
Abstract
Out of eight Gyps species in the world, three residents (G. bengalensis, G. indicus, G. tenuirostris) and two migratory (G. fulvus, G. himalayensis) inhabit Indian forests and other landscapes. While Himalayan and Eurasian Gyps are near threatened and least concerned species, respectively, the resident Gyps are critically endangered. They are facing modification in habitats caused by anthropogenic factors and are enduring climate change. The impact of climate change has been insufficiently studied. The aim of this study is to predict current and future habitat suitability for these vultures shaped by bioenvironmental factors using maximum-entropy species distribution modelling. Seventy-one robust predictions and models, species and scenario wise (AUC 0.780 to 0.981, TSS 0.478 to 0.852 and CBI 0.978 to 0.997) were generated. Whole Indian landscape (3,287,263 km2) was categorised into unsuitable, moderately suitable and highly suitable habitats and analysed floristic region-wise. There was a reasonable change in habitat suitability which showed a trend of decrease in the suitable area in the future (3287 to 65,745 km2). The key environmental variables shaping current and future habitat included land use/land cover, annual mean temperature (bio1), precipitation of coldest quarter (bio19), precipitation seasonality (bio15) and precipitation of warmest quarter (bio18). Our results on the potential habitat in different floristic regions and projected change in future habitat will aid national and regional managers to design proactive approaches towards conservation of endangered Gyps vultures. Management interventions like in situ conservation, habitat maintenance, advance planning of habitat improvement, expansion of favourable area and protection of suitable area have been proposed.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Vultures are among the most threatened birds throughout the world with a majority of the species at risk (Botha et al. 2017). India is endowed with six resident (bearded vulture, Egyptian vulture, Indian vulture, red-headed vulture, slender-billed vulture and white-rumped vulture) and three wintering (cinereous vulture, Eurasian griffon and Himalayan griffon) old-world vultures (MoEFCC 2020). Five of these Gyps vultures (Indian vulture Gyps indicus = INV, slender-billed vulture Gyps tenuirostris = SBV, white-rumped vulture Gyps bengalensis = WRV, Eurasian griffon Gyps fulvus = EGV, Himalayan griffon Gyps himalayensis = HGV) are phylogenetically closer in comparison to four distant, non-Gyps vultures having different competitive behaviour. The former is comparatively more social and has the advantage of getting information early about carcass presence and stronger group defence but are disadvantaged when it comes to poisoned carcasses, where they are at higher risk. Therefore, the management requirements of these two groups differ significantly (Campbell 2016).
The populations of vultures, in general, have diminished or they have been eliminated in some of their distribution ranges due to habitat loss and other reasons like lack of food resources, exposure to livestock contaminated with drugs and direct persecution like poison or shooting (Green et al. 2004; Ogada et al. 2011; Ilanloo et al. 2020). Decimation of the population of vultures in India was also reported due to shelter destruction along with food shortage and poor breeding success (Chhangani and Mohnot 2004). Hurtt et al. (2011) reviewed that habitat loss is the primary cause of species extinctions. However, Ilanloo et al. (2020) identified climate change as a major factor for species extinction along with changes in habitat distribution within the short span of a few decades. As reviewed by Thapa et al. (2021), climate change may cause redistributions of species directly through changes in temperature and water availability, and indirectly through further habitat modification.
An important step in the recovery of the diminished vulture population (Prakash et al. 2012, 2019; Galligan et al. 2020, 2021; UPFD and BNHS 2021) will be a gain in the knowledge of current and possible future habitats of vultures and their management. For this, ecological modelling having the potential to fill the knowledge gaps regarding species distribution, may provide insight into the impact of climate change and aid in conservation planning (Rodríguez et al. 2007; Mateo et al. 2013). Therefore, species distribution modelling (SDM) could be applied as a tool in determining environmental covariates of habitats, mapping of suitable habitats and predicting the impact of climate change on it (Angelieri et al. 2016). Many SDM algorithms exist, but machine learning approaches have gained popularity due to their ability to fit responses with high predictive performance (Elith and Graham 2009). MaxEnt is one such machine learning algorithm used the most by the ecological modelling community (Merow et al. 2013; Sesink-Clee et al. 2015; Morales et al. 2017; Mohammadi et al. 2019; Urbina-Cardona et al. 2019; Vu et al. 2019) due to its high reliability and statistical robustness among well-established SDMs (Phillips et al. 2004; Elith et al. 2006; Summers et al. 2012; Banag et al. 2015).
Under the above premise, the present study is aimed at the following using MaxEnt SDM: (a) identifying dominant environmental factors shaping the habitat of Gyps vultures; (b) predicting and mapping the current habitat suitability; (c) assessing predicted change in future habitat expanse due to climate change; and (d) proposing broad management interventions based on habitat quality.
Methods
Study area
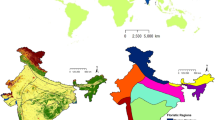
This study covers whole India, a country once known for widespread and abundant populations of Gyps species (Gadhvi and Dodia 2006; Baral et al. 2013), now threatened (IUCN 2022). Topographical heterogeneity, climatic variation, land use/land cover (LULC) pattern, historical occurrence of Gyps vultures and delineation of the floristic regions (FRs) of the study area are presented in Fig. 1 and Fig. 2 along with some detailed features in Supplementary Table 1. Range of elevation, mean annual temperature and mean annual precipitation of the study area is − 1 to 8583 m, − 33.8 to 30.0 °C and 33 to 9312 mm, respectively. The forests in different regions provide nesting and foraging habitats to the vultures due to presence of large trees, mountain cliffs and wildlife population. Outside forests also there is a sizable population of livestock which is another source of carcasses (NDDB 2019).

Location, physiographic and climatic details of the study area, India. Top row: Geographic position of India in the world. Middle row: Land use/Land cover and elevation gradients, Bottom row: Mean annual precipitation and Temperature ranges

Vegetation based divisions of India and occurrences of Gyps vultures. Top row: different floristic regions and occurrence locations of Gyps bengalensis (white-rumped vulture), Bottom row: occurrence locations of Gyps indicus (indian vulture) and Gyps tenuirostris (slender-billed vulture), and Gyps himalayensis (Himalayan griffon) and Gyps fulvus (Eurasian griffon)
Data collection and processing
Presence-only species distribution model like MaxEnt requires species occurrence locations and their environmental covariates. Therefore, nesting, roosting and foraging presence sites of the studied species were collected from published literature (Supplementary Table 2), citizen science repositories ebird (http://www.ebird.org, Sullivan et al. 2009) and iNaturalist (http://www.inaturalist.org, iNaturalist users and Ueda 2020), and author’s field works. Since spatial filtering, also known as spatial rarefying (Brown et al. 2017), improves the performance of ecological niche models by reducing sampling bias (Boria et al. 2014), the occurrence data were cleaned, duplicates were removed and then rarefied. Out of several rarefying distances tried, 4 km gave optimum results in the present case. Original occurrence points of different Gyps species, 2767 (EGV), 10,653 (HGV), 5533 (INV), 617 (SBV) and 4939 (WRV), were reduced after duplicate removal and spatial rarefication to 529 (EGV), 1131 (HGV), 876 (INV), 114 (SBV) and 1163 (WRV), respectively.
The environmental variables like temperature, precipitation, LULC and elevation determine vultures’ habitat in general and shelter in particular (Jha and Jha 2021b). Therefore, predictor environmental covariates like bioclimatic variables required by SDM were downloaded at 30 arc second resolution from https://www.worldclim.org/ (1970–2000; Fick and Hijmans 2017), LULC from Copernicus Global Land Service at 100 m resolution (Buchhorn et al. 2020) and elevation from SRTM 1 Arc second global data from https://earthexpoler.usgs.gov/ (USGS EROS 2018). These layers were resampled at 30 arc second spatial resolution before plugging in MaxEnt. The categorical variable, LULC, available in 23 fine classes was reduced to six (forest, water, scrubland, agriculture, built-up, wasteland) for the present study as suggested by Jha and Jha (2021b). All these environmental variables may possibly have collinearity among them and ignoring the multicollinearity may lead to false ecological conclusions in modelling the spatial distribution of a species (Heikkinen and Luoto 2006; de Frutos et al. 2007). Therefore, Pearson correlation analysis was carried out at ± 0.8 threshold to identify and remove colinear covariates using “Remove highly correlated variables” tool of SDM tool box (Brown et al. 2017). The variables were input alphabetically. Highly correlated variables namely, bio4, bio5, bio6, bio8, bio9, bio10, bio11, bio13, bio16, bio17 and Elevation were eliminated. For further improvement of the models, bias file was prepared using Correcting latitudinal background selection biases tool of SDM tool box (Brown et al. 2017). This was done for minimising overfitting and avoid sampling habitat outside of a species’ known occurrence and account for collection sampling biases with coordinate data (Brown et al. 2017).
Climate model selection
There are several GCMs available with Representative Concentration Pathways (RCP2.6, RCP4.5, RCP6.0, RCP8.5) for different climate scenarios (Stocker 2014) but there is no consensus on which makes the most accurate one. However, Sutton et al. (2015) recommended ensemble of multiple GCMs to reinforce the accuracy of the projections. This is a very common approach for habitat projection (Stiels and Schidelko 2018). Therefore, we chose commonly used GCMs and downloaded precalculated 30 arc second data from WorldClim (Fick and Hijmans 2017): CCSM4 (de Luis et al. 2019), HadGEM2AO (Ahmad et al. 2019) and MIROC5 (Sony et al. 2018). Two RCPs namely, RCP4.5 (moderate) and RCP8.5 (extreme) (de Luis et al. 2019; Vu et al. 2019), for short-term (2041–2060 represented by year 2050) and long-term (2061–2080 represented by year 2070) prediction were chosen based on the hypothesis that a sharp cut in CO2 emission will not happen (Lane 2018) and lower emission scenarios will be unlikely (Manning et al. 2010). Prediction averaging of the three GCMs was done in order to increase model accuracy of habitat suitability area (Dormann et al. 2018).
