
Abstract
Vultures are an indispensable part of a healthy and balanced ecosystem. Vultures migrate from breeding to wintering grounds to access resources which are otherwise unavailable. Despite the possible effect of climate change, their wintering habitat is not sufficiently studied. We studied wintering habitats of two Eurasian vultures—Griffon Vulture (Gyps fulvus; EGV) and Cinereous Vulture (Aegypius monachus; EBV) in northern India. Habitat distribution model (Maxent) and global circulation models (CCSM4, HadGEM2AO, MIROC5) were used to predict current and future (RCP2.6, RCP4.5 and RCP8.5 for 2050 and 2070) habitats of both the species. We identified influential habitat variables to determine current and future habitat suitability and suggested appropriate conservation measures. Model prediction showed that mean temperature of the driest quarter and precipitation of the wettest month were the two major influential variables with highest contribution, though landcover also played an important role. Suitable area expanse for EBV (12%) was smaller than EGV (38%) in the available 240,928 km2. The predictive suitability map showed the dynamics of suitable and unsuitable habitats indicating range expansion (3–16%) and contraction (1–6%). Their shift was multidirectional occurring towards the Northwest, East, Southwest and Southeast. Across the scenario, impact of climate change was positive (net gain in suitable area) in most of the cases. The findings show that the Tarai districts need improvement in habitat management to provide safe roosting and foraging sites for Eurasian vultures with a large migratory pattern.
Zusammenfassung
Wie sicher sind die Überwinterungsgebiete eurasischer Geier beim Klimawandel noch? Eine Studie zu Habitatmodellen in Nordindien.
Geier sind ein unverzichtbarer Bestandteil eines gesunden und ausgewogenen Ökosystems; sie ziehen von den Brutgebieten in die Winterquartiere, um Zugang zu Nahrungsquellen zu haben, die ansonsten nicht zur Verfügung stehen. Obwohl sich der Klimawandel vermutlich auf die Überwinterungsgebiete auswirkt, sind diese nicht ausreichend untersucht. Wir untersuchten die Winterquartiere zweier eurasischer Geierarten, Gänsegeier (Gyps fulvus; EGV) und Mönchsgeier (Aegypius monachus; EBV), in Nordindien. Hierfür wurden das Habitatverteilungsmodell (Maxent) und Modelle für die globale Zirkulation (CCSM4, HadGEM2AO, MIROC5) zur Vorhersage der aktuellen und zukünftigen (RCP2.6, RCP4.5 und RCP8.5 für 2050 und 2070) Lebensräume beider Arten verwendet. Wir bestimmten die maßgeblichen Merkmale dieser Lebensräume, um sie auf ihre aktuelle und künftige Eignung zu prüfen und geeignete Maßnahmen zu ihrer Erhaltung vorzuschlagen. Die Vorhersage aus den Modellen zeigte, dass die mittlere Temperatur des trockensten Quartals und der Niederschlag des feuchtesten Monats die beiden wichtigsten Einflussfaktoren waren, wobei auch die Bodendecke eine wichtige Rolle spielte. Auf den verfügbaren 240.928 km2 war die geeignete Fläche für EBV (12 %) kleiner als die für EGV (38 %). Die Landkarte der vorhergesagten Eignung zeigte die Dynamik geeigneter und ungeeigneter Habitate, die auf eine Ausweitung (3 % bis 16 %) und eine Verringerung (1 % bis 6 %) des Verbreitungsgebiets hinweist. Die Verschiebung erfolgte in mehrere Richtungen: nach Nordwesten, Osten, Südwesten und Südosten. Für fast alle Szenarien waren in den meisten Fällen die Auswirkungen des Klimawandels positiv: ein Nettogewinn an geeigneter Fläche. Die Ergebnisse zeigen, dass in den Tarai-Distrikten die Gestaltung und das Management der Lebensräume verbessert werden müssen, um sichere Schlaf- und Futterplätze für die über weite Entfernungen ziehenden eurasischen Geier zu schaffen.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
India has been a party to the Convention on Conservation of Migratory Species, an environmental treaty of the UN that provides a global platform for the conservation and sustainable use of migratory animals and their habitats. It is committed to the “Urgent need to step up actions for vultures in Africa and Eurasia” (Botha et al. 2024) and elsewhere. Two of the three migratory vultures in India, Cinereous Vultures (Aegypius monachus Linnaeus 1766 or Eurasian Black Vulture), hereafter EBV, and (Eurasian) Griffon Vultures (Gyps fulvus Hablizl 1783), hereafter EGV, are Eurasian species that migrate to spend the non-breeding season in northern African and Eurasian countries including India (BirdLife International 2021a,b). They are commonly found in north, west and central Indian states during winters (Jha 2015; MoEFCC 2020; Jha and Jha 2021; Ram et al. 2022).
EBV is categorised as “Near threatened” resident with declining population in Asia and Europe (BirdLife International 2021a). This species has been reported to migrate within Asian countries (Kang et al. 2019). EGV is categorised as “Least Concerned” resident with increasing population in Africa, Europe and Asia (BirdLife International 2021b). A recent telemetry study recorded the movement of EGV between India and Russia through central Asian republics (Ram et al. 2022).
These species are exposed to several threats such as energy production and mining (renewable energy), transportation and service corridors (utility and service lines), biological resource use (hunting and trapping terrestrial animals), human intrusions and disturbance (recreational activities), natural system modifications, climate change and weather extremes during their residency including wintering grounds (BirdLife International 2021a,b). Batbayar et al. (2008) reviewed that one of the Eurasian vultures faced a lack of suitable habitat in the wintering grounds in several Asian countries. As Jha et al. (2022) explicate, wintering involves spending significant time in improved living conditions in comparison to the bird’s original home. Survival entails that sites must have abundant food, shelter, minimum disturbance, and optimal weather conditions (Clausen et al. 2018).
