
Abstract
As obligate scavengers, vultures are important to ecosystem health but their numbers are declining globally. A major cause may be habitat loss due to anthropogenic or natural factors. Four threatened and endangered non-Gyps vultures (Bearded, Cinereous, Egyptian, Red-headed) found in many other countries also inhabit diverse floristic landscapes of India. This study aimed to determine present habitat expanse and the projected changes in habitat in future, identify vital habitat influencing factors, and suggest conservation strategies. Species Distribution Model Maxent, presence locations and bioclimate data for the present, and short- and long-term future were used and predictions were made for these four species. To increase the accuracy, uncertainties were removed, ensemble models were created using three GCMs and data for two RCPs (RCP4.5, RCP8.5) across two future tenures. All the models had strong predictability (AUC: 0.759–0.966, TSS: 0.445–0.866, and CBI: 0.986–1.000). With respect to habitat suitability across the landscapes, in the present-day scenario, 5%, 10%, 18% and 48% of the area were found suitable for Bearded, Cinereous, Red-headed, and Egyptian vultures, respectively, against 3.28 million km2. This expanse fluctuated due to the changing climate in future scenarios, considerably large patches undergoing either loss or gain in suitability. The three most vital bioclimatic variables for habitat prediction were bio19 (Precipitation of coldest quarter), bio01 (Mean annual temperature), and bio07 (Temperature annual range). The data generated could be useful in developing conservation strategies. Consistently suitable area could be used for establishing vulture protection area and vulnerable areas for habitat improvement.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
India is one of the tropical countries where scavenging is mostly done by vultures. The vultures that inhabit India and found in other countries can be categorized as non-Gyps and Gyps vultures on account of their behavior and taxonomy. The former is comparatively less social and has the advantage of lower risk of poisoning at carcass but a minor disadvantage of late information about carcass presence. Therefore, the management requirement of these two groups differs from each other (Campbell 2015). However, out of four monotypic non-Gyps species found in this country, three (Gypaetus barbatus Linnaeus 1758, Bearded Vulture = BRV; Neophron percnopterus Linnaeus 1758, Egyptian Vulture = EGV; Sarcogyps calvus Scopoli 1786, Red-headed Vulture = RHV) are residents while one (Aegypius monachus Linnaeus 1766, Cinereous vulture = CNV) is wintering (MoEFCC 2020). Two (BRV and CNV) have Nearly Threatened status of conservation while, EGV is Endangered and RHV is Critically Endangered (IUCN 2021). Cuthbert et al. (2006) reported an 80% decline in EGV and 91% in RHV during 1991–2003 survey and Acharya et al. (2010) recorded 73–80% decrease in BRV during 2002–2008 survey in India and neighboring countries. Moreover, population of all these vultures is decreasing globally and their population trend has been marked as declining. The estimated lowest global number of these non-Gyps vultures is around 33,000 to 75,000 only (IUCN 2021). Further lowering of such a small number is a serious cause of concern resulting in the loss of an important group of obligatory scavengers. This would in turn affect ecosystem services and balance through ecological processes which control energy flow in ecosystems (Almaraz et al. 2022).
India is a huge country which could be subdivided into smaller landscapes as floristic regions. These landscapes have varied phytogeographical features in altitude, temperature and precipitation ranges, and vegetation structure (Sharma 2005). Vegetated landscapes like temperate coniferous forests, tropical and subtropical broadleaved forests, subtropical scrublands, and agroforests support population of many vulture species (Chhangani 2007; Ramesh et al. 2011; Thakur and Narang 2012; Jha et al. 2022) indicating their suitability. Vulture populations have declined in recent decades (Prakash et al. 2017) for which several hypotheses have been proposed. Apart from poisoning and persecution (Ogada et al. 2011; Ilanloo et al. 2020), a major concern in India has been shelter destruction along with food shortage and poor breeding success (Chhangani and Mohnot 2004). Habitat loss and climate change may cause species extinction along with change in habitat distribution and redistribution of species (Ilanloo et al. 2020; Thapa et al. 2021). Bertrand et al. (2011) and Kupika et al. (2018) further elaborated that climate parameters, such as humidity, precipitation, and temperature, have significant effects on quantity and quality of the habitat and spatial distribution of animal species. Vulture species distribution is also dependent on physical features of habitat, such as vegetation cover, cliff, and water presence (Herrero et al. 2006; Bosch et al. 2014; Jha and Jha 2021). At the same time, climatic factors like the rainfall patterns influence the success of vulture breeding (Bridgeford and Bridgeford 2003; Virani et al. 2012) and temperature change also governs the reproduction of vultures, directly causing stress (Chaudhry 2007 and Schultz 2007 in Phipps et al. 2017, Bamford et al. 2009, Midgley and Bond 2015).
The threat to vulture population and the reasons for their further decline warrant conservation management for which knowledge of habitat suitability, habitat controlling factors, and changes in suitability status in future is a precondition. Such information could be collected using species distribution modeling (Angelieri et al. 2016). Numerous algorithms are available for species distribution modeling (Li and Wang 2013; Jha et al. 2022), but MaxEnt has gained a significant advantage because it uses both continuous and categorical, and presence data as input variables (Eastmann 2016; Morales et al. 2017). MaxEnt is popular in data and resource scarce regions especially in developing countries (Banag et al. 2015; Kaky et al. 2020). Some of the other advantages of this algorithm are better performance than many other models (Elith et al. 2006; Banag et al. 2015), ease of use, and functionality (Morales et al. 2017; Mohammadi et al. 2019), requires presence only data and small sample size especially for the prediction of distribution for rare and threatened species (Wisz et al. 2008; Kumar and Stohlgren 2009), creates a spatially explicit map for habitat suitability, measures importance of each environmental variable (Elith et al. 2011; Groff et al. 2014), and can project habitat losses and gains in future under climate change (Phillips et al. 2006; Elith et al. 2011).
Keeping the requirements of vulture conservation in view, the present study used MaxEnt and aimed at (1) determining habitat suitability types and expanse in the varied floristic landscapes of India, (2) identifying habitat determining major environmental factors, (3) assessing the changes in suitable habitats in near and distant future, and (4) suggesting some conservation measures.
