
Abstract
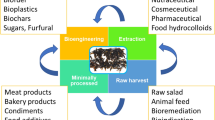
Seaweed is more than the wrap that keeps rice together in sushi. Seaweed biomass is already used for a wide range of other products in food, including stabilising agents. Biorefineries with seaweed as feedstock are attracting worldwide interest and include low-volume, high value-added products and vice versa. Scientific research on bioactive compounds in seaweed usually takes place on just a few species and compounds. This paper reviews worldwide research on bioactive compounds, mainly of nine genera or species of seaweed, which are also available in European temperate Atlantic waters, i.e. Laminaria sp., Fucus sp., Ascophyllum nodosum, Chondrus crispus, Porphyra sp., Ulva sp., Sargassum sp., Gracilaria sp. and Palmaria palmata. In addition, Undaria pinnatifida is included in this review as this is globally one of the most commonly produced, investigated and available species. Fewer examples of other species abundant worldwide have also been included. This review will supply fundamental information for biorefineries in Atlantic Europe using seaweed as feedstock. Preliminary selection of one or several candidate seaweed species will be possible based on the summary tables and previous research described in this review. This applies either to the choice of high value-added bioactive products to be exploited in an available species or to the choice of seaweed species when a bioactive compound is desired. Data are presented in tables with species, effect and test organism (if present) with examples of uses to enhance comparisons. In addition, scientific experiments performed on seaweed used as animal feed are presented, and EU, US and Japanese legislation on functional foods is reviewed.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Whilst food has long been used to improve health, our knowledge of the relationship between food components and health is now being used to improve food. Although most foods can be considered “functional”, in the context of this review, the term is reserved for foods and food components that have been demonstrated to provide specific health benefits beyond basic nutrition. Widespread interest in select foods that might promote health has resulted in the use of the term “functional foods”. Food and nutrition science has moved from identifying and correcting nutritional deficiencies to designing foods that promote optimal health and reduce the risk of disease. Today’s science and technology can be used to provide many additional functional foods, and future scientific and technological advances promise an even greater range of health benefits for consumers. Functional foods can provide health benefits by reducing the risk of chronic diseases and enhancing the ability to manage chronic diseases, thus improving the quality of life. Functional foods also can promote growth and development and enhance performance.
Seaweeds are known for their richness in polysaccharides, minerals and certain vitamins (Arasaki and Arasaki 1983), but they also contain bioactive substances like polysaccharides, proteins, lipids and polyphenols, with antibacterial, antiviral and antifungal properties, as well as many others (Kumar et al. 2008b). This gives seaweed great potential as a supplement in functional food or for the extraction of compounds. Physiologically active substances in marine algae are classified into two types based on the difference in the mechanisms: (1) non-absorbed high-molecular materials like dietary fibres and (2) low-molecular materials, which are absorbed and which affect the maintenance of human homeostasis directly (Murata and Nakazoe 2001).
Nutritional elements
The moisture content of fresh marine algae is very high and can account for up to 94% of the biomass (Table 1). Though, like other plants, marine algae contain nutritional elements such as proteins, lipids, carbohydrates, vitamins and minerals, the content of these elements varies depending on season and the area of production (Connan et al. 2004; Khan et al. 2007; Marinho-Soriano et al. 2006; Murata and Nakazoe 2001; Zubia et al. 2008). Researchers from the Norwegian Institute of Seaweed Research have been studying seasonal variations of the biochemical composition of the large brown seaweed species in particular since the 1950s. The ash content in seaweed is high compared to vegetables (Table 1; Murata and Nakazoe 2001). The ash content includes macro-minerals and trace elements. Ash content of blades of Saccharina latissima, Laminaria spp. and Alaria esculenta is lowest during September, October and November and highest during spring (February to June); however, it varies only slightly in the stipes over the year. Dry weight was lowest from January to March and highest in July to September for these species (Haug and Jensen 1954; Jensen and Haug 1956). These seasonal and environmental variations in the composition of seaweed make generalisations impossible. Nevertheless, this review summarises scientific data on the biochemical composition and bioactivity of compounds in seaweed, although some data might just provide the concentration as a snapshot and not the seasonal variations, for example.
Polysaccharides
Marine algae contain large amounts of polysaccharides, notably cell wall structural, but also mycopolysaccharides and storage polysaccharides (Kumar et al. 2008b; Murata and Nakazoe 2001). Polysaccharides are polymers of simple sugars (monosaccharides) linked together by glycosidic bonds, and they have numerous commercial applications in products such as stabilisers, thickeners, emulsifiers, food, feed, beverages, etc. (McHugh 1987; Tseng 2001). The total polysaccharide concentrations in the seaweed species of interest range from 4% to 76% of dry weight (Table 2). The highest contents are found in species such as Ascophyllum, Porphyra and Palmaria; however, green seaweed species such as Ulva also have a high content, up to 65% of dry weight.
Seaweeds are low in calories from a nutritional perspective. The lipid content is low, and even though the carbohydrate content is high, most of this is dietary fibres and not taken up by the human body. However, dietary fibres are good for human health as they make an excellent intestinal environment (Mouritsen 2009).
The cell wall polysaccharides mainly consist of cellulose and hemicelluloses, neutral polysaccharides, and are thought to physically support the thallus in water. The building blocks needed to support the thalli of seaweed in water are less rigid/strong compared to terrestrial plants and trees. The cellulose and hemicellulose content of the seaweed species of interest in this review is 2–10% and 9% dry weight, respectively. Lignin is only found in Ulva sp. at concentrations of 3% dry weight (Table 2).
The cell wall and storage polysaccharides are species-specific, and examples of concentrations are given in Table 3. Green algae contain sulphuric acid polysaccharides, sulphated galactans and xylans, brown algae alginic acid, fucoidan (sulphated fucose), laminarin (β-1,3 glucan) and sargassan and red algae agars, carrageenans, xylans, floridean starch (amylopectin-like glucan), water-soluble sulphated galactan, as well as porphyran as mucopolysaccharides located in the intercellular spaces (Table 3; Kumar et al. 2008b; Murata and Nakazoe 2001). Contents of both total and species-specific polysaccharides show seasonal variations. The mannitol content varied markedly in the fronds of Saccharina and Laminaria species, with maximum amounts found during summer and autumn, from June to November. Laminaran showed extreme variations during the year, with very small amounts or none at all in February to June and maximum in September to November (Haug and Jensen 1954; Jensen and Haug 1956). The maximum content of alginic acid in the fronds of Saccharina and Laminaria was generally found from March to June and the minimum from September to October (Haug and Jensen 1954). However, the highest contents of alginic acid were found during winter in other seasonal studies on Laminaria from the same areas in Norway (Jensen and Haug 1956).
Further investigations on the hydrolysates of some brown algae showed complex mixtures of monosaccharides. The components of galactose, glucose, mannose, fructose, xylose, fucose and arabinose were found in the total sugars in the hydrolysates. The glucose content was 65%, 30% and 20% of the total sugars in an autumn sample of 50 individual plants of Saccharina, Fucus (serratus and spiralis) and Ascophyllum, respectively (Jensen 1956).
Several other polysaccharides are present in and utilised from seaweed, e.g. furcellaran, funoran, ascophyllan and sargassan; however, these are not described in this review. Seaweed polysaccharides are separated into dietary fibres, hydrocolloids, etc. in the following paragraphs, even though the polysaccharides belong to more than just one of the functional groups.
Dietary fibres
The dietary fibres are very diverse in composition and chemical structure as well as in their physicochemical properties, their ability to be fermented by the colonic flora, and their biological effects on animal and human cells (Lahaye and Kaeffer 1997). Edible seaweed contain 33–62% total fibres on a dry weight basis, which is higher than the levels found in higher plants, and these fibres are rich in soluble fractions (Lahaye 1991; Dawczynski et al. 2007). The dietary fibres included in marine algae are classified into two types, i.e. insoluble such as cellulose, mannans and xylan, and water-soluble dietary fibres such as agars, alginic acid, furonan, laminaran and porphyran. The total content of dietary fibres is 58% dry weight for Undaria, 50% for Fucus, 30% for Porphyra and 29% for Saccharina (Murata and Nakazoe 2001). Fucus and Laminaria have the highest content of insoluble dietary fibres (40% and 27%, respectively), and Undaria pinnatifida (wakame), Chondrus and Porphyra have the highest content of soluble dietary fibres (15–22%; Fleury and Lahaye 1991).
The undigested polysaccharides of seaweed can form important sources of dietary fibres, although they might modify the digestibility of dietary protein and minerals. Apparent digestibility and retention coefficients of Ca, Mg, Fe, Na and K were lower in seaweed-fed rats (Urbano and Goñi 2002). The seaweed dietary fibres contain some valuable nutrients and substances, and there has been a deal of interest in seaweed meal, functional foods and nutraceuticals for human consumption (McHugh 2003) because, among other things, polysaccharides show anti-tumour and anti-herpetitic bioactivity; they are potent as an anticoagulant and decrease low-density lipid (LDL)-cholesterols in rats (hypercholesterolemia); they prevent obesity, large intestine cancer and diabetes; and they have antiviral activities (Lee et al. 2004; Murata and Nakazoe 2001; Amano et al. 2005; Athukorala et al. 2007; Ghosh et al. 2009; Murata and Nakazoe 2001; Ye et al. 2008; Tables 4, 5 and 6). Moreover, glucose availability and absorption are delayed in the proximal small intestine after the addition of soluble fibres, thus reducing postprandial glucose levels (Jenkins et al. 1978). Water-insoluble polysaccharides (celluloses) are mainly associated with a decrease in digestive tract transit time (see also later chapter on the digestibility of polysaccharides; Mabeau and Fleurence 1993).
Alginates
Alginates were discovered in the 1880s by a British pharmacist, E.C.C. Stanford; industrial production began in California in 1929. Nowadays, the key industrial producers of alginates are the US, Great Britain, Norway, Canada, France, Japan and China; the annual sales volume of alginates in the world is about 26,500 t out of a total volume of 86,000 t hydrocolloids (Bixler and Porse 2010). Production of carbohydrates that dissolve in water is a growing industry. The three major groups of phycocolloids are alginates, carrageenan and agar, and these are used for applications such as thickening aqueous solutions, forming gels and water-soluble films, and stabilising products such as ice cream, tooth paste, mayonnaise, etc. (FAO 2008; Tseng 2001).
Algins/alginates are extracted from brown seaweed and are available in both acid and salt forms. The acid form is a linear polyuronic acid and is referred to as alginic acid, whereas the salt form is an important cell wall component in all brown seaweed, constituting up to 40–47% of the dry weight of algal biomass (Table 4;Arasaki and Arasaki 1983; Rasmussen and Morrissey 2007). Various industries related to food processing, pharmaceuticals, feed and cosmetics use alginic acid extracted from Saccharina and Undaria. It has been reported that alginic acid leads to a decrease in the concentration of cholesterol, exerts an anti-hypertension effect, can prevent absorption of toxic chemical substances, and plays a major role as dietary fibre for the maintenance of animal and human health (Kim and Lee 2008; Murata and Nakazoe 2001; Nishide and Uchida 2003). These dietary polysaccharides are not found in any land plants. They help protect against potential carcinogens and they clear the digestive system and protect the surface membranes of the stomach and intestine. They absorb substances such as cholesterol, which are then eliminated from the digestive system (Burtin 2003; Ito and Tsuchida 1972) and result in hypocholesterolemic and hypolipidemic responses (Kiriyama et al. 1969; Lamela et al. 1989; Panlasigui et al. 2003). This is often coupled with an increase in the faecal cholesterol content and a hypoglycaemic response (Dumelod et al. 1999; Ito and Tsuchida 1972; Nishide et al. 1993).
Alginates, fucoidans and laminarin extracts were tested against nine bacteria, including Escherichia coli, Staphylococcus, Salmonella and Listeria. They appeared to be effective against E. coli and Staphylococcus. Sodium alginate seemed to demonstrate a strong antibacterial element. It not only binds but also kills the bacteria. Studies conducted on seaweed extracts found that fucoidan appeared to function as a good prebiotic (a substance that encourages the growth of beneficial bacteria in the intestines). An anti-inflammatory effect from some of the extracts has also been found, and so far, no toxic effects have emerged in use for human health (Hennequart 2007). Furthermore, alginates with molecular weights ≥50 kDa could prevent obesity, hypocholesterolemia and diabetes (Kimura et al. 1996). Clinical observations of volunteers who were 25–30% overweight showed that alginate, a drug containing alginic acid, significantly decreased body weight (Zee et al. 1991). In type II diabetes treatment, taking 5 g of sodium alginate every morning was found to prevent a postprandial increase of glucose, insulin, and C-peptide levels and slowed down gastric transit (Torsdottir et al. 1991). Meal supplemented with 5% kelp alginates decreased glucose absorption balance over 8 h in pigs. Similar studies have been done on rats and humans (Vaugelade et al. 2000). Another health effect is that the binding property of alginic acid to divalent metallic ions is correlated to the degree of the gelation or precipitation in the range of Ba<Pb<Cu<Sr<Cd<Ca<Zn<Ni<Co<Mn<Fe<Mg. No intestinal enzymes can digest alginic acid. This means that heavy metals taken into the human body are gelated or rendered insoluble by alginic acid in the intestines and cannot be absorbed into the body tissue (Arasaki and Arasaki 1983).
In several countries such as the US, Germany, Japan, Belgium and Canada, the use of alginic acid and its derivatives for the treatment of gastritis and gastroduodenal ulcers, as well as the use of alginates as anti-ulcer remedies, is protected by patents (Bogentoff 1981; Borgo 1984; Reckitt and Colman Products Ltd. 1999; Sheth 1967). Several products of alginate-containing drugs have been shown to effectively suppress postprandial (after eating) and acidic refluxes, binding of bile acids and duodenal ulcers in humans. Examples are “Gaviscon” (sodium alginate, sodium bicarbonate and calcium carbonate), “Algitec” (sodium alginate and cimetidine, an H2 antagonist) and “Gastralgin” (alginic acid, sodium alginate, aluminium hydroxide, magnesium hydroxide and calcium carbonate; Khotimchenko et al. 2001; Washington and Denton 1995). Clinical trials showed that sodium alginate promotes the regeneration of the mucous membrane in the stomach, suppresses inflammation, eradicates colonies of Helicobacter pylori in the mucous membrane and normalises non-specific resistance of the latter in 4- to 15-year-old children. It also promotes restoration of the intestinal biocenosis (Miroshnichenko et al. 1998). Other studies show positive dietary effects of alginates on faecal microbial fauna, changes in concentrations of compounds and acids, and prebiotic properties that can promote health (Terada et al. 1995; Wang et al. 2006).
Sodium alginate is often used as a powder, either pure or mixed with other drugs, on septic wounds. The polysaccharide base stimulates reparative processes, prepares the wound for scarring, and displays protective and coating effects, shielding mucous membranes and damaged skin against irritation from unfavourable environments. Calcium alginate promotes the proliferation of fibroblasts and inhibits the proliferation of microvascular endothelial cells and keratinocytes (Doyle et al. 1996; Glyantsev et al. 1993; Swinyard and Pathak 1985). Profound wound healing effects have also been reported for a gelatine–alginate sponge impregnated with antiseptics and antibiotics (Choi et al. 1999).
Alginates absorbing haemostatic effect has also been exploited in surgery. Gauze dressings, cotton, swabs and special materials impregnated with a solution of sodium alginate are produced and used for external use and for application onto bleeding points during abdominal operations on parenchymatous organs (Khotimchenko et al. 2001; Savitskaya 1986). Studies on the effect of alginate on prothrombotic blood coagulation and platelet activation have shown that the degree of these effects depends on the ratio between the mannuronic and guluronic chains in the molecule, as well as on the concentration of calcium. However, a zinc ion containing alginate was shown to have the most profound haemostatic effects (Segal et al. 1998). A “poraprezinc–sodium alginate suspension” has been suggested as a high-performance mixture for the treatment of severe gingivostomatitis (cold sores) complicated by haemorrhagic erosions and ulcers (Katayama et al. 1998). When applied to the tooth surface, alginate fibres swell to form a gel-like substance, a matrix for coagulation. Alginate dressings are used to pack sinuses, fistulas and tooth cavities (Reynolds and Prasad 1982).
Furthermore, the algins have anticancer properties (Murata and Nakazoe 2001), and a bioactive food additive “Detoxal”, containing calcium alginate, has antitoxic effects on hepatitis. This drug decreases the content of lipid peroxidation products and normalises the concentrations of lipids and glycogen in the liver (Khotimchenko et al. 2001). Mannuronate surfactants derived from alginate can be applicable in fields as cosmetics, health and agrochemistry (Benvegnu and Sassi 2010).
Carrageenans
Carrageenans are a group of biomolecules composed of linear polysaccharide chains with sulphate half-esters attached to the sugar unit. These properties allow carrageenans to dissolve in water, form highly viscous solutions and remain stable over a wide pH range. There are three general forms (kappa, lambda and iota), each with its own gelling property (Rasmussen and Morrissey 2007). Chondrus crispus and Kappaphycus sp. contain up to 71% and 88% of carrageenan, respectively (Chopin et al. 1999; Rodrigueza and Montaño 2007). Food applications for carrageenans (E 407) are many, including canned foods, dessert mousses, salad dressings, bakery fillings, ice cream, instant desserts and canned pet foods. Industrial applications for purified extracts of carrageenans are equally diverse. They are used in the brewing industry for clarifying beer, wines and honeys, although less commonly than previously. From a human health perspective, it has been reported that carrageenans have anti-tumour and antiviral properties (Skoler-Karpoff et al. 2008; Vlieghe et al. 2002; Yan et al. 2004; Zhou et al. 2006b). Furthermore, Irish moss or carrageen (C. crispus and Mastocarpus stellatus) has a large number of medical applications, some of which date from the 1830s. It is still used in Ireland to make traditional medicinal teas and cough medicines to combat colds, bronchitis and chronic coughs. It is said to be particularly useful for dislodging mucus and has antiviral properties. Carrageenans are also used as suspension agents and stabilisers in other drugs, lotions and medicinal creams. Other medical applications are as an anticoagulant in blood products and for the treatment of bowel problems such as diarrhoea, constipation and dysentery. They are also used to make internal poultices to control stomach ulcers (Morrissey et al. 2001).
New research on the biocide properties shows that applications of carrageenan gels from C. crispus may block the transmission of the HIV virus as well as other STD viruses such as gonorrhoea, genital warts and the herpes simplex virus (HSV; Caceres et al. 2000; Carlucci et al. 1997; Luescher-Mattli 2003; Shanmugam and Mody 2000; Witvrouw and DeClercq 1997). In addition, carrageenans are good candidates for use as vaginal microbicides because they do not exhibit significant levels of cytotoxicity or anticoagulant activity (Buck et al. 2006; Zeitlin et al. 1997). Results of sexual lubricant gels raised the possibility that the use of such lubricant products, or condoms lubricated with carrageenan-based gels, could block the sexual transmission of human papillomavirus (HPV) types that can cause cervical cancer and genital warts. However, carrageenan inhibition of herpes simplex virus and HIV-1 infectivity were demonstrated as about 1,000-fold higher than the IC50’s observed for genital HPVs in vitro (Witvrouw and DeClercq 1997; Luescher-Mattli 2003). A carrageenan-based vaginal microbicide called Carraguard has been shown to block HIV and other sexually transmitted diseases in vitro. Massive clinical trials by the Population Council Centre began in two severely affected African countries—Botswana and South Africa—in 2002. Carraguard entered phase III clinical trials involving 6,000 non-pregnant, HIV-negative women in South Africa and Botswana in 2003 (Spieler 2002). This trial showed that the microbiocide Carraguard® was safe for vaginal use, but did not demonstrate that it is effective in preventing male-to-female HIV transmission during vaginal intercourse (Population Council 2008).
Many reports of anticoagulant activity and inhibited platelet aggregation of carrageenan exist (Hawkins et al. 1962; Hawkins and Leonard 1963; Kindness et al. 1979). Among the carrageenan types, λ-carrageenan (primarily from C. crispus) has approximately twice the activity of unfractionated carrageenan and four times the activity of κ-carrageenan (E. cottoni and Eucheuma spinosum). The most active carrageenan has approximately one fifteenth the activity of heparin (Hawkins et al. 1962), but the sulphated galactan from Grateloupa indica collected from Indian waters exhibited an anticoagulant activity as potent as heparin (Sen et al. 1994).
The principal basis of the anticoagulant activity of carrageenan appeared to be an anti-thrombotic property. λ-Carrageenan showed greater anti-thrombotic activity than κ-carrageenan, probably due to its higher sulphate content, whereas the activity of the unfractionated material remained between the two. It was found that the toxicity of carrageenans depended on the molecular weight and not the sulphate content. Similar results were obtained with λ-carrageenan of Phyllophora brodiaei which gave the highest blood anticoagulant activity (Sen et al. 1994). In addition, the hypoglycaemic effect of carrageenan may prove useful in the prevention and management of metabolic conditions such as diabetes (Dumelod et al. 1999).The use of carrageenan for food applications started almost 600 years ago. Due to its long and safe history of use, carrageenan is generally recognised as safe (GRAS) by experts from the US Food and Drug Administration (21 CFR 182.7255) and is approved as a food additive (21 CFR 172.620). The WHO Joint Expert Committee of Food Additives has concluded that it is not necessary to specify an acceptable daily intake limit for carrageenans (van de Velde et al. 2002). Although carrageenans are used widely as a food ingredient, they are also used in experimental research in animals where they induce pleurisy and ulceration of the colon (Noa et al. 2000). Furthermore, carrageenans can cause reproducible inflammatory reaction and remain a standard irritant for examining acute inflammation and anti-inflammatory drugs. Two test systems are used widely for the evaluation of non-steroidal anti-inflammatory drugs and cyclooxygenase activity: (1) the carrageenan air pouch model and (2) the carrageenan-induced rat paw oedema assay (Dannhardt and Kiefer 2001). The role of carrageenans in the promotion of colorectal ulceration formation is controversial, and much seems to depend on the molecular weight of the carrageenan used. The International Agency for Research on Cancer classified degraded carrageenan as a possible human carcinogen, but native carrageenan remains unclassified in relation to a causative agent of human colon cancer and has GRAS status (Carthew 2002; Tobacman 2001).
Agar
Agar is a mixture of polysaccharides, which can be composed of agarose and agropectin, with similar structural and functional properties as carrageenans. It is extracted from red seaweed such as Gelidium spp. and Gracilaria spp. (FAO 2008; Rasmussen and Morrissey 2007; Jeon et al. 2005). The agar content in Gracilaria species can reach 31% (Table 4). Agar–agar is a typical and traditional food material in Japan and is used for cooking and Japanese-style confectionary. In addition, agar–agar is used in the manufacture of capsules for medical applications and as a medium for cell cultures, etc. It has been reported that agar–agar leads to decreases in the concentration of blood glucose and exerts an anti-aggregation effect on red blood cells. It has also been reported to affect the absorption of ultraviolet rays (Murata and Nakazoe 2001). Anti-tumour activity was also found in an agar-type polysaccharide from cold water extraction of another Gracilaria species, and hydrolysates of agar resulted in agaro-oligosaccharides with activity against α-glucosidase and antioxidant ability (Chen et al. 2005; Fernandez et al. 1989). Agarose can be separated from the agar with a yield of 42%, and the agar content varied seasonally from 26% to 42% in Gelidium spp. in another experiment (Mouradi-Givernaud et al. 1992; Jeon et al. 2005). Agaro-oligosaccharides have also been shown to suppress the production of a pro-inflammatory cytokine and an enzyme associated with the production of nitric oxide (Enoki et al. 2003).
Fucoidan/fucans/fucanoids
Fucoidans are a group of polysaccharides (fucans) primarily composed of sulphated l-fucose with <10% of other monosaccharides. They are widely found in the cell walls of brown seaweed, but not in other algae or higher plants (Berteau and Mulloy 2003). Fucoidan is considered as a cell wall-reinforcing molecule and seems to be associated with protection against the effects of desiccation when the seaweed is exposed at low tide. Fucoidans were first isolated by Kylin almost one century ago and have interesting bioactivities (Kylin 1913). Fucus vesiculosus contains the highest concentration of fucoidans (up to 20% on a dry weight basis) (Table 5). Fucanoids can make up more than 40% of dry weight of isolated algal cell walls and can easily be extracted using either hot water or an acid solution (Berteau and Mulloy 2003). Fucoidan is viscous in very low concentrations and susceptible to breakdown by diluted acids and bases.
A Tasmanian company, Marinova Pty Ltd., is able to supply commercial volumes of fucoidan extract and their derivates, formulated to purity levels of up to 95%. All fucoidans of Undaria, Lessonia, Macrocystis, Cladosiphon, Durvillea, Laminaria, Ecklonia, Fucus, Sargassum, Ascophyllum and Alaria are Halal- and Kosher-certified. Marinova has isolated fucoidans from a range of species (species-specific) and can provide characterised fractions for either investigational research or as ingredients for nutraceutical and cosmetic applications. Different therapeutic profiles are primarily due to the molecular structure. The company has developed the Maritech™ cold water extraction process which maintains the integrity of fucoidans and produces nature-equivalent high-molecular-weight molecules with optimal bioactivity. Solvent-based extraction, which is commonly used, causes degradation of fucoidans and limits the activity of these molecules in biological assays (Marinova 2010).
Although the major physiological purposes of fucans in the algae are not thoroughly understood, they are known to possess numerous biological properties with potential human health applications (Berteau and Mulloy 2003). The list of bioactivity of fucoidan for human health is long fucoidan found in seaweed such as Undaria and Laminaria shows anticoagulant, antiviral and anticancer properties (Table 5; Chevolot et al. 1999; Zhuang et al. 1995).
Fucoidan preparations have been proposed as an alternative to the injectable anticoagulant heparin because fucoidan originates from plant matter and is less likely to contain infectious agents such as viruses (Berteau and Mulloy 2003). No toxicological changes were observed when 300 mg kg−1 body wt day−1 fucoidan was administered to rats; however, significantly prolonged blood-clotting times were observed when concentrations were increased threefold (Li et al. 2005). The biological activity (e.g. antioxidant and anticoagulant) of sulphated polysaccharides is not only related to molecular weight and sulphated ester content (role in the charge of the molecule), as previously determined, but also to glucuronic acid and fucose content, together with the position of the sulphate groups on the sugar resides (Berteau and Mulloy 2003; Li et al. 2005; Zhao et al. 2008). A large molecular weight is required to achieve anticoagulant activity as fucoidan needs a long sugar chain in order to be able to bind the thrombin (coagulation protein in the blood stream). Some researchers have measured fucoidan’s molecular weight at approximately 100 kDa, whilst others have observed a molecular weight of 1,600 kDa (Rioux et al. 2007). The native fucoidan from Lessonia vadosa with a molecular weight of 320 kDa showed good anticoagulant activity compared to a smaller depolymerised fraction with a molecular weight of 32 kDa, which presented weaker anticoagulant activity (Li et al. 2008a). Some structural features of fucoidan are most likely required for certain specific activities.