Distribution modelling
After processing presence locations and environmental variables in ArcGIS, they were used as the input of the MaxEnt model and predictions were made for the different scenarios. The model was run at default MaxEnt settings with a change in the run type, which was set as bootstrap with 10 replicates per prediction and a random test percentage of 25. Feature type used for model training was auto features and the number of background points was 10,000. Six model entities were studied (5 Gyps species and 1 aggregated all species of Gyps or “superspecies”). For each of the 5 species, 2 models (with 10 replicates) were trained: with-LULC and without-LULC models, while for the superspecies entity, only one model was trained (without LULC for comparison). Predictions for the present period were made by all of the models while predictions for the future periods were made by only the without-LULC model due to unknown long-term dynamics of landcover. Climatic predictors used for the future predictions were updated for the corresponding time period.
Tabulation and mapping
Area under receiver operator curve (AUC) values and variable contribution were tabulated, maps were reclassified and jackknife bar charts and response curves were analysed to present the results. Other two model evaluators true skill statistics (TSS) and continuous Boyce index (CBI) were also computed in R using SSDM (Schmitt et al. 2017) and modEvA (Barbosa et al. 2013) packages, respectively. Most of the studies considered reclassification of continuous MaxEnt output range (0–1) into four classes (Vu et al. 2019; Zhang et al. 2019) in a smaller study area. Proposed classification for raptors including vulture by Zhang et al. (2019) was fine tuned in the present study into three categories of suitability (0–0.3 as unsuitable; 0.3–0.6 as moderate and 0.6–1.0 as high). This was done keeping into view coarse scaling in large study area (25 times larger than Zhang et al. 2019) based on previous field experiences (Jha and Jha 2020, 2021a, b). All MaxEnt outputs were also separately reclassified into two categories, unsuitable (0–0.30) and suitable (0.3–1.0), for calculation of future loss or gain in habitat area. The present prediction was then compared with each of the future scenarios to estimate the suitable area lost or gained as a result of climate change. ArcGIS 10.5 and Microsoft Excel were used to process the results. The huge study area (3.28 million km2) was divided into floristic regions of India (Sharma 2005). A map of different floristic regions was digitised using the data from Sharma (2005) and the output maps were analysed accordingly.
Results
Model predictors
The selected non-colinear covariates (Pearson coefficient ± 0.8) namely, bio1, bio2, bio3, bio7, bio12, bio14, bio15, bio18, bio19 and LULC contributed in varying proportions to prediction in different species, as detailed in the covariate contribution table of MaxEnt output. However, in the case of present models with LULC, the average cumulative contribution for all the species (top three covariates) was found to be 71.2% (range 65 to 86%). For the top five covariates, this was found to be 88.5% (range 86 to 97%). Without LULC, it differed marginally, 74.2% (range 62.9 to 91.6%) for the top three covariates and 89.2% (range 84.6 to 96.5%) for the top five covariates. Considering all the species together, the top five rankers or dominant variables (based on modified Likert ranking method; Bhattacherjee 2012), in decreasing order of importance, for without LULC models were bio1, bio19, bio15, bio18 and bio14. Species wise top three covariates were bio19, bio7 and bio18 (EGV); bio1, bio19 and bio14 (HGV); bio15, bio14 and bio1 (INV); bio18, bio1 and bio14 (SBV); and bio15, bio1 and bio12 (WRV). In the case of with LULC models, this order changed which is described in Supplementary Box 1. However, the jackknife chart of training gain (Figs. 3 and 4) showed some variation in the ranking of variables when compared to the variable contribution table. Response charts of LULC components are presented in Supplementary Fig. 1.

Jackknife chart of variable importance in resident Gyps species. Note dark blue, light blue and red bars in jackknife chart (top indian vulture, middle slender-billed vulture and bottom white-rumped vulture) showing importance with only variable, without variable and with all variables, respectively

Jackknife chart of variable importance in migratory Gyps species. Note dark blue, light blue and red bars in jackknife chart (top: Himalayan griffon and bottom: Eurasian griffon) showing importance with only variable, without variable and with all variables, respectively
The response curves between dominant environmental variables and distribution probability of resident Gyps vultures (G. bengalensis, G. indicus, G. tenuirostris) and migratory (G. fulvus, G. himalayensis) drawn by MaxEnt reflected varied responses (Figs. 5 and 6). However, the most dominant climatic covariate bio1 (Annual mean temperature) showed a narrow range (around 24 °C) as the most suitable temperature for INV, SBV, WRV and EGV but HGV favoured a wider range towards lower temperature (1–24 °C). Bio18 (precipitation of warmest quarter), the next dominant variable, presented positive relationship in precipitation increase (500 mm onwards) with occurrence probability in all the Gyps species except G. indicus with inverse relation. Bio19 (precipitation of coldest quarter), yet another dominant covariate, recorded that in the case of resident Gyps this (precipitation, 200 mm onwards) had decreasing but migratory vultures had increasing correlation. Response of precipitation seasonality (bio15) towards species occurrence reflected positive relation in all the Gyps except minor variation of first decrease and then increase in the case of G. himalayensis.

Response curves of environmental parameters (rows 1 to 4; bio1 (annual mean temperature), bio18 (precipitation of warmest quarter), bio19 (precipitation of coldest quarter), bio15 (precipitation seasonality), respectively) in resident Gyps vultures (indian vulture, slender-billed vulture, white-rumped vulture)

Response curves of environmental parameters (rows 1 to 4; bio1 (annual mean temperature), bio18 (precipitation of warmest quarter), bio19 (precipitation of coldest quarter), bio15 (precipitation seasonality), respectively) in migratory Gyps vultures (Eurasian griffon, Himalayan griffon)
Model performance
The distribution density of 10,036 occurrence points in different landcover classes was found to be 0.0112 km−2 (built-up area), 0.0064 km−2 (forest), 0.0054 km−2 (waterbody), 0.0050 km−2 (scrubland), 0.0013 km−2 (wasteland) and 0.0010 km−2 (agriculture). Using unique points for each species, 70 species-based predictions (EGV; HGV; INV; SBV and WRV) and one all vultures (superspecies) prediction for habitat assessment were developed. Species wise details of model evaluators (AUC, TSS, CBI) are presented in Supplementary Table 3. The AUC, TSS and CBI values for models without LULC ranged between 0.781 and 0.976, 0.478 and 0.852, and 0.978 and 0.997, respectively. The CBI charts of different species are presented in Supplementary Fig. 2.
Habitat and floristic regions’ suitability
Habitat suitability (pan-India) and floristic region’s (vegetation based regional units) suitability in terms of area for different Gyps species are presented in Fig. 7 and Supplementary Table 4 and 5, respectively. All Gyps area suitability map is presented as Supplementary Fig. 3. Habitat suitability area of different species and floristic regions, modelled with LULC, are presented in Supplementary Tables 6 and 7, respectively. The projected pan-India suitable area for different species, in the present without LULC, in decreasing order was 50.2% (WRV), 28.4% (INV), 23.2% (EGV), 10.5% (HGV) and 3.9% (SBV) in the total available area of 3.287 million km2. However, some areas were found to be overlapping among different species. When LULC was included as a modelling parameter, the area availability decreased from climatic projection by 12.1% (WRV), 6.5% (EGV), 4.5% (INV), 0.6% (SBV) and 0.1% (HGV), respectively, indicating a greater role of LULC in the former three species.

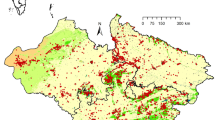
Habitat suitability classes distribution (yellow = unsuitable, blue = moderate, pink = high) of Gyps vultures in different floristic regions of India. Top row: Gyps indicus and Gyps bengalensis. Middle row: Gyps tenuirostris. Bottom row Gyps himalayensis and Gyps fulvus. Distribution of Gyps tenuirostris and Gyps himalayensis may be seen confined to northern floristic regions with limited expanse
As regards the vegetation based regional suitability, suitable habitat for Gyps was found in all the FRs though in varying sizes. The habitat of resident vultures was spread broadly all over the country from the northern-most region, Western Himalaya (33.636°N), to southern-most region, Malabar (09.179°N) while the habitat of wintering vultures was confined from Western Himalaya (35.837°N) to West Indian Plain and Central India (22.631°N). Among residents, Gyps tenuirostris (30.497°N–25.198°N) and, among wintering, Gyps himalayensis (35.348°N–25.565°N) had the narrowest habitat belts.
Since none of the species of vultures were reported from the Andaman and Nicobar and other Islands during the considered study period, we ignored this FR for the purpose of this study. Though all other FRs of India had suitable habitats for Gyps vultures, resident and wintering vultures showed some preferences when seen at the species level. Himalayan griffon remained confined to the northern FRs (Assam, Eastern Himalaya and Western Himalaya) but EGV expanded its presence further up to the central FRs (Assam, Central India, Eastern Himalaya, Gangetic Plain, West Indian Plain and Western Himalaya). The southern peninsular region of India lacked the presence of wintering griffons. Resident Gyps species showed better adaptation as indicated by the presence of suitable habitats in the southern region. Like HGV, SBV was also confined to the northern regions but in a much narrower belt of the Himalayan tarai plain. Unlike SBV, other two resident Gyps had a wider distribution. For example, INV had suitable habitats in all the floristic regions except Assam and Deccan; and WRV was present through all the floristic regions.