Materials and methods
Study area and landscapes
This study covers the whole of India spread over a geographical area of 3.28 million km2 encompassing hills, plateau, and plains interspersed with forests, water bodies, scrubland, agriculture, built-up areas, and wasteland. Non-Gyps vultures are known to utilize such areas for nesting and foraging (Ramesh et al. 2011; Prakash et al. 2017; Jha et al. 2020). The ranges of environmental features of the country are: elevation (− 1 to 8583 m), mean annual temperature (− 33.8 to 30.0 °C), and mean annual precipitation (33–9312 mm) (Fick and Hijmans 2017; USGS EROS 2018).
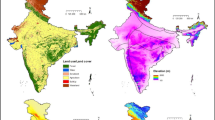
After modeling the habitat, the whole study area was divided into nine smaller units based on unique vegetation structure known as floristic landscapes (FLs or floristic region, sensu Sharma 2005). This has been done for simplified and vulture biology-based interpretation of results since, different types of forests are known to play their role in vulture habitation and foraging, and ease of conservation strategy implementation (Jha et al. 2020). However, these landscapes are dispersed in temperate mountains, subtropical plains and plateau, and tropical Indian Ocean. All the study area-related features and vulture locations are presented in Fig. 1, Supplementary Table 1, and Supplementary Fig. 1.

Maps showing location, physiographic and landscapes details of the study area, India. Top row: geographic position of India in the world. Bottom row: land use/land cover and floristic landscape delineation (color figure online)
Data collection and processing
Occurrence data of non-Gyps vultures were collected from field surveys in Uttar Pradesh, Madhya Pradesh, and Rajasthan, published literature (Supplementary Table 2), and citizen science repositories, namely eBird (http://www.ebird.org Sullivan et al. 2009) and iNaturalist (http://www.inaturalist.org iNaturalist users & Ueda 2020). Cleaning of occurrence data was carried out by removing duplicates. This was further subjected to spatial filtering (at 4 km) to improve the performance of models by reducing sampling bias (Brown et al. 2017). The finally reduced occurrence locations (473 BRV, 319 CNV, 2261 EGV and 555 RHV from 2585 BRV, 1991 CNV, 15,580 EGV, and 3598 RHV, respectively) were the model input.
The bioclimatic variables for the present (1970–2000) and future (2041–2060 represented by the year 2050 and 2061–2080 represented by the year 2070) were downloaded at 30 arc second resolution from https://www.worldclim.org/ (Fick and Hijmans 2017). The categorical land-use landcover (LULC) was sourced from Copernicus Global Land Service at 100 m resolution (Buchhorn et al. 2020). LULC available in 23 classes was reclassified into six broad categories (Forest, Water, Scrubland, Agriculture, Built-up, and Wasteland). Elevation SRTM 1 Arc second global data were downloaded from https://earthexpoler.usgs.gov/ (USGS EROS 2018). Since the data varied in spatial resolution and projection, they were resampled at 30 arc second spatial resolution and WGS 1984 projection. Bioclimatic and bioenvironmental variables used in this study are compiled in Supplementary Table 3. Collinearity effects among such variables were reduced by subjecting them to Pearson’s correlation test with the coefficient set at ± 0.8 (SDM toolbox, Brown et al. 2017). To improve the performance of models further, a bias file was used (Cao et al. 2016; Baloch et al. 2020) to minimize overfitting and avoid sampling habitat outside of a species’ known occurrence and account for collection sampling biases with coordinate data. The bias file also ensured selection of non-appearance points (background points) by limiting it to feasible areas of dispersal through “Background Selection: Sample by Buffered MCP” tool (Brown et al. 2017, SDM tool box).
Species distribution and climate models
MaxEnt 3.4.1 was used to make the present and future predictions from processed data. The random test percentage was set at 25 for training each model. The algorithm run type set at Bootstrap was run using 10 replicates (Dong et al. 2019; Mori et al. 2020) in 500 iterations, with a convergence threshold of 0.00001 and 10,000 maximum background points. For the present, eight models (four each species with and without LULC) were developed. Forty-eight future predictions using GCMs (CCSM4, HadGEM2A, MIROC5 for RCP4.5, RCP8.5 of four species and two terms) were averaged and total 16 predictions were made. Averaging was done using Raster Calculator tool of ArcGIS 10.5. To assess the strength of the models, three model evaluators Area Under receiver operator characteristic Curve (AUC), True skill statistics (TSS), and continuous Boyce index (CBI) were used in this study. Stacked Species Distribution Modeling (Schmitt et al. 2017) and modEvA (Barbosa et al. 2013) packages in R and were used for TSS and CBI, respectively.
Habitat classification and vital variable identification
Whole Indian landscape (3,287,263 km2) was categorized into different suitability classes and analyzed floristic region-wise. For this, the heatmaps obtained from MaxEnt modeling indexed between 0 and 1 were reclassified in to three suitability classes as per ranges 0–0.3 as Unsuitable, 0.3–0.6 as Moderate, and 0.6–1.0 as High for this study. This was guided by Zhang et al. (2019a)’s classification for vulture and other raptors with modifications since the current study area was too large and a coarse classification was considered good enough. For studying future habitat dynamics, the heatmaps reclassification was done as unsuitable (0–0.3) and suitable (0.3–1.0) only. This was used to calculate stable and unstable areas (areas undergoing habitat loss and gain) in future scenarios. In order to assess loss and gain in suitable area, future climatic predictions were compared with present climatic projection using Raster Calculator. This revealed area category change (suitable to unsuitable and vice versa) and indicated area dynamics. To further understand the habitats of vultures in different landscapes, floristic landscape and species-wise area calculation was done.
Some of the non-collinear variables which contributed the most (generally between three to five dominant ones; Zhang et al. 2019b; Anoop et al. 2020; Gao et al. 2021) in habitat determination were considered vital variables (sensu Zhang et al. 2020). These vital variables and their percentage contribution were obtained from MaxEnt generated ‘variable contribution table’.
Results
Models and performance
Altogether 56 predictions based on 10 non-collinear variables (bio01, bio02, bio03, bio07, bio12, bio14, bio15, bio18, bio19, and LULC) were developed for identification of dominant variables and habitat prediction. Species-wise four models each with and without LULC were generated for the present scenario. Apart from this, two short- and long-term future predictions of medium (RCP4.5) and extreme (RCP 8.5) emission pathways were also built. All these future models were without LULC. Values of AUC of the models for the present ranged between 0.763 and 0.964. Values for other model evaluators TSS and CBI ranged between 0.445 and 0.866, and 0.986 and 1.000, respectively (Supplementary Tables 4 and 5).
