
Abstract
Ecologically and economically important obligate scavengers like vultures are under threat of extinction in the old world. Several resident and migratory vulture sites and individuals are hosted by the Gangetic-Thar-Deccan region of India with varied landscapes. The landscape is under threat from anthropogenic activities and climate change impacting the habitat. Therefore, habitat suitability of vultures was analysed using species distribution model, MaxEnt, ensemble of global circulation models (CCSM4, HadGEM2AO and MIROC5), citizen science and expert collected data. Altogether, 51 models were developed and their robustness was assessed to be good for conservation purpose (AUC range 0.719–0.906). Predicted unsuitable and suitable area categories of all vultures, resident vultures and migratory vultures were identified for the present and future years (2050 and 2070) under moderate and extreme emission scenarios (RCP 4.5 and RCP 8.5). The short-term and long-term area suitability change varied between 1 and 3%. Area suitability differences were also noticed among larger (global) and smaller (local) geographical areas. The bioenvironmental parameters (land use, land cover and human footprint) played a major role in habitat determination in the current scenario. Bioclimatic factors, like precipitation parameters (precipitation seasonality bio 15 and annual precipitation bio12) and temperature parameters (isothermality bio 3 and temperature seasonality bio04), were the main model determining covariates for future prediction. An earlier hypothesis of higher suitability of forest and lower suitability of agriculture area tested in this study stood modified. Implications of the results are discussed, and conservation strategies are suggested with an advice of global strategy and local execution.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Vultures, the most extinction-prone avian group in the world, are obligate scavengers and play an essential role in recycling carrion in terrestrial ecosystems (Şekercioğlu et al., 2004; Straub et al., 2015). Out of nine old world vultures found in India, five resident vultures are critically endangered, one endangered and one in the not-threatened category. Two migratory vultures are in the not-threatened and one in the least-concerned category, thus making the residents a greater cause of concern than the migrants. However, the population trend for all these, except two migratory griffons is decreasing (IUCN, 2020), including those with a massive decline in the past reported by Prakash et al. (2003). Therefore, they are expected to pose severe ecological and socio-economic implications (Pain et al., 2003; Markandya et al., 2008). In the past, they have faced survival crises in all the three continents where they are dominant. In Africa, they are known to suffer direct poisoning, while in Asia, in particular in the Indian subcontinent, indirect poisoning and other anthropogenic activities have led to a change in habitat. Despite that, the role of altering habitat and its impact on their conservation has not been explored sufficiently.
The habitat of vultures could be defined as a 3-dimensional space in which this avian species can survive, grow and breed well or complete its life cycle successfully (Liu et al., 2017). This space comprises the biotic and abiotic components having interactive functions among themselves. A combination of feed-, roost- and nest-linked covariates decide the suitability of the habitat. Such covariates could be trees in the forest, cliffs in mountains, ungulate population, water bodies, countryside, temperature, precipitation, anthropogenic activities, etc. A suitable habitat is known to promote the survival of wild animals (Wu et al., 2016) while habitat degradation is regarded as one of the most important factors in the loss of biodiversity (Jiao et al., 2016). Additionally, climate change has been identified as a major cause of habitat degradation or loss (Newbold et al., 2015; Liu et al., 2017). Global climate change in association with anthropogenic factors has influenced the decline in vulture populations (Phipps et al., 2017; Anoop et al., 2020).
However, vultures have been seen to adapt well in forests, mountains and agricultural fields for roosting, breeding and foraging depending on the species. They are known to occupy a large landscape with varied land cover combination. It has been seen that vultures have preferences among different types of land cover (Jha & Jha, 2020, 2021). Therefore, it should be interesting to know their acceptance of a combination of different categories of land use land cover (LULC) forming their habitat or more specifically their niche. Predicted effects of climate change across the landscape on the future distribution of habitats for the species, could help land managers mitigate any potential threats to their habitats (Peterson et al., 2008; Zhang et al., 2019).
Habitat projection can be done using ecological niche modelling, a tool for identifying geographic and ecological areas suitable for species persistence based on the environmental variables of known occurrence sites (Phillips & Elith, 2010). It has the potential to fill the knowledge gap regarding species distributions (Lewison et al., 2012; Mateo et al., 2013) and provide an insight into the impacts of climate change (Peterson, 2006; Ingenloff, 2017). Ecological niche or species distribution models provide immense opportunities to identify suitable habitats for current and future climate change scenarios (Osborne & Seddon, 2012; D’Elia et al., 2015). Furthermore, there is a growing interest in utilizing niche models in spatial conservation planning for vultures in different parts of the world (D’Elia et al., 2015; Phipps et al., 2017). Maximum entropy algorithm (MaxEnt) has been abundantly used in recent years, owing to its efficiency for such modelling in raptors including vulture species (Ilanloo et al., 2020; Ortiz-Urbina et al., 2020; Saenz-Jimenez et al., 2020).
Although India remains a stronghold of vultures in the Asian region, the Gangetic-Thar-Deccan region (GTDR) within the country supports a bulk of their population, migratory as well as residents (Jha, 2018). GTDR is a mosaic of agriculture, forest and aquatic landscapes with varied climatic conditions. With this context in view, we aimed to study the habitats of vultures as a group, including resident and migratory in GDTR, using MaxEnt modelling along the following lines: (1) estimating the current expanse of habitat with suitability classes, (2) predicting potential future habitat in the short and long term, (3) identifying vulnerable areas and determining loss and gain due to climate impact and (4) finding out variation in global and local scale modelling.
Materials and methods
Study area
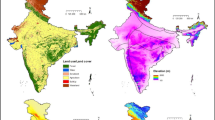
The Gangetic-Thar-Deccan region (GTDR) of India is composed of vast swathes of moist and dry plains and hills and plateau land supporting a major part of the vulture population in the country. Three selected provinces (Fig. 1), the administrative units forming this region, are Uttar Pradesh (UP, Gangetic plain), Rajasthan (RAJ, Thar desert) and Madhya Pradesh (MP, Deccan trap). They were chosen on account of varying LULC, different geoclimatic features and the high abundance of vultures (Kale, 2014; Jha, 2018). Although all the three provinces have significant agriculture land, MP also has a sizable expanse of forests. Rajasthan and UP have low forest cover with dominating agriculture land use but the former is dryer while the latter is wetter. On the basis of expanse (Table 1) and with vulture-preferred LULC (Jha & Jha, 2020) in mind, the landscape of these states may be categorised as forest-agriculture-water (MP), agriculture-forest-water (RAJ) and agriculture-water-forest (UP). The average temperature also varies in these states. Figure 2 and Table 1 provide LULC and other environmental features. The livestock population, an indicator of potential food availability (cattle, buffalo, sheep, goat, pig, etc.), is the highest in UP followed by RAJ and MP.

Vulture site clouds (red) in the study area with different land use land cover background in Rajasthan (top left), Uttar Pradesh (top right) and Madhya Pradesh (bottom). Inset projecting the location of study area in India

Climatic and altitudinal variation and existing land use land cover features of the study area providing niche to the vultures. Shrubland, agriculture and forest dominance may be noted in Rajasthan, Uttar Pradesh and Madhya Pradesh, respectively
Occurrence data collection
Species occurrence data used in this study was sourced from field surveys using the synchronised counting method in UP in 2010–2011 and in MP in 2016 and a transect survey in RAJ in 2017 (Jha, 2015, 2017; Jha et al., 2021). A single transect survey was not considered to be a good representative of a large state like RAJ, so it was supplemented with data from published literature (Chhangani, 2009; Saran & Purohit, 2012). Some records in published literature lacked geocoordinates. To remedy this, Google Earth™ was used for georeferencing such locations as suggested by Ahmad et al. (2019). Since quantity of data is considered more important than spatial biases (Gaul et al., 2020), citizen science data was also included. This brought the benefit of historical record to the study, which provides better prediction of range shift (Jackson et al., 2015; Carvalho et al., 2020). Citizen science repositories like e-bird (http://www.ebird.org) and i-naturalist (http://www.inaturalist.org) were used for collecting long-range historical observations in both time and space. Biases such as repeat observations at popular sites, concentrated occurrence in favoured regions as well as latitudinal bias were rectified using advanced tools of duplicate removal, occurrence rarefication and bias file preparation. Finally, vulture sighting records used in the study were temporo-spatially distributed over a study area of 892,190 km2 and a span of three decades (1990–2020).