On account of species richness, the most suited region is Western Himalaya (all the five species) and least suited is Deccan (two species only, INV and WRV). The floristic regions Assam, Eastern Himalaya and West Indian Plain supported four species while Central India, Gangetic Plain and Malabar harboured three species. However, these species had a suitable area expanse at 47% of 3.287 million km2. Though overlapping among the species, their suitable area was distributed in different floristic regions in decreasing order: Central India (24%), Western Himalaya (15.9%), West Indian Plain (14.4%), Gangetic Plain (10.5%), Deccan (10.1%), Eastern Himalaya (9.6%), Malabar (8.2%) and Assam (7.3%). This indicated that Central India is the most important floristic region for Gyps species.
Impact of climate change on habitat
Predicted future emission scenarios RCP4.5 and RCP8.5 belonging to (i) the short term, i.e. year 2050, and (ii) the long term, i.e. 2070 (Figs. 8, 9 and 10, 11 and Supplementary Table 4), when compared with the present climatic model, showed changes in both unsuitable and suitable areas. Out of four, a minimum of three scenarios showed a decreasing trend in area suitability in EGV, HGV, INV and WRV. In the case of SBV, all four scenarios showed a decrease from the present prediction.

Short term projected habitat suitability (yellow = unsuitable, blue = moderate, pink = high) of resident Gyps vultures under moderate (RCP 4.5) and extreme (RCP 8.5) scenarios. Top row: Gyps indicus. Middle row: Gyps tenuirostris. Bottom row: Gyps bengalensis

Short-term projected habitat suitability (yellow = unsuitable, blue = moderate, pink = high) of wintering Gyps vultures under moderate (RCP 4.5) and extreme (RCP 8.5) scenarios. Top row: Gyps fulvus. Bottom row: Gyps himalayensis

Long-term projected habitat suitability (yellow = unsuitable, blue = moderate, pink = high) of resident Gyps vultures under moderate (RCP 4.5) and extreme (RCP 8.5) scenarios. Top row: Gyps indicus. Middle row: Gyps tenuirostris. Bottom row: Gyps bengalensis

Long-term projected habitat suitability (yellow = unsuitable, blue = moderate, pink = high) of wintering Gyps vultures under moderate (RCP 4.5) and extreme (RCP 8.5) scenarios. Top row: Gyps fulvus. Bottom row: Gyps himalayensis
The “stable” area (suitable as well as unsuitable) along with “loss” of suitable area and “gain” from unsuitable area in future scenarios is presented in Supplementary Table 8. Both suitable and unsuitable area of the present prediction recorded a change in their status since their parts converted to unsuitable and suitable category, respectively, in both short and long terms. Eurasian griffon, SBV and INV showed a reduction in suitable area for both the RCPs and terms, while WRV showed a decreasing trend (except 2050 RCP4.5), and HGV showed an increasing trend (except 2050 RCP4.5). The amount of change was meagre between 0.1 and 2% but still sizable in expanse (3287 to 65,745 km2). The conversion dynamics are depicted species-wise and residency class-wise in Supplementary Figs. 4 and 5 migratory/wintering, and Supplementary Figs. 6 to 8 resident Gyps vultures.
The change in future habitat, especially the loss of suitable habitat, was observed to be the lowest in the Himalayan region (Western Himalaya and Eastern Himalaya) and the Nilgiri mountains (Malabar and Deccan) for any vulture. Most of the changes or habitat conversion were seen in the plains and at the fringes of suitable or unsuitable areas for all the species found therein. The West Indian Plain, Central India and Gangetic Plain FRs were found more prone to loss.
Discussion
This study has produced habitat suitability maps derived from statistical models for five Gyps species. Naoroji (2006), referring to the limitation of existing range maps, has recorded that exact distribution of a species depends on availability of suitable habitat. Bustamante and Seoane (2004) have further suggested that SDM generated maps are improvement of traditional range maps based on broad survey of species presence. This study also provides the first account of a national baseline of species-wise suitable projected habitat of Gyps vultures and possible changes in the future in different floristic regions using field surveys in > 17% of total area considered, citizen science and published occurrence records, bioclimatic variables and SDM. The resulting analysis has shown a concern for suitable area showing a decreasing trend in the future for critically endangered resident species. Though citizen science data is considered unstructured, integration of expert survey data with the filtered citizen science data has been in vogue and is known to result in improved inference, predictive ability and ultimately with increased extent of inference of the structured surveys or expert data (Robinson et al. 2020).
Our study is based on MaxEnt algorithm which uses presence-only data. Using presence-only data to calibrate distribution models has some known drawbacks which may limit model performance (Brotons et al. 2004; Alatawi et al. 2020). Importantly, presence-only methods probably over-estimate species occurrence, because locations predicted to be suitable may not in fact be occupied, as a result of limited species dispersal. As a result, using presence-absence data is strongly recommended whenever available (Brotons et al. 2004). However, presence-only records often the only available information about species occurrences, and these are still informative about the true underlying distribution (Zaniewski et al. 2002). Despite using presence-only data, Maxent has been shown to perform well, generating predictive models even with biased data and small sample sizes (Hernandez et al. 2006; Pearson et al. 2007; Wisz et al. 2008). Kaky et al. (2020) have also reviewed that though MaxEnt models were criticised by some researchers earlier, it continued to be frequently used to fit models across many different taxa, geographical areas, time periods and environmental scenarios (Achour and Kalboussi 2020; Anoop et al. 2020; Anand et al. 2021; Cable et al. 2021; Dobrev and Popgeorgiev 2021; Gao et al. 2021; Gao and Shi 2021; Grimshaw et al. 2021; Jha and Jha 2021a,b,c; Mushtaq et al. 2021; Oliveira et al. 2021; Panthi et al. 2021, among others). MaxEnt has numerous advantages like (1) it can work with a small sample size especially rare and threatened species (Hernandez et al. 2006; Wisz et al. 2008; Kumar and Stohlgren 2009; Abolmaali et al. 2018), (2) it is easy to use and very useful when presence-absence data collection is impractical (Phillips et al. 2006; Kumar and Stohlgren 2009; Angelieri et al. 2016), (3) both categorical and continuous environmental layers can be applied in this software and (4) it measures importance of each environmental variable using the jackknife test, in terms of gain (Elith et al. 2011; Groff et al. 2014).
Despite the universal use of MaxEnt software, Cobos et al. (2019) and de Andrade et al. (2020) have recently suggested use of R packages which allows robust processes of modelling and straightforward construction of complex ecological niche models. Therefore, it is advisable that R packages, such as “kuenm,” “ENMTML,” “maxnet,” and “dismo,” may be used for prediction improvement in future modelling. It is further proposed that future models must contain modelled future LULC for enhanced accuracy in prediction, since climatic prediction may be an overestimation without LULC (Jha and Jha 2021b). Preston et al. (2008) also stated that distribution models predicting species responses to climate change included mostly climate variables and rarely the biotic interactions.
Habitat determinants
Bioclimatic habitat determinants used in this study sourced from WorldClim follow the dynamic approach. Bede-Fazekas and Somodi (2020) recently discovered potential traps in the use of the widely applied dynamic approach but simultaneously stated that this approach cannot be ignored also. The models presented here were with good to excellent prediction power due to uncertainty removal (De Marco and Nobrega 2018). MaxEnt variable contribution table showed that major determinants of suitable habitat for Gyps species in the study area are land use/land cover (LULC), annual mean temperature (bio1) and temporal variants of precipitation, e.g. bio19 (precipitation of coldest quarter), bio15 (precipitation seasonality) and bio18 (precipitation of warmest quarter). This finding is in general agreement with Herrero et al. (2006) which states that the influence of vegetation cover on the distribution of animal species, in providing food and shelter, also acts as a limiting factor to the spread of species. Freeman et al. (2019) also had a similar observation suggesting that forest cover is a more vital driver as compared to climate for the current distribution of the target species.
However, climate variables, particularly rainfall and temperature, generally influence habitat quantity and quality affecting the structure, composition and dynamics of wildlife species (Kupika et al. 2018). Jha and Jha (2020) also reported LULC as the most prominent determinant of the distribution of different vulture species, followed by isothermality, and precipitation seasonality in Central India. The findings of this study broadly concur with this but differ in order of preference of these determinants in different species when considered on a much larger scale at country level. More so, several studies suggested the influence of bioclimatic variables in habitat determination in raptors (Gschweng et al. 2012; Liminana et al. 2012) and vultures (Zhang et al. 2019; Anoop et al. 2020) although the set of covariates were not similar in these different and distant localities.
Moreover, the lowest spatiotemporal occupancy in agricultural landscape (0.0010 km−2) and highest in built-up area (0.0112 km−2) followed by forest (0.0064 km−2), water (0.0054 km−2), scrubland (0.0050 km−2) and wasteland (0.0013 km−2) could be speculated to play a role in the prediction of habitat suitability corresponding to their importance. As suggested by response bars, built-up area, land covered by buildings and other manmade structures (Buchhorn et al. 2020) including road, railway, paved land and urban park (Venter et al. 2016) favours the Gyps species in providing foraging materials, for example, accidental carcasses, direct disposal of dead animals, feed through slaughterhouses and bone mills etc. Forest is the next favourable area for nesting as well as foraging while agricultural landscape is the least suitable for lack of nesting sites.
Response curves indicated varied impact of the covariates on vulture presence. Habitat suitability is a product of interaction among numerous covariates in different quantities (grades), not a function of any single variable (Richard et al. 2018; Jha and Jha 2021b). Quite a few among these could be following Liebig’s Law of the Minimum (a covariate behaving as limiting factor). Hence, considering a single variable in isolation may be misleading as the species choose their habitat based on the interaction of several factors (Golterman 1975; Jha and Jha 2021b). However, a thumb rule can be drawn from response curves that increase in mean annual temperature beyond 24 °C produces stress conditions. Another stressing factor is precipitation in coldest quarter (bio19) beyond 200 mm for resident Gyps, though migratory ones have no such impact. However, precipitation in warmest quarter (bio18) beyond 500 mm enhances the probability of occurrence for all the Gyps species except G. indicus. Further generalisation reveals that precipitation seasonality (bio15) will be useful for residency but mean annual temperature beyond 30 °C may become intolerable.