Vital habitat variables
The covariates contributed in varied proportions in model predictions in different species. However, the cumulative average contribution of the top three covariates was 66% (range 52–89%) and for the top five covariates was 84% (range 73–93%) in the case of present projection with LULC. Without LULC, it differed marginally, 69% and 83%, respectively in the sets of three and five covariates. Considering all the species together, the top five rankers or dominant variables (based on modified Likert ranking method; Bhattacherjee 2012) in decreasing order in models without LULC were bio19, bio01, bio07, bio15, and bio14, while in models with LULC, they were bio19, LULC, bio15, bio1, and bio07. Species-wise, most important covariates for BRV and EGV were bio01 and for CNV and RHV was bio19 in the case of models without LULC. In models with LULC, this changed to bio01 (BRV), bio19 (CNV), and bio15 (EGV and RHV). The jackknife charts of training gain (Supplementary Fig. 2) were broadly similar in the ranking (importance) of variables in relation to variable contribution table. Within the LULC, built-up and water were very important but agriculture the least important components (Supplementary Fig. 3). Supplementary Figs. 4 and 5 depict the relationship between vital climatic factors of habitats of non-Gyps species and probability of vulture occurrence (i.e., a particular range of temperature and precipitation affecting habitat suitability positively or negatively). Bearded Vulture showed different trends than other three vultures.
Floristic landscape suitability
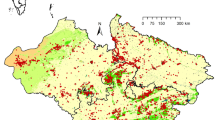
Suitable non-Gyps habitat was found in all the FLs in present projection (except Andaman and Nicobar and other Islands which was ignored due to non-reporting of vultures from this landscape). The habitat of resident non-Gyps vultures (BRV, EGV, and RHV) was spread broadly all over the country from the northernmost Western Himalaya landscape to the southernmost Malabar landscape, but the wintering vulture (CNV) habitat was mainly confined to Western Himalaya landscape, West Indian Plain landscape, Central India landscape, Malabar landscape and Eastern Himalaya landscape. However, BRV habitat was restricted to Western Himalaya landscape only while EGV habitat was spread everywhere except Eastern Himalaya landscape and Assam landscape. RHV habitat was scattered in all the landscapes (Fig. 2 and Supplementary Figs. 6–10). The future scenarios of suitable habitat presence of these vultures in different landscapes were similar to the present, but the expanse got changed.

Current habitat distribution of non-Gyps vultures projected without LULC i.e., with only climatic variables (color figure online)
Suitable habitats
It was a general trend that in all the species, unsuitable area was much larger than suitable area in both the predictions of the present (with and without LULC). Within the suitable habitats, moderately suitable area was much larger than highly suitable area in all the cases (Table 1). However, suitable habitat area under present prediction (without LULC) in decreasing order was 47.92%, 18.04%, 10.35%, and 5.10% out of 3.28 million km2 for EGV, RHV, CNV, and BRV, respectively. The corresponding figures for the prediction with LULC were 42.86%, 13.60%, 8.63%, and 5.01, respectively, showing a decrease in area.
Suitable habitats of the near and far future predicted on the basis of variables without LULC using the two RCPs (4.5 and 8.5), compared with present (without LULC) prediction indicated change in area of suitable habitats. BRV and CNV showed mixed trends in the change in area expanse over the two terms. RHV showed increasing trend (exception 2050-RCP8.5), but EGV showed area decrease in all scenarios.
Bearded Vulture showed the most specific preference for suitable habitats present mainly in Western Himalaya landscape. It has minor potential in Eastern Himalaya and Malabar (14–35 km2) but completely absent in other landscapes. RHV and EGV recorded the presence of suitable habitat in all the landscapes, but in general, the former had lesser suitable area than the latter. CNV, the migratory species, also appeared in all the FLs with much lesser occupancy as compared to RHV and EGV (Supplementary Table 6 and 7).
Habitat dynamics
The habitat of all non-Gyps species (BRV, CNV, EGV, and RHV) showed change in area and locality in future scenarios as compared to the present without LULC (Table 2 and Fig. 3, Supplementary Figs. 11–14). In all the four species, parts of the present habitat were predicted to undergo change in terms of gain or loss in area. Suitable habitats are projected to lose area and become unsuitable while also gaining some areas from the unsuitable landscape. Therefore, net loss or net gain is the first indicator of the impact of changing climate. There was a clear-cut trend of net loss in the case of EGV and BRV, but the latter had an exception in scenario 2070-RCP4.5. The case of RHV was reverse, where there was a clear trend of gain similar to CNV, the latter having an exception in scenario 2070-RCP4.5. However, in rounded numerical terms, EGV showed net loss between 1 and 3% (between 11,469 and 40,327 km2) and RHV showed net gain between 2 and 6% (between 10,690 and 32,327 km2) across the scenarios. BRV showed net loss between 1 and 5% (between 1375 and 7928 km2) except net gain of 1% (1884 km2) in 2070-RCP4.5 scenario and CNV showed net gain between 5 and 11% (between 16,143 and 34,675 km2) except net loss of 11% (32,973 km2) in 2070-RCP4.5 scenario.

Habitat suitability dynamics of non-Gyps vultures. Note the gain (green polygon) in suitable area from unsuitable habitat and the loss (red polygon) of suitable area from suitable habitat. Unsuitable (yellow) and suitable (blue) area are unchanged area (color figure online)
Discussion
Model robustness
AUC is widely used for assessing the accuracy of models but some researchers pointed out its inadequacy (Lobo et al. 2008) and suggested use of TSS and CBI (Allouche et al. 2006; Hirzel et al. 2006). Model evaluator values, respectively, for these indicators, above 0.7 (Swets 1988), 0.4 (Allouche et al. 2006), and 0.7 (Hirzel et al. 2006) are acceptable. Therefore, our predictions with AUC (0.763–0.964), TSS (0.445–0.866), and CBI (0.986–1.000) values are acceptable and good for conservation planning (Hosmer and Lemeshow 2004; Zeng et al. 2015).