Fucoidan stimulates the immuno system in several ways (Table 5), and the numerous important biological effects of fucoidans are related to their ability to modify cell surface properties (Usov et al. 2001). Oral intake of the fucoidans present in dietary brown seaweed might take the protective effects through direct inhibition of viral replication and stimulation of the immune system (innate and adaptive) functions (Hayashi et al. 2008). Fucoidan has been found to restore the immune functions of immune-suppressed mice, act as an immunomodulator directly on macrophage, T lymphocyte, B cell, natural killer (NK) cells (Wang et al. 1994), promote the recovery of immunologic function in irradiated rats (Wu et al. 2003), induce the production of interleukin 1 (IL-1) and interferon-γ (IFN-γ) in vitro, and promote the primary antibody response in sheep red blood cells in vivo (Yang et al. 1995).
The mechanism of antiviral activities of fucoidan is to inhibit viral sorption so as to inhibit viral-induced syncytium formation (Mandal et al. 2007). Sulphate is necessary for the antiviral activity, and sulphate located at C-4 of (1-3)-linked fucopyranosyl units appears to be very important for the anti-herpetic activity of fucoidan (Mandal et al. 2007). Some antiviral properties of sulphated fucans have also been characterised, for example, inhibition of infection of HIV, HSV (Iqbal et al. 2000; Mandal et al. 2007; Witvrouw and DeClercq 1997), poliovirus III, adenovirus III, ECH06 virus, coxsackie B3 virus, coxsackie A16 (Li et al. 1995), cytomegalovirus and bovine viral diarrhoea virus (Iqbal et al. 2000).
Fucoidan is known to have anti-tumour effects, but its mode of action is not fully understood. A study done by Alekseyenko et al. 2007 demonstrated that when 10 mg/kg of fucoidan was administered in mice with transplanted Lewis lung adenocarcinoma, it produced moderate anti-tumour and anti-metastatic effects (Li et al. 2008a, b). These polyanionic polysaccharides have anti-angiogenesis, antiproliferation for tumour cells, they inhibit tumour growth and reduce tumour size (Ellouali et al. 1993; Li et al. 2008a), inhibit tumour cell adhesion to various substrata (Liu et al. 2005), and have direct anticancer effects on human HS-Sultan cells through caspase and ERK pathways (Aisa et al. 2005).
Besides directly inhibiting the growth of tumour cells, fucoidans can also restrain the development and diffusion of tumour cells through enhancing the body’s immunomodulatory activities because fucoidan mediates tumour destruction through type 1 T-helper (Th1) cell and NK cell responses (Maruyama et al. 2006). In addition, at a dose of 25 mg kg−1, fucoidan potentiated the toxic effect of cyclophosphamide used to treat various types of cancer and some autoimmune disorders (Alekseyenko et al. 2007).
Many studies suggest that fucoidan has potential for use as an anti-inflammatory agent. A study showed that fucoidan treatment led to less severe symptoms in the early stages of Staphylococcus aureus-triggered arthritis in mice, but delayed phagocyte recruitment and decreased clearance of the bacterium (Verdrengh et al. 2000). Additionally, injection of fucoidan into sensitised mice before hapten challenge can reduce contact hypersensitivity reactions (Nasu et al. 1997). Furthermore, recruitment of leukocytes into cerebrospinal fluid in a meningitis model is reduced by fucoidan (Granert et al. 1999), as is IL-1 production in a similar model (Ostergaard et al. 2000).
Fucoidan can act as a ligand for either L- or P-selectins, both of which interact with the sulphated oligosaccharides, and this interaction has physiological consequences that could be therapeutically beneficial (Omata et al. 1997). Selectins are a group of lectins (sugar-binding proteins) that interact with oligosaccharides clustered on cell surfaces during the margination and rolling of leukocytes prior to firm adhesion, extravasation and migration to sites of infection (Lasky 1995).
In addition, fucoidan is an excellent natural antioxidant and presents significant antioxidant activity in experiments in vitro. In recent years, sulphated polysaccharides from the marine algae Porphyra haitanesis (Zhang et al. 2003), Ulva pertusa (Qi et al. 2005a, b), F. vesiculosus (Ruperez et al. 2002), Laminaria japonica (Xue et al. 2004) and Ecklonia kurome (Hu et al. 2001) have been demonstrated to possess antioxidant activity. There are few reports, however, detailing the relationship between the structure and antioxidant activity of sulphated polysaccharides from marine algae. Fucan showed low antioxidant activity relative to fucoidan (Rocha de Souza et al. 2007), and as mentioned previously, the ratio of sulphate content/fucose and the molecular weight were effective indicators of the antioxidant activity of the samples (Wang et al. 2008). Fucoidan may have potential for preventing free radical-mediated diseases such as Alzheimer’s and the ageing process. Previously, fucoidan was extracted from L. japonica, a commercially important algae species in China. Three sulphated polysaccharide fractions were successfully isolated through anion exchange column chromatography and had their antioxidant activities investigated employing various established in vitro systems, including superoxide and hydroxyl radical scavenging activity, chelating ability and reducing power (Wang et al. 2008). All fractions were more effective than the unprocessed fucoidan. Two galactose-rich fractions had the most potent scavenging activity against superoxide (generated in the PMS-NADH system) and hydroxyl radicals, with EC50 values of 1.7 μg mL−1 and 1.42 mg mL−1, respectively. One of these fractions also showed the strongest ferrous ion-chelating ability at 0.76 mg mL−1 (Wang et al. 2008). Additionally, fucoidan (homofucan) from F. vesiculosus and fucans (heterofucans) from Padina gymnospora had an inhibitory effect on the formation of hydroxyl radical and superoxide radical (Rocha de Souza et al. 2007).
Mannitol
Mannitol is an important sugar alcohol which is present in many species of brown algae, especially in Laminaria and Ecklonia. The mannitol content is subject to wide seasonal fluctuations and varies with environment. Mannitol is the sugar alcohol corresponding to mannose. It usually constitutes <10% of dry weight in both Ascophyllum nodosum and Laminaria hyperborea stipe. In autumn fronds of L. hyperborea, however, the content may be as high as 25% of dry weight (Table 6).
Applications of mannitol are extremely diverse. It is used in pharmaceuticals, in making chewing gum, in the paint and varnish industry, in leather and paper manufacture, in the plastics industry and in the production of explosives. The US, the UK, France and Japan are the main centres of production. Mannitol can be used in a variety of foods, candies and chocolate-flavoured compound coatings because it can replace sucrose to make sugar-free compound coatings. Sugar-free chocolates are especially popular for people with diabetes, a growing problem in modern society. It is used as a flavour enhancer because of its sweet and pleasantly cool taste. Mannitol can be used to maintain the proper moisture level in foods so as to increase shelf-life and stability because it is non-hygroscopic and chemically inert. Mannitol is the preferred excipient for chewable tablets due to its favourable feel in the mouth. It is non-carcinogenic and can be used in paediatric and geriatric food products as it will not contribute to tooth decay (Nabors 2004).
Laminarin
Laminarin is found in the fronds of Laminaria/Saccharina and, to a lesser extent, in Ascophyllum and Fucus species and Undaria. The content varies seasonally and with habitat and can reach up to 32% of dry weight (Table 6).
Commercial applications of the extract have so far been limited, although some progress has been made in France as an antiviral in agricultural applications (Goemar 2010) or as dietary fibre (Deville et al. 2004). Especially the use of laminarin as substratum for prebiotic bacteria seems to have a good commercial application (Deville et al. 2004). Laminarin does not gel or form any viscous solution, and its main potential appears to lie in medical and pharmaceutical uses. It has been shown to be a safe surgical dusting powder and may have value as a tumour-inhibiting agent and, in the form of a sulphate ester, as an anticoagulant (Miao et al. 1999). The presence of anticoagulant activity in brown algae was first reported in 1941, when Laminaria showed anticoagulant properties with its active compound being located in the holdfasts (Shanmugam and Mody 2000). There are about 60 brown algal species identified to have blood anticoagulant properties. Laminarin only shows anticoagulant activity after structural modifications such as sulphation, reduction or oxidation. The anticoagulant activity is improved chemically by increasing the degree of sulphation (Shanmugam and Mody 2000).
Preparations containing 1→3:1→6-β-d-glucans, laminarin and fucoidan are manufactured by the health industry and marketed for their beneficial properties on the immune system. The producers of these tablets cite numerous papers discussing the biological activity of these glucans.
Laminarin provides protection against infection by bacterial pathogens and protection against severe irradiation, boosts the immune system by increasing the B cells and helper T cells, reduces cholesterol levels in serum, and lowers systolic blood pressure, among other effects (Table 6; Hoffmane et al. 1995) lower the levels of total cholesterol, free cholesterol, triglyceride and phospholipid in the liver (Miao et al. 1999; Renn et al. 1994a, b). The hypocholesterolemic and hypolipidemic responses are noted to be due to reduced cholesterol absorption in the gut (Kiriyama et al. 1969; Lamela et al. 1989; Panlasigui et al. 2003). This is often coupled with an increase in the faecal cholesterol content and a hypoglycaemic response (Dumelod et al. 1999; Ito and Tsuchida 1972; Nishide et al. 1993). A high level of LDL-cholesterol can lead to plaque forming and clog arteries and lead to cardiovascular diseases and heart attacks or strokes, a major cause of disease in the US. Laminarin as a potential cancer therapeutic is under intensive investigation (Miao et al. 1999).
Ulvan, porphyran and xylans
The name ulvan is derived from the original terms ulvin and ulvacin introduced by Kylin in reference to different fractions of Ulva lactuca water-soluble sulphated polysaccharides. It is now being used to refer to polysaccharides from members of the Ulvales, mainly Ulva and Enteromorpha spp. Ulvans are highly charged sulphated polyelectrolytes composed mainly of rhamnose, uronic acid and xylose as the main monomer sugars and containing a common constituting disaccharide: the aldobiuronic acid, [→4)-d-glucuronic acid-(1→4)-l-rhamnose3-sulphate-(1→]. Iduronic acid is also a constituent sugar. The average molecular weight of ulvans ranges from 189 to 8,200 kDa (Lahaye 1998). The cell wall polysaccharides of ulvales represent 38–54% of dry algal matter (Lahaye 1998). Two major kinds have been identified: water-soluble ulvan and insoluble cellulose-like material.
The mechanism of gel formation is unique among polysaccharide hydrogels. It is very complex and not yet fully understood. The viscosity of ulvan solutions as isolated polysaccharides has been compared to that of arabic gum. Whether ulvans present other functional properties of this gum remains to be established. The gelling properties of ulvans are affected by boric acid, divalent cations and pH. They are thermoreversible without thermal hysteresis. The gelling properties can be of interest for applications where gel formation needs to be precisely controlled (by pH or temperature), like the release of trapped molecules or particles under specific conditions (Percival and McDowell 1990; Lahaye et al. 1998). As already mentioned, highly absorbent, biodegradable hydrocolloid wound dressings limit wound secretions and minimise bacterial contamination. Polysaccharide fibres trapped in a wound are readily biodegraded. In the context of bovine spongiform encephalopathy (mad cow disease) and other prion contamination diseases, macromolecular materials from algal biomasses such as ulvans can constitute an effective and low-cost alternative to meat-derived products because their rheological and gelling properties make them suitable substitutes for gelatin and related compounds (Choi et al. 1999).
Ulvans are a source of sugars for the synthesis of fine chemicals. In particular, they are a potential source of iduronic acid, the only occurrence to date of this rare uronic acid in plants (Lahaye and Ray 1996). Iduronic acid is used in the synthesis of heparin fragment analogues with anti-thrombotic activities, and obtaining it requires a lengthy synthetic procedure that could be more cost-effectively replaced by a natural source (Lahaye 1998). Oligosaccharides from Ulva could be used as reference compounds for analysing biologically active domains of glycosaminoglycan-like heparin. The use of oligo-ulvans as anticoagulant agents could be expected since other, rarer, sulphated polysaccharides, like dermatan sulphate or fucan in brown algae, have shown this anti-thrombinic activity. Regular oligomers from ulvans may provide better-tolerated anti-thrombinic drugs (Paradossi et al. 2002).
Rhamnose, a major component of ulvans, is a rare sugar used as a precursor for the synthesis of aroma compounds. Combinatorial libraries in glycopeptide mimetics are another example of the use of l-rhamnose in the pharmaceutical industry. The production of rhamnose from Monostroma, a Japanese species of Codiales, has been patented. Rare sulphated sugars such as rhamnose 3-sulphate and xylose 2-sulphate are also of interest (Lahaye and Robic 2007).
Other potential applications of ulvan oligomers and polymers are related to their biological properties. Recent studies have demonstrated that ulvans and their oligosaccharides were able to modify the adhesion and proliferation of normal and tumoural human colonic cells as well as the expression of transforming growth factors and surface glycosyl markers related to cellular differentiation (Lahaye and Robic 2007). Earlier work demonstrated strain-specific anti-influenza activities of ulvan from U. lactuca, and the use of rhamnan, rhamnose and oligomers from desulphated Monostroma ulvans has been patented for the treatment of gastric ulcers (Fujiwara-Arasaki et al. 1984; Nagaoka et al. 2003).
Oligomers from seaweed species such as Laminaria spp. or Fucus serratus have also been studied as plant elicitors. These are natural compounds which stimulate the natural defences of plants. Several products derived from brown algae are already marketed worldwide. The success is because of their size and availability rather than their chemical composition. Ulva cell walls bind heavy metals and ulvans are the main contributors with 2.8–3.77 meq g−1 polysaccharide. The ion exchange property of ulvans explains why they have been chosen as bioindicators for monitoring heavy metal pollution in coastal waters (Nagaoka et al. 1999).
In red algae, the fibrillar network is made of low crystalline cellulose, mannan or xylan and represents only about 10% of cell wall weight. It can also contain minor amounts of sulphated glucans, mannoglycans and complex galactans. Most of our current knowledge of red algal cell wall polysaccharides is on the gelling and thickening water-soluble galactans, agars and carrageenans used in various applications. Unlike most red seaweed generally studied, P. palmata does not produce matrix galactans, but instead (10/4)- and (10/3)-linked-b-d-xylan together with a minor amount of fibrillar cellulose and b-(10/4)-d-xylan. Xylans can be 35% of the dry weight of Palmaria (Lahaye et al. 2003). Xylans have not yet been of economic interest, and only few applications are known.
Species of Porphyra contain a sulphated polysaccharide called porphyran, a complex galactan. Porphyran is a dietary fibre of good quality and chemically resembles agar. A powder consisting of 20% nori mixed with a basic diet given orally to rats prevented 1,2-dimethylhydrazine-induced intestinal carcinogenesis. Porphyran showed appreciable anti-tumour activity against Meth-A fibrosarcoma. In addition, it can significantly lower the artificially enhanced level of hypertension and blood cholesterol in rats (Noda 1993).
Digestibility of polysaccharides
The majority of edible seaweed fibres are soluble anionic polysaccharides which are little-degraded or not fermented by the human colonic microflora (Lahaye and Kaeffer 1997). The amount of dietary fibres in marine algae not digested by the human digestive tract is higher than that of other food materials (Murata and Nakazoe 2001). Most of the total algal fibres disappeared after 24 h (range 60–76%) in in vitro fermentation of, e.g. Laminaria digitata and U. pinnatifida using human faecal flora. However, unlike the reference substrate (sugar beet fibres), the algal fibres were not completely metabolised to short-chain fatty acids (SCFA; range 47–62%). Among the purified algal fibres, disappearance of laminarins was approximately 90% and metabolism to SCFA was approximately 85%, in close agreement with the fermentation pattern of reference fibres. Sulphated fucans were not degraded. Sodium alginates (Na alginates) exhibited a fermentation pattern quite similar to that of the whole algal fibres, with a more pronounced discrepancy between disappearance and production of SCFA: disappearance was approximately 83%, but metabolism was only approximately 57%. Laminarin seemed to be a modulator of the intestinal metabolism by its effects on mucus composition, intestinal pH and short-chain fatty acid production, especially butyrate. The characteristic fermentation pattern of the total fibres from the brown algae investigated was attributed to the peculiar fermentation of alginates (Michel et al. 1996; Deville et al. 2007).
Phycolloids are more or less degraded following adaptation of the human microflora, but none of the seaweed polysaccharides have been shown to be metabolised, although some may be partly absorbed. Nothing is known about the fate of other algal polysaccharides in the human digestive tract, except that they cannot be digested by human endogenous enzymes. Results of fermentation in vitro with human faecal bacteria indicate that brown seaweed fibres exhibit an original fermentation pathway (Mabeau and Fleurence 1993). Carrageenan is a good source of soluble fibre (Burtin 2003). Rats excrete carrageenan quantitatively in the faeces if it is administered in the diet at levels of 2–20%, and it therefore has no direct nutritive value (Hawkins and Yaphe 1965). Weight gain was significantly reduced, especially at higher levels. Furthermore, food efficiency showed interference with utilisation of other nutrients in the diet. Only 10–15% appeared digestible from faecal examination (Hawkins and Yaphe 1965). An experimental feeding with L. digitata seaweed extract in pig resulted in a higher production of butyric acid in the caecum and colon compared to the control group (Reilly et al. 2008). Butyrate is a beneficial metabolite for intestinal bacteria because it is quickly metabolised by colonoyctes and accounts for about 70% of the total energy consumption of the colon (Reilly et al. 2008). Therefore, it is desirable to promote butyrate production in the colon by laminarin fermentation.
The particular chemical structure of ulvan (and of Ulva) is responsible for the resistance of this polysaccharide to colonic bacterial fermentation. Consumption of dietary fibres from Ulva sp. could be expected to act mainly as bullring agents with little effect on nutrient metabolism due to colonic bacterial fermentation products (short-chain fatty acids; Bobin-Dubigeon et al. 1997). All soluble fibre fractions of P. palmata consisted of linear beta-1,4/beta1,3 mixed linked xylans containing similar amounts of 1,4 linkages (70.5–80.2%). The insoluble fibres contained essentially 1,4-linked xylans with some 1,3-linked xylose and a small amount of 1,4-linked glucose (cellulose). Soluble fibres were fermented within 6 h by human faecal bacteria into short-chain fatty acids (Lahaye et al. 1993).
Proteins, peptides and amino acids
Although the structure and biological properties of seaweed proteins are still poorly documented, the amino acid compositions of several species have been known for a long time (Murata and Nakazoe 2001).
The non-protein nitrogen is thought to be an intermediate or final product of nitrogen metabolism in plant tissue, and this fraction consists of amino acids, peptides, amines, and nucleotides and accounts for 10–20% of the nitrogen in seaweed (Arasaki and Arasaki 1983). Investigations found the biochemical composition of total nitrogen to be 57% in F. vesiculosus, of which 7–8% were the total nitrogen peptides and 10% free amino acids (Smith and Young 1953).
Habitat and especially seasonal variation have an effect on proteins, peptides and amino acids in seaweed (Arasaki and Arasaki 1983; Haug and Jensen 1954).
Proteins
The protein fraction of seaweed varies with the species, but is generally small in brown seaweed. Undaria has the maximal content of 24% dry weight, followed by several species (Sargassum, Fucus and Laminaria/Saccharina) with maxima around 17–21% and with Ascophyllum with the lowest content (maximum 10% of dry weight; Table 7). Higher protein contents are recorded for green and red seaweed. Proteins can represent up to 35%, 44% and 44% of dry matter in P. palmata (dulse), Porphyra tenera (nori) and Ulva spp., respectively. These levels are comparable to those found in high-protein vegetables such as soybeans, in which proteins make up 40% of the dry mass (Murata and Nakazoe 2001). The protein content in the fronds of Saccharina and Laminaria species and A. esculenta showed a pronounced maximum during the period from February to May, the young parts of the Saccharina and Laminaria species fronds being considerably richer than the old parts (Haug and Jensen 1954; Jensen and Haug 1956).
Most seaweed species contain all the essential amino acids and are a rich source of the acidic amino acids, aspartic acid and glutamic acid (Fleurence 2004). Whilst threonine, lysine, tryptophan, sulphur amino acids (cysteine and methionine) and histidine have been perceived as limiting amino acids in algal proteins, their general levels in algal proteins are higher than those found in terrestrial plants (Galland-Irmouli et al. 1999). The proteins from P. tenera exhibit an amino acid composition close to that of ovalbumin (Mabeau and Fleurence 1993). The combined glutamic acid and aspartic acid level accounts for 22–44% wet weight of the total amino acid fraction in Fucus sp. and 39–41% in Sargassum sp., whilst both amino acids together represent 18% wet weight of the total amino acid content of L. digitata proteins. An analogous result was reported for A. nodosum (Augier and Santimone 1978; Fleurence 2004; Munda 1977). Brown seaweed proteins have also been described as being a rich source of threonine, valine, leucine, lysine, glycine and alanine, with amino acids such as cysteine, methionine, histidine, tryptophan and tyrosine recorded at lower levels (Augier and Santimone 1978; Dawczynski et al. 2007; Fujiwara-Arasaki et al. 1984).
Red seaweed proteins in general, as with other seaweed groups, possess high quantities of the acidic amino acids, glutamic acid and aspartic acid, whilst basic amino acids are less dominant. However, some reports have shown that red algae in general appear to contain lower quantities of glutamic acid and aspartic acid compared to the other two algal groups (Fleurence 2004). Quantities range from 7.9% of total amino acids in C. crispus, 14% in P. tenera and up to 28% in P. palmata (Fujiwara-Arasaki et al. 1984; Galland-Irmouli et al. 1999; Young and Smith 1958). Ulva armoricana contains high levels of the amino acid proline (approximately 5–11%) of total amino acids, whilst U. pertusa is reported to have an arginine content of 15% (Fujiwara-Arasaki et al. 1984; Fleurence et al. 1995).
Bioactive lectins are found in macroalgal species such as Ulva sp., Eucheuma spp. and Gracilaria sp. (Table 8). Lectins are a structurally diverse group of carbohydrate-binding proteins of non-immune origin found in a wide range of organisms (Hori et al. 2000). Lectins purified from the green marine algae Codium fragile subsp. tomentosoides have been produced and sold commercially (Smit 2004). Lectins interact with specific glycan structures linked to soluble and membrane-bound glycoconjugates. It is these protein–carbohydrate interactions that are responsible for lectin involvement in numerous biological processes such as host–pathogen interactions, cell–cell communication, induction of apoptosis, cancer metastasis and differentiation, as well as recognising and binding carbohydrates (Hori et al. 2000). Furthermore, lectin has been found to increase the agglutination of blood cells (erythrocytes) and is also useful in the detection of disease-related alterations of glycan synthesis, including infectious agents such as viruses, bacteria, fungi and parasites (Bird et al. 1993; Cardozo et al. 2007; Murata and Nakazoe 2001). Other bioactive properties exhibited by marine algal lectins include antibiotic, mitogenic, cytotoxic, anti-nociceptive, anti-inflammatory, anti-adhesion and anti-HIV activities (Bird et al. 1993; Mori et al. 2005; Smit 2004). Lectins from three Eucheuma species, E. serra, E. amakusaensis and E. cottonii have shown strong mitogenic activity on mouse and human lymphocytes, whilst a marine glycoprotein E. serra agglutinin (ESA) derived from E. serra exhibits a cytotoxic effect against several cancer cell lines, such as colon cancer Colo201 cells and cervical cancer HeLa (Kawakubo et al. 1997, 1999; Sugahara et al. 2001). It was also reported that ESA-2, a lectin isolated and characterised from E. serra, suppressed colonic carcinogenesis in mice when orally administered and exhibited growth inhibition of 35 human cancer cell lines (Hori et al. 2007). This phycolectin was found to inhibit adenosine diphosphate and collagen-induced human platelet aggregation in a dose-dependent manner.
Antibacterial lectins have also been isolated from E. serra and Galaxaura marginata (Liao et al. 2003). These carbohydrate-binding proteins exhibit antibacterial activity against the fish pathogen Vibrio vulnificus. Aggregation and immobilisation of V. vulnificus cells were observed following incubation of the cells with glycoprotein molecules because lectins recognise and bind to complementary carbohydrates or glycoproteins on either the cell walls or plasma membranes of the associated bacteria cells. As a result, the microorganisms are immobilised and further growth and multiplication is prevented (Liao et al. 2003).
Mucin-binding agglutinin and isoagglutinin glycoproteins isolated from two different species of red algae possessed anti-inflammatory activity and showed mitogenic activity on mouse spleen lymphocytes, and they inhibited the growth of mouse leukaemia cells L1210 and mouse FM3A tumour cells (Bitencourt et al. 2008; Hori et al. 1988).
Phycobiliproteins are described later.
Peptides
The light-harvesting complex fucoxanthin, from brown algae, has been found to contain four 20-kDa polypeptides. The gene coding sequences for these polypeptides (lhcf genes) show homology to those found in Macrocystis pyrifera and L. digitata (two Laminariales) and indicate that these proteins are probably similar in all brown algae (Douady et al. 1994; Martino et al. 2004). Eisenine is isolated from Eisenia bicyclis; octaglutamic acid and l-pyrrolidonoyl-l-glutamyl-l-glutamine are isolated from Pelvetia canaliculata; carnosine in Acanthophora delilei and many peptides, including glutamic and aspartic acid, are found in L. japonica (Arasaki and Arasaki 1983). The two antioxidant peptides, carnosine and glutathione, that are generally present in high concentrations in animal muscle have also been found in marine macroalgae (Arasaki and Arasaki 1983; Shiu and Lee 2005).
The depsipeptide kahalalide F from Bryopsis sp. is active in the treatment of lung cancer, tumours and AIDS (Smit 2004). In addition, ten dipeptides from hot water extracts of U. pinnatifida reduced blood pressure in spontaneously hypertensive rats and showed angiotensin-converting enzyme (ACE) inhibitory activity (Sato et al. 2002; Suetsuna et al. 2004). Peptides with ACE inhibitory activity have been isolated from Porphyra yezoensis and Hizikia fusiformis (Suetsuna 1998a, b). Aqueous extracts of ten green algae and 19 brown algae extracted at 70°C also exhibited ACE inhibitory activity, with the highest activity associated with Ecklonia cava (Cha et al. 2006). In addition, plasma and hepatic cholesterol-reducing actions were observed when feed containing hydrolysates was administered to rats. This feed was also shown to improve rat hepatic function and reduce plasma glucose levels. Furthermore, fractions of the pepsin hydrolysates were reported to have superoxide dismutase (SOD)-like and antioxidant activity in a linoleic acid system.
