eds) Crystallography in Molecular Biology. Plenum Press, New York, p 229
Rubin B (1994) Struct. Biol. 1: 568
Heliwell JR, Helliwell M (1996) Chem. Commun. 1595
Fersht A (1977) Enzyme Structure and Mechanism. Freeman, San Francisco, p 15
Schrag JD, Cygler M (1993) J. Mol. Biol. 230: 575
Grochulski P, Li Y, Schrag JD, Bouthillier F, Smith P, Harrison D, Rubin B, Cygler M (1993) J. Biol. Chem. 268: 12843
Uppenberg J, Hansen MT, Patkar S, Jones TA (1994) Structure 2: 293
Noble MEM, Cleasby A, Johnson LN, Egmond MR, Frenken LGJ (1993) FEBS Lett. 331: 123
Burkert U, Allinger NL (1982) Molecular mechanics; In: Caserio MC (ed) ACS Monograph, vol 177. ACS, Washington
Norin M, Hult K, Mattson A, Norin T (1993) Biocatalysis 7: 131
Fitzpatrick PA, Ringe D, Klibanov AM (1992) Biotechnol. Bioeng. 40: 735
Ortiz de Montellano PR, Fruetel JA, Collins JR, Camper DL, Loew GH (1991) J. Am. Chem. Soc. 113: 3195
Oberhauser T, Faber K, Griengl H (1989) Tetrahedron 45: 1679
Kazlauskas RJ, Weissfloch ANE, Rappaport AT, Cuccia LA (1991) J. Org. Chem. 56: 2656
Karasaki Y, Ohno M (1978) J. Biochem. (Tokyo) 84: 531
Ahmed SN, Kazlauskas RJ, Morinville AH, Grochulski P, Schrag JD, Cygler M (1994) Biocatalysis 9: 209
Provencher L, Wynn H, Jones JB, Krawczyk AR (1993) Tetrahedron Asymmetry 4: 2025
Desnuelle P (1972) The lipases. In: Boyer PO (ed) The Enzymes, vol 7. Academic Press, New York, p 575
Wooley P, Petersen SB (eds) (1994) Lipases, their Structure, Biochemistry and Applications. Cambridge UP, Cambridge
Macrae AR (1983) J. Am. Oil Chem. Soc. 60: 291
Hanson M (1990) Oils Fats Int. 5: 29
Nielsen T (1985) Fette Seifen Anstrichmittel 87: 15
Jaeger KE, Reetz MT (1998) Trends Biotechnol. 16: 396
Schmid RD, Verger R (1998) Angew. Chem. Int. Ed. 37: 1608
Alberghina L, Schmidt RD, Verger R (eds) (1991) Lipases: Structure, Mechanism and Genetic Engineering. GBF Monographs, vol 16. Verlag Chemie, Weinheim
Ransac S, Carriere F, Rogalska E, Verger R, Marguet F, Buono G, Pinho Melo E, Cabral JMS, Egloff MPE, van Tilbeurgh H, Cambillau C (1996) The Kinetics, Specificites and Structural Features of Lipases. In: Op den Kamp AF (ed) Molecular Dynamics of Biomembranes. NATO ASI Ser, vol H 96. Springer, Heidelberg, pp 265–304
Sarda L, Desnuelle P (1958) Biochim. Biophys. Acta 30: 513
Schonheyder F, Volqvartz K (1945) Acta Physiol. Scand. 9: 57
Verger R (1997) Trends Biotechnol. 15: 32
Theil F (1995) Chem. Rev. 95: 2203
Bianchi D, Cesti P (1990) J. Org. Chem. 55: 5657
Iriuchijima S, Kojima N (1981) J. Chem. Soc., Chem. Commun. 185
Derewenda ZS, Wei Y (1995) J. Am. Chem. Soc. 117: 2104
Nagao Y, Kume M, Wakabayashi RC, Nakamura T, Ochiai M (1989) Chem. Lett. 239
Kazlauskas RJ, Weissfloch ANE (1997) J. Mol. Catal. B 3: 65
Kato K, Gong Y, Tanaka S, Katayama M, Kimoto H (1999) Biotechnol. Lett. 21: 457
Mugford P, Wagner U, Jiang Y, Faber K, Kazlauskas RJ (2008) Angew. Chem. Int. Ed. 47: 8782
Effenberger F, Gutterer B, Ziegler T, Eckhart E, Aichholz R (1991) Liebigs Ann. Chem. 47
Brockerhoff H (1968) Biochim. Biophys. Acta 159: 296
Brockmann HL (1981) Methods Enzymol. 71: 619
Desnuelle P (1961) Adv. Enzymol. 23: 129
Servi S (1999) Top. Curr. Chem. 200: 127
D’Arrigo P, Servi S (1997) Trends Biotechnol. 15: 90
Hultin PG, Jones JB (1992) Tetrahedron Lett. 33: 1399
Wimmer Z (1992) Tetrahedron 48: 8431
Claßen A, Wershofen S, Yusufoglu A, Scharf HD (1987) Liebigs Ann. Chem. 629
Jones JB, Hinks RS (1987) Can. J. Chem. 65: 704
Kloosterman H, Mosmuller EWJ, Schoemaker HE, Meijer EM (1987) Tetrahedron Lett. 28: 2989
Shaw JF, Klibanov AM (1987) Biotechnol. Bioeng. 29: 648
Sweers HM, Wong CH (1986) J. Am. Chem. Soc. 108: 6421
Hennen WJ, Sweers HM, Wang YF, Wong CH (1988) J. Org. Chem. 53: 4939
Waldmann H, Sebastian D (1994) Chem. Rev. 94: 911
Ballesteros A, Bernabé M, Cruzado C, Martin-Lomas M, Otero C (1989) Tetrahedron 45: 7077
Guibé-Jampel E, Rousseau G, Salaun J (1987) J. Chem. Soc., Chem. Commun. 1080
Cohen SG, Milovanovic A (1968) J. Am. Chem. Soc. 90: 3495
Laumen K, Schneider M (1985) Tetrahedron Lett. 26: 2073
Hemmerle H, Gais HJ (1987) Tetrahedron Lett. 28: 3471
Banfi L, Guanti G (1993) Synthesis 1029
Guanti G, Banfi L, Narisano E (1990) Tetrahedron Asymmetry 1: 721
Guanti G, Narisano E, Podgorski T, Thea S, Williams A (1990) Tetrahedron 46: 7081
Patel RN, Robison RS, Szarka LJ (1990) Appl. Microbiol. Biotechnol. 34: 10
Kerscher V, Kreiser W (1987) Tetrahedron Lett. 28: 531
Breitgoff D, Laumen K, Schneider MP (1986) J. Chem. Soc., Chem. Commun. 1523
Gao Y, Hanson RM, Klunder JM, Ko SY, Masamune H, Sharpless KB (1987) J. Am. Chem. Soc. 109: 5765
Marples BA, Roger-Evans M (1989) Tetrahedron Lett. 30: 261
Ladner WE, Whitesides GM (1984) J. Am. Chem. Soc. 106: 7250
Ramos-Tombo GM, Schär HP, Fernandez i Busquets X, Ghisalba O (1986) Tetrahedron Lett. 27: 5707
Bornemann S, Crout DHG, Dalton H, Hutchinson DW (1992) Biocatalysis 5: 297
Palomo JM, Segura RL, Mateo C, Terreni M, Guisan JM, Fernandez-Lafuente R (2005) Tetrahedron Asymmetry 16: 869
Cotterill IC, Sutherland AG, Roberts SM, Grobbauer R, Spreitz J, Faber K (1991) J. Chem. Soc., Perkin Trans. 1, 1365
Cotterill IC, Dorman G, Faber K, Jaouhari R, Roberts SM, Scheinmann F, Spreitz J, Sutherland AG, Winders JA, Wakefield BJ (1990) J. Chem. Soc., Chem. Commun. 1661
De Jersey J, Zerner B (1969) Biochemistry 8: 1967
Crich JZ, Brieva R, Marquart P, Gu RL, Flemming S, Sih CJ (1993) J. Org. Chem. 58: 3252
Cotterill IC, Finch H, Reynolds DP, Roberts SM, Rzepa HS, Short KM, Slawin AMZ, Wallis CJ, Williams DJ (1988) J. Chem. Soc., Chem. Commun. 470
Naemura K, Matsumura T, Komatsu M, Hirose Y, Chikamatsu H (1988) J. Chem. Soc., Chem. Commun. 239
Pearson AJ, Lai YS (1988) J. Chem. Soc., Chem. Commun. 442
Pearson AJ, Lai YS, Lu W, Pinkerton AA (1989) J. Org. Chem. 54: 3882
Gautier A, Vial C, Morel C, Lander M, Näf F (1987) Helv. Chim. Acta 70: 2039
Pawlak JL, Berchtold GA (1987) J. Org. Chem. 52: 1765
Sugai T, Kakeya H, Ohta H (1990) J. Org. Chem. 55: 4643
Kitazume T, Sato T, Kobayashi T, Lin JT (1986) J. Org. Chem. 51: 1003
Abramowicz DA, Keese CR (1989) Biotechnol. Bioeng. 33: 149
Hoshino O, Itoh K, Umezawa B, Akita H, Oishi T (1988) Tetrahedron Lett. 29: 567
Sugai T, Kakeya H, Ohta H, Morooka M, Ohba S (1989) Tetrahedron 45: 6135
Pottie M, Van der Eycken J, Vandevalle M, Dewanckele JM, Röper H (1989) Tetrahedron Lett. 30: 5319
Dumortier L, Van der Eycken J, Vandewalle M (1989) Tetrahedron Lett. 30: 3201
Klempier N, Geymeyer P, Stadler P, Faber K, Griengl H (1990) Tetrahedron Asymmetry 1: 111
Yamaguchi Y, Komatsu O, Moroe T (1976) J. Agric. Chem. Soc. Jpn. 50: 619
Cygler M, Grochulski P, Kazlauskas RJ, Schrag JD, Bouthillier F, Rubin B, Serreqi AN, Gupta AK (1994) J. Am. Chem. Soc. 116: 3180
Hönig H, Seufer-Wasserthal P (1990) Synthesis 1137
Pai YC, Fang JM, Wu SH (1991) J. Org. Chem. 59: 6018
Saf R, Faber K, Penn G, Griengl H (1988) Tetrahedron 44: 389
Königsberger K, Faber K, Marschner C, Penn G, Baumgartner P, Griengl H (1989) Tetrahedron 45: 673
Hult K, Norin T (1993) Indian J. Chem. 32B: 123
Wu SH, Guo ZW, Sih CJ (1990) J. Am. Chem. Soc. 112: 1990
Allenmark S, Ohlsson A (1992) Biocatalysis 6: 211
Colton IJ, Ahmed SN, Kazlauskas RJ (1995) J. Org. Chem. 60: 212
Uppenberg J, Patkar S, Bergfors T, Jones TA (1994) J. Mol. Biol. 235: 790
Patkar SA, Björkling F, Zyndel M, Schulein M, Svendsen A, Heldt-Hansen HP, Gormsen E (1993) Indian J. Chem. 32B: 76
Rogalska E, Cudrey C, Ferrato F, Verger R (1993) Chirality 5: 24
Hoegh I, Patkar S, Halkier T, Hansen MT (1995) Can. J. Bot. 73 (Suppl 1): S869
Anderson EM, Larsson KM, Kirk O (1998) Biocatalysis Biotrans. 16: 181
Martinelle M, Hult K (1995) Biochim. Biophys. Acta 1251: 191
Uppenberg J, Öhrner N, Norin M, Hult K, Kleywegt GJ, Patkar S, Waagen V, Anthonsen T, Jones A (1995) Biochemistry 34: 16838
Holla EW, Rebenstock HP, Napierski B, Beck G (1996) Synthesis 823
Ohtani T, Nakatsukasa H, Kamezawa M, Tachibana H, Naoshima Y (1997) J. Mol. Catal. B 4: 53
Sanchez VM, Rebolledo F, Gotor V (1997) Tetrahedron Asymmetry 8: 37
Konegawa T, Ohtsuka Y, Ikeda H, Sugai T, Ohta H (1997) Synlett. 1297
Adam W, Diaz MT, Saha-Möller CR (1998) Tetrahedron Asymmetry 9: 791
Mulvihill MJ, Gage JL, Miller MJ (1998) J. Org. Chem. 63: 3357
Kingery-Wood J, Johnson JS (1996) Tetrahedron Lett. 37: 3975
Hansen TV, Waagen V, Partali V, Anthonsen HW, Anthonsen T (1995) Tetrahedron Asymmetry 6: 499
Waagen V, Hollingsaeter I, Partali V, Thorstad O, Anthonsen T (1993) Tetrahedron Asymmetry 4: 2265
Kato K, Katayama M, Fujii S, Kimoto H (1996) J. Ferment. Bioeng. 82: 355
Xie ZF (1991) Tetrahedron Asymmetry 2: 733
Schrag JD, Li Y, Cygler M, Lang D, Burgdorf T, Hecht HJ, Schmid R, Schomburg D, Rydel TJ, Oliver JD, Strickland LC, Dunaway CM, Larson SB, Day J, McPherson A (1997) Structure 5: 187
Kim KK, Song HK, Shin DH, Hwang KY, Suh SW (1996) Structure 5: 173
Xie ZF, Nakamura I, Suemune H, Sakai K (1988) J. Chem. Soc., Chem. Commun. 966
Xie ZF, Sakai K (1989) Chem. Pharm. Bull. 37: 1650
Seemayer R, Schneider MP (1990) J. Chem. Soc., Perkin Trans. 1, 2359
Laumen K, Schneider MP (1988) J. Chem. Soc., Chem. Commun. 598
Kalaritis P, Regenye RW, Partridge JJ, Coffen DL (1990) J. Org. Chem. 55: 812
Klempier N, Geymayer P, Stadler P, Faber K, Griengl H (1990) Tetrahedron Asymmetry 1: 111
Kloosterman M, Kierkels JGT, Guit RPM, Vleugels LFW, Gelade ETF, van den Tweel WJJ, Elferink VHM, Hulshof LA, Kamphuis J (1991) Lipases: biotransformations, active site models and kinetics. In: Alberghina L, Schmidt RD, Verger R (eds) Lipases: Structure, Mechanism and Genetic Engineering GBF Monographs, vol 16. Verlag Chemie, Weinheim, p 187
Lemke K, Lemke M, Theil F (1997) J. Org. Chem. 62: 6268
Johnson CR, Adams JP, Bis SJ, De Jong RL, Golebiowski A, Medich JR, Penning TD, Senanayake CH, Steensma DH, Van Zandt MC (1993) Indian J. Chem. 32B: 140
Hughes DL, Bergan JJ, Amato JS, Bhupathy M, Leazer JL, McNamara JM, Sidler DR, Reider PJ, Grabowski EJJ (1990) J. Org. Chem. 55: 6252
Mizuguchi E, Takemoto M, Achiwa K (1993) Tetrahedron Asymmetry 4: 1961
Ors M, Morcuende A, Jimenez-Vacas MI, Valverde S, Herradon B (1996) Synlett. 449
Soriente A, Laudisio G, Giorgano M, Sodano G (1995) Tetrahedron Asymmetry 6: 859
Burgess K, Henderson I (1989) Tetrahedron Lett. 30: 3633
Ghiringhelli O (1983) Tetrahedron Lett. 24: 287
Hanessian S, Kloss J (1985) Tetrahedron Lett. 26: 1261
Bianchi D, Cesti P, Golini P (1989) Tetrahedron 45: 869
Itoh T, Takagi Y, Nishiyama S (1991) J. Org. Chem. 56: 1521
Allen JV, Williams JMJ (1996) Tetrahedron Lett. 37: 1859
Scilimati A, Ngooi T K, Sih CJ (1998) Tetrahedron Lett. 29: 4927
Bänzinger M, Griffiths GJ, McGarrity JF (1993) Tetrahedron Asymmetry 4: 723
Liang S, Paquette L A (1990) Tetrahedron Asymmetry 1: 445
Waldinger C, Schneider M, Botta M, Corelli F, Summa V (1996) Tetrahedron Asymmetry 7: 1485
Patel RN, Banerjee A, Ko RY, Howell JM, Li WS, Comezoglu FT, Partyka RA, Szarka L (1994) Biotechnol. Appl. Biochem. 20: 23
Takano S, Yamane T, Takahashi M, Ogasawara K (1992) Tetrahedron Asymmetry 3: 837
Takano S, Yamane T, Takahashi M, Ogasawara K (1992) Synlett 410
Gaucher A, Ollivier J, Marguerite J, Paugam R, Salaun J (1994) Can. J. Chem. 72: 1312
Xie ZF, Suemune H, Sakai K (1993) Tetrahedron Asymmetry 4: 973
Huge Jensen B, Galluzzo DR, Jensen RG (1987) Lipids 22: 559
Chan C, Cox PB, Roberts SM (1988) J. Chem. Soc., Chem. Commun. 971
Estermann H, Prasad K, Shapiro MJ (1990) Tetrahedron Lett. 31: 445
Hilgenfeld R, Saenger W (1982) Top. Curr. Chem. 101: 1
Naemura K, Takahashi N, Chikamatsu H (1988) Chem. Lett. 1717
Janes LE, Kazlauskas RJ (1997) Tetrahedron: Asymmetry 8: 3719
Itoh T, Kuroda K, Tomosada M, Takagi Y (1991) J. Org. Chem. 56: 797
Tinapp P (1971) Chem. Ber. 104: 2266
Satoh T, Suzuki S, Suzuki Y, Miyaji Y, Imai Z (1969) Tetrahedron Lett. 10: 4555
Effenberger F (1994) Angew. Chem. Int. Ed. 33: 1555
Mitsuda S, Yamamoto H, Umemura T, Hirohara H, Nabeshima S (1990) Agric. Biol. Chem. 54: 2907
Effenberger F, Stelzer U (1993) Chem. Ber. 126: 779
Mitsuda S, Nabeshima S, Hirohara H (1989) Appl. Microbiol. Biotechnol. 31: 334
Okumura S, Iwai M, Tsujisaka Y (1979) Biochim. Biophys. Acta 575: 156
Jensen RG (1974) Lipids 9: 149
Martinez C, Abergel C, Cambillau C, de Geus P, Lauwereys M (1991) Crystallographic study of a recombinant cutinase from Fusarium solani pisi. In: Alberghina L, Schmidt RD, Verger R (eds) Lipases: Structure, Mechanism and Genetic Engineering. GBF Monographs, vol 16. Verlag Chemie, Weinheim, p 67
Martinez C, Nicholas A, van Tilbeurgh H, Egloff MP, Cudrey C, Verger R, Cambillau C (1994) Biochemistry 33: 83
Martinez C, De Geus P, Lauwereys M, Matthysens G, Cambillau C (1992) Nature 356: 615
Dumortier L, Liu P, Dobbelaere S, Van der Eycken J, Vandewalle M (1992) Synlett. 243
Holmberg E, Hult K (1991) Biotechnol. Lett. 323
Holmberg E, Szmulik P, Norin T, Hult K (1989) Biocatalysis 2: 217
Itoh T, Ohira E, Takagi Y, Nishiyama S, Nakamura K (1991) Bull. Chem. Soc. Jpn. 64: 624
Bamann E, Laeverenz P (1930) Ber. dtsch. chem. Ges. 63: 394
Inada Y, Yoshimoto T, Matsushima A, Saito Y (1986) Trends Biotechnol. 4: 68
Offord RE (1987) Protein Eng. 1: 151
Lundblad RL (1991) Chemical Reagents for Protein Modification, 2nd edn. CRC Press, Boca Raton
Gu QM, Sih CJ (1992) Biocatalysis 6: 115
Tuomi WV, Kazlauskas RJ (1999) J. Org. Chem. 64: 2638
Lin G, Lin WY, Shieh CT (1998) Tetrahedron Lett. 39: 8881
Shimizu S, Kataoka M (1996) Chimia 50: 409
Novick NJ, Tyler ME (1982) J. Bacteriol. 149: 364
Dong YH, Wang LH, Xu JL, Zhang HB, Zhang XF, Zhang LH (2001) Nature 411: 813
Gutman AL, Zuobi K, Guibe-Jampel E (1990) Tetrahedron Lett. 31: 2037
Kataoka M, Honda K, Shimizu S (2000) Eur. J. Biochem. 267: 3
Honda K, Kataoka K, Shimizu S (2002) Appl. Microbiol. Biotechnol. 60: 288
Onakunle OA, Knowles CJ, Bunch AW (1997) Enzyme Microb. Technol. 21: 245
Honda K, Tsubioi H, Minetoki T, Nose H, Sakamoto K, Kataoka M, Shimizu S (2005) Appl. Microbiol. Biotechnol. 66: 520
Kesseler M, Friedrich T, Höffken H W, Hasuer B (2002) Adv. Synth. Catal. 344: 1103
Fujii H, Koyama T, Ogura K (1982) Biochim. Biophys. Acta 712: 716
Durrwachter JR, Wong CH (1988) J. Org. Chem. 53: 4175
Straub A, Effenberger F, Fischer P (1990) J. Org. Chem. 55: 3926
Bednarski MD, Simon ES, Bischofberger N, Fessner WD, Kim MJ, Lees W, Saito T, Waldmann H, Whitesides GM (1989) J. Am. Chem. Soc. 111: 627
Schultz M, Waldmann H, Kunz H, Vogt W (1990) Liebigs Ann. Chem. 1019
Scollar MP, Sigal G, Klibanov AM (1985) Biotechnol. Bioeng. 27: 247
van Henk T, Hartog AF, Ruijssenaars HJ, Kerkman R, Schoemaker HE, Wever R (2007) Adv. Synth. Catal. 349: 1349
Herdewijn P, Balzarini J, De Clerq E, Vanderhaege H (1985) J. Med. Chem. 28: 1385
Borthwick AD, Butt S, Biggadike K, Exall AM, Roberts SM, Youds PM, Kirk BE, Booth BR, Cameron JM, Cox SW, Marr CLP, Shill MD (1988) J. Chem. Soc., Chem. Commun. 656