Vital model predictors
The current habitat characterization of non-Gyps vultures in our study was governed by precipitation (bio19 and bio15), LULC, and temperature (bio1 and bio07), the top five vital contributors. This is in general agreement with earlier studies on raptors and scavengers (Phipps et al. 2017; Zhang et al. 2019a) that the bioclimatic variables influence habitat determination. As also noted in other studies that precipitation (Virani et al. 2012), vegetation and landscape (Hernandez-Baz et al. 2016; Cable et al. 2021), and temperature (Chaudhry 2007, Corovic et al. 2018) regulate the species distribution. However, species-wise variation in the most influential covariates of models without and with LULC in the present study indicated that habitat suitability was not a function of any single variable, rather it was a product of interaction among numerous covariates in different quantities (Jha and Jha 2021). This can further be substantiated from responses of bioclimatic factors to vulture’s occurrence probability (Supplementary Figs. 4 and 5). For example, precipitation of the coldest quarter (bio19), > 100 mm had a positive effect on BRV, almost no effect on CNV but a negative effect on EGV and RHV. Similarly, annual mean temperature (bio1) initially had the positive effect from 18 to 25 °C, depending on the species. Overall, similarity in responses between EGV and RHV could be explained by their habit of being resident species of warmer plains. Marginal differences of CNV from these two may be due to its residency in colder climates but spending warmer winters in the foothills and plains. BRV showing an entirely different response may be assigned to its residency in colder climates of higher altitude.
The LULC components also varied with the non-Gyps species studied, but order of preference was contrasted with Jha and Jha (2020) where forest and waterbody were the top contributors instead of built-up area. Only similarity in these two studies was the least importance of agriculture fields. However, study of Jha and Jha (2021) supported the present findings—built-up the most influential landcover covariate of the habitat.
Habitat suitability and landscapes
Present models predicted varied habitat expanse between with LULC and without LULC (Fig. 2 and Supplementary Fig. 6; and Table 1). The former had a lower amount of suitable area than the latter which could be attributed to the fact that the climatic umbrella is mostly larger than the environmental one due to the specific requirement of a niche e.g., trees/cliffs, water, ungulate/cattle concentration, etc. (Jha and Jha 2021). This implied that the actual suitable habitat area was an overestimation in the case of without LULC or climate-only models. It was assumed that such overestimation would also be in the cases of all the future models which are climate-only in nature. Nevertheless, this limitation finds corroboration in Preston et al. (2008)’s argument that ‘this was the general case with most distribution models predicting species responses to climate change which included climate variables and rarely the biotic interactions’. It must be noted that LULC has important role in habitat determination. Therefore, it should not be ignored. It can neither be assumed as a constant in future models since LULC (tree cover) is changing every year in floristic landscapes (ISFR 2021). Further, it is predicted to undergo changes itself due to climate crisis (Ravindranath et al. 2006).
Out of the four non-Gyps species, EGV had maximum suitable area followed by RHV, CNV and BRV in decreasing order. Similarly, their spread in different landscapes also indicated toward higher to lower number of landscapes coverage (Table 1 and Supplementary Table 6, and Fig. 2 and Supplementary Fig. 6). This indicated their habitat preference and area availability in the country. For example, BRV is a high-altitude bird (Subedi et al. 2018); therefore, they were largely confined in a limited area in Western Himalaya landscape. In contrast, RHV, a low-altitude species, requiring tall trees for nesting (Sinha et al. 2017) was spread in all the landscapes. The EGV, primarily a low-altitude and cliff-nesting species with high plasticity in adaptation, are known to invade urban area for food (Bahadur et al. 2019) along with nesting on tall structures like overhead water tank, tall buildings (Kumar et al. 2020). These features gave them the opportunity to spread in all most all the landscapes (exception Assam) with much larger area as compared to RHV. The CNV, being a migratory species only spent winters in northern and western India and required only roosting areas. A common assumption for all the four species is that there was sufficient availability of food in the vulture inhabiting area.
Habitat dynamics and climate impact
Ilanloo et al. (2020) identified climate change as a major factor for changes in habitat distribution within the short span of a few decades. Similar to this, in our findings, in comparison to current climatic scenario, all the four cases of future (2050-RCP4.5, 2050-RCP8.5, 2070-RCP4.5, and 2070-RCP8.5) predictions showed some loss of suitable habitat within three to five decades. Such shrinkage in habitat is reported earlier in birds (Banda and Tassie 2018), vultures (Saenz-Jiminez et al. 2020) and other organisms (Kazmi et al. 2022). However, this loss had been offset due to addition of suitable area (net gain; RHV, CNV with one exception) in one outcome and remained unaffected (net loss; EGV, BRV with one exception) in another outcome. The contrasting change in suitable habitat i.e., expansion of area due to climate change is also reported in birds (Gaudreau et al 2018). A review by Santini et al. (2021) presented wider coverage of literature indicating contraction and expansion of habitat area due to climate change. However, from our study, it can be hypothesized, sensu lato, that the damaging impact of changing climate on species distribution could be selective. While EGV and BRV are projected to face some difficulty, RHV and CNV could be benefited by climate change. Comparison from a recent study, (Jha and Jha 2023) showed that these resident non-Gyps (EGV and BRV except RHV) have similar habitat constraints as the Gyps residents (Indian Vulture, Slender-billed Vulture, and White-rumped Vulture). The migratory non-Gyps (CNV) and Gyps (Eurasian Griffon and Himalayan Griffon) showed mixed trend. This could be due to specific niche and habitat requirement of different or a group of species and transformation of landscapes (Hassan and Ismail 2017; Byrne et al. 2019; Hill et al. 2021).
Management implication
Effective management of wildlife populations requires sound knowledge of species distributions (or habitat suitability) and associated threats (Passadore et al. 2018). Though suitable areas (moderate and highly suitable) of non-Gyps species are overlapping, total suitable area in different FLs was in the following decreasing order: Central India > West Indian Plains > Western Himalaya > Gangetic Plains > Deccan > Malabar > Eastern Himalaya > Assam. However, highly suitable area was present only in Central India, Western Indian Plains, Western Himalaya, Gangetic Plains, and Deccan. These two different types of area (highly suitable and suitable) could be used for implementing different conservation strategies like, in situ conservation, reintroduction, and habitat rehabilitation (Khosravi et al. 2016; Anoop et al. 2020). Overlapping suitable areas of different vultures should receive high order of protection, and be treated as inviolate, as suggested by Zhang et al. (2019a). The areas identified as unstable (lost and gained) could be used for habitat improvement by ensuring nesting and foraging resources for further use by vultures in expanding their territory for increasing population. In general, development of reserves, protection of large trees, adoption of agroforestry, etc. could be useful in model predicted areas to attempt a reversal of the endangered status, to some extent, of indigenous vultures in different FLs. This may also help the wintering vultures spending their time in secured environment. Further, these management implications may be fine-tuned based on the results after further tweaking of the present study modeling in light of the fact that the climate change has maximum impact on natural resources like water, agriculture, and forests (Patasaraiya et al. 2021) which will significantly affect the habitat structure leading to loss of shelter and alimentary resources.