A mitogenic hexapeptide, SECMA 1, has been isolated from Ulva sp. This peptide was shown to be involved in the modulation of cell proliferation-associated molecules, proteoglycans and glycosaminoglycans in human foreskin fibroblasts (Ennamany et al. 1998). In addition, a rare cyclic pentapeptide, Galaxamide, obtained from the marine alga Galaxaura filamentous has been shown to exhibit anti-proliferative activity against human epithelial cancer cell lines (Xu et al. 2008).
A number of oligopeptides that act like analogues of the neurotransmitter γ-aminobutyric acid (GABA) have been discovered in red algae. These analogue compounds bind to GABA sub(A) receptors, and some are even more active than GABA itself. The proposed applications for these oligopeptides are in both the diagnosis and therapy of certain brain and central nervous system disorders caused by the malfunction of systems controlled by GABA (Aneiros and Garateix 2004).
Free amino acids
The free amino acid fraction of seaweed is mainly composed of alanine, amino butyric acid, taurine, omithine, citruiline and hydroxyproline. Levels differ according to the species. The characteristic taste of nori is caused by the large amounts of the three amino acids: alanine, glutamic acid and glycine. In P. tenera, alanine amounting to about 25% of the total free amino acids probably accounts for the faintly sweet taste (McHugh 2003).
Ulva contains large amounts of cysteinolic acid, cysteic acid, praline, glutamic acid and chondrine, and Undaria contains abundant alanine, glycine and praline, whereas the arginine content is high in Gracilaria. In addition, such amino acids as citrulline, praline, taurine, arginine and ornithine accumulate in cells (Arasaki and Arasaki 1983). Certain species have a high level of arginine (U. pertusa, U. pinnatifida and P. tenera) or glycine (P. tenera). The major amino acids in C. crispus are arginine (34%) and amide N (10% of total N). Furthermore, citrulline and ornithine (6% and 7% of total N, respectively) have been established as occurring both in the free state and in simple and complex peptides in C. crispus, but not in the insoluble protein (Table 9; Young and Smith 1958).
The eight essential amino acids (cystine, isoleucine, leucine, lysine, methionine, phenylalanine, tyrosine and valine) cannot be synthesised by animals, nor can they be replaced by other “less valuable” building blocks. Generally, the first limiting amino acid in most of the proteins of marine algae is lysine, one of the essential amino acids (Murata and Nakazoe 2001), and the various edible seaweed exhibit similar essential amino acid patterns (Mabeau and Fleurence 1993). All essential amino acids were detected in some brown and red seaweed species, and red species featured uniquely high concentrations of taurine when compared to brown seaweed varieties (Dawczynski et al. 2007). Taurine is mostly found in fish, shellfish and seaweed. The taurine content is at a level of 400 mg g−1 dry weight in S. latissima and P. tenera, which is also the case in lobster, crab, shellfish and squid (Murata and Nakazoe 2001). Taurine is not a true amino acid due to the lack of a carboxyl group, but contains a sulfonated acid group instead. This amino acid is not used as building blocks in proteins but is found in, e.g. the gall bladder, where it works as an emulsifier which binds and eases the uptake of lipids. Taurine is important for the formation of bile by which cholesterols are bound. Taurine thereby facilitates reduction of cholesterol content in the blood of humans (Mouritsen 2009). In addition to its requirement for a number of biological and physiological functions, taurine has been identified as having a number of health-promoting properties, e.g. anti-hypertensive, hypocholesterolemic and antioxidant properties, to name but a few (Lourenço and Camilo 2002). Taurine leads to decreases in the concentration of cholesterol in both the serum and liver of rats, prevention effect of vascular diseases, chronic hepatitis and diabetes (Mochizuki et al. 1999).
In addition to taurine, other bioactive amino acids such as laminine, kainoids and mycosporine-like amino acids have been found in marine macroalgae. Laminine, a choline-like basic amino acid, has been isolated from Laminaria angustata and Chondria armata (Bhakuni and Rawat 2005). This amino acid was reported to depress the contraction of excited smooth muscles, whilst laminine monocitrate was shown to exert a transitory hypotensive effect (Bhakuni and Rawat 2005). The kainoid amino acids, kainic and domoic acids have also been found in numerous algal species, but extracted from Digenea simplex and commercialised (Smit 2004). Several preparations of kainic acids are available, and these include ‘Digenin’ and ‘Helminal’ (Bhakuni and Rawat 2005). Kainoids are a unique group of unusual amino acids which are related structurally to, and have similar functions to, the acidic amino acids aspartic acid and glutamic acid. They have attracted considerable interest due to their high insecticidal, anthelmintic and neuroexcitatory properties (Parsons 1996). Moreover, D. simplex and C. armata extracts containing domoic and kainic acid have been used as anthelmintic agents for centuries in Japan for the treatment of ascariasis (a disease in humans caused by the parasitic roundworm; Bhakuni and Rawat 2005; Parsons 1996; Smit 2004). These compounds are currently being used in research associated with neurophysiological disorders such as Alzheimer’s and Parkinson’s disease and epilepsy (Smit 2004).
Nutritional value and digestibility of seaweed proteins
The nutritional value of proteins referred to as “amino acid score” is evaluated based on the composition of essential amino acids. The amino acid score of the proteins of the marine algae ranges from 60 to 100, a value higher than that of the proteins in cereal and vegetables. The amino acid score of proteins in Porphyra and Undaria was 91 and 100, respectively, the same as that of animal foods (Murata and Nakazoe 2001).
The in vivo digestibility of seaweed proteins is not well documented, and available studies about their assimilation by humans have not provided conclusive results. However, several researchers have described a high rate of seaweed protein degradation in vitro by proteolytic enzymes such as pepsin, pancreatin and pronase, with pronase as the most digestive enzyme. For instance, the digestibility in vitro of proteins from P. tenera, U. pinnatifida and U. pertusa is 78%, 87% and 95%, respectively (Fleurence 1999). Digestibility of U. lactuca protein concentrates was 86%, determined by a multi-enzyme method using different trypsins and peptidases (Wong and Cheung 2001c). However, for brown seaweed species, the high phenolic content might limit protein availability in vivo and thus moderate in vitro figures. This situation is probably not found for the green and red seaweed which possess low levels of phenols and higher protein content (Mabeau and Fleurence 1993).
Lipids
Lipids are a broad group of naturally occurring molecules which includes fats, waxes, sterols, fat-soluble vitamins (such as vitamins A, D, E and K), mono-, di- and triacylglycerols, diglycerides, phospholipids, and others. Lipids account for about 40% of the total calories in most industrialised countries; they reduce the bulk of the diet because of the concentrated source of energy and are indispensable from the human diet as they form the major sources of essential fatty acids and contain fat-soluble substances like vitamins and carotenoids. Vegetable oils contribute 70% of the dietary lipids, whilst marine oils contribute about 2% of the global oil production, with the remainder coming from fats from land animals. The per capita consumption of marine oils is approximately 1 g day−1. Vegetable oils usually consist of saturated and unsaturated fatty acids with 16- and 18-carbon chain lengths (C16 and C18), whilst marine oils mainly consist of C14–C22 fatty acids (Narayan et al. 2006).
Murata and Nakazoe (2001) state that phospholipids are the main source of lipids in marine algae, contrary to Le Tutour (1990), Bhaskar et al. (2004) and Khotimchenko (2005) who claim that the glycolipids are the major lipid class in all seaweed, followed by neutral and phospholipids. Besides fatty acids, the unsaponifiable fraction of seaweed contains carotenoids, such as β-carotene, lutein and violaxanthin in red and green algae and fucoxanthin in brown algae, as well as tocopherols and sterols (Table 10; see also later chapters; Jensen 1969b). Lipids represent up to 4.5% of the seaweed on a dry weight basis, and this content is lower than that of other marine organisms. Their contribution as a food energy source thus appears to be low (Table 11; Mabeau and Fleurence 1993; Murata and Nakazoe 2001). Like other biochemical components, the fatty acid content varies with the season and other environmental factors. It is generally known that algae can accumulate polyunsaturated fatty acids (PUFAs) when there is a decrease in the environmental temperature (Khotimchenko 1991). Aquatic species that live in colder waters generally contain larger quantities of PUFAs (Narayan et al. 2006), and the maximum content of lipids in the fronds of Saccharina, Laminaria species and A. esculenta was generally found in winter (Haug and Jensen 1954). However, the total lipids of Fucus sp. were most abundant in summer, with highest levels recorded in August. A attached specimen of Gracilaria accumulated twice as much lipid compared to the unattached form, but with no difference in fatty acid composition (Khotimchenko and Levchenko 1997; Kim et al. 1996).
Induction of omega-3 fatty acid production has been reported in seaweed. Cell suspension cultures in photobioreactors of S. latissima produced three sulphate fatty acids deriving from an n-6 lipoxygenase oxidation (15-hydroxy-5,8,11,13-eicosatetraenoic acid (15-HETE), 13-hydroxy-6,9,11,15-octadecatetraenoic acid (13-HODTA), and 13-hydroxy-9,11-octadecadienoic acid (13-HODE). The yields of 15-HETE, 13-HODTA and 13-HODE ranged from 100 to 1,000 μg product g−1 dry biomass, but addition of linoleic and γ-linolenic acid in the medium increased the yield of all three sulphate fatty acids by up to 400% (Rorrer et al. 1997).
The lack of studies of the bioavailability of algal lipids currently limits their nutritional evaluation (Mabeau and Fleurence 1993).
Fatty acids
Marine-based long-chain PUFAs (LC-PUFA) have 20 or more carbons with two or more double bonds from the methyl (omega) terminus. The omega-3 LC-PUFAs (n-3 LC-PUFA) have the first double bond located in the third carbon from the methyl terminus and can contain up to six double bonds. They are of particular interest in respect to functional food (Rasmussen and Morrissey 2007). PUFAs are occasionally classified into two families because of their metabolic connections. One is the linoleic acid family (n-6 fatty acid) and the other is the α-linolenic acid family (n-3 fatty acid; Table 11). Marine lipids contain substantial amounts of LC PUFAs, with n-3 fatty acids as the significant component and mono-unsaturated fatty acids. Eicosapentaenoic acid (EPA; 20:5 n-3) and docosahexaenoic acid (DHA; 22:6 n-3) are the two important fatty acids of marine lipids, along with the precursor α-linolenic acid (ALA; 18:3 n-3) and docosapentaenoic acid (22:5 n-3). Both EPA and DHA are basically derived from ALA through elongation and desaturation (Narayan et al. 2006). These LC-PUFAs EPA and DHA have been shown to cause significant biochemical and physiological changes in the body. The main effects of n-3 fatty acids on human health can be divided into three main categories: (1) their essentiality as structural components of cell and organelle membranes; (2) their significant role in lowering blood lipids; and (3) their role as precursors for mediating biochemical and physiological responses. Humans have to directly intake the ALA, EPA and/or DHA to obtain physiological effects of EPA and DHA. The major sources of EPA and DHA in human diets are marine products. Terrestrial plants can produce small to moderate amounts of ALA; however, the mammalian system cannot synthesise ALA. Major sources of EPA and DHA are unicellular phytoplankton and seaweed. EPA and DHA are accumulated in fish and other marine animals that consume algae and get passed on to other species through the food chain. It was found that individuals consuming a normal diet had low tissue levels of EPA and DHA, whilst those who consumed fish had higher levels in their tissue (Narayan et al. 2006). Generally, the marine n-3 polyunsaturated fatty acids exert anti-arteriosclerosis, anti-hypertension, anti-inflammation, immunoregulation effects, etc. (Khan et al. 2007; Maeda et al. 2005; Plaza et al. 2008).
The predominant fatty acid in various seaweed products is EPA (C20:5, n-3), which is at concentrations as high as 50% of the total fatty acid content (Dawczynski et al. 2007; Murata and Nakazoe 2001). Marine algae also contain the n-3 PUFA 18:4 n-3, which is not included in other organisms. Notably, red seaweed species contain significant quantities of polyunsaturated fatty acids such as EPA and arachidonic acid (20:4). The green algal species are unusual in containing 16:4 varying from 4.9% to 23.1% of the total fatty acids; 16:0, 18:1 and 18:3 acids have also been found in high amounts. Unsaturated fatty acids predominate in all the brown seaweed studied and saturated fatty acids in the red seaweed, but both groups are balanced sources of n-3 and n-6 acids. Even though seaweed is processed (canned and dried), the products are left with substantial nutritional value of protein, ash, and n-3 and n-6 fatty acid contents (Mabeau and Fleurence 1993; Sanchez-Machado et al. 2004a). Algae are therefore a good source of EPA n-3 (Plaza et al. 2008) and an important source of supply of n-3 PUFAs for the maintenance of health (Murata and Nakazoe 2001). It is suggested that algal 18:4 n-3 affects the immune system in humans (Ishihara et al. 1998). In accordance with Table 11, Palmaria and Sargassum have the best n-6/n-3 ratio (0.1 and 0.3, respectively) and also some of the highest total lipid contents of 3.8% and 3.9% respectively. Gracilaria, Ulva and Fucus have the highest contents of oleic acid (31%, 27% and 25%, respectively).
Phospholipids
Phospholipids (PLs) consist of fatty acids and a phosphate-containing compound attached to glycerol or the amino alcohol sphingosine, resulting in compounds with fat-soluble and water-soluble regions. Cell membranes utilise the dual hydrophilic and hydrophobic characteristic of PLs to maintain structure and transport materials. Glycerol-containing PLs include phosphatidic acid, phosphatidylcholine (PC), phophatidylethanolamine (PE), phophatidylinositol and phosphatidylserine. Sphingomyelin (SPH), the other major PL, consists of sphingosine and PC. Phospholipids contribute to lipoprotein formation in the liver, nervous system conduction and protection, memory storage and muscle control. Choline, a major component of PC and SPH, functions as a methyl donor and a precursor to the neurotransmitter acetylcholine (Miraglio 2009).
The amount of phospholipids in various red seaweed species varies from 10% to 21% of the total lipid (0.5–2.6 mg g−1 dry weight). The major phospholipids appear to be PC (62–78%) and PG (10–23%). The presence of unidentified polar phospholipids vary from 2.7% to 10%, and PE and DPG are found as minor phospholipids (Dembitsky and Rozentsvet 1990). The major phospholipids in F. vesiculosus and A. nodosum are phosphatidylethanolamine, but these represent <10% of the total acyl lipids (Jones and Harwood 1992).
Phospholipids in diet act as natural emulsifiers and as such facilitate and ease the digestion and absorption of fatty acids, cholesterol and other lipophilic nutrients. A producer of marine phopholipids explains that these phospholipids have many advantages compared with fish oils because they are much more resistant to oxidation (rancidity), have much higher contents of the physiologically important EPA and DHA, provide these fatty acids with much better bioavailability and have much better spectrum of health benefits for humans and animals (Sørensen 2009).
Glycolipids
Glycolipids are carbohydrate-attached lipids. Their role is to provide energy and also serve as markers for cellular recognition because of their association with cell membranes.
The red algae contained almost equal quantities of monoglycosyldiacylglycerol (MGDG), diglycosyldiacylglycerol (DGDG) and sulphaquinovosyldiacyl-glycerol, with some exceptions. MGDG and DGDG were the major glycolipids in green seaweed. Glycolipid contents of total lipid extracts of red seaweed range from 16 to 32 μmol g−1 dry weight, whilst those in green algae vary from 11 to 32 μmol g−1 dry weight. Examination of the composition of glycolipids in some brown algae showed that MGDG content varied from 26% to 47%, DGDG content from 20% to 44% and sulphaquinovosylglycerol content from 18% to 52% of total glycolipids (Dembitsky et al. 1990, 1991). Glycolipid content in two Fucus species made up 14% of the ester-soluble lipids (Dembitsky et al. 1990)
The major acyl lipids in F. vesiculosus and A. nodosum are monogalactosyldiacylglycerol, digalactosyldiacylglycerol, sulphoquinovosyldiacylglycerol and trimethyl-beta-alaninediacylglycerol (Jones and Harwood 1992). The highest concentration of saturated and mono-unsaturated fatty acids was found in sulfoquinovosyldiacylglycerides for species of Palmaria, Undaria and Ulva (Khotimchenko 2003).
Sterols
Sterols occur naturally in plants, animals and fungi, with the most familiar type of animal sterol being cholesterol. Cholesterol is vital to cellular function where it affects the fluidity of the animal cell membrane and serves as a secondary messenger in developmental signalling. Furthermore, cholesterol is a precursor to fat-soluble vitamins and steroid hormones. Content and type of sterols vary with the seaweed species. Green seaweed species contain 28-isofucocholesterol, cholesterol, 24-methylene-cholesterol and β-sitosterol, whilst brown seaweed contains fucosterol, cholesterol and brassicasterol. Red seaweed contains desmosterol, cholesterol, sitosterol, fucosterol and chalinasterol (Sanchez-Machado et al. 2004b; Whittaker et al. 2000). The predominant sterol, fucosterol, in the brown seaweed (Laminaria and Undaria) makes up 83–97% of total sterol content (662–2,320 μg g−1 dry weight) and desmosterol in red seaweed (Palmaria and Porphyra), 87–93% of total sterol content (87–337 μg g−1 dry weight). However, the red seaweed C. crispus has cholesterol as its major sterol (Kumar et al. 2008b; Sanchez-Machado et al. 2004b).
It is reported that plant sterols such as β-sitosterol and fucosterol lead to the decrease of the concentration of cholesterol in the serum in experimental animals and humans (Whittaker et al. 2000).
Pigments
Chlorophylls
Chlorophylls are green lipid-soluble pigments found in all algae, higher plants or cyanobacteria that carry out photosynthesis. Chlorophyll a is essential in the reaction centre of the thylakoid, light-harvesting structures in which photosynthesis is carried out (Lobban and Harrison 1994; Rasmussen and Morrissey 2007). Chlorophyll is known to be converted into pheophytin, pyropheophytin and pheophorbide in processed vegetable food and following ingestion by humans. These derivates show anti-mutagenic effect and may play a significant role in cancer prevention. The cellular uptake and inhibition of myeloma cell multiplicity were found to be greater for pheophorbide than for pheophytin. Calculated on the amount of cell-associated chlorophyll derivative, however, pheophytin was more cytostatic/cytotoxic than pheophorbide (Chernomorsky et al. 1999).
The pigment content (violaxanthin, fucoxanthin, carotene and chlorophyll a) in Ascophyllum was high in March to June (peak in May) and with minimum levels in July to November. This seasonal variation was not so distinct in F. serratus, although it showed somewhat lower levels during autumn (Jensen 1966). Preliminary studies on chlorophyll showed that the content was three times higher in seaweed growing in harbour districts compared to seaweed from an open sea locality (Larsen and Haug 1958). The chlorophyll a content was 565–2,000 mg kg−1 on a dry weight basis in the brown species reported (Table 12).
Carotenoids
Carotenoids are nature’s most widespread pigments, and they are present in all algae, higher plants and many photosynthetic bacteria. They represent photosynthetic pigments in the red, orange or yellow wavelengths. Carotenoids are linear polyenes that function both as light energy harvesters and as antioxidants that inactivate reactive oxygen species formed by exposure to light and air (von Elbe and Schwartz 1996). Carotenoids are tetraterpenes, carotenes are hydrocarbons, and xanthophylls contain one or more oxygen molecules (Table 12; Lobban and Harrison 1994). Green seaweed species include β-carotene, lutein, violaxanthin, neoxanthin and zeaxanthin, whilst red seaweed species contain mainly α- and β-carotene, lutein and zeaxanthin. β-carotene, violaxanthin and fucoxanthin are present in brown seaweed species (Haugan and Liaaen-Jensen 1994).
Some carotenoids also function as vitamins that are classified by their biological and chemical activity, not their structure, and each “vitamin” may therefore refer to several vitamer compounds. Vitamers are often interconverted in the body. Vitamins have diverse biochemical functions, including function as hormones, antioxidants, and mediators of cell signalling and regulators of cell and tissue growth and differentiation.
The β-carotene (with provitamin A activity) content ranges from 36 to 4,500 mg kg−1 (or ppm) dry weight, with Porphyra with the highest content and P. palmata with the second highest of 456 ppm according to Table 12. The carotenes in Palmaria show seasonal variations, with the highest content in summer (420 ppm) and lowest in winter (37 ppm; Yuan 2008).
The antioxidant properties of the algal carotenoids have also been shown to play a role in preventing man pathologies linked to oxidative stress (Okuzumi et al. 1993, Yan et al. 1999). Furthermore, experimental studies strongly suggest that β-carotene could prevent the onset of cancers, especially lung cancer (Astorg 1997). However, other studies suggest that β-carotene could induce lung cancer in smokers. Recent studies have shown the correlation between a diet rich in carotenoids and a diminishing risk of cardiovascular disease, cancers (β-carotene, lycopene), as well as opthalmological diseases (lutein, zeaxanthin; Hosokawa et al. 1999).
Fucoxanthin is a xanthophyll and has a unique structure, including an unusual allenic bond and 5,6-monoepoxide in its molecule (Maeda et al. 2008b). Fucoxanthin is one of the most abundant carotenoids in nature. The content in seaweed varies during season and life cycle. It is quite stable in the presence of organic ingredients, apart from surviving the drying process and storage at ambient temperature. In pure form, fucoxanthin is vulnerable to oxidation (Haugan and Liaaen-Jensen 1994). The total carotenoid content of F. serratus was found to be aproximately 0.08% of the dried extracted cells, and fucoxanthin comprises about 70% of the total carotenoid (Chapman 1970; Haugan and Liaaen-Jensen 1989). In accordance with Table 12, the fucoxanthin content ranges from 172 to 720 mg kg−1 dry weight in the brown seaweed species, with maximal concentration in F. serratus.
Waste parts of cultivated L. japonica (kombu) are a good biosource for fucoxanthin extraction. Discards from processing (stipe, holdfast and blade material) can be used as a source for fucoxanthin extraction. These discards have similar content of fucoxanthin (178–196 mg kg−1 fresh weight). The recovery ratio of fucoxanthin has been shown to reach 82%, and a total of 1,490 g of fucoxanthin was obtained from 10 t of kombu waste. The fucoxanthin obtained was stable and reduced by only 2% at 4°C in 6 months of storage (Kanazawa et al. 2008). Fucoxanthin from fresh Undaria is mostly found as the geometrical isomer all-trans (∼88%). Trans forms are more stable; however, cis forms of fucoxanthin were found to exert a higher inhibitory effect compared to their trans counterparts on human leukaemia (HL-60) cells and colon cancer (Caco-2) cells. Uptake and incorporation of the trans form of fucoxanthin into cellular lipids were faster compared to cis counterparts (Nakazawa et al. 2009). Fucoxanthin is easily converted to fucoxanthinol in human intestinal cells and in mice (suggesting that the active form of fucoxanthin in a biological system would be fucoxanthinol; Sugawara et al. 2002).
Experiments have shown that fucoxanthin from brown seaweed Undaria significantly reduces the viability of human prostate cancer cells and significantly reduces the percentage of tumour-bearing mice and the average number of tumours per mouse when given in the drinking water (Table 12; Kotake-Nara et al. 2001; Okuzumi et al. 1993). Other studies demonstrated anticancer effects, including inhibition of the proliferation of human leukaemia cell line (HL-60), and induced their apoptosis (Hosokawa et al. 1999; Miyashita and Hosokawa 2008).
Maeda et al. (2005) demonstrated a reduction in white adipose tissue (storage tissue for fat) in rats and obese and diabetic model mice when fed with fucoxanthin extracted from brown seaweed. Furthermore, the diet containing fucoxanthin suppressed body weight gains in mice and induced the expression of the thermogenesis protein, uncoupling protein 1 (UCP1), in white adipose tissue.
Purified fucoxanthin inhibited the cytoplasmic lipid accumulation in 3T3-L1 that can be induced by, e.g. insulin, and in a dose-dependent manner significantly attenuated the expression of lipid metabolic genes in 3T3-L1 adipocytes. These results suggest that fucoxanthin is an effective natural food constituent to prevent obesity (Miyashita and Hosokawa 2008).
Tocopherol is another strong antioxidant and part of the carotenoid family, and the content can fluctuate strongly, e.g. in L. digitata. The concentration ranges from trace levels in winter to 2% of dry weight in summer. Porphyra also contains high contents of vitamin E (tocols), followed by Ulva, Ascophyllum and Fucus species (Table 13). Vitamin E includes α-, β- and δ-tocopherol (Kumar et al. 2008b). The species of tocols in some seaweed species are given in Table 13.
Phycobiliproteins
Unlike chlorophylls and carotenoids, phycobiliproteins are water-soluble and form particles (phycobilisomes) on the surface of the thylakoids rather than being embedded in the membranes. Phycobiliproteins consist of pigmented phycobilins, which are linear tetrapyrroles. Different combinations of the two principal phycobilins—phycoerythrobilin (red) and phycocyanobilin (blue)—which absorb different regions of wavelengths give different absorption spectra (Lobban and Harrison 1994). Besides the phycoerythrin and phycocyanin, phycobiliproteins can also be divided into allophycocyanin and phycoerythrocyanin. Within phycobilisomes, phycobiliproteins play an important role in the photosynthetic process of the three types of algae—Rhodophyta, Cyanophyta and Cyptophyta (Aneiros and Garateix 2004; Chronakis et al. 2000). The additional photosynthetic pigments make light harvesting possible in deep waters as surface light wavelengths for some colours are almost completely absorbed below 10 m (Voet et al. 1999).
Phycobiliproteins, in particular phycoerythrin, can constitute a major proportion of the red algal cell proteins, with levels of 1.2% and 0.5% of dry weight reported for P. palmata and Gracilaria tikvahiae, respectively (Chronakis et al. 2000; Wang et al. 2002). Phycobiliproteins are currently being used as natural colourants for food and cosmetic applications and are used in foods such as chewing gum and dairy products in addition to lipsticks and eyeliners (Sekar and Chandramohan 2008). Niohan Siber Hegner Ltd. and Dainippon Ink & Chemical Inc. are two companies in Tokyo that market the phycobiliprotein phycocyanin as a natural food colourant (Houghton 1996). Different phycobiliproteins have been shown to exhibit antioxidant, anti-inflammatory, neuroprotective, hypocholesterolemic, hepatoprotective, antiviral, anti-tumour, liver-protecting, atherosclerosis treatment, serum lipid-reducing and lipase inhibition activities (Sekar and Chandramohan 2008).