Langer RS, Hamilton BC, Gardner CR, Archer MD, Colton CC (1976) AIChE J. 22: 1079
Chenault HK, Simon ES, Whitesides GM (1988) Biotechnol. Gen. Eng. Rev. 6: 221
Pradines A, Klaébé A, Périé J, Paul F, Monsan P (1991) Enzyme Microb. Technol. 13: 19
Wong CH, Haynie SL, Whitesides GM (1983) J. Am. Chem. Soc. 105: 115
Hirschbein BL, Mazenod FP, Whitesides GM (1982) J. Org. Chem. 47: 3765
Simon ES, Grabowski S, Whitesides GM (1989) J. Am. Chem. Soc. 111: 8920
Crans DC, Whitesides GM (1983) J. Org. Chem. 48: 3130
Bolte J, Whitesides GM (1984) Bioorg. Chem. 12: 170
Kameda A, Shiba T, Kawazoe Y, Satoh Y, Ihara Y, Munekata M, Ishige K, Noguchi T (2001) J. Biosci. Bioeng. 91: 557
van Herk T, Hartog AF, van der Burg AM, Wever R (2005) Adv. Synth. Catal. 347: 1155
Kazlauskas RJ, Whitesides GM (1985) J. Org. Chem. 50: 1069
Marshall DL (1973) Biotechnol. Bioeng. 15: 447
Augé C, Mathieu C, Mérienne C (1986) Carbohydr. Res. 151: 147
Wong CH, Haynie SL, Whitesides GM (1982) J. Org. Chem. 5416
Cantoni GL (1975) Ann. Rev. Biochem. 44: 435
Baughn RL, Adalsteinsson O, Whitesides GM (1978) J. Am. Chem. Soc. 100: 304
Le Grand DM, Roberts SM (1993) J. Chem. Soc., Chem. Commun. 1284
Drueckhammer DG, Wong CH (1985) J. Org. Chem. 50: 5912
Chenault HK, Mandes RF, Hornberger KR (1997) J. Org. Chem. 62: 331
Pollak A, Baughn RL, Whitesides GM (1977) J. Am. Chem. Soc. 77: 2366
Wong CH, Whitesides GM (1981) J. Am. Chem. Soc. 103: 4890
Wong CH, Whitesides GM (1983) J. Org. Chem. 48: 3199
van Herk T, Hartog AF, Schoemaker H E, Wever R (2006) J. Org. Chem. 71: 6244
Gross A, Abril O, Lewis JM, Geresh S, Whitesides GM (1983) J. Am. Chem. Soc. 105: 7428
Thorner JW, Paulus H (1973) In: The Enzymes; Boyer PD (ed) Academic Press, New York, vol 8, p 487
Rios-Mercadillo VM, Whitesides GM (1979) J. Am. Chem. Soc. 101: 5829
Crans DC, Whitesides GM (1985) J. Am. Chem. Soc. 107: 7019
Crans DC, Whitesides GM (1985) J. Am. Chem. Soc. 107: 7008
Chenault HK, Chafin LF, Liehr S (1998) J. Org. Chem. 63: 4039
Eibl H (1980) Chem. Phys. Lipids 26: 405
Vasilenko I, Dekruijff B, Verkleij A (1982) Biochim. Biophys. Acta 685: 144
Schurig V, Betschinger F (1992) Chem. Rev. 92: 873
Kolb HC, Van Nieuenhze MS, Sharpless KB (1994) Chem. Rev. 94: 2483
Scott JW (1984) Chiral carbon fragments and their use in synthesis. In: Scott JW, Morrison JD (eds) Asymmetric Synthesis, vol 4. Academic Press, Orlando, p 5
Finn MG, Sharpless KB (1985) On the mechanism of asymmetric epoxidation with titanium-tartrate catalysts. In: Scott JW, Morrison JD (eds) Asymmetric Synthesis, vol 5. Academic Press, Orlando, p 247
Jacobsen EN, Zhang W, Muci AR, Ecker JR, Deng L (1991) J. Am. Chem. Soc. 113: 7063
Pedragosa-Moreau S, Archelas A, Furstoss R (1995) Bull. Chim. Soc. Fr. 132: 769
de Bont JAM (1993) Tetrahedron Asymmetry 4: 1331
Onumonu AN, Colocoussi N, Matthews C, Woodland MP, Leak DJ (1994) Biocatalysis 10: 211
Besse P, Veschambre H (1994) Tetrahedron 50: 8885
Hartmans S (1989) FEMS Microbiol. Rev. 63: 235
Tokunaga M, Larrow JF, Kakiuchi F, Jacobsen EN (1997) Science 277: 936
Lu AYH, Miwa GT (1980) Ann. Rev. Pharmacol. Toxicol. 20: 513
Seidegard J, De Pierre JW (1983) Biochim. Biophys. Acta 695: 251
Armstrong RN (1987) Crit. Rev. Biochem. 22: 39
Hanzlik RP, Heidemann S, Smith D (1978) Biochem. Biophys. Res. Commun. 82: 310
Arand M, Wagner H, Oesch F (1996) J. Biol. Chem. 271: 4223
Nardini M, Ridder IS, Rozeboom HJ, Kalk KH, Rink R, Janssen DB, Dijkstra BW (1999) J. Biol. Chem. 274: 14579
Tzeng HF, Laughlin LT, Lin S, Armstrong RN (1996) J. Am. Chem. Soc. 118: 9436
DuBois GC, Appella E, Levin W, Lu AYH, Jerina DM (1978) J. Biol. Chem. 253: 2932
Janssen DB, Pries F, van der Ploeg JR (1994) Annu. Rev. Microbiol. 48: 163
Verschueren KHG, Seljee F, Rozeboom HJ, Kalk KH, Dijkstra BW (1993) Nature 363: 693
Withers SG, Warren RAJ, Street IP, Rupitz K, Kempton JB, Aebersold R (1990) J. Am. Chem. Soc. 112: 5887
Arand M, Hallberg BM, Zou J, Bergfors T, Oesch F, van der Werf M, de Bont JAM, Jones TA, Mowbray SL (2003) EMBO J. 22: 2583
Bellucci G, Chiappe C, Cordoni A, Marioni F (1994) Tetrahedron Lett. 35: 4219
Escoffier B, Prome JC (1989) Bioorg. Chem. 17: 53
Mischitz M, Mirtl C, Saf R, Faber K (1996) Tetrahedron Asymmetry 7: 2041
Moussou P, Archelas A, Baratti J, Furstoss R (1998) Tetrahedron Asymmetry 9: 1539
Daiboun T, Elalaoui MA, Thaler-Dao H, Chavis C, Maury G (1993) Biocatalysis 7: 227
Lu AYH, Levin W (1978) Methods Enzymol. 52: 193
Berti G (1986) Enantio- and diastereoselectivity of microsomal epoxide hydrolase: potential applications to the preparation of non-racemic epoxides and diols. In: Schneider MP (ed) Enzymes as Catalysts in Organic Synthesis, NATO ASI Series, vol 178. Reidel, Dordrecht, p 349
Bellucci G, Chiappe C, Marioni F (1989) J. Chem. Soc., Perkin Trans. 1, 2369
Bellucci G, Capitani I, Chiappe C, Marioni F (1989) J. Chem. Soc., Chem. Commun. 1170
Watabe T, Suzuki S (1972) Biochem. Biophys. Res. Commun. 46: 1120
Weijers CAGM, De Haan A, De Bont JAM (1988) Appl. Microbiol. Biotechnol. 27: 337
Yamada Y, Kikuzaki H, Nakatani N (1992) Biosci. Biotechnol. Biochem. 56: 153
Kolattukudy PE, Brown L (1975) Arch. Biochem. Biophys. 166: 599
Kolattokudy PE, Brown L (1975) Arch. Biochem. Biophys. 166: 599
Pedragosa-Moreau S, Archelas A, Furstoss R (1996) Tetrahedron Lett. 37: 3319
Misawa E, Chan Kwo Chion CKC, Archer IV, Woodland MC, Zhou NY, Carter SF, Widdowson DA, Leak DJ (1998) Eur. J. Biochem. 253: 173
Morisseau C, Archelas A, Guitton C, Faucher D, Furstoss R (1999) Eur. J. Biochem. 263: 386
van der Werf MJ, Overkamp KM, de Bont JAM (1998) J. Bacteriol. 180: 5052
Mischitz M, Faber K, Willetts A (1995) Biotechnol. Lett. 17: 893
Kroutil W, Genzel Y, Pietzsch M, Syldatk C, Faber K (1998) J. Biotechnol. 61: 143
Svaving J, de Bont JAM (1998) Enzyme Microb. Technol. 22: 19
Archer IVJ (1997) Tetrahedron 53: 15617
Weijers CAGM, de Bont JAM (1999) J. Mol. Catal. B 6: 199
Orru RVA, Archelas A, Furstoss R, Faber K (1999) Adv. Biochem. Eng. Biotechnol. 63: 145
Faber K, Mischitz M, Kroutil W (1996) Acta Chem. Scand. 50: 249
Orru RVA, Faber K (1999) Curr. Opinion Chem. Biol. 3: 16
Botes AL, Steenkamp JA, Letloenyane MZ, van Dyk MS (1998) Biotechnol. Lett. 20: 427
Weijers CAGM (1997) Tetrahedron Asymmetry 8: 639
Botes AL, Weijers CAGM, van Dyk MS (1998) Biotechnol. Lett. 20: 421
Weijers CAGM, Botes AL, van Dyk MS, de Bont JAM (1998) Tetrahedron Asymmetry 9: 467
Morisseau C, Nellaiah H, Archelas A, Furstoss R, Baratti JC (1997) Enzyme Microb. Technol. 20: 446
Spelberg JHL, Rink R, Kellogg RM, Janssen DB (1998) Tetrahedron Asymmetry 9: 459
Grogan G, Rippe C, Willetts A (1997) J. Mol. Catal. B 3: 253
Pedragosa-Moreau S, Archelas A, Furstoss R (1996) Tetrahedron 52: 4593
Mischitz M, Kroutil W, Wandel U, Faber K (1995) Tetrahedron Asymmetry 6: 1261
Moussou P, Archelas A, Furstoss R (1998) Tetrahedron 54: 1563
Kroutil W, Mischitz M, Plachota P, Faber K (1996) Tetrahedron Lett. 37: 8379
Mischitz M, Faber K (1996) Synlett. 978
Archer IVJ, Leak DJ, Widdowson DA (1996) Tetrahedron Lett. 37: 8819
Steinreiber A, Mayer SF, Saf R, Faber K (2001) Tetrahedron Asymmetry 12: 1519
Imai K, Marumo S, Mori K (1974) J. Am. Chem. Soc. 96: 5925
Pedragosa-Moreau S, Archelas A, Furstoss R (1993) J. Org. Chem. 58: 5533
Orru RVA, Mayer SF, Kroutil W, Faber K (1998) Tetrahedron 54: 859
Kroutil W, Mischitz M, Faber K (1997) J. Chem. Soc., Perkin Trans. 1, 3629
Legras JL, Chuzel G, Arnaud A, Galzy P (1990) World Microbiol. Biotechnol. 6: 83
Solomonson LP (1981) Cyanide as a metabolic inhibitor. In: Vennesland B et al. (eds) Cyanide in Biology. Academic Press, London, p 11
Legras JL, Jory M, Arnaud A, Galzy P (1990) Appl. Microbiol. Biotechnol. 33: 529
Jallageas JC (1980) Adv. Biochem. Eng. 14: 1
Meth-Cohn O, Wang MX (1995) Tetrahedron Lett. 36: 9561
Hjort CM, Godtfredsen SE, Emborg C (1990) J. Chem. Technol. Biotechnol. 48: 217
Thompson LA, Knowles CJ, Linton EA, Wyatt JM (1988) Chem. Brit. 900
Nagasawa T, Yamada H (1989) Trends Biotechnol. 7: 153
Arnaud A, Galzy P, Jallageas JC (1976) Folia Microbiol. 21: 178
Nagasawa T, Takeuchi K, Yamada H (1988) Biochem. Biophys. Res. Commun. 155: 1008
Nagasawa T, Nanba H, Ryuno K, Takeuchi K, Yamada H (1987) Eur. J. Biochem. 162: 691
Brennan BA, Alms G, Nelson MJ, Durney LT, Scarrow RC (1996) J. Am. Chem. Soc. 118: 9194
Sugiura Y, Kuwahara J, Nagasawa T, Yamada H (1987) J. Am. Chem. Soc. 109: 5848
Nagume T (1991) J. Mol. Biol. 220: 221
Song L, Wang M, Shi J, Xue Z, Wang MX, Qian S (2007) Biochem. Biophys. Res. Commun. 362: 319
Nagashima S, Nakasako M, Dohmae N, Tsujimura M, Takio K, Odaka M, Yohda M, Kamiya N, Endo I (1998) Nature Struct. Biol. 5: 347
Huang W, Jia J, Cummings J, Nelson M, Schneider G, Lindqvist Y (1997) Structure 5: 691
Miyanaga A, Fushinobu S, Ito K, Wakagi T (2001) Biochem. Biophys. Res. Commun. 288: 1169
Endo I, Odaka M, Yohda M (1999) Trends Biotechnol. 17: 244
Odaka M, Fujii K, Hoshino M, Noguchi T, Tsujimura M, Nagashima S, Yohda M, Nagamune T, Inoue Y, Endo I (1997) J. Am. Chem. Soc. 119: 3785
Desai LV, Zimmer M (2004) J. Chem. Soc. Dalton Trans. 872
Kobayashi M, Shimizu S (1998) Nature Biotechnol. 16: 733
Kobayashi M, Shimizu S (2000) Curr. Opinion Chem. Biol. 4: 95
Asano Y, Fujishiro K, Tani Y, Yamada H (1982) Agric. Biol. Chem. 46: 1165
Layh N, Parratt J, Willetts A (1998) J. Mol. Catal. B 5: 476
Brenner C (2002) Curr. Opinion Struct. Biol. 12: 775
Kobayashi M, Goda M, Shimizu S (1998) Biochem. Biophys. Res. Commun. 253: 662
Ingvorsen K, Yde B, Godtfredsen SE, Tsuchiya RT (1988) Microbial hydrolysis of organic nitriles and amides. In: Cyanide Compounds in Biology. Ciba Foundation Symp. 140. Wiley, Chichester, p 16
Ohta H (1996) Chimia 50: 434
Wyatt JM, Linton EA (1988) The industrial potential of microbial nitrile biochemistry. In: Cyanide Compounds in Biology. Ciba Foundation Symp. 140, Wiley, Chichester, p 32
Nagasawa T, Yamada H (1990) Pure Appl. Chem. 62: 1441
De Raadt A, Klempier N, Faber K, Griengl H (1992) Microbial and enzymatic transformation of nitriles. In: Servi S (ed) Microbial Reagents in Organic Synthesis., NATO ASI Series C, vol 381. Kluwer, Dordrecht, p 209
Nagasawa T, Yamada H (1990) Large-scale bioconversion of nitriles into useful amides and acids. In: Abramowicz DA (ed) Biocatalysis.Van Nostrand Reinhold, New York, p 277
Sugai T, Yamazaki T, Yokohama M, Ohta H (1997) Biosci. Biotechnol. Biochem. 61: 1419
Crosby J, Moilliet J, Parratt JS, Turner NJ (1994) J. Chem. Soc., Perkin Trans. 1, 1679
Nazly N, Knowles CJ, Beardsmore AJ, Naylor WT, Corcoran EG (1983) J. Chem. Technol. Biotechnol. 33: 119
Knowles CJ, Wyatt JM (1988) World Biotechnol. Rep. 1: 60
Wyatt JM (1988) Microbiol. Sci. 5: 186
Ingvorsen K, Hojer-Pedersen B, Godtfredsen SE (1991) Appl. Environ. Microbiol. 57: 1783
Maestracci M, Thiéry A, Arnaud A, Galzy P (1988) Indian J. Microbiol. 28: 34
Bui K, Arnaud A, Galzy P (1982) Enzyme Microb. Technol. 4: 195
Lee CY, Chang HN (1990) Biotechnol. Lett. 12: 23
Asano Y, Yasuda T, Tani Y, Yamada H (1982) Agric. Biol. Chem. 46: 1183
Ryuno K, Nagasawa T, Yamada H (1988) Agric. Biol. Chem. 52: 1813
Nagasawa T, Yamada H (1988) Pure Appl. Chem. 62: 1441
Mauger J, Nagasawa T, Yamada H (1989) Tetrahedron 45: 1347
Mauger J, Nagasawa T, Yamada H (1988) J. Biotechnol. 8: 87
Nagasawa T, Mathew CD, Mauger J, Yamada H (1988) Appl. Environ. Microbiol. 54: 1766
Petersen M, Kiener A (1999) Green Chem. 4: 99
Kobayashi M, Nagasawa T, Yanaka N, Yamada H (1989) Biotechnol. Lett. 11: 27
Kobayashi M, Yanaka N, Nagasawa T, Yamada H (1990) J. Antibiot. 43: 1316
Mathew CD, Nagasawa T, Kobayashi M, Yamada H (1988) Appl. Environ. Microbiol. 54: 1030
Vaughan PA, Cheetham PSJ, Knowles CJ (1988) J. Gen. Microbiol. 134: 1099
Nagasawa T, Yamada H, Kobayashi M (1988) Appl. Microbiol. Biotechnol. 29: 231
Bengis-Garber C, Gutman AL (1988) Tetrahedron Lett. 29: 2589
Kobayashi M, Nagasawa T, Yamada H (1988) Appl. Microbiol. Biotechnol. 29: 231
Nishise H, Kurihara M, Tani Y (1987) Agric. Biol. Chem. 51: 2613
Meth-Cohn O, Wang MX (1997) J. Chem. Soc., Perkin Trans. 1, 3197
Taylor SK, Chmiel NH, Simons LJ, Vyvyan JR (1996) J. Org. Chem. 61: 9084
Kieny-L’Homme MP, Arnaud A, Galzy P (1981) J. Gen. Appl. Microbiol. 27: 307
Yokoyama M, Sugai T, Ohta H (1993) Tetrahedron Asymmetry 6: 1081
Stolz A, Trott S, Binder M, Bauer R, Hirrlinger B, Layh N, Knackmuss HJ (1998) J. Mol. Catal. B 5: 137
Maddrell SJ, Turner NJ, Kerridge A, Willetts AJ, Crosby J (1996) Tetrahedron Lett. 37: 6001
Robertson DE, Chaplin JA, DeSantis G, Podar M, Madden M, Chi E, Richardson T, Milan A, Miller M, Weiner DP, Wong K, McQuaid J, Farwell B, Preston L A, Tan X, Snead MA, Keller M, Mathur E, Kretz PL, Burk MJ, Short JM (2004) Appl. Environ. Microbiol. 70: 2429
Robertson DE, Steer BA (2004) Curr. Opinion Chem. Biol. 8: 141
DeSantis G, Zhu Z, Greenberg A, Wong K, Chaplin J, Hanson SR, Farwell B, Nicholson LW, Rand CL, Weiner DP, Robertson DE, Burk MJ (2002) J. Am. Chem. Soc. 124: 9024
Furuhashi et al. (1992) Appl. Microbiol. Biotechnol. 37: 184
Fukuda Y, Harada T, Izumi Y (1973) J. Ferment. Technol. 51: 393
Yamamoto K, Oishi K, Fujimatsu I, Komatsu I (1991) Appl. Environ. Microbiol. 57: 3028
Yamamoto K, Ueno Y, Otsubo K, Kawakami K, Komatsu K (1990) Appl. Environ. Microbiol. 56: 3125
Gröger H (2001) Adv. Synth. Catal. 343: 547
Macadam AM, Knowles CJ (1985) Biotechnol. Lett. 7: 865
Arnaud A, Galzy P, Jallageas JC (1980) Bull. Soc. Chim. Fr. II: 87
Bhalla TC, Miura A, Wakamoto A, Ohba Y, Furuhashi K (1992) Appl. Microbiol. Biotechnol. 37: 184
Choi SY, Goo YM (1986) Arch. Pharm. Res. 9: 45
Effenberger F, Böhme J (1994) Bioorg. Med. Chem. 2: 715
Martinkova L, Stolz A, Knackmuss HJ (1996) Biotechnol. Lett. 18: 1073
Yamamoto K, Komatsu KI (1991) Agric. Biol. Chem. 55: 1459
Fallon R D, Stieglitz B, Turner I (1997) Appl. Microbiol. Biotechnol. 47: 156
Kakeya H, Sakai N, Sugai T, Ohta H (1991) Tetrahedron Lett. 32: 1343
Bianchi D, Bosetti A, Cesti P, Franzosi G, Spezia S (1991) Biotechnol. Lett. 13: 241
Osprian I, Jarret C, Strauss U, Kroutil W, Orru RVA, Felfer U, Willetts AJ, Faber K (1999) J. Mol. Catal. B 6: 555
Layh N, Willetts A (1998) Biotechnol. Lett. 20: 329
References to Sect. 2.2
May SW, Padgette SR (1983) Biotechnology 677
Jones JB, Beck JF (1976) Asymmetric syntheses and resolutions using enzymes. In: Jones JB, Sih CJ, Perlman D (eds) Applications of Biochemical Systems in Organic Chemistry. Wiley, New York, p 236
Hummel W, Kula MR (1989) Eur. J. Biochem. 184: 1
Willner I, Mandler D (1989) Enzyme Microb. Technol. 11: 467
Chenault HK, Whitesides GM (1987) Appl. Biochem. Biotechnol. 14: 147
Wichmann R, Vasic-Racki D (2005) Adv. Biochem. Eng. Biotechnol. 92: 225
van der Donk WA, Zhao HM (2003) Curr. Opinion Biotechnol. 14: 421
Jones JB, Sneddon DW, Higgins W, Lewis AJ (1972) J. Chem. Soc., Chem. Commun. 856