Conclusion
This study provided the first assessment of vital environmental factors and prediction of habitat suitability and dynamics due to climate change in non-Gyps vultures using SDM in India landscape-wise. Our models with high predictive power are dependable for use in conservation management. For further improvement, inclusion of future LULC in predictions would be needed since climate-only models do not represent the complete biotic and ecological niche of the species. Despite this limitation in future models, even climatic-only models are pragmatic enough to rely upon since they provide baseline data.
All the four cases of future predictions showed some loss of suitable habitat within three to five decades but was offset by addition of suitable area. In general, RHV and CNV had net gain while EGV and BRV had net loss. This led to a hypothesis that the damaging impact of changing climate on species distribution could be selective: EGV and BRV facing difficulty while RHV and CNV getting benefited by climate change. Apart from this result, the information about stable area (suitable as well as unsuitable), unstable area, or dynamic part of the stable area (loss and gain) may guide the managers or policy-makers for instant and pre-emptive actions in order to strategize the conservation activities. Habitat management activities may be taken up in the unstable area to control the degradation or ameliorate the would-be degraded area. Such areas could be used for in situ conservation and reintroduction of vulture species.
Data availability
The datasets generated during and/or analyzed during the current study are available in the manuscript and supplementary.
References
Acharya R, Cuthbert R, Baral HS, Chaudhary A (2010) Rapid decline of bearded vultures Gypaetus barbatus in upper mustang. Nepal Forktail 26:117–120
Allouche O, Tsoar A, Kadmon R (2006) Assessing the accuracy of species distribution models: prevalence, kappa and the true skill statistic (TSS). J Appl Ecol 43:1223–1232. https://doi.org/10.1111/j.1365-2664.2006.01214.x
Almaraz P, Martínez F, Morales-Reyes Z, Sanchez-Zapata JA, Blanco G (2022) Long-term demographic dynamics of a keystone scavenger disrupted by human-induced shifts in food availability. Ecol Appl 2022:e2579. https://doi.org/10.1002/eap.2579
Angelieri CCS, Adams-Hosking C, Ferraz KMPMB, de Souza MP, McAlpine CA (2016) Using species distribution models to predict potential landscape restoration effects on puma conservation. PLoS ONE 11(1):e0145232. https://doi.org/10.1371/journal.pone.0145232
Anoop NR, Babu S, Nagarajan R, Sen S (2020) Identifying suitable reintroduction sites for the White-rumped Vulture (Gyps bengalensis) in India’s Western Ghats using niche models and habitat requirements. Ecol Eng 158(2020):106034. https://doi.org/10.1016/j.ecoleng.2020.106034
Araujo MB, Pearson RG, Thuiller W, Erhard M (2005) Validation of species-climate impact models under climate change. Glob Change Biol 11:1504–1513. https://doi.org/10.1111/j.1365-2486.2005.01000.x
Bahadur KCK, Koju NP, Bhusal KP, Low M, Ghimire SK, Ranabhat R, Panthi S (2019) Factors influencing the presence of the endangered Egyptian vulture Neophron percnopterus in Rukum, Nepal. Glob Ecol Conserv 20:e00727. https://doi.org/10.1016/j.gecco.2019.e00727
Baloch MN, Fan J, Haseeb M, Zhang R (2020) Mapping potential distribution of Spodoptera frugiperda (Lepidoptera: Noctuidae) in Central Asia. InSects 11(3):172. https://doi.org/10.3390/insects11030172
Bamford AJ, Monadjem A, Hardy ICW (2009) An effect of vegetation structure on carcass exploitation by vultures in an African savanna. Ostrich 80:135–137
Banag C, Thrippleton T, Alejandro GJ, Reineking B, Liede-Schumann S (2015) Bioclimatic niches of selected endemic Ixora species on the Philippines: predicting habitat suitability due to climate change. Plant Ecol 216:1325–1340. https://doi.org/10.1007/s11258-015-0512-6
Banda LB, Tassie N (2018) Modeling the distribution of four-bird species under climate change in Ethiopia. Ethiop J Biol Sci 17(1):1–17
Barbosa AM, Real R, Munoz AR, Brown JA (2013) New measures for assessing model equilibrium and prediction mismatch in species distribution models. Divers Distrib 19:1333–1338. https://doi.org/10.1111/ddi.12100
Bertrand R, Lenoir J, Piedallu C, Riofrío-Dillon G, de Ruray P, Vidal C, Pierrat JC, Gégout JN (2011) Changes in plant community composition lag behind climate warming in lowland forests. Nature 479:517–520. https://doi.org/10.1038/nature10548
Bhattacherjee A (2012) Social science research: principles, methods, and practices. Textb Collect 3. http://scholarcommons.usf.edu/oa_textbooks/3;2012
Bosch J, Mardones F, Pérez A, de la Torre A, Muñoz MJ (2014) A maximum entropy model for predicting wild boar distribution in Spain. Span J Agric Res 12(4):984–999. https://doi.org/10.5424/sjar/2014124-5717
Bridgeford P, Bridgeford M (2003) Ten years of monitoring breeding Lappet-faced Vultures Torgos tracheliotos in the Namib-Naukluft Park, Namibia. Vulture News 48:3–11
Brown JL, Bennett JR, French CM (2017) SDMtoolbox 2.0: the next generation Python-based GIS toolkit for landscape genetic, bio-geographic and species distribution model analyses. PeerJ 5:e4095. https://doi.org/10.7717/peerj.4095
Buchhorn M, Smets B, Bertels L, De Roo B, Lesiv M, Tsendbazar N-E, Herold M, Fritz S (2020) Copernicus Global Land Service: Land Cover 100m: collection 3: epoch 2019: Globe. https://doi.org/10.5281/zenodo.3939050
Byrne ME, Holland AE, Turner KL, Bryan AL, Beasley JC (2019) Using multiple data sources to investigate foraging niche partitioning in sympatric obligate avian scavengers. Ecosphere 10(1):e02548. https://doi.org/10.1002/ecs2.2548