Iodine
Marine algae are known to be high in mineral content and are used as feed and food supplements to supply minerals. As mentioned previously, marine algae have 10–100 times higher mineral content than traditional vegetables (Arasaki and Arasaki 1983; Nisizawa 2002). Ash and thereby the mineral content of the species of interest reaches levels of up to 55% on a dry weight basis (Table 1). Ash content in most land vegetables is usually much lower than in seaweed. For example, sweet corn has a content of 2.6%, whilst spinach has an exceptionally high mineral content of 20% (Rupérez 2002). Some of the minerals are necessary for our health, whilst some are toxic in varying degrees. The mineral composition varies according to phylum as well as various other factors such as seasonal, environmental, geographical and physiological variations. Nevertheless, this review will not focus on the contents of all minerals and vitamins despite their importance in human health. However, the important mineral iodine needs some attention in this review as the concentration of iodine can reach very high levels in certain brown algae. Generally, the iodine content reached up to 1.2% of dry weight in these species (Table 14).
Laminaria japonica is an excellent source of iodine, and it has been used in China for centuries as a dietary iodine supplement to prevent goitre (underactive thyroids, myxoedema) and has also been used for medical and food purposes for over 1,500 years in China, Japan and Korea. Most of the kombu is dried and eaten directly in soups, salads and tea or used to make secondary products with various seasonings (sugar, salt, soy sauce; Lobban and Harrison 1994). Furthermore, kelps were used as raw material for the extraction of iodine in Ireland during the seventeenth century (Morrissey et al. 2001).
The results of a study on the feasibility of naturally iodised salt indicated that the level of iodine in seaweed is constant, consumer-acceptable and of equal bioavailability (Mabeau and Fleurence 1993). Species of the genus Laminaria are the strongest iodine accumulators among all living systems, and the accumulation can be up to 30,000 times larger than the surrounding evironment (Table 14; Bartsch et al. 2008). The uptake of dietary iodide by the human and animal thyroid glands leading to thyroid hormone formation is a well-established phenomenon. L. digitata is widely used as a health supplement for myxoedema and for the treatment of goitre (Müssig 2009). An anti-tumourogenic role of U. pinnatifida has been reported (Funahashi et al. 1999; Maruyama et al. 2003; Takahashi et al. 2000). This Asian kelp species or its equivalent iodine content inhibited tumourogenesis in rats with carcinogen-induced mammary tumours, although the mechanism of action is not understood (Funahashi et al. 2001). It has also been suggested that the high dietary seaweed content may account for the relatively low prevalence of breast cancer in Japanese women (Cann 2006). Nevertheless, too much iodine in sensitive persons can trigger a hyperactive thyroid gland and may cause goitre, similar to the myxoedema reaction. Excessive seaweed consumption was found to be frequently associated with goitre, hypothyroidism and Hashimoto’s thyroiditis in countries where marine algae are traditionally used as food, whereas iodine-induced thyrotoxicosis following consumption of seaweed or seaweed-containing dietary supplements appears to be rare (Cloughley et al. 2008).
Phenols and other phorotannins
Phenols, sometimes called phenolics, are a class of chemical compounds consisting of a hydroxyl group (–OH) bonded directly to an aromatic hydrocarbon group. The simplest of the class is phenol, the parent compound used as a disinfectant and for chemical synthesis. Polyphenols from terrestrial plants are derived from gallic and ellagic acid, whereas the algal polyphenols are derived from polymerised phloroglucinol units (1,3,5-trihydroxybenzene). Phlorotannins are a group of phenolic compounds. Phlorotannins constitute an extremely heterogeneous group of molecules (structure and polymerisation degree heterogeneity) providing a wide range of potential biological activity (Table 15; Burtin 2003). Phlorotannins are localised in physodes which are membrane-bound cytoplasmic vesicles, and the fusion of physodes with cell membranes results in a secretion of phlorotannins (Bartsch et al. 2008; Li et al. 2009; Lüder and Clayton 2004). The size of the physodes is about 2,500 μm in F. serratus and L. hyperborea, and appearance does not differ much. The reducing power (up to 3.0 meq g−1 dry weight) increased with increasing physode volume (3–11% of the volume of the entire seaweed tissue of F. serratus, F. vesiculosus and A. nodosum; Baardseth 1958). Reducing power also correlated positively with the dry matter content (correlation coefficient of 0.73; Larsen and Haug 1958).
Green and red seaweed have low concentrations of phenols (Mabeau and Fleurence 1993) compared to brown seaweed species, which have high concentrations of the phenol group phlorotannin. Phenol content varies from <1% to 14% of dry seaweed biomass, with Ascophyllum and Fucus as the species with the highest content (14% and 12%, respectively; Table 15). Concentrations of polyphenols exhibit seasonal variations, but also vary within the different parts of thalli, such as old versus new thalli, basal part or frond (Johnson and Mann 1986). Polyphenol content shows a significantly temporal correlation with the reproductive state of the algae. The polyphenol content in Ascophyllum is at a minimum (∼9–10% of dry weight) during the period of maximal fruit body shedding (May) and reaches a maximum (∼12–14%) during the “winter season”. In F. vesiculosus, the minimum (∼8–10%) is 1–2 months later, just before the period of maximum fertility. Thereafter, the content rises to a maximum (∼11–13%) during the period of sterility (August to March; Ragan and Jensen 1978). This does not, however, correspond to the seasonal phenolic content shown in another study by Connan et al. (2004). In that study, the highest phenolic content in F. vesiculosus is during summer. Seasonal variations in phenolic content are more pronounced in some seaweed species like F. vesiculosus and Himanthalia elongata (sea spaghetti) compared to that of L. digitata (Connan et al. 2004). Seasonal studies on the bromophenol content of U lactuca showed that the levels increased dramatically at the end of the summer whilst they decreased during the rest of the year. The bromophenols may protect the seaweed from strong light intensities experienced during the summer (Pedersen et al. 1996; Flodin et al. 1999). The main phenolic compound of marine origin is bromophenol as it is found at high levels in various types of seaweed such as Polysiphonia urceolata (Li et al. 2007).
The levels of antioxidants present in algae can also be affected by a number of other parameters such as location and salinity. Sampath-Wiley et al. (2008) examined the effect of sun exposure and emersion on Porphyra umbilicalis and found that seaweed located in upper intertidal regions during the summer contained higher levels of antioxidants than submerged seaweed. Submersion protected against environmental stresses caused by moisture loss and exposure (Sampath-Wiley et al. 2008). The reducing power of polyphenols was ten times higher in species of Ascophyllum from open coast localities than from specimens from fjords near river outlets, probably because of differences in salinity (Baardseth 1958; Larsen and Haug 1958).
Preliminary studies of polyphenols in seaweed species collected in Denmark show that species of Fucus showed the highest antioxidative activity compared to other brown algal species, e.g. Sargassum and Laminaria. The antioxidant activity was tested with four methods: in vitro established systems such as antioxidant activity in liposome model systems; 1,1-diphenyyl-2-picrylhydrazyl (DPPH) radical scavenging activity; reducing power; and metal chelating activity. Water extracts had the highest antioxidant activity compared to ethanol extracts. Extracts were made from freeze-dried seaweed samples (Farvin et al. 2010).
Drying has no influence on the reducing power of the Fucaceae, but this is not the case for Laminaria and Ascophyllum species. In these species, the reducing power is usually markedly lower in dry samples (Haug and Larsen 1958). Antioxidants from the red seaweed K. alvarezii were extracted and a phenolic content of approx. 2% was found in the chloroform/methanol (2:1) extracts using the Folin–Ciocalteau method (Kumar et al. 2008c). The ethanol extract had the best DPPH scavenging ability (IC50 = 3.03 mg mL−1) and most potent inhibitory effect against lipid peroxidation, whereas the methanol extract exhibited the best metal ion-chelating ability (IC50 = 3.08 mg mL−1) and also had the greatest reducing power. Li et al. (2007) extracted four bromophenol compounds from a red alga P. urceolata; each was found to have DPPH-scavenging ability, with one compound displaying an IC50 value of 9.67 μM.
Polyphenols (fucol, fucophlorethol, fucodiphloroethol G and ergosterol) and the phenolic compound phlorotannin are abundant in brown macroalgae, such as Sargassum, Fucus and A. nodosum, and have strong antioxidant effects (Table 16).
Investigations on crude extracts and phenol content show that the reducing power and hydroxyl radical scavenging activity of some seaweed species (Eucheuma, Kappaphycus and Turbinaria conoides) are found to be higher compared to standard antioxidant (a-tocopherol). In several experiments, the total phenol content of several seaweed species was significantly different in three out of six seaweed species tested. The in vitro antioxidant activities of methanol extracts of all six species tested exhibited dose dependency and increased with increasing concentration of the extract (Ganesan et al. 2008; Kumar et al. 2008a). Studies have shown that there is a synergistic effect with vitamin E (tocopherol; Le Tutour 1990).
Polyphenols are of cosmeceutical and pharmacological value because of their antioxidative activity, and they have also shown other effects such as radiation protective, antibiotic and antidiabetic. Several of these effects were tested in bacteria, cell cultures, rodents and even in humans with regard to sexual performance and desire (Table 16). Certain polyphenols work as preventative medicines for problems such as cardiovascular diseases, cancers, arthritis and autoimmune disorders by helping to protect tissues against oxidative stress (Garbisa et al. 2001; Maliakal et al. 2001; Kang et al. 2003). In addition, polyphenols are found as anti-inflammatory agent and have anti-allergic effect and antibacterial activity (e.g. anti-Staphylococcus activity) together with other broad therapeutic perspectives (Table 16; Li et al. 2009; Zubia et al. 2008; Sailler and Glombitza 1999).
Oral tests on rats and humans with a commercially available sample containing 30% polyphenols and 70% dietary fibres indicated good oral absorption of the compounds and fast binding to the luminal surface of the blood vessels (Kang et al. 2003). Polyphenols have been traditionally thought of as having low bioavailability, but some studies have noted a significant increase in the antioxidant capacity of plasma following ingestion of these compounds in foods and beverages (Arts et al. 2001; Kalt et al. 2000). Epidemiological studies also note an association between a high intake of dietary polyphenols and lowered incidence of various chronic diseases (Arts et al. 2001). Therefore, it could be suggested that phlorotannins would be potential candidates for the development of unique natural antioxidants for further industrial applications as functional foods, cosmetics and pharmaceuticals (Li et al. 2009).
Besides phloroglucinol, there are many other flavenoids and their glycosides present in green, brown and red algae, such as catechin and epicatechin (up to 4.77 mg g−1 dry weight) or the flavonol glycoside rutin (up to 11.4 mg g−1 dry weight) in Porphyra. The flavone glycoside hespiridin is present in green, brown and red seaweeds and is especially high in certain green algae at 117 mg g−1 dry weight.
Halogenated compounds
Halogenated compounds are produced naturally, mainly by red and brown seaweed, dispelling the widespread notion that these chemicals are only of man-made origin (Butler and Carter-Franklin 2004; Flodin et al. 1999). Halogenated compounds are dispersed in several different classes of primary and secondary metabolites, including indoles, terpenes, acetogenins, phenols, fatty acids and volatile halogenated hydrocarbons (e.g. bromoform, chloroform, dibromomethane; Butler and Carter-Franklin 2004; Dembitsky and Srebnik 2002; Flodin et al. 1999). In many cases, they possess biological activities of pharmacological interest, including antibacterial (Vairappan et al. 2001) and anti-tumoural (Fuller et al. 1992). The most notable producers of halogenated compounds in the marine environment belong to the genus Laurencia (Rhodophyta; Faulkner 2001; Wright et al. 2003). The compounds are predominantly derivates of esquiterpenes diterpenes, triterpenes, acetogenins, fatty acids and brominated indoles. In addition to antimicrobial and cytotoxic properties, Laurencia compounds may also play multifunctional ecological roles such as acting as a feeding defendant (Brito et al. 2002; Flodin et al. 1999; Iliopoulou et al. 2002; Suzuki et al. 2002).
Polyhalogenated monoterpenes from red algae of the genera Plocamium and Chondrococcus exhibit a wide range of pharmacological activities, including antimicrobial, anti-tubercular and anticancer activities (Blunt et al. 2003; Darias et al. 2001; Flodin et al. 1999; Fuller et al. 1992; Knott et al. 2005). Brominated fatty acids are synthesised by Asparagopsis and Bonnemaisonia (Dembitsky and Srebnik 2002). The presence of halogen atoms (F, Cl, Br or I) in the fatty alkyl chain causes significant changes in their physicochemical characteristics, increasing their reactivity and changing the conformation of biological membranes (Dembitsky and Srebnik 2002).
Extracts or powder of seaweed
Various extraction methods have been used to release identified and unidentified bioactive substances from marine algae. Powder derived from dried seaweed has also been tested.
Antioxidant effect
The result of 16 lipophilic extracts suggests that seaweed can be considered as a potential source for the extraction of lipophilic antioxidants, which might be used as dietary supplements or in production in the food industry. The results indicated an increase in antioxidative property with increasing content of unsaturated fatty acid (Huang and Wang 2004). Sargassum species might be a valuable source of natural antioxidants containing both water- and fat-soluble antioxidative components, preventing oxidative damage of food oils (Siriwardhana et al. 2003, 2004). Further antioxidative effects of compounds from macroalgae with potential in applications in human health and nutrition are described by Cornish and Garbary (2010).
Zaragoza et al. (2008) investigated the antioxidant behaviour of two extracts from brown seaweed F. vesiculosus using either 30–35% ethanol or 50–70% ethanol. The 30–35% ethanol extract exhibited more potent antioxidant behaviour than the 50–70% ethanol extract, showing superior DPPH, superoxide, peroxyl and ABST radical scavenging ability. The 50–70% ethanol extract was found to increase the reducing power and superoxide scavenging ability in the plasma of laboratory rats. The 50–70% ethanol extract was also found to stimulate the immune system of the animals tested by increasing the Cu–Zn SOD activity by approx. 32% (Dembitsky and Srebnik 2002).
Peroxidation of fatty acids
Addition of 1% or 2% Undaria powder to rat diets decreased liver and serum triglycerol levels, respectively, compared to control. It is known that the synthesis and oxidation of fatty acids by the liver control the concentration of triacylglycerol in the serum and liver. The enzyme activities involved in the synthesis or the oxidation of fatty acids show changes in rats with Undaria powder in diets. Addition of 5–10% Undaria powder decreased glucose-6-phosphate dehydrogenase activity; however, 10% wakame increased several activities of other enzymes and co-enzymes. These results indicate that the decrease in concentration of triacylglycerol in serum and liver by dietary Undaria powder in rats is due to the promotion of fatty acid oxidation in rat liver. Therefore, Undaria may play a role in the prevention and treatment of arteriosclerosis. In addition, since the decrease in concentration of triacylglycerol in the serum and liver by dietary Undaria is associated with the promotion of oxidation of fatty acids in the liver, Undaria may become a food material for the prevention of obesity (Table 17; Murata et al. 1999). Furthermore, fish oil in rat diets was found to decrease the concentration of triacylglycerols in serum and liver significantly compared to control, but this decrease became even more conspicuous in a diet with both fish oil and Undaria powder (Murata et al. 2002). Methanol extract from the brown algae Sargassum inhibits oxidation in rat liver homogenates (Kumar et al. 2008b; Mori et al. 2003).
Other effects
Extracts of seaweed with different solvents such as water, ethanol, dichloromethane and chloroform have shown antibacterial, anti-inflammatory and anti-pathogenic effects (Table 17).
The antibacterial effect of seaweed extracts was seen in several Gram-negative bacteria strains, and the extracts were non-toxic against non-targeted larvae (Andersson et al. 1983; Zubia et al. 2008). Anti-inflammatory effect was seen in oedema and erythema of mice, but there was also decreased motor activity in rodents when seaweed extract was added to their diet (Andersson et al. 1983). Another study on rodents improved the survival of stroke-prone hypertensive rats when given 5% seaweed powder in the diet (Ikeda et al. 2003). Similarly, Undaria powder supplement in rat diet (5% wet weight) attenuated the development of hypertension and its related diseases and improved survival rates (Ikeda et al. 2003). Methanol or enzymatic extracts of Sargassum species were furthermore preventive against DNA damage (Cho et al. 2007; Park et al. 2005). The latter effect was also seen by enzymatic extracts of other brown seaweed (Heo et al. 2005). Seaweed extract stimuli in plant science and agriculture is described in recent publications such as Fan et al. (2011) and the comprehensive review by Craigie (2010).
Feed supplement
Animals such as sheep, cattle and horses that live in the coastal areas are known to actively graze seaweed from the shore. Today, seaweed is dried and milled to a fine seaweed meal, in particular from A. nodosum due to the harvesting history of this species. Analyses show that it contains useful amounts of minerals (K, P, Mg, Ca, Na, chlorine and S), trace metals and vitamins. Because most of the carbohydrates and proteins are not digestible, the nutritional value of seaweed has traditionally been assumed to be in its contribution of minerals, trace elements and vitamins to the diet of animals. The feeding value has been assessed in Norway as having only 30% of the value of grains. Feeding trials with added Ascophyllum meal to poultry and pigs showed an increase in the iodine content of the eggs from the poultry, and no effect was observed in pigs. However, positive results were reported with increases in milk production in dairy cows, whilst sheep were able to maintain weight better during winter feeding, with increases in lambs’ birth weight and greater wool production (McHugh 2003; Hertz 2008, personal communication on the birth weight of lambs).
Recent research on seaweed used as animal feed supplement of either protein, polysaccharides or enriched with PUFAs has given promising results. Moreover, fishmeal protein can be supplemented with no effect or even with a beneficial effect on growth, feed conversion ratio and flesh colour if supplemented with seaweed (Table 18). Soler-Vila et al. (2009) have shown that Porphyra dioica can effectively be included in diets for rainbow trout at up to 10%, without significant negative effects on weight gain and growth performance. The positive pigmentation effect of the fish flesh by adding the Porphyra meal to the feed is of considerable interest to the organic salmon-farming industry. Other research showed that when Ulva meal replaced 5% of the diet lipids, the growth performance, feed efficiency, nutrient utilisation and body composition of Nile tilapia were improved (Ergün et al. 2008). In addition, seaweed meal elevated body weight gain of sea bream and tended to increase feed efficiency and muscle protein deposition. Algae-fed groups were higher in liver glycogen and triglyceride accumulation in muscle. Feeding Porphyra showed the most pronounced effects on growth and energy accumulation, followed by Ascophyllum and Ulva. The results suggest the practical efficacy of using algae as a feed additive for the effective use of nutrients in cultured sea bream (Mustafa et al. 1995).
Many experiments have been performed on seaweed additions in the diets of pigs (including weanling and grower-finisher). When piglets move from suckling to a solid diet, stomach-soling and intestinal problems often appear. Seaweed extracts fed to pigs have been shown to provide a dietary means to improve gut health and potentially reduce pathogen carriage in finishing pigs. However, the effect on growth performance in healthy animals was negative (Gardiner et al. 2008).
Undesirable compounds
Heavy metals
Besides health-promoting and stimulating compounds in seaweed, other metals, minerals and bioactives may pose a negative effect on health. Not only desireable minerals but also metals in toxic concentrations, heavy metals and other compounds are concentrated in seaweed. For example, lead (Pb), mercury (Hg) cadmium (Cd), copper (Cu), manganese (Mn), zinc (Zn) and the explosive compound trinitrotoluene that may be present in the ambient environment (Besada et al. 2009; Cruz-Uribe et al. 2007; Greger et al. 2007; Mehta and Gaur 2005; Ronnberg et al. 1990; Sandau et al. 1996). Concentration factors of 103–104 for the above heavy metals and even up to 106 can be reached for chromium (Cr; references in Lobban and Harrison 1994). This accumulating effect present in seaweed can also be exploited to remediate for undesired compounds in sea and wastewater or in the use of seaweed as a bioindicator (Boubonari et al. 2008; Cruz-Uribe et al. 2007; Mehta and Gaur 2005). However, undesired metals and compounds should be taken into account and tested if seaweed species are to be used as a feed and food product. The concentrations of, e.g., heavy metals depend on the surrounding environment, and variations can be very local.
Inorganic arsenic (As) accumulation in seaweed is an example of another problem because it has an effect on the human nervous system. Marine organisms take up the inorganic arsenic, but usually, the biological system transforms it into an organic and non-toxic form of arsenic. Arsenic concentrations are generally considerably higher in brown seaweed than in red or green (Francesconi and Edmonds 1996). In most species of seaweed, the concentrations are below 54 mg kg−1 dry weight, and 5-10% of the total arsenic is organic, primarily in the form of various arseno-sugars (Table 19; Almela et al. 2002; Francesconi and Edmonds 1996). Fucus readily accumulates arsenate and transforms it into several arsenic compounds depending on the exposure concentration and other major arsenic species. Arsenite, methylarsonate and dimethylarsinate can be transformed to e.g. arsenosugars (Geiszinger et al. 2001). However, the seaweed species Sargassum fusiforme (and possibly other species in the Sargassum family) concentrate large amounts of the inorganic form (88 mg kg−1 dry weight, up 72% of total As content; Table 19). This edible brown seaweed, S. fusiforme (former Hizikia fusiformes), is sold as Hijiki or Hiziki from Japan, and the S. muticum may only contain 38–75% of their arsenic in organic forms (Almela et al. 2002; Shinagawa et al. 1983; Yasui et al. 1978). Arsenic in Sargassum is removed by 89–92% (wet weight) by the cooking process, or effectively removed just by soaking edible brown seaweed. These experiments were tested on mice (Ichikawa et al. 2006). However, this effect has not been observed in vegetables where cooking is only of a very limited value as a means of reducing metal concentrations (arsenic, cadmium, mercury and lead; Perelló et al. 2008). Figures of these compounds in seaweed sold as food or in coastal seaweed in Ireland are shown in Table 19.
Kainic acid
Kainic acid is a neurotoxin that is similar to domoic acid (a neurotoxin which causes amnesic shellfish poisoning), an amino acid associated with certain harmful algal blooms. This acid is a natural marine acid present in some seaweed, and it is a potent central nervous system stimulant. Kainic acid was originally isolated from the seaweed called “Kainin-sou” or “Makuri” (D. simplex) in Japan in 1953. “Kainin-sou” is used as an anthelmintic in Japan. Moreover, it has been developed as the prototype neuroexcitatory amino acid for the induction of seizures in experimental animals, at typical doses of 10–30 mg kg−1 in mice. Because of recent supply shortages, the price of kainic acid has risen significantly (Ramsey et al. 1994). Three different companies quote prices ranging from US $3 mg−1 to US $12 mg−1 for 99% pure kainic acid (glutamate-free). P. palmata var. sarniensis seems to have some levels of kainic acid which, after metabolite extraction, produced 0.72% kainic acid. Kainic acid can be extracted from these sources by treatment with a mixture of chloroform and methanol (1:2, v/v).
The concentration of kainic acid was higher in wild-harvested populations of P. palmata (12 and 130 mg kg−1 dry weight in Ireland and France, respectively) compared to cultivated material (2.5 mg g−1 dry weight). Experiments have shown that this neurotoxin is not transferred to abalone fed on Palmaria (Lüning 2008).
Legislation, bioactive molecules and seaweed as food
Seaweed species and their components must meet certain consumer safety regulations. The seaweed species authorised for human consumption have been defined in France (Table 20). Moreover, maximum allowed levels of toxic minerals (lead, cadmium, tin, mercury, mineral arsenic and iodine) have been defined for all edible seaweed. These low levels are considered a high guarantee of food safety; in fact, heavy metal norms for certain fish and shellfish are much higher (e.g. 1 mg kg−1 dry weight for mercury in tuna in the USA; 0.7 mg kg−1 in France; Mabeau and Fleurence 1993).
No legislation regarding contaminants in algae food products is currently present in Spain. Some samples analysed from legal seaweed food products did, however, reveal cadmium and inorganic arsenic levels higher than those permitted by legislation in other countries (Table 21). Given the high concentrations of inorganic arsenic found in S. fusiforme, a daily consumption of 1.7 g of the product would reach the Provisional Tolerable Weekly Intake recommended by the WHO for an average body weight of 68 kg (Almela et al. 2002). The maximal content of neurotoxins (not specified) is 20 ppm on a fresh weight basis for mussels, but no limits seem to be mentioned for seaweed, e.g. kainic acid in Palmaria sp. (Lüning 2008).
Seaweed species are not subject to particular regulations in other European countries, but are considered as food if put on market as food or food ingredient and consumed to a significant degree before May 15 1997, and therefore not considered as novel food. However, other specific legislation may restrict the placing on the market in some Member States. As an example, Fucus serratus and Ulva sp. would be considered novel food in Denmark (European Commission 1997). Algal products have to comply with the upper level limits given in Table 21.
International legislation on food supplements, fortified and functional foods
Functional foods are continuing to grow in popularity around the world, but there is currently no universal definition of the category. Functional food is a category recognised by the industry only and is a virtual category in legal terms in that there is no specific “functional food” legislation. Authorisation and labelling of “functional food” falls under existing legislation that governs all food.
Functional foods originated in the Far East as an effort to reduce increasing health costs in the public sector. Industrial functional foods have blossomed, first in Japan and then in the US, and these contries have the majority of the value; only a modest increase has been achieved in the EU of eight billion euros. The key reason for this is the incoherent EU legislation because no European legislation exists specifically for the health benefits (efficacy and health claims). The combined US, Western European and Asia Pacific functional food and drink market are worth US $72.3 billion, and the market is forecast to grow at a compound annual growth rate of 5.7% between 2007 and 2012. Asia will remain the largest functional food market, and it is forecast to continue to outperform the US and European markets in the foreseeable future. The buoyant growth throughout the sector is being driven by consumers who are increasingly looking for products that offer a solution to both long-term and short-term health problems (Gruenwald 2008; Korver et al. 2008).
The situation in the EU contrasts with Japan and the US. An American company deals only with one regulatory body, the Food and Drug Administration (FDA), and the same applies for Japan. In Europe, however, a variety of national regulators have to be dealt with. The European and US markets are quite different in structure, market positioning, consumer and reseller attitudes and expectations, regulations, labelling and claims, value chain, and even product specifications and packaging. Furthermore, there is no GRAS protocol in Europe, and many functional foods have to obtain “novel foods” approval, a potentially lengthy and expensive process with no guarantee of success.