Climate change is causing changes in global climate-related parameters, mainly temperature and precipitation. Increase in global temperatures is predicted to continue (IPCC 2021), influencing other climatic parameters and events. Northern India is particularly susceptible to climate change, experiencing diverse impacts such as heatwaves, altered rainfall patterns, a rise in the frequency of extreme weather events etc. (Awasthi et al. 2023). These may affect raptors, or vultures, in various ways, including changes in distribution ranges, migration, abundance, population dynamics, etc. (Martinez-Ruiz et al. 2023). A La Nina induced drought has already shown its impact on vultures of Northwestern India (Hall et al. 2012). Heat stress is known to affect the mortality of cattle (Das et al. 2016), in turn creating a reduction of food stock for vultures. Following the predicted acceleration of climate change, some raptor species will become more vulnerable to the increased variation in climate conditions and extreme weather patterns. Moreover, the threat to migratory birds has grown due to habitat destruction and so it is imperative that their habitat and its conditions be studied to facilitate their conservation (Yong et al. 2021; Jha et al. 2022).
Habitat (distribution) models, also known as species distribution models (SDMs) or ecological niche models (Elith and Leathwick 2009), are used for many applications in ecology and conservation. These models are among the most frequently used methods in studying avian ecology, evolution, biogeography, and conservation. They have been applied to identifying suitable habitats, predicting the impact of climate change on distribution, etc. in vulture species (Anoop et al. 2020; Jha and Jha 2024; Subedi et al. 2023 etc.). Out of a host of SDMs developed, Maxent was considered more useful (Kaky et al. 2020) and applied in the northern Indian context (Jha et al. 2022; Jha and Jha 2024).
Based on the background literature, in this study, we aimed to identify the habitat variables controlling the residential suitability of EBV and EGV in northern India, determine the extent of suitable habitats for these vultures, and assess their susceptibility to climate change. Additionally, we aimed to develop potential management implications and conservation strategies to mitigate any identified threats to their habitats and populations.
Materials and methods
Study sites, vulture species and occurrence data
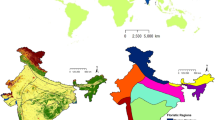
Uttar Pradesh, a north Indian state where wintering of EBV and EGV is reported (Naoroji 2006), was selected for the study. This state lies between 23°52′ N and 30°24′ N latitude, and 77°5′ E and 84°38′ E longitude (Fig. 1). Its geographical area of 240,928 km2 is divided into four ecozones: Tarai, Bundelkhand–Vindhyan, Semi-arid and Gangetic with a varying combination of vegetation, temperature, and precipitation. It has a humid subtropical dry winter climate (Jha 2015). The ranges of elevation, mean annual temperature and mean annual precipitation are 23 m–935 m, 21.01–26.56 °C and 601 mm–2145 mm, respectively (Fick and Hijmans 2017).

Study area location map (inset) with land-use landcover, transect survey routes and occurrence points (Modified: Jha and Jha 2024)
A vulture occurrence (transect) survey was conducted during the winters of 2020 and 2021 along the tracks (Fig. 1). This was decided after consulting the literature, experts, and local forest staff. Occurrence data were also procured from a published record—Jha (2015). The data pool was further supplemented using citizen science repositories, eBird and iNaturalist (Sullivan et al. 2009; iNaturalist users and Ueda 2020). The data were finally subjected to cleaning, duplicate removal, and spatial rarefication using SDM toolbox of ArcGIS 10.5 (Brown et al. 2017). Low number of final occurrence records for modelling was taken care of using bias file for generating background points as also done recently by Peterson et al. (2024) with six to eight data points.
Current and future climate envelope
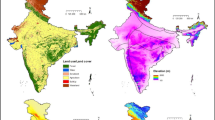
Biologically significant 19 climatic layers of current day, for three global circulation models (CCSM4, HadGEM2AO and MIROC5) of optimistic, intermediate, and pessimistic emission scenarios (RCP2.6, RCP4.5 and RCP8.5) (Anoop et al. 2020) were downloaded from www.worldclim.org (Fick and Hijmans 2017) at 30 Arc seconds. Elevation data from Earth Resources Observation and Science (EROS) (USGS EROS 2018) and two sets of normalised differentiated vegetation index (NDVI) data, winter (January) and summer (June), (Didan 2015) at 250 m resolution were downloaded from www.earthexplorer.usgs.gov. Land use and land cover (LULC) was downloaded at 100 m resolution from www.land.copernicus.eu (Buchhorn et al. 2020). LULC types were merged and the map was reclassified into six classes (Forest, Water, Scrubland, Agriculture, Built-Up and Wasteland) as the present study did not require a detailed classification (Jha and Jha 2021). All the layers were resampled to 1 km (30 Arc seconds) spatial resolution using ArcGIS 10.5. Finally, these variables (electronic supplementary material Table S1) were subjected to Pearson collinearity test to eliminate multicollinearity among variables at ± 0.7 threshold. For future projections, LULC and NDVI were not used since it is very challenging to get such layers. Moreover, bioclimatic variables are most used and often important for modelling geographic distribution of a species (Ardestani et al. 2021) and spatial distribution of a species is highly correlated to temperature and precipitation (Gaudreau et al. 2018). Zegarra et al. (2020) further recorded that the climate represents part of the niche environment and it is possible to predict suitable area of a species.
Model selection and run
Apart from the reasons mentioned in the introduction, selection of Maxent was based on its merits: (i) it required only presence data to generate the probability of presence (Kumar and Stohlgren 2009), (ii) it could work with a small sample size (Abolmaali et al. 2018), (iii) it outperformed other methods when the number of geographic records was scarce (Elith et al. 2006), (iv) both, categorical and continuous environmental layers, could be applied in this software, (v) it could measure the importance of each environmental variable using the jackknife test, in terms of gain (Elith et al. 2011). Maxent programme options were set at default (e.g. auto features) except as suggested in Dong et al. (2019). The random test was set at 25% for training and 75% for testing. The algorithm run type was Bootstrap with 10 replicates, 500 iterations, 0.00001 convergence threshold, and 10,000 background points.