Model robustness
Despite the weakness of AUC as an inadequate model evaluator (Lobo et al. 2008; Fourcade et al. 2014; Jiménez-Valverde 2014), model performance is commonly evaluated by AUC values of the Receiver Operating Characteristic (DeLong et al. 1988), especially MaxEnt (Gao and Shi 2021). For example, it was preferred in recent studies on animals (Achour and Kalboussi 2020; Mori et al. 2020), birds (Tehrani et al. 2021), raptors (Regos et al. 2021) and vultures (Khwarahm et al. 2021), since AUC is one of those statistics which provides a good information to judge the model performance where only presence data are used (Proosdij et al. 2016; Anand et al. 2021). Although AUC is widely applied (Abolmali et al. 2018; Abdelaal et al. 2019; de Luis et al. 2019; Achour and Kalboussi 2020; Mori et al. 2020; Anand et al. 2021; Jha and Jha 2021b), many agree that it tends to be overoptimistic (Lobo et al. 2008; Shabani et al. 2016), and hence, it is often complemented by another measure of model goodness. Therefore, CBI and TSS may be used as additional and better assessment as suggested by Allouche et al. (2006), Breiner et al. (2015), Manzoor et al. (2018) and Shabani et al. (2018). The reason for this could be as suggested by Lobo et al. (2008, 2010) that AUC is influenced by species prevalence but TSS has been widely advocated as a suitable discrimination metric less dependent on prevalence (Allouche et al. 2006; Somodi et al. 2017). Additionally, Sun et al. (2021) suggested that AUC is suitable for evaluating models built based on presence-absence data, and the CBI (Hirzel et al. 2006) evaluates presence-only models such as MaxEnt used in this study.
Model performance could be assessed by their categories as suggested in different studies. For example, AUC closest to a value of 1 would be a perfect model and AUC = 0.5 would indicate that the model performed no better than random (Barragan -Barrera et al. 2019). However, models are excellent with AUC > 0.9, good with AUC between 0.8 and 0.9, fair with AUC 0.7–0.8 and uninformative with AUC < 0.7 (Swets 1988; Araújo et al. 2005). TSS values are categorised as excellent with > 0.8, good between 0.6 and 0.8, fair with 0.4 and 0.6, poor or no predictive ability with < 0.4 (Rew et al. 2020). The Boyce index varies between − 1 and + 1. Positive values indicate a model in which predictions are consistent with the distribution of presences in the evaluation data set, values close to 0 mean that the model is not different from a random model, and negative values indicate counter predictions (Hirzel et al. 2006; Di Cola et al. 2017). All our predictions (AUC between 0.781 and 0.976, TSS between 0.478 and 0.852 and CBI between 0.978 and 0.997) are very useful and considered suitable for conservation planning (Pearce and Ferrier 2000).
The models for the present, both with and without LULC, predicted results which varied in the expanse of habitat. The former had a lesser spread of suitable area than the latter which could be attributed to the fact that the environmental umbrella is mostly smaller than the climatic umbrella due to the specific requirements of a niche, e.g. trees/cliffs, water and ungulate/cattle concentration (Jha and Jha 2021a,b). This implied that the actual suitable habitat area was an overestimation in the case of bioclimatic models. This was similar across all the future models which are bioclimatic in nature also supported by Preston et al. (2008).
Habitat availability
Species distribution modelling was used to predict the habitat suitability, shaped by environmental factors, which was then reclassified in two classes. The first was the class comprising moderately and highly suitable areas and the second comprising unsuitable area for different species of Gyps in India, for each floristic region. It is apparent that HGV and SBV have restricted suitable habitats in the north which also overlaps other species (WRV, EGV). The habitat of INV is distributed mainly in the central and south-western part, mostly overlapping with WRV everywhere and EGV in northern part. This indicates lower availability of projected suitable area kilometrage per species due to inter species competition in particular regions for shelter, territory and available food. Possible reason for the overlap was assigned to availability of ample source of carrion and relatively low availability of nest sites (Ferguson-Lees and Christie 2001). For example, large vultures like Lammergeier and Himalayan griffon are reported to coexist in closer proximity along with Saker Falcon (Katzner et al. 2004). However, overlapping areas due to sympatric nature of the species should be highly valued and protected (Zhang et al. 2019) to maximise multispecies conservation. Ground verification of selected sites was done in Western Himalaya tarai, Gangetic Plain, Central India and West Indian Plain (Madhya Pradesh, Uttar Pradesh and Rajasthan). It was observed that WRV and SBV preferred trees while INV chose cliff nesting surrounded by forests. This agreed with many studies (Majgaonkar et al. 2018; Jha et al. 2021) in different provinces but with a couple of exceptions (Khatri 2015; Navaneethan et al. 2015). Vultures showed an affinity to localities closer to water sources, mostly rivers or large waterbodies. Anoop et al. (2020) also recorded the use of riparian forested area in the Western Ghats (Malabar region) being used by WRV for nesting. Kumar et al. (2014) and Misher et al. (2017) had similar observations of cliff nesting in INV along a river in Central India. Nests were mostly found in areas free from disturbance but cases of nesting near human settlements was recorded in the case of WRV. Such plasticity was also reported elsewhere (Bahadur et al. 2019).
Impact of climate change
Climate is considered a primary factor in constraining the distribution of plant species (Banag et al. 2015). This could be a possible reason for the change in habitat area in the future in this study, since other anthropogenic factors have not been considered in modelling. The impact is evident from the change in habitat expanse in the short- and long-term future. As a result of loss of suitable area and gain from unsuitable area, a trend of net loss is observed in different scenarios of emission and term tenures. Different observations of a decrease in suitable habitat of wildlife species in general (McDonald et al. 2019) and vultures specifically (Ilanloo et al. 2020) support our finding. However, such an expansion and contraction of habitat area may have a bearing on the Gyps species population as habitat is an important place for the survival, reproduction and population development of any organism. Any changes in the quantity, quality and distribution of habitat have a wide range of effects on spatial dynamics and can directly affect the distribution, quantity and survival rate of organisms (Zhang et al. 2019). Nevertheless, such areas require advance planning to mitigate the loss and exploit the gains. Another point of interest of conservation should be the highland areas where there was the lowest change in habitat due to climate change as also observed by Banda and Tassie (2018) in endemic bird species.
However, the highest number of floristic region (FR) as well as the largest area coverage in decreasing order, i.e. WRV (1,253,037 km2, 8 FR) > INV (786,121 km2, 6 FR) > EGV (548,181 km2, 4 FR) > HGV (343,094 km2, 3 FR) > SBV (108,091 km2, 3 FR), indicated better adaptability and lower vulnerability to varied bioenvironmental conditions of the former ones than the latter ones. Therefore, priority must be given to conservation of species with lower adaptability or higher vulnerability against climate change.
Management implications
Predictive models are a useful tool for wildlife managers to make better decisions about biodiversity management and conservation (Rodriguez et al. 2007). This study provided potential habitat areas for different Gyps species based on predictive modelling under different scenarios. All the models are robust enough to be replicated but a point of concern could be the future predictions which do not consider land use while it is evident that there is a constant change in forest, agricultural, built-up and other landcover. Nevertheless, due to a timely intervention in the management of forests, a positive shift, i.e. increase in (very dense forest and) open forest but decrease in moderately dense forest (ISFR 2021), was seen. Therefore, it is assumed that there may not be a negative change in vulture habitat or at least it may remain unchanged on this account.
However, the prediction of suitable habitat for Gyps vultures combined and species-wise independently differed in the floristic regions. Highly suitable area in the all-species combined prediction was reduced when compared with species individually; for example, SBV in Assam, INV in Central India, WRV in Central India and West Indian Plain, and EGV in West Indian Plain had much larger expanses of suitable area. This is due to the fact that the algorithm of the SDM works on specific requirement of a species. In the case of all Gyps vultures, common minimum suitability criteria were considered and a projection was made. Such a prediction indicated that species-wise management planning should be a better option than combined conservation efforts wherever feasible. Nevertheless, in Indian context, all species information is equally important since the Forest Departments look after overall vulture conservation.
Keeping the above in view, the predictions from this study could be used for conservation planning in the study area. As regards the planning structure, Indian governance is federal in nature where the responsibility of making a standard policy for conservation, if needed, rests with the Centre while states are responsible for implementing this strategy at a local level. In such a scenario, a large research study area becomes useful for the formulation of a common management strategy by the central government for the states. The following measures could be considered based on the above findings.
This study provided the expanse of suitable area in different floristic regions for different species. However, it is important to focus on land use in the area for any conservation programme. For example, in the case of reintroduction of a species, availability of nesting, roosting, and foraging area all become important. The agricultural land falling in suitable areas may provide only foraging opportunity, while forests, tall trees, or cliffs, depending on species, would be a must for nesting and roosting requirement.
