
Abstract
Eggplant (Solanum melongena L.) is a major vegetable crop widely grown in tropics and subtropics. However, eggplant production is subject to high losses from biotic and abiotic stress. Eggplant is exposed to a broad range of biotic stresses such as nematodes, wilt diseases, eggplant fruit and shoot borer, two-spotted spider mites, whitefly and aphids. These biotic challenges reduce significantly yields, fruit quality, shelf-life, and nutritional content in eggplants. Farmers mostly rely on pesticides to control biotic stress. Breeders from public and private sector have performed some efforts for the development of pest-resistant varieties. To date, some disease resistance genes have been utilized in commercial cultivars, but much less progress has been achieved for arthropods resistance. In this book chapter, we review the basic information of the crop, major different biotic stresses in eggplant, genetic resources of resistance, traditional and marker assisted breeding, molecular mapping and cloning of resistance genes, genomics-assisted breeding, as well as genetic engineering for resistance traits. Furthermore, brief accounts on social, political and regulatory issues and future perspectives for this crop are highlighted.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
4.1 Introduction
Eggplant (Solanum melongena L.) is an economically important vegetable crop worldwide, with a total production of more than 52 million tonsand net value of over $21.4 billion according to statistics from the Food and Agriculture Organization of the United Nation (FAOSTAT 2017). Asia alone produces more than 90% of the global production of eggplant. As of FAOSTAT (2017), China (32.8 mill tons), India (12.5 mill tons), Egypt (1.3 mill tons), Turkey (0.82 mill tons) and Iran (0.75 mill tons) are the five leading eggplant producing countries in the world.
Eggplant (2n = 2x = 24), also known as brinjal eggplant, is native to the Old World and was first domesticated over 4,000 years ago in South East Asia (Meyer et al. 2012). Two other cultivated eggplants (S. aethiopicum and S. macrocarpon), which are members of the nightshade (Solanaceae) family are also grown. Brinjal eggplant (S. melongena) is a well known species worldwide, but the scarlet eggplant (S. aethiopicum L.) and the gboma eggplants (S. macrocarpon L.), have local importance in tropical Africa (Daunay and Hazra 2012). Solanaceae family includes approximately 3,000 species distributed in 90 genera. Solanum is the largest genus, which includes around 1,500 species such as eggplant and other globally important crops like potato (S. tuberosum L.) or tomato (S. lycopersicum L.). Solanum melongena and S.macrocarpon belong to section Melongena (Lester and Daunay 2003; Lester et al. 2011), whereas S. aethiopicum belongs to section Oliganthes (Lester 1986).
Eggplant is a low-calorievegetable crop and contributes to a healthydiet of consumers. It is a very good source of dietary fiber (both soluble and insoluble) and considered amongst the healthiest vegetables for their high content in vitamins, minerals and bioactive constituents for human health (Taher et al. 2017). The phytonutrients found in eggplant fruits include various phenolic compounds, such caffeic and chlorogenic acid and flavonoids such as nasunin. The benefits attributed to these compounds include antioxidant and antimicrobial activity. In many African countries, leaves and roots of African eggplant are used as medicines for the treatment of several ailments such as high blood pressure; cure wounds, diabetes, or inflammatory tumours (Oboh et al. 2005).
Eggplant production is severely constrained by biotic and abiotic stresses in tropics and subtropics. The most common abiotic stress includes high and low temperatures, salinity, and drought, and biotic stress includes nematodes, wilt diseases caused mostly by Ralstonia solanacearum, Fusarium oxysporum f. sp. melongenae, Verticillium dahliae, and insect pests such as eggplant fruit and shoot borer (Leucinodes orbonalis Guenée), whitefly (Bemisia tabaci Gennadius), two-spotted spider mites (Tetranychus urticae Koch), leafhopper (Amrasca devastans Distant), and aphids (Aphis gossypii Glover) (Taher et al. 2017). Cultivated eggplants are related to a large number of wild species which are rich sources of variation for breeding programs, in particular for traits related to adaptation to climate change including disease and insect resistance (Rotino et al. 2014).