Jensen MA, Elving PJ (1984) Biochim. Biophys. Acta 764: 310
Wienkamp R, Steckhan E (1982) Angew. Chem. Int. Ed. 21: 782
Simon H, Bader J, Günther H, Neumann S, Thanos J (1985) Angew. Chem. Int. Ed. 24: 539
Mandler D, Willner I (1986) J. Chem. Soc., Perkin Trans. 2, 805
Jones JB, Taylor KE (1976) Can. J. Chem. 54: 2069
Legoy MD, Laretta-Garde V, LeMoullec JM, Ergan F, Thomas D (1980) Biochimie 62: 341
Julliard M, Le Petit J, Ritz P (1986) Biotechnol. Bioeng. 28: 1774
van Eys J (1961) J. Biol. Chem. 236: 1531
Gupta NK, Robinson WG (1966) Biochim. Biophys. Acta 118: 431
Wang SS, King CK (1979) Adv. Biochem. Eng. Biotechnol. 12: 119
Karabatsos GL, Fleming JS, Hsi N, Abeles RH (1966) J. Am. Chem. Soc. 88: 849
Stampfer W, Kosjek B, Moitzi C, Kroutil W, Faber K (2002) Angew. Chem. Int. Ed. 41: 1014
Levy HR, Loewus FA, Vennesland B (1957) J. Am. Chem. Soc. 79: 2949
Tischer W, Tiemeyer W, Simon H (1980) Biochimie 62: 331
Wichmann R, Wandrey C, Bückmann AF, Kula MR (1981) Biotechnol. Bioeng. 23: 2789
Slusarczyk H, Pohl M, Kula MR (1998) In: Ballesteros A, Plou FJ, Iborra JL, Halling P (eds) Stability and Stabilisation of Biocatalysts. Elsevier: Amsterdam, p 331
Shaked Z, Whitesides GM (1980) J. Am. Chem. Soc. 102: 7104
Hummel W, Schütte H, Schmidt E, Wandrey C, Kula MR (1987) Appl. Microbiol. Biotechnol. 26: 409
Weuster-Botz D, Paschold H, Striegel B, Gieren H, Kula M-R, Wandrey C (1994) Chem. Ing. Tech. 17: 131
Seelbach K, Riebel B, Hummel W, Kula MR, Tishkov VI, Egorov AM, Wandrey C, Kragl U (1996) Tetrahedron Lett. 37: 1377
Tishkov VI, Galkin AG, Fedorchuk VV, Savitsky PA, Rojkova AM, Gieren H, Kula MR (1999) Biotechnol. Bioeng. 64: 187
Tishkov VI, Galkin AG, Marchenko GN, Tsygankov YD, Egorov AM (1993) Biotechnol. Appl. Biochem. 18: 201
Vandecasteele J-P, Lemal J (1980) Bull. Soc. Chim. Fr. 101
Wong C-H, Drueckhammer DG, Sweers HM (1985) J. Am. Chem. Soc. 107: 4028
Pollak A, Blumenfeld H, Wax M, Baughn RL, Whitesides GM (1980) J. Am. Chem. Soc. 102: 6324
Hirschbein BL, Whitesides GM (1982) J. Am. Chem. Soc. 104: 4458
Wong CH, Gordon J, Cooney CL, Whitesides GM (1981) J. Org. Chem. 46: 4676
Guiseley KB, Ruoff PM (1961) J. Org. Chem. 26: 1248
Johannes TW, Woodyer RD, Zhao H (2007) Biotechnol. Bioeng. 96: 18
Vrtis JM, White AK, Metcalf WW, van der Donk WA (2002) Angew. Chem. Int. Ed. 41: 3257
Costas AM, White A K, Metcalf WW (2001) J. Biol. Chem. 276: 17429
Woodyer RD, van der Donk W A, Zhao HM (2003) Biochemistry 42: 11604
Woodyer R, D van der Donk W A, Zhao HM (2006) Comb. Chem. High Throughput Screen. 9: 237
Johannes TW, Woodyer RD, Zhao HM (2005) Appl. Environ. Microbiol. 71: 5728
Dodds DR, Jones JB (1982) J. Chem. Soc., Chem. Commun. 1080
Wang SS, King C-K (1979) Adv. Biochem. Eng. 12: 119
Mansson MO, Larsson PO, Mosbach K (1982) Methods Enzymol. 89: 457
Wong CH, Whitesides GM (1983) J. Am. Chem. Soc. 105: 5012
Utaka M, Yano T, Ema T, Sakai T (1996) Chem. Lett. 1079
Danielsson B, Winquist F, Malpote JY, Mosbach K (1982) Biotechnol. Lett. 4: 673
Payen B, Segui M, Monsan P, Schneider K, Friedrich CG, Schlegel HG (1983) Biotechnol. Lett. 5: 463
Lee LG, Whitesides GM (1986) J. Org. Chem. 51: 25
Carrea G, Bovara R, Longhi R, Riva S (1985) Enzyme Microb. Technol. 7: 597
Matos JR, Wong CH (1986) J. Org. Chem. 51: 2388
Bednarski MD, Chenault HK, Simon ES, Whitesides GM (1987) J. Am. Chem. Soc. 109: 1283
Morokutti A, Lyskowski A, Sollner S, Pointner E, Fitzpatrick TB, Kratky C, Gruber K, Macheroux P (2005) Biochemistry 44: 13724
Riebel BR, Gibbs PR, Wellborn WB, Bommarius AS (2003) Adv. Synth. Catal. 345: 707
Ross R P, Claiborne A (1992) J. Mol. Biol. 227: 658
Ward DE, Donelly CJ, Mullendore ME, van der Oost J, de Vos WM, Crane EJ (2001) Eur. J. Biochem. 268: 5816
Riebel BR, Gibbs PR, Wellborn WB, Bommarius AS (2002) Adv. Synth. Catal. 3454: 1156
Lemière GL, Lepoivre JA, Alderweireldt FC (1985) Tetrahedron Lett. 26: 4527
Lemière GL (1986) Alcohol dehydrogenase catalysed oxidoreduction reactions in organic chemistry. In: Schneider MP (ed) Enzymes as Catalysts in Organic Synthesis, NATO ASI Series C, vol 178. Reidel, Dordrecht, p 19
Devaux-Basseguy R, Bergel A, Comtat M (1997) Enzyme Microb. Technol. 20: 248
Hummel W (1997) Adv. Biochem. Eng. Biotechnol. 58: 145
Hummel W (1999) New alcohol dehydrogenases for the synthesis of chiral compounds. In: Scheper T (ed) New Enzymes for Organic Synthesis. Springer, Heidelberg, pp 145–184
Prelog V (1964) Pure Appl. Chem. 9: 119
Peters J, Minuth T, Kula MR (1993) Biocatalysis 8: 31
Hou CT, Patel R, Barnabe N, Marczak I (1981) Eur. J. Biochem. 119: 359
Bradshaw CW, Hummel W, Wong CH (1992) J. Org. Chem. 57: 1532
Hummel W (1990) Appl. Microbiol. Biotechnol. 34: 15
Kula MR, Kragl U (2000) Dehydrogenases in the synthesis of chiral compounds. In: Patel RN (ed) Stereoselective Biocatalysis. Marcel Dekker, New York, p 839
Nakamura K, Miyai T, Kawai J, Nakajima N, Ohno A (1990) Tetrahedron Lett. 31: 1159
MacLeod R, Prosser H, Fikentscher L, Lanyi J, Mosher HS (1964) Biochemistry 3: 838
Prelog V (1964) Colloqu. Ges. Physiol. Chem. 14: 288
Bradshaw CW, Fu H, Shen GJ, Wong CH (1992) J. Org. Chem. 57: 1526
Roberts SM (1988) Enzymes as catalysts in organic synthesis; In: Cooper A, Houben JL, Chien LC (eds) The Enzyme Catalysis Process. NATO ASI Series A, vol 178. Reidel, Dordrecht, p 443
Lepoivre JA (1984) Janssen Chim. Acta 2: 20
Plant A (1991) Pharm. Manufact. Rev., March: 5
Olson LP, Luo J, Almarsson Ö, Bruice TC (1996) Biochemistry 35: 9782
Cedergen-Zeppezauer ES, Andersson I, Ottonello S (1985) Biochemistry 24: 4000
Ganzhorn AJ, Green DW, Hershey AD, Gould RM, Plapp BV (1987) J. Biol. Chem. 262: 3754
Jones JB, Schwartz HM (1981) Can. J. Chem. 59: 1574
Van Osselaer TA, Lemière GL, Merckx EM, Lepoivre JA, Alderweireldt FC (1978) Bull. Soc. Chim. Belg. 87: 799
Jones JB, Takemura T (1984) Can. J. Chem. 62: 77
Davies J, Jones JB (1979) J. Am. Chem. Soc. 101: 5405
Lam LKP, Gair IA, Jones JB (1988) J. Org. Chem. 53: 1611
Krawczyk AR, Jones JB (1989) J. Org. Chem. 54: 1795
Irwin AJ, Jones JB (1976) J. Am. Chem. Soc. 98: 8476
Sadozai SK, Merckx EM, Van De Val AJ, Lemière GL, Esmans EL, Lepoivre JA, Alderweireldt FC (1982) Bull. Soc. Chim. Belg. 91: 163
Nakazaki M, Chikamatsu H, Fujii T, Sasaki Y, Ao S (1983) J. Org. Chem. 48: 4337
Nakazaki M, Chikamatsu H, Naemura K, Suzuki T, Iwasaki M, Sasaki Y, Fujii T (1981) J. Org. Chem. 46: 2726
Nakazaki M, Chikamatsu H, Sasaki Y (1983) J. Org. Chem. 48: 2506
Matos JR, Smith MB, Wong CH (1985) Bioorg. Chem. 13: 121
Takemura T, Jones JB (1983) J. Org. Chem. 48: 791
Haslegrave JA, Jones JB (1982) J. Am. Chem. Soc. 104: 4666
Fries RW, Bohlken DP, Plapp BV (1979) J. Med. Chem. 22: 356
Dodds DR, Jones JB (1988) J. Am. Chem. Soc. 110: 577
Yamazaki Y, Hosono K (1988) Tetrahedron Lett. 29: 5769
Yamazaki Y, Hosono K (1989) Tetrahedron Lett. 30: 5313
Horjales E, Bränden CI (1985) J. Biol. Chem. 260: 15445
Jones JB, Jakovac IJ (1982) Can. J. Chem. 60: 19
Nakazaki M, Chikamatsu H, Naemura K, Sasaki Y, Fujii T (1980) J. Chem. Soc., Chem. Commun. 626
Lemière GL, Van Osselaer TA, Lepoivre JA, Alderweireldt FC (1982) J. Chem. Soc., Perkin Trans. 2, 1123
Secundo F, Phillips RS (1996) Enzyme Microb. Technol. 19: 487
Keinan E, Sinha SC, Sinha-Bagchi A (1991) J. Chem. Soc., Perkin Trans. 1, 3333
Keinan E, Seth KK, Lamed R, Ghirlando R, Singh SP (1990) Biocatalysis 3: 57
Rothig TR, Kulbe KD, Buckmann F, Carrea G (1990) Biotechnol. Lett. 12: 353
Keinan E, Sinha SC, Singh SP (1991) Tetrahedron 47: 4631
Edegger K, Stampfer W, Seisser B, Faber K, Mayer SF, Oehrlein R, Hafner A, Kroutil W (2006) Eur. J. Org. Chem. 1904
Kosjek B, Stampfer W, Pogorevc M, Goessler W, Faber K, Kroutil W (2004) Biotechnol. Bioeng. 86: 55
Stampfer W, Kosjek B, Faber K, Kroutil W (2003) J. Org. Chem. 68: 402
Drueckhammer DG, Sadozai SK, Wong CH, Roberts SM (1987) Enzyme Microb. Technol. 9: 564
Keinan E, Seth KK, Lamed R (1986) J. Am. Chem. Soc. 108: 3474
De Amici M, De Micheli C, Carrea G, Spezia S (1989) J. Org. Chem. 54: 2646
Drueckhammer DG, Barbas III CF, Nozaki K, Wong CH (1988) J. Org. Chem. 53: 1607
Kelly DR, Lewis JD (1991) J. Chem. Soc., Chem. Commun. 1330
Crans D, Marshman RW, Nielsen R, Felty I (1993) J. Org. Chem. 58: 2244
Schubert T, Hummerl W, Kula MR, Müller M (2001) Eur. J. Org. Chem. 4181
Kim MJ, Whitesides GM (1988) J. Am. Chem. Soc. 110: 2959
Kim M-J, Kim JY (1991) J. Chem. Soc., Chem. Commun. 326
Luyten MA, Bur D, Wynn H, Parris W, Gold M, Frieson JD, Jones JB (1989) J. Am. Chem. Soc. 111: 6800
Casy G, Lee TV, Lovell H (1992) Tetrahedron Lett. 33: 817
Schütte H, Hummel H, Kula MR (1984) Appl. Microbiol. Biotechnol. 19: 167
Hummel W, Schütte H, Kula MR (1985) Appl. Microbiol. Biotechnol. 21: 7
De Amici M, De Micheli C, Molteni G, Pitrè D, Carrea G, Riva S, Spezia S, Zetta L (1991) J. Org. Chem. 56: 67
Butt S, Davies HG, Dawson MJ, Lawrence GC, Leaver J, Roberts SM, Turner MK, Wakefield BJ, Wall WF, Winders JA (1987) J. Chem. Soc., Perkin Trans. 1, 903
Riva S, Ottolina G, Carrea G, Danieli B (1989) J. Chem. Soc., Perkin Trans. 1, 2073
Carrea G, Colombi F, Mazzola G, Cremonesi P, Antonini E (1979) Biotechnol. Bioeng. 21: 39
Butt S, Davies HG, Dawson MJ, Lawrence GC, Leaver J, Roberts SM, Turner MK, Wakefield BJ, Wall WF, Winders JA (1985) Tetrahedron Lett. 26: 5077
Leaver J, Gartenmann TCC, Roberts SM, Turner MK (1987) In: Laane C, Tramper J, Lilly MD (eds) Biocatalysis in Organic Media. p 411, Elsevier, Amsterdam
Nakamura K, Yoneda T, Miyai T, Ushio K, Oka S, Ohno A (1988) Tetrahedron Lett. 29: 2453
Fontana A (1984) Thermophilic Enzymes and Their Potential Use in Biotechnology. Dechema, Weinheim, p 221
Bryant FO, Wiegel J, Ljungdahl LG (1988) Appl. Environ. Microbiol. 54: 460
Pham VT, Phillips RS, Ljungdahl LG (1989) J. Am. Chem. Soc. 111: 1935
Daniel RM, Bragger J, Morgan HW (1990) Enzymes from extreme environments. In: Abramowicz DA (ed) Biocatalysis. Van Nostrand Reinhold, New York, p 243
Willaert JJ, Lemière GL, Joris LA, Lepoivre JA, Alderweideldt FC (1988) Bioorg. Chem. 16: 223
Pham VT, Phillips RS (1990) J. Am. Chem. Soc. 112: 3629
Chen CS, Zhou BN, Girdaukas G, Shieh WR, VanMiddlesworth F, Gopalan AS, Sih CJ (1984) Bioorg. Chem. 12: 98
Shieh WR, Gopalan AS, Sih CJ (1985) J. Am. Chem. Soc. 107: 2993
Hoffmann RW, Ladner W, Helbig W (1984) Liebigs Ann. Chem. 1170
Nakamura K, Ushio K, Oka S, Ohno A (1984) Tetrahedron Lett. 25: 3979
Fuganti C, Grasselli P, Casati P, Carmeno M (1985) Tetrahedron Lett. 26: 101
Nakamura K, Kawai Y, Oka S, Ohno A (1989) Tetrahedron Lett. 30: 2245
Christen M, Crout DHG (1987) Bioreactors and Biotransformations, In: Moody GW, Baker PB (eds) Elsevier, London, p 213
Sakai T, Nakamura T, Fukuda K, Amano E, Utaka M, Takeda A (1986) Bull. Chem. Soc. Jpn. 59: 3185
Buisson D, Azerad R, Sanner C, Larcheveque M (1991) Tetrahedron Asymmetry 2: 987
Ushio K, Inoue K, Nakamura K, Oka S, Ohno A (1986) Tetrahedron Lett. 27: 2657
Kometani T, Kitatsuji E, Matsuno R (1991) Agric. Biol. Chem. 55: 867
Dahl AC, Madsen JO (1998) Tetrahedron Asymmetry 9: 4395
Buisson D, Azerad R (1986) Tetrahedron Lett. 27: 2631
Seebach D, Züger MF, Giovannini F, Sonnleitner B, Fiechter A (1984) Angew. Chem. Int. Ed. 23: 151
Servi S (1990) Synthesis 1
Kometani T, Yoshii H, Matsuno R (1996) J. Mol. Catal. B 1: 45
Sih CJ, Chen CS (1984) Angew. Chem. Int. Ed. 23: 570
Ward OP, Young CS (1990) Enzyme Microb. Technol. 12: 482
Csuk R , Glänzer B (1991) Chem. Rev. 91: 49
Neuberg C, Lewite A (1918) Biochem. Z. 91: 257
Neuberg C (1949) Adv. Carbohydr. Chem. 4: 75
Ticozzi C, Zanarotti A (1988) Tetrahedron Lett. 29: 6167
Dondoni A, Fantin G, Fogagnolo M, Mastellari A, Medici A, Nefrini E, Pedrini P (1988) Gazz. Chim. Ital. 118: 211
Bucchiarelli M, Forni A, Moretti I, Prati F, Torre G, Resnati G, Bravo P (1989) Tetrahedron 45: 7505
Bernardi R, Bravo P, Cardillo R, Ghiringhelli D, Resnati G (1988) J. Chem. Soc., Perkin Trans. 1: 2831
Kitazume T, Kobayashi T (1987) Synthesis 87
Kitazume T, Nakayama Y (1986) J. Org. Chem. 51: 2795
Takano S, Yanase M, Sekiguchi Y, Ogasawara K (1987) Tetrahedron Lett. 28: 1783
Bucchiarelli M, Forni A, Moretti I, Torre G (1983) Synthesis 897
Kitazume T, Lin JT, (1987) J. Fluorine Chem. 34: 461
Fujisawa T, Hayashi H, Kishioka Y (1987) Chem. Lett. 129
Seebach D, Roggo S, Maetzke T, Braunschweiger H, Cerkus J, Krieger M (1987) Helv. Chim. Acta 70: 1605
Nakamura K, Inoue Y, Shibahara J, Oka S, Ohno A (1988) Tetrahedron Lett. 29: 4769
Guette JP, Spassky N (1972) Bull. Soc. Chim. Fr. 4217
Levene PA, Walti A (1943) Org. Synth., Coll. Vol. II: 545
Takano S (1987) Pure Appl. Chem. 59: 353
Itoh T, Yoshinaka A, Sato T, Fujisawa T (1985) Chem. Lett. 1679
Kozikowski AP, Mugrage BB, Li CS, Felder L (1986) Tetrahedron Lett. 27: 4817
Bernardi R, Ghiringhelli D (1987) J. Org. Chem. 52: 5021
Top S, Jaouen G, Gillois J, Baldoli C, Maiorana S (1988) J. Chem. Soc., Chem. Commun. 1284
Yamazaki Y, Hosono K (1988) Agric. Biol. Chem. 52: 3239
Syldatk C, Andree H, Stoffregen A, Wagner F, Stumpf B, Ernst L, Zilch H, Tacke R (1987) Appl. Microbiol. Biotechnol. 27: 152
Yamazaki Y, Kobayashi H (1993) Chem. Express 8: 97
Le Drian C, Greene AE (1982) J. Am. Chem. Soc. 104: 5473
Belan A, Bolte J, Fauve A, Gourcy JG, Veschambre H (1987) J. Org. Chem. 52: 256
Hirama M, Nakamine T, Ito S (1984) Chem. Lett. 1381
Deshong P, Lin M-T, Perez JJ (1986) Tetrahedron Lett. 27: 2091
Tschaen DM, Fuentes LM, Lynch JE, Laswell WL, Volante RP, Shinkai I (1988) Tetrahedron Lett. 29: 2779
Mori K (1989) Tetrahedron 45: 3233
Kramer A, Pfader H (1982) Helv. Chim. Acta 65: 293
Sih CJ, Zhou B, Gopalan AS, Shieh WR, VanMiddlesworth F (1983) Strategies for Controlling the Stereochemical Course of Yeast Reductions. In: Bartmann W, Trost BM (eds) Selectivity – a Goal for Synthetic Efficiency. Proc. 14th Workshop Conference Hoechst. Verlag Chemie, Weinheim, p 250
Heidlas J, Engel KH, Tressl R (1991) Enzyme Microb. Technol. 13: 817
Nakamura K, Inoue K, Ushio K, Oka S, Ohno A (1987) Chem. Lett. 679
Nakamura K, Kawai Y, Oka S, Ohno A (1989) Bull. Chem. Soc. Jpn. 62: 875
Nakamura K, Higaki M, Ushio K, Oka S, Ohno A (1985) Tetrahedron Lett. 26: 4213
Chibata I, Tosa T, Sato T (1974) Appl. Microbiol. 27: 878
Nakamura K, Kawai Y, Ohno A (1990) Tetrahedron Lett. 31: 267
Hayakawa R, Nozawa K, Kimura K, Shimizu M (1999) Tetrahedron 55: 7519
Ushio K, Hada J, Tanaka Y, Ebara K (1993) Enzyme Microb. Technol. 15: 222
Miya H, Kawada M, Sugiyama Y (1996) Biosci. Biotechnol. Biochem. 60: 95
Arnone A, Biagnini G, Cardillo R, Resnati G, Begue JP, Bonnet-Delpon D, Kornilov A (1996) Tetrahedron Lett. 37: 3903
Deol B, Ridley D, Simpson G (1976) Aust. J. Chem. 29: 2459
Nakamura K, Miyai T, Nozaki K, Ushio K, Ohno A (1986) Tetrahedron Lett. 27: 3155
Fujisawa T, Itoh T, Sato T (1984) Tetrahedron Lett. 25: 5083
Buisson D, Henrot S, Larcheveque M, Azerad R (1987) Tetrahedron Lett. 28: 5033