Vulture centric development of existing suitable areas must be carried out. The concept of Vulture Protection Area (overlapping areas of high suitability for multiple species) and Vulture Conservation Area (species wise high suitability area) must be introduced in order to secure fruitful conservation. This must include roosting, nesting and foraging area for vultures/species in the core zone and foraging area in the periphery for buffer function. Going by the area prediction, and availability of sufficient nesting structures, feasibility of Vulture Conservation Areas may be explored in different floristic zones, for example, SBV Protection Area in the eastern part of Eastern Himalayas, INV-WRV Conservation Area in the eastern and western part of Central India. The area expanse of such reserves could be as large as possible after taking into consideration threats and opportunities, since vultures are highly mobile organisms and are capable of long-distance foraging trips, up to 100 km (Moleón et al. 2020). Within these reserves and outside, if feasible, suggested activities could be (i) in situ conservation of vultures in highly suitable areas; (ii) habitat maintenance for expansion of territory in moderately suitable areas; (iii) planning in advance for habitat improvement in unstable areas since changing climate can cause changes in the geographic distribution of the amount and quality of habitat (Holyoak and Heath 2016); (iv) expansion of favourable area for vultures which have shown habit plasticity (Genero et al. 2020), an agroforestry model could also be adopted by including trees of moderate height for shelter and rearing of medium sized vertebrates for food (Hiraldo et al. 1991; Chhangani 2007) in the areas where only foraging is possible; and (v) above all there is a need to protect the landscape (moderate and highly suitable area) against human-induced large-scale habitat change like, deforestation for development, in order to slow down species extinction (Kentie et al. 2018).
Conclusion
Though the models generated in this study based on bioclimatic variables could be further improved by incorporating bioenvironmental variable like dynamic LULC layers for future prediction, they are strong enough and should be used as a starting point for immediate management planning of Gyps vulture conservation in various floristic regions.
Our study of Gyps vultures’ habitat suitability impacted by impending climate change identified habitat variables and provided delimitation of stable and unstable habitats of suitable and unsuitable nature in Indian context for the first time. This has direct implication on management of imperilled vultures since stable and suitable area could be used for in situ conservation and reintroduction of the species. Unstable area could be used for habitat improvement by ensuring nesting and foraging resources for further use by vultures in expanding their territory for increasing population. In general, development of reserves, protection of large trees, adoption of agroforestry etc. could be useful in model predicted areas to attempt a reversal of the endangered status, to some extent, of indigenous vultures in different floristic regions. The study also indicated better adaptability and lower vulnerability to varied bioenvironmental conditions of the different Gyps species. Therefore, priority must be given to conservation of species with lower adaptability or higher vulnerability against climate change.
Data availability
This manuscript has data included as electronic supplementary material.
References
Abdelaal M, Fois M, Fenu G, Bachhetta G (2019) Using MaxEnt modeling to predict the potential distribution of the endemic plant Rosa arabica Crép. in Egypt. Ecol Inf 50:68–75. https://doi.org/10.1016/j.ecoinf.2019.01.003
Abolmaali MRS, Tarkesh M, Bashari H (2018) MaxEnt modeling for predicting suitable habitats and identifying the effects of climate change on a threatened species, Daphne mucronata, in central Iran. Ecol Inf 43:116–123. https://doi.org/10.1016/j.ecoinf.2017.10.002
Achour H, Kalboussi M (2020) Modelling and mapping the current and future potential habitats of the Algero-Tunisian endemic newt Pleurodeles nebulosus under climate change. Eur J Wildl Res 66:61. https://doi.org/10.1007/s10344-020-01386-x
Ahmad R, Khuroo AA, Hamid M, Charles B, Rashid I (2019) Predicting invasion potential and niche dynamics of Parthenium hysterophorus (congress grass) in India under projected climate change. Biodiv Conserv 28:2319–2344. https://doi.org/10.1007/s10531-019-01775-y
Alatawi AS, Gilbert F, Reader T (2020) Modelling terrestrial reptile species richness, distributions and habitat suitability in Saudi Arabia. J Arid Environ 178:104153. https://doi.org/10.1016/j.jaridenv.2020.104153
Allouche O, Tsoar A, Kadmon R (2006) Assessing the accuracy of species distribution models: prevalence, kappa and the true skill statistic (TSS). J Appl Ecol 43:1223–1232. https://doi.org/10.1111/j.1365-2664.2006.01214.x
Anand V, Oinam B, Singh IH (2021) Predicting the current and future potential spatial distribution of endangered Rucervus eldii eldii (Sangai) using MaxEnt model. Environ Monit Assess 193:147. https://doi.org/10.1007/s10661-021-08950-1
Angelieri CCS, Adams-Hosking C, de Ferraz KMPMB, de Souza MP, McAlpine CA (2016) Using species distribution models to predict potential landscape restoration effects on puma conservation. PLoS One 11:e0145232. https://doi.org/10.1371/journal.pone.0145232
Anoop NR, Babu S, Nagarajan R, Sen S (2020) Identifying suitable reintroduction sites for the white-rumped vulture (Gyps bengalensis) in India’s Western Ghats using niche models and habitat requirements. Ecol Eng 158:106034. https://doi.org/10.1016/j.ecoleng.2020.106034
Araújo MB, Pearson RG, Thuiller W, Erhard M (2005) Validation of species-climate impact models under climate change. Global Chang Biol 11:1504–1513. https://doi.org/10.1111/j.1365-2486.2005.01000.x
Bahadur KCK, Koju NP, Bhusal KP, Low M, Ghimire SK, Ranabhat R, Panthi S (2019) Factors influencing the presence of the endangered Egyptian vulture Neophron percnopterus in Rukum, Nepal. Global Ecol Conserv 20:e00727. https://doi.org/10.1016/j.gecco.2019.e00727
Banag C, Thrippleton T, Alejandro GJ, Reineking B, Liede-Schumann S (2015) Bioclimatic niches of selected endemic Ixora species on the Philippines: predicting habitat suitability due to climate change. Plant Ecol 216:1325–1340. https://doi.org/10.1007/s11258-015-0512-6
Banda LB, Tassie N (2018) Modeling the distribution of four-bird species under climate change in Ethiopia. Ethiopian J Biol Sci 17:1–17
Baral N, Nagy C, Crain BJ, Gautam R (2013) Population viability analysis of critically endangered white-rumped vultures Gyps bengalensis. Endang Species Res 21:65–76. https://doi.org/10.3354/esr00511
Barbosa AM, Real R, Munoz AR, Brown JA (2013) New measures for assessing model equilibrium and prediction mismatch in species distribution models. Divers Distrib 19:1333–1338. https://doi.org/10.1111/ddi.12100
Barragan-Barrera DC, do Amaral KB, Chávez-Carreño PA et al (2019) Ecological niche modeling of three species of Stenella dolphins in the Caribbean Basin, with application to the Seaflower Biosphere Reserve. Front Mar Sci 6:10. https://doi.org/10.3389/fmars.2019.00010
Bede-Fazekas Á, Somodi I (2020) The way bioclimatic variables are calculated has impact on potential distribution models. Methods Ecol Evol 11:1559–1570. https://doi.org/10.1111/2041-210X.13488
Bhattacherjee A (2012) Social science research: principles, methods, and practices. Textbooks Collection. 3. http://scholarcommons.usf.edu/oa_textbooks/3. Accessed 24 February 2023
Boria RA, Olson LE, Goodman SM, Anderson RP (2014) Spatial filtering to reduce sampling bias can improve the performance of ecological niche models. Ecol Model 275:73–77. https://doi.org/10.1016/j.ecolmodel.2013.12.012
Botha AJ, Andevski J, Bowden CGR, Gudka M, Safford RJ, Tavares J, Williams NP (2017) Multi-species action plan to conserve African-Eurasian vultures. CMS Raptors MOU Technical Publication No. 5. CMS Technical Series No. 3. Coordinating Unit of the CMS Raptors MOU, United Arab Emirates
Breiner FT, Guisan A, Bergamini A, Nobis MP (2015) Overcoming limitation of modeling rare species by using ensembles of small models. Methods Ecol Evol 6:1210–1218. https://doi.org/10.1111/2041-210X.12403
Brotons L, Thuiller W, Araujo MB, Hirzel AH (2004) Presence-absence versus presence-only modelling methods for predicting bird habitat suitability. Ecography 27:437–448. https://doi.org/10.1111/j.0906-7590.2004.03764.x
Brown JL, Bennett JR, French CM (2017) SDMtoolbox 2.0: the next generation Python-based GIS toolkit for landscape genetic, bio-geographic and species distribution model analyses. PeerJ 5:e4095. https://doi.org/10.7717/peerj.4095
Buchhorn M, Smets B, Bertels L, De Roo B, Lesiv M, Tsendbazar N-E, Herold M, Fritz S (2020) Copernicus Global Land Service: land cover 100m: collection 3: epoch 2019: Globe. 10.5281/zenodo.3939050. Accessed 24 February 2023