Europe
The European Commission Concerted Action on Functional Food Science in Europe regards a food as functional if it is satisfactorily demonstrated to affect beneficially one or more target functions in the body, beyond adequate nutritional effects, in a way that is relevant to either an improved state of health and well-being and/or reduction of risk of disease. In this context, functional foods are not pills or capsules, but must remain foods, and they must demonstrate their effects in amounts that can normally be expected to be consumed in the diet (EAS 2008).
United States
The US FDA does not provide a legal definition for the term ‘functional foods’, and the term is currently used primarily as a marketing idiom for the category. The FDA is still reviewing comments on how functional foods should be regulated before considering further actions (Heller 2009). Comments filed with the FDA follow a public hearing held in 2006 designed to examine whether the current regulatory framework is suitable for the overview of functional foods or whether the FDA needs to change the way it regulates this new category. At the public hearing, the agency took into account recommendations from the Government Accountability Office, the International Life Sciences Institute, the Institute of Food Technologists (IFT) and a citizen’s petition from advocacy group Center for Science in the Public Interest. The FDA is currently in the process of evaluating all comments submitted on this issue, and once it has done this, the FDA will consider further action, if necessary. However, no timeline has been issued (Heller 2009). A number of working definitions have been developed though, by different organisations, including the American Dietetic Association (ADA), the International Food Information Council (IFIC), and the IFT. ADA classifies all foods as functional at some physiological level because they provide nutrients or other substances that furnish energy, sustain growth or maintain/repair vital processes. However, functional foods move beyond necessity to provide additional health benefits that may reduce disease risk and/or promote optimal health. The IFIC considers functional foods to include any food or food component that may have health benefits other than basic nutrition. These substances provide essential nutrients, often beyond quantities necessary for normal maintenance, growth and development, and/or other biologically active components that impart health benefits or desirable physiological effects.
“Medical foods” and “foods for special dietary use” are considered to be two subcategories of functional foods in the US.
Medical foods are defined by the Orphan Drug Act. “A food which is formulated to be consumed or administered enterally under the supervision of a physician and which is intended for the specific dietary management of a disease or condition for which distinctive nutritional requirements, based on recognised scientific principles, are established by medical evaluation”.
Foods for special dietary use are defined by the Federal Food, Drug, and Cosmetic Act (Section 411(c)(3)) as “a particular use for which a food purports or is represented to be used, including but not limited to the following: 1. Supplying a special dietary need that exists by reason of a physical, physiological, pathological, or other condition….; 2. Supplying a vitamin, mineral, or other ingredient for use by humans to supplement the diet by increasing the total dietary intake. 3. Supplying a special dietary need by reason of being a food for use as the sole item of the diet…”. Examples of such include infant foods, hypoallergenic foods such as gluten-free and lactose-free foods, and foods offered for reducing weight (Heller 2009).
Canada
Health Canada defines functional foods as “similar in appearance to, or may be, a conventional food, is consumed as part of a usual diet, and is demonstrated to have physiological benefits and/or reduce the risk of chronic disease beyond basic nutritional functions”.
Japan
Japan is the only country that recognises functional foods as a distinct category, and the Japanese functional food market is now one of the most advanced in the world. Known as “foods for specified health use”, these are foods composed of functional ingredients that affect the structure and/or function of the body and are used to maintain or regulate specific health conditions, such as gastrointestinal health, blood pressure and blood cholesterol levels (EAS 2008).
Food supplements
Ireland
The 2003 Irish food supplements regulation (S.I. No. 539) lays down specific rules for the use of vitamins and minerals in food supplements. Herb and other substances are not specifically covered at this stage (EAS 2008). The Irish regulation lists the vitamin and mineral substances permitted for use in food supplements, in line with those listed in Annex II of the EU Food Supplement Directive. The Irish Food Safety Authority (FSAI) has granted derogations for 210 substances until the end of 2009. No minimum or maximum levels have been set in the 2003 regulations; however, the Irish authorities indicate that the level should not exceed 1× the recommended daily intake as listed in the 2005 nutrition labelling for foodstuffs regulations (S.I. No. 65 of 2005). Bioactive substances are evaluated on a case-by-case basis and classified as foods or medicines by the Irish Medicine Board (IMB). When substances are to be considered safe, the FSAI generally does not object to their use as a food supplement. To this purpose, the IMB also communicates ingredient/product classification decisions to the FSAI. The FSAI may ask the IMB for an opinion, in case of doubt. According to IMB guidelines, amino acids may only be used at levels usually present in a normal diet. This is, however, not enforced, except for tryptophan (EAS 2008).
The regulations require a food supplement product to be notified by forwarding a completed notification form, a model of the label and any relevant references. This requirement applies when the product is manufactured and is being placed on the Irish market for the first time. Once notification has taken place, food products can be placed on the market and sold through a variety of outlets, such as supermarkets, pharmacies, drugstores and health food stores.
Fortified and functional foods
European Union
The new EU Regulation 1925/2006 on the addition of vitamins and minerals and certain other substances to food came into force in Ireland in July 2007. The legislation currently provides clear rules for the addition of vitamins and minerals to food, whilst herbal ingredients and other substances are not explicitly mentioned in the Regulation. There is currently no additional national legislation on food fortification or functional foods (EAS 2008). With regard to functional foods, amino acids, fish oil and lactic bacteria can generally be used in foods as long as the resulting product is safe. There is no notification requirement for foods with added substances, and products can be sold via all outlets.
The underlying principle of all legislative developments is that the food manufacturer must be able to substantiate every health claim on any functional food that is marketed. The question of “what is sufficient scientific evidence?” remains a problem (Korver et al. 2008). Although the safety aspects of new functional foods are covered in the EU Novel Foods legislation (258/97/EC as amended by 1829/2003/EC and 1882/2003/EC), no European legislation exists specifically for the health benefits (efficacy and health claims). Until there is such legislation, EU Member States will each continue to have their own set of rules and regulations. A proposal was submitted by the European Commission in 2003 on nutrition and health claims to better inform the consumer and harmonise the market. It was hoped that this would transpose into law by 2005 (Korver et al. 2008). Negotiations between EU Member States about a legislative framework for health claims have been going on for many years. At the crux of the problem are the different views on functional foods in various EU countries. For instance, in some countries, especially in Southern Europe, the attitude on functional foods is generally fairly positive and similar to the US. In other countries, functional foods tend to be seen as medicines, and in Germany, for example, legislators have tried to push them into the pharmaceutical regulatory system. Scandinavia is traditionally very reserved towards industry involvement and wants safeguards against possible abuse (Korver et al. 2008). Nevertheless, the regulation was adopted as Regulation (EC) No. 1924/2006 of the European Parliament and of the Council on Nutrition and Health Claims made on foods by the end of 2006. This Regulation lays down harmonised rules for the use of nutrition and health claims and contributes to a high level of consumer protection. It ensures that any claim made on a food label in the EU is clear, accurate and substantiated, enabling consumers to make informed and meaningful choices. The Regulation also aims to ensure fair competition and promote and protect innovation in the area of food. Regulation EC No. 1924/2006 on nutrition and health claims made on foods came into force on 19 January 2007 and has been applied from 1 July 2007.
Health claims
A “health claim” means any claim that states, suggests or implies that a relationship exists between a food category, a food or one of its constituents and health. The nutrition and health claims Regulation 1924/2006 does what the name says and is something that innovative food industries and those carrying out research and development should be aware of because it has a major bearing on the marketability of a potential food or ingredient. A ‘positive list’ of well-established claims should be elaborated within 3 years of the Regulation entering into force. This list should describe the role of a nutrient or of another substance in growth, development and the normal functions of the body, which are based on generally accepted scientific data and well understood by the average consumer. As an example, the claim that calcium plays an important role in the strengthening of teeth and bones would be permitted. For more novel claims, scientific evaluation and premarketing approval will be required. Implied health claims will not be allowed, for example claims which make reference to general non-specific benefits of the nutrient or food for overall good health. Claims will not be permitted that make reference to the advice of doctors, other health professionals, their professional associations or charities, or suggest that health could be affected by not consuming the food (EAS 2009).
Proactive companies seeking to keep ahead of the competition will be assisted by a European Commission decision to fast track health claims based on newly developed science and a decision to give exclusive use for 5 years of claims requiring proprietary data protection (EAS 2009). Article 13.5 allows data protection, which means that competitors must have similar data of their own to submit in order to make the same claim. The decision to allow these claims to be submitted in February 2008 rather than waiting until the final cutoff in 2010 will boost industry competitiveness on the European Union food market. Claims which are not based on proprietary data, but which are currently in use, should already be scientifically justified. This makes it difficult to say the claim is based on newly developed scientific evidence. Such claims should be introduced into the Article 13 list before the deadline of 31 January 2009. If they are not, then the companies would either have to withdraw their claim when the claims list is established in 2010 or in the intervening time submit an application to have it approved under the Article 13.5 procedure (EAS 2009).
If a company makes a nutrition or health claim about a product, it must comply with Regulation 1924/2006 on nutrition and health claims made on foods. Permitted nutrition claims are listed in the Annex to the legislation. A list of approved health claims and conditions for their use should have been published by the European Commission by 31 January 2010 at the latest, but it has not yet been released. Health claims not on this list will not be permitted.
It is the food business operator’s responsibility to comply with this Regulation. Claims that are not compliant will not be permitted.
Novel food authorisation in the EU
A novel food is any food or ingredient that does not have a significant history of consumption in the EU prior to 15 May 1997 (European Commission 1997). This legislation is currently being revised, but will be in place for another year or so yet. To introduce a novel food or ingredient to the EU market, prior authorisation is required through this Regulation, which includes a safety assessment carried out by a Member State and possibly also by the European Food Safety Authority. Final authorisation is completed by a voting process (Comitology) by Member States.
Authorisation to market a novel food in Europe, which includes manufacturing processes, requires a rigorous assessment of toxicological, nutritional, compositional and other relevant data by the competent authorities of EU Member States. A new food does not qualify as a ‘novel’ food and may be placed on the market within the EU after notification to the European Commission if it is proven to be substantially equivalent to one already on the market. However, a food or food manufacturing process is considered ‘novel’ if it has not been available within the EU (at least one Member State) to a significant degree prior to 15 May 1997 (Novel Food Regulation EC 258/97; European Commission 2007). The authorisation process is a multi-step procedure with defined time limits coordinated by the European Commission (DG SANCO) and involves the applicant and EU Member States with their expert advisory personnel (O’Mahony 2009, personal communication). This process is outlined in Fig. 1.

Procedure for authorisation of novel food within EU (FSAI 2010)
The FSAI became the competent authority for novel foods, taking over from the Department of Health and Children on 1 January 2001. The FSAI is a consumer-oriented, science-based independent body and draws on the expertise of scientists throughout the country for advice. The FSAI is responsible for the premarketing safety assessment of any novel food to be marketed in Ireland and Europe (Fig. 2). Whilst Ireland has not yet had any direct applications, it has reviewed and provided comments with the help of its scientific advisers on novel food assessments by other Member States. The FSAI is also responsible for ensuring compliance with novel food regulations in Ireland, including appropriate labelling laws. The FSAI handles all aspects of novel foods, including application assessments, regular surveys of the marketplace, and dissemination of information on procedures and legislation (FSAI 2010).

Management of novel foods (FSAI 2010)
One to two novel food applications are assessed on average each month by EU Member States, whilst a number of others are simply notified to the Commission as being substantially equivalent. To have a novel food marketed within the EU, a company must prepare a dossier detailing prescribed safety-related scientific data and present it to a lead Member State along with an assessment fee that varies between Member States. An initial assessment of the data is carried out by the ‘lead’ Member State that presents a report to the Commission, which in turn provides it for other Member States. The summary report and initial assessment are then critically reviewed by Member States before a decision is made. A positive decision means that the applicant is free to market the product within the EU. Reasoned objections, or a request for further assessment, lead to the application being referred to the Standing Committee on Food who may seek advice from the Scientific Committee on Food before making its own decision through a qualified majority vote. It is referred to the Council of Ministers who may decide to accept or reject a Commission proposal by qualified majority if the Standing Committee on Food fails to reach a decision. The Commission will be free to implement its proposal as it stands if the Council of Ministers fails to successfully address the issue within 90 days.
Novel foods containing, or derived from, genetically modified organisms (GMOs) have attracted much attention in recent years. This notoriety has subsided despite the de facto moratorium that has been in place on such foods, resulting in numerous foods derived from GMOs (GM food) being stranded in the authorisation process. In the meantime, other non-GM novel food applications are being assessed and, whilst most are eventually authorised, a small number have been rejected. However, the FSAI anticipates this to be an active area for the foreseeable future with the expected evolution of current and pending EU novel food regulations and the increasing numbers of foods and food processes seeking a marketplace within the EU. The constant vigilance and heightened awareness of the EU and its Member States on the issue of novel foods can be justified by consumer concerns about the food supply and in light of serious food scares of the recent past (O’Mahony 2009, personal communication). Legislation has appeared in piecemeal fashion over the years as and when required. What usually happens is that consolidated legislation is developed which simplifies matters into one or a few documents, but advances in food production and processing are still growing, so consolidation is still a long way.
The FSAI has interaction with the IMB. Unlike most food legislation, medicine legislation is not as harmonised in the EU. Therefore, interpretation can vary between Member States, whilst there is common EU medicinal legislation. What this means is that what one Member State considers a medicine, another may not. When the IMB considers an ingredient medicinal, they have the prerogative to bring it under medicines control, though not a written rule. A claim that a food can bring about the treatment or cure of a disease automatically brings the product under medicine legislation. The nutrition and health claim legislation will only affect the marketing of a product and not its legal status as long as medicinal claims are not made.
Food supplements (certain vitamins and minerals) are covered by Directive 2002/46, and to be able to use these ingredients in dietary supplements, they must be listed in the annex to this legislation. To introduce new vitamins and minerals into supplements would firstly require a safety assessment and authorisation under the Novel Food Regulation 258/97. Those supplements containing non-vitamin or mineral ingredients (herbal supplements) would fall under novel food legislation (Novel Food Regulation 258/97).
The Food Fortification Regulation 1925/2006 sets out standards of purity and other conditions for the addition of vitamins, minerals and certain other substances to foods (fortification). Only vitamins and minerals (and their formulations) listed in the Annex may be added to foods, and to add a new one would require authorisation through novel food legislation (1925/2006).
Summary and conclusion
This review summarises the potential of bioactive molecules in species that could be of interest for utilisation for health and functional food applications. The compounds of interest need to be identified before making choices of which species to exploit because one species may be high in one compound, but low in another.
Most of the polysaccharides stimulate human health, for example by the creation of a better intestinal environment or because of their antiviral, anti-tumour and hypocholesterolemic and hypolipidemic properties. As an example, fucans are in large concentrations in large brown seaweed species. These are easy to isolate and have numerous health benefits, which give them the potential to serve as a valuable bioactive ingredient in natural health foods. The choice of species to exploit the content of bioactive compounds relies on the aim and property of the polysaccharide. Proteins, peptides and amino acids from seaweed have shown positive bioactive effects in the treatment of diabetes, cancer, and AIDS and the prevention of vascular diseases. The amino acid profile of some seaweed species is similar to that of animal foods. Extracts of valuable amino acids for feed supplement could be of potential because essential amino acids cannot be replaced by other compounds. The essential amino acids need to be present in the right quantities in order to make proteins. The genera Ulva, Laminaria, Saccharina, Sargassum and Chondrus have some of the best profiles for the amino acids needed in fish feed. Seaweed lipids have high percentages of the omega-3 fatty acids and have been tested positive as immune stimulants, whilst the fucosterol content decreases the concentration of cholesterol in animal and humans. In addition to this, other general effects of PUFAs include anti-arteriosclerosis, anti-hypertension, anti-inflammation and immunoregulation. The pigments have shown bioactivity effect towards, e.g. cancer, obesity, decreasing blood glucose and as an antioxidant. Fucus, Ascophyllum and Sargassum are some of the best sources of phenols for antioxidant purposes. The phlorotannins show bioactive effects such as antioxidant activity, radiation protection, antibiotics, anti-diabetes, etc. Phlorotannins would be potential candidates for the development of natural antioxidants for further industrial applications such as functional foods, cosmetics and pharmaceuticals.
Powder and extracts of seaweed have shown bioactive effects such as antioxidant, peroxidation of fatty acids, antibacterial, anti-fungical and anti-inflammatory. Almost all genera of seaweed have shown effects. Effects on animals when seaweeds are used as ingredients in food have been interesting, including higher milk yield in cattle, increased growth rate of lambs, increased iodine concentration in tissues and change in intestinal bacterial communities of pigs, and accumulation of triglycerides in sea bream. In several of the experiments, growth performances are increased or at least there are no negative effects. Seaweed species are potentially good candidates as feed supplements, with beneficial effects on health or the desired properties of the animal and product.
However, seaweed not only accumulates the desired minerals and trace metals but also accumulates undesirable metals and heavy metals from the surrounding environment. This can be exploited, and seaweed can be used as a biofilter. Kainic acid and high inorganic arsenic are toxic and accumulated in species such as P. palmata and Sargassum sp., respectively.
This review finishes with rules and regulations in respect to heavy metals in seaweed for human consumption and other EU regulations on food supplements and health claims. There are different views in respect to seaweed as food. For example, edible seaweed species have been approved as a human food product in France and are recognised by the EU. Legislative aspects are compared with the current situations in the US, Japan and some other major players in the functional food markets.
Clearly, it is not easy to market a bioactive component derived from seaweed because one has to adhere to the current legislation, which is continuously changing and not very transparent. Other licences are required to source material for processing and extraction and to deal with waste issues that might arise from extraction. In order to market seaweed and/or bioactive compounds derived from seaweed as novel or functional food and make health claims, one needs proven scientific evidence and an extensive scientific dossier needs to be submitted to the EU. This is often an expensive exercise.
Legislation is complicated and will become even more complicated with time, according to experts at food safety authorities. The high cost of scientific dossier (if a new activity or compound is discovered) makes it difficult to further develop seaweed as a functional food ingredient. Nevertheless, seaweed as a whole food product, or bioactive compounds extracted from seaweed, show a tremendous potential for human health and should be further developed and exploited.
References
Aisa Y, Miyakawa Y, Nakazato T, Shibata H, Saito K, Ikeda Y, Kizaki M (2005) Fucoidan induces apoptosis of human HS-Sultan cells accompanied by activation of caspase-3 and down-regulation of ERK pathways. Am J Hematol 78:7–14
Alekseyenko TV, Zhanayeva SY, Venediktova AA, Zvyagintseva TN, Kuznetsova TA, Besednova NN, Korolenko TA (2007) Antitumor and antimetastatic activity of fucoidan, a sulfated polysaccharide isolated from the Okhotsk sea Fucus evanescens brown alga. Bull Exp Biol Med 143:730–732