Output data and processing
Models were evaluated using area under receiver operating characteristic curve (AUC) and continuous Boyce index (CBI). Variable contribution and importance were used as such but rasterised maps were reclassified into unsuitable (0–0.25), low suitable (0.25–0.5), moderate suitable (0.5–0.75), and highly suitable (0.75–1) categories of habitat suitability following Jha and Jha (2024). The rasterised maps were again reclassified into unsuitable (0–0.25) and suitable (0.25–1.0) categories for future and present scenarios. Using these, suitable area loss and gain categorical maps were also prepared to estimate the area dynamics and expanse in future scenarios. ArcGIS 10.5 was used for map generation, area calculation and centroid marking.
Results
Original sample pool of occurrence record for EBV and EGV was reduced from 35 and 36 to 16 and 17, respectively, after cleaning, duplicate removal, and spatial rarefication of data. Pearson test of collinearity resulted in selection of noncolinear variables such as, bio1, bio2, bio3, bio8, bio9, bio13, bio14, bio15, elevation, LULC, and NDVI. Model strength in terms of AUC was 0.93 (EBV) and 0.84 (EGV), and in terms of CBI it was 0.691 (EBV) and 0.703 (EGV).
Bioclimatic variables and species’ response
Jackknife chart (electronic supplementary material Fig. S1) suggested that four most important variables were bio13, bio9, bio1 and bio2 (EBV), and bio13, bio1, bio9 and bio8 (EGV). But, as per variable contribution table, three vital variables (sensu Zhang et al. 2020) with a ~ 90% contribution to niche characterisation (electronic supplementary material Table S2) were bio9, bio13, and bio2 (EBV) and bio9, bio1 and bio13 (EGV). Two common vital variables, bio9 and bio13, contributed 66% (EGV) to 81% (EBV) in habitat determination. In future scenarios, the contribution of bioclimatic variables was similar. After supplementing the biophysical variables to the current, LULC showed its importance as the fourth variable in both the vultures. Within LULC, forest and waterbody were prominent. However, increasing mean temperature of driest quarter (bio9) decreased the presence or suitability of the habitat of both the vultures. Increase in precipitation of wettest month (bio13) increased the presence probability in both the species (Fig. 2). For the presence probability 50% and above, the range of mean temperature of the driest quarter was 16–21 °C in EBV, while it was slightly wider in the case of EGV (16–25 °C). The range of precipitation of the wettest month was around 400 mm–700 mm in both cases.

Response of vultures (Eurasian Black Vulture and Eurasian Griffon Vulture) or habitat suitability towards major bioclimatic habitat variables (bio9 and bio13)
Current and future habitat suitability
As per the prediction of the bioclimatic model, suitable area in current condition was 12% (EBV) and 38% (EGV) against a total area 240,928 km2 (Fig. 3). Suitable area in bioenvironmental and bioclimatic model prediction was almost similar. It showed approximately 1% area difference: negative in EBV while positive in EGV. Within the suitable categories, the quantum of areas of low, moderate, and high suitability was in decreasing order (electronic supplementary material Table S3). In the near future, EBV showed an increasing trend in suitable area (exception RCP4.5) while EGV demonstrated a decreasing trend (exception RCP4.5). In the distant future, both vultures showed increasing trends across the emission pathways. Figures 4 and 5 may be seen for pictorial understanding, though the differences are not visually significantly appreciable.

Bioclimatic habitat suitability distribution of Eurasian black vulture and Eurasian griffon vulture in Uttar Pradesh

Maps depicting area suitability classes under different scenarios of near future

Maps depicting area suitability classes under different scenarios of distant future
Habitat suitability dynamics
We found changes in the niche or habitat of the wintering species in the future with respect to the current projection. They are discussed in terms of area expansion (gain), area regression (loss), area conservation/ preservation (stable suitable habitat) and out-niche (stable unsuitable habitat) (sensu Adjonou et al. 2020). The major part of current habitat for both the vultures, suitable (11–12% in EBV and 32–38% in EGV) and unsuitable (85–88% in EBV and 46–61% in EGV), will remain stable but a smaller area, will undergo changes in the future due to changed climatic conditions (electronic supplementary material Table S4 and Figs. 6–7). The model shows maximum expansion or gain up to 3% (RCP2.6) but almost no contraction or loss, exception (RCP4.5), up to < 1% in suitable area in the near future in EBV. In EGV, there will be expansion (maximum up to 2% in RCP4.5) as well as contraction (maximum up to 6% in RCP8.5) in suitable area in the near future across scenarios. In the distant future, across the emission scenarios, there will be an expansion of suitable area (around 1%) as well as a contraction (much < 1%) in suitable area in EBV. In the case of EGV, the situation is similar but the quantum of expansion and contraction in suitable area is much higher (gain 6–16%; loss 1–3%).

Maps depicting area dynamics (contraction and expansion) under different scenarios of near future

Maps depicting area dynamics (contraction and expansion) under different scenarios of distant future
Vector and scalar movement of the centroid of suitable area in different future scenarios with respect to current scenario is presented in Fig. 8. Centroid movement was observed in EBV towards the southwest, northwest, and southeast directions, while in EGV, it was towards southwest, northwest, and East. Scalar movement was found to be 4 km–13 km in the former, while 6 km–38 km in the latter (electronic supplementary material Table S5).