Bustamante J, Seoane J (2004) Predicting the distribution of four species of raptors (Aves: Accipitridae) in southern Spain: statistical models work better than existing maps. J Biogeogr 31:295–306. https://doi.org/10.1046/j.0305-0270.2003.01006.x
Cable AB, O’Keefe JM, Deppe JL, Hohoff TC, Taylor SJ, Davis MA (2021) Habitat suitability and connectivity modelling reveal priority areas for Indiana bat (Myotis sodalis) conservation in a complex habitat mosaic. Landsc Ecol 36:119–137. https://doi.org/10.1007/s10980-020-01125-2
Campbell MO (2016) Vultures, their evolution, ecology and conservation. CRC Press, Taylor & Francis Group, Boca Raton, London, New York
Chhangani AK (2007) Sightings and nesting sites of red-headed vulture Sarcogyps calvus in Rajasthan, India. Indian Birds 3:218–221
Chhangani AK, Mohnot SM (2004) Is diclofenac the only cause of vulture decline? Curr Sci 87:1496–1497
Cobos ME, Peterson AT, Barve N, Osorio-Olvera L (2019) kuenm: an R package for detailed development of ecological niche models using Maxent. PeerJ 7:e6281. https://doi.org/10.7717/peerj.6281
de Andrade AFA, Velazco SJE, De Marco Júnior P (2020) ENMTML: an R package for a straightforward construction of complex ecological niche models. Environ Model Softw 125:104615. https://doi.org/10.1016/j.envsoft.2019.104615
de Frutos A, Olea PP, Vera R (2007) Analyzing and modelling spatial distribution of summering lesser kestrel: the role of spatial autocorrelation. Ecol Model 200:33–44. https://doi.org/10.1016/j.ecolmodel.2006.07.007
de Luis M, Alvarez-Jiminez J, Martınez-Labarga JM, Bartolome C (2019) Four climate change scenarios for Gypsophila bermejoi G Lopez (Caryophyllaceae) to address whether bioclimatic and soil suitability will overlap in the future. PLoS One 14:e0218160. https://doi.org/10.1371/journal.pone.0218160
De Marco P, Nobrega CC (2018) Evaluating collinearity effects on species distribution models: an approach based on virtual species simulation. PLoS One 13:e0202403. https://doi.org/10.1371/journal.pone.0202403
DeLong ER, DeLong DM, Clarke-Pearson DL (1988) Comparing the areas under two or more correlated receiver operating characteristic curves: a nonparametric approach. Biometrics 44:837–845. https://doi.org/10.2307/2531595
Di Cola V, Broennimann O, Petitpierre B, Breiner FT, D’Amen M, Randin C, Engler R, Pottier J, Pio D, Dubuis A, Pellissier L, Mateo RG, Hordijk W, Salamin N, Guisan A (2017) Ecospat: an R package to support spatial analyses and modelling of species niches and distributions. Ecography 40:774–787. https://doi.org/10.1111/ecog.02671
Dobrev D, Popgeorgiev G (2021) Modeling breeding habitat suitability of griffon vultures (Gyps fulvus) in Bulgaria and conservation planning. Nwest J Zool 17:281–287
Dormann CF, Calabrese JM, Guillera-Arroita G et al (2018) Model averaging in ecology: a review of Bayesian, information-theoretic, and tactical approaches for predictive inference. Ecol Monogr 88:485–504. https://doi.org/10.1002/ecm.1309
Elith J, Graham CH (2009) Do they? How do they? Why do they differ? On finding reasons for differing performances of species distribution models. Ecography 32:66–77. https://doi.org/10.1111/j.1600-0587.2008.05505.x
Elith J, Graham CH, Anderson RP et al (2006) Novel methods improve prediction of species’ distributions from occurrence data. Ecography 29:129–151. https://doi.org/10.1111/j.2006.0906-7590.04596
Elith J, Phillips SJ, Hastie T, Dudik M, Ye C, Yates CJ (2011) A statistical explanation of MaxEnt for ecologists. Divers Distrib 1:43–57. https://doi.org/10.1111/j.1472-4642.2010.00725.x
Ferguson-Lees J, Christie DA (2001) Raptors of the world. Houghton Mifflin, Boston
Fick SE, Hijmans RJ (2017) WorldClim 2: new 1km spatial resolution climate surfaces for global land areas. Int J Climatol 37:4302–4315. https://doi.org/10.1002/joc.5086
Fourcade Y, Engler J, Rödder D, Secondi J (2014) Mapping species distributions with MAXENT using a geographically biased sample of presence data: a performance assessment of methods for correcting sampling bias. PLoS One 9:e97122. https://doi.org/10.1371/journal.pone.0097122
Freeman B, Jimenez-Garcia D, Barca B, Grainger M (2019) Using remotely sensed and climate data to predict the current and potential future geographic distribution of a bird at multiple scales: the case of Agelastes meleagrides, a western African forest endemic. Avian Res 10:22. https://doi.org/10.1186/s40657-019-0160-y
Gadhvi IR, Dodia PP (2006) Indian white-backed vultures Gyps bengalensis nesting in Mahuva, Bhavnagar district, Gujarat, India. Indian Birds 2:36
Galligan TH, Bhusal KP, Paudel K, Chapagain D, Joshi AB, Chaudhary IP, Chaudhary A, Baral HS, Cuthbert RJ, Green RE (2020) Partial recovery of critically endangered Gyps vulture populations in Nepal. Bird Conserv Int 30:87–102. https://doi.org/10.1017/S0959270919000169
Galligan TH, Mallord JW, Prakash VM et al (2021) Trends in the availability of the vulture-toxic drug, diclofenac, and other NSAIDs in South Asia, as revealed by covert pharmacy surveys. Bird Conserv Int 31:337–353. https://doi.org/10.1017/S0959270920000477
Gao T, Shi J (2021) The potential global distribution of Sirex juvencus (Hymenoptera: Siricidae) under near current and future climatic conditions as predicted by the Maximum Entropy model. Insects 12:222. https://doi.org/10.3390/insects1203022
Gao T, Xu Q, Liu Y, Zhao J, Shi J (2021) Predicting the potential geographic distribution of Sirex nitobei in China under climate change using Maximum Entropy model. Forests 12:151. https://doi.org/10.3390/f12020151
Genero F, Franchini M, Fanin Y, Filacorda S (2020) Spatial ecology of non-breeding Eurasian griffon vultures Gyps fulvus in relation to natural and artificial food availability. Bird Study 67:53–70. https://doi.org/10.1080/00063657.2020.1734534
Golterman HL (1975) Physiological limnology: an approach to the physiology of lake ecosystem. Developments in Water Science Series. Elsevier Publishing company. https://doi.org/10.1016/s0167-5648(08)71058-X
Green RE, Newton I, Schultz S, Cunningham AA, Gilbert M, Pain DJ, Prakash V (2004) Diclofenac poisoning as a cause of vulture population declines across the Indian subcontinent. J Appl Ecol 41:793–800. https://doi.org/10.1111/J.0021-8901.2004.00954.X
Grimshaw JR, Ray DA, Stevens RD (2021) Ecological niche models for bat species of greatest conservation need in Louisiana. Museum of Texas Tech University Number 378, 19 October 2021 https://www.depts.ttu.edu/nsrl/publications/downloads/OP378.pdf. Accessed 24 Feb 2023
Groff LA, Marks SB, Hayes MP (2014) Using ecological niche models to direct rare amphibian surveys: a case study using the Oregon spotted frog (Rana pretiosa). Herpetol Conserv Biol 9:354–368
Gschweng M, Kalko EKV, Berthold P, Fiedler W, Fahr J (2012) Multi-temporal distribution modelling with satellite tracking data: predicting responses of a long distance migrant to changing environmental conditions. J Appl Ecol 49:803–813. https://doi.org/10.1111/j.1365-2664.2012.02170.x
Heikkinen RK, Luoto M (2006) Methods and uncertainties in bioclimatic envelope modelling under climate change. Prog Phys Geogr 6:751–777. https://doi.org/10.1177/0309133306071957
Hernandez PA, Graham CH, Master LL, Albert DL (2006) The effect of sample size and species characteristics on performance of different species distribution modelling methods. Ecography 29:773–785. https://doi.org/10.1111/j.0906-7590.2006.04700.x
Herrero J, García-Serrano A, Couto S, Ortuño V, García-González R (2006) Diet of wild boar Sus scrofa L. and crop damage in an intensive agroecosystem. Eur J Wildl Res 52:245–250. https://doi.org/10.1007/s10344-006-0045-3
Hiraldo F, Blanco JC, Bustamante J (1991) Unspecialised exploitation of small carcasses by birds. Bird Study 38:200–207. https://doi.org/10.1080/00063659109477089
Hirzel AH, Le Laya G, Helfera V, Randina C, Guisan A (2006) Evaluating the ability of habitat suitability models to predict species presences. Ecol Model 199:142–152. https://doi.org/10.1016/j.ecolmodel.2006.05.017
Holyoak M, Heath SK (2016) The integration of climate change, spatial dynamics, and habitat fragmentation: a conceptual overview. Integr Zool 11:40–59. https://doi.org/10.1111/1749-4877.12167
Hurtt GC, Chini LP, Frolking S et al (2011) Harmonization of land-use scenarios for the period 1500–2100: 600 years of global gridded annual land-use transitions, wood harvest, and resulting secondary lands. Clim Chang 109:117–161. https://doi.org/10.1007/s10584-011-0153-2
Ilanloo SS, Khani A, Kafash A, Valizadegan N, Ashrafi S, Loercher F, Ebrahimi E, Yousefi M (2020) Applying opportunistic observations to model current and future suitability of the Kopet Dagh Mountains for a Near Threatened avian scavenger. Avian Biol Res 14:18–26. https://doi.org/10.1177/1758155920962750
iNaturalist users and Ueda K (2020) iNaturalist Research-grade Observations. iNaturalist.org. Occurrence dataset. https://doi.org/10.15468/ab3s5x. Accessed 23 Oct 2020
ISFR (2021) India State of Forest Report. Forest Survey of India (MoEFCC), Dehradun, India
IUCN (2022) The IUCN red list of threatened species. https://www.iucnredlist.org/ Accessed 03 Aug 2022