Almela C, Algora S, Benito V, Clemente MJ, Devesa V, Suner MA, Velez D, Montoro R (2002) Heavy metal, total arsenic, and inorganic arsenic contents of algae food products. J Agric Food Chem 50:918–923
Amano H, Kakinuma M, Coury DA, Ohno H, Hara T (2005) Effect of a seaweed mixture on serum lipid level and platelet aggregation in rats. Fish Sci 71:1160–1166
Andersson L, Lidgren G, Bohlin L, Magni L, Ogren S, Afzelius L (1983) Studies of Swedish marine organisms. 1. Screening of biological-activity. Act Pharm Suec 20:401–414
Aneiros A, Garateix A (2004) Bioactive peptides from marine sources: pharmacological properties and isolation procedures. J Chromatogr B Analyt Technol Biomed Life Sci 803:41–53
Arasaki S, Arasaki T (1983) Low calorie, high nutrition vegetables from the sea to help you look and feel better. Japan Publications, Tokyo, 196 pp
Archer GS (2005) Reducing stress in sheep by feeding the seaweed Ascophyllum nodosum. Texas A&M University, 129 pp
Arnold TM, Targett NM (2000) Evidence for metabolic turnover of polyphenolics in tropical brown algae. J Chem Ecol 26:1393–1410
Arts ICW, Hollman PCH, De Mesquita HBB, Feskens EJM, Kromhout D (2001) Dietary catechins and epithelial cancer incidence: the Zutphen Elderly Study. Int J Cancer 92:298–302
Astorg P (1997) Food carotenoids and cancer prevention: an overview of current research. Trends Food Sci Technol 8:406–413
Athukorala Y, Lee KW, Kim SK, Jeon YJ (2007) Anticoagulant activity of marine green and brown algae collected from Jeju Island in Korea. Bioresour Technol 98:1711–1716
Augier H, Santimone M (1978) Contribution to study of composition in total nitrogen, proteins and proteinic amino-acids of different parts of thallus of Laminaria digitata (Huds) Lamour in scope of its industrial and agricultural exploitation. Bull Soc Phyc France 110:19–28
Baardseth E (1958) A method of estimating the physode content in brown algae-reprint of report 20. Papers Presented at the Third International Seaweed Symposium by the Staff of the Norwegian Institute of Seaweed Research, Norwegian Institute of Seaweed Research, Trondheim. Report 20, pp 1–6
Baardseth E, Haug A (1953) Individual variation of some constituents in brown algae, and reliability of analytical results. Norwegian Institute of Seaweed Research, Akademisk Trykningssentral, Oslo. Report 2, pp 1–23
Bansemir A, Blume M, Schröder S, Lindequist U (2006) Screening of cultivated seaweeds for antibacterial activity against fish pathogenic bacteria. Aquaculture 252:79–84
Barbarino E, Lourenço S (2005) An evaluation of methods for extraction and quantification of protein from marine macro- and microalgae. J Appl Phycol 17:447–460
Bartsch I, Wienke C, Bischof K, Buchholz CM, Buck BH, Eggert A, Feuerpfeil P, Hanelt D, Jacobsen S, Karez R et al (2008) The genus Laminaria sensu lato: recent insights and developments. Eur J Phycol 43:1–86
Benvegnu T, Sassi J-F (2010) Oligomannuronates from seaweeds as renewable sources for the development of green surfactants. Top Curr Chem 294:143–164
Berteau O, Mulloy B (2003) Sulfated fucans, fresh perspectives: structures, functions, and biological properties of sulfated fucans and an overview of enzymes active toward this class of polysaccharide. Glycobiology 13:29R–40R
Besada V, Andrade JM, Schultze F, Gonzalez JJ (2009) Heavy metals in edible seaweeds commercialised for human consumption. J Mar Syst 75:305–313
Bhakuni DS, Rawat DS (2005) Bioactive metabolites of marine algae, fungi and bacteria. In: Bioactive marine natural products. Co-published by Anamaya Publishers and Springer, New Delhi, pp 1–25
Bhaskar N, Hosokawa M, Miyashita K (2004) Comparative evaluation of fatty acid composition of different Sargassum (Fucales, Phaeophyta) species harvested from temperature and tropical waters. J Aquat Food Prod Technol 3:53–70
Bird KT, Chiles TC, Longley RE, Kendrick AF, Kinkema MD (1993) Agglutinins from marine macroalgae of the southeastern United States. J Appl Phycol 5:213–218
Bitencourt FD, Figueiredo JG, Mota MRL, Bezerra CCR, Silvestre PP, Nascimento VMR, KS SAH, Nagano CS, Saker-Sampaio S et al (2008) Antinociceptive and anti-inflammatory effects of a mucin-binding agglutinin isolated from the red marine alga Hypnea cervicornis. Naunyn-Schmiedebergs Arch Pharmacol 377:139–148
Bixler HJ, Porse H (2010) A decade of change in the seaweed hydrocolloids industry. J Appl Phycol. doi:10.1007/s10811-010-9529-3
Black WAP (1950) The seasonal variation in the cellulose content of the common Scottish Laminariaceae and Fucaceae. J Mar Biol Assoc UK 29:379–387
Blunt JW, Copp BR, Munro MHG, Northcote PT, Prinsep MR (2003) Marine natural products. Nat Prod Rep 20:1–48
Bobin-Dubigeon C, Lahaye M, Barry JL (1997) Human colonic bacterial degradability of dietary fibres from sea-lettuce (Ulva sp.). J Sci Food Agric 73:149–159
Bogentoff CB (1981) Antacid composition for neutralizing stomach acid. Germany Patent No. 2,722,484
Borgo E (1984) Pharmaceutical compositions for the treatment of gastric hyperacidity, gastritis and gastroduodenal ulcers. Canada Patent No. 1,176,984
Boubonari T, Malea P, Kevrekidis T (2008) The green seaweed Ulva rigida as a bioindicator of metals (Zn, Cu, Pb and Cd) in a low-salinity coastal environment. Bot Mar 51:472–484
Brito I, Cueto M, Díaz-Marrero AR, Darias J, Martin AS (2002) Oxachamigrenes, new halogenated sesquiterpenes from Laurencia obtusa. J Nat Prod 65:946–948
Bruhn A, Rasmussen MB, Olesen B, Worm T (2008) Kommerciel dyrkning af carrageentang (Chondrus crispus) i danske farvande. Danmarks Miljøundersøgelser, Silkeborg, 62 pp (in Danish)
Buck CB, Thompson CD, Roberts JN, Muller M, Lowy DR, Schiller JT (2006) Carrageenan is a potent inhibitor of papillomavirus infection. PLoS Pathog 2:671–680
Burtin P (2003) Nutritional value of seaweeds. Electron J Environ Agric Food Chem 2:498–503
Butler A, Carter-Franklin JN (2004) The role of vanadium bromoperoxidase in the biosynthesis of halogenated marine natural products. Nat Prod Rep 21:180–188
Caceres PJ, Carlucci MJ, Damonte EB, Matsuhiro B, Zuniga EA (2000) Carrageenans from Chilean samples of Stenogramme interrupta (Phyllophoraceae): structural analysis and biological activity. Phytochemistry 53:81–86
Cann SAH (2006) Hypothesis: dietary iodine intake in the etiology of cardiovascular disease. J Am Coll Nutr 25:1–11
Cardozo KHM, Guaratini T, Barros MP, Falcão VR, Tonon AP, Lopes NP, Campos S, Torres MA, Souza AO, Colepicolo P et al (2007) Metabolites from algae with economical impact. Comp Biochem Physiol C: Toxicol Pharmacol 146:60–78
Carlucci MJ, Pujol CA, Ciancia M, Noseda MD, Matulewicz MC, Damonte EB, Cerezo AS (1997) Antiherpetic and anticoagulant properties of carrageenans from the red seaweed Gigartina skottsbergii and their cyclized derivatives: correlation between structure and biological activity. Int J Biol Macromol 20:97–105
Carthew P (2002) Safety of carrageenan in foods. Environ Health Perspect 110:A176
Carvalho MDT, Harada LM, Gidlund M, Ketelhuth DFJ, Boschcov P, Quintao ECR (2002) Macrophages take up triacylglycerol-rich emulsions at a faster rate upon co-incubation with native and modified LDL: an investigation on the role of natural chylomicrons in atherosclerosis. J Cell Biochem 84:309–323
Cérantola S, Breton F, Ar Gall E, Deslandes E (2006) Co-occurrence and antioxidant activities of fucol and fucophlorethol classes of polymeric phenols in Fucus spiralis. Bot Mar 49:347–351
Cha SH, Ahn GN, Heo SJ, Kim KN, Lee KW, Song CB, Cho SK, Jeon YJ (2006) Screening of extracts from marine green and brown algae in Jeju for potential marine angiotensin-I converting enzyme (ACE) inhibitory activity. J Korean Soc Food Sci Nutr 35:307–314
Chapman VJ (1970) Seaweeds and their uses, 2nd edn. Methuen, London, 304 pp
Chen H-M, Zheng L, Yan X-J (2005) The preparation and bioactivity research of agaro-oligosaccharides. Food Technol Biotechnol 43:29–36
Chernomorsky S, Segelman A, Poretz RD (1999) Effect of dietary chlorophyll derivatives on mutagenesis and tumor cell growth. Tera Carc Mut 19:313–322
Chevolot L, Foucault A, Chaubet F, Kervarec N, Sinquin C, Fisher AM, Boisson-Vidal C (1999) Further data on the structure of brown seaweed fucans: relationships with anticoagulant activity. Carbohydr Res 319:154–165
Cho SH, Kang SE, Cho JY, Kim AR, Park SM, Hong YK, Ahn DH (2007) The antioxidant properties of brown seaweed (Sargassum siliquastrum) extracts. J Med Food 10:479–485
Choi YS, Hong SR, Lee YM, Song KW, Park MH, Nam YS (1999) Study on gelatin-containing artificial skin: I. Preparation and characteristics of novel gelatin–alginate sponge. Biomaterials 20:409–417
Chopin T, Gallant T, Davison I (1995) Phosphorus and nitrogen nutrition in Chondrus crispus (Rhodophyta): effects on total phosphorus and nitrogen content, carrageenan production, and photosynthetic pigments and metabolism. J Phycol 31:283–293
Chopin T, Sharp G, Belyea E, Semple R, Jones D (1999) Open-water aquaculture of the red alga Chondrus crispus in Prince Edward Island, Canada. Hydrobiologia 398/399:417–425
Chronakis IS, Galatanu AN, Nylander T, Lindman B (2000) The behaviour of protein preparations from blue-green algae (Spirulina platensis strain Pacifica) at the air/water interface. Colloid Surface Physicochem Eng Aspect 173:181–192
Cloughley G, Burbridge E, Kraan S, Guiry MD, Smyth PPA (2008) Iodine uptake and efflux by macroalgae. 11th International Conference on Applied Phycology, Galway, Ireland, p 81
Connan S, Goulard F, Stiger V, Deslandes E, Ar Gall E (2004) Interspecific and temporal variation in phlorotannin levels in an assemblage of brown algae. Bot Mar 47:410–416
Connan S, Delisle F, Deslandes E, Ar Gall E (2006) Intra-thallus phlorotannin content and antioxidant activity in Phaeophyceae of temperate waters. Bot Mar 49:39–46
Constantinides P, Cairns A, Werner A (1954) Antilipemic activity of sulfated polysaccharides. Arch Int Pharmacodyn Thér 99:334–345
Cornish ML, Garbary DJ (2010) Antioxidants from macroalgae: potential applications in human health and nutrition. Algae 25:000–000. doi:10.4490/algae.2010.25.4.000
Craigie JS (2010) Seaweed extract stimuli in plant science and agriculture. J Appl Phycol. doi:10.1007/z10811-010-9560-4
Cruz-Uribe O, Cheney DP, Rorrer GL (2007) Comparison of TNT removal from seawater by three marine macroalgae. Chemosphere 67:1469–1476
Dannhardt G, Kiefer W (2001) Cyclooxygenase inhibitors—current status and future prospects. Eur J Med Chem 36:109–126
Darias J, Rovirosa J, San Martin A, Diaz AR, Dorta E, Cueto M (2001) Furoplocamioids A-C, novel polyhalogenated furanoid monoterpenes from Plocamium cartilagineum. J Nat Prod 64:1383–1387
Dawczynski C, Schubert R, Jahreis G (2007) Amino acids, fatty acids, and dietary fibre in edible seaweed products. Food Chem 103:891–899
Dembitsky VM, Rozentsvet OA (1990) Phospholipid composition of some marine red algae. Phytochemistry 29:3149–3152
Dembitsky VM, Srebnik M (2002) Natural halogenated fatty acids: their analogues and derivatives. Prog Lipid Res 41:315–367
Dembitsky VM, Rozentsvet OA, Pechenkina EE (1990) Glycolipids, phospholipids and fatty-acids of brown-algae species. Phytochemistry 29:3417–3421
Dembitsky VM, Pechenkinashubina EE, Rozentsvet OA (1991) Glycolipids and fatty-acids of some seaweeds and marine grasses from the Black Sea. Phytochemistry 30:2279–2283
Deville C, Damas J, Forget P, Dandrifosse G, Peulen O (2004) Laminarin in the dietary fibre concept. J Sci Food Agric 84:1030–1038
Deville C, Gharbi M, Dandrifosse G, Peulen O (2007) Study on the effects of laminarin, a polysaccharide from seaweed, on gut characteristics. J Sci Food Agric 87:1717–1725
Dierick N, Ovyn A, De Smet S (2009) Effect of feeding intact brown seaweed Ascophyllum nodosum on some digestive parameters and on iodine content in edible tissues in pigs. J Sci Food Agric 89:584–594
Dietrich CP, Farias GGM, de Abreu LRD, Leite EL, da Silva LF, Nader HB (1995) A new approach for the characterization of polysaccharides from algae: presence of four main acidic polysaccharides in three species of the class Phaeophyceae. Plant Sci 108:143–153
Doh-ura K, Kuge T, Uomoto M, Nishizawa K, Kawasaki Y, Iha M (2007) Prophylactic effect of dietary seaweed fucoidan against enteral prion infection. Antimicrob Agents Chemother 51:2274–2277
Doi K, Tsuji K (1998) Dietary fibers: basics and critics. Asakura Shoten, Tokyo, 426 pp (in Japanese)
Douady D, Rousseau B, Caron L (1994) Fucoxanthin-chlorophyll a/c light-harvesting complexes of Laminaria saccharina: partial amino acid sequences and arrangement in thylakoid membranes. Biochemistry 33:3165–3170
Doyle JW, Roth TP, Smith RM, Li YQ, Dunn RM (1996) Effect of calcium alginate on cellular wound healing processes modeled in vitro. J Biomed Mater Res 32:561–568
Dubber D, Harder T (2008) Extracts of Ceramium rubrum, Mastocarpus stellatus and Laminaria digitata inhibit growth of marine and fish pathogenic bacteria at ecologically realistic concentrations. Aquaculture 274:196–200
Dumelod BD, Ramirez RPB, Tiangson CLP, Barrios EB, Panlasigui LN (1999) Carbohydrate availability of arroz caldo with lambda-carrageenan. Int J Food Sci Nutr 50:283–289
Durcan J, Cave R, Tyrrell L, McGovern, Stengel DB (2010) Arsenic levels in selected red, green and brown macroalgae from Western Ireland. XX Seaweed Symposium, p 121
Durig J, Bruhn T, Zurborn KH, Gutensohn K, Bruhn HD, Beress L (1997) Anticoagulant fucoidan fractions from Fucus vesiculosus induce platelet activation in vitro. Thromb Res 85:479–491
Dworjanyn SA, Pirozzi I, Liu W (2007) The effect of the addition of algae feeding stimulants to artificial diets for the sea urchin Tripneustes gratilla. Aquaculture 273:624–633
European Advisory Services (EAS) (2008) Marketing food supplements, fortified and functional foods in Europe. Legislation and practice 2008, 250 pp
EAS (2009) Regulation 1170/2009 amending Directive 2002/46/EC and Regulation 1925/2006 as regards the lists of vitamin and minerals and their forms that can be added to foods, including food supplements. http://www.eas.eu/Legislation.aspx?download=56, accessed 6 April 2010
Ellouali M, Boissonvidal C, Durand P, Jozefonvicz J (1993) Antitumor-activity of low-molecular-weight fucans extracted from brown seaweed Ascophyllum nodosum. Anticancer Res 13:2011–2019
Ennamany R, Saboureau D, Mekideche N, Creppy EE (1998) SECMA 1 (R), a mitogenic hexapeptide from Ulva algeae modulates the production of proteoglycans and glycosaminoglycans in human foreskin fibroblast. Hum Exp Toxicol 17:18–22
Enoki T, Sagawa H, Tominaga T, Nishiyama E, Komyama N, Sakai T, Yu FG, Ikai K, Kato I (2003) Drugs, foods or drinks with the use of algae-derived physiologically active substances. US Patent 0,105,029
Ergün S, Soyutürk M, Güroy B, Güroy D, Merrifield D (2008) Influence of Ulva meal on growth, feed utilization, and body composition of juvenile Nile tilapia (Oreochromis ániloticus) at two levels of dietary lipid. Aquac Int 17:355–361
EU (2008) Kommisionens forordning (EF) Nr. 629/2008, af 2. juli 2008, om ændring af forordning (EF) nr. 1881/2006 om fastsættelse af grænseværdier for bestemte forurenende stoffer i fødevarer. Den Europæiske Unions Tidende, pp 6–9 (in Danish)
European Commission (1997) Novel Food Catalogue. http://ec.europa.eu/food/food/biotechnology/novelfood/nfnetweb/mod_search/index.cfm on December 14 2010
Fan D, Hodges DM, Zhang J, Kirby CW, Ji X, Locke SJ, Critchley AT, Prithiviraj B (2011) Commercial extract of the brown seaweed Ascophyllum nodosum enhances phenolic antioxidant content of spinach (Spinacia oleracea L.) which protects Caenorhabditis elegans against oxidative and thermal stress. Food Chem 124:195–202
FAO (2008) FAO statistics. http://wwwfaoorg/figis/servlet/static?dom=root&xml=aquaculture/indexxml, accessed 2 February 2009
Farvin KHS, Holdt SL, Jacobsen C (2010) Phenolic composition and in vitro antioxidant activities of selected species of seaweeds from Danish coast. 101st American Oil Chemists Society Annual Meeting and Expo, Phoenix, Arizona. May 16–19
Faulkner DJ (2001) Marine natural products. Nat Prod Rep 18:1–49
Fernandez LE, Valiente OG, Mainardi V, Bello JL, Velez H, Rosado A (1989) Isolation and characterization of an antitumor active agar-type polysaccharide of Gracilaria dominguensis. Carbohydr Res 190:77–83
Fleurence J (1999) Seaweed proteins: biochemical, nutritional aspects and potential uses. Trends Food Sci Technol 10:25–28
Fleurence J (2004) Seaweed proteins. In: Yada RY (ed) Proteins in food processing. Woodhead Publishing, Cambridge, pp 197–213
Fleurence J, LeCoeur C, Mabeau S, Maurice M, Landrein A (1995) Comparison of different extractive procedures for proteins from the edible seaweeds Ulva rigida and Ulva rotundata. J Appl Phycol 7:577–582
Fleury N, Lahaye M (1991) Chemical and physicochemical characterization of fibers from Laminaria digitata (Kombu Breton)—a physiological approach. J Sci Food Agric 55:389–400
Flodin C, Helidoniotis F, Whitfield FB (1999) Seasonal variation in bromophenol content and bromoperoxidase activity in Ulva lactuca. Phytochemistry 51:135–138
Foster GG, Hodgson AN (1998) Consumption and apparent dry matter digestibility of six intertidal macroalgae by Turbo sarmaticus (Mollusca: Vetigastropoda: Turbinidae). Aquaculture 167:211–227
Foti MC (2007) Antioxidant properties of phenols. J Pharm Pharmacol 59:1673–1685
Francesconi KA, Edmonds JS (1996) Arsenic and marine organisms. Adv Inorg Chem 44:147–189
Freile-Pelegrín Y, Morales JL (2004) Antibacterial activity in marine algae from the coast of Yucatan, Mexico. Bot Mar 47:140–146
Freile-Pelegrín Y, Robledo D (1997) Effects of season on the agar content and chemical characteristics of Gracilaria cornea from Yucatan, Mexico. Bot Mar 40:285–290
FSAI (2010) http://www.fsai.ie/legislation/food_legislation/novel_foods/application_procedure.html, accessed July 2010
Fujiwara-Arasaki T, Mino N, Kuroda M (1984) The protein value in human nutrition of edible marine algae in Japan. Hydrobiologia 116/117:513–516
Fuller RW, Cardellina JH, Kato Y, Brinen LS, Clardy J, Snader KM, Boyd MR (1992) A pentahalogenated monoterpene from the red alga Portieria hornemannii produces a novel cytotoxicity profile against a diverse panel of human tumor cell lines. J Med Chem 35:3007–3011
Funahashi H, Imai T, Tanaka Y, Tsukamura K, Hayakawa Y, Kikumori T, Mase T, Itoh T, Nishikawa M, Hayashi H et al (1999) Wakame seaweed suppresses the proliferation of 7,12-dimethylbenz(a)anthracene-induced mammary tumors in rats. Jpn J Cancer Res 90:922–927
Funahashi H, Imai T, Mase T, Sekiya M, Yokoi K, Hayashi H, Shibata A, Hayashi T, Nishikawa M, Suda N et al (2001) Seaweed prevents breast cancer? Jpn J Cancer Res 92:483–487
Gahan DA, Lynch MB, Callan JJ, O’Sullivan JT, O’Doherty JV (2009) Performance of weanling piglets offered low-, medium- or high-lactose diets supplemented with a seaweed extract from Laminaria spp. Animal 3:24–31
Galland-Irmouli AV, Fleurence J, Lamghari R, Luçon M, Rouxel C, Barbaroux O, Bronowicki JP, Villaume C, Guéant JL (1999) Nutritional value of proteins from edible seaweed Palmaria palmata (dulse). J Nutr Biochem 10:353–359
Ganesan P, Kumar CS, Bhaskar N (2008) Antioxidant properties of methanol extract and its solvent fractions obtained from selected Indian red seaweeds. Bioresour Technol 99:2717–2723
Garbisa S, Sartor L, Biggin S, Salvato B, Benelli R, Albini A (2001) Tumor gelatinases and invasion inhibited by the green tea flavanol epigallocatechin-3-gallate. Cancer 91:822–832
Gardiner GE, Campbell AJ, O’Doherty JV, Pierce E, Lynch PB, Leonard FC, Stanton C, Ross RP, Lawlor PG (2008) Effect of Ascophyllum nodosum extract on growth performance, digestibility, carcass characteristics and selected intestinal microflora populations of grower-finisher pigs. Anim Feed Sci Technol 141:259–273
Geiszinger A, Goessler W, Pedersen SN, Francesconi KA (2001) Arsenic biotransformation by the brown macroalga Fucus serratus. Environ Toxicol Chem 20:2255–2262
Gessner F (1971) Wasserpermeabilität und Photosynthese bei marinen Algen. Bot Mar 14:29–31
Ghosh T, Chattopadhyay K, Marschall M, Karmakar P, Mandal P, Ray B (2009) Focus on antivirally active sulfated polysaccharides: from structure–activity analysis to clinical evaluation. Glycobiology 19:2–15
Ginzberg A, Cohen M, Sod-Moriah UA, Shany S, Rosenshtrauch A, Arad S (2000) Chickens fed with biomass of the red microalga Porphyridium sp. have reduced blood cholesterol level and modified fatty acid composition in egg yolk. J Appl Phycol 12:325–330
Glyantsev S, Annaev A, Savvina TV (1993) Morphological basis for selecting composition and structure of biologically active compounds based on sodium alginate for wound treatment. Byull Eksp Biol Med 115:65–67
Goemar (2010) http://www.goemar.fr, accessed 5 June 2010
Granert C, Raud J, Waage A, Lindquist L (1999) Effects of polysaccharide fucoidin on cerebrospinal fluid interleukin-1 and tumor necrosis factor alpha in pneumococcal meningitis in the rabbit. Infect Immun 67:2071–2074
Gravett RB (2000) The effect of Ascophyllum nodosum on immune function, performance, and carcass characteristics of sheep and cattle. Master’s thesis, Graduate Faculty of Texas Tech University, pp 1–86
Greger M, Malm T, Kautsky L (2007) Heavy metal transfer from composted macroalgae to crops. Eur J Agron 26:257–265
Gruenwald J (2008) http://www.entrepreneur.com/tradejournals/article/189971352.html, accessed 8 January 2010
Hammerstrom K, Dethier MN, Duggins DO (1998) Rapid phlorotannin induction and relaxation in five Washington kelps. Mar Ecol Prog Ser 165:263–305
Han JG, Syed AQ, Kwon M, Ha JH, Lee HY (2008) Antioxident, immunomodulatory and anticancer activity of fucoidan isolated from Fucus vesiculosus. J Biotechnol 136:571
Haug A, Jensen A (1954) Seasonal variation in the chemical composition of Alaria esculenta, Laminaria saccharina, Laminaria hyperborea and Laminaria digitata from Northern Norway, Norwegian Institute of Seaweed Research. Akademisk Trykningssentral, Blindern, Norway. Report 4, pp 1–14
Haug A, Larsen B (1957) Carotene content of seaweed and seaweed meal. Norwegian Institute of Seaweed Research. Akademisk Trykningssentral, Blindern, Norway. Report 16, pp 1–19
Haug A, Larsen B (1958) Brunfarging av tangmel og tangekstrakter [The formation of brown coloured substances in seaweed and seaweed extracts]. Norwegian Institute of Seaweed Research. NTH Trykk, Norway. Report 22, pp 1–18
Haugan JA, Liaaen-Jensen S (1989) Algal carotenoids.43. Improved isolation procedure for fucoxanthin. Phytochemistry 28:2797–2798
Haugan JA, Liaaen-Jensen S (1994) Algal carotenoids.54. Carotenoids of brown-algae (Phaeophyceae). Biochem Syst Ecol 22:31–41
Hawkins WW, Leonard VG (1963) Antithrombic activity of carrageenan in human blood. Can J Physiol Biochem 41:1325–1327
Hawkins WW, Yaphe W (1965) Carrageenan as a dietary constituent for rat—faecal excretion nitrogen absorption and growth. Can J Biochem 43:479–484
Hawkins WW, Leonard VG, Maxwell JE, Rastogi KS (1962) A study of the prolonged intake of small amounts of EDTA on the utilization of low dietary levels of calcium and iron by the rat. Can J Physiol Biochem 40:391
Hayashi L, Yokoya NS, Ostini S, Pereira RTL, Braga ES, Oliveira EC (2008) Nutrients removed by Kappaphycus alvarezii (Rhodophyta, Solieriaceae) in integrated cultivation with fishes in re-circulating water. Aquaculture 277:185–191
Heller L (2009) Functional foods: what they mean around the world. http://www.nutraingredients-usa.com/Industry/Functional-foods-What-they-mean-around-the-world, accessed July 2010
Hellio C, De La Broise D, Dufosse L, Le Gal Y, Bourgougnon N (2001) Inhibition of marine bacteria by extracts of macroalgae: potential use for environmentally friendly antifouling paints. Mar Environ Res 52:231–247
Hemmingson J, Falshaw R, Furneaux R, Thompson K (2006) Structure and antiviral activity of the galactofucan sulfates extracted from Undaria pinnatifida (Phaeophyta). J Appl Phycol 18:185–193
Hennequart F (2007) Seaweed applications in human health/nutrition: the example of algal extracts as functional ingredients in novel beverages. 4th International Symposium Health and Sea, Granville, France. http://www.sante-mer.com/pdf_dwnld/O17_Presentation_H_HENNEQUART.pdf, accessed 26 October 2010
Heo SJ, Jeon YJ (2009) Protective effect of fucoxanthin isolated from Sargassum siliquastrum on UV-B induced cell damage. J Photochem Photobiol B 65:101–107
Heo SJ, Park EJ, Lee KW, Jeon YJ (2005) Antioxidant activities of enzymatic extracts from brown seaweeds. Bioresour Technol 96:1613–1623
Herbreteau F, Coiffard LJM, Derrien A, De Roeck-Holtzhauer Y (1997) The fatty acid composition of five species of macroalgae. Bot Mar 40:25–27
Hoebler C, Guillon F, Darcy-Vrillon B, Vaugelade P, Lahaye M, Worthington E, Duee PH, Barry JL (2000) Supplementation of pig diet with algal fibre changes the chemical and physicochemical characteristics of digesta. J Sci Food Agric 80:1357–1364
Hoffmane R, Paper DH, Donaldson J, Alban S, Franz G (1995) Characterization of a laminarin sulfate which inhibits basic fibroblast growth-factor binding and endothelial-cell proliferation. J Cell Sci 108:3591–3598