Little breeding efforts have been made in eggplant compared to other major Solanaceous crops such as tomato (S. lycopersicum), potato (S. tuberosum), and pepper (Capsicum annuum) (Daunay and Hazra 2012), mostly because production is overwhelmingly concentrated in developing countries where investments in breeding are often reduced. Current eggplant breeding programs focuses on development of F1 hybrids with high-yield and fruit quality as well as resistance to abiotic and biotic stresses.
4.2 Description on Different Biotic Stresses
Eggplant is one of the most important vegetables in tropics and subtropics, but production is subject to high losses due todiseases and insect pests. In addition, biotic stress reduces fruit quality, shelf-life, and nutritional content of eggplant. Farmers often depend on pesticides to control diseases in the absence of resistant cultivars. Intensive pesticide use in eggplant poses health hazards to growers and their families, the environment, and consumers; as well as increases the cost of production, which makes this vegetable expensive for poor consumers (Ramasamy 2009).Resistant cultivars are among the cheapest, simplest, and most environmentally safe ways to manage diseases and insect pests. The major biotic stress include bacterial wilt, Fusarium wilt, Verticillium wilt, eggplant leaf spot, brinjal little leaf, eggplant fruit and shoot borer, spider mites, leafhopper and whitefly.
4.2.1 Bacterial Wilt
The disease is caused by Ralstonia solanacearum (Smith) Yabuuchi et al., which has many hosts in the nightshade family including tomato, pepper and potato. It is a soil-borne bacterium and infection takes place through roots and especially damaged root tissue. Infection may follow infected planting material, contaminated tools or irrigation water. High temperature increases the growth of the pathogen and therefore the bacterial wilt is a problem mainly in the tropics and subtropics. Symptoms are seen as wilting but then the roots and stems are already damaged and the losses can be fatal Genin et al. (2012). Cultural methods as crop rotation and hygiene measures are important, as chemical methods are not effective due to the location of the pathogen deep inside the xylem or the soil. Usingresistant cultivars and rootstocks for grafting has been a more reliable strategy, but requires access to such germplasm and a screening and breeding pipeline (Huet 2014; Keatinge et al. 2014). At the World Vegetable Center, accessions from eggplant and its wild relatives have been screened for resistance to bacterial wilt with promising results (AVRDC 1999; Namisy et al. 2019). Especially the wild relatives are interesting here, which have beenconfirmed in several studies (see Genetic Resources Section for details). A challenge however is how to move from trait discovery to resistant cultivars, as it is hampered by polygenic inheritance and linkage drag with traits associated with the wild species (e.g., Boshou 2005).
4.2.2 Fusarium Wilt
The disease is caused by Fusarium oxysporum f. sp. melongenae (Fomg), which is a soil-borne pathogen that also is a problem in other Solanaceous crops. Symptoms are discoloration of leaves that later develops into wilting plants with severe stem and root damages. Cultivation methods with raised beds that promote soil water drainage may reduce the infection. The pathogen can survive as chlamydospores over time in soil and plant debris. Crop rotation with non-hosts will however reduce the population. Disinfection of equipments and removal of infected derbies is important hygiene measures if infection is found. Fungicide treatment is difficult to apply as the pathogen is located deep inside the root and stem. Fusarium wilt resistant varieties is a way forward and resistance genes have been identified in both cultivated and wild eggplants and a review of these sources is provided under the Genetic Resources Section in this chapter. Resistant rootstocks could be of great value, as grafted plants are presenting a good level of resistance.
4.2.3 Verticillium Wilt
The disease is caused by the fungus Verticillium dahliae Kleb., which has many host plants and can survive for several years as microsclerotia in the soil. Symptoms are curling leaves with discolouration, and early senescence and dieback of plants. Crop rotation with at least two years of non-host crops like wheat, corn and barley may help, as removing plant derbies and other hygiene measures like clean planting material. Soil fumigation or pre-plant fungicide treatments are methods used. Resistant varieties would be a good alternative, but so far, there are not many varieties released with proper resistance. Resistance sources have been identified but these are found in wild eggplant species, which means that a lot of breeding effort is needed. Grafting eggplants onto resistant tomato rootstocks to suppress the infection is an alternative tested (Liu et al. 2009).