Cable AB, O’Keefe JM, Deppe JL, Hohoff TC, Taylor SJ, Davis MA (2021) Habitat suitability and connectivity modeling reveal priority areas for Indiana bat (Myotis sodalis) conservation in a complex habitat mosaic. Landsc Ecol 36:119–137. https://doi.org/10.1007/s10980-020-01125-2
Campbell MO (2015) Vultures, their evolution, ecology and conservation. CRC Press, Taylor & Francis Group, Boca Raton, London, New York
Cao B, Bai C, Zhang L, Li G, Mao M (2016) Modeling habitat distribution of Cornus officinalis with Maxent modeling and fuzzy logics in China. J Plant Ecol 9(6):742–751. https://doi.org/10.1093/jpe/rtw009
Chaudhry MJI (2007) Are Cape Vultues (Gyps coprotheres) feeling the heat? Behavioural differences at north and south facing colonies in South Africa. University of Cape Town, Cape Town
Chhangani AK (2007) Sightings and nesting sites of red-headed vulture Sarcogyps calvus in Rajasthan, India. Indian Birds 3:218–221
Chhangani AK, Mohnot SM (2004) Is diclofenac the only cause of vulture decline? Curr Sci 87(11):1496–1497
Ćorović J, Popović M, Cogălniceanu D, Carretero MA, Crnobrnja-Isailović J (2018) Distribution of the meadow lizard in Europe and its realized ecological niche model. J Nat Hist 52(29–30):1909–1925. https://doi.org/10.1080/00222933.2018.1502829
Cuthbert R, Green RE, Ranade S, Saravanan SS, Pain DJ, Cunningham AA, Prakash V (2006) Rapid population declines of Egyptian Vulture Neophron percnopterus and Red-headed Vulture Sarcogyps calvus in India. Anim Conserv 9:349–354. https://doi.org/10.1111/j.1469-1795.2006.00041.x
de Frutos A, Olea PP, Vera R (2007) Analyzing and modelling spatial distribution of summering lesser kestrel: the role of spatial autocorrelation. Ecol Model 200:33–44. https://doi.org/10.1016/j.ecolmodel.2006.07.007
Diarrassouba A, Gnagbo A, Kouakou YC, Campbell G, Tiedoué MR, Tondossama A, Kühl HS (2019) Koné I (2019) Differential response of seven duiker species to human activities in Taï National Park. Côte D’ivoire Afr J Ecol 00:1–11. https://doi.org/10.1111/aje.12680
Dong X, Chu Y, Gu X, Huang Q, Zhang J, Bai W (2019) Suitable habitat prediction of Sichuan snub-nosed monkeys (Rhinopithecus roxellana) and its implications for conservation in Baihe Nature Reserve, Sichuan, China. Environ Sci Pollut Res 26:32374–32384. https://doi.org/10.1007/s11356-019-06369-3
Eastmann RJ (2016) TerrSet habitat and biodiversity modeller manual. Clark Labs. https://clarklabs.org/wp-content/uploads/2020/05/Terrset-Manual.pdf
Elith J, Graham CH, Anderson RP et al (2006) Novel methods improve prediction of species’ distributions from occurrence data. Ecography 29:129–151. https://doi.org/10.1111/j.2006.0906-7590.04596
Elith J, Phillip SJ, Hastie T, Dudík M, Chee YE, Yates CJ (2011) A statistical explanation of MaxEnt for ecologists. Divers Distrib 17(1):43–57. https://doi.org/10.1111/j.1472-4642.2010.00725.x
Ferrier S (2002) Mapping spatial pattern in biodiversity for regional conservation planning: where to from here? Syst Biol 51:331–363
Fick SE, Hijmans RJ (2017) WorldClim 2: new 1km spatial resolution climate surfaces for global land areas. Int J Climatol 37(12):4302–4315. https://doi.org/10.1002/joc.5086
Gao T, Xu Q, Liu Y, Zhao J, Shi J (2021) Predicting the potential geographic distribution of Sirex nitobei in China under climate change using maximum entropy model. Forests 12:151. https://doi.org/10.3390/f12020151
Gaudreau J, Perez LID, Harati S (2018) Towards modelling future trends of Quebec’s boreal birds’ species distribution under climate change. Int J Geo-Inf 7:335. https://doi.org/10.3390/ijgi7090335
Groff LA, Marks SB, Hayes MP (2014) Using ecological niche models to direct rare amphibian surveys: a case study using the Oregon spotted frog (Rana pretiosa). Herpetol Conserv Biol 9:354–368
Hassan TA, Ismail OA (2017) Identification of vulture species around galagu station in Dinder national park February 2017. Biodivers Int J 1(6):72–75. https://doi.org/10.15406/bij.2017.01.00032
Heikkinen RK, Luoto M, Araújo MB, Virkkala R, Thuiller W, Sykes MT (2006) Methods and uncertainties in bioclimatic envelope modelling under climate change. Prog Phys Geogr 30(6):1–27. https://doi.org/10.1177/0309133306071957
Hernandez-Baz F, Romo H, Gonzalez JM, Hernandez MJM, Pastrana RG (2016) Maximum entropy niche-based modeling (Maxent) of potential geographical distribution of Coreura albicosta (Lepidoptera: Erebidae: Ctenuchina) in Mexico, Florida. Entomologist 99(3):376–380. https://doi.org/10.1653/024.099.0306
Herrero J, García-Serrano A, Couto S, Ortuño V, García-González R (2006) Diet of wild boar Sus scrofa L. and crop damage in an intensive agroecosystem. Eur J Wildl Res 52:245–250. https://doi.org/10.1007/s10344-006-0045-3
Hill JE, Kellner KF, Kluever BM, Avery ML, Humphrey JS, Tillman EA, DeVault TL, Belant JL (2021) Landscape transformations produce favorable roosting conditions for Turkey vultures and black vultures. Sci Rep 11:14793. https://doi.org/10.1038/s41598-021-94045-3
Hirzel AH, Le Laya G, Helfera V, Randina C, Guisan A (2006) Evaluating the ability of habitat suitability models to predict species presences. Ecol Model 99:142–152. https://doi.org/10.1016/j.ecolmodel.2006.05.017
Hosmer DW, Lemeshow S (2004) Applied logistic regression, 2nd edn. Wiley, New York
Ilanloo SS, Khani A, Kafash A, Valizadegan N, Ashrafi S, Loercher F, Ebrahimi E, Yousefi M (2020) Applying opportunistic observations to model current and future suitability of the Kopet Dagh Mountains for a Near Threatened avian scavenger. Avian Biol Res 14:18–26. https://doi.org/10.1177/1758155920962750
iNaturalist users, Ueda K (2020). iNaturalist research-grade observations. iNaturalist.org. Occurrence dataset. https://doi.org/10.15468/ab3s5x accessed via GBIF.org on 2020-10-23
ISFR (2021) India State of Forest Report. Forest Survey of India (MoEFCC), Dehradun, India.