Vultures’ grouping
The occurrence records containing data of eight species of vultures found across the study area were grouped as per the management strategy of the Forest Department, which tends to focus on vultures as a single group, not species wise, for ease of management. However, due to differing ecological requirements, it is more relevant to group vultures as per their residency period. The resident species need intensive care around the year including the breeding and fledgling periods while migratory vultures require simple care for a maximum of five wintering months per year. Keeping in mind these facts, they were grouped into three categories: resident vultures (Egyptian, Indian, red-headed, slender-billed and white-rumped), migratory vultures (Cinereous, Himalayan Griffon and Eurasian Griffon) and all vultures combined. Although there exists the possibility of interspecific competition due to species differences influencing the species distribution (Jackson, 1981; Baah-Acheamfour et al., 2017), such groupings were suggested/followed by others ( Holland et al., 2019; Genero et al., 2020), considering their sympatric nature, low abundance of migratory vultures in the entire population and partial temporal sharing of landscape, higher carrying capacity of available habitat, mutually exclusive niche and ecological plasticity in some species.
Environmental parameters
Environmental covariates chosen for the study was LULC providing shelter and foraging grounds. Land cover data was procured as MODIS product (https://earthexplorer.usgs.gov/), which was created using supervised classification for global maps of land cover at annual time steps and 500-m spatial resolution for 2001–present (Friedl & Sulla-Menashe, 2019). Seventeen land cover classes were reduced to broader six classes (forests, water, agriculture, built up area, wasteland and shrubland) as the present study did not demand details like different types of forests, grassland, Savannah and agriculture land. Normalised differential vegetation index (NDVI) was used as a proxy to ungulate forage availability for vultures (Santangeli et al., 2018). Human Footprint (HFP) was used as a proxy for human disturbance or pressure on natural systems (Di Marco & Santini, 2015; Gallardo et al., 2015; Newbold et al., 2015; Venter et al., 2016a). Elevation and slope were also used since cliff dwelling vultures prefer lower elevation, steep slope and western aspect (Liberatori & Penteriani, 2001; Marinković et al., 2012).
Habitat projection models
For habitat suitability projection a species distribution model (SDM), MaxEnt was chosen due to its high popularity on account of various advantages reviewed in Jha and Jha (2021). There are several global circulation models (GCM) available for the projection of future scenarios. While all these are understood to project accurate scenarios, there is no consensus among them. Hence, ensemble modelling which is recommended in other studies for accuracy (Sutton et al., 2015; Ashrafzadeh et al., 2019), was used by selecting CCSM4 (Ashrafzadeh et al., 2019), HadGEM2AO (Ahmad et al., 2019) and MIROC5 (Sony et al., 2018). Two RCPs, viz., RCP4.5 and RCP8.5 (Arumoogum et al., 2019), for these GCMs were chosen for this study.
Sample and covariates’ layer preparation
A series of conversion of raw data obtained from various sources, their use in MaxEnt model and the outputs are summarised in Fig. 3. The inputs for the MaxEnt model were (i) bioclimatic and bioenvironmental layers and (ii) sample or presence records in the form of coordinate data. Duplicate sites were removed from the occurrence list in Microsoft Excel using the remove duplicates tool. Multiple presence points obtained from various sources (19,660 vultures, 16,838 residents, 2822 migrants) were reduced to 4854, 4726 and 561, respectively, after duplicate removal. Finally, such unique sample records were rarefied using SDM toolbox (spatially rarefy occurrence data) and fixing rarefying resolution to 4 km keeping in view shelter sites defined in Jha et al. (2020). The final sample layer consisted of 3291 (all vultures), 3191 (residents) and 303 (migrants) unique occurrence points.

Global and local models
Individual provinces are the administrative units of vulture management in India as the Forest Department of each province is responsible for the protection and conservation of vultures in their jurisdiction. The global projection of suitable area differs from local projections (Jha & Jha, 2021, unpublished). Therefore, two different types of models, global and local, were generated to analyse the difference in global and local suitability of habitat. Global models were generated for the whole study area or all three select provinces together. Local models were also developed for individual provinces separately using the same covariates. The bioenvironmental models included climatic as well as other predictors (LULC, HFP, NDVI, elevation and aspect) while bioclimatic models included only climatic predictors (precipitation and temperature variants, viz. bio01 to bio19).
Habitat suitability and mapping
The MaxEnt index (0–1) was reclassified into four suitability classes, e.g., unsuitable (0.0–0.25), low (0.25–0.50), moderate (0.50–0.75) and high (0.75–1.00), as used for raptors by Zhang et al. (2019). The reclassified maps were used to calculate the area under these classes. For assessing vulnerable area changes in the future, i.e., area loss from good (suitable) to bad (unsuitable) area and area gain from bad to good area, the above classification was slightly modified. The unsuitable area was kept unchanged but low, moderate and high suitability classes were merged into one group resulting onto only two classes, unsuitable and suitable. Using the second set of reclassified maps, the present-day scenario was compared to different future projections. The resultant maps had four different classes viz., unsuitable, suitable (both remaining stable), loss and gain (the area undergoing a change). Reclassification and area calculation were performed on the MaxEnt heatmaps using ArcGIS 10.5. The same software along with Microsoft Excel was used for area and other calculations.
Results
Model robustness
Based on the Pearson coefficient set at ±0.8, the 10 bioclimatic and six bioenvironmental non-colinear variables (bio01 annual mean temperature, bio02 mean diurnal range (mean of monthly (max temp–min temp)), bio03 Isothermality (bio2/bio7) (×100), bio04 temperature seasonality (standard deviation × 100), bio09 mean temperature of driest quarter, bio12 annual precipitation, bio14 precipitation of driest month, bio15 precipitation seasonality (coefficient of variation), bio17 precipitation of driest quarter, bio18 precipitation of warmest quarter, LULC (forest, water, built-up, agriculture, wasteland, scrubland), NDVI (January, June), elevation, HFP and aspect) were extracted for generating the models. Fourteen global models, each for the three groups, all vultures, resident vultures and migrant vultures, were developed. These fourteen models were three GCMs (CCSM4, HadGEM and MIROC) for two RCPs (4.5 and 8.5) and two terms (2050 and 2070) in addition to current bioclimatic and bioenvironmental models for each group. The range of area under curve (AUC) for these three groups of vultures were 0.719–0.790 (0.726 ± 0.017), 0.718–0.796 (0.728 ± 0.019) and 0.852–0.906 (0.865 ± 0.011), respectively. Similarly, nine local models of current environmental scenarios (3 provinces and 3 groups) were developed. Their AUC ranged between 0.793 and 0.940, where the average AUC of all vultures, residents and migrants was 0.838 ± 0.034, 0.845 ± 0.031 and 0.932 ± 0.011, respectively.
Variable contribution
The models of current habitat prediction reflected a higher importance of bioenvironmental variables than simply bioclimatic variables. This was the case with three out of the top six variables that were considered. HFP, LULC and elevation remained the top three contributors for all vultures and resident vultures and among the top five contributors for migratory vultures. The quantitative values of these three contributors were 66% (all vultures), 67% (residents) and 58% (migratory), respectively. However, the first two variables (HFP and LULC) had the lion’s share, with 57%, 59% and 51%, respectively. Among the ten non-colinear bioclimatic variables, the top three (bio03, bio15, bio04 or bio12) contributed 60%, 57% and 54% and the top five (bio03, bio15, bio04, bio12, bio14 or bio02) contributed 85%, 82% and 77% for all vultures, resident vultures and migratory vultures, respectively. The range values (Hijmans et al., 2005) of the current bioclimatic components in the study area were 6.55–18.50 °C (bio02; mean diurnal range of temperature), 36.16–54.29 °C (bio03; isothermality), 361.67–818.56 °C (bio04; temperature seasonality), 104–2124 mm (bio12; annual precipitation), 0–12 mm (bio14; precipitation of driest month) and 103.61–175.05 mm (bio15; precipitation seasonality). Among future scenarios (where bioenvironmental variables were not considered) the top five climatic variables were bio03, bio04, bio15, bio14 and bio18 or bio12 for all vultures; bio03, bio15, bio04, bio12 and bio14 or bio18 for residents; while bio12, bio04, bio15, bio03 and bio18 or bio02 for migrants.