Maps showing centroids of suitable area under future scenarios with reference to current. Zoomed portion of the map elucidates vector and scaler shift
Discussion
Model improvement and evaluation
SDMs are subject to many sources of uncertainty, limiting their application in research and practice (Tessarolo et al. 2021). We improved our models by introducing data cleaning, duplicate removal, and spatial rarefication in intensively collected multiple source occurrence data (Brown et al. 2017). Though sample size does matter in modelling, our sample size for EBV (16) and EGV (17), was supported by Hernandez et al. (2006) who could produce useful models with as few as five to ten positive observations, and Maxent’s inbuilt feature of maximising a lower sample size. However, higher sample size (20–30) can overcome this limitation (Jiménez-Valverde 2020). Collinearity impact of environmental parameters was reduced by discarding colinear variables (bio4, bio5, bio6, bio7, bio10, bio11, bio12, bio16, bio17, bio18, and bio19) using Pearson test of collinearity (Manzoor et al. 2018). For correction of latitudinal bias, a bias file was used during the model run (Brown et al. 2017). Sources of uncertainty in future climate data due to differences between GCMs and natural climate variability were addressed using an ensemble of three GCMs (Gould et al. 2014). Incorporating future NDVI and LULC was challenging but for current day prediction, they were taken to address proxy of food (Santangeli et al. 2018) and shelter quality, respectively. Inclusion of summer NDVI in SDM improves the prediction of habitat suitability (Salas et al. 2017; Konatowska et al. 2023), though both the NDVI in the present case were of low significance (4–5%) in niche contribution. Therefore, it is argued that animal density should be used as a variable instead of the proxy.
For model strength evaluation, we did not depend fully on commonly used evaluator AUC, which is criticised for inadequacy but considered acceptable for presence–absence data (Lobo et al. 2008). We also used CBI which is useful for presence only data model (Hirzel et al. 2006), such as Maxent. These evaluators indicated that our models were in the good to excellent categories, therefore, the predictions analysed below are reliable and can be useful for the development of conservation strategies (Pearce and Ferrier 2000).
Predictor variables and species’ response
Bioclimatic as well as biophysical variables played their role to varying degrees in niche determination. For example, in the bioenvironmental model (bioclimatic + biophysical variables), LULC was the fourth important variable as per the jackknife test but contributed to 26% (EBV) and 20% (EGV) of the niche. Contrastingly, the first important variable in EBV (bio13) and second important variable in EGV (bio1) contributed 19% and 20%, respectively. Plausible explanation of this jumbling up of the position (importance versus quantum of contribution) may be due to interaction among the variables and some could play a limiting role in habitat determination. Jha and Jha (2021) have affirmed that habitat suitability was not a function of a single variable, rather it was a product of the interaction among numerous covariates in different quantities (grades) in which quite a few could be following Liebig’s law of the minimum (Golterman, 1975). However, Gschweng et al. (2012) and Liminana et al. (2012) have noted the influence of bioclimatic variables in habitat determination. Herrero et al. (2006) and Bosch et al. (2014) have also asserted that vegetation cover or forested area is reported to influence the distribution of an animal species since it determines the land’s ability to supply food and/or shelter to animals. Therefore, landcover functions as a limiting factor for the spread of a species. Nevertheless, the role of the two most important variables (bio9 and bio13) cannot be ignored because temperature and precipitation have a significant influence on the distribution of terrestrial vertebrate fauna. These two factors synthesise the flows of energy and water in the ecosystem and substantially limit the global distribution of these fauna (Bosch et al. 2014).
In a broader sense, bio9, bio13, and LULC should be the variables of primary concern for habitat determination of EBV and EGV. For want of future LULC data, only bioclimatic models generated for six scenarios suggested that these two bioclimatic variables would remain the top two contributors of the niche (electronic supplementary material Table S2), always contributing around 80% in EBV and around 57–66% in EGV. The influence of bioclimatic variables in habitat determination has also been noted in earlier studies on raptors and scavengers (Gschweng et al. 2012; Liminana et al. 2012; Phipps et al. 2017; Zhang et al. 2019; Anoop et al. 2020). However, since the residency of migratory vultures is confined to winter months, the presence of vultures would be controlled directly by bio9 but bio13 would govern the occurrence indirectly by providing rains to waterbodies (for direct consumption) and to vegetation for maintaining productivity.
Current and future habitat suitability
Availability of three times more suitable envelop for EGV than EBV is not in agreement with Jha et al. (2022) where EBV has more suitable area than EGV. This difference may be due to the modelling approach adopted, ensemble in the latter instead of standalone Maxent. However, present study indicates that EGV is adaptable to a wider set of climatic conditions than EBV, since the former is physiologically marginally plastic. Irrespective of the unknown carrying capacity of suitable areas, it can be deduced that the studied area is currently safe for a minimum of 29–52 EBV and 85–216 EGV individuals as assessed by Jha (2022). Compared with Jha (2015)’s estimation a decade ago, it is apparent that the arrival of wintering vulture individuals has increased. Though raptors are limited by food (Newton 2003) in addition to their roosting requirements and the assumed availability of the same, the common or overlapping suitable areas of EBV and EGV were mainly in the Tarai ecozone. The area of EGV was extended in adjoining Gangetic plain ecozone which is slightly warmer. This may be corroborated with a wider range of variable condition adaptability for similar presence/ occurrence of species or degree of habitat suitability, for example, mean temperature of driest quarter (November to February in studied area) range 16–21 °C in EBV while 16–25 °C in EGV. Notably, one of the home areas (Kazakhstan for EGV) has a similar temperature range between April and October months (Harris et al. 2020), which is the permanent stay period at home. Within India, these species prefer the northern plains as compared to the high altitudes due to a presumed shortage of resources in high altitude states during the harsh winter months.
Habitat dynamics
It is critical to understand how the spatial distribution or habitat of a species changes under the changing climate for protecting biodiversity and formulating effective policies (Liang et al. 2021). In terms of suitable habitat availability in any scenario, EBV is in a less advantageous position in northern India as compared to EGV, since EBV must face a higher expanse of unsuitable (additional 26%) area. But in terms of adapting to habitat contraction, EGV could face more stress because a larger portion of its habitat may undergo changes. This is under the assumption that the future arrival of EBV or EGV may not find some of the previous sites to which they have had fidelity to be still suitable. However, in the long run, net change (contraction or expansion) would be more relevant from a proactive habitat management point of view.