Akita H, Furuichi A, Koshoji H, Horikoshi K, Oishi T (1983) Chem. Pharm. Bull. 31: 4376
Frater G, Müller U, Günther W (1984) Tetrahedron 40: 1269
VanMiddlesworth F, Sih CJ (1987) Biocatalysis 1: 117
Shieh WR, Sih CJ (1993) Tetrahedron Asymmetry 4: 1259
Hoffmann RW, Helbig W, Ladner W (1982) Tetrahedron Lett. 23: 3479
Chenevert R, Thiboutot S (1986) Can. J. Chem. 64: 1599
Ohta H, Ozaki K, Tsuchihashi G (1986) Agric. Biol. Chem. 50: 2499
Nakamura K, Kawai Y, Ohno A (1991) Tetrahedron Lett. 32: 2927
Nakamura K, Miyai T, Nozaki K, Ushio K, Oka S, Ohno A (1986) Tetrahedron Lett. 27: 3155
Buisson D, Azerad R, Sanner C, Larcheveque M (1990) Biocatalysis 3: 85
Buisson D, Sanner C, Larcheveque M, Azerad R (1987) Tetrahedron Lett. 28: 3939
Cabon O, Buisson D, Lacheveque M, Azerad R (1995) Tetrahedron Asymmetry 6: 2199
Nishida T, Matsumae H, Machida I, Shibatani T (1995) Biocatalysis Biotrans. 12: 205
Sato T, Tsurumaki M, Fujisawa T (1986) Chem. Lett. 1367
Besse P, Veschambre H (1993) Tetrahedron Asymmetry 4: 1271
Soukup M, Wipf B, Hochuli E, Leuenberger HGW (1987) Helv. Chim. Acta 70: 232
Nakamura K, Inoue K, Ushio K, Oka S, Ohno A (1988) J. Org. Chem. 53: 2598
Brooks DW, Mazdiyasni H, Chakrabarti S (1984) Tetrahedron Lett. 25: 1241
Brooks DW, Woods KW (1987) J. Org. Chem. 52: 2036
Brooks DW, Mazdiyasni H, Grothaus PG (1987) J. Org. Chem. 52: 3223
Brooks DW, Mazdiyasni H, Sallay P (1985) J. Org. Chem. 50: 3411
Fujisawa T, Kojima E, Sato T (1987) Chem. Lett. 2227
Takeshita M, Sato T (1989) Chem. Pharm. Bull. 37: 1085
Kieslich K (1976) Microbial Transformations of Non-Steroid Cyclic Compounds. Thieme, Stuttgart
Besse P, Sokoltchik T, Veschambre H (1998) Tetrahedron Asymmetry 9: 4441
Tidswell EC, Salter GJ, Kell DB, Morris JG (1997) Enzyme Microb. Technol. 21: 143
Wipf B, Kupfer E, Bertazzi R, Leuenberger HGW (1983) Helv. Chim. Acta 66: 485
Bernardi R, Cardillo R, Ghiringhelli D, de Pavo V (1987) J. Chem. Soc., Perkin Trans. 1, 1607
Ikeda H, Sato E, Sugai T, Ohta H (1996) Tetrahedron 52: 8113
Fujisawa T, Onogawa Y, Sato A, Mitsuya T, Shimizu M (1998) Tetrahedron 54: 4267
Wei ZL, Li ZY, Lin GQ (1998) Tetrahedron 54: 13059
Fantin G, Fogagnolo M, Giovannini PP, Medici A, Pedrini P, Gardini F, Lanciotti R (1996) Tetrahedron 52: 3547
Akakabe Y, Naoshima Y (1993) Phytochemistry 32: 1189
Bruni R, Fantin G, Maietti S, Medici A, Pedrini P, Sacchetti G (2006) Tetrahedron Asymmetry 17: 2287
Vicenzi JT, Zmijewski MJ, Reinhard MR, Landen BE, Muth WL, Marler PG (1997) Enzyme Microb. Technol. 20: 494
Hummel W (1997) Biochem. Eng. 58: 145
Patel RN, Banerjee A, McNamee CG, Brzozowski D, Hanson RL, Szarka LJ (1993) Enzyme Microb. Technol. 15: 1014
Haberland J, Hummel W, Daußmann T, Liese A (2002) Org. Proc. Res. Dev. 6: 458
Schmidt E, Ghisalba O, Gygax D, Sedelmeier G (1992) J. Biotechnol. 24: 315
Kataoka M, Kita K, Wada M, Yasohara Y, Hasegawa J, Shimizu S (2003) Appl. Microbiol. Biotechnol. 62: 437
Carnell AJ (1999) Adv. Biochem. Eng. Biotechnol. 63: 57
Voss CV, Gruber CC, Kroutil W (2010) Synlett 991
Gadler P, Glueck S M, Kroutil W, Nestl BM, Larissegger-Schnell B, Ueberbacher BT, Wallner SR, Faber K (2006) Biochem. Soc. Trans 34: 296
Hasegawa J, Ogura M, Tsuda S, Maemoto S, Kutsuki H, Ohashi T (1990) Agric. Biol. Chem. 54: 1819
Azerad R, Buisson D (1992) In: Servi S (ed) Microbial Reagents in Organic Synthesis. NATO ASI Series C, vol 381, p. 421, Kluwer, Dordrecht
Buisson D, Azerad R, Sanner C, Lacheveque M (1992) Biocatalysis 5: 249
Nakamura K, Inoue Y, Matsuda T, Ohno A (1995) Tetrahedron Lett. 36: 6263
Fantin G, Fogagnolo M, Giovannini PP, Medici A, Pedrini, P (1995) Tetrahedron Asymmetry 6: 3047
Takahashi E, Nakamichi K, Furui M (1995) J. Ferment. Bioeng. 80: 247
Setyahadi S, Harada E, Mori N, Kitamoto Y (1998) J. Mol. Catal. B 4: 205
Azerad R, Buisson D (1992) Stereocontrolled reduction of β-ketoesters with Geotrichum candidum. In: Servi S (ed) Microbial Reagents in Organic Synthesis. NATO ASI Series C, vol 381. Kluwer, Dordrecht, pp 421–440
Voss CV, Gruber CC, Faber K, Knaus T, Macheroux P, Kroutil W (2008) J. Am. Chem. Soc. 130: 13969
Voss CV, Gruber CC, Kroutil W (2008) Angew. Chem. Int. Ed. 47: 741
Ogawa J, Xie SX, Shimizu S (1999) Biotechnol. Lett. 21: 331
Brown GM (1971) In: Florkin M, Stotz EH (eds) Comprehensive Biochemistry, vol. 21. Elsevier, New York, p 73.
Shimizu S, Hattori S, Hata H, Yamada Y (1987) Appl. Environ. Microbiol. 53: 519
Shimizu S, Hattori S, Hata H, Yamada H (1987) Enzyme Microb. Technol. 9: 411
Page PCB, Carnell AJ, McKenzie MJ (1998) Synlett. 774
Matsumura S, Kawai Y, Takahashi Y, Toshima K (1994) Biotechnol. Lett. 16: 485
Carnell AJ, Iacazio G, Roberts SM, Willetts AJ (1994) Tetrahedron Lett. 35: 331
Moon Kim B, Guare JP, Hanifin CM, Arford-Bickerstaff DJ, Vacca JP, Ball RG (1994) Tetrahedron Lett. 35: 5153
Ohshima T, Soda K (2000) Amino acid dehydrogenases and their applications. In: Patel RN (ed) Stereoselective Biocatalysis. Marcel Dekker, New York, p. 877
Sekimoto T, Matsuyama T, Fukui T, Tanizawa K (1993) J. Biol. Chem. 268: 27039
Bommarius AS, Schwarm M, Drauz K (1998) J. Mol. Catal. B: Enzym. 5: 1
Krix G, Bommarius AS, Drauz K, Kottenhahn M, Schwarm M, Kula MR (1997) J. Biotechnol. 53: 29
Kragl U, Vasic-Racki D, Wandrey C (1996) Bioproc. Eng. 14: 291
Yang JW, Hechavarria Fonseca MT, Vignola N, List B (2005) Angew. Chem. Int. Ed. 44:108
Williams RE, Bruce NC (2002) Microbiology 148: 1607
Steinbacher S, Stumpf M, Weinkauf S, Rohdich F, Bacher A, Simon H (2002) Enoate reductase family. In: Chapman SK, Perham RN, Scrutton NS (eds) Flavins and Flavoproteins. Weber, p 941
Warburg O, Christian W (1933) Biochem. Z. 266: 377
Barna T, Messiha HL, Petosa C, Bruce NC, Scrutton NS, Moody PCE (2002) J. Biol. Chem. 277: 30976
Schaller F, Biesgen C, Müssig C, Altmann T, Weiler EW (2000) Planta 210: 979
Vaz ADN, Chakraborty S, Massey V (1995) Biochemistry 34: 4246
Snape Jr, N. Walkley A, Morby AP, Nicklin S, White GF (1997) J. Bacteriol. 179: 7796
Nishino SF, Spain JC (1993) Appl. Environ. Microbiol. 59: 2520
Barna TM, Khan H, Bruce NC, Barsukov I, Scrutton NS, Moody PCE (2001) J. Mol. Biol. 310: 433
Kohli RM, Massey V (1998) J. Biol. Chem. 273: 32763
Shimoda K, Ito DI, Izumi S, Hirata T (1996) J. Chem. Soc. Perkin Trans. 1, 4: 355
Bougioukou DJ, Stewart JD (2008) J. Am. Chem. Soc. 130: 7655
Schlieben NH, Niefind K, Müller J, Riebel B, Hummel W, Schomburg D (2005) J. Mol. Biol. 349: 801
Kurata A, Kurihara T, Kamachi H, Esaki N (2004) Tetrahedron Asymmetry 15: 2837
Kataoka M, Kotaka A, Hasegawa A, Wada M, Yoshizumi A, Nakamori S, Shimizu S (2002) Biosci. Biotechnol. Biochem. 66: 2651
Williams RE, Rathbone DA, Scrutton NS, Bruce NC (2004) Appl. Environ. Microbiol. 70: 3566
Fuganti C, Grasselli P (1979) J. Chem. Soc. Chem. Commun. 995
Fischer FG, Wiedemann O (1934) Liebigs Ann. Chem. 513: 260
Desrut M, Kergomard A, Renard MF, Veschambre H (1981) Tetrahedron 37: 3825
Simon H, White H, Lebertz H, Thanos I (1987) Angew. Chem. Int. Ed. 26: 785
Simon H (1993) Indian J. Chem. 32B: 170
Hauer B, Stuermer R, Hall M, Faber K (2007) Curr. Opinion Chem. Biol. 11: 203
Hall M, Stueckler C, Kroutil W, Macheroux P, Faber K (2007) Angew. Chem. Int. Ed. 46: 3934
Toogood HS, Gardiner JM, Scrutton NS (2010) ChemCatChem 2: 892
Hall M, Stueckler C, Ehammer H, Pointner E, Oberdorfer G, Gruber K, Hauer B, Stuermer R, Kroutil W, Macheroux P, Faber K (2008) Adv. Synth. Catal. 350: 411
Hall M, Stueckler C, Hauer B, Stuermer R, Friedrich T, Breuer M, Kroutil W, Faber K (2008) Eur. J. Org. Chem. 1511
Mueller NJ, Stueckler C, Hauer B, Baudendistel N, Housden H, Bruce NC, Faber K (2010) Adv. Synth. Catal. 352: 387
Swiderska MA, Stewart JD (2006) J. Mol. Catal. B: Enzym. 42: 52
Swiderska MA, Stewart JD (2006) Org. Lett. 8: 6131
Fuganti C, Grasselli P (1989) Baker’s yeast mediated synthesis of natural products. In: Whitaker JR, Sonnet PE (eds) Biocatalysis in Agricultural Biotechnology, ACS Symp. Ser. 389. ACS, Washington, p 359
Suemune H, Hayashi N, Funakoshi K, Akita H, Oishi T, Sakai K (1985) Chem. Pharm. Bull. 33: 2168
Mueller A, Stuermer R, Hauer B, Rosche B (2007) Angew. Chem. Int. Ed. 46: 3316
Gil G, Ferre E, Barre M, Le Petit J (1988) Tetrahedron Lett. 29: 3797
Fuganti C, Grasselli P (1982) J. Chem. Soc., Chem. Commun. 205
Fuganti C, Grasselli P, Servi S (1983) J. Chem. Soc., Perkin Trans. 1, 241
Sato T, Hanayama K, Fujisawa T (1988) Tetrahedron Lett. 29: 2197
Kergomard A, Renard MF, Veschambre H (1982) J. Org. Chem. 47: 792
Sih CJ, Heather JB, Sood R, Price P, Peruzzotti G, Lee HFH, Lee SS (1975) J. Am. Chem. Soc. 97: 865
Durchschein K, Ferreira-da Silva B, Wallner S, Macheroux P, Kroutil W, Glueck S M, Faber K (2010) Green Chem. 12: 616
Ohta H, Ozaki K, Tsuchihashi G (1987) Chem. Lett. 191
Ferraboschi P, Grisenti P, Casati R, Fiecchi A, Santaniello E (1987) J. Chem. Soc., Perkin Trans. 1, 1743
Kitazume T, Ishikawa N (1984) Chem. Lett. 587
Gramatica P, Manitto P, Monti D, Speranza G (1987) Tetrahedron 43: 4481
Gramatica P, Manitto P, Ranzi BM, Delbianco A, Francavilla M (1983) Experientia 38: 775
Gramatica P, Manitto P, Monti D, Speranza G (1988) Tetrahedron 44: 1299
Leuenberger HG, Boguth W, Widmer E, Zell R (1976) Helv. Chim. Acta 59: 1832
Ohta H, Kobayashi N, Ozaki K (1989) J. Org. Chem. 54: 1802
Hall M, Stueckler C, Ehammer H, Pointner E, Kroutil W, Macheroux P, Faber K (2007) Org. Lett. 9: 5409
Utaka M, Konishi S, Mizuoka A, Ohkubo T, Sakai T, Tsuboi S, Takeda A (1989) J. Org. Chem. 54: 4989
Utaka M, Konishi S, Okubo T, Tsuboi S, Takeda A (1987) Tetrahedron Lett. 28: 1447
Takabe K, Hiyoshi H, Sawada H, Tanaka M, Miyazaki A, Yamada T, Kitagiri T, Yoda H (1992) Tetrahedron Asymmetry 3: 1399
Somers WAC, van Hartingsveldt W, Stigter ECA, van der Lugt JP (1997) Trends Biotechnol. 15: 495
References to Sect. 2.3
Glueck SM, Gümüs S, Fabian WMF, Faber K (2010) Chem. Soc. Rev. 39: 313
Fang JM, Lin CH, Bradshaw CW, Wong CH (1995) J. Chem. Soc., Perkin Trans. 1, 967
Schmid RD, Urlacher VB (eds) (2007) Modern Biooxidation Methods. Wiley-VCH, Weinheim
Fonken GS, Johnson RA (1972) Chemical oxidations with microorganisms. In: Belew JS (ed) Oxidation in Organic Chemistry, vol 2. Marcel Dekker, New York, p 185
Reichstein T (1934) Helv. Chim. Acta 17: 996
Sato K, Yamada Y, Aida K, Uemura T (1967) Agric. Biol. Chem. 31: 877
Touster O, Shaw DRD (1962) Physiol. Rev. 42: 181
Bernhauer K, Knobloch H (1940) Biochem. Z. 303: 308
Kaufmann H, Reichstein T (1967) Helv. Chim. Acta 50: 2280
Jones JB, Taylor KE (1976) Can. J. Chem. 54: 2969 and 2974
Irwin AJ, Jones JB (1977) J. Am. Chem. Soc. 99: 1625
Jones JB, Jacovac IJ (1990) Org. Synth., coll. vol. 7: 406
Drueckhammer DG, Riddle VW, Wong CH (1985) J. Org. Chem. 50: 5387
Wong C-H, Matos JR (1985) J. Org. Chem. 50: 1992
Irwin AJ, Jones JB (1977) J. Am. Chem. Soc. 99: 556
Jones JB, Lok KP (1979) Can. J. Chem. 57: 1025
Ng GSY, Yuan LC, Jakovac IJ, Jones JB (1984) Tetrahedron 40: 1235
Lok KP, Jakovac IJ, Jones JB (1985) J. Am. Chem. Soc. 107: 2521
Holland HL (1992) Organic Synthesis with Oxidative Enzymes. Verlag Chemie, Weinheim
Walsh C (1979) Enzymatic Reaction Mechanisms. Freeman, San Francisco, p 501
Dalton H (1980) Adv. Appl. Microbiol. 26: 71
Hayashi O (ed) (1974) The Molecular Mechanism of Oxygen Activation. Academic Press, New York
Gunsalus IC, Pederson TC, Sligar SG (1975) Ann. Rev. Microbiol. 377
Dawson JH (1988) Science 240: 433
Hou CT (1986) Biotechnol. Gen. Eng. Rev. 4: 145
Dunford HB (1982) Adv. Inorg. Biochem. 4: 41
Richter G (1983) Glucose oxidase. In: Godfrey T, Reichelt J (eds) Industrial Enzymology. Nature Press, New York
Szwajcer E, Brodelius P, Mosbach K (1982) Enzyme Microb. Technol. 4: 409
Dix TA, Benkovic SS (1988) Acc. Chem. Res. 21: 101
Massey V (2000) Biochem. Soc. Trans. 28: 283
Ziegler DM (1990) Trends Pharmacol. Sci. 11: 321
Massey V (1994) J. Biol. Chem. 269: 22459
Sato R, Omura T (eds) (1978) Cytochrome P-450. Academic Press, New York
Ortiz de Montellano PR (ed) (1986) Cytochrome P-450. Plenum Press, New York
Takemori S (1987) Trends Biochem. Sci. 12: 118
Dawson JH, Sono M (1987) Chem. Rev. 87: 1255
Müller HG (1990) Biocatalysis 4: 11
Alexander LS, Goff HM (1982) J. Chem. Educ. 59: 179
Poulos TL, Finzel BC, Howard AJ (1986) Biochemistry 25: 5314
Poulos TL, Finzel BC, Gunsalus IC, Wagner GC, Kraut J (1985) J. Biol. Chem. 260: 16122
Grinberg AV, Hannemann F, Schiffler B, Muller J, Heinemann U, Bernhardt R (2000) Proteins 40: 590
Bernhardt R (2006) J. Biotechnol. 124: 128
Ortiz de Montanello PR (ed) (2005) Cytochrome P450: Structure, Mechanism and Biochemistry, 3rd ed. Kluwer/Plenum Press, New York
Ryerson CC, Ballou DP, Walsh C (1982) Biochemistry 21: 2644
Visser CM (1983) Eur. J. Biochem. 135: 543
Ghisla S, Massey V (1989) Eur. J. Biochem. 181: 1
Entsch B, van Berkel WJ (1995) FASEB J 9: 476
Jadan AP, Moonen MJH, Boeren SA, Golovleva LA, Rietjens IMCM, van Berkel WJH (2004) Adv. Synth. Catal. 346: 376
Schmid A, Vereyken I, Held M, Witholt B (2001) J. Mol. Catal. B: Enzym. 11: 455
Meyer A, Schmid A, Held M, Westphal A H, Rothlisberger M, Kohler HPE, van Berkel WJH, Witholt B (2002) J. Biol. Chem. 277: 5575
Meyer A, Held M, Schmid A, Kohler HPE, Witholt B (2003) Biotechnol. Bioeng. 81: 518
Otto K, Hofstetter K, Rothlisberger M, Witholt B, Schmid A (2004) J. Bacteriol. 186: 5292
Hollmann F, Hofstetter K, Habicher T, Hauer B, Schmid A (2005) J. Am. Chem. Soc. 127: 6540
Mansuy D, Battioni P (1989) In: Hill CL (ed) Activation and Functionalisation of Alkanes. Wiley, New York, p 195
Johnson RA (1978) Oxygenations with micro-organisms. In: Trahanovsky WS (ed) Oxidation in Organic Synthesis, part C. Academic Press, New York, p 131
Fonken G, Johnson RA (1972) Chemical Oxidations with Microorganisms. Marcel Dekker, New York
Kieslich K (1984) In: Rehm HJ, Reed G (eds) Biotechnology, vol 6a. Verlag Chemie, Weinheim, p 1
Kieslich K (1980) Bull. Soc. Chim. Fr. 11: 9
Sariaslani FS (1989) Crit. Rev. Biotechnol. 9: 171
Bruce NC, French CE, Hailes AM, Long MT, Rathbone DA (1995) Trends Biotechnol. 13: 200
Breslow R (1980) Acc. Chem. Res. 13: 170
Fossey J, Lefort D, Massoudi M, Nedelec JY, Sorba J (1985) Can. J. Chem. 63: 678
Barton DHR, Kalley F, Ozbalik N, Young E, Balavoine G (1989) J. Am. Chem. Soc. 111: 7144
Holland HL (1999) Curr. Opinion Chem. Biol. 3: 22
Mansuy D (1990) Pure Appl. Chem. 62: 741
Kieslich K (1969) Synthesis 120
Kolot FB (1983) Process Biochem. 19
Marsheck WJ (1971) Progr. Ind. Microbiol. 10: 49
Holland HL (1984) Acc. Chem. Res. 17: 389
Holland HL (1982) Chem. Soc. Rev. 11: 371
Sedlaczek L (1988) Crit. Rev. Biotechnol. 7: 186
Weiler EW, Droste M, Eberle J, Halfmann HJ, Weber A (1987) Appl. Microbiol. Biotechnol. 27: 252
Perlman D, Titius E, Fried J (1952) J. Am. Chem. Soc. 74: 2126
Davies HG, Green RH, Kelly DR, Roberts SM (1989) Biotransformations in Preparative Organic Chemistry. Academic Press, London, p 175