IUCN (2021) IUCN red list of threatened species. https://www.iucnredlist.org/. Accessed 10 May 2021.
Jha KK, Jha R (2020) Habitat suitability mapping for migratory and resident vultures: a case of Indian stronghold and species distribution model. J Wildl Biodivers 4(3):91–111. https://doi.org/10.22120/jwb.2020.120246.1111
Jha R, Jha KK (2021) Habitat prediction modelling for vulture conservation in Gangetic Thar Deccan region of India. Environ Monit Assess 193(8):532. https://doi.org/10.1007/s10661-021-09323-4
Jha R, Jha KK (2023) Environmental factors shaping habitat suitability of Gyps vultures: climate change impact modelling for conservation in India. Ornithol Res. https://doi.org/10.1007/s43388-023-00124-6
Jha KK, Campbell MO, Jha R (2020) Vultures, their population status and some ecological aspects in an Indian stronghold. Notul Sci Biol 12(1):124–142. https://doi.org/10.15835/nsb12110547
Jha R, Kanaujia A, Jha KK (2022) Wintering habitat modelling for conservation of Eurasian vultures in northern India. Nova Geodesia 2(1):22. https://doi.org/10.55779/ng2122
Kaky E, Nolan V, Alatawi A, Gilbert F (2020) A comparison between Ensemble and MaxEnt species distribution modelling approaches for conservation: a case study with Egyptian medicinal plants. Ecol Inform 60:101150. https://doi.org/10.1016/j.ecoinf.2020.101150
Kazmi FA, Shafique F, Hassan MU, Khalid S, Ali N, Akbar N, Batool K, Khalid M, Khawajah S (2022) Ecological impacts of climate change on the snow leopard (Panthera unica) in South Asia. Braz J Biol 82:e240219. https://doi.org/10.1590/1519-6984.240219
Khosravi R, Hemami M-R, Malekian M, Flint AL, Flint LE (2016) Maxent modeling for predicting potential distribution of goitered gazelle in central Iran: the effect of extent and grain size on performance of the model. Turk J Zool 40:574–585
Kumar C, Kaleka AS, Thind SK (2020) Observations on breeding behaviour of a pair of endangered Egyptian Vultures Neophron percnopterus (Linnaeus, 1758) over three breeding seasons in the plains of Punjab, India. J Threat Taxa 12(9):16013–16020. https://doi.org/10.11609/jott.4539.12.9.16013-16020
Kumar S, Stohlgren TJ (2009) MaxEnt modeling for predicting suitable habitat for threatened and endangered tree Canacomyrica monticola in New Caledonia. J Ecol Nat Environ 1:94–98
Kupika OL, Gandiwa E, Kativu S, Nhamo G (2018) Impacts of climate change and climate variability on wildlife resources in Southern Africa: experience from selected protected areas in Zimbabwe. In: Sen B, Grillo O (eds) Selected studies in biodiversity. IntechOpen, London. https://doi.org/10.5772/intechopen.70470
Li X, Wang Y (2013) Applying various algorithms for species distribution modelling. Integr Zool 8:124–135. https://doi.org/10.1111/1749-4877.12000
Lobo JM, Jiménez-Valverde A, Real R (2008) AUC: a misleading measure of the performance of predictive distribution models. Glob Ecol Biogeogr 17:145–151. https://doi.org/10.1111/j.1466-8238.2007.00358.x
Midgley GF, Bond WJ (2015) Future of African terrestrial biodiversity and ecosystems under anthropogenic climate change. Nat Clim Chang 5:823–829
MoEFCC (2020) Action Plan for Vulture Conservation in India, 2020–2025. Ministry of Environment, Forest and Climate Change Government of India, New Delhi
Mohammadi S, Ebrahimi E, Shahriari MM, Bosso L (2019) Modelling current and future potential distributions of two desert jerboas under climate change in Iran. Ecol Inf 52:7–13. https://doi.org/10.1016/j.ecoinf.2019.04.003
Morales NS, Fernández IC, Baca-González V (2017) MaxEnt’s parameter configuration and small samples: are we paying attention to recommendations? A systematic review. PeerJ 5:e3093. https://doi.org/10.7717/peerj.3093
Mori GM, Castillo EB, Guzmán CT, Sánchez DAC, Valqui BKG, Oliva M, Bandopadhyay S, López RS, Briceño NBR (2020) Predictive modelling of current and future potential distribution of the spectacled bear (Tremarctos ornatus) in Amazonas, Northeast Peru. Animals 10:1816. https://doi.org/10.3390/ani10101816
Mushtaq S, Reshi ZA, Shah MA, Charles B (2021) Modelled distribution of an invasive alien plant species differs at different spatiotemporal scales under changing climate: a case study of Parthenium hysterophorus L. Trop Ecol 62:398–417. https://doi.org/10.1007/s42965-020-00135-0
Ogada DL, Keesing F, Virani MZ (2011) Dropping dead: causes and consequences of vulture population declines worldwide. Ann N Y Acad Sci. https://doi.org/10.1111/j.1749-6632.2011.06293.x
Passadore C, Möller LM, Diaz-Aguirre F, Parra GJ (2018) Modelling dolphin distribution to inform future spatial conservation decisions in a marine protected area. Sci Rep 8:15659. https://doi.org/10.1038/s41598-018-34095-2
Patasaraiya MK, Devi RM, Sinha B, Bisaria J, Saran S, Jaiswal R (2021) Understanding the resilience of sal and teak forests to climate variability using NDVI and EVI time series. For Sci 67(2):192–204. https://doi.org/10.1093/forsci/fxaa051
Phillips SJ, Anderson RP, Schapire RE (2006) Maximum entropy modelling of species geographic distribution. Ecol Model 190:231–259. https://doi.org/10.1016/j.ecolmodel.2005.03.026
Phipps WL, Diekmannb M, MacTavishc LM, Mendelsohnd JM, Naidoo V, Wolter K, Yarnell RW (2017) Due south: a first assessment of the potential impacts of climate change on Cape vulture occurrence. Biol Cons 210:16–25. https://doi.org/10.1016/j.biocon.2017.03.028