For the nine models, which focused on local prediction, LULC emerged as the most important variable (21.3% for all vultures, 21.2% for residents and 30.03% for migrants). The other most important variable was HFP whose contribution was at par with LULC for all vultures (21.2%) and residents (21.8%). However, for migrants, HFP (6.26%) was displaced to the sixth spot by climatic factors (bio18, bio03, bio09 and bio15). Response curves of these most influential variables and Jackknife bar charts showing relative importance of variables are presented in Figs. 4 and 5.

Response bars and curves of influential bioenvironmental and bioclimatic variables in current scenario for different vulture groups in respective columns

Relative contribution of different variables in the current habitat prediction of all vultures, residents and migrants (top to bottom in two columns) under bioenvironmental (left) and bioclimatic (right) scenarios
Habitat suitability
The suitability area projected by three different GCMs varied and were therefore averaged. Such ensemble area under different suitability categories and its delineation in the study area under different scenarios is presented in Table 2 and Figs. 6 and 7, respectively. Maps of future scenarios are presented in Figs. 8 and 9. The breakup of the total area studied (892,190 km2), varied among different suitability categories, vulture groups and time period. Under the current environment scenario, suitable area for all vultures was 56%, while it was 82% under the current climate-only scenario. In the short-term and long-term scenario, the suitable area ranged from 80 to 81% which was marginally lower. For the resident category, in the current scenario, environment versus climatic trend remained similar to all vultures but the area expanse was found to have decreased by 2%. Suitable area was 54% for the environmental scenario versus 80% for the only-climate scenario. The short term and long term suitable area for residents ranged between 79 and 82%. For the migratory group, suitable area in the environment and climate scenario was highly reduced as compared to other groups, 18% and 34%, respectively. In the short-term and long-term scenarios, this area ranged between 33 and 34%.

Maps showing different categories of current habitat suitability for all vulture species combined under two sets of covariates. Difference in suitability area may be noted in bioclimatic (left) and bioenvironmental (right) scenario, latter showing higher unsuitable area

Maps showing different categories of current habitat suitability for resident and migrant species combined under two sets of covariates. Differences in suitability areas may be noted in residents versus migrants (left and right) as well as bioclimatic versus bioenvironmental (top and bottom) scenarios, the latter showing higher unsuitable area in both the cases

Maps showing suitability area categories of resident vultures in medium (RCP 4.5) and high (RCP 8.5) emission scenarios for short term (2050) and long term (2070) periods. Although minor, but a significant area change under different categories and emission could be noticed minutely. This may be correlated with Table 2

Maps showing suitability area categories of migrant vultures in medium (RCP 4.5) and high (RCP 8.5) emission scenarios for short term (2050) and long term (2070) periods. Although minor, but a significant area change under different categories and emission could be noticed minutely. This may be correlated with Table 2
However, looking at the area change from current expanse under different emission and term scenarios in different vulture groups indicated a mixed trend. The unsuitable area of all vultures increased from the present for both the terms in moderate emission scenario, but in the extreme emission scenario, this increased in the short term and decreased in the long term. Resident vultures showed reverse pattern in moderate emission but followed a similar pattern in extreme emission. The migrant vultures exhibited a clear pattern of decrease in moderate emission and increase in extreme emission in both the terms.
The current stable habitat area (unsuitable and suitable) is predicted to change partially in the future. The unstable area (from suitable to unsuitable, i.e., loss and vice versa, i.e., gain) is also predicted to undergo change in the short- and long-term future under two emission scenarios. This is presented quantitatively in Table 3 and pictorially represented in Figs. 10 and 11. While a linear trend in the net change in area may not be evident, it is clear that large swathes of area will undergo change in habitat suitability in the future.

Maps showing suitability area change (gain and loss) under medium (RCP 4.5) and high (RCP 8.5) emission scenarios for short term period (2050) in resident (upper row) and migrant (lower row) vultures. It may be noted that habitat of migrant vultures suffers more changes under different emission conditions

Maps showing suitability area change (gain and loss) under medium (RCP 4.5) and high (RCP 8.5) emission scenarios for short-term period (2070) in resident (upper row) and migrant (lower row) vultures. It may be noted that habitat of migrant vultures suffers more changes under different emission conditions
However, when we looked at the net change in unstable area, the trend varied in different groups and scenarios. For all vultures, there will be loss in the moderate scenario in both the long and short term, but in the extreme scenario, there will be loss in the short term and gain in the long term. For resident vultures, there will be gain in both the terms in moderate emission but loss in the short term and gain in the long term in extreme emission. For the migratory group, there will be gain in moderate emission and loss in extreme emission in both terms.
Land use, land cover and area suitability
Major land cover types available for shelter, foraging and water requirement of vultures were agriculture land, forest and water bodies (Table 1). The total area of forests in the studied landscape is 131,070 km2, and the agriculture area is 533,240 km2, almost four times that of the former (ISFR, 2017). The present records of vultures showed that they favoured forests, agriculture and the vicinity of water bodies. As per the output of the model, forests and water bodies played a major role while agriculture played a minor role in determining suitable area. Habitat suitability (HS) in the whole study area for all vultures showed that 95% of the forest area and only 53% of the agriculture area were suitable. The percentage suitability showed a similar trend favouring forest areas when the three states (which have forests, agriculture areas and water bodies in different proportions) were separated from within the model. It was found that arid landscape (RAJ) had lower suitability in both forest and agricultural land (68% forest and 45% agriculture) in relation to the relatively moist landscapes of UP (96% forests and 54% agriculture) and MP (97% forests and 59% agriculture). Resident vultures, they had similar patterns but the quantum was marginally lower in all the cases by 1 to 6% in forest and 0% to 4% in agriculture. However, in the case of migratory vultures, area suitability was very low in all the cases, such as only 69% of forest and 5% of suitable agriculture area in the whole landscape under study. In each province within the model, HS varied between 50–78% (forests) and 4–8% (agriculture). The quantum of HS was found to be different when the province range was considered on a local basis i.e., independently run model per province (different from the global above). This range was much reduced and was recorded at 47–94% (forests) and 27–43% (agriculture) for all vultures. The area for resident vultures was reduced from the all vultures model by 0–5% (forest) and 0–4% (agriculture). Similarly, the suitability range for migrants was further reduced from residents by 17–32% (forest) and 20–34% (agriculture). Figures 12 and 13 represent the comparison of global versus local projection of habitat suitability in resident and migrant vultures as a group.

Maps showing suitability area categories in current bioenvironmental scenario of whole study area based on all presence records and bioenvironmental covariates of large area (global projection: top left) for resident vultures. Remaining maps are of its three territorial components based on smaller and individual state (local) with its own bioenvironmental covariates. Expanse of suitability area of individual state (Uttar Pradesh: top right, Madhya Pradesh: bottom right and Rajasthan: bottom left) may be compared with corresponding component in global projection. Minute observation reflects higher unsuitable area in local models

Maps showing suitability area categories in current bioenvironmental scenario of whole study area based on all presence records and bioenvironmental covariates of large area (global projection: top left) for migrant vultures. Remaining maps are of its three territorial components based on smaller and individual state (local) with its own bioenvironmental covariates. Expanse of suitability area of individual state (Uttar Pradesh: top right, Madhya Pradesh: bottom right and Rajasthan: bottom left) may be compared with corresponding component in global projection. Minute observation reflects higher unsuitable area in local models
Discussion
Occurrence and LULC data sources
Occurrence data based on a survey by the authors was limited temporally and spatially. Any model requires data that span over time and space to yield better prediction with higher accuracy. Therefore, to cover the gaps of presence data in the present study, citizen science was employed as advocated by Milanesi et al. (2020) who assert that citizen science data can be correctly used to develop SDMs with high predictive accuracy or at least improved fit (Roy-Dufresne et al., 2019). The citizen science data repositories provided a high number of occurrences over a large area and time span (Ramesh et al., 2017). Though such data were opportunistic and bias prone, advance tools and statistical technology were used (Jackson et al., 2015; Croft et al., 2019) for cleanup. Removal of duplicates and spatial rarefication were useful for reducing the potential disadvantages of citizen science data. The use of published literature (McDonald et al., 2019) about vulture occurrence data collected by experts and our field survey in the study area were also added to the samples to offset above weakness in citizen science data to some extent. This has been done in earlier work for use in distribution modelling (Corovic et al., 2018).