Since the RCP2.6 scenario is optimistic and ideal and unlikely to be achieved, we should focus on intermediate scenario RCP4.5 which is pessimistic but likely possible if emissions continue to remain uncontrolled (Manning et al. 2010; Lane 2018). Across the scenarios, changes in the expanse of habitat are reported earlier too (Liang et al. 2021). The decrease of habitat area is the main threat to biodiversity caused by climate change (Taubert et al. 2018). However, net gain in suitable habitat is always beneficial for the vultures to allow them to spread over a larger area if abundance is increased. That means EBV will have RCP8.5 more useful in the near future showing a net gain, though RCP2.6 conditions would still be better. In the distant future, RCP4.5 condition would be followed by RCP8.5 and RCP2.6. However, EGV showed a net loss in RCP8.5 and RCP2.6, but a marginal gain in RCP4.5. In the distant future, EGV would have the highest advantage of net gain in RCP4.5 followed by RCP2.6 and RCP8.5. Overall, climate change would favour the wintering vultures in northern India.
Centroid or the core of suitable area was identified to analyse the distance and direction of the shift of EBV and EGV habitats. The current core was in the moist deciduous area which would move in various directions (southwest, northwest, east and southeast) in the same climatic zone. This is contrary to the general belief of poleward shifting (Parmesan and Yohe 2003) but in agreement with VanDerWal et al. (2013)’s multidirectional shift. Shift in habitat due to climate change has been reported in vector as well as scalar forms (Shen et al. 2021). They (Shen et al. 2021) have also reported a reversal in the direction of shift as in our case.
Management implication
Our study projected a good expanse of stable as well as dynamic suitable area for wintering vultures (28,781 km2–35,980 km2 for EBV, 92,557 km2–128, 155 km2 for EGV), mainly confined to forested and surrounding areas in the studied northern state of India. The vast stretches of agriculture land with very sparse tree density were mostly unsuitable. Such habitats available for vulture use should be managed judiciously in their favour keeping their residency need in view. Stable habitat would need long-term maintenance, while unstable habitat would require anticipated mitigation to maintain their suitability. Foraging stock would also need to be maintained within and outside the suitable area.
Since migrating to a new destination allows the birds to escape competition and the harsh winter climate of their original home or breeding territory (Yamac and Bilgin 2012), their requirement at the new home is safe roosting and foraging for the winter months (November to February, many a times extended to March) along with protection from poisoning and infrastructure threats. The EBV spend much of their time soaring overhead in search of food and perch more often on trees. The EGV roosts on large cliffs and can forage outside woodland (BirdLife International 2021a,b). Therefore, logging activities and other human-induced disturbance should be restricted during the wintering season. Suitable roost trees must be identified and preserved as a vital part of the long-term conservation management of the area (Yamac 2007). Jha (2015)’s identification of some roosting / nesting trees (Anogeisus latifolia, Bombax ceiba, Dalbergia sissoo, Ficus religiosa, F. racemosa, Haldina cordifolia, Tamarindus indica, and Tectona grandis) in the study region could be targeted for this. Anthropogenic disturbances, mainly mining or quarrying, should never be allowed in the vicinity of roosting cliffs since birds show fidelity to wintering sites and visit them year after year until altered severely (Fox et al. 1994; Clausen et al. 2018).
Though vultures prefer to roost in forested areas, they also wander towards agriculture landscape in search of carcasses. It would be pertinent to develop an agroforestry system in suitable or unsuitable areas adjoining the forests. This would provide the double advantage of producing domestic mammals as potential food and relatively smaller trees for roosting, if needed (Jha et al. 2022).
The wintering vultures have the benefit of overlapping residency areas with the indigenous or resident vultures. Though sympatry enhances the competition for resources, EBV and EGV would always get the opportunity of using resources from the habitat of resident vultures which are already being managed well, at least in the protected areas (PAs) of the state. Considering the important role of PAs in vulture protection (Jha et al. 2021), it is advisable for the policy makers to consider increasing the forested area under the PAs network which is limited in the study area.
Conclusion
It is essential to protect EBV and EGV from any threat: known or unknown. This study for the first time analysed the impact of climate change on the wintering habitat of these two migratory vulture species in northern India, far away from their breeding home, using maximum entropy principle. With strong prediction power, the Maxent models projected stable as well as unstable areas of suitability along with unsuitability in current time and future scenarios. They showed scalar as well as vector shift in suitable habitats which was in consonance with previous studies. Data from this study indicated significant flux in the habitat of EGV. The habitat of EBV was relatively less dynamic. They were mainly guided by mean temperature of driest quarter and precipitation of wettest month and limited by land-use cover. Since wintering areas are important reservoirs of actively breeding population during unfavourable climates in their home range, habitat protection is essential. In this light, the information generated from this study can be used for effective management of these winter visitors in their temporary habitat, especially due to the changing dynamics of this habitat.
Data availability
All data used in the study are available in the manuscript or associated supplementary material.