Holdt SL (2009) Nutrient reduction in aquaculture waste by macroalgae production. PhD dissertation, University of Southern Denmark, 80 pp
Hori K, Ikegami S, Miyazawa K, Ito K (1988) Mitogenic and antineoplastic isoagglutinins from the red alga Solieria robusta. Phytochemistry 27:2063–2067
Hori K, Matsubara K, Miyazawa K (2000) Primary structures of two hemagglutinins from the marine red alga, Hypnea japonica. Biochim Biophys Acta Gen Subj 147:226–236
Hori K, Sato Y, Ito K, Fujiwara Y, Iwamoto Y, Makino H, Kawakubo A (2007) Strict specificity for high-mannose type N-glycans and primary structure of a red alga Eucheuma serra lectin. Glycobiology 17:479–491
Horn SJ (2000) Bioenergy from brown seaweeds. PhD dissertation, Department of Biotechnology, Norwegian University of Science and Technology, NTNU, Norway, 82 pp
Hosokawa M, Wanezaki S, Miyauchi K, Kunihara H, Kohno H, Kawabata J, Odashima S, Takahashi K (1999) Apoptosis-inducing effect of fucoxanthin on human leukemia cell line HIL-60. Food Sci Technol Res 5:243–246
Houghton JD (1996) Haems and bilins. In: Houghton JD, Hendry GAF (eds) Natural food colorants, 2nd edn. Blackie (Chapman and Hall), Glasgow, pp 157–196
Houston MC (2005) Nutraceuticals, vitamins, antioxidants, and minerals in the prevention and treatment of hypertension. Prog Cardiovasc Dis 47:396–449
Hu JF, Geng MY, Zhang JT, Jiang HD (2001) An in vitro study of the structure–activity relationships of sulfated polysaccharide from brown algae to its antioxidant effect. J Asian Nat Prod Res 3:353–358
Huang HL, Wang BG (2004) Antioxidant capacity and lipophilic content of seaweeds collected from the Qingdao coastline. J Agric Food Chem 52:4993–4997
Ichikawa S, Kamoshida M, Hanaoka K, Hamano M, Maitani T, Kaise T (2006) Decrease of arsenic in edible brown algae Hijikia fusiforme by the cooking process. Appl Organomet Chem 20:585–590
Ikeda K, Kitamura A, Machida H, Watanabe M, Negishi H, Hiraoka J, Nakano T (2003) Effect of Undaria pinnatifida (wakame) on the development of cerebrovascular diseases in stroke-prone spontaneously hypertensive rats. Clin Exp Pharmacol Physiol 30:44–48
Iliopoulou D, Roussis V, Pannecouque C, De Clercq E, Vagias C (2002) Halogenated sesquiterpenes from the red alga Laurencia obtusa. Tetrahedron 58:6749–6755
Indergaard M, Minsaas J (1991) Seaweed resources in Europe: uses and potential. In: Guiry MD, Blunden G (eds) Animal and human nutrition. Wiley, Chichester, pp 21–64
Iqbal M, Flick-Smith H, McCauley JW (2000) Interactions of bovine viral diarrhoea virus glycoprotein E-rns with cell surface glycosaminoglycans. J Gen Virol 81:451–459
Ishihara K, Murata M, Kaneniwa M, Saito H, Shinohara K, Maeda-Yamamoto M (1998) Inhibition of icosanoid production in MC/9 mouse mast cells by n-3 polyunsaturated fatty acids isolated from edible marine algae. Biosci Biotechnol Biochem 62:1412–1415
Ishihara K, Murata M, Kaneniwa M, Saito H, Komatsu W, Shinohara K (2000) Purification of stearidonic acid (18: 4(n-3)) and hexadecatetraenoic acid (16: 4(n-3)) from algal fatty acid with lipase and medium pressure liquid chromatography. Biosci Biotechnol Biochem 64:2454–2457
Ito K, Tsuchida Y (1972) The effect of algal polysaccharides on depressing of plasma cholesterol level in rats. Proceedings of the7th International Seaweed Symposium, pp 451–455
Je J-Y, Park PJ, Kim EK, Park JS, Yoon HD, Kim KR, Ahn CB (2009) Antioxidant activity of enzymatic extracts from the brown seaweed Undaria pinnatifida by electron spin resonance spectroscopy. LWT Food Sci Technol 42:874–878
Jenkins DJA, Wolever TMS, Leeds AR, Gassull MA, Haisman P, Dilawari J, Goff DV, Metz GL, Alberti KGMM (1978) Dietary fibers, fiber analogs, and glucose-tolerance—importance of viscosity. Br Med J 1:1392–1394
Jensen A (1956) Component sugars of some common brown algae. Norwegian Institute of Seaweed Research, Akademisk Trykningssentral, Blindern, Oslo. Report 9, pp 1–8
Jensen A (1960) Produksjon av tangmel. Norwegian Institute of Seaweed Research, N.T.H. TRYKK. Report 24, pp 1–23
Jensen A (1966) Carotenoids of Norwegian brown seaweeds and of seaweed meals. Norwegian Institute of Seaweed Research, TAPIR. Report 31, pp 1–138
Jensen A (1969a) Tocopherol content of seaweed and seaweed meal. 2. Individual, diurnal and seasonal variations in some Fucaceae. J Sci Food Agric 20:454–458
Jensen A (1969b) Tocopherol content of seaweed and seaweed meal. 3. Influence of processing and storage on content of tocopherols, carotenoids and ascorbic acid in seawood meal. J Sci Food Agric 20:622–626
Jensen A, Haug A (1956) Geographical and seasonal variation in the chemical composition of Laminaria hyperborea and Laminaria digitata from the Norwegian coast. Norwegian Institute of Seaweed Research, Akademisk Trykningssentral, Blindern, Oslo. Report 14, pp 1–8
Jensen A, Nebb H, Sæter EA (1968) The value of Norwegian seaweed meal as a mineral supplement for dairy cows. Norwegian Institute of Seaweed Research, TAPIR. Report 32, pp 1–35
Jeon Y-J, Athukorala Y, Lee J (2005) Characterization of agarose product from agar using DMSO. Algae 20:61–67
Johnson CR, Mann KH (1986) The importance of plant defence abilities to the structure of subtidal seaweed communities: the kelp Laminaria longicruris de la Pylaie survives grazing by the snail Lacuna vincta (Montagu) at high population densities. J Exp Mar Biol Ecol 97:231–267
Jones AL, Harwood JL (1992) Lipid composition of the brown algae Fucus vesiculosus and Ascophyllum nodosum. Phytochemistry 31:3397–3403
Joseph J, Niggemann B, Zaenker KS, Entschladen F (2002) The neurotransmitter γ-aminobutyric acid is an inhibitory regulator for the migration of SW 480 colon carcinoma cells. Cancer Res 62:6467–6469
Kaeffer B, Benard C, Lahaye M, Blottiere HM, Cherbut C (1999) Biological properties of ulvan, a new source of green seaweed sulfated polysaccharides, on cultured normal and cancerous colonic epithelial tells. Planta Med 65:527–531
Kalt W, McDonald JE, Donner H (2000) Anthocyanins, phenolics, and antioxidant capacity of processed lowbush blueberry products. J Food Sci 65:390–393
Kanazawa K, Ozaki Y, Hashimoto T, Das SK, Matsushita S, Hirano M, Okada T, Komoto A, Mori N, Nakatsuka M (2008) Commercial-scale preparation of biofunctional fucoxanthin from waste parts of brown sea algae Laminalia japonica. Food Sci Technol Res 14:573–582
Kang K, Park Y, Hwang HJ, Kim SH, Lee JG, Shin HC (2003) Antioxidative properties of brown algae polyphenolics and their perspectives as chemopreventive agents against vascular risk factors. Arch Pharm Res 26:286–293
Katayama S, Ohshita J, Sugaya K, Hirano M, Momose Y, Yamamura S (1998) New medicinal treatment for severe gingivostomatitis. Int J Mol Med 2:675–679
Kawakubo AL, Makino H, Ohnishi J, Hirohara H, Hori K (1997) The marine red alga Eucheuma serra J. Agardh, a high yielding source of two isolectins. J Appl Phycol 9:331–338
Kawakubo A, Makino H, Ohnishi J, Hirohara H, Hori K (1999) Occurrence of highly yielded lectins homologous within the genus Eucheuma. J Appl Phycol 11:149–156
Khan MNA, Cho JY, Lee MC, Kang JY, Park NG, Fujii H, Hong YK (2007) Isolation of two anti-inflammatory and one pro-inflammatory polyunsaturated fatty acids from the brown seaweed Undaria pinnatifida. J Agric Food Chem 55:6984–6988
Khan MNA, Suk-Choi J, Lee MC, Kim E, Nam TJ, Fujii H, Hong YK (2008) Anti-inflammatory activities of methanol extracts from various seaweed species. J Environ Biol 29:465–469
Khotimchenko SV (1991) Fatty-acid composition of 7 Sargassum species. Phytochemistry 30:2639–2641
Khotimchenko SV (2003) The fatty acid composition of glycolipids of marine macrophytes. Russ J Mar Biol 29:126–128
Khotimchenko SV (2005) Lipids from the marine alga Gracilaria verrucosa. Chem Nat Compd 41:285–288
Khotimchenko SV, Levchenko EV (1997) Lipids of the red alga Gracilaria verrucosa (Huds.) Papenf. Bot Mar 40:541–545
Khotimchenko YS, Kovalev VV, Savchenko OV, Ziganshina OA (2001) Physical–chemical properties, physiological activity, and usage of alginates, the polysaccharides of brown algae. Russ J Mar Biol 27:53–64
Kim IH, Lee JH (2008) Antimicrobial activities against methicillin-resistant Staphylococcus aureus from macroalgae. J Ind Eng Chem 14:568–572
Kim MK, JP D, Thomas JC, Giraud G (1996) Seasonal variations of triacylglycerols and fatty acids in Fucus serratus. Phytochemistry 43:49–55
Kimura Y, Watanabe K, Okuda H (1996) Effects of soluble sodium alginate on cholesterol excretion and glucose tolerance in rats. J Ethnopharmacol 54:47–54
Kindness G, Williamson FB, Long WF (1979) Effect of polyanetholesulphonic acid and xylan sulphate on antithrombin III activity. Biochem Biophys Res Commun 13:1062–1068
Kiriyama S, Okazaki Y, Yoshida A (1969) Hypocholesterolemic effect of polysaccharides and polysaccharide-rich foodstuffs in cholesterol-fed rats. J Nutr 97:382–388
Klinkenberg-Knol EC, Festen HP, Meuwissen SG (1995) Pharmacological management of gastro-oesophageal reflux disease. Drugs 49:495–710
Knott MG, Mkwananzi H, Arendse CE, Hendricks DT, Bolton JJ, Beukes DR (2005) Plocoralides A-C, polyhalogenated monoterpenes from the marine alga Plocamium corallorhiza. Phytochemistry 66:1108–1112
Korver O, Kühn MC, Richardson DP (2008) The functional food dossier; Building solid health claims. Practical industrial guide. Foodlink Forum, Bennekom, the Netherlands, 185 pp
Kotake-Nara E, Kushiro M, Zhang H, Sugawara T, Miyashita K, Nagao A (2001) Carotenoids affect proliferation of human prostate cancer cells. J Nutr 131:3303–3306
Kumar CS, Ganesan P, Bhaskar N (2008a) In vitro antioxidant activities of three selected brown seaweeds of India. Food Chem 107:707–713
Kumar CS, Ganesan P, Suresh PV, Bhaskar N (2008b) Seaweeds as a source of nutritionally beneficial compounds—a review. J Food Sci Technol 45:1–13
Kumar KS, Ganesane K, Rao PVS (2008c) Antioxidant potential of solvent extracts of Kappaphycus alvarezii (Doty) Doty—an edible seaweed. Food Chem 107:289–295
Kuznetsova TA, Krylova NV, Besednova NN, Vasiléva VN, Zviagintseva TN, Krashevskii SV, Eliakova LA (1994) The effect of translam on the natural resistance indices of the irradiated organism. Radiats Biol Radioecol 34:236–239
Kylin H (1913) Zur Biochemie der Meeresalgen. Hoppe-Seyler’s Z Physiol Chem 83:171–197
Lahaye M (1991) Marine-algae as sources of fibers—determination of soluble and insoluble dietary fiber contents in some sea vegetables. J Sci Food Agric 54:587–594
Lahaye M (1998) NMR spectroscopic characterisation of oligosaccharides from two Ulva rigida ulvan samples (Ulvales, Chlorophyta) degraded by a lyase. Carbohydr Res 314:1–12
Lahaye M, Kaeffer B (1997) Seaweed dietary fibres: structure, physico-chemical and biological properties relevant to intestinal physiology. Sci Aliment 17:563–584
Lahaye M, Ray B (1996) Cell-wall polysaccharides from the marine green alga Ulva rigida (Ulvales, Chlorophyta)—NMR analysis of ulvan oligosaccharides. Carbohydr Res 283:161–173
Lahaye M, Robic A (2007) Structure and functional properties of ulvan, a polysaccharide from green seaweeds. Biomacromolecules 8:1765–1774
Lahaye M, Michel C, Barry JL (1993) Chemical, physicochemical and in vitro fermentation characteristics of dietary-fibers from Palmaria palmata (L.) Kuntze. Food Chem 47:29–36
Lahaye M, Inizan F, Vigouroux J (1998) NMR analysis of the chemical structure of ulvan and of ulvan–boron complex formation. Carbohydr Polym 36:239–249
Lahaye M, Rondeau-Mouro C, Deniaud E, Buleon A (2003) Solid-state 13C NMR spectroscopy studies of xylans in the cell wall of Palmaria palmata (L. Kuntze, Rhodophyta). Carbohydr Res 338:1559–1569
Lam C, Stang A, Harder T (2008) Planktonic bacteria and fungi are selectively eliminated by exposure to marine macroalgae in close proximity. FEMS Microbiol Ecol 63:283–291
Lamare MD, Wing SR (2001) Calorific content of New Zealand marine macrophytes. NZ J Mar Freshwat Res 35:335–341
Lamela M, Anca J, Villar R, Otero J, Calleja JM (1989) Hypoglycemic activity of several seaweed extracts. J Ethnopharmacol 27:35–43
Larsen B, Haug A (1958) The influence of habitat on the chemical composition of Ascophyllum nodosum (L.) Le Jol.-reprint of report 20. Norwegian Institute of Seaweed Research. Trondheim, Norway, pp 29–38
Lasky LA (1995) Selectin–carbohydrate interactions and the initiation of the inflammatory response. Annu Rev Biochem 64:113–139
Le Tutour B (1990) Antioxidative activities of algal extracts, synergistic effect with vitamin E. Phytochemistry 29:3759–3765
Le Tutour B, Benslimane F, MPe G, Saadan B, Quemeneur F (1998) Antioxidant and pro-oxidant activities of the brown algae, Laminaria digitata, Himanthalia elongata, Fucus vesiculosus, Fucus serratus and Ascophyllum nodosum. J Appl Phycol 10:121–129
Lee JB, Hayashi K, Hashimoto M, Nakano T, Hayashi T (2004) Novel antiviral fucoidan from sporophyll of Undaria pinnatifida (Mekabu). Chem Pharm Bull 52:1091–1094
Li F, Tian TC, Shi YC (1995) Study on anti virus effect of fucoidan in vitro. J Norman Bethune Univ Med Sci 21:255–257
Li N, Zhang QB, Song JM (2005) Toxicological evaluation of fucoidan extracted from Laminaria japonica in Wistar rats. Food Chem Toxicol 43:421–426
Li K, Li XM, Ji NY, Wang BG (2007) Natural bromophenols from the marine red alga Polysiphonia urceolata (Rhodomelaceae): structural elucidation and DPPH radical-scavenging activity. Bioorg Med Chem 15:6627–6631
Li B, Lu F, Wei XJ, Zhao RX (2008a) Fucoidan: structure and bioactivity. Molecules 13:1671–1695
Li Y, Qian ZJ, Le QT, Kim MM, Kim SK (2008b) Bioactive phloroglucinol derivatives isolated from an edible marine brown alga, Ecklonia cava. J Biotechnol 136:578
Li Y, Qian ZJ, Ryu B, Lee SH, Kim MM, Kim SK (2009) Chemical components and its antioxidant properties in vitro: an edible marine brown alga, Ecklonia cava. Bioorg Med Chem 17:1963–1973
Liao WR, Lin JY, Shieh WY, Jeng WL, Huang R (2003) Antibiotic activity of lectins from marine algae against marine vibrios. J Ind Microbiol Biotechnol 30:433–439
Lim SN, Cheung PCK, Ooi VEC, Ang PO (2002) Evaluation of antioxidative activity of extracts from a brown seaweed, Sargassum siliquastrum. J Agric Food Chem 50:3862–3866
Liu RM, Bignon J, Haroun-Bouhedja F, Bittoun P, Vassy J, Fermandjian S, Wdzieczak-Bakala J, Boisson-Vidal C (2005) Inhibitory effect of fucoidan on the adhesion of adenocarcinoma cells to fibronectin. Anticancer Res 25:2129–2133
Lobban CS, Harrison PJ (1994) Seaweed ecology and physiology. Cambridge University Press, Cambridge, 384 pp
Lourenço R, Camilo ME (2002) Taurine: a conditionally essential amino acid in humans? An overview in health and disease. Nutr Hosp 17:262–270
Lüder UH, Clayton MN (2004) Induction of phlorotannins in the brown macroalga Ecklonia radiata (Laminariales, Phaeophyta) in response to simulated herbivory—the first microscopic study. Planta 218:928–937
Lüning (2008) Integrated macroalgae-oyster aquaculture on a north sea island: seasonal productivity of the brown alga Laminaria saccharina and the red algae Palmaria palmata; Solieria chordalis, Gracilaria vermiculophylla, and the use of these seaweeds in human nutrition or as raw material for the cosmetics industry. 11th International Conference on Applied Phycology, Galway, Ireland. June 22–27
Luescher-Mattli M (2003) Algae, a possible source for new drugs in the treatment of HIV and other viral diseases. Curr Med Chem 2:219–225
Mabeau S, Fleurence J (1993) Seaweed in food products: biochemical and nutritional aspects. Trends Food Sci Technol 4:103–107
MacArtain P, Gill CIR, Brooks M, Campbell R, Rowland IR (2007) Nutritional value of edible seaweeds. Nutr Rev 65:535–543
Maeda H, Hosokawa M, Sashima T, Funayama K, Miyashita K (2005) Fucoxanthin from edible seaweed, Undaria pinnatifida, shows antiobesity effect through UCP1 expression in white adipose tissues. Biochem Biophys Res Commun 332:392–397
Maeda H, Tsukui T, Sashima T, Hosokawa M, Miyashita K (2008a) Seaweed carotenoid, fucoxanthin, as a multi-functional nutrient. Asia Pac J Clin Nutr 17:196–199
Maeda H, Hosokawa M, Sashima T, Miyashita K (2008b) Antiobesity effect of fucoxanthin from edible seaweeds and its multibiological functions. ACS Symp Ser 993:376–388
Mai K, Mercer JP, Donlon J (1994) Comparative studies on the nutrition of two species of abalone, Haliotis tuberculata L. and Haliotis discus Hannai Ino. II. Amino acid composition of abalone and six species of macroalgae with an assessment of their nutritional-value. Aquaculture 128:115–130
Maliakal PP, Coville PF, Wanwimolruk S (2001) Tea consumption modulates hepatic drug metabolizing enzymes in Wistar rats. J Pharm Pharmacol 53:569–577
Mandal P, Mateu CG, Chattopadhyay K, Pujol CA, Damonte EB, Ray B (2007) Structural features and antiviral activity of sulphated fucans from the brown seaweed Cystoseira indica. Antiviral Chem Chemother 18:153–162
Marais MF, Joseleau JP (2001) A fucoidan fraction from Ascophyllum nodosum. Carbohydr Res 336:155–159
Marinho-Soriano E, Fonseca PC, Carneiro MAA, Moreira WSC (2006) Seasonal variation in the chemical composition of two tropical seaweeds. Bioresour Technol 97:2402–2406
Marinova (2010) Nutraceuticals from Maritech extracts. http://www.marinova.com.au/Nutraceuticals.html on December 13 2010
Marsham S, Scott GW, Tobin ML (2007) Comparison of nutritive chemistry of a range of temperate seaweeds. Food Chem 100:1331–1336
Martino AD, Douady D, Quinet-Szely M, Rousseau B, Crépineau F, Apt K, Caron L (2004) The light-harvesting antenna of brown algae. Highly homologous proteins encoded by a multigene family. Eur J Biochem 267:5540–5549
Maruyama H, Tamauchi H, Hashimoto M, Nakano T (2003) Antitumor activity and immune response of Mekabu fucoidan extracted from sporophyll of Undaria pinnatifida. In Vivo 17:245–249
Maruyama H, Tamauchi H, Iizuka M, Nakano T (2006) The role of NK cells in antitumor activity of dietary fucoidan from Undoria pinnotifida sporophylls (Mekabu). Planta Med 72:1415–1417
Maruyama H, Tanaka M, Hashimoto M, Inoue M, Sasahara T (2007) The suppressive effect of Mekabu fucoidan on an attachment of Cryptosporidium parvum oocysts to the intestinal epithelial cells in neonatal mice. Life Sci 80:775–781
Matsubara K, Sumi H, Hori K (1996) Platelet aggregation is inhibited by phycolectins. Experientia 52:540–543
Matsubara K, Xue C, Zhao X, Mori M, Sugawara T, Hirata T (2005) Effects of middle molecular weight fucoidans on in vitro and ex vivo angiogenesis of endothelial cells. Int J Mol Med 15:695–699
Mayer AMS, Hamann MT (2004) Marine pharmacology in 2000: marine compounds with antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiplatelet, antituberculosis, and antiviral activities; affecting the cardiovascular, immune, and nervous systems and other miscellaneous mechanisms of action. Mar Biotechnol 6:37–52
Mayer AMS, Rodríguez AD, Berlinck RGS, Hamann MT (2007) Marine pharmacology in 2003–4: marine compounds with anthelmintic antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiplatelet, antiprotozoal, antituberculosis, and antiviral activities; affecting the cardiovascular, immune and nervous systems, and other miscellaneous mechanisms of action. Comp Biochem Physiol C 145:553–581
McHugh DJ (1987) Production and utilization of products from commercial seaweeds. FAO Fisheries Technical Paper No. 288, pp 1–189
McHugh DJ (2003) A guide to the seaweed industry. FAO Fish Tech Pap 441, Rome, Italy, 105 pp
Mehta SK, Gaur JP (2005) Use of algae for removing heavy metal ions from wastewater: progress and prospects. Crit Rev Biotechnol 25:113–152
Miao HQ, Elkin M, Aingorn E, Ishai-Michaeli R, Stein CA, Vlodavsky I (1999) Inhibition of heparanase activity and tumor metastasis by laminarin sulfate and synthetic phosphorothioate oligodeoxynucleotides. Int J Cancer 83:424–431
Michel C, Macfarlane GT (1996) Digestive fates of soluble polysaccharides from marine macroalgae: involvement of the colonic microflora and physiological consequences for the host. J Appl Bacteriol 80:349–369
Michel C, Lahaye M, Bonnet C, Mabeau S, Barry JL (1996) In vitro fermentation by human faecal bacteria of total and purified dietary fibres from brown seaweeds. Br J Nutr 75:263–280
Miki W (1991) Biological functions and activities of animal carotenoids. Pure Appl Chem 63:141–146
Militante JD, Lombardini JB (2002) Treatment of hypertension with oral taurine: experimental and clinical studies. Amino Acids 23:381–393
Miraglio AM (2009) Phospholipids: structure plus functionality. http://www foodproductdesign com/articles/2006/03/phospholipids-structure-plus-functionality.aspx, accessed 5 June 2010
Mishra VK, Temelli F, Ooraikul B, Shacklock PF, Craigie JS (1993) Lipids of the red alga, Palmaria palmata. Bot Mar 36:169–174
Miyashita H, Hosokawa M (2008) Beneficial health effects of seaweed carotenoid, fucoxanthin. In: Barrow C, Shahidi F (eds) Marine nutraceuticals and functional foods. CRC, Boca Raton, pp 297–320
Mochizuki H, Takido J, Oda H, Yokogoshi H (1999) Improving effect of dietary taurine on marked hypercholesterolemia induced by a high-cholesterol diet in streptozotocin-induced diabetic rats. Biosci Biotechnol Biochem 63:1984–1987
Morgan K, Wright J, Simpson F (1980) Review of chemical constituents of the red alga Palmaria palmata (dulse). Econ Bot 34:27–50
Mori J, Matsunaga T, Takahashi S, Hasegawa C, Saito H (2003) Inhibitory activity on lipid peroxidation of extracts from marine brown alga. Phytother Res 17:549–551
Mori T, O’Keefe BR, Sowder RC, Bringans S, Gardella R, Berg S, Cochran P, Turpin JA, Buckheit RW, McMahon JB et al (2005) Isolation and characterization of griffithsin, a novel HIV-inactivating protein, from the red alga Griffithsia sp. J Biol Chem 280:9345–9353
Morrissey J, Kraan S, Guiry MD (2001) A guide to commercially important seaweeds on the Irish coast. Bord Iascaigh Mhara, Dublin, 66 pp
Morse ANC (1991) GABA-mimetic peptides from marine algae and cyanobacteria as potential diagnostic and therapeutic agents. In: Thompson MF, Sarojini R, Nagabhushanam R (eds) Bioactive compounds from marine organisms. Oxford & IBH Publishing, New Delhi, pp 167–172
Mouradi-Givernaud A, Givernaud T, Morvan H, Cosson J (1992) Agar from Gelidium latifolium (Rhodophyceae, Gelidiales)—biochemical composition and seasonal variations. Bot Mar 35:153–159
Mouritsen OG (2009) Tang-grøntsager fra havet. Nyt Nordisk Forlag, Arnold Busck, Copenhagen, 284 pp (in Danish)
Munda IM (1977) Differences in amino acid composition of estuarine and marine fucoids. Aquat Bot 3:273–280
Murata M, Nakazoe J (2001) Production and use of marine algae in Japan. Jpn Agr Res Q 35:281–290
Murata M, Ishihara K, Saito H (1999) Hepatic fatty acid oxidation enzyme activities are stimulated in rats fed the brown seaweed, Undaria pinnatifida (wakame). J Nutr 129:146–151
Murata M, Sano Y, Ishihara K, Uchida M (2002) Dietary fish oil and Undaria pinnatifida (wakame) synergistically decrease rat serum and liver triacylglycerol. J Nutr 132:742–747
Müssig K (2009) Iodine-induced toxic effects due to seaweed consumption. In: Preedy VR, Burrow GN, Watson R (eds) Comprehensive handbook of iodine. Elsevier, New York, pp 897–908
Mustafa MG, Wakamatsu S, Takeda TA, Umino T, Nakagawa H (1995) Effects of algae meal as feed additive on growth, feed efficiency, and body composition in red sea bream. Fish Sci 61:25–28
Nabors LOB (2004) Alternative sweeteners. Agro Food Industry Hi-Tech 15:39–41
Nagaoka M, Shibata H, Kimura-Takagi I, Hashimoto S, Kimura K, Makino T, Aiyama R, Ueyama S, Yokokura T (1999) Structural study of fucoidan from Cladosiphon okamuranus Tokida. Glycoconj J 16:19–26
Nagaoka M, Shibata H, Kimura I, Hashimoto S (2003) Oligosaccharide derivatives and process for producing the same. United States Patent No. 6,645,940
Nagayama K, Shibata T, Fujimoto K, Honjo T, Nakamura T (2003) Algicidal effect of phlorotannins from the brown alga Ecklonia kurome on red tide microalgae. Aquaculture 218:601–611
Nakagawa H, Kasahara S (1986) Effect of Ulva meal supplement to diet on the lipid metabolism of red sea bream. Bull Jpn Soc Sci Fish 52:1887–1893
Nakagawa H, Umino T, Tasaka Y (1997) Usefulness of Ascophyllum meal as a feed additive for red sea bream, Pagrus major. Aquaculture 151:275–281
Nakamura T, Nagayama K, Uchida K, Tanaka R (1996) Antioxidant activity of phlorotannins isolated from the brown alga Eisenia bicyclis. Fish Sci 62:923–926
Nakazawa Y, Sashima T, Hosokawa M, Miyashita K (2009) Comparative evaluation of growth inhibitory effect of stereoisomers of fucoxanthin in human cancer cell lines. J Funct Foods 1:88–97
Narayan B, Miyashita K, Hosakawa M (2006) Physiological effects of eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA)—a review. Food Rev Int 22:291–307
Nasu T, Fukuda Y, Nagahira K, Kawashima H, Noguchi C, Nakanishi T (1997) Fucoidin, a potent inhibitor of L-selectin function, reduces contact hypersensitivity reaction in mice. Immunol Lett 59:47–51
Nishide E, Uchida H (2003) Effects of Ulva powder on the ingestion and excretion of cholesterol in rats. In: Chapman ARO, Anderson RJ, Vreeland VJ, Davison IR (eds) Proceedings of the 17th International Seaweed Symposium. Oxford University Press, Oxford, pp 165–168