Open source LULC data provided by NASA-LPDAAC (Friedl & Sulla-Menashe, 2019) was used in our study since locally generated data by the National Remote Sensing Centre (NRSC), Indian Space Research Organisation exclusively for India is not freely accessible. The global data had some limitations as compared to NRSC as its spatial resolution was inferior and it lacked comprehensive ground truthing on a level same as that of NRSC. Moreover, this data also had finely divided classes, which, being broader in nature, were not required for the present study. The reclassification of LULC (17 classes merged to six) was good enough for use in vulture habitat prediction.
Model evaluation
There are a few model performance indicators out of which we used most popular AUC that quantified the significance of the curve and its values to determine model accuracy (Hameed et al., 2020). The local models (AUC 0.838–0.932) were marginally improved over global models (AUC 0.718–0.728). Within global models, different vulture groups showed varied performance indicating a successive improvement trend viz., all vultures (0.726) < resident vultures (0.728) < migratory vultures (0.865). The performance of these models appeared satisfactory, since a model with AUC value closer to 1 is considered to be a perfect performer while 0.5 is seen as no better than random (Phillips et al., 2006; Bosch et al., 2014). However, the lowest (0.718) and highest (0.932) AUC fell in the good and very good categories as per model evaluation classification (models: very good = AUC > 0.9; good = AUC 0.7–0.9 and uninformative = AUC < 0.7) suggested by Baldwin (2009). Thus, the robustness of our models was useful in achieving the conservation objective. This is because the lower end AUC values of good model prediction category are well accepted (Songer et al., 2012; Bosch et al., 2014) and considered suitable for conservation planning (Zeng et al., 2015).
As for the quality of models, the comparison of prediction variation in habitat area expanse of bioclimatic and bioenvironmental models is pertinent at this point. Our current models predicted results with LULC and without LULC which differed by 24% with the former being lower. This could be attributed to the fact that the climatic umbrella is generally larger than the environmental one due to the specific requirement of a niche, e.g., trees/cliffs, water and ungulate/cattle concentration (Jha and Jha 2021). This implied that the actual suitable habitat area was an overestimation in the case of bioclimatic models. This is of significance to future models since they are all climatic and could be considered an overestimation. Preston et al. (2008) argued that this was usually the case with most distribution models predicting species responses to climate change, which included climate variables and, rarely, the biotic interactions.
Habitat drivers
Both bioclimatic and bioenvironmental covariates played their role in habitat expanse determination in the present study, but a few environmental parameters contributed more than the dominant climatic factors. This is in concurrence with the results of Freeman et al. (2019) suggesting that forest cover, rather than climate, was the major driver of the forest dwelling guineafowl’s current distribution. Herrero et al. (2006) also made a similar deduction stating that vegetation cover influenced the distribution of an animal species more than any other factor since it determines the ability of the land to supply food and/or shelter for animals. However, when the interaction of vultures with only bioclimatic factors was modelled, it returned a much larger suitable habitat area than with bioenvironmental factors. This was similar to Jha and Jha (2021) but in contradiction to Gama et al. (2015). The results remained the same even when the vultures were grouped into resident and migrant categories. The role of different components of LULC, an important component of environmental factors, in model predictions differed from earlier findings of Jha and Jha (2020, 2021). They predicted forest and water body as the most important covariates for habitat suitability with built-up area being less important. In contradiction to this, our study found built-up area to be at the top while forest and water body played second and third fiddle. This could possibly be due to the presence of a much larger expanse of built-up area and a larger population of eco-plastic species of vulture in the present study area.
However, HFP, another component of environmental predictors, was of the highest importance in habitat composition of vultures, even above LULC (built-up, forest, water body, etc.). This may be due to Venter et al. (2016b) model of HFP, which includes the extent of built environments, other than cropland, pastureland, human population density, nighttime lights, railways, roads and navigable waterways, which are part of both favourable and/or unfavourable environment. Though vultures are known to be affected negatively by anthropogenic disturbances (Chomba & M’Simuko, 2013; Arkumarev et al., 2018), some prefer to set up colonies in the vicinity of human habitation (Jha, 2015; Henriques et al., 2018) and some embrace rural garbage dumps and surrounding area (McGrady et al., 2018; Bahadur et al., 2019). Interestingly, the total percentage contribution of HFP and LULC in model prediction in the present study was almost similar to only LULC in Jha and Jha (2020)’s study. This could be speculated to be a result of the duplication and distributed weightage of some factors in the group of categorical parameters like HFP and LULC. It is interesting to note the hypothesis of Cable et al. (2021) in this regard. They state that in the case of cave dwelling and tree harbouring bats, landscape suitability improving factors included limited agriculture, more forest cover, forest edge, proximity to medium-sized water bodies, lower elevations and limited urban development. This is pertinent to our case where field verification and expert consultation revealed that the most suitable habitat for vultures is forested landscape with interspersed water bodies since it can provide both shelter and opportunity for foraging. Contrastingly, agriculture was found to be less suitable since this provided mainly foraging and very little shelter opportunity on small trees. Ramesh et al. (2011) and Navaneethan et al. (2015) had also reported very low sighting of vultures in agriculture areas as compared to different types of forests and scrubland.
Other than physical features (LULC), which provide shelter and water, climate components also played an important role in habitat determination of vultures. This corresponds with the affirmation of Bosch et al. (2014) that temperature and precipitation have a significant influence on the distribution of terrestrial vertebrate fauna since these two factors synthesise the flows of energy and water in the ecosystem and substantially limit the global distribution of biodiversity. The present study suggested that the components of temperature and precipitation contribute in habitat determination. Among the top five contributors were isothermality (bio03), temperature seasonality (bio04), precipitation seasonality (bio15), annual precipitation (bio12) and precipitation of driest month (bio14) in the current climatic models. In the models for the future, the role of mean diurnal range (bio02) and precipitation of warmest quarter (bio18) could be detected in a few models. The remaining variables had very little role to play in vulture habitat delineation in the study area. However, the influence of bioclimatic variables in habitat determination, in general, has been noted in other studies on raptors (Gschweng et al., 2012; Liminana et al., 2012), although the set of covariates were not similar in different and distant localities of vultures (Phipps et al., 2017; Zhang et al., 2019; Anoop et al., 2020).
Habitat suitability was not a function of any single variable, rather it was a product of interaction among numerous covariates in different quantities (grades) in which quite a few could be following Liebig’s Law of the Minimum (Golterman, 1975). Therefore, considering a single variable in isolation may be misleading as the species choose their habitat based on the interaction of several factors. For example, dry and hot area (Thar and its adjoining locality), which is favourable for vultures, would also need to have good shelter and a water body close by. From the response curves (Fig. 4; bio12, bio03, bio15, bio04), we get an indication that vultures preferred drier areas, and their presence increased with the increase in seasonality. Interpreting the curves as per Shelford’s Law (Golterman, 1975), the optimum temperature zone of tolerance for resident vultures may be 24 to 42 °C. Higher temperatures up to 46 °C would be a zone of thermal stress, and vultures may not tolerate temperatures beyond 46 °C (zone of intolerance).
Habitat dynamics
An assessment of overall decreasing tendency in the present study in the total habitat area of resident as well as migratory vultures in the short and long term with respect to current climatic scenario, though nominal (1 to 3%), corroborated with the earlier findings of reduction in suitable area in different species (Jiménez-García & Peterson, 2019; McDonald et al., 2019), particularly in the Bearded Vulture in West Asia (Ilanloo et al., 2020) and in the Black Vulture and the Andean Condor in the new world (Saenz-Jimenez et al., 2020). However, our finding was contradicted in Phipps et al. (2017) in the Cape Vulture in Southern Africa. Such variation may be explained by the varying region-specific niche characteristic, which is driven by factors extrinsic to the species, such as the spatial distribution of climatic conditions (Lin et al., 2019). Holyoak and Heath (2016) and Kupika et al. (2018) also suggested that temperature and rainfall display complex temporal variation changing from place to place across geographies and climate variables, particularly rainfall and temperature, generally influenced habitat quantity and quality. The GTDR of India is an intensively used landscape due to urban development and economically driven land use as suggested by Teeffelen et al. (2012). Therefore, this region is more susceptible to the impact of climate change on habitat suitability, which might result in a shift in area in the future as documented in other studies (Pires et al., 2018; Trautmann, 2018).