Peterson DH, Murray HC, Eppstein SH, Reineke LM, Weintraub A, Meister PD, Leigh HM (1952) J. Am. Chem. Soc. 74: 5933
Fried J, Thoma RW, Gerke JR, Herz JE, Donin MN, Perlman D (1952) J. Am. Chem. Soc. 74: 3692
Sawada S, Kulprecha S, Nilubol N, Yoshida T, Kinoshita S, Taguchi H (1982) Appl. Environ. Microbiol. 44: 1249
Gbewonyo K, Buckland BC, Lilly MD (1991) Biotechnol. Bioeng. 37: 1101
Cohen N, Eichel WF, Lopersti RJ, Neukom C, Saucy G (1976) J. Org. Chem. 41: 3505
Branca Q, Fischli A (1977) Helv. Chim. Acta 60: 925
Evans DA, Sacks CE, Kleschick WA, Taber TR (1979) J. Am. Chem. Soc. 101: 6789
Ohashi T, Hasegawa J (1992) New preparative methods for optically active β-hydroxycarboxylic acids. In: Collins, AN, Sheldrake GN, Crosby J (eds) Chirality in Industry. Wiley, New York, p 249
Goodhue CT, Schaeffer JR (1971) Biotechnol. Bioeng. 13: 203
Aberhart DJ (1977) Bioorg. Chem. 6: 191
Jung H, Jung K, Kleber HP (1993) Adv. Biochem. Eng. Biotechnol. 50: 21
Ciegler A (1974) Microbial transformations of terpenes. In: CRC Handbook of Microbiology, vol 4. CRC Press, Boca Raton, pp 449–458
Krasnobajew V (1984) Terpenoids. In: Rehm HJ, Reed G (eds) Biotechnology, vol 6a. Verlag Chemie: Weinheim, p 97
Lamare V, Furstoss R (1990) Tetrahedron 46: 4109
Rosazza JPN, Steffens JJ, Sariaslani S, Goswami A, Beale JM, Reeg S, Chapman R (1987) Appl. Environ. Microbiol. 53: 2482
Liu WG, Goswami A, Steffek RP, Chapman RL, Sariaslani FS, Steffens JJ, Rosazza JPN (1988) J. Org. Chem. 53: 5700
Archelas A, Furstoss R, Waegell B, le Petit J, Deveze L (1984) Tetrahedron 40: 355
Johnson RA, Herr ME, Murray HC, Reineke LM (1971) J. Am. Chem. Soc. 93: 4880
Furstoss R, Archelas A, Waegell B, le Petit J, Deveze L (1981) Tetrahedron Lett. 22: 445
Johnson RA, Herr ME, Murray HC, Fonken GS (1970) J. Org. Chem. 35: 622
Archelas A, Fourneron JD, Furstoss R (1988) Tetrahedron Lett. 29: 6611
Fonken GS, Herr ME, Murray HC, Reineke LM (1968) J. Org. Chem. 33: 3182
Archelas A, Fourneron JD, Furstoss R (1988) J. Org. Chem. 53: 1797
Johnson RA, Herr ME, Murray HC, Fonken GS (1968) J. Org. Chem. 33: 3217
Furstoss R, Archelas A, Fourneron JD, Vigne B (1986) A model for the hydroxylation site of the fungus Beauveria sulfurescens. In: Schneider MP (ed) Enzymes as Catalysts in Organic Synthesis, NATO ASI Series C, vol 178. Reidel, Dordrecht, p 361
Holland HL, Morris TA, Nava PJ, Zabic M (1999) Tetrahedron 55: 7441
de Raadt A, Griengl H, Petsch M, Plachota P, Schoo N, Weber H, Braunegg G, Kopper I, Kreiner M, Zeiser A (1996) Tetrahedron Asymmetry 7: 491
de Raadt A, Griengl H, Petsch M, Plachota P, Schoo N, Weber H, Braunegg G, Kopper I, Kreiner M, Zeiser A, Kieslich K(1996) Tetrahedron Asymmetry 7: 467
de Raadt A, Griengl H, Petsch M, Plachota P, Schoo N, Weber H, Braunegg G, Kopper I, Kreiner M, Zeiser A (1996) Tetrahedron Asymmetry 7: 473
Braunegg G, de Raadt A, Feichtenhofer S, Griengl H, Kopper I, Lehmann A, Weber H (1999) Angew. Chem. Int. Ed. 38: 2763
Olah GA, Ernst TD (1989) J. Org. Chem. 54: 1204
Komiyam AM (1989) J. Chem. Soc., Perkin Trans. 1, 2031
Zimmer H, Lankin DC, Horgan SW (1971) Chem. Rev. 71: 229
Powlowski JB, Dagley S, Massey V, Ballou DP (1987) J. Biol. Chem. 262: 69
Wiseman A, King DJ (1982) Topics Enzymol. Ferment. Biotechnol. 6: 151
Boyd DR, Campbell RM, Craig HC, Watson CG, Daly JW, Jerina DM (1976) J. Chem. Soc., Perkin Trans. 1, 2438
Schreuder HA, Hol WGJ, Drenth J (1988) J. Biol. Chem. 263: 3131
Klibanov AM, Berman Z, Alberti BN (1981) J. Am. Chem. Soc. 103: 6263
Doddema HJ (1988) Biotechnol. Bioeng. 32: 716
Kazandjian RZ, Klibanov AM (1985) J. Am. Chem. Soc. 107: 5448
Vigne B, Archelas A, Furstoss R (1991) Tetrahedron 47: 1447
Yoshioka H, Nagasawa T, Hasegawa R, Yamada H (1990) Biotechnol. Lett. 679
Theriault RJ, Longfield TH (1973) Appl. Microbiol. 25: 606
Glöckler R, Roduit JP (1996) Chimia 50: 413
Watson GK, Houghton C, Cain RB (1974) Biochem. J. 140: 265
Hoeks FWJMM, Meyer HP, Quarroz D, Helwig M, Lehky P (1990) Scale-up of the process for the biotransformation of nicotinic acid into 6-hydroxynicotinic acid. In: Copping LG, Martin RE, Pickett JA, Bucke C, Bunch AW (eds) Opportunities in Biotransformations. Elsevier, London, p 67
Pasutto FM, Singh NN, Jamali F, Coutts RT, Abuzar S (1987) J. Pharm. Sci. 76: 177
Kolb HC, VanNieuwenhze MS, Sharpless KB (1994) Chem. Rev. 94: 2483
Johnson RA, Sharpless KB (1993) In: Ojima I (ed) Catalytic Asymmetric Synthesis. Verlag Chemie, New York, pp 103–158
Pfenninger A (1986) Synthesis 89
Konishi K, Oda K, Nishida K, Aida T, Inoue S (1992) J. Am. Chem. Soc. 114: 1313
Leak DJ, Aikens PJ, Seyed-Mahmoudian M (1992) Trends Biotechnol. 10: 256
Weijers CAGM, de Haan A, de Bont JAM (1988) Microbiol. Sci. 5: 156
May SW (1979) Enzyme Microb. Technol. 1: 15
Furuhashi K (1986) Econ. Eng. Rev. 18 (7/8): 21
Abraham WR, Stumpf B, Arfmann HA (1990) J. Essent. Oil Res. 2: 251
Habets-Ctützen AQH, Carlier SJN, de Bont JAM, Wistuba D, Schurig V, Hartmans S, Tramper J (1985) Enzyme Microb. Technol. 7: 17
Furuhashi K (1986) Chem. Econ. Eng. Rev. 18 (7–8): 21
de Smet MJ, Witholt B, Wynberg H (1981) J. Org. Chem. 46: 3128
Takahashi O, Umezawa J, Furuhashi K, Takagi M (1989) Tetrahedron Lett. 30: 1583
Furuhashi K (1992) Biological methods to optically active epoxides. In: Collins AN, Sheldrake GN, Crosby J (eds) Chirality in Industry. Wiley, New York, p 167
White RF, Birnbaum J, Meyer RT, ten Broeke J, Chemerda JM, Demain AL (1971) Appl. Microbiol. 22: 55
Peterson JA, Basu D, Coon MJ (1966) J. Biol. Chem. 241: 5162
de Smet MJ, Witholt B, Wynberg H (1983) Enzyme Microb. Technol. 5: 352
Jurtshuk P, Cardini GE (1972) Crit. Rev. Microbiol. 1: 254
Fu H, Newcomb M, Wong CH (1991) J. Am. Chem. Soc. 113: 5878
Katopodis AG, Wimalasena K, Lee J, May SW (1984) J. Am. Chem. Soc. 106: 7928
May SW, Abbott BJ (1973) J. Biol. Chem. 248: 1725
May SW (1976) Catal. Org. Synth. 4: 101
May SW, Schwartz RD, Abbott BJ, Zaborsky OR (1975) Biochim. Biophys. Acta 403: 245
Habets-Crützen AQH, de Bont JAM (1985) Appl. Microbiol. Biotechnol. 22: 428
Brink LES, Tramper J (1987) Enzyme Microb. Technol. 9: 612
Brink LES, Tramper J (1985) Biotechnol. Bioeng. 27: 1258
Hou CT, Patel R, Laskin AI, Barnabe N, Barist I (1983) Appl. Environ. Microbiol. 46: 171
Furuhashi K (1981) Ferment. Ind. 39: 1029
Ohta H, Tetsukawa H (1979) Agric. Biol. Chem. 43: 2099
Johnstone SL, Phillips GT, Robertson BW, Watts PD, Bertola MA, Koger HS, Marx AF (1987) Stereoselective synthesis of (S)-β-blockers via microbially produced epoxide intermediates. In: Laane C, Tramper J, Lilly MD (eds) Biocatalysis in Organic Media. Elsevier, Amsterdam, p 387
Fu H, Shen GJ, Wong CH (1991) Recl. Trav. Chim. Pays-Bas 110: 167
Howe R, Shanks RG (1966) Nature 210: 1336
Weijers CAGM, van Ginkel CG, de Bont JAM (1988) Enzyme Microb. Technol. 10: 214
Schmid A, Hofstetter K, Freiten H-J, Hollmann F, Witholt B (2001) Adv. Synth. Catal. 343: 752
Panke S, Held W, Wubbolts MG, Witholt B, Schmid A (2002) Biotechnol. Bioeng. 80: 33
Bravo P, Resnati G, Viani F (1985) Tetrahedron Lett. 26: 2913
Solladie G (1981) Synthesis 185
Goldberg SI, Sahli MS (1967) J. Org. Chem. 32: 2059
Solladie G, Demailly G, Greck C (1985) J. Org. Chem. 50: 1552
Kagan HB, Dunach E, Nemeck C, Pitchen P, Samuel O, Zhao S (1985) Pure Appl. Chem. 57: 1922
Holland HL (1988) Chem. Rev. 88: 473
Phillips RS, May SW (1981) Enzyme Microb. Technol. 3: 9
Allen CCR, Boyd DR, Dalton H, Sharma ND, Haughey SA, McMordie RAS, McMurray BT, Sheldrake GN, Sproule K (1995) J. Chem. Soc., Chem. Commun. 119
Dodson RM, Newman N, Tsuchiya HM (1962) J. Org. Chem. 27: 2707
Auret BJ, Boyd DR, Henbest HB, Watson CG, Balenovic K, Polak V, Johanides V, Divjak S (1974) Phytochemistry 13: 65
Ohta H, Okamoto Y, Tsuchihashi G (1985) Agric. Biol. Chem. 49: 2229
Ohta H, Matsumoto S, Okamoto Y, Sugai T (1989) Chem. Lett., 625
Holland HL, Brown FM, Lakshmaiah G, Larsen BG, Patel M (1997) Tetrahedron Asymmetry 8: 683
Holland HL, Carter IM (1983) Bioorg. Chem. 12: 1
Buist PH, Marecak DM, Partington ET, Skala P (1990) J. Org. Chem. 55: 5667
Beecher J, Brackenridge I, Roberts SM, Tang J, Willetts AJ (1995) J. Chem. Soc., Perkin Trans. 1, 1641
Ohta H, Okamoto Y, Tsuchihashi G (1985) Agric. Biol. Chem. 49: 671
Abushanab E, Reed D, Suzuki F, Sih CJ (1978) Tetrahedron Lett. 19: 3415
Holland HL, Pöpperl H, Ninniss RW, Chenchaiah PC (1985) Can. J. Chem. 63: 1118
Poje M, Nota O, Balenovic K (1980) Tetrahedron 36: 1895
Auret BJ, Boyd DR, Breen F, Greene RME, Robinson PM (1981) J. Chem. Soc., Perkin Trans. 1, 930
Auret BJ, Boyd DR, Cassidy ES, Turley F, Drake AF, Mason SF (1983) J. Chem. Soc., Chem. Commun. 282
Okamoto Y, Ohta H, Tsuchihashi G (1986) Chem. Lett. 2049
Colonna S, Gaggero N, Pasta P, Ottolina G (1996) Chem. Commun. 2303
Krow GR (1981) Tetrahedron 37: 2697
Mimoun H (1982) Angew. Chem. Int. Ed. 21: 734
Criegee R (1948) Liebigs Ann. Chem. 560: 127
Lee JB, Uff BC (1967) Quart. Rev. 21: 429
Schwab JM, Li WB, Thomas LP (1983) J. Am. Chem. Soc. 105: 4800
Gunsalus IC, Peterson TC, Sligar SG (1975) Ann. Rev. Biochem. 44: 377
Walsh CT, Chen YCJ (1988) Angew. Chem. Int. Ed. 27: 333
Roberts SM, Wan PWH (1998) J. Mol. Catal. B 4: 111
Bolm C, Schlingloff G, Weickhardt K (1994) Angew. Chem. Int. Ed. 33: 1848
Abril O, Ryerson CC, Walsh C, Whitesides GM (1989) Bioorg. Chem. 17: 41
Nealson KH, Hastings JW (1979) Microbiol. Rev. 43: 496
Donoghue NA, Norris DB, Trudgill PW (1976) Eur. J. Biochem. 63: 175
Britton LN, Markavetz AJ (1977) J. Biol. Chem. 252: 8561
Trower MK, Buckland RM, Griffin M (1989) Eur. J. Biochem. 181: 199
Torres Pazmino DE, Snajdrova R, Baas BJ, Ghobrial M, Mihovilovic MD, Fraaije MW (2008) Angew. Chem. Int. Ed. 47: 2275
Alphand V, Archelas A, Furstoss R (1990) J. Org. Chem. 55: 347
Taschner MJ, Black DJ (1988) J. Am. Chem. Soc. 110: 6892
Taschner MJ, Black DJ, Chen QZ (1993) Tetrahedron Asymmetry 4: 1387
Kelly DR, Knowles CJ, Mahdi JG, Taylor IN, Wright MA (1995) J. Chem. Soc., Chem. Commun. 729
Ottolina G, Carrea G, Colonna S, Rückemann A (1996) Tetrahedron Asymmetry 7: 1123
Ouazzani-Chahdi J, Buisson D, Azerad R (1987) Tetrahedron Lett. 28: 1109
Alphand V, Archelas A, Furstoss R (1990) Biocatalysis 3: 73
Levitt MS, Newton RF, Roberts SM, Willetts AJ (1990) J. Chem. Soc., Chem. Commun. 619
Alphand V, Archelas A, Furstoss R (1989) Tetrahedron Lett. 30: 3663
Carnell AJ, Roberts SM, Sik V, Willetts AJ (1990) J. Chem. Soc., Chem. Commun. 1438
Doig SD, Simpson H, Alphand V, Furstoss R, Woodley JM (2003) Enzyme Microb. Technol. 32: 347
Alphand V, Carrea G, Wohlgemuth R, Furstoss R, Woodley J M (2003) Trends Biotechnol. 21: 318
Stark D, von Stockar U (2003) Adv. Biochem. Eng. Biotechnol. 80: 149
Vicenci JT, Zmijewski MJ, Reinhard MR, Landen BE, Muth WL, Marler PG (1997) Enzyme Microb. Technol. 29: 494
Alphand V, Furstoss R (1992) J. Org. Chem. 57: 1306
Petit F, Furstoss R (1993) Tetrahedron Asymmetry 4: 1341
Grogan G, Roberts SM, Wan P, Willetts AJ (1993) Biotechnol. Lett. 15: 913
Willetts A (1997) Trends Biotechnol. 15: 55
Kelly DR, Knowles CJ, Mahdi JG, Wright MA, Taylor IN, Roberts SM, Wan PWH, Grogan G, Pedragosa-Moreau S, Willetts AJ (1996) Chem. Commun. 2333
Grogan G, Roberts SM, Willetts AJ (1993) J. Chem. Soc., Chem. Commun. 699
Mihovilovic MD, Müller B, Stanetty P (2002) Eur. J. Org. Chem. 3711
Kayser M, Chen G, Stewart J (1999) Synlett. 153
Stewart JD, Reed KW, Kayser MM (1996) J. Chem. Soc., Perkin Trans. 1, 755
Kayser MM, Chen G, Stewart JD (1998) J. Org. Chem. 63: 7103
Stewart JD, Reed KW, Martinez CA, Zhu J, Chen G, Kayser MM (1998) J. Am. Chem. Soc. 120: 3541
Stewart JD (1998) Curr. Org. Chem. 2: 211
Kyte BC, Rouviere P, Cheng Q, Stewart JD (2004) J. Org. Chem. 69: 12
Willetts AJ, Knowles CJ, Levitt MS, Roberts SM, Sandey H, Shipston NF (1991) J. Chem. Soc., Perkin Trans. 1, 1608
Gibson DT, Parales RE (2000) Curr. Opin. Biotechnol. 11: 236
Butler CS, Mason JR (1997) Adv. Microb. Physiol. 38: 47
Hamberg M (1996) Acta Chem. Scand. 50: 219
Yagi K (ed) (1982) Lipid Peroxides in Biology and Medicine. Academic Press, New York
Subramanian V, Sugumaran M, Vaidyanathan CS (1978) J. Ind. Inst. Sci. 60: 143
Jeffrey H, Yeh HJC, Jerina DM, Patel TR, Davey JF, Gibson DT (1975) Biochemistry 14: 575
Axelrod B (1974) ACS Adv. Chem. Ser. 136: 324
Vick BA, Zimmerman DC (1984) Plant. Physiol. 75: 458
Corey EJ, Nagata R (1987) J. Am. Chem. Soc. 109: 8107
Theorell H, Holman RT, Akeson A (1947) Acta Chem. Scand. 1: 571
Finnazzi-Agro A, Avigliano L, Veldink GA, Vliegenhart JFG, Boldingh J (1973) Biochim. Biophys. Acta 326: 462
Axelrod B, Cheesbrough TM, Laakso TM (1981) Methods Enzymol. 71: 441
Funk Jr MO, Andre JC, Otsuki T (1987) Biochemistry 26: 6880
Van Os CPA, Vente M, Vliegenhart JFG (1979) Biochim. Biophys. Acta 547: 103
Corey EJ, Albright JO, Burton AE, Hashimoto S (1980) J. Am. Chem. Soc. 102: 1435
Corey EJ, Nagata R (1987) Tetrahedron Lett. 28: 5391
Iacazio G, Langrand G, Baratti J, Buono G, Triantaphylides C (1990) J. Org. Chem. 55: 1690
Gunstone FD (1979) In: Barton DHR, Ollis WD, Haslam E (eds) Comprehensive Organic Chemistry. Pergamon Press, New York, p 587
Datcheva VK, Kiss K, Solomon L, Kyler KS (1991) J. Am. Chem. Soc. 113: 270
Novak MJ (1999) Bioorg. Med. Chem. Lett. 9: 31
Martini D, Buono G, Iacazio G (1996) J. Org. Chem. 61: 9062
Zhang P, Kyler KS (1989) J. Am. Chem. Soc. 111: 9241
Akakabe Y, Matsui K, Kajiwara T (1999) Tetrahedron Lett. 40: 1137
Shine WE, Stumpf PK (1974) Arch. Biochem. Biophys. 162: 147
Adam W, Boland W, Hartmann-Schreier J, Humpf HU, Lazarus M, Saffert A, Saha-Möller CR, Schreier P (1998) J. Am. Chem. Soc. 120: 11044
Adam W, Lazarus M, Saha-Möller CR, Schreier P (1996) Tetrahedron Asymmetry 7: 2287
Smith MR, Ratledge C (1989) Appl. Microbiol. Biotechnol. 32: 68
Zylstra GJ, Gibson DT (1989) J. Biol. Chem. 264: 14940
Gibson DT, Koch JR, Kallio RE (1968) Biochemistry 7: 3795
Brazier AJ, Lilly MD, Herbert AB (1990) Enzyme Microb. Technol. 12: 90
Parales RE, Resnick SM (2007) Application of aromatic hydrocarbon dioxygenases. In: Patel R N (ed) Biocatalysis in the Pharmaceutical and Biotechnological Industries. CRC Press, Boca Raton, p 299
Crosby J (1991) Tetrahedron 47: 4789
Ribbons DW, Evans CT, Rossiter JT, Taylor SCJ, Thomas SD, Widdowson DA, Williams DJ (1990) Biotechnology and biodegradation. In: Kamely D, Chakrabarty A, Omenn GS (eds) Advances in Applied Biotechnology Series, vol 4. Gulf Publ. Co., Houston, p 213
Widdowson DA, Ribbons DW (1990) Janssen Chim. Acta 8 (3): 3
Ballard DHG, Courtis A, Shirley IM, Taylor SC (1988) Macromolecules 21: 294
Sheldrake GN (1992) Biologically derived arene cis-dihydrodiols as synthetic building blocks. In: Collins AN, Sheldrake GN, Crosby J (eds) Chirality in Industry. Wiley, New York, p 127
Taylor SC, Ribbons DW, Slawin AMZ, Widdowson DA, Williams DJ (1987) Tetrahedron Lett. 28: 6391