Nishide E, Anzai H, Uchida N (1993) Effects of alginates on the ingestion and excretion of cholesterol in the rat. J Appl Phycol 5:207–211
Nishino T, Nishioka C, Ura H, Nagumo T (1994) Isolation and partial characterization of a novel amino sugar-containing fucan sulfate from commercial Fucus vesiculosus fucoidan. Carbohydr Res 255:213–224
Nisizawa K (2002) Seaweed Kaiso, bountiful harvest from the seas. Sustenance for health and well-being by preventing common life-style related diseases. Kochi University, Kochi, 106 pp
Noa M, Mas R, Carbajal D, Valdes S (2000) Effect of D-002 on acetic acid-induced colitis in rats at single and repeated doses. Pharmacol Res 41:391–395
Noda H (1993) Health benefits and nutritional properties of nori. J Appl Phycol 5:255–258
Okuzumi J, Takahashi T, Yamane T, Kitao Y, Inagake M, Ohya K, Nishino H, Tanaka Y (1993) Inhibitory effects of fucoxanthin, a natural carotenoid, on N-ethyl-N′-nitro-N-nitrosoguanidine-induced mouse duodenal carcinogenesis. Cancer Lett 68:159–168
Omata M, Matsui N, Inomata N, Ohno T (1997) Protective effects of polysaccharide fucoidin on myocardial ischemia–reperfusion injury in rats. J Cardiovasc Pharmacol 30:717–724
Ortiz J, Romero N, Robert P, Araya J, Lopez-Hernández J, Bozzo C, Navarrete E, Osorio A, Rios A (2006) Dietary fiber, amino acid, fatty acid and tocopherol contents of the edible seaweeds Ulva lactuca and Durvillaea antarctica. Food Chem 99:98–104
Ostergaard C, Yieng-Kow RV, Benfield T, Frimodt-Moller N, Espersen F, Lundgren JD (2000) Inhibition of leukocyte entry into the brain by the selectin blocker fucoidin decreases interleukin-1 (IL-1) levels but increases IL-8 levels in cerebrospinal fluid during experimental pneumococcal meningitis in rabbits. Infect Immun 68:3153–3157
Panlasigui LN, Baello OQ, Dimatangal JM, Dumelod BD (2003) Blood cholesterol and lipid-lowering effects of carrageenan on human volunteers. Asia Pac J Clin Nutr 12:209–214
Paradossi G, Cavalieri F, Chiessi E (2002) A conformational study on the algal polysaccharide ulvan. Macromolecules 35:6404–6411
Park PJ, Heo SJ, Park EJ, Kim SK, Byun HG, Jeon BT, Jeon YJ (2005) Reactive oxygen scavenging effect of enzymatic extracts from Sargassum thunbergii. J Agric Food Chem 53:6666–6672
Parsons AF (1996) Recent developments in kainoid amino acid chemistry. Tetrahedron 52:4149–4174
Pavia H, Åberg P (1996) Spatial variation in polyphenolic content of Ascophyllum nodosum (Fucales, Phaeophyta). Hydrobiologia 326/327:199–203
Pavia H, Cervin G, Lindgren A, Åberg P (1997) Effects of UV-B radiation and simulated herbivory on phlorotannins in the brown alga Ascophyllum nodosum. Mar Ecol Prog Ser 157:139–146
Pedersen M, Collen J, Abrahamsson K, Ekdahl A (1996) Production of halocarbons from seaweeds: an oxidative stress reaction? Sci Mar 60:257–263
Percival E, McDowell RH (1990) Algal polysaccharides. In: Dey PM (ed) Methods in plant biochemistry, volume 2: carbohydrates. Academic, London, pp 523–547
Perelló G, Martí-Cid R, Llobet JM, Domingo JL (2008) Effects of various cooking processes on the concentrations of arsenic, cadmium, mercury, and lead in foods. J Agric Food Chem 56:11262–11269
Plaza M, Cifuentes A, Ibáñez E (2008) In the search of new functional food ingredients from algae. Trends Food Sci Technol 19:31–39
Plouguerne E, Hellio C, Deslandes E, Veron B, Stiger-Pouvreau V (2008) Anti-microfouling activities in extracts of two invasive algae: Grateloupia turuturu and Sargassum muticum. Bot Mar 51:202–208
Population Council (2008) Trial shows anti-HIV microbicide is safe, but does not prove it effective. http://www.popcouncil.org/mediacenter/newsreleases/2008_CarraguardResults.asp on December 12 2010
Qi HM, Zhang QB, Zhao TT, Chen R, Zhang H, Niu XZ, Li Z (2005a) Antioxidant activity of different sulfate content derivatives of polysaccharide extracted from Ulva pertusa (Chlorophyta) in vitro. Int J Biol Macromol 37:195–199
Qi HM, Zhao TT, Zhang QB, Li Z, Zhao ZQ, Xing R (2005b) Antioxidant activity of different molecular weight sulfated polysaccharides from Ulva pertusa Kjellm (Chlorophyta). J Appl Phycol 17:527–534
Ragan MA, Jensen A (1978) Quantitative studies on brown algal phenols. II. Seasonal variation in polyphenol content of Ascophyllum nodosum (L.) Le Jol. and Fucus vesiculosus (L.). J Exp Mar Biol Ecol 34:245–258
Ramsey UP, Bird CJ, Shacklock PF, Laycock MV, Wright JL (1994) Kainic acid and 1′-hydroxykainic acid from Palmariales. Nat Toxins 2:286–292
Rasmussen RS, Morrissey MT (2007) Marine biotechnology for production of food ingredients. In: Taylor SL (ed) Advances in food and nutrition research, vol 52. Elsevier, New York, pp 237–292
Reckitt and Colman Products Ltd (1999) Formulations for the treatment of gastro-oesophageal reflux. US Patent No. 6,348,502
Reilly P, O’Doherty JV, Pierce KM, Callan JJ, O’Sullivan JT, Sweeney T (2008) The effects of seaweed extract inclusion on gut morphology, selected intestinal microbiota, nutrient digestibility, volatile fatty acid concentrations and the immune status of the weaned pig. Animal 2:1465–1473
Renn DW, Noda H, Amano H, Nishino T, Nishizana K (1994a) Antihypertensive and antihyperlipidemic effects of funoran. Fish Sci 60:423–427
Renn DW, Noda H, Amano H, Nishino T, Nishizana K (1994b) Study on hypertensive and antihyperlipidemic effect of marine algae. Fish Sci 60:83–88
Reynolds JEF, Prasad AB (1982) Martindale the extra pharmacopoeia. 28th Pharmaceutical Press, London, 735 pp
Riccardi BA, Fahrenbach MJ (1965) Effect of guar gum and pectin on serum and liver lipids of cholesterol-fed rats. Proc Soc Exp Biol Med 124:749–752
Rioux LE, Turgeon SL, Beaulieu M (2007) Characterization of polysaccharides extracted from brown seaweeds. Carbohydr Polym 69:530–537
Robertson WV, Schwartz B (1953) Ascorbic acid and the formation of collagen. J Biol Chem 201:689–696
Robledo D, Freile-Pelegrín Y (1997) Chemical and mineral composition of six potentially edible seaweed species of Yucatan. Bot Mar 40:301–306
Rocha de Souza M, Marques C, Guerra Dore C, Ferreira da Silva F, Oliveira Rocha H, Leite E (2007) Antioxidant activities of sulfated polysaccharides from brown and red seaweeds. J Appl Phycol 19:153–160
Rodrigueza MRC, Montaño MNE (2007) Bioremediation potential of three carrageenophytes cultivated in tanks with seawater from fish farms. J Appl Phycol 19:755–762
Ronnberg O, Adjers K, Ruokolahti C, Bondestam M (1990) Fucus vesiculosus as an indicator of heavy metal availability in a fish farm recipient in the northern Baltic Sea. Mar Pollut Bull 21:388–392
Rorrer GL, Yoo HD, Huang YM, Hayden C, Gerwick WH (1997) Production of hydroxy fatty acids by cell suspension cultures of the marine brown alga Laminaria saccharina. Phytochemistry 46:871–877
Rupérez P (2002) Mineral content of edible marine seaweeds. Food Chem 79:23–26
Rupérez P, Saura-Calixto F (2001) Dietary fibre and physicochemical properties of edible Spanish seaweeds. Eur Food Res Technol 212:349–354
Ruperez P, Ahrazem O, Leal JA (2002) Potential antioxidant capacity of sulfated polysaccharides from the edible marine brown seaweed Fucus vesiculosus. J Agric Food Chem 50:840–845
Sailler B, Glombitza KW (1999) Phlorethols and fucophlorethols from the brown alga Cystophora retroflexa. Phytochemistry 50:869–881
Sampath-Wiley P, Neefus CD, Jahnke LS (2008) Seasonal effects of sun exposure and emersion on intertidal seaweed physiology: fluctuations in antioxidant contents, photosynthetic pigments and photosynthetic efficiency in the red alga Porphyra umbilicalis Kützing (Rhodophyta, Bangiales). J Exp Mar Biol Ecol 361:83–91
Sanchez-Machado DI, Lopez-Cervantes J, Lopez-Hernandez J, Paseiro-Losada P (2004a) Fatty acids, total lipid, protein and ash contents of processed edible seaweeds. Food Chem 85:439–444
Sanchez-Machado DI, Lopez-Hernandez J, Paseiro-Losada P, Lopez-Cervantes J (2004b) An HPLC method for the quantification of sterols in edible seaweeds. Biomed Chromatogr 18:183–190
Sandau E, Sandau P, Pulz O, Zimmermann M (1996) Heavy metal sorption by marine algae and algae by-products. Acta Biotechnol 16:103–119
Sathivel A, Raghavendran HRB, Srinivasan P, Devaki T (2008) Anti-peroxidative and anti-hyperlipidemic nature of Ulva lactuca crude polysaccharide on d-galactosamine induced hepatitis in rats. Food Chem Toxicol 46:3262–3267
Sato M, Hosokawa T, Yamaguchi T, Nakano T, Muramoto K, Kahara T, Funayama K, Kobayashi A, Nakano T (2002) Angiotensin I-converting enzyme inhibitory peptides derived from wakame (Undaria pinnatifida) and their antihypertensive effect in spontaneously hypertensive rats. J Agric Food Chem 50:6245–6252
Satoh K, Nakagawa H, Kasahara S (1987) Effect of Ulva meal supplementation on disease resistance of red sea bream. Nippon Suisan Gakkaishi 53:1115–1120
Savitskaya IM (1986) Trial of a local hemostatic with alginate base. Klin Khirurgiya 3:39–40
Schaeffer DJ, Krylov VS (2000) Anti-HIV activity of extracts and compounds from algae and cyanobacteria. Ecotoxicol Environ Saf 45:208–227
Segal HC, Hunt BJ, Gilding K (1998) The effects of alginate and non-alginate wound dressings on blood coagulation and platelet activation. J Biomater Appl 12:249–257
Sekar S, Chandramohan M (2008) Phycobiliproteins as a commodity: trends in applied research, patents and commercialization. J Appl Phycol 20:113–136
Sen AK, Das AK, Banerji N, Siddhanta AK, Mody KH, Ramavat BK, Chauhan VD, Vedasiromoni JR, Ganguly DK (1994) A new sulfated polysaccharide with potent blood anti-coagulant activity from the red seaweed Grateloupia Indica. Int J Biol Macromol 16:279–280
Shanmugam M, Mody KH (2000) Heparinoid-active sulphated polysaccharides from marine algae as potential blood anticoagulant agents. Curr Sci 79:1672–1683
Sheth BB (1967) Alginate containing antacid compositions. US Patent No. 3,326,755
Shinagawa A, Shiomi K, Yamanaka H, Kicuchi T (1983) Selective determination of inorganic arsenic(III), (V) and organic arsenic in marine organisms. Bull Jpn Soc Sci Fish 49:75–78
Shiu CT, Lee TM (2005) Ultraviolet-B-induced oxidative stress and responses of the ascorbate–glutathione cycle in a marine macroalga Ulva fasciata. J Exp Bot 56:2851–2865
Sieburth JM, Conover JT (1965) Sargassum tannin, an antibiotic which retards fouling. Nature 208:52–53
Simpson FJ, Shacklock PF (1979) The cultivation of Chondrus crispus. Effect of temperature on growth and carrageenan production. Bot Mar 22:295–298
Siriwardhana N, Lee KW, Kim SH, Ha JW, Jeon YJ (2003) Antioxidant activity of Hizikia fusiformis on reactive oxygen species scavenging and lipid peroxidation inhibition. Food Sci Technol Int 9:339–346
Siriwardhana N, Lee KW, Kim SH, Ha JH, Park GT, Jeon YJ (2004) Lipid peroxidation inhibitory effects of Hizikia fusiformis methanolic extract on fish oil and linoleic acid. Food Sci Technol Int 10:65–72
Skoler-Karpoff S, Ramjee G, Ahmed K, Altini L, Plagianos MG, Friedland B, Govender S, Kock AD, Cassim N, Palanee T et al (2008) Efficacy of Carraguard for prevention of HIV infection in women in South Africa: a randomised, double-blind, placebo-controlled trial. Lancet 372:1977–1987
Slezka IE, Miroshichenko VA, Vostrikova OG, Ziganshina OA (1998) Applications of bioactive compounds from marine organisms in atherosclerosis prophylaxis in children, new biomedical technologies using bioactive additives. Proceedings of All Russian Conference. IMKVL Siberian Branch, Ross Akad Med Nauk, Vladivostok, pp 90–94
Smit AJ (2004) Medicinal and pharmaceutical uses of seaweed natural products: a review. J Appl Phycol 16:245–262
Smith DG, Young EG (1953) On the nitrogenous constituents of Fucus vesiculosus. J Biol Chem 205:849–858
Soler-Vila A, Coughlan S, Guiry MD, Kraan S (2009) The red alga Porphyra dioica as a fish-feed ingredient for rainbow trout (Oncorhynchus mykiss): effects on growth, feed efficiency and carcass composition. J Appl Phycol 21:617–624
Sørensen HO (2009) Marine phospholipids. Int Aqua Feed 12:14–15
Sæter EA, Jensen A (1957) Forsøk med tangmel som tilskuddsfôr til sau [Experiments with seaweed meal as supplement to rations for sheep—summary in English]. Norwegian Institute of Seaweed Research. Akademisk Trykningssentral, Blinder, Oslo. Report 17, pp 1–30
Spieler R (2002) Seaweed compound’s anti-HIV efficacy will be tested in southern Africa. Lancet 359:1675
Stahl W, Sies H (2005) Bioactivity and protective effects of natural carotenoids. BBA-Mol Basis Disease 1740:101–107
Stahl W, Ale-Agha N, Polidori MC (2002) Non-antioxidant properties of carotenoids. Biol Chem 383:553–558
Suetsuna K (1998a) Purification and identification of angiotensin I converting enzyme inhibitors from the red alga Porphyra yezoensis. J Mar Biotechnol 6:163–167
Suetsuna K (1998b) Separation and identification of angiotensin I-converting enzyme inhibitory peptides from peptic digest of Hizikia fusiformis protein. Nippon Suisan Gakkaishi 64:862–866
Suetsuna K, Maekawa K, Chen JR (2004) Antihypertensive effects of Undaria pinnatifida (wakame) peptide on blood pressure in spontaneously hypertensive rats. J Nutr Biochem 15:267–272
Sugahara T, Ohama Y, Fukuda A, Hayashi M, Kawakubo A, Kato K (2001) The cytotoxic effect of Eucheuma serra agglutinin (ESA) on cancer cells and its application to molecular probe for drug delivery system using lipid vesicles. Cytotechnology 36:93–99
Sugawara T, Baskaran V, Tsuzuki W, Nagao A (2002) Brown algae fucoxanthin is hydrolyzed to fucoxanthinol during absorption by Caco-2 human intestinal cells and mice. J Nutr 132:946–951
Suzuki M, Daitoh M, Vairappan CS, Abe T, Masuda M (2002) Brominated metabolites from an Okinawan Laurencia intricate. Phytochemistry 60:861–867
Swinyard EA, Pathak MA (1985) Surface-acting drugs. In: Gilman AG, Goodman LS, Rall TW (eds) The pharmacological basis of therapeutics, 7th edn. Macmillan, New York, pp 946–958
Takahashi N, Ojika M, Dogasaki C, Nisizawa M, Fukuoka H, Sahara H, Sato N, Mori M, Kikuchi K (2000) Substance isolated from the kelp rhizoid identified as l-tryptophan shows high inhibition of breast cancer. Gan To Kagaku Ryoho 27:251–255
Tasende MG (2000) Fatty acid and sterol composition of gametophytes and sporophytes of Chondrus crispus (Gigartinaceae, Rhodophyta). Sci Mar 64:421–426
Terada A, Hara H, Mitsuoka T (1995) Effect of dietary alginate on the faecal microbiota and faecal metabolic activity in humans. Microb Ecol Health Dis 8:259–266
Tititudorancea (2009) http://www tititudorancea com/z/nutrition_facts_undaria_spp_seaweed_wakame_raw htm, accessed 2 February 2009
Tobacman JK (2001) Review of harmful gastrointestinal effects of carrageenan in animal experiments. Environ Health Perspect 109:983–994
Torsdottir I, Alpsten M, Holm G, Sandberg AS, Tolli J (1991) A small dose of soluble alginate-fiber affects postprandial glycemia and gastric-emptying in humans with diabetes. J Nutr 121:795–799
Tseng CK (2001) Algal biotechnology industries and research activities in China. J Appl Phycol 13:375–380
Tsuchiya Y (1969) Comparative hypocholesterolemic activities of marine algae. Proceeding of the International Seaweed Symposium 6:747–757
Urbano MG, Goñi I (2002) Bioavailability of nutrients in rats fed on edible seaweeds, nori (Porphyra tenera) and wakame (Undaria pinnatifida), as a source of dietary fibre. Food Chem 76:281–286
Usov AI, Smirnova GP, Klochkova NG (2001) Polysaccharides of algae: 55. Polysaccharide composition of several brown algae from Kamchatka. Russ J Bioorgan Chem 27:395–399
Vadas S, Wright WA, Beal BF (2004) Biomass and productivity of intertidal rockweeds (Ascophyllum nodosum LeJolis) in Cobscook Bay. Northeastern Naturalist 11:123–142
Vairappan CS, Suzuki M, Abe T, Masuda M (2001) Halogenated metabolites with antibacterial activity from the Okinawan Laurencia species. Phytochemistry 58:517–523
Valente LMP, Gouveia A, Rema P, Matos J, Gomes EF, Pinto IS (2006) Evaluation of three seaweeds Gracilaria bursa-pastoris, Ulva rigida and Gracilaria cornea as dietary ingredients in European sea bass (Dicentrarchus labrax) juveniles. Aquaculture 252:85–91
van de Velde F, Lourenco ND, Pinheiro HM, Bakker M (2002) Carrageenan: a food-grade and biocompatible support for immobilisation techniques. Adv Synth Catal 344:815–835
van Netten C, Hoption Cann SA, Morley DR, van Netten JP (2000) Elemental and radioactive analysis of commercially available seaweed. Sci Total Environ 255:169–175
Vaugelade P, Hoebler C, Bernard F, Guillon F, Lahaye M, Duee PH, Darcy-Vrillon B (2000) Non-starch polysaccharides extracted from seaweed can modulate intestinal absorption of glucose and insulin response in the pig. Reprod Nutr Dev 40:33–47
Ventura MR, Castañón JIR (1998) The nutritive value of seaweed (Ulva lactuca) for goats. Small Rumin Res 29:325–327
Verdrengh M, Erlandsson-Harris H, Tarkowski A (2000) Role of selectins in experimental Staphylococcus aureus-induced arthritis. Eur J Immunol 30:1606–1613
Vlieghe P, Clerc T, Pannecouque C, Witvrouw M, De Clercq E, Salles JP, Kraus JL (2002) Synthesis of new covalently bound kappa-carrageenan-AZT conjugates with improved anti-HIV activities. J Med Chem 45:1275–1283
Voet D, Voet J, Pratt CW (1999) Fundamentals of biochemistry. Wiley, New York, 1095 pp
von Elbe JH, Schwartz SJ (1996) Colorants. In: Fennema OR (ed) Food chemistry. Marcel Dekker, New York, pp 651–722
Wahbeh MI (1997) Amino acid and fatty acid profiles of four species of macroalgae from Aqaba and their suitability for use in fish diets. Aquaculture 159:101–109
Wang WT, Zhou JH, Xing ST (1994) Immunomodulating action of marine algae sulfated polysaccharides on normal and immunosuppressed mice. Chin J Pharm Toxicol 8:199–202
Wang GC, Sun HB, Fan X, Tseng CK (2002) Large-scale isolation and purification of R-phycoerythrin from red alga Palmaria palmata using the expanded bed adsorption method. Acta Bot Sin 44:541–546
Wang Y, Han F, Hu B, Li JB, Yu WG (2006) In vivo prebiotic properties of alginate oligosaccharides prepared through enzymatic hydrolysis of alginate. Nutr Res 26:597–603
Wang J, Zhang QB, Zhang ZS, Li Z (2008) Antioxidant activity of sulfated polysaccharide fractions extracted from Laminaria japonica. Int J Biol Macromol 42:127–132
Washington N, Denton G (1995) Effect of alginate and alginate–cimetidine combination therapy on stimulated postprandial gastro-oesophageal reflux. J Pharm Pharmacol 47:879–882
Weinberger F (2007) Pathogen-induced defense and innate immunity in macroalgae. Biol Bull 213:290–302
Wen X, Peng CL, Zhou HC, Lin ZF, Lin GZ, Chen SW, Li P (2006) Nutritional composition and assessment of Gracilaria lemaneiformis Bory. J Integr Plant Biol 48:1047–1053
Whittaker MH, Frankos VH, Wolterbeek AMP, Waalkens-Berendsen DH (2000) Effects of dietary phytosterols on cholesterol metabolism and atherosclerosis: clinical and experimental evidence. Am J Med 109:600–601
Wikipedia (2010) www.wikipedia.com, accessed 25 June 2010
Witvrouw M, DeClercq E (1997) Sulfated polysaccharides extracted from sea algae as potential antiviral drugs. Gen Pharmacol Vasc Syst 29:497–511
Wong KH, Cheung PCK (2000) Nutritional evaluation of some subtropical red and green seaweeds. Part I—proximate composition, amino acid profiles and some physico-chemical properties. Food Chem 71:475–482
Wong K, Cheung PC (2001a) Influence of drying treatment on three Sargassum species. 2. Protein extractability, in vitro protein digestibility and amino acid profile of protein concentrates. J Appl Phycol 13:51–58
Wong K, Cheung PC (2001b) Influence of drying treatment on three Sargassum species. 1. Proximate composition, amino acid profile and some physico-chemical properties. J Appl Phycol 13:43–50
Wong KH, Cheung PCK (2001c) Nutritional evaluation of some subtropical red and green seaweeds. Part II. In vitro protein digestibility and amino acid profiles of protein concentrates. Food Chem 72:11–17
Wright AD, Goclik E, Konig GM (2003) Three new sesquiterpenes from the red alga Laurencia perforata. J Nat Prod 66:435–437
Wu XW, Yang ML, Huang XL, Yan J, Luo Q (2003) Effect of fucoidan on splenic lymphocyte apoptosis induced by radiation. Chin J Radiol Med Prot 23:43–50
Xu WJ, Liao XJ, Xu SH, Diao JZ, Du B, Zhou XL, Pan SS (2008) Isolation, structure determination, and synthesis of galaxamide, a rare cytotoxic cyclic pentapeptide from a marine algae Galaxaura filamentosa. Org Lett 10:4569–4572
Xue CH, Chen L, Li ZJ, Cai YP, Lin H, Fang Y (2004) Antioxidative activities of low molecular fucoidans from kelp Laminaria japonica. Dev Food Sci 42:139–145
Yamamoto I, Takahashi M, Tamura E, Maruyama H, Mori H (1984) Antitumor-activity of edible marine-algae—effect of crude fucoidan fractions prepared from edible brown seaweeds against L-1210 leukemia. Hydrobiologia 116:145–148
Yan XJ, Chuda Y, Suzuki M, Nagata T (1999) Fucoxanthin as the major antioxidant in Hijikia fusiformis, a common edible seaweed. Biosci Biotechnol Biochem 63:605–607
Yan XJ, Zheng L, Chen HM, Lin W, Zhang WW (2004) Enriched accumulation and biotransformation of selenium in the edible seaweed Laminaria japonica. J Agric Food Chem 52:6460–6464
Yang XL, Sun JY, Xu HN (1995) An experimental study on immunoregulatory effect of fucoidan. Chin J Mar Drugs 14:909–913
Yasui A, Tsutsumi C, Toda S (1978) Selective determination of inorganic arsenic(III), (V) and organic arsenic in biological materials by solvent extraction atomic absorption spectrophotometry. Agric Biol Chem 42:2139–2145
Ye H, Wang K, Zhou C, Liu J, Zeng X (2008) Purification, antitumor and antioxidant activities in vitro of polysaccharides from the brown seaweed Sargassum pallidum. Food Chem 111:428–432
Ying P, Shakibaei M, Patankar MS, Clavijo P, Beavis RC, Clark GF, Frevert U (1997) The malaria circumsporozoite protein: Interaction of the conserved regions I and II-plus with heparin-like oligosaccharides in heparan sulfate. Exp Parasitol 85:168–182
Yone Y, Furuichi M, Urano K (1986) Effects of wakame Undaria pinnatifida and Ascophyllum nodosum on absorption of dietary nutrients, and blood sugar and plasma free amino-N levels of red sea bream. Bull Jpn Soc Sci Fish 52:1817–1819
Young EG, Smith DG (1958) Amino acids, peptides, and proteins of Irish moss, Chondrus crispus. J Biol Chem 233:406–410
Yuan YV (2008) Marine algal constituents. In: Barrow C, Shahidi F (eds) Marine neutraceuticals and functional foods. CRC, Boca Raton, pp 297–320
Yuan YV, Walsh NA (2006) Antioxidant and antiproliferative activities of extracts from a variety of edible seaweeds. Food Chem Toxicol 44:1144–1150
Yuan YV, Carrington MF, Walsh NA (2005) Extracts from dulse (Palmaria palmata) are effective antioxidants and inhibitors of cell proliferation in vitro. Food Chem Toxicol 43:1073–1081
Zaragoza MC, Lopez D, Saiz MP, Poquet M, Perez J, Puig-Parellada P, Marmol F, Simonetti P, Gardana C, Lerat Y et al. (2008) Toxicity and antioxidant activity in vitro and in vivo of two Fucus vesiculosus extracts. J Agric Food Chem 56:7773–7780
Zee S (1991) Body weight loss with the aid of alginic acid. Arc Intern Med 3-4:113–114
Zeitlin L, Whaley KJ, Hegarty TA, Moench TR, Cone RA (1997) Tests of vaginal microbicides in the mouse genital herpes model. Contraception 56:329–335
Zeitoun P, Salmon L, Bouche O, Jolly, D, Thiefin G (1998) Outcome of erosive/ulcerative reflux oesophagitis in 181 consecutive patients 5 years after diagnosis. Ital J Gastroenterol Hepatol 30:470–474
Zhang QB, Li N, Zhou GF, Lu XL, Xu ZH, Li Z (2003) In vivo antioxidant activity of polysaccharide fraction from Porphyra haitanesis (Rhodephyta) in aging mice. Pharmacol Res 48:151–155
Zhang JZ, Tiller C, Shen JK, Wang C, Girouard GS, Dennis D, Barrow CJ, Miao MS, Ewart HS (2007) Antidiabetic properties of polysaccharide- and polyphenolic-enriched fractions from the brown seaweed Ascophyllum nodosum. Can J Physiol Pharmacol 85:1116–1123
Zhao X, Xue CH, Li BF (2008) Study of antioxidant activities of sulfated polysaccharides from Laminaria japonica. J Appl Phycol 20:431–436
Zhou G, Sheng W, Yao W, Wang C (2006a) Effect of low molecular [lambda]-carrageenan from Chondrus ocellatus on antitumor H-22 activity of 5-Fu. Pharmacol Res 53:129–134
Zhou Y, Yang HS, Hu HY, Liu Y, Mao YZ, Zhou H, Xu XL, Zhang FS (2006b) Bioremediation potential of the macroalga Gracilaria lemaneiformis (Rhodophyta) integrated into fed fish culture in coastal waters of North China. Aquaculture 252:264–276
Zhuang C, Itoh H, Mizuno T, Ito H (1995) Antitumor active fucoidan from the brown seaweed, Umitoranoo (Sargassum thunbergii). Biosci Biotechnol Biochem 59:563–567
Zou Y, Qian ZJ, Li Y, Kim MM, Lee SH, Kim SK (2008) Antioxidant effects of phlorotannins isolated from Ishige okamurae in free radical-mediated oxidative systems. J Biotechnol 136:S579–S580
Zubia M, Payri C, Deslandes E (2008) Alginate, mannitol, phenolic compounds and biological activities of two range-extending brown algae, Sargassum mangarevense and Turbinaria ornata (Phaeophyta: Fucales), from Tahiti (French Polynesia). J Appl Phycol 20:1033–1043
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Holdt, S.L., Kraan, S. Bioactive compounds in seaweed: functional food applications and legislation. J Appl Phycol 23, 543–597 (2011). https://doi.org/10.1007/s10811-010-9632-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10811-010-9632-5