A reduction in suitable habitat area for resident and migratory vultures was found in the three independently modelled provinces of UP, MP and RAJ under the current bioclimatic scenario as compared to the whole study area combined when modelled separately. This could be attributed to the smaller bioclimatic envelopes in the former (i.e. one single province) than the larger envelope of the three provinces combined. This was possibly due to varied range of temperature (especially isothermality, bio03 and temperature seasonality, bio04) and precipitation (especially annual precipitation, bio12 and precipitation seasonality, bio15) components in all the four unique territories. In an earlier study (Songer et al., 2012), precipitation and temperature seasonality were found to influence the habitat significantly. Additional factors which make each envelope specific are varied LULC features, such as different types and extent of forests, agricultural field, water bodies and built-up area availability. These habitat-influencing features are reported in vultures by Jha and Jha (2020) on a localised scale in Central India. Moreover, comparing old world vulture habitat suitability in Central India with India as a whole in a global study by Santangeli et al. (2019), it is evident that a global or large scale study, which is broadened or generalised in nature, has a larger suitable area whereas a small scale study, which is specialised, has smaller suitable area. This bears concordance with the present study.
A nominal decrease in the total suitable area in 50 years may not be alarming at the moment, but it can be a cause of concern in the future, keeping in view the uncertain control over increasing emissions. This may be exacerbated by rapid anthropogenic change in shelter conditions along with possible loss of mammalian biodiversity (Forister et al., 2010). New climatic conditions will change biophysical features, such as change in hydrology, vegetation shift and composition (Ravindranath et al., 2006). In addition, anthropogenic activities will also change the current LULC, for example, construction of development structures or increase in built-up area, thinning of vegetation and over exploitation of other natural resources. However, the proactive management of such areas might be one solution, if the new occurrence of a species of high conservation concern and/or climatic risk can be predicted with high certainty (Trautmann, 2018).
Management implications
Holyoak and Heath (2016) hypothesised that climate change may impact the abundance and occupancy of a population directly and indirectly through change in habitat quality, quantity and distribution and habitat dynamics. Therefore, one of the conservation requirements of biodiversity could be the precise assessment of species ranges or suitable habitat availability and knowing the distribution limiting factors to mitigate their negative impact (Fourcade et al., 2014). Our results provide the availability of different categories of suitable habitat area and unstable habitat area, which could be managed in favour of vulture conservation by adopting various strategies. Moderate and highly suitable areas which are stable even in the long run should be given top priority in the maintenance of an environment free of any anthropogenic activities which may disturb breeding and territory expansion activities. In situ conservation of resident vultures must be ensured. Such an area, especially that containing breeding or nesting colonies, should be protected by implementing strict provisions and be treated like a vulture sanctuary. Any isolated and mature trees which could be potential nesting trees for the future should not be logged off under any circumstances (Poirazidis et al., 2004; Jha & Jha, 2021). Habitat degradation must not be allowed in suitable areas. Instead, habitat improvement measures such as banning quarrying and mining, tree felling and lopping, must be implemented to provide the most favourable conditions for vulture nesting and roosting. Activities which cause disturbance such as film shooting and tourism should be highly regulated. In the not suitable areas surrounding the forests or forest fringes, landholders must be encouraged to adopt agri-horticulture and agri-silviculture with animal husbandry. This will provide medium-sized trees and domestic animals as resources for vulture outside forests. Ecologically plastic vultures (Arrondo et al., 2018; Genero et al., 2020) are known to utilise trees such as coconut, toddy palm, mango and Indian mesquite (Khejari) for nesting and roosting, and they consume smaller mammals as feed (Chhangani, 2007; Kambale, 2011). Such expansion may help migratory vultures as well since suitable area as per their requirements is already small. Unstable areas, lost and gained habitat due to climate impact, must receive timely and proper treatment either to improve or to utilise them in the future. In general, carcass disposal from Gaushala and Panjarpole (cow caring centres) and at Panchayat (community) dumping sites which are mostly found in not suitable or less suitable area predicted in the study must be coordinated to ensure safe and secure food supply (Chhangani, 2007; Saran & Purohit, 2012). Vultures must be protected from injury and death due to electricity towers and wind turbines by using reflectors (Purohit & Saran, 2013) and rescue and rehabilitation should also be ensured (Chhangani, 2009).
Conclusion
The present study also hypothesised that forest area is highly suitable as compared to agriculture area as vulture habitat. However, the agriculture area suitability in GTDR has increased manyfold in comparison to earlier studies conducted on a smaller scale (Jha, 2015; Jha & Jha, 2020). This could possibly be due to the incorporation of a higher number of foraging sites in modelling from agriculture field and built-up area collected over a long period through citizen science.
Our study provided comprehensive details of vulture habitat suitability in the present in GTDR based on species distribution modelling. It also identified area more vulnerable to climate change in the future. It further indicated that vultures can survive well in moderate to harsh climate and varied landscape from moist and dry deciduous forests to arid forest and scrubland surrounded by agriculture and agroforest area with interspersed water bodies and sufficient supply of food. We also suggested some management interventions for vulture conservation. Such management may be strategised at the global level but planned and executed at the local level.
Like any other MaxEnt climate prediction models, our models are a basic reference for predicting future habitats which may not be a close biological reality (Merow et al., 2013; Coxen et al., 2017). However, to further enhance the accuracy of model prediction, even though the models were quite strong, India-specific LULC data should be used. Since NDVI used as a proxy of food presence is not contributing much to the model prediction, possibly due to use of LULC, it may be excluded, and actual food availability (presence of ungulates and other animals) may be used to replace this parameter. Also important is the use of modelled short-term and long-term LULC for future predictions, since LULC contribution is much higher than climatic covariates.
Availability of data and material
Available in the manuscript (sources cited).
References
Ahmad, R., Khuroo, A. A., Hamid, M., Charles, B., & Rashid, I. (2019). Predicting invasion potential and niche dynamics of Parthenium hysterophorus (Congress grass) in India under projected climate change. Biodiversity and Conservation, 28(8–9), 2319–2344. https://doi.org/10.1007/s10531-019-01775-y
Anoop, N. R., Babu, S., Nagarajan, R., & Sen, S. (2020). Identifying suitable reintroduction sites for the White-rumped Vulture (Gyps bengalensis) in India’s Western Ghats using niche models and habitat requirements. Ecological Engineering, 158(2020), 106034. https://doi.org/10.1016/j.ecoleng.2020.106034
Arkumarev, V., Dobrev, V., Stoychev, S., Dobrev, D., Demerdzhiev, D., & Nikolov, S. C. (2018). Breeding performance and population trend of the Egyptian Vulture Neophron percnopterus in Bulgaria: Conservation implications. Ornis Fennica, 95, 00–00. Retrieved 09 February 2021 from https://www.researchgate.net/publication/326988690
Arrondo, E., Moleón, M., Cortés-Avizanda, A., Jiménez, J., Beja, P., Sánchez-Zapata, J. A., & Donazar, J. A. (2018). Invisible barriers: Differential sanitary regulations constrain vulture movements across country borders. Biological Conservation, 219, 46–52. https://doi.org/10.1016/j.biocon.2017.12.039
Arumoogum, N., Schoeman, M. C., & Ramdhani, S. (2019). The relative influence of abiotic and biotic factors on suitable habitat of Old World fruit bats under current and future climate scenarios. Mammalian Biology, 98, 188–200.
Ashrafzadeh, M. R., Naghipour, A. A., Haidarian, M., Kusza, S., & Pilliod, D. S. (2019). Effects of climate change on habitat and connectivity for populations of a vulnerable, endemic salamander in Iran. Global Ecology and Conservation, 19, e00637. https://doi.org/10.1016/j.gecco.2019.e00637
Baah-Acheamfour, M., Bourque, C.P.-A., Meng, F.-R., & Swift, D. E. (2017). Incorporating interspecific competition into species-distribution mapping by upward scaling of small-scale model projections to the landscape. PLoS One, 12(2), e0171487. https://doi.org/10.1371/journal.pone.0171487
Bahadur, K. C. K., Koju, N. P., Bhusal, K. P., Low, M., Ghimire, S. K., Ranabhat, R., & Panthi, S. (2019). Factors influencing the presence of the endangered Egyptian vulture Neophron percnopterus in Rukum, Nepal. Global Ecology and Conservation, 20(2019), e00727. https://doi.org/10.1016/j.gecco.2019.e00727
Baldwin, R. A. (2009). Use of maximum entropy modelling in wildlife research. Entropy, 11(4), 854–866.
Bosch, J., Mardones, F., Pérez, A., la Torre, A. D., & Muñoz, A. J. (2014). A maximum entropy model for predicting wild boar distribution in Spain. Spanish Journal of Agricultural Research, 12(4), 984–999.