Rossiter JT, Williams SR, Cass AEG, Ribbons DW (1987) Tetrahedron Lett. 28: 5173
Boyd DR, Sharma ND, Hand MV, Groocock MR, Kerley NA, Dalton H, Chima J, Sheldrake GN (1993) J. Chem. Soc., Chem. Commun. 974
Ribbons DW, Cass AEG, Rossiter JT, Taylor SJC, Woodland MP, Widdowson DA, Williams SR, Baker PB, Martin RE (1987) J. Fluorine Chem. 37: 299
Geary PJ, Pryce RJ, Roberts SM, Ryback G, Winders JA (1990) J. Chem. Soc., Chem. Commun. 204
Deluca ME, Hudlicky T (1990) Tetrahedron Lett. 31: 13
Wackett LP, Kwart LD, Gibson DT (1988) Biochemistry 27: 1360
Boyd DR, Sharma ND, Boyle R, Malone JF, Chima J, Dalton H (1993) Tetrahedron Asymmetry 4: 1307
Allen CCR, Boyd DR, Dalton H, Sharma ND, Brannigan I, Kerley NA, Sheldrake GN, Taylor SS (1995) J. Chem. Soc., Chem. Commun. 117
Boyd DR, Sharma ND, Boyle R, McMurry BT, Evans TA, Malone JF, Dalton H, Chima J, Sheldrake GN (1993) J. Chem. Soc., Chem. Commun. 49
Lakshman MK, Chaturvedi S, Zaijc B, Gibson DT, Resnick SM (1998) Synthesis 1352
Hudlicky T, Boros EE, Boros CH (1993) Tetrahedron Asymmetry 4: 1365
Boyd DR, Sharma ND, Bowers NI, Brannigan IN, Groocock MR, Malone JF, McConville G, Allen CCR (2005) Adv. Synth. Catal. 347: 1081
Ley SV, Sternfeld F (1989) Tetrahedron 45: 3463
Ley SV, Parra M, Redgrave AJ, Sternfeld F (1990) Tetrahedron 46: 4995
Hudlicky T, Luna H, Barbieri G, Kwart LD (1988) J. Am. Chem. Soc. 110: 4735
Hudlicky T, Luna H, Price JD, Rulin F (1989) Tetrahedron Lett. 30: 4053
Hudlicky T, Price JD (1990) Synlett. 159
Hudlicky T, Luna H, Price JD, Rulin F (1990) J. Org. Chem. 55: 4683
Mermod N, Harayamas S, Timmis KN (1986) Bio/Technology 4: 321
Strukul G (ed) (1992) Catalytic Oxidations with Hydrogen Peroxide as Oxidant. Kluwer, Dordrecht
Butler A, Walker JV (1993) Chem. Rev. 93: 1937
Kutney JP (1991) Synlett. 11
Nakayama T, Amachi T (1999) J. Mol. Catal. B 6: 185
Flohe L (1979) CIBA Found. Symp. 65: 95
de Boer E, Y van Kooyk, MGM Tromp, Plat H, Wever R (1986) Biochim. Biophys. Acta 869: 48
Butler A (1998) Curr. Opinion Chem. Biol. 2: 279
Kuwahara M, Glenn JK, Morgan MA, Gold MH (1984) FEBS Lett. 169: 247
Dolin MI (1957) J. Biol. Chem. 225: 557
Dunford HB (1991) Horseradish peroxidase: structure and kinetic properties. In: Everse J, Everse KE, Grisham MB (eds) Peroxidases in Chemistry and Biology. CRC Press, Boca Raton, p 1
Anni H, Yonetani T (1992) Mechanism of action of peroxidases. In: Sigel H, Sigel A (eds) Metal Ions in Biological Systems: Degradation of Environmental Pollutants by Microorganisms and their Related Metalloenzymes. Marcel Dekker, New York, p 219
Ortiz de Montellano PR (1992) Annu. Rev. Pharm. Toxicol. 32: 89
Berglund GI, Carlsson GH, Smith A T, Szöke H, Hensiksen A, Hajdu J (2002) Nature 417: 463
Colonna S, Gaggero N, Richelmi C, Pasta P (1999) Trends Biotechnol. 17: 163
Adam W, Lazarus M, Saha-Möller CR, Weichold O, Hoch U, Häring D, Schreier P (1999) Adv. Biochem. Eng. Biotechnol. 63: 73
van Deurzen MPJ, van Rantwijk F, Sheldon RA (1997) Tetrahedron 53: 13183
Dordick JS (1992) Trends Biotechnol. 10: 287
Uyama H, Kurioka H, Sugihara J, Kobayashi S (1996) Bull. Chem. Soc. Jpn. 69: 189
Kobayashi S, Shoda S, Uyama H (1995) Adv. Polym. Sci. 121: 1
Fukunishi K, Kitada K, Naito I (1991) Synthesis 237
Schmitt MM, Schüler E, Braun M, Häring D, Schreier P (1998) Tetrahedron Lett. 39: 2945
Littlechild J (1999) Curr. Opinion Chem. Biol. 3: 28
Franssen MCR (1994) Biocatalysis 10: 87
Shaw PD, Hager LP (1961) J. Biol. Chem. 236: 1626
Blanke SR, Hager LP (1989) Biotechnol. Lett. 11: 769
Sundaramoothy M, Terner J, Poulos TL (1995) Structure 3: 1367
McCarthy MB, White RE (1983) J. Biol. Chem. 258: 9153
Miller VP, Tschirret-Guth RA, Ortiz de Montellano PG(1995) Arch. Biochem. Biophys. 319: 333
Zaks A, Dodds DR (1995) J. Am. Chem. Soc. 117: 10419
Hu S, Hager LP (1999) J. Am. Chem. Soc. 121: 872
Seelbach K, van Deurzen MPJ, van Rantwijk F, Sheldon RA, Kragl U (1997) Biotechnol. Bioeng. 55: 283
van Deurzen MPJ, van Rantwijk F, Sheldon RA (1996) J. Mol. Catal. B 2: 33
Hofmann B, Tolzer S, Pelletier I, Altenbuchner J, van Pee K-H (1998) J. Mol. Biol. 279: 889
Kirk O, Conrad LS (1999) Angew. Chem. Int. Ed. 38: 977
Hager LP, Lakner FJ, Basavapathruni A (1998) J. Mol. Catal. B 5: 95
Hu S, Hager LP (1999) Tetrahedron Lett. 40: 1641
Lakner FJ, Cain KP, Hager LP (1997) J. Am. Chem. Soc. 119: 443
Allain EJ, Hager LP, Deng L, Jacobsen EN (1993) J. Am. Chem. Soc. 115: 4415
Dexter AF, Lakner FJ, Campbell RA, Hager LP (1995) J. Am. Chem. Soc. 117: 6412
Lakner FJ, Hager LP (1996) J. Org. Chem. 61: 3923
Colonna S, Gaggero N, Casella L, Carrea G, Pasta P (1993) Tetrahedron Asymmetry 4: 1325
Kobayashi S, Nakano M, Kimura T, Schaap AP (1987) Biochemistry 26: 5019
Colonna S, Gaggero N, Casella L, Carrea G, Pasta P (1992) Tetrahedron Asymmetry 3: 95
Colonna S, Gaggero N, Manfredi A, Casella L, Gullotti M, Carrea G, Pasta P (1990) Biochemistry 29: 10465
Andersson M, Willetts A, Allenmark S (1997) J. Org. Chem. 62: 8455
Allenmark SG, Andersson MA (1996) Tetrahedron Asymmetry 7: 1089
Andersson MA, Allenmark SG (1998) Tetrahedron 54: 15293
Höft E, Hamann HJ, Kunath A, Adam W, Hoch U, Saha-Möller CR, Schreier P (1995) Tetrahedron Asymmetry 6: 603
Adam W, Fell RT, Hoch U, Saha-Möller CR, Schreier P (1995) Tetrahedron Asymmetry 6: 1047
Adam W, Lazarus M, Hoch U, Korb MN, Saha-Möller CR, Schreier P (1998) J. Org. Chem. 63: 6123
Adam W, Hoch U, Lazarus M, Saha-Möller CR, Schreier P (1995) J. Am. Chem. Soc. 117: 11898
Adam W, Boss B, Harmsen D, Lukacs Z, Saha-Möller CR, Schreier P (1998) J. Org. Chem. 63: 7598
Adam W, Mock-Knoblauch C, Saha-Möller CR (1999) J. Org. Chem. 64: 4834
van Deurzen MPJ, Seelbach K, van Rantwijk F, Kragl U, Sheldon RA (1997) Biocatalysis 15: 1
Pasta P, Carrea G, Monzani E, Gaggero N, Colonna S (1999) Biotechnol. Bioeng. 62: 489
van der Velde F, van Rantwijk F, Sheldon RA (1999) J. Mol. Catal. B 6: 453
References to Sect. 2.4
Evans DA, Nelson JV, Taber TR (1982) Topics Stereochem. 13: 1
Heathcock CH (1984) Asymm. Synthesis 3: 111
Mukaiyama T (1982) Org. React. 28: 203
Paterson I, Goodman JM, Lister MA, Schumann RC, McClure CK, Norcross RD (1990) Tetrahedron 46: 4663
Masamune S, Choy W, Peterson J, Sita LR (1986) Angew. Chem. Int. Ed. 24: 1
Kazmeier U (2005) Angew. Chem. Int. Ed. 44: 2186
Fessner W-D (1992) Kontakte (Merck) (3) 3
Fessner W-D (1993) Kontakte (Merck) (1) 23
Toone EJ, Simon ES, Bednarski MD, Whitesides, GM (1989) Tetrahedron 45: 5365
Gijsen HJM, Qiao L, Fitz W, Wong CH (1996) Chem. Rev. 96: 443
Wong CH (1993) Chimia 47: 127
Drueckhammer DG, Hennen WJ, Pederson RL, Barbas CF, Gautheron CM, Krach T, Wong CH (1991) Synthesis 499
Look GC, Fotsch CH, Wong CH (1993) Acc. Chem. Res. 26: 182
Wong CH (1990) Aldolases in organic synthesis. In: Abramowicz DA (ed) Biocatalysis. Van Nostrand Reinhold, New York, p 319
Dean SM, Greenberg WA, Wong CH (2007) Adv. Synth. Catal. 349: 1308
Fessner WD, Helaine V (2001) Curr. Opinion Biotechnol. 12: 574
Seoane G (2000) Curr. Org. Chem. 4: 283
Machajewski TD, Wong CH (2000) Angew. Chem. Int. Ed. 39: 1352
Fessner WD, Jennewein S (2007) Biotechnological applications of aldolases. In: Patel R N (ed) Biocatalysis in the Pharmaceutical and Biotechnology Industries. CRC Press, Boca Raton, p 363
Clapes P, Fessner WD, Sprenger GA, Samland AK (2010) Curr. Opin. Chem. Biol. 14: 154
Meyerhof O, Lohmann K (1934) Biochem. Z. 271: 89
Horecker L, Tsolas O, Lai CY (1972) In: Boyer PD (ed) The Enzymes, vol 7. Academic Press, New York, p 213
Brockamp HP, Kula MR (1990) Appl. Microbiol. Biotechnol. 34: 287
von der Osten CH, Sinskey AJ, Barbas III CF, Pederson RL, Wang YF, Wong C-H (1989) J. Am. Chem. Soc. 111: 3924
Fessner WD, Schneider A, Held H, Sinerius G, Walter C, Hixon M, Schloss JV (1996) Angew. Chem., Int. Ed. Engl. 35: 2219
Dreyer MK, Schulz GE (1993) J. Mol. Biol. 231: 549
Shelton MC, Cotterill IC, Novak STA, Poonawala RM, Sudarshan S, Toone EJ (1996) J. Am. Chem. Soc. 118: 2117
Schoevaart R, van Rantwijk F, Sheldon RA (1999) Tetrahedron Asymmetry 10: 705
Jones JKN, Sephton HH (1960) Can. J. Chem. 38: 753
Jones JKN, Kelly RB (1956) Can. J. Chem. 34: 95
Horecker BL, Smyrniotis PZ (1952) J. Am. Chem. Soc. 74: 2123
Huang PC, Miller ON (1958) J. Biol. Chem. 330: 805
Kajimoto T, Chen L, Liu KKC, Wong C-H (1991) J. Am. Chem. Soc. 113: 6678
Effenberger F, Straub A, Null V (1992) Liebigs Ann Chem. 1297
Wong C-H, Mazenod FP, Whitesides GM (1983) J. Org. Chem. 3493
Bednarski MD, Waldmann HJ, Whitesides GM (1986) Tetrahedron Lett. 27: 5807
Jones JKN, Matheson NK (1959) Can. J. Chem. 37: 1754
Gorin PAJ, Hough L, Jones JKN (1953) J. Chem. Soc. 2140
Lehninger AL, Sice J (1955) J. Am. Chem. Soc. 77: 5343
Charalampous FC (1954) J. Biol. Chem. 211: 249
Duncan R, Drueckhammer DG (1996) J. Org. Chem. 61: 438
Fessner WD, Sinerius G (1994) Angew. Chem. Int. Ed. 33: 209
Simon ES, Plante R, Whitesides GM (1989) Appl. Biochem. Biotechnol. 22: 169
Bischofberger N, Waldmann H, Saito T, Simon ES, Lees W, Bednarski MD, Whitesides GM (1988) J. Org. Chem. 53: 3457
Arth HL, Fessner WD (1997) Carbohydr. Res. 305: 131
Liu KKC, Kajimoto T, Chen L, Zhong Z, Ichikawa Y, Wong CH (1991) J. Org. Chem. 56: 6280
Ziegler T, Straub A, Effenberger F (1988) Angew. Chem. Int. Ed. 27: 716
Schultz M, Waldmann H, Kunz H, Vogt W (1990) Liebigs Ann. Chem. 1010
Lees WJ, Whitesides GM (1993) J. Org. Chem. 58: 1887
Durrwachter JR, Drueckhammer DG, Nozaki K, Sweers HM, Wong CH (1986) J. Am. Chem. Soc. 108: 7812
Fessner WD (1992) A building block strategy for asymmetric synthesis: the DHAP-aldolases. In: Servi S (ed) Microbial Reagents in Organic Synthesis. Kluwer, Dordrecht, p 43
Ozaki A, Toone EJ, von der Osten CH, Sinskey AJ, Whitesides GM (1990) J. Am. Chem. Soc. 112: 4970
Pederson RL, Esker J, Wong CH (1991) Tetrahedron 47: 2643
Effenberger F, Straub A (1987) Tetrahedron Lett. 28: 1641
Gefflaut T, Lemaire M, Valentin ML, Bolte J (1997) J. Org. Chem. 62: 5920
Fessner WD, Walter C (1992) Angew. Chem. Int. Ed. 31: 614
Lagunas R, Sols A (1968) FEBS Lett. 1: 32
Drueckhammer DG, Durrwachter JR, Pederson RL, Crans DC, Daniels L, Wong CH (1989) J. Org. Chem. 54: 70
Suhiyama M, Hong Z, Whalen LJ, Greenberg WA, Wong CH (2006) Adv. Synth. Catal. 348: 2555
Schürmann M, Sprenger GA (2001) J. Biol. Chem. 276: 11055
Schürmann M, Schürmann M, Sprenger GA (2002) J. Mol. Catal. B: Enzym. 19–20: 247
Sugiyama M, Hong Z, Liang PH, Dean SM, Whalen LJ, Greenberg WA, Wong CH (2007) J. Am. Chem. Soc. 129: 14811
Castillo JA, Guerard-Helaine C, Gutierrez M, Garrabou X, Sancelme M, Schürmann M, Inoue T, Helaine V, Charmantray F, Gefflaut T, Hecquet L, Joglar J, Clapes P, Sprenger GA, Lemaire M (2010) Adv. Synth. Catal. 352: 1039
Castillo JA, Calveras J, Casas J, Mitjans M, Vinardell MP, Parella T, Inoue T, Sprenger GA, Joglar J, Clapes P (2006) Org. Lett. 8: 6067
Simon ES, Bednarski MD, Whitesides GM (1988) J. Am. Chem. Soc. 110: 7159
Uchida Y, Tsukada Y, Sugimori T (1985) Agric. Biol. Chem. 49: 181
Kragl U, Kittelmann M, Ghisalba O, Wandrey C (1995) Ann. NY Acad. Sci. 750: 300
Kragl U, Gygax D, Ghisalba O, Wandrey C (1991) Angew. Chem. Int. Ed. 30: 827
Maru I, Ohnishi J. Ohta H, Tsukuda Y (1998) Carbohydr. Res. 306: 575
Mahmoudian M, Noble D, Drake CS, Middleton RF, Montgomery DS, Piercey JE, Ramlakhan D, Todd M, Dawson MJ (1997) Enzyme Microb. Technol. 20: 393
Schauer R (1985) Trends Biochem. Sci. 10: 357
Ota Y, Shimosaka M, Murata K, Tsudaka Y, Kimura A (1986) Appl. Microbiol. Biotechnol. 24: 386
Brunetti P, Jourdian GW, Roseman S (1962) J. Biol. Chem. 237: 2447
Brossmer R, Rose U, Kasper D, Smith TL, Grasmuk H, Unger FM (1980) Biochem. Biophys. Res. Commun. 96: 1282
Augé C, David S, Gautheron C, Malleron A, Cavayre B (1988) New J. Chem. 12: 733
Kim MJ, Hennen WJ, Sweers HM, Wong CH (1988) J. Am. Chem. Soc. 110: 6481
Lin CH, Sugai T, Halcomb RL, Ichikawa Y, Wong CH (1992) J. Am. Chem. Soc. 114: 10138
Augé C, Gautheron C, David S, Malleron A, Cavayé B, Bouxom B (1990) Tetrahedron 46: 201
Bednarski MD, Crans DC, DiCosmio R, Simon ES, Stein PD, Whitesides GM, Schneider M (1988) Tetrahedron Lett. 29: 427
Sugai T, Shen GJ, Ichikawa Y, Wong CH (1993) J. Am. Chem. Soc. 115: 413
Henderson DP, Cotterill IC, Shelton MC, Toone EJ (1998) J. Org. Chem. 63: 906
Reimer LM, Conley DL, Pompliano DL, Frost JW (1986) J. Am. Chem. Soc. 108: 8010
Allen ST, Heintzelman, GR, Toone EJ (1992) J. Org. Chem. 57: 426
Barbas III CF, Wang YF, Wong CH (1990) J. Am. Chem. Soc. 112: 2013
Wong CH, Garcia-Junceda E, Chen L, Blanco O, Gijsen HJM, Stennsma DH (1995) J. Am. Chem. Soc. 117: 3333
Greenberg WA, Varvak A, Hanson SR, Wong K, Huang H, Chen P, Burk MJ (2004) Proc. Nat. Acad. Sci. USA 101: 5788
DeSantis G, Liu J, Clark DP, Heine A, Wilson JA, Wong CH (2003) Bioorg. Med. Chem. 11: 43
Müller M (2005) Angew. Chem. Int. Ed. 44: 362
Gijsen HJM, Wong CH (1995) J. Am. Chem. Soc. 117: 2947
Gijsen HJM, Wong CH (1995) J. Am. Chem. Soc. 117: 7585
Lotz BT, Gasparski CM, Peterson K, Miller MJ (1990) J. Chem. Soc., Chem. Commun. 1107
Steinreiber J, Fesko K, Reisinger C, Schürmann M, van Assema F, Wolberg M, Mink D, Griengl H (2007) Tetrahedron 63: 918
Liu JQ, Dairi T, Itoh N, Kataoka M, Shimizu S, Yamada H (2000) J. Mol. Catal. B 10: 107
Saeed A, Young DW (1992) Tetrahedron 48: 2507
Vassilev VP, Uchiyama T, Kajimoto T, Wong CH (1995) Tetrahedron Lett. 36: 4081
Kimura T, Vassilev VP, Shen GJ, Wong CH (1997) J. Am. Chem. Soc. 119: 11734
Steinreiber J, Fesko K, Mayer C, Reisinger C, Schürmann M, Griengl H (2007) Tetrahedron 63: 8088
Steinreiber J, Schürmann M, van Assema F, Wolberg M, Fesko K, Reisinger C, Mink D, Griengl H (2007) Adv. Synth. Catal. 349: 1379
Lohmann W, Schuster G (1937) Biochem. Z. 294: 188
Müller M, Gocke D, Pohl M (2009) FEBS J. 276: 2894
Pohl M, Lingen B, Müller M (2002) Chem. Eur. J. 8: 5289
Pohl M, Sprenger GA, Müller M (2004) Curr. Opinion Biotechnol. 15: 335
Frank RAW, Leeper FJ (2007) Cell. Mol. Life Sci. 64: 892
Breslow R (1957) J. Am. Chem. Soc. 79: 1762
Kern D, Kern G, Neef H, Tittmann K, Killenberg-Jabs M, Wikner C, Schneider G, Hübner G (1997) Science 275: 67
Lintner CJ, Liebig HJ (1913) Z. physiol. Chem. 88: 109
Neuberg C, Hirsch J (1921) Biochem. Z. 115: 282
Hildebrandt G, Klavehn W (1930) Ger. Patent 548459
Pohl M (1997) Adv. Biochem. Eng. Biotechnol. 58: 16
Fuganti C, Grasselli P (1989) Baker’s yeast-mediated synthesis on natural products. In: Whitaker JR, Sonnet PE (eds) Biocatalysis in Agricultural Biotechnology. ACS Symp. Ser., ACS, Washington DC, p 359
Fuganti C, Grasselli P (1977) Chem. Ind. 983
Crout DHG, Dalton H, Hutchinson DW, Miyagoshi M (1991) J. Chem. Soc., Perkin Trans. 1, 1329
Pohl M (1999) Protein design on pyruvate decarboxylase (PDC) by site-directed mutagenesis. In: Scheper T (ed) New Enzymes for Organic Synthesis. Springer, Berlin Heidelberg New York, p 15