Prakash V, Galligan TH, Chakraborty SS, Dave R, Kulkarni MD, Prakash N, Shringarpure RN, Ranade SP, Green RE (2017) Recent changes in populations of critically endangered Gyps vultures in India. Bird Conserv Int 29(1):55–70. https://doi.org/10.1017/S0959270917000545
Preston KL, Rotenberry JT, Redak RA, Michael F, Allen MF (2008) Habitat shifts of endangered species under altered climate conditions: importance of biotic interactions. Global Chang Biol 14:2501–2515. https://doi.org/10.1111/j.1365-2486.2008.01671.x
Ramesh T, Sankar K, Qureshi Q (2011) Status of vultures in Mudumalai Tiger Reserve, Western Ghats, India. Forktail 27:96–97
Ravindranath NH, Joshi NV, Sukumar R, Saxena A (2006) Impact of climate change on forest in India. Curr Sci 90(3):354–361
Saenz-Jimenez F, Rojas-Soto O, Perez-Torres J, Martinez-Meyer E, Sheppard JK (2020) Effects of climate change and human influence in the distribution and range overlap between two widely distributed avian scavengers. Bird Conserv Int 31(1):77–95. https://doi.org/10.1017/S0959270920000271
Santini L, Benítez-López A, Maiorano L, Čengić M, Huijbregts MAJ (2021) Assessing the reliability of species distribution projections in climate change research. Divers Distrib 27:1035–1050. https://doi.org/10.1111/ddi.13252
Schmitt S, Pouteau R, Justeau D, de Boisseu F, Birnbaum P (2017) SSDM: an R package to predict distribution of species richness and composition based on stacked species distribution models. Methods Ecol Evol 8:1795–1803. https://doi.org/10.1111/2041-210X.12841
Schultz P (2007) Does bush encroachment impact foraging success of the critically endangered Namibian Population of the Cape Vulture Gyps coprotheres? University of Cape Town. http://hdl.handle.net/20.500.11892/49885
Sharma PD (2005) Ecology and environment. Rastogi Publications, Meerut
Sinha A, Kumar A, Kanaujia A (2017) Red-Headed vulture: a solitary scavenger. Int J Recent Sci Res 8(7):18737–18741
Subedi TR, Virani MZ, Gurung S, Buij R, Baral HS, Buechley ER, Anadón JD, Sah SAM (2018) Estimation of population density of bearded vultures using line-transect distance sampling and identification of perceived threats in the Annapurna Himalaya range of Nepal. J Rapt Res 52(4):443–453. https://doi.org/10.3356/JRR-18-25.1
Sullivan BL, Wood CL, Iliff MJ, Bonney RE, Fink D, Kelling S (2009) eBird: a citizen-based bird observation network in the biological sciences. Biol Conserv 142:2282–2292. https://doi.org/10.1016/j.biocon.2009.05.006
Thakur ML, Narang SK (2012) Population status and habitat-use pattern of Indian whitebacked Vulture (Gyps bengalensis) in Himachal Pradesh, India. J Ecol Nat Environ 4(7):173-180.
Thapa S, Baral S, Hu Y, Huang Z, Yue Y, Dhakal M, Jnawali SR, Chettri N, Racey PA, Yu W, Wu Y (2021) Will climate change impact distribution of bats in Nepal Himalayas? Glob Ecol Conserv 26:e01483. https://doi.org/10.1016/j.gecco.2021.e01483
USGS EROS (2018) Shuttle radar topography mission 1 arc-second global. https://doi.org/10.5066/F7PR7TFT
Veloz SD (2009) Spatially autocorrelated sampling falsely inflates measures of accuracy for presence-only niche models. J Biogeogr 36:2290–2299
Virani MZ, Monadjem A, Thomsett S, Kendall C (2012) Seasonal variation in breeding Ruppell’s Vultures Gyps rueppellii at Kwenia, southern Kenya and implications for conservation. Bird Conserv Int 22:260–269
Wisz MS, Hijmans RJ, Li J, Peterson AT, Graham CH, Guisan A, NCEAS PSDWG (2008) Effects of sample size on the performance of species distribution models. Divers Distrib 14:763–773. https://doi.org/10.1111/j.1472-4642.2008.00482.x
Zeng Q, Zhang Y, Sun G, Duo H, Wen L, Lei G (2015) Using species distribution model to estimate the wintering population size of the endangered scaly-sided merganser in China. PLoS ONE 10(2):e0117307. https://doi.org/10.1371/journal.pone.0117307
Zhang J, Jiang F, Li G, Qin W, Li S, Gao H, Cai Z, Lin G, Zhang T (2019a) Maxent modeling for predicting the spatial distribution of three raptors in the Sanjiangyuan National Park, China. Ecol Evol 9:6643–6654. https://doi.org/10.1002/ece3.5243
Zhang K, Zhang Y, Tao J (2019b) Predicting the potential distribution of Paeonia veitchii (Paeoniaceae) in China by incorporating climate change into a Maxent model. Forests 10:190. https://doi.org/10.3390/f10020190
Zhang K, Zhang Y, Jia D, Tao J (2020) Species distribution modeling of Sassafras tzumu and implications for forest management. Sustainability 12:4132. https://doi.org/10.3390/su12104132
Acknowledgements
Not applicable. The experiment complies with the current laws of the country.
Funding
The authors declare that no funds, grants, or other support was received during the preparation of this manuscript.
Author information
Authors and Affiliations
Contributions
Both the authors contributed to the study conception and design. Material preparation, data collection, and analysis were performed by both the authors. The first draft of the manuscript was written by RJ and KKJ commented on previous versions of the manuscript. Both the authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Jha, R., Jha, K.K. Prediction of habitat suitability dynamics and environmental factors of non-Gyps vultures for conservation in floristic landscapes of India. Landscape Ecol Eng 20, 19–31 (2024). https://doi.org/10.1007/s11355-023-00575-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11355-023-00575-5