Cable, A. B., O’Keefe, J. M., Deppe, J. L., Hohoff, T. C., Taylor, S. J., & Davis, M. A. (2021). Habitat suitability and connectivity modeling reveal priority areas for Indiana bat (Myotis sodalis) conservation in a complex habitat mosaic. Landscape Ecology, 36, 119–137. https://doi.org/10.1007/s10980-020-01125-2
Carvalho, J. S., Graham, B., Bocksberger, G., Maisels, F., Williamson, E.A., Wich, S., Sop, T., Amarasekaran, B., Barca, B., Barrie, A., Bergl, R .A., Boesch, C., Boesch, H., Brncic, T. M., Buys, B., Chancellor, R., Danquah, E., Doumbé, O. A., Le-Duc, S. Y., … & Kuhl, H. S. (2020). Predicting range shifts of African apes under global change scenarios. Retrieved 10 February 2021 from https://www.researchgate.net/publication/342476747 https://doi.org/10.1101/2020.06.25.168815
Chhangani, A. K. (2007). Sightings and nesting sites of Red-headed Vulture Sarcogyps calvus in Rajasthan, India. Indian Birds, 3(6), 218–221.
Chhangani, A. K. (2009). Status of vulture population in Rajasthan, India. Indian Forester, 135(2), 239–240.
Chomba, C., & M’Simuko, E. (2013). Nesting patterns of raptors; White backed vulture (Gyps africanus) and African fish eagle (Haliaeetus vocifer), in Lochinvar National Park on the Kafue Flats, Zambia. Open Journal of Ecology, 3(5), 325–330. https://doi.org/10.4236/oje.2013.35037
Corovic, J., Popovic, M., Cogalniceanu, D., Carretero, M. A., & Crnobrnja-Isailovic, J. (2018). Distribution of the meadow lizard in Europe and its realized ecological niche model. Journal of Natural History, 52(29–30), 1909–1925. https://doi.org/10.1080/00222933.2018.1502829
Coxen, C. L., Frey, J. K., Carleton, S. A., & Collins, D. P. (2017). Species distribution models for a migratory bird based on citizen science and satellite tracking data. Global Ecology and Conservation, 11, 298e311.
Croft, S., Ward, A. I., Aegerter, J. N., & Smith, G. C. (2019). Modeling current and potential distributions of mammal species using presence-only data: A case study on British deer. Ecology and Evolution, 2019, 1–12. https://doi.org/10.1002/ece3.5424
D’Elia, J., Haig, S. M., Johnson, M., Marcot, B. G., & Young, R. (2015). Activity-specific ecological niche models for planning reintroductions of California condors (Gymnogyps californianus). Biological Conservation, 184, 90–99.
Di Marco, M., & Santini, L. (2015). Human pressures predict species’ geographic range size better than biological traits. Global Change Biology, 21, 2169–2178.
Forister, M. L., McCall, A. C., Sanders, N. J., Fordyce, J. A., Thorne, J. H., O’Brien, J., Waetjen, D. P., & Shapiro, A. M. (2010). Compounded effects of climate change and habitat alteration shift patterns of butterfly diversity. PNAS, 107(5), 2088–2092. https://www.pnas.org/cgi/doi/ https://doi.org/10.1073/pnas.0909686107
Fourcade, Y., Engler, J. O., Rodder, D., & Secondi, J. (2014). Mapping species distributions with MAXENT using a geographically biased sample of presence data: A performance assessment of methods for correcting sampling bias. PLoS One, 9(5), e97122. https://doi.org/10.1371/journal.pone.0097122
Friedl, M., & Sulla-Menashe, D. (2019). MCD12Q1 MODIS/Terra+aqua land cover type yearly L3 global 500 m SIN Grid V006 . NASA EOSDIS Land Processes DAAC. Retrieved 25 September 2020 from https://doi.org/10.5067/MODIS/MCD12Q1.006
Freeman, B., Jimenez-Garcia, D., Barca, B., & Grainger, M. (2019). Using remotely sensed and climate data to predict the current and potential future geographic distribution of a bird at multiple scales: The case of Agelastes meleagrides, a western African forest endemic. Avian Research, 10, 22. https://doi.org/10.1186/s40657-019-0160-y
Gallardo, B., Zieritz, A., & Aldridge, D. C. (2015). The importance of the human footprint in shaping the global distribution of terrestrial, freshwater and marine invaders. PLoS One, 10(5), e0125801. https://doi.org/10.1371/journal.pone.0125801
Gama, M., Crespo, D., Dolbeth, M., & Anastacio, P. (2015). Predicting global habitat suitability for Corbicula fluminea using species distribution models: The importance of different environmental datasets. Ecological Modelling, 319, 163–169. https://doi.org/10.1016/j.ecolmodel.2015.06.001
Gaul, W., Sadykova, D., White, H. J., León-Sánchez, L., Caplat, P., Emmerson, M. C., & Yearsley, J. M. (2020). Data quantity is more important than its spatial bias for predictive species distribution modelling. Retrieved 10 February 2021 from https://www.researchgate.net/publication/341706726 https://doi.org/10.1101/2020.05.24.113415
Genero, F., Franchini, M., Fanin, Y., & Filacorda, S. (2020). Spatial ecology of non-breeding Eurasian Griffon vultures Gyps fulvus in relation to natural and artificial food availability. Bird Study, 67(1), 53–70.
Golterman, H. L. (1975). Physiological Limnology: An approach to the physiology of lake ecosystem (Eds.). Developments in Water Science Series. Elsevier Publishing company. https://doi.org/10.1016/s0167-5648(08)71058-X
Gschweng, M., Kalko, E. K. V., Berthold, P., Fiedler, W., & Fahr, J. (2012). Multi-temporal distribution modelling with satellite tracking data: predicting responses of a long distance migrant to changing environmental conditions. Journal of Applied Ecology, 49, 803–813.
Hameed, S., Din, Ju., Ali, H., Kabir, M., Younas, M., Rehman, E., Bischof, R., & Nawaz, M. A. (2020). Identifying priority landscapes for conservation of snow leopards in Pakistan. PLoS One, 15(11), e0228832. https://doi.org/10.1371/journal.pone.0228832
Henriques, M., Granadeiro, J. P., Monteiro, H., Nuno, A., Lecoq, M., Cardoso, P., Regalla, A., & Catry, P. (2018). Not in wilderness: African vulture strongholds remain in areas with high human density. PLoS One, 13(1), e0190594. https://doi.org/10.1371/journal.pone.0190594
Herrero, J., García-Serrano, A., Couto, S., Ortuño, V., & García-González, R. (2006). Diet of wild boar Sus scrofa L. and crop damage in an intensive agroecosystem. European Journal of Wildlife Research, 52, 245–250.
Hijmans, R. J., Cameron, S. E., Parra, J. L., Jones, P. G., & Jarvis, A. (2005). Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology, 25, 1965–1978. https://doi.org/10.1002/joc.1276
Holland, A. E., Byrne, M. E., Hepinstall-Cymerman, J., Bryan, A. L., DeVault, T. L., Rhodes, O. E., & Beasley, J. C. (2019). Evidence of niche differentiation for two sympatric vulture species in the Southeastern United States. Movement Ecology, 7, 31. https://doi.org/10.1186/s40462-019-0179-z
Holyoak, M., & Heath, S. K. (2016). The integration of climate change, spatial dynamics, and habitat fragmentation: A conceptual overview. Integrative Zoology, 11, 40–59. https://doi.org/10.1111/1749-4877.12167
Ilanloo, S.S., Khani, A., Kafash, A., Valizadegan, N., Ashrafi, S., Loercher. F., Ebrahimi, E. & Yousefi, M. (2020). Applying opportunistic observations to model current and future suitability of the Kopet Dagh Mountains for a near threatened avian scavenger. Avian Biology Research, 00(0). Retrieved 10 February 2021 from https://www.researchgate.net/publication/346416388 https://doi.org/10.1177/1758155920962750
ISFR. (2017). India State of Forest Report. Forest Survey of India (MoEFCC), Dehradun, India.
Ingenloff, K. (2017). Biologically informed ecological niche models for an example pelagic, highly mobile species. European Journal of Ecology, 3(1), 55–75. https://doi.org/10.1515/eje-2017-0006
IUCN. (2020). IUCN Redlist. Retrieved 24 December 2020 from https://www.iucnredlist.org/search?query=bearded%20vulture&searchType=species
Jackson, J. B. C. (1981). Interspecific competition and species’ distributions: The ghosts of theories and data past. Theoretical Ecology, 21(4), 889–901.