References
Abolmaali MRS, Tarkesh M, Bashari H (2018) MaxEnt modeling for predicting suitable habitats and identifying the effects of climate change on a threatened species, Daphne mucronata, in central Iran. Ecol Inf 43:116–123
Adjonou K, Abotsi KE, Segla KN, Rabiou H, Houetchegnon T, Sourou KNB, Johnson BN, Nougbodé Ouinsavi CAI, Kokutse AD, Mahamane A, Kokou K (2020) Vulnerability of African Rosewood (Pterocarpus erinaceus, Fabaceae) natural stands to climate change and implications for silviculture in West Africa. Heliyon 6(6):e04031. https://doi.org/10.1016/j.heliyon.2020.e04031
Anoop NR, Babu S, Nagarajan R, Sen S (2020) Identifying suitable reintroduction sites for the White-rumped Vulture (Gyps bengalensis) in India’s Western Ghats using niche models and habitat requirements. Ecol Eng 158:106034. https://doi.org/10.1016/j.ecoleng.2020.106034
Ardestani EG, Rigi H, Honarbakhsh A (2021) Predicting optimal habitats of Haloxylon persicum for ecosystem restoration using ensemble ecological niche modeling under climate change in southeast Iran. Restor Ecol 29:e13492. https://doi.org/10.1111/rec.13492
Awasthi A, Pattnayak KC, Tandon A, Sarkar A, Chakraborty M (2023) Implications of climate change on surface temperature in North Indian states: evidence from CMIP6 model ensembles. Front Environ Sci 1:1264757. https://doi.org/10.3389/fenvs.2023.1264757
Batbayar N, Reading R, Kenny D, Natsagdorj T, Kee P (2008) Migration and movement patterns of cinereous vultures in Mongolia. Falco 32:5–7
BirdLife International (2021a) Aegypius monachus. The IUCN red list of threatened species 2021: e.T22695231A154915043. https://doi.org/10.2305/IUCN.UK.2021-3.RLTS.T22695231A154915043.en. Accessed 21 Apr 2024
BirdLife International (2021b) Gyps fulvus. The IUCN red list of threatened species 2021: e.T22695219A157719127. https://doi.org/10.2305/IUCN.UK.2021-3.RLTS.T22695219A157719127.en. Accessed 21 Apr 2024
Bosch J, Mardones F, Perez A, Torre AL, Munoz MJ (2014) A maximum entropy model for predicting wild boar distribution in Spain. Span J Agricul Res 12(4):984–999. https://doi.org/10.5424/sjar/2014124-5717
Botha A, Doherty J, Weston J, Andevski J, Safford R, Tavares J, Bowden C, Matsvimbo F, Sebele L, Al-Hasani I, Gallo-Orsi U (2024) Report on the mid-term implementation review of the vulture multispecies action plan to conserve African-Eurasian vultures. CMS Raptors MOU Secretariat, Abu Dhabi. https://www.cms.int/en/publication/mid-term-implementation-review-multi-species-action-plan-conserve-african-eurasian. Accessed 09 Jul 2024
Brown JL, Bennett JR, French CM (2017) SDM tollbox 2.0: the next generation python-based GIS toolkit for landscape, genetic, biogeographic and species distribution model analysis. Peer J 5:e4095. https://doi.org/10.7717/peerj.4095
Buchhorn M, Smets B, Bertels L, De Roo B, Lesiv M, Tsendbazar N-E, Herold M, Fritz S (2020) Copernicus global land service: land cover 100m: collection 3: epoch 2019: Globe. Copernicus Land Service: Global Land Cover / Land Use. https://doi.org/10.5281/zenodo.3939050. Accessed 08 August 2019
Clausen KK, Madsen J, Cottaar F, Kuijken E, Verscheure C (2018) Highly dynamic wintering strategies in migratory geese: coping with environmental change. Global Change Biol 24:3214–3225. https://doi.org/10.1111/gcb.14061
Das R, Sailo L, Verma N, Bharti P, Saikia J, Imtiwati KR (2016) Impact of heat stress on health and performance of dairy animals: a review. Vet World 9:260–268. https://doi.org/10.14202/vetworld.2016.260-268
Didan K (2015) MOD13A3 MODIS/terra vegetation indices monthly L3 global 1km SIN grid V006. NASA EOSDIS Land Processes DAAC. https://doi.org/10.5067/MODIS/MOD13A3.006.Accessed08August2019
Dong X, Chu Y, Gu X, Huang Q, Zhang J, Bai W (2019) Suitable habitat prediction of Sichuan snub-nosed monkeys (Rhinopithecus roxellana) and its implications for conservation in Baihe nature reserve, Sichuan, China. Environ Sci Pollut Res 26:32374–32384. https://doi.org/10.1007/s11356-019-06369-3
Elith J, Leathwick JR (2009) Species distribution models: ecological explanation and prediction across space and time. Ann Rev Ecol Evol Syst 40:677–697. https://doi.org/10.1146/annurev.ecolsys.110308.120159
Elith J, Graham CH, Anderson RP et al (2006) Novel methods improve prediction of species’ distributions from occurrence data. Ecography 29:129–151
Elith J, Phillips SJ, Hastie T, Dudik M, Chee YE, Yates CJ (2011) A statistical explanation of max-ent for ecologists. Divers Distrib 1:43–57. https://doi.org/10.1111/j.1472-4642.2010.00725.x
Fick SE, Hijmans RJ (2017) WorldClim 2: new 1 km spatial resolution climate surfaces for global land areas. Int Jour Climatol 37:4302–4315. https://doi.org/10.1002/joc.5086
Fox AD, Mitchell C, Stewart A, Fletcher JD, Turner JV, Boyd H, Shimmings P, Salmon DG, Haines WG, Tomlinson C (1994) Winter movements and site fidelity of pink-footed geese Anser brachyrhynchus ringed in Britain, with particular emphasis on those marked in Lancashire. Bird Study 41:221–234. https://doi.org/10.1080/00063659409477222
Gaudreau J, Perez L, Harati S (2018) Towards modelling future trends of quebec’s boreal birds’ species distribution under climate change. Int J Geo-Inf 7:335. https://doi.org/10.3390/ijgi7090335
Golterman HL (1975) Physiological limnology: an approach to the physiology of lake ecosystem (eds). Developments in water science series. Elsevier Publishing company. https://doi.org/10.1016/s0167-5648(08)71058-X
Gould SF, Beeton NJ, Harris RM, Hutchinson MF, Lechner AM, Porfirio LL, Mackey BG (2014) A tool for simulating and communicating uncertainty when modelling species distributions under future climates. Ecol Evol 4:4798–4811. https://doi.org/10.1002/ece3.1319