Sprenger GA, Pohl M (1999) J. Mol. Catal. B 6: 145
Fuganti C, Grasselli P, Poli G, Servi S, Zorzella A (1988) J. Chem. Soc., Chem. Commun. 1619
Fuganti C, Grasselli P, Servi S, Spreafico F, Zirotti C (1984) J. Org. Chem. 49: 4087
Suomalainen H, Linnahalme T (1966) Arch. Biochem. Biophys. 114: 502
Neuberg C, Liebermann L (1921) Biochem. Z. 121: 311
Behrens M, Ivanoff N (1926) Biochem. Z. 169: 478
Fronza G, Fuganti C, Grasselli P, Poli G, Servi S (1990) Biocatalysis 3: 51
Fronza G, Fuganti C, Majori L, Pedrocchi-Fantoni G, Spreafico F (1982) J. Org. Chem. 47: 3289
Fuganti C, Grasselli P, Spreafico F, Zirotti C (1984) J. Org. Chem. 49: 543
Pohl M, Lingen B, Müller M (2002) Chem. Eur. J. 8: 5288
Kren V, Crout DHG, Dalton H, Hutchinson DW, König W, Turner MM, Dean G, Thomson N (1993) J. Chem. Soc., Chem. Commun. 341
Bornemann S, Crout DHG, Dalton H, Hutchinson DW, Dean G, Thomson N, Turner MM (1993) J. Chem. Soc., Perkin Trans. 1, 309
Dünkelmann P, Kolter-Jung D, Nitsche A, Demir AS, Siegert P, Lingen B, Baumann M, Pohl M, Müller M (2002) J. Am. Chem. Soc. 124: 12084
Pohl M, Gocke D, Müller M (2009) Thiamine-based enzymes for biotransformations. In: Crabtree RH (ed) Handbook of Green Chemistry, vol. 3: Biocatalysis. Wiley-VCH, Weinheim, p 75
Iding H, Siegert P, Mesch K, Pohl M (1998) Biochim. Biophys. Acta 1385: 307
Baykal A, Chakraborty S, Dodoo A, Jordan F (2006) Bioorg. Chem. 34: 380
Bringer-Meyer S, Sahm H (1988) Biocatalysis 1: 321
Neuser F, Zorn H, Berger RG (2000) Z. Naturforsch. 55: 560
Neuser F, Zorn H, Berger RG (2000) J. Agric. Food Chem. 48: 6191
Kurniadi T, Bel-Rhlid R, Fay LB, Juillerat MA, Berger RG (2003) J. Agric. Food Chem. 51: 3103
Siegert P, McLeish MJ, Baumann M, Iding H, Kneen MM, Kenyon GL, Pohl M (2005) Prot. Eng. Des. Sel. 18: 345
Iding H, Dünnwald T, Greiner L, Liese A, Müller M, Siegert P, Grötzinger J, Demir AS, Pohl M (2000) Chem. Eur. J. 6: 1483
Demir AS, Dünnwald T, Iding H, Pohl M, Müller M (1999) Tetrahedron Asymmetry 10: 4769
Janzen E, Müller M, Kolter-Jung D, Kneen MM, McLeish M J, Pohl M (2006) Bioorg. Chem. 34: 345
Demir AS, Ayhan P, Sopaci SB (2007) Clean 35: 406
Dünkelmann P, Pohl M, Müller M (2004) Chimica Oggi/Chem. Today 22: 24
Demir AS, Ayan P, Igdir A C, Guygu AN (2004) Tetrahedron 60: 6509
Demir AS, Pohl M, Janzen E, Müller M (2001) J. Chem. Soc. Perkin Trans. 1, 633
Demir AS, Sesenoglu O, Eren E, Hosrik B, Pohl M, Janzen E, Kolter D, Feldmann R, Dünkelmann P, Müller M (2002) Adv. Synth. Catal. 344: 96
Cosp A, Dresen C, Pohl M, Walter L, Röhr C, Müller M (2008) Adv. Synth. Catal. 350: 759
Thunberg L, Backstrom G, Lindahl U (1982) Carbohydr. Res. 100: 393
Morris KG, Smith MEB, Turner NJ, Lilly MD, Mitra RK, Woodley JM (1996) Tetrahedron Asymmetry 7: 2185
Demuynck C, Bolte J, Hecquet L, Dalmas V (1991) Tetrahedron Lett. 32: 5085
Humphrey AJ, Turner NJ, McCague R, Taylor SCJ (1995) J. Chem. Soc., Chem. Commun. 2475
Bolte J, Demuynck C, Samaki H (1987) Tetrahedron Lett. 28: 5525
Kobori Y, Myles DC, Whitesides GM (1992) J. Org. Chem. 57: 5899
Mocali A, Aldinucci D, Paoletti F (1985) Carbohydr. Res. 143: 288
Turner NJ (2000) Curr. Opinion Biotechnol. 11: 527
Datta A, Racker E (1961) J. Biol. Chem. 236: 617
Villafranca J, Axelrod B (1971) J. Biol. Chem. 246: 3126
Hobbs GR, Lilly MD, Turner NJ, Ward JM, Willetts AJ, Woodley JM (1993) J. Chem. Soc., Perkin Trans. 1, 165
Hobbs GR, Mitra RK, Chauhan RP, Woodley JM, Lilly MD (1996) J. Biotechnol. 45: 173
Effenberger F, Null V, Ziegler T (1992) Tetrahedron Lett. 33: 5157
Gocke D, Nguyen C L, Pohl M, Stillger T, Walter L, Müller M (2007) Adv. Synth. Catal. 349: 1425
Kitazume T, Ishikawa N (1984) Chem. Lett. 1815
Kitazume T, Ikeya T, Murata K (1986) J. Chem. Soc., Chem. Commun. 1331
Svedendahl M, Jovanovic B, Fransson L, Berglund P (2009) ChemCatChem 1: 252
Strohmeier GA, Sovic T, Steinkellner G, Hartner FS, Andryushkova A, Purkarthofer T, Glieder A, Gruber K, Griengl H (2009) Tetrahedron 65: 5663
Svedendahl M, Hult K, Berglund P (2005) J. Am. Chem. Soc. 127: 17988
Carlqvist P, Svedendahl M, Branneby C, Hult K, Brinck T, Berglund P (2005) ChemBioChem 6: 331
References to Sect. 2.5
Sheldon RA (2000) Pure Appl. Chem. 72: 1233
Sheldon RA (1997) J. Chem. Technol. Biotechnol. 68: 381
Smitskamp-Wilms E, Brussee J, van der Gen A, van Scharrenburg GJM, Sloothaak JB (1991) Rec. Trav. Chim. Pays-Bas 110: 209
Johnson DV, Griengl H (1999) Adv. Biochem. Eng. Biotechnol. 63: 31
Effenberger FX (1992) (R)- and (S)-cyanohydrins – their enzymatic synthesis and their reactions. In: Servi S (ed) Microbial Reagents in Organic Synthesis. Kluwer, Dordrecht, p 25
Kruse CG (1992) Chiral cyanohydrins – their manufacture and utility as chiral building blocks. In: Collins AN, Sheldrake GN, Crosby J (eds) Chirality in Industry. Wiley, New York, p 279
van Scharrenburg GJM, Sloothaak JB, Kruse CG, Smitskamp-Wilms E (1993) Ind. J. Chem. 32B: 16
Rosenthaler I (1908) Biochem. Z. 14: 238
Effenberger F (1994) Angew. Chem., Int. Ed. 33: 1555
Matsuo T, Nishioka T, Hirano M, Suzuki Y, Tsushima K, Itaya N, Yoshioka H (1980) Pestic. Sci. 202
Krepski LR, Jensen KM, Heilmann SM, Rasmussen JK (1986) Synthesis 301
Jackson WR, Jacobs HA, Matthews BR, Jayatilake GS, Watson KG (1990) Tetrahedron Lett. 31: 1447
Brussee J, van Benthem RATM, Kruse CG, van der Gen A (1990) Tetrahedron Asymmetry 1: 163
Effenberger F, Stelzer U (1991) Angew. Chem. Int. Ed. 30: 873
Nahrstedt A (1985) Pl. Syst. Evol. 150: 35
Fechter MH, Griengl H (2004) Food Technol. Biotechnol. 42: 287
Sharma M, Sharma NN, Bhalla TC (2005) Enzyme Microb. Technol. 37: 279
Effenberger F, Förster S, Wajant H, (2000) Curr. Opinion Biotechnol. 11: 532
Griengl H, Schwab H, Fechter M (2000) Trends Biotechnol. 18: 252
Hochuli E (1983) Helv. Chim. Acta 66: 489
Jorns MS, Ballenger C, Kinney G, Pokora A, Vargo D (1983) J. Biol. Chem. 258: 8561
Kiljunen E, Kanerva LT (1997) Tetrahedron Asymmetry 8: 1225
Becker W, Pfeil E (1964) Naturwissensch. 51: 193
Niedermeyer U, Kula MR (1990) Angew. Chem. Int. Ed. 29: 386
Klempier N, Griengl H, Hayn M (1993) Tetrahedron Lett. 34: 4769
Förster S, Roos J, Effenberger F, Wajant H, Sprauer A (1996) Angew. Chem. Int. Ed. 35: 437
Effenberger F, Hörsch B, Förster S, Ziegler T (1990) Tetrahedron Lett. 31: 1249
Seely MK, Criddle RS, Conn EE (1966) J. Biol. Chem. 241: 4457
Selmar D, Lieberei R, Biehl B, Conn EE (1989) Physiol. Plantarum 75: 97
Klempier N, Pichler U, Griengl H (1995) Tetrahedron Asymmetry 6: 845
Kuroki GW, Conn EE (1989) Proc. Natl. Acad. Sci. USA 86: 6978
Bourquelot E, Danjou E (1905) J. Pharm. Chim. 22: 219
Hughes J, De Carvalho FJP, Hughes MA (1994) Arch. Biochem. Biophys. 311: 496
Wagner UG, Hasslacher M, Griengl H, Schwab H, Kratky C (1996) Structure 4: 811
Gruber K, Kratky C (2004) J. Polym. Sci [A] 42: 479
Zuegg J, Gruber K, Gugganig M, Wagner UG, Kratky C(1999) Protein Sci. 8: 1990
Brussee J, Loos WT, Kruse CG, van der Gen A (1990) Tetrahedron 46: 979
Effenberger F, Ziegler T, Förster S (1987) Angew. Chem. Int. Ed. 26: 458
Ziegler T, Hörsch B, Effenberger F (1990) Synthesis 575
Becker W, Pfeil E (1966) J. Am. Chem. Soc. 88: 4299
Becker W, Pfeil E (1966) Biochem. Z. 346: 301
Effenberger F, Hörsch B, Weingart F, Ziegler T, Kühner S (1991) Tetrahedron Lett. 32: 2605
Effenberger F, Heid S (1995) Tetrahedron Asymmetry 6: 2945
Weis R, Gaisberger R, Skranc W, Gruber K, Glieder A (2005) Angew. Chem. Int. Ed. 44: 4700
Glieder A, Weis R, Skranc W, Poechlauer P, Dreveny I, Majer S, Wubbolts M, Schwab H, Gruber K (2003) Angew. Chem. Int. Ed. 42, 4815
Griengl H, Hickel A, Johnson DV, Kratky C, Schmidt M, Schwab H (1997) Chem. Commun. 1933
Sheldon RA, Schoemaker HE, Kamphuis J, Boesten WHJ, Meijer EM (1988) Enzymatic methods for the industrial synthesis of optically active compounds. In: Ariens EJ, van Rensen JJS, Welling W (eds) Stereoselectivity of Pestcides. Elsevier, Amsterdam, p 409
Wehtje E, Adlercreutz P, Mattiasson B (1990) Biotechnol. Bioeng. 36: 39
Ognyanov VI, Datcheva VK, Kyler KS (1991) J. Am. Chem. Soc. 113: 6992
Menendez E, Brieva R, Rebolledo F, Gotor V (1995) J. Chem. Soc., Chem. Commun. 989
Purkarthofer T, Gruber K, Gruber-Khadjawi M, Waich K, Skranc W, Mink D, Griengl H (2006) Angew. Chem. Int. Ed. 45: 3454
Gruber-Khadjawi M, Purkarthofer T, Skranc W, Griengl H (2007) Adv. Synth. Catal. 349: 1445
Hill RL, Teipel JW (1971) Fumarase and Crotonase. In: Boyer P (ed) The Enzymes, vol 5, Academic Press, New York, p 539
Hanson KR, Havir EA (1972) The enzymic elimination of ammonia. In: Boyer P (ed) The Enzymes, vol. 7. Academic Press, New York, p 75
Botting NP, Akhtar M, Cohen MA, Gani D (1987) J. Chem. Soc., Chem. Commun. 1371
Nuiry II, Hermes JD, Weiss PM, Chen C, Cook PF (1984) Biochemistry 23: 5168
Woods SA, Miles JS, Roberts RE, Guest JR (1986) Biochem. J. 237: 547
Findeis MA, Whitesides GM (1987) J. Org. Chem. 52: 2838
Michielsen MJF, Frielink C, Wijffels RH, Tramper J, Beeftink HH (2000) J. Biotechnol. 79: 13
van der Werf M, van den Tweel W, Hermans S (1992) Appl. Environ. Microbiol. 58: 2854
Subramanian SS, Rao MRR (1968) J. Biol. Chem. 243: 2367
Mattey M (1992) Crit. Rev. Biotechnol. 12: 87
Tosa T, Shibatani T (1995) Ann. NY Acad. Sci. 750: 364
Kieslich K (1991) Acta Biotechnol. 11: 559
Hasegawa J, Ogura M, Kanema H, Noda N, Kawaharada H, Watanabe K (1982) J. Ferment. Technol. 60: 501
Obon JM, Maiquez JR, Canovas M, Kleber HP, Iborra JL (1997) Enzyme Microb. Technol. 21: 531
Meyer HP (1993) Chimia 47: 123
Hoeks FWJMM, Muehle J, Boehlen L, Psenicka I (1996) Chem. Eng. J. 61: 53
Fronza G, Fuganti C, Grasselli P, Poli G, Servi S (1988) J. Org. Chem. 53: 6154
Abraham WR, Arfmann HA (1989) Appl. Microbiol. Biotechnol. 32: 295
Holland HL, Gu JX (1998) Biotechnol. Lett. 20: 1125
Chibata I, Tosa T, Sato T (1976) Methods Enzymol. 44: 739
Kumagai H (2000) Adv. Chem. Eng. Biotechnol. 69: 71
Terasawa M, Yukawa H, Takayama Y (1985) Proc. Biochem. 20: 124
Yamagata H, Terasawa M, Yukawa H (1994) Catal. Today 22: 621
Shi W, Dunbar J, Jayasekera MMK, Viola RE, Farber GK (1997) Biochemistry 36: 9136
Viola RE (2000) Adv. Enzymol. Relat. Areas Mol. Biol. 74: 295
Fujii T, Sakai H, Kawata Y, Hata Y (2003) J. Mol. Biol. 328: 635
Weiner B, Poelarends GJ, Janssen DB, Feringa BL (2008) Chem. Eur. J. 14: 10094
Gulzar MS, Akhtar M, Gani D (1997) J. Chem. Soc., Perkin Trans. 1, 649
Emery TF (1963) Biochemistry 2: 1041
Akhtar M, Botting NP, Cohen MA, Gani D (1987) Tetrahedron 43: 5899
Akhtar M, Cohen MA, Gani D (1986) J. Chem. Soc., Chem. Commun. 1290
Breuer M, Hauer B (2003) Curr. Opinion Biotechnol. 14: 570
Steele CL, Chen Y, Dougherty BA, Li W, Hofstead S, Lam KS, Xing Z, Chiang SJ (2005) Arch. Biochem. Biophys. 438: 1
Christianson CV, Montavon TJ, Festin GM, Cooke HA, Shen B, Bruner SD (2007) J. Am. Chem. Soc. 129: 15744
Szymanski W, Wu B, Weiner B, de Wildeman S, Feringa BL, Janssen DB (2009) J. Org. Chem. 74: 9152
Wu B, Szymanski W, Wietzes P, de Wildeman S, Poelarends GJ, Feringa BL, Janssen DB (2009) ChemBioChem 10: 338
References to Sect. 2.6
Kennedy JF, White CA (1983) Bioactive Carbohydrates. Ellis Horwood, West Sussex
Ginsburg V, Robbins PW (eds) (1984) Biology of Carbohydrates. Wiley, New York
Karlsson KA (1989) Ann. Rev. Biochem. 58: 309
Hakomori S (1984) Ann. Rev. Immunol. 2: 103
Laine RA (1994) Glycobiology 4: 759
Boons GJ (1996) Tetrahedron 52: 1095
Nilsson KGI (1988) Trends Biotechnol. 6: 256
Wong CH (1996) Acta Chem. Scand. 50: 211
Okamoto K, Goto T (1990) Tetrahedron 46: 5835
Gigg J, Gigg R (1990) Topics Curr. Chem. 154: 77
Cote GL, Tao BY (1990) Glycoconjugate J. 7: 145
Wong CH, Halcomb RL, Ichikawa Y, Kajimoto T (1995) Angew. Chem. Int. Ed. 34: 521
Edelman J (1956) Adv. Enzymol. 17: 189
Leloir LF (1971) Science 172: 1299
Sharon N (1975) Complex Carbohydrates. Addison-Wesley, Reading, MA
Schachter H, Roseman S (1980) In: Lennarz WJ (ed) The Biochemistry of Glycoproteins and Proteoglycans. Plenum Press, New York, p 85
Beyer TA, Sadler JE, Rearick JI, Paulson JC, Hill RL (1981) Adv. Enzymol. 52: 23
Öhrlein R (1999) Topics Curr. Chem. 200: 227
Elling L (1999) Glycobiotechnologhy: enzymes for the synthesis of nucleotide sugars. In: Scheper T (ed) New Enzymes for Organic Synthesis. Springer, Berlin Heidelberg New York, p 89
Elling L (1997) Adv. Biochem. Eng. Biotechnol. 58: 89
Utagawa T (1999) J. Mol. Catal. B 6: 215
Wong CH (1996) Practical synthesis of oligosaccharides based on glycosyl transferases and glycosylphosphites. In: Khan SH, O’Neill RA (eds) Modern Methods in Carbohydrate Synthesis, vol 19. Harwood, Amsterdam, p 467
Sadler JE, Beyer TA, Oppenheimer CL, Paulson JC, Prieels JP, Rearick JI, Hill RL (1982) Methods Enzymol. 83: 458
Aoki D, Appert HE, Johnson D, Wong SS, Fukuda MN (1990) EMBO J. 9: 3171
Ginsburg V (1964) Adv. Enzymol. 26: 35
Kochetkov NK, Shibaev VN (1973) Adv. Carbohydr. Chem. Biochem. 28: 307
Palcic MM, Srivastava OP, Hindsgaul O (1987) Carbohydr. Res. 159: 315
Berliner LJ, Robinson RD (1982) Biochemistry 21: 6340
Lambright DG, Lee TK, Wong SS (1985) Biochemistry 24: 910
Augé C, David S, Mathieu C, Gautheron C (1984) Tetrahedron Lett. 25: 1467
Srivastava OP, Hindsgaul O, Shoreibah M, Pierce M (1988) Carbohydr. Res. 179: 137
Palcic MM, Venot AP, Ratcliffe RM, Hindsgaul O (1989) Carbohydr. Res. 190: 1
Nunez HA, Barker R (1980) Biochemistry 19: 489
David S, Augé C (1987) Pure Appl. Chem. 59: 1501
Wong CH, Haynie SL, Whitesides GM (1982) J. Org. Chem. 47: 5416
Goedl C, Schwarz A, Mueller M, Brecker L, Nidetzky B (2008) Carbohydr. Res. 343: 2032
Kitaoka M, Hayashi K (2002) Trends Glycosci. Glycotechnol. 14: 35
Lairson LL, Withers SG (2004) Chem. Commun. 2243
Haynie SL, Whitesides GM (1990) Appl. Biochem. Biotechnol. 23: 155
Goedl C, Sawangwan T, Wildberger P, Nidetzky B (2010) Biocatal. Biotransform. 28: 10
Sawangwan T, Goedl C, Nidetzky B (2009) Org. Biomol. Chem. 7: 4267
Goedl C, Sawangwan T, Mueller M, Schwarz A, Nidetzky B (2008) Angew. Chem. Int. Ed. 47: 10086
Henrissat B, Davies G (1997) Curr. Opinion Struct. Biol. 7: 637
Wallenfels K, Weil R (1972) In: Boyer PD (ed) The Enzymes, vol VII. Academic Press, New York, p 618
Pan SC (1970) Biochemistry 9: 1833
Scigelova M, Singh S, Crout DHG (1999) J. Mol. Catal. B 6: 483
Crout DHG, Vic G (1998) Curr. Opinion Chem. Biol. 2: 98
Nilsson KGI (1996) Synthesis with glycosidases. In: Khan SH, O’Neill RA (eds) Modern Methods in Carbohydrate Synthesis, vol 21. Harwood, Amsterdam, p 518
van Rantwijk F, Woudenberg-van Oosterom M, Sheldon RA (1999) J. Mol. Catal. B 6: 511
Bourquelot E, Bridel M (1913) Ann. Chim. Phys. 29: 145
Wang Q, Graham RW, Trimbur D, Warren RAJ, Withers SG (1994) J. Am. Chem. Soc. 116: 11594
Post CB, Karplus M (1986) J. Am. Chem. Soc. 108: 1317
Huang X, Tanaka KSE, Bennet AJ (1997) J. Am. Chem. Soc. 119: 11147
Chiba S (1997) Biosci. Biotechnol. Biochem. 61: 1233
Withers SG, Street IP (1988) J. Am. Chem. Soc. 110: 8551