Jackson, M. M., Gergel, S. E., & Martin, K. (2015). Citizen science and field survey observations provide comparable results for mapping Vancouver Island White-tailed Ptarmigan (Lagopus leucura saxatilis) distributions. Biological Conservation, 181, 162–172.
Jha, K. K. (2015). Distribution of vultures in Uttar Pradesh. India. Journal of Threatened Taxa, 7(1), 6750–6763.
Jha, K. K. (2017). Vulture atlas Central India MP. Indian Institute of Forest Management, Bhopal, India.
Jha, K. K. (2018). Mapping and management of vultures in an Indian stronghold. In M.O. Campbell, Geomatics and Conservation Biology. (Eds.), 45-75. Nova Science Publishers, New York.
Jha, K. K., Campbell, M. O., & Jha, R. (2020). Vultures, their population status and some ecological aspects in an Indian stronghold. Notulae Scientia Biologicae, 12(1), 124–142.
Jha, K. K., & Jha, R. (2020). Habitat suitability mapping for migratory and resident vultures: A case of Indian stronghold and species distribution model. Journal of Wildlife and Biodiversity, 4(3), 91–111.
Jha, K. K., & Jha, R. (2021). Study of vulture habitat suitability and impact of climate change in Central India using MaxEnt. Journal of Resources and Ecology, 12(1), 30–42. https://doi.org/10.5814/j.issn.1674-764x.2021.01.004
Jha, K. K., Jha, R., & Campbell, M. O. (2021). The distribution, nesting habits and status of threatened vulture species in protected areas of Central India. Ecological Questions, 32(2021)2. https://doi.org/10.12775/EQ.2021.020
Jiao, S., Zeng, Q., Sun, G., & Lei, G. (2016). Improving conservation of cranes by modeling potential wintering distributions in China. Journal of Resources and Ecology, 7(1), 44–50.
Jiménez-García, D., & Peterson, A. T. (2019). Climate change impact on endangered cloud forest tree species in Mexico (Impacto del cambio climático sobre las especies de árboles amenazadas del bosque mesófilo en México). Revista Mexicana de Biodiversidad, 90 (2019), e902781 2. https://doi.org/10.22201/ib.20078706e.2019.90.2781
Kale, V. S. (2014). Landscapes and landforms of India. Springer.
Kambale, A. A. (2011). A study on breeding behaviour of oriental white backed vulture (Gyps bengalensis) in Anjarle and Deobag, Maharashtra. Wildlife Institute of India, Dehradun.
Kupika, O. L., Gandiwa, E., Kativu, S., & Nhamo, G. (2018). Impacts of climate change and climate variability on wildlife resources in Southern Africa: Experience from selected protected areas in Zimbabwe. In B. Sen, & O. Grillo (Eds.), Selected studies in biodiversity. IntechOpen. https://doi.org/10.5772/intechopen.70470
Lewison, R., Oro, D., Godley, B., Underhill, L., Bearhop, S., Wilson, R. P., Ainley, D., Arcos, J. M., Boersma, P. D., Borboroglu, P. G., Boulinier, T., Frederiksen, M., Genovart, M., González-Solís, J., Green, J. A., Grémillet, D., Hamer, K. C., Hilton, G. M., Hyrenbach, K. D., & Yorio, P. (2012). Research priorities for seabirds: Improving conservation and management in the 21st century. Endangered Species Research, 17, 93–121.
Liberatori, F., & Penteriani, V. (2001). A long-term analysis of the declining population of the Egyptian vulture in the Italian peninsula: Distribution, habitat preference, productivity and conservation implications. Biological Conservation, 101, 381–389.
Liminana, R., Soutullo, A., Arroyo, B., & Urios, V. (2012). Protected areas do not fulfil the wintering habitat needs of the trans-Saharan migratory Montagu’s harrier. Biological Conservation, 145, 62–69.
Lin, L.-H., Zhu, X.-M., Du, Y., Fang, M.-C., & Ji, X. (2019). Global, regional, and cladistic patterns of variation in climatic niche breadths in terrestrial elapid snakes. Current Zoology, 65(1), 1–9. https://doi.org/10.1093/cz/zoy026
Liu, L., Zhao, Z., Zhang, Y., & Wu, X. (2017). Using MaxEnt model to predict suitable habitat changes for key protected species in Koshi Basin, Central Himalayas. Journal of Resources and Ecology, 8(1), 77–87.
Markandya, A., Taylor, T., Longo, A., Murty, M. N., Murty, S., & Dhavala, K. (2008). Counting the cost of vulture decline—An appraisal of the human health and other benefits of vultures in India. Ecological Economics, 67, 194–204.
Marinković, S. P., Orlandić, L. B., Skorić, S. B., & Karadžić, B. D. (2012). Nest-site preference of griffon vulture (Gyps Fulvus) in Herzegovina. Archives of Biological Science, 64(1), 385–392. https://doi.org/10.2298/ABS1201385M
Mateo, R. G., De La Estrella, M., Felicísimo, Á. M., Munoz, J., & Guisan, A. (2013). A new spin on a compositionalist predictive modelling framework for conservation planning: a tropical case study in Ecuador. Biological Conservation, 160, 150–161.
McDonald, M. M., Johnson, S. M., Henry, E. R., & Cunneyworth, P. M. K. (2019). Differences between ecological niches in northern and southern populations of Angolan black and white colobus monkeys (Colobus angolensis palliatus and Colobus angolensis sharpei) throughout Kenya and Tanzania. American Journal of Primatology, 2019, e22975. https://doi.org/10.1002/ajp.22975
McGrady, M. J., Karelus, D. L., Rayaleh, H. A., Willson, M. S., Meyburg, B.-U., Oli, M. K., & Bildsten, K. (2018). Home ranges and movements of Egyptian Vultures Neophron percnopterus in relation to rubbish dumps in Oman and the Horn of Africa. Bird Study, 65(4), 544–556. https://doi.org/10.1080/00063657.2018.1561648
Merow, C., Smith, M. J., & Silander, J. A. (2013). A practical guide to MaxEnt for modeling species’ distributions: what it does, and why inputs and settings matter. Ecography, 36, 1058e1069.
Milanesi, P., Mori, E., & Menchetti, M. (2020). Observer-oriented approach improves species distribution models from citizen science data. Ecology and Evolution, 00, 1–11. https://doi.org/10.1002/ece3.6832
Navaneethan, B., Sankar, K., Qureshi, Q., & Manjrekar, M. (2015). The status of vultures in Bandhavgarh Tiger Reserve, Madhya Pradesh, central India. Journal of Threatened Taxa, 7(14), 8134–8138. https://doi.org/10.11609/jott.2428.7.14.8134-8138
Newbold, T., Hudson, L. N., Hill, S. L. L., Contu, S., Lysenko, I., Senior, R. A., Börger, L., Bennett, D. J., Choimes, A., Collen, B., Day, J., Palma, A. D., Díaz, S., Echeverria-Londoño, S., Edgar, M. J., Feldman, A., Garon, M., Harrison, M. L. K., Alhusseini, T., & Purvis, Andy. (2015). Global effects of land use on local terrestrial biodiversity. Nature, 520, 45–50.
Ortiz-Urbina, E., Diaz-Balteiro, L., & Iglesias-Merchan, C. (2020). Influence of anthropogenic noise for predicting cinereous vulture nest distribution. Sustainability, 12, 503. https://doi.org/10.3390/su12020503
Osborne, P. E., & Seddon, P. J. (2012). Selecting suitable habitats for reintroductions: variation, change and the role of species distribution modelling. In J.G., Ewen, D.P., Armstrong, K.A., Parker, P.J. Seddon, (Eds.). Reintroduction Biology: Interacting Science and Management, 73–104. United Kingdom, Blackwell Publishing Ltd.
Pain, D. J., Cunningham, A. A., Donald, P. F., Duckworth, J. W., Houston, D. C., Katzner, T., et al. (2003). Gyps vulture declines in Asia: Temperospatial trends, causes and impacts. Conservation Biology, 17, 661–671.
Peterson, A. T. (2006). Uses and requirements of ecological niche models and related distribution models. Biodiversity Informatics, 3, 59–72.