Gschweng M, Kalko EKV, Berthold P, Fiedler W, Fahr J (2012) Multi-temporal distribution modelling with satellite tracking data: predicting responses of a long distance migrant to changing environmental conditions. J App Ecol 49:803–813. https://doi.org/10.1111/j.1365-2664.2012.02170.x
Hall JC, Chhangani AK, Waite TA, Hamilton IM (2012) The impacts of La Niña-induced drought on Indian vulture Gyps indicus populations in western Rajasthan. Bird Conserv Int 22:247–259. https://doi.org/10.1017/S0959270911000232
Harris I, Osborn TJ, Jones PD, Lister DH (2020) Version 4 of the CRU TS monthly high-resolution gridded multivariate climate dataset. Sci Data 7:109. https://worldclim.org/data/monthlywth.html
Herrero J, Garcia-Serrano A, Couto S, Ortuno V, Garcia-Gonzalez R (2006) Diet of wild boar Sus scrofa L. and crop damage in an intensive agroecosystem. Eur J Wildl Res 52:245–250. https://doi.org/10.1007/s10344-006-0045-3
Hernandez PA, Graham CH, Master LL, Albert DL (2006) The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 29:773–785. https://doi.org/10.1111/j.0906-7590.2006.04700.x
Hirzel AH, Le Laya G, Helfera V, Randina C, Guisan A (2006) Evaluating the ability of habitat suitability models to predict species presences. Ecol Model 199:142–152. https://doi.org/10.1016/j.ecolmodel.2006.05.017
iNaturalist users, Ueda K (2020) iNaturalist Research-grade Observations. iNaturalist.org. Occurrence dataset. https://doi.org/10.15468/ab3s5x. Accessed 23 Oct 2020
IPCC (Intergovernmental Panel on Climate Change) (2021) Summary for policymakers. In: Zhou B (ed) MassonDelmotte V, Zhai P, Pirani A, Connors SL, Pe´an C, Berger S, Caud N, Chen Y, Goldfarb L, Gomis MI, Huang M, Leitzell K, Lonnoy E, Matthews JBR, Maycock TK, Waterfield T, Yelek O¸ Yu R. Climate Change 2021: The physical science basis Contribution of Working I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge
Jha KK (2015) Distribution of vultures in Uttar Pradesh, India. J Threat Taxa 7:6750–6763. https://doi.org/10.11609/JoTT.o3319.6750-63
Jha R, Jha KK (2021) Habitat prediction modelling for vulture conservation in Gangetic-Thar-Deccan region of India. Environ Monit Assess 193:532. https://doi.org/10.1007/s10661-021-09323-4
Jha R, Jha KK (2024) Projected future of Slender-billed Vulture: habitat distribution modelling and population study in northern India. Biologia 79:223–238. https://doi.org/10.1007/s11756-023-01523-2
Jha KK, Jha R, Campbell MO (2021) The distribution, nesting habits and status of threatened vulture species in protected areas of central India. Ecol Ques 32:7–22. https://doi.org/10.12775/EQ.2021.20
Jha R, Kanaujia A, Jha KK (2022) Wintering habitat modelling for conservation of Eurasian vultures in northern India. Nova Geod. https://doi.org/10.55779/ng2122
Jha R (2022) Sociocultural aspects, spatial distribution, decadal change in populations and impact of climate crisis on habitats of Uttar Pradesh. In: Ph D dissertation, University of Lucknow, India
Jiménez-Valverde A (2020) Sample size for the evaluation of presence-absence models. Ecol Ind 114:106289
Kaky E, Nolan V, Alatawi A, Gilbert F (2020) A comparison between ensemble and MaxEnt species distribution modelling approaches for conservation: a case study with Egyptian medicinal plants. Ecol Inf 60:101150. https://doi.org/10.1016/j.ecoinf.2020.101150
Kang J-H, Hyun B-R, Kim IK, Lee H, Lee J-K, Hwang H-S, Eom T-K, Rhim SJ (2019) Movement and home range of cinereous vulture Aegypius monachus during the wintering and summering periods in east Asia. Turk J Zool 43(3):7. https://doi.org/10.3906/zoo-1807-3
Konatowska M, Młynarczyk A, Kowalewski W, Rutkowski P (2023) NDVI as a potential tool for forecasting changes in geographical range of sycamore (Acer pseudoplatanus L.). Sci Rep 13:19818. https://doi.org/10.1038/s41598-023-46301-x
Kumar S, Stohlgren TJ (2009) MaxEnt modeling for predicting suitable habitat for threatened and endangered tree Canacomyrica monticola in New Caledonia. J Ecol Nat Environ 1:94‒98. https://academicjournals.org/journal/JENE/article-full-text-pdf/C1CDB822968
Lane J-E (2018) Climate crisis and the “We”: An essay in deconstruction. IJMSR 6:34‒43. https://www.arcjournals.org/international-journal-of-managerial-studies-and-research/volume-6-issue-7/4
Liang J, Peng Y, Zhu Z, Li X, Xing W, Li X, Yan M, Yuan Y (2021) Impacts of changing climate on the distribution of migratory birds in China: Habitat change and population centroid shift. Ecol Ind 127:107729. https://doi.org/10.1016/j.ecolind.2021.107729
Liminana R, Soutullo A, Arroyo B, Urios V (2012) Protected areas do not fulfil the wintering habitat needs of the trans-Saharan migratory Montagu’s Harrier. Biol Conserv 145:62–69. https://doi.org/10.1016/j.biocon.2011.10.009
Lobo JM, Jimenez-Valverde A, Real R (2008) AUC: a misleading measure of the performance of predictive distribution models. Glob Ecol Biogeogr 17:145–151. https://doi.org/10.1111/j.1466-8238.2007.00358.x
Manning MR, Edmonds J, Emori S, Grubler A, Hibbard K, Joos F, Kainuma M, Keeling RF, Kram T, Manning AC, Meinshausen M, Moss R, Nakicenovic N, Riahi K, Rose SK, Smith S, Swart R, van Vuuren DP (2010) Misrepresentation of the IPCC CO2 emission scenarios. Nat Geosci 3:376–377. https://doi.org/10.1038/ngeo880
Manzoor SA, Griffiths G, Lukac M (2018) Species distribution model transferability and model grain size—finer may not always be better. Sci Rep 8:7168. https://doi.org/10.1038/s41598-018-25437-1
Martínez-Ruiz M, Dykstra CR, Booms TL, Henderson MT (2023) Conservation letter: effects of global climate change on raptors. J Rapt Res 57:92–105. https://doi.org/10.3356/JRR-22-75