Koshland DE (1953) Biol. Rev. 28: 416
Heightman TD, Vasella AT (1999) Angew. Chem. Int. Ed. 38: 750
Sinnot ML (1990) Chem. Rev. 90: 1171
Wang Q, Withers SG (1995) J. Am. Chem. Soc. 117: 10137
Veibel S (1936) Enzymologia 1: 124
Li YT (1967) J. Biol. Chem. 242: 5474
Johansson E, Hedbys L, Mosbach K, Larsson PO, Gunnarson A, Svensson S (1989) Enzyme Microb. Technol. 11: 347
Likolov ZL, Meagher MM, Reilly PJ (1989) Biotechnol. Bioeng. 34: 694
Rastall RA, Bartlett TJ, Adlard MW, Bucke C (1990) The production of hetero-oligosaccharides using glycosidases. In: Copping LG, Martin RE, Pickett JA, Bucke C, Bunch AW (eds) Opportunities in Biotransformations. Elsevier, London, p 47
Vulfson EN, Patel R, Beecher JE, Andrews AT, Law BA (1990) Enzyme Microb. Technol. 12: 950
Beecher JE, Andrews AT, Vulfson EN (1990) Enzyme Microb. Technol. 12: 955
Fujimoto H, Nishida H, Ajisaka K (1988) Agric. Biol. Chem. 52: 1345
Kren V, Thiem J (1997) Chem. Soc. Rev. 26: 463
Vocadlo DJ, Withers SG (2000) In: Ernst B, Hart GW, Sinay P (eds) Carbohydrates in Chemistry and Biology. Wiley-VCH, Weinheim 2: 723
Husakova L, Riva S, Casali M, Nicotra S, Kuzma M, Hunkova Z, Kren V (2001) Carbohydr. Res. 331: 143
Fialova P, Weignerova L, Rauvolfova J, Prikrylova V, Pisvejcova A, Ettrich R, Kuzma M, Sedmera P, Kren V (2004) Tetrahedron 60: 693
Gold AM, Osber MP (1971) Biochem. Biophys. Res. Commun. 42: 469
Williams SJ, Withers SG (2000) Carbohydr. Res. 327: 27
Fialova P, Carmona AT, Robina I, Ettrich R, Sedmera P, Prikrylova V, Petraskova-Husakova L, Kren V (2005) Tetrahedron Lett. 46: 8715
Day AG, Withers SG (1986) Biochem. Cell Biol. 64: 914
Yasukochi T, Fukase K, Kusumoto S (1999) Tetrahedron Lett. 40: 6591
Chiffoleau-Giraud V, Spangenberg P, Rabiller C (1997) Tetrahedron Asymmetry 8: 2017
Kobayashi S, Kiyosada T, Shoda S (1997) Tetrahedron Lett. 38: 2111
Nilsson KGI (1990) Asymmetric synthesis of complex oligosaccharides. In: Copping LG, Martin RE, Pickett JA, Bucke C, Bunch AW (eds) Opportunities in Biotransformations. Elsevier, London, p 131
Nilsson KGI (1990) Carbohydr. Res. 204: 79
Nilsson KGI (1988) Ann. NY Acad. Sci. USA 542: 383
Nilsson KGI (1987) A comparison of the enzyme-catalysed formation of peptides and oligosaccharides in various hydroorganic solutions using the nonequilibrium approach. In: Laane C, Tramper J, Lilly MD (eds) Biocatalysis in Organic Media. Elsevier, Amsterdam, p 369
Nilsson KGI (1987) Carbohydr. Res. 167: 95
Nilsson KGI (1989) Carbohydr. Res. 188: 9
Nilsson KGI (1988) Carbohydr. Res. 180: 53
Crout DHG, Howarth OW, Singh S, Swoboda BEP, Critchley P, Gibson WT (1991) J. Chem. Soc., Chem. Commun. 1550
Huber RE, Gaunt MT, Hurlburt KL (1984) Arch. Biochem. Biophys. 234: 151
Ooi Y, Mitsuo N, Satoh T (1985) Chem. Pharm. Bull. 33: 5547
Mitsuo N, Takeichi H, Satoh T (1984) Chem. Pharm. Bull. 32: 1183
Trincone A, Improta R, Nucci R, Rossi M, Giambacorta A (1994) Biocatalysis 10: 195
Björkling F, Godtfredsen SE (1988) Tetrahedron Lett. 44: 2957
Gais HJ, Zeissler A, Maidonis P (1988) Tetrahedron Lett. 29: 5743
Crout DHG, MacManus DA, Critchley P (1991) J. Chem. Soc., Chem. Commun. 376
Matsumura S, Yamazaki H, Toshima K (1997) Biotechnol. Lett. 19: 583
Kengen SWM, Luesink EJ, Stams AJM, Zehnder AJB (1993) Eur. J. Biochem. 213: 305
Perugino G, Trincone A, Rossi M, Moracci M (2004) Trends Biotechnol. 22: 31
Williams SJ, Withers SG (2002) Austr. J. Chem. 55: 3
Jahn M, Chen H, Muellegger J, Marles J, Warren RAJ, Withers SG (2004) Chem. Commun. 274
Perugino G, Cobucci-Ponzano B, Rossi M, Moracci M (2005) Adv. Synth. Catal. 347: 941
Jahn M, Marles J, Warren RAJ, Withers SG (2003) Angew. Chem. Int. Ed. 42: 352
Kim YW, Fox DT, Hekmat O, Kantner T, McIntosh LP, Warren RAJ, Withers SG (2006) Org. Biomol. Chem. 4: 2025
Turner N J, Truppo M D (2010) Biocatalytic routes to non-racemic chiral amines, In: Nugent TC (ed) Chital Amine Synthesis. Wiley-VCH, Weinheim, p 431
Koszelewski D, Tauber K, Faber K, Kroutil W (2010) Trends Biotechnol. 28: 324
Zhu D, Hua L (2009) Biotechnol. J. 4: 1420
Höhne M, Bornscheuer UT (2009) ChemCatChem 1: 42
Stewart JD (2001) Curr. Opinion Chem. Biol. 5: 120
Ager DJ, Li T, Pantaleone DP, Senkpeil RF, Taylor PP, Fotheringham IG (2001) J. Mol. Catal. B: Enzym. 11: 199
Noe FF, Nickerson WJ (1958) J. Bacteriol. 75: 674
Kim K H (1964) J. Biol. Chem. 239: 783
Matcham GW, Bowen ARS (1996) Chim. Oggi 14: 20
Cho BK, Park HY, Seo JH, Kinnera K, Lee BS, Kim BG (2004) Biotechnol. Bioeng. 88: 512
Mehta PK, Hale TI, Christen P (1993) Eur. J. Biochem. 214: 549
Hwang BY, Cho BK, Yun H, Koteshwar K, Kim BG (2005) J. Mol. Catal. B: Enzym. 37: 47
Shin JS, Kim BG (2001) Biosci. Biotechnol. Biochem. 65: 1782
Iwasaki A, Yamada Y, Ikenaka Y, Hasegawa J (2003) Biotechnol. Lett. 25: 1843
Koszelewski D, Lavandera I, Clay D, Rozzell D, Kroutil W (2008) Adv. Synth. Catal. 350: 2761
Hwang BY, Kim BG (2004) Enzyme Microb. Technol. 34: 429
Yun H, Cho BK, Kim BG, (2004) Biotechnol. Bioeng. 87: 772
Smithies K, Smith MEB, Kaulmann U, Galman JL, Ward JM, Hailes HC (2009) Tetrahedron Asymmetry 20: 570
Kaulmann U, Smithies K, Smith MEB, Hailes H C, Ward JM (2007) Enzyme Microb. Technol. 41: 628
Ingram CU, Bommer M, Smith MEB, Dalby PA, Ward JM, Hailes HC, Lye GJ (2007) Biotechnol. Bioeng. 96: 559
Hanson RL, Davis BL, Chen Y, Goldberg SL, Parker WL, Tully TP, Montana MA, Patel RN (2008) Adv. Synth. Catal. 350: 1367
Yun H, Lim S, Cho BK, Kim BG, (2004) Appl. Environ. Microbiol. 70: 2529
Koszelewski D, Göritzer M, Clay D, Seisser B, Kroutil W (2010) ChemCatChem 2: 73
Shin JS. Kim BG (1998) Biotechnol. Bioeng. 60: 534
Matcham G, Bhatia M, Lang W, Lewis C, Nelson R, Wang A, Wu W (1999) Chimia 53: 584
Helaine V, Bolte J (1999) Eur. J. Org. Chem. 3403
Li T, Kootstra AB, Fotheringham IG (2002) Org. Proc. Res. Dev. 6: 533
Lo HH, Hsu SK, Lin WD, Chan NL, Hsu WH (2005) Biotechnol. Progr. 21: 411
Hoene M, Kuehl S, Robins K, Bornscheuer UT (2008) ChemBioChem 9: 363
Hwang JY, Park J, Seo JH, Cha M, Cho BK, Kim J, BG Kim (2009) Biotechnol. Bioeng. 102: 1323
Yun H, Kim BG (2008) Biosci. Biotechnol. Biochem. 72: 3030
Truppo MD, Rozzell JD, Moore JC, Turner NJ (2009) Org. Biomol. Chem. 7: 395
Koszelewski D, Lavandera I, Clay D, Guebitz GM, Rozzell D, Kroutil W (2008) Angew. Chem. Int. Ed. 47: 9337
References to Sect. 2.7
Petty MA (1961) Bacteriol. Rev. 25: 111
Field JA, Verhagen FJM, de Jong E (1995) Trends Biotechnol. 13: 451
Gschwend PM, MacFarlane JK, Newman KA (1985) Science 227: 1033
Siuda JF, De Barnardis JF (1973) Lloydia 36: 107
Fowden L (1968) Proc. Royal Soc. London B 171: 5
Fenical W (1979) Recent Adv. Phytochem. 13: 219
Wever R, Krenn BE (1990) Vanadium Haloperoxidases. In: Chasteen ND (ed) Vanadium in Biological Systems. Kluwer, Dordrecht, p 81
Krenn BE, Tromp MGM, Wever R (1989) J. Biol. Chem. 264: 19287
Faulkner DJ (1984) Nat. Prod. Rep. 1: 251
Morris DR, Hager LP (1966) J. Biol. Chem. 241: 1763
van Pée KH (1990) Kontakte (Merck) 41
Neidleman SL, Geigert J (1986) Biohalogenation. Ellis Horwood, Chichester
Franssen MCR, van der Plas HC (1992) Adv. Appl. Microbiol. 37: 41
Neidleman SL (1980) Hydrocarbon Proc. 60: 135
van Pee KH (2001) Arch. Microbiol. 175: 250
Fujimori D G, Walsh C T (2007) Curr. Opinion Chem. Biol. 11: 553
Anderson JLR, Chapman SK (2006) Mol. BioSyst. 2: 350
Murphy C D (2006) Nat. Prod. Rep. 23: 147
van Pee KH, Patallo EP (2006) Appl. Microbiol. Biotechnol. 70: 631
van Pee KH, Flecks S, Patallo EP (2007) Chimica Oggi 25: 22
Krebs K, Fujimori D G, Walsh CT, Bollinger Jr JM, (2007) Acc. Chem. Res. 40: 484
van Pee KH (1990) Biocatalysis 4: 1
Neidleman SL, Geigert J (1987) Endeavour 11: 5
Hofrichter M, Ullrich R (2006) Appl. Microbiol. Biotechnol. 71: 276
Itoh N, Izumi Y, Yamada H (1985) J. Biol. Chem. 261: 5194
van Pée KH, Lingens F (1985) J. Bacteriol. 161: 1171
Wiesner W, van Pée KH, Lingens F (1985) Hoppe-Seylers Z. Physiol. Chem. 366: 1085
van Pée KH, Lingens F (1985) J. Gen. Microbiol. 131: 1911
Wagner AP, Psarrou E, Wagner LP (1983) Anal. Biochem. 129: 326
Fukuzawa A, Aye M, Murai A (1990) Chem. Lett. 1579
Thomas JA, Morris DR, Hager LP (1970) J. Biol. Chem. 245: 3135
Yamada H, Itoh N, Izumi Y (1985) J. Biol. Chem. 260: 11962
Libby RD, Shedd AL, Phipps AK, Beachy TM, Gerstberger SM (1992) J. Biol. Chem. 267: 1769
Wagenknecht HA, Woggon WD (1997) Chem. Biol. 4: 367
Turk J, Henderson WR, Klebanoff SJ, Hubbard WC (1983) Biochim. Biophys. Acta 751: 189
Boeynaems JM, Watson JT, Oates JA, Hubbard WC (1981) Lipids 16: 323
Geigert J, Neidleman SL, Dalietos DJ, DeWitt SK (1983) Appl. Environ. Microbiol. 45: 1575
Lee TD, Geigert J, Dalietos DJ, Hirano DS (1983) Biochem. Biophys. Res. Commun. 110: 880
Neidleman SL, Oberc MA (1968) J. Bacteriol. 95: 2424
Levine SD, Neidleman SL, Oberc MA (1968) Tetrahedron 24: 2979
Geigert J, Neidleman SL, Dalietos DJ, DeWitt SK (1983) Appl. Environ. Microbiol. 45: 366
Neidleman SL, Levin SD (1968) Tetrahedron Lett. 9: 4057
Ramakrishnan K, Oppenhuizen ME, Saunders S, Fisher J (1983) Biochemistry 22: 3271
Geigert J, Neidleman SL, Dalietos DJ (1983) J. Biol. Chem. 258: 2273
Neidleman SL, Cohen AI, Dean L (1969) Biotechnol. Bioeng. 2: 1227
van Pée KH, Lingens F (1984) FEBS Lett. 173: 5
Itoh N, Izumi Y, Yamada H (1987) Biochemistry 26: 282
Jerina D, Guroff G, Daly J (1968) Arch. Biochem. Biophys. 124: 612
Matkovics B, Rakonczay Z, Rajki SE, Balaspiri L (1971) Steroidologia 2: 77
Corbett MD, Chipko BR, Batchelor AO (1980) Biochem. J. 187: 893
Loo TL, Burger JW, Adamson RH (1964) Proc. Soc. Exp. Biol. Med. 114: 60
Beissner RS, Guilford WJ, Coates RM, Hager LP (1981) Biochemistry 20: 3724
Libby RD, Thomas JA, Kaiser LW, Hager LP (1982) J. Biol. Chem. 257: 5030
Franssen MCR, van der Plas HC (1984) Recl. Trav. Chim. Pays-Bas 103: 99
Neidleman SL, Diassi PA, Junta B, Palmere RM, Pan SC (1966) Tetrahedron Lett. 7: 5337
Theiler R, Cook JC, Hager LP, Siuda JF (1978) Science 202: 1094
Zaks A, Yabannavar AV, Dodds DR, Evans CA, Das PR, Malchow R (1996) J. Org. Chem. 61: 8692
Grisham MB, Jefferson MM, Metton DF, Thomas EL (1984) J. Biol. Chem. 259: 10404
Silverstein RM, Hager LP (1974) Biochemistry 13: 5069
Tsan M-F (1982) J. Cell. Physiol. 111: 49
Lal R, Saxena DM (1982) Microbiol. Rev. 46: 95
Alexander M (1977) Introduction to Soil Microbiology. Wiley, Chichester, p. 438
Ghisalba O (1983) Experientia 39: 1247
Rothmel RK, Chakrabarty AM (1990) Pure Appl. Chem. 62: 769
Müller R, Lingens F (1986) Angew. Chem. Int. Ed. 25: 778
Vogel TM, Criddle CS, McCarthy PL (1987) Environ. Sci. Technol. 21: 722
Castro CE, Wade RS, Belser NO (1985) Biochemistry 24: 204
Chacko CI, Lockwood JL, Zabik M (1966) Science 154: 893
Markus A, Klages V, Krauss S, Lingens F (1984) J. Bacteriol. 160: 618
Yoshida M, Fujita T, Kurihara N, Nakajima M (1985) Pest. Biochem. Biophysiol. 23: 1
Leisinger T, Bader R (1993) Chimia 47: 116
Hardman DJ (1991) Crit. Rev. Biotechnol. 11:1
Weightman AJ, Weightman AL, Slater JH (1982) J. Gen. Microbiol. 128: 1755
Soda K, Kurihara T, Liu J-Q, Nardi-Dei V, Park C, Miyagi M, Tsunasawa S, Esaki N (1996) Pure Appl. Chem. 68: 2097
Motosugi K, Esaki N, Soda K (1982) Agric. Biol. Chem. 46: 837
Allison N, Skinner AJ, Cooper RA (1983) J. Gen. Microbiol. 129: 1283
Kawasaki H, Miyoshi K, Tonomura K (1981) Agric. Biol. Chem. 45: 543
Walker JRL, Lien BC (1981) Soil Biol. Biochem. 13: 231
Kawasaki H, Tone N, Tonomura K (1981) Agric. Biol. Chem. 45: 35
Onda M, Motosugi K, Nakajima H (1990) Agric. Biol. Chem. 54: 3031
Tsang JSH, Sallis PJ, Bull AT, Hardman DJ (1988) Arch. Microbiol. 150: 441
Little M, Williams PA (1971) Eur. J. Biochem. 21: 99
Vyazmensky M, Geresh S (1998) Enzyme Microb. Technol. 22: 323
Cambou B, Klibanov AM (1984) Appl. Biochem. Biotechnol. 9: 255
Taylor SC (1990) (S)-2-Chloropropanoic acid by biotransformation. In: Copping LG, Martin RE, Pickett JA, Bucke C, Bunch AW (eds) Opportunities in Biotransformations. Elsevier, London, p 170
Smith JM, Harrison K, Colby J (1990) J. Gen. Microbiol. 136: 881
Barth PT, Bolton L, Thomson JC (1992) J. Bacteriol. 174: 2612
Taylor SC (1997) (S)-2-Chloropropanoic Acid: Developments in Its Industrial Manufacture. In: Collins AN, Sheldrake GN, Crosby J (eds) Chirality in Industry II. Wiley, Chichester, p 207
Hasan AKMQ, Takata H, Esaki N, Soda K (1991) Biotechnol. Bioeng. 38: 1114
van den Wijngaard AJ, Reuvekamp PTW, Janssen DB (1991) J. Bacteriol. 173: 124
Castro CE, Bartnicki EW (1968) Biochemistry 7: 3213
Geigert J, Neidleman SL, Liu TN, DeWitt SK, Panschar BM, Dalietos DJ, Siegel ER (1983) Appl. Environ. Microbiol. 45: 1148
Nagasawa T, Nakamura T, Yu F, Watanabe I, Yamada H (1992) Appl. Microbiol. Biotechnol. 36: 478
Assis HMS, Sallis PJ, Bull AT, Hardman DJ (1998) Enzyme Microb. Technol. 22: 568
Assis HMS, Bull AT, Hardman DJ (1998) Enzyme Microb. Technol. 22: 545
Kasai N, Tsujimura K, Unoura K, Suzuki T (1990) Agric. Biol. Chem. 54: 3185
Nakamura T, Yu F, Mizunashi W, Watanabe I (1991) Agric. Biol. Chem. 55: 1931
Nakamura T, Nagasawa T, Yu F, Watanabe I, Yamada H (1992) J. Bacteriol. 174: 7613
Kasai N, Tsujimura K, Unoura K, Suzuki T (1992) J. Ind. Microbiol. 9: 97
Kasai N, Suzuki T, Furukawa Y (1998) J. Mol. Catal. B 4: 237
Suzuki T, Kasai N (1991) Bioorg. Med. Chem. Lett. 1: 343
Suzuki T, Kasai N, Minamiura N (1994) Tetrahedron Asymmetry 5: 239
Kasai N, Suzuki T (2002) Adv. Synth. Catal. 345: 437
de Vries EJ, Janssen DB (2003) Curr. Opinion Biotechnol. 14: 414
Tang L, Lutje Spelberg JH, Fraaije MW, Janssen DB (2003) Biochemistry 42: 5378
De Jong RM, Dijkstra BW (2003) Curr. Opinion Struct. Biol. 13: 722
Haak RM, Tarabiono C, Janssen DB, Minnaard AJ, de Vries J G, Feringa BL (2007) Org. Biomol. Chem. 5: 318
Elenkov MM, Hoeffken HW, Tang L, Hauer B, Janssen DB (2007) Adv. Synth. Catal. 349: 2279
Hasnaoui G, Lutje Spelberg JH, de Vries E, Tang L, Hauer B, Janssen DB (2005) Tetrahedron Asymmetry 16: 1685
Elenkov MM, Hauer B, Janssen DB (2006) Adv. Synth. Catal. 348: 579
Hasnaoui-Dijoux G, Elenkov MM, Lutje Spelberg JH, Hauer B, Janssen DB (2008) ChemBioChem 9: 1048
Fuchs M, Simeo Y, Ueberbacher BT, Mautner B, Netscher T, Faber K (2009) Eur. J. Org. Chem. 833
Fox RJ, Davis SC, Mundorff EC, Newman LM, Gavrilovic V, Ma SK, Chung LM, Ching C, Tam S, Muley S, Grate J, Gruber J, Whitman JC, Sheldon RA, Huisman GW (2007) Nature Biotechnol. 25: 338
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Copyright information
© 2011 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Faber, K. (2011). Biocatalytic Applications. In: Biotransformations in Organic Chemistry. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-17393-6_2
Download citation
DOI: https://doi.org/10.1007/978-3-642-17393-6_2
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-17392-9
Online ISBN: 978-3-642-17393-6
eBook Packages: Chemistry and Materials ScienceChemistry and Material Science (R0)