Jha KK (2018) Mapping and management of vultures in an Indian stronghold. In: Campbell MO (ed) Geomatics and conservation biology. Nova Science Publishers, New York, pp 45–75
Jha KK, Jha R (2020) Habitat suitability mapping for migratory and resident vultures: a case of Indian stronghold and species distribution model. J Wildl Biodiv 4:91–111. https://doi.org/10.22120/jwb.2020.120246.1111
Jha KK, Jha R (2021a) Study of vulture habitat suitability and impact of climate change in Central India using MaxEnt. J Resour Ecol 12:30–42. https://doi.org/10.5814/j.issn.1674-764x.2021.01.004
Jha R, Jha KK (2021b) Habitat prediction modelling for vulture conservation in Gangetic-Thar-Deccan region of India. Environ Monit Assess 193:532. https://doi.org/10.1007/s10661-021-09323-4
Jha R, Jha KK (2021c) What could be the present and future habitats of resident and wintering vultures for conservation in Indian subcontinent. In: Campbell MO, Jha KK (eds) Critical research techniques in habitat and animal ecology: examples from India. Nova Science Publishers, New York pp, pp 287–327
Jha KK, Campbell MO, Jha R (2020) Vultures, their population status and some ecological aspects in an Indian stronghold. Not Sci Biol 12:124–142. https://doi.org/10.15835/nsb12110547
Jha KK, Jha R, Campbell MO (2021) The distribution, nesting habits and status of threatened vulture species in protected areas of Central India. Ecol Questions 32:7–22
Jiménez-Valverde A (2014) Threshold-dependence as a desirable attribute for discrimination assessment: implications for the evaluation of species distribution models. Biodivers Conserv 23:369–385. https://doi.org/10.1007/s10531-013-0606-1
Kaky E, Nolan V, Alatawi A, Gilbert F (2020) A comparison between Ensemble and MaxEnt species distribution modelling approaches for conservation: a case study with Egyptian medicinal plants. Ecol Inf 60:101150. https://doi.org/10.1016/j.ecoinf.2020.101150
Katzner TE, Lai CH, Gardiner JD, Foggin JM, Pearson D, Smith AT (2004) Adjacent nesting by lammergeier Gypaetus barbatus and Himalayan griffon Gyps himalayensis on the Tibetan Plateau, China. Forktail 20:94–96
Kentie R, Coulson T, Hooijmeijer JCEW, Howison RA, Loonstra AHJ, Verhoeven MA, Both C, Piersma T (2018) Warming springs and habitat alteration interact to impact timing of breeding and population dynamics in a migratory bird. Global Chang Biol 24:5292–5303. https://doi.org/10.1111/gcb.14406
Khatri PC (2015) First nesting of critically endangered vulture in Bikaner: the nest site record of long billed vulture (Gyps indicus) in Kolayat tehsil, Bikaner. Int J Inn Res Rev 3:8–13
Khwarahm NR, Ararat K, Qade S, Al-Quraishi AMF (2021) Modelling habitat suitability for the breeding Egyptian vulture (Neophron percnopterus) in the Kurdistan Region of Iraq. Iran J Sci Technol Trans A 45:1519–1530. https://doi.org/10.1007/s40995-021-01150-z
Kumar S, Stohlgren TJ (2009) MaxEnt modeling for predicting suitable habitat for threatened and endangered tree Canacomyrica monticola in New Caledonia. J Ecol Nat Environ 1:94–98
Kumar S, Meena H, Jangid PK, Nama KS (2014) Current status of vulture population in Chambal Valley of Kota, Rajasthan. Int J Pure Appl Biosci 2:224–228
Kupika OL, Gandiwa E, Kativu,S, Nhamo G (2018) Impacts of climate change and climate variability on wildlife resources in Southern Africa: experience from selected protected areas in Zimbabwe. In: Sen B, Grillo O (eds) Selected studies in biodiversity. IntechOpen. https://doi.org/10.5772/intechopen.70470
Lane JE (2018) Climate crisis and the “We”: an essay in deconstruction. Int J Manag Stud Res 6:34–43. https://doi.org/10.22158/jepf.v4n3p208
Lemes P, Loyola RD (2013) Accommodating species climate-forced dispersal and uncertainties in spatial conservation planning. PLoS One 8:e54323. https://doi.org/10.1371/journal.pone.0054323
Liminana R, Soutullo A, Arroyo B, Urios V (2012) Protected areas do not fulfil the wintering habitat needs of the trans-Saharan migratory Montagu’s harrier. Biol Conserv 145:62–69. https://doi.org/10.1016/j.biocon.2011.10.009
Lobo JM, Jiménez-Valverde A, Real R (2008) AUC: a misleading measure of the performance of predictive distribution models. Global Ecol Biogeogr 17:145–151. https://doi.org/10.1111/j.1466-8238.2007.00358.x
Lobo JM, Jiménez-Valverde A, Hortal J (2010) The uncertain nature of absences and their importance in species distribution modelling. Ecography 33:103–114. https://doi.org/10.1111/j.1600-0587.2009.06039.x
Majgaonkar I, Bowden CGR, Quader S (2018) Nesting success and nest-site selection of white-rumped vultures (Gyps bengalensis) in western Maharashtra, India. J Raptor Res 52:431–442. https://doi.org/10.3356/JRR-17-26.1
Manning MR, Edmonds J, Emori S et al (2010) Misrepresentation of the IPCC CO2 emission scenarios. Nat Geosci 3:376–377. https://doi.org/10.1038/ngeo880
Manzoor SA, Griffiths G, Lukac M (2018) Species distribution model transferability and model grain size – finer may not always be better. Sci Rep 8:7168. https://doi.org/10.1038/s41598-018-25437-1
Mateo RG, De La Estrella M, Felicísimo ÁM, Munoz J, Guisan A (2013) A new spin on a compositionalist predictive modelling framework for conservation planning: a tropical case study in Ecuador. Biol Conserv 160:150–161. https://doi.org/10.1016/j.biocon.2013.01.014
Mateo-Tomas P, Olea PP (2010) Anticipating knowledge to inform species management: predicting spatially explicit habitat suitability of a colonial vulture spreading its range. PLoS One 5:e12374. https://doi.org/10.1371/journal.pone.0012374
McDonald MM, Johnson SM, Henry ER, Cunneyworth PMK (2019) Differences between ecological niches in northern and southern populations of Angolan black and white colobus monkeys (Colobus angolensis palliatus and Colobus angolensis sharpei) throughout Kenya and Tanzania. Am J Primatol 2019:e22975. https://doi.org/10.1002/ajp.22975
Merow C, Smith MJ, Silander JA Jr (2013) A practical guide to MaxEnt for modelling species’ distributions: what it does, and why inputs and settings matter. Ecography 36:1058–1069. https://doi.org/10.1111/j.1600-0587.2013.07872.x
Misher C, Bajpai H, Bhattarai S, Sharma P, Sharma R, Kumar N (2017) Observations on the breeding of Indian long-billed vultures Gyps indicus at Gapernath, Chambal River in Rajasthan, India. Vulture News 72:14–21. https://doi.org/10.4314/vulnew.v72i1.2
MoEFCC (2020) Action Plan for Vulture Conservation in India, 2020‐2025. Ministry of Environment, Forest and Climate Change Government of India, New Delhi
Mohammadi S, Ebrahimi E, Shahriari MM, Bosso L (2019) Modelling current and future potential distributions of two desert jerboas under climate change in Iran. Ecol Inf 52:7–13. https://doi.org/10.1016/j.ecoinf.2019.04.003
Moleón M, Cortés-Avizanda A, Pérez-García JM, Bautista J, Geoghegan C, Carrete M, Amar A, Sánchez-Zapata JA, Donázar JA (2020) Distribution of avian scavengers inside and outside of protected areas: contrasting patterns between two areas of Spain and South Africa. Biodivers Conserv 29:3349–3368. https://doi.org/10.1007/s10531-020-02027-0
Morales N, Fernández IC, Baca-González V (2017) MaxEnt’s parameter configuration and small samples: are we paying attention to recommendations? A systematic review. PeerJ 5:e3093. https://doi.org/10.7717/peerj.3093
Mori GM, Castillo EB, Guzmán CT, Sánchez DAC, Valqui BKG, Oliva M, Bandopadhyay S, Salas López R, Rojas Briceño NB (2020) Predictive modelling of current and future potential distribution of the spectacled bear (Tremarctos ornatus) in Amazonas, Northeast Peru. Animals 10:1816. https://doi.org/10.3390/ani10101816
Mushtaq S, Reshi ZA, Shah MA, Charles B (2021) Modelled distribution of an invasive alien plant species differs at different spatiotemporal scales under changing climate: a case study of Parthenium hysterophorus L. Trop Ecol 62:398–417. https://doi.org/10.1007/s42965-020-00135-0
Naoroji R (2006) Birds of prey of the Indian subcontinent. Om Books International, New Delhi
Navaneethan B, Sankar K, Qureshi Q, Manjrekar M (2015) The status of vultures in Bandhavgarh Tiger Reserve, Madhya Pradesh, central India. J Threat Taxa 7:8134–8138. https://doi.org/10.11609/jott.2428.7.14.8134-8138
NDDB (2019) Livestock population in India by Species. National Dairy Development Board. https://www.nddb.coop/information/stats/pop. Accessed 31 Jul 2022
Ogada DL, Keesing F, Virani MZ (2011) Dropping dead: causes and consequences of vulture population declines worldwide. Ann N Y Acad Sci 1249:57–71. https://doi.org/10.1111/j.1749-6632.2011.06293.x
Oliveira MdR, Szabo JK, Júnior AdS, Guedes NMR, Tomas WM, Camilo AR, Padovane CR, Peterson AT, Garcia LC (2021) Lack of protected areas and future habitat loss threaten the hyacinth macaw (Anodorhynchus hyacinthinus) and its main food and nesting resources. Ibis 163:1217–1234. https://doi.org/10.1111/ibi.12982
Panthi S, Pariyar S, Low M (2021) Factors influencing the global distribution of the endangered Egyptian vulture. Sci Rep 11:21901. https://doi.org/10.1038/s41598-021-01504-y
Pearce J, Ferrier S (2000) An evaluation of alternative algorithms for fitting species distribution models using logistic regression. Ecol Model 128:127–147. https://doi.org/10.1016/S0304-3800(99)00227-6