MoEFCC (2020) Action Plan for Vulture Conservation in India, 2020–2025. In: Ministry of Environment, Forest and Climate Change, Government of India, New Delhi
Naoroji R (2006) Birds of prey of the Indian subcontinent. Om Books International, Noida
Newton I (2003) The role of natural factors in the limitation of bird of prey numbers: a brief review of the evidence. In: Thompson DBA (ed) Birds of prey in a changing environment. Scottish Natural Heritage/The Stationary Office, Edinburgh, pp 5–23
Parmesan C, Yohe G (2003) A globally coherent fingerprint of climate change impacts across natural systems. Nature 421:37–42. https://doi.org/10.1038/nature01286
Pearce J, Ferrier S (2000) An evaluation of alternative algorithms for fitting species distribution models using logistic regression. Ecol Model 128:127–147. https://doi.org/10.1016/S0304-3800(99)00227-6
Petersen MJ, Ortiz Cano HG, Gomez T, Johnson RL, Anderson VJ, Petersen SL (2024) Maxent predictive species distribution models and model accuracy assessment for two species of Psilochalcis Kieffer (Hymenoptera: Chalcididae) occurring in the eastern great basin of Utah, USA. Diversity 16:348. https://doi.org/10.3390/d16060348
Phipps WL, Diekmann M, MacTavish LM, Mendelsohn JM, Naidoo V, Wolter K, Yarnell RW (2017) Due South: a first assessment of the potential impacts of climate change on cape vulture occurrence. Biol Conserv 210:16–25. https://doi.org/10.1016/j.biocon.2017.03.028
Ram M, Sahu A, Tikadar S, Gadhavi D, Rather TA, Jhala L, Zala Y (2022) Home range, movement and activity patterns of six vulture species using satellite telemetry in Saurashtra landscape, Gujarat, India. Ecologies 3:492–507. https://doi.org/10.3390/ecologies3040035
Salas EAL, Valdez R, Michel S (2017) Summer and winter habitat suitability of Marco Polo argali in southeastern Tajikistan: a modelling approach. Heliyon 3:e00445. https://doi.org/10.1016/j.heliyon.2017.e00445
Santangeli A, Spiegel O, Bridgeford P, Girardello M (2018) Synergistic effect of land-use and vegetation greenness on vulture nestling body condition in arid ecosystems. Sci Rep 8:13027. https://doi.org/10.1038/s41598-018-31344-2
Shen T, Yu H, Wang Y-Z (2021) Assessing the impacts of climate change and habitat suitability on the distribution and quality of medicinal plant using multiple information integration: take Gentiana rigescens as an example. Ecol Indic 123:107376. https://doi.org/10.1016/j.ecolind.2021.107376
Sullivan BL, Wood CL, Iliff MJ, Bonney RE, Fink D, Kelling S (2009) eBird: a citizen-based bird observation network in the biological sciences. Biol Conserv 142:12282–12292. https://doi.org/10.1016/j.biocon.2009.05.006
Taubert F, Fischer R, Groeneveld J, Lehmann S, Muller MS, Rodig E, Wiegand T, Huth A (2018) Global patterns of tropical forest fragmentation. Nature 554:519–534. https://doi.org/10.1038/nature25508
Tessarolo G, Ladle RJ, Lobo JM, Rangel TF, Hortal J (2021) Using maps of biogeographical ignorance to reveal the uncertainty in distributional data hidden in species distribution models. Ecography 44:1743–1755. https://doi.org/10.1111/ecog.05793
VanDerWal J, Murphy HT, Kutt AS, Perkins GC, Bateman BL, Perry JJ, Reside AE (2013) Focus on poleward shifts in species’ distribution underestimates the fingerprint of climate change. Nat Clim Chang 3:239–243. https://doi.org/10.1038/nclimate1688
Yamaç E (2007) Roosting tree selection of Cinereous Vulture Aegypius monachus in breeding season in Turkey. Podoes 2:30–36. https://www.wesca.net/Podoces/Podoces2.1/PODOCES_2(1)_Cinereous_Vulture.pdf. Accessed 17 Oct 2023
Yamaç E, Bilgin CC (2012) Post-fledging movements of cinereous vultures Aegypius monachus in Turkey revealed by GPS telemetry. Ardea 100:149–156. https://doi.org/10.5253/078.100.0206
Yong DL, Heim W, Chowdhury SU et al (2021) The state of migratory land birds in the east Asian flyway: distributions, threats, and conservation needs. Front Ecol Evol 9:613172. https://doi.org/10.3389/fevo.2021.613172
Zegarra O, Pacheco J, Pacheco V (2020) Distributional patterns of the Brazilian free-tailed Bat Tadarida brasiliensis in the Peruvian territory. Therya 11:495–507. https://doi.org/10.12933/therya-20-995
Zhang J, Jiang F, Li G, Qin W, Li S, Gao H, Cai Z, Lin G, Zhang T (2019) MaxEnt modeling for predicting the spatial distribution of three raptors in the Sanjiangyuan National Park, China. Ecol Evol 9:6643–6654. https://doi.org/10.1002/ece3.5243
Zhang P, Dong X, Grenouillet G, Lek S, Zheng Y, Chang J (2020) Species range shifts in response to climate change and human pressure for the world’s largest amphibian. Sci Tot Environ 735:139543. https://doi.org/10.1016/j.scitotenv.2020.139543
Funding
The authors did not receive support from any organisation for the submitted work.
Author information
Authors and Affiliations
Contributions
All the authors contributed to the study conception and design. Material preparation, data collection and analysis were performed by Radhika Jha and Kaushalendra Kumar Jha. The first draft of the manuscript was written by Radhika Jha, and all the authors commented on previous versions of the manuscript. All the authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Ethical approval
This is an observational study and no ethical approval is required.
Additional information
Communicated by C. T. Downs.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Jha, R., Jha, K.K. Undergoing climate change, how safe the wintering habitats of Eurasian vultures would remain: habitat modelling study in northern India. J Ornithol (2024). https://doi.org/10.1007/s10336-024-02214-8
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10336-024-02214-8