Peterson, A. T., Papeş, M., & Soberón, J. (2008). Rethinking receiver operating characteristic analysis applications in ecological niche modeling. Ecological Modelling, 213, 63–72.
Phillips, S. J., Anderson, R. P., & Schapire, R. E. (2006). Maximum entropy modelling of species geographic distribution. Ecological Modelling, 190, 231–259.
Phillips, S. J., & Elith, J. (2010). POC plots: calibrating species distribution models with presence-only data. Ecology, 91(8), 2476–84. https://doi.org/10.1890/09-0760.1
Phipps, W. L., Diekmann, M., MacTavish, L. M., Mendelsohn, J. M., Naidoo, V., Wolter, K., & Yarnell, R. W. (2017). Due South: A first assessment of the potential impacts of climate change on Cape vulture occurrence. Biological Conservation, 210, 16–25. https://doi.org/10.1016/j.biocon.2017.03.028
Pires, M. M., Périco, E., Renner, S., & Sahlén, G. (2018). Predicting the effects of future climate change on the distribution of an endemic damselfly (Odonata, Coenagrionidae) in subtropical South American grasslands. Journal of Insect Conservation, 22, 303–319. https://doi.org/10.1007/s10841-018-0063-y
Poirazidis, K., Goutner, V., Skartsi, T., & Stamou, G. (2004). Modelling nesting habitat as a conservation tool for the Eurasian black vulture (Aegypius monachus) in Dadia Nature Reserve, northeastern Greece. Biological Conservation, 118, 235–248. https://doi.org/10.1016/j.biocon.2003.08.016
Prakash, V., Pain, D. J., Cunningham, A. A., Donald, P. F., Prakash, N., Verma, A., Gargi, R., Sivakumar, S., & Rahmani, A. R. (2003). Catastrophic collapse of Indian white-backed Gyps bengalensis and long-billed Gyps indicus vulture populations. Biological Conservation, 109, 381–390.
Preston, K. L., Rotenberry, J. T., Redak, R. A., Michael, F., & Allen, M. F. (2008). Habitat shifts of endangered species under altered climate conditions: Importance of biotic interactions. Global Change Biology, 14, 2501–2515. https://doi.org/10.1111/j.1365-2486.2008.01671.x
Purohit, A., & Saran, R. (2013). Population status and feeding behavior of Cinereous vulture (Aegypus monachus): Dynamics and implications for the species conservation in and around Jodhpur. World Journal of Zoology, 8(3), 312–318. https://doi.org/10.5829/idosi.wjz.2013.8.3.74148
Ramesh, T., Sankar, K., & Qureshi, Q. (2011). Status of vultures in Mudumalai Tiger Reserve, Western Ghats, India. Forktail, 27, 96–97.
Ramesh, V., Gopalakrishna, T., Barve, S., & Melnick, D. J. (2017). IUCN greatly underestimates threat levels of endemic birds in the Western Ghats. Biological Conservation, 210, 205–221.
Ravindranath, N. H., Joshi, N. V., Sukumar, R., & Saxena, A. (2006). Impact of climate change on forest in India. Current Science, 90(3), 354–361.
Roy-Dufresne, E., Saltré, F., Cooke, B. D., Mellin, C., Mutze, G., Cox, T., & Fordham, D. A. (2019). Modeling the distribution of a wide-ranging invasive species using the sampling efforts of expert and citizen scientists. Ecology and Evolution, 9(19), 11053–11063. https://doi.org/10.1002/ece3.5609
Santangeli, A., Girardello, M., Buechley, E., Botha, A., Di Minin, E., & Moilanen, A. (2019). Priority areas for conservation of Old World vultures. Conservation Biology, 33(5), 1056–1065. https://doi.org/10.1111/cobi.13282
Saenz-Jimenez, F., Rojas-Soto, O., Perez-Torres, J., Martinez-Meyer, E., & Sheppard, J. K. (2020). Effects of climate change and human influence in the distribution and range overlap between two widely distributed avian scavengers. Bird Conservation International (2020), 1-19. Retrieved 11 February 2021 from https://www.researchgate.net/publication/342012019 https://doi.org/10.1017/S0959270920000271
Santangeli, A., Spiegel, O., Bridgeford, P., & Girardello, M. (2018). Synergistic effect of land-use and vegetation greenness on vulture nestling body condition in arid ecosystems. Scientific Reports, 8, 13027. https://doi.org/10.1038/s41598-018-31344-2
Saran, R. P., & Purohit, A. (2012). Eco-transformation and electrocution: A major concern for the decline in vulture population in and around Jodhpur. International Journal of Conservation Science, 3(2), 111–118.
Şekercioğlu, C. H., Daily, G. C., & Ehrlich, P. R. (2004). Ecosystem consequences of bird declines. Proceedings of National Academy of Science, 101, 18042–18047.
Songer, M., Delion, M., Biggs, A. & Huang, Q. (2012). Modeling impacts of climate change on giant panda habitat. International Journal of Ecology, 108752, 12 pages. Retrieved 11 February 2021 from https://www.researchgate.net/publication/235798056 https://doi.org/10.1155/2012/108752
Sony, R. K., Sen, S., Kumar, S., Sen, M., & Jayahari, K. M. (2018). Niche models inform the effects of climate change on the endangered Nilgiri Tahr (Nilgiritragus hylocrius) populations in the southern Western Ghats, India. Ecological Engineering, 120, 355–363.
Straub, M. H., Kelly, T. R., Rideout, B. A., Eng, C., Wynne, J., Braun, J., & Johnson, C. K. (2015). Seroepidemiologic survey of potential pathogens in obligate and facultative scavenging avian species in California. Plos One, 10(11), e0143018. https://doi.org/10.1371/journal.pone.0143018
Sutton, W. B., Barrett, K., Moody, A. T., Loftin, C. S., de Maynadier, P. G., & Nanjappa, P. (2015). Predicted changes in climatic niche and climate refugia of conservation priority salamander species in the northeastern United States. Forests, 6, 1–26. https://doi.org/10.3390/f6010001
Teeffelen, A. J. A. V., Vos, C. C., & Opdam, P. (2012). Species in a dynamic world: Consequences of habitat network dynamics on conservation planning. Biological Conservation, 153, 239–253.
Trautmann, S. (2018). Climate change impacts on bird species. In D. Tietze (Ed.), Bird Species, Fascinating Life Sciences. Springer Open. https://doi.org/10.1007/978-3-319-91689-7_12
Venter, O., Sanderson, E. W., Magrach, A., Allan, J. R., Beher, J., Jones, K. R., Possingham, H. P., Laurance, W. F., Wood, P., Fekete, B. M., Levy, M. A., & Watson, J. E. M. (2016a). Data descriptor: Global terrestrial human footprint maps for 1993 and 2009. Scientific Data, 3, 160067. https://doi.org/10.1038/sdata.2016.67
Venter, O., Sanderson, E. W., Magrach, A., Allan, J. R., Beher, J., Jones, K. R., Possingham, H. P., Laurance, W. F., Wood, P., Fekete, B. M., Levy, M. A., & Watson, J. E. M. (2016b). Sixteen years of change in the global terrestrial human footprint and implications for biodiversity conservation. Nature Communications, 7, 12558. https://doi.org/10.1038/ncomms12558
Wu, Q., Wang, L., Zhu, R., Yang, Y. B., Jin, H. Y., & Zou, H. F. (2016). Nesting habitat suitability analysis of red-crowned crane in Zhalong Nature Reserve based on MAXENT modeling. Acta Ecologica Sinica, 36(12), 1–7.
Zeng, Q., Zhang, Y., Sun, G., Duo, H., Wen, L., & Lei, G. (2015). Using species distribution model to estimate the wintering population size of the endangered scaly-sided merganser in China. PLoS One, 10(2), e0117307. https://doi.org/10.1371/journal.pone.0117307
Zhang, J., Jiang, F., Li, G., Qin, W., Li, S., Gao, H., Cai, Z., Lin, G., & Zhang, T. (2019). MaxEnt modeling for predicting the spatial distribution of three raptors in the Sanjiangyuan National Park, China. Ecology and Evolution, 9, 6643–6654. https://doi.org/10.1002/ece3.5243
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Jha, R., Jha, K.K. Habitat prediction modelling for vulture conservation in Gangetic-Thar-Deccan region of India. Environ Monit Assess 193, 532 (2021). https://doi.org/10.1007/s10661-021-09323-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10661-021-09323-4