Pearson RG, Raxworthy CJ, Nakamura M, Peterson AT (2007) Predicting species distributions from small numbers of occurrence records: a test case using cryptic geckos in Madagascar. J Biogeogr 34:102–117. https://doi.org/10.1111/j.1365-2699.2006.01594.x
Phillips SJ, Dudík M (2008) Modeling of species distributions with MaxEnt: new extensions and a comprehensive evaluation. Ecography 31:161–175. https://doi.org/10.1111/j.0906-7590.2008.5203.x
Phillips SJ, Anderson RP, Schapire RE (2006) Maximum entropy modelling of species geographic distribution. Ecol Model 190:231–259. https://doi.org/10.1016/j.ecolmodel.2005.03.026
Phillips SJ, Dudík M, Schapire RE (2004) A maximum entropy approach to species distribution modeling. In: Greiner R, Schuurmans D (eds) Proceedings of the 21st International Conference on Machine Learning. Alberta, Canada, pp 655–662
Prakash V, Bishwakarma MC, Chaudhary A, Cuthbert R, Dave R, Kulkarni M, Kumar S, Paudel K, Ranade S, Shringarpure R, Green RE (2012) The population decline of Gyps vultures in India and Nepal has slowed since veterinary use of diclofenac was banned. PLoS One 7:e49118. https://doi.org/10.1371/journal.pone.0049118
Prakash V, Galligan TH, Chakraborty SS, Dave R, Kulkarni MD, Prakash N, Shringarpure RN, Ranade SP, Green RE (2019) Recent changes in populations of critically endangered Gyps vultures in India. Bird Conserv Int 29:55–70. https://doi.org/10.1017/S0959270917000545
Preston KL, Rotenberry JT, Redak RA, Michael F, Allen MF (2008) Habitat shifts of endangered species under altered climate conditions: importance of biotic interactions. Global Chang Biol 14:2501–2515. https://doi.org/10.1111/j.1365-2486.2008.01671.x
Proosdij AJ, Sosef M, Wieringa J, Raes N (2016) Minimum required number of specimen records to develop accurate species distribution models. Ecography 39:542–552. https://doi.org/10.1111/ecog.01509
Regos A, Tapia L, Gil-Carrera A, Domínguez J (2021) Caution is needed when using niche models to infer changes in species abundance: the case of two sympatric raptor populations. Animals 11:2020. https://doi.org/10.3390/ani11072020
Rew J, Cho Y, Moon J, Hwang E (2020) Habitat suitability estimation using a two-stage ensemble approach. Remote Sens 12:1475. https://doi.org/10.3390/rs12091475
Richard K, Abdel-Rahman EM, Mohamed SA, Ekesi S, Borgemeister C, Landmann T (2018) Importance of remotely-sensed vegetation variables for predicting the spatial distribution of African citrus triozid (Trioza erytreae) in Kenya. Int J Geo-Inf 7:429. https://doi.org/10.3390/ijgi7110429
Robinson OJ, Ruiz-Gutierrez V, Reynolds MD, Golet GH, Strimas-Mackey M, Fink D (2020) Integrating citizen science data with expert surveys increases accuracy and spatial extent of species distribution models. Divers Distrib 26:976–986. https://doi.org/10.1111/ddi.13068
Rodríguez JP, Brotons L, Bustamante J, Seoane J (2007) The application of predictive modelling of species distribution to biodiversity conservation. Divers Distrib 13:243–251. https://doi.org/10.1111/j.1472-4642.2007.00356.x
Schmitt S, Pouteau R, Justeau D, de Boisseu F, Birnbaum P (2017) SSDM: an R package to predict distribution of species richness and composition based on stacked species distribution models. Methods Ecol Evol 8:1795–1803. https://doi.org/10.1111/2041-210X.12841
Sesink-Clee PR, Abwe EE, Ambahe R et al (2015) Chimpanzee population structure in Cameroon and Nigeria is associated with habitat variation that may be lost under climate change. BMC Ecol Evol 15:2. https://doi.org/10.1186/s12862-014-0275-z
Shabani F, Kumar L, Ahmadi M (2016) A comparison of absolute performance of different correlative and mechanistic species distribution models in an independent area. Ecol Evol 6:5973–5986. https://doi.org/10.1002/ece3.2332
Shabani F, Kumar L, Ahmadi M (2018) Assessing accuracy methods of species distribution models: AUC, specificity, sensitivity and the true skill statistic. Global J Hum Soc Sci b 18:1–13
Sharma PD (2005) Ecology and environment. Rastogi Publications, Meerut, India
Somodi I, Lepesi N, Botta-Dukát Z (2017) Prevalence dependence in model goodness measures with special emphasis on true skill statistics. Ecol Evol 7:863–872. https://doi.org/10.1002/ece3.2654
Sony RK, Sen S, Kumar S, Sen M, Jayahari KM (2018) Niche models inform the effects of climate change on the endangered Nilgiri tahr (Nilgiritragus hylocrius) populations in the southern Western Ghats, India. Ecol Eng 120:355–363. https://doi.org/10.1016/j.ecoleng.2018.06.017
Stiels D, Schidelko K (2018) Modeling avian distributions and niches: insights into invasions and speciation in birds. In: Tietze DT (ed) Bird species, fascinating life sciences. Springer, Cham, pp 147–164
Stocker T (2014) Climate change 2013: the physical science basis. Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, UK
Sullivan BL, Wood CL, Iliff MJ, Bonney RE, Fink D, Kelling S (2009) EBird: a citizen-based bird observation network in the biological sciences. Biol Conserv 142:12282–12292. https://doi.org/10.1016/j.biocon.2009.05.006
Summers DM, Bryan BA, Crossman ND, Myer WS (2012) Species vulnerability to climate change: impacts on spatial conservation priorities and species representation. Global Chang Biol 18:2335–2348. https://doi.org/10.1111/j.1365-2486.2012.02700.x
Sun X, Long Z, Jia J (2021) A multi-scale Maxent approach to model habitat suitability for the giant pandas in the Qionglai Mountain, China. Global Ecol Conserv 30:e01766. https://doi.org/10.1016/j.gecco.2021.e01766
Sutton WB, Barrett K, Moody AT, Loftin CS, de Maynadier PG, Nanjappa P (2015) Predicted changes in climatic niche and climate refugia of conservation priority salamander species in the northeastern United States. Forests 6:1–26. https://doi.org/10.3390/f6010001
Swets JA (1988) Measuring the accuracy of diagnostic systems. Science 240:1285–1293. https://doi.org/10.1126/science.3287615
Tehrani NA, Naimi B, Jaboyedoff M (2021) Modelling current and future species distribution of breeding birds as regional essential biodiversity variables (SD EBVs): a bird perspective in Swiss Alps. Global Ecol Conserv 27:e01596. https://doi.org/10.1016/j.gecco.2021.e01596
Thapa S, Baral S, Hu Y, Huang Z, Yue Y, Dhakal M, Jnawali SR, Chettri N, Racey PA, Yu W, Wu Y (2021) Will climate change impact distribution of bats in Nepal Himalayas? Global Ecol Conserv 26:e01483. https://doi.org/10.1016/j.gecco.2021.e01483
Thomas CD, Cameron A, Green RE et al (2004) Extinction risk from climate change. Nature 427:145–148. https://doi.org/10.1038/nature02121
UPFD, BNHS (2021) Determination of the status and distribution of vultures in Uttar Pradesh. Uttar Pradesh, Lucknow and Bombay Natural History Society, Mumbai, Forest Department
Urbina-Cardona N, Blair ME, Londono MC, Loyola R, Velasquez-Tibata J, Morales-Devia H (2019) Species distribution modelling in Latin America: a 25-year retrospective review. Trop Conserv Sci 12:1–19. https://doi.org/10.1177/1940082919854058
USGS EROS (2018) Shuttle radar topography Mission 1 Arc-Second Global. https://doi.org/10.5066/F7PR7TFT. Accessed 24 February 2023
Venter O, Sanderson EW, Magrach A, Allan JR, Beher J, Jones KR, Possingham HP, Laurance WF, Wood P, Fekete BM, Levy MA, Watson JEM (2016) Data descriptor: global terrestrial human footprint maps for 1993 and 2009. Sci Data 3:160067. https://doi.org/10.1038/sdata.2016.67
Vu TT, Tran DV, Tran HTP, Nguyen MD, Do TA, Ta NT, Cao HT, Pham NT, Phan DV (2019) An assessment of the impact of climate change on the distribution of the grey-shanked douc Pygathrix cinerea using an ecological niche model. Primates 61:267–275. https://doi.org/10.1007/s10329-019-00763-8
Wisz MS, Hijmans RJ, Li J, Peterson AT, Graham CH, Guisan A, NCEAS PSDWG (2008) Effects of sample size on the performance of species distribution models. Divers Distrib 14:763–773. https://doi.org/10.1111/j.1472-4642.2008.00482.x
Zaniewski AE, Lehmann A, Overton JM (2002) Predicting species spatial distributions using presence-only data: a case study of native New Zealand ferns. Ecol Model 157:261–280. https://doi.org/10.1016/S0304-3800(02)00199-0
Zhang J, Jiang F, Li G, Qin W, Li S, Gao H, Cai Z, Lin G, Zhang T (2019) Maxent modelling for predicting the spatial distribution of three raptors in the Sanjiangyuan National Park, China. Ecol Evol 9:6643–6654. https://doi.org/10.1002/ece3.5243
Acknowledgements
The authors would like to record appreciation to Dibyendu Biswas, Researcher, Wildlife Institute of India, for his contribution in addressing some observations of the reviewers. We also acknowledge the efforts of reviewers in improving the manuscript.
Author information
Authors and Affiliations
Contributions
Both the authors have contributed equally.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Communicated by Rafael A. Dias (Associate Editor).
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Jha, R., Jha, K.K. Environmental factors shaping habitat suitability of Gyps vultures: climate change impact modelling for conservation in India. Ornithol. Res. 31, 119–140 (2023). https://doi.org/10.1007/s43388-023-00124-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s43388-023-00124-6