4.2.4 Eggplant Leaf Spot
The disease is caused by Pseudocercospora egenula (Syd.) U. Braun & Crous, which is a fungus. Symptoms start as round yellow spots, as they grow larger, the shape becomes irregular, merge and turn brown, and the older leaves die. Spores are spread by wind and rain splash. Locally, the disease can be a large problem and fungicides are applied. Some differences in tolerance among varieties arereported but the disease is not fully mapped (Liang et al. 2016; Vaghefi et al. 2016).
4.2.5 Brinjal Little Leaf
Brinjal little leaf is caused by a phytoplasma. Symptoms are small leaves that turn yellow and later the whole plant is affected, including fruit setting and yield causing considerable economic losses (Rao et al. 2010). Phytoplasma belonging to six groups, and from different parts of the world, have been reported to infect eggplant (Kumar et al. 2017; Kumari et al 2019). The pathogen is transmitted via leafhoppers. Other Solanum crops but also weeds may serve as host plants. To reduce the problem, vectors but also certainweeds should be controlled, but there are no direct chemical treatments of plant phytoplasmas. Tolerant varieties have been developed (Chakrabarti and Choudhury 1975) but not many publications on resistance screening and recent breeding are available.
4.2.6 Eggplant Fruit and Shoot Borer
Eggplant fruit and shoot borer is major pest on eggplant throughout the tropics in Asia and Africa. The larva feeds on the tender shoots, flower buds, flowers, and fruits. It also tunnels inside the shoot and feeds on the inner contents, resulting in wilting of young shoots, followed by drying and dropoff, which slows plant growth. In addition, the larva feeds inside the fruit and creates tunnels filled with frass and fecal pellets, which makes the fruit unmatketable and unfit for consumption. Farmers are mostly applying large quantities of insecticides to control the pest, which has resulted in the development of pesticide resistance in insects. Tolerance to this pest has been reported in some local varieties in India such as Pusa Purple Long, Pusa Purple Cluster, Pusa Purple Round, Aushey, Shyamla Dhepa, Banaras Long Purple, Arka Kesav, Arka Kusmakar, Punjab Barsati, Punjab Chamkila, Kalyanpur-2 and Gote-2 (Parker et al. 1995; Alam et al. 2003; Shivalingaswamy and Satpathy 2007).
4.2.7 Spider Mites
The two-spotted spider mite [T. urticae Koch (Acari: Tetranychidae)] is a very distructive pest in eggplant worldwide. This pest mostly prefers to live in colonies on the underside of leaves, and high temperatures and dry conditions are suitable for the multiplication and reprodution. Adults suck the chlorophyll, nutrients, and water from the leaf cells with their piercing and sucking mouthparts which causes foliar damage with tiny white or yellow spots on the leaves. Under heavy infestation, the mites move to the tip of the leaf or top of the plant and result in leaf discoloration, often called bronzing and drop off, and will finally lead to stunting or plant death. Chemical control of spider mites is often costly, and the excessive use of pesticides harms human health and environment. Resistant cultivars are not available and biological control by natural enemies could be used in protected cultivation but is not feasible in the field (Taher et al. 2019).
4.2.8 Leafhopper
The leafhopper [Amrasca biguttula (Ishida) (Hemiptera: Cicadellidae)] is an eggplant pest in several countries in Asia. Both nymphs and adults suck the sap from the undersides of leaves causing small and yellow patches, followed by crinkling, curling, bronzing, and drying, or “hopper burn” in severe attacks. High infestation also causes a reduction in yield. Leafhoppers may transmit viruses and phytoplasma that causes the so-called little leaf disease. Management of leafhopper might include use of tolerant eggplant varities such as Manjari Gota, Vaishali, Mukta Kesi, Round Green, and Kalyanipur T3, growing okra as a trap crop along the borders of an eggplant field, and using natural predators such as ladybird beetles and green lacewings which are highly efficient in preying on leafhopper nymphs and adults.
4.2.9 Whitefly
The sweetpotato whitefly (Bemisia tabaci Gennadius) can cause considerable direct and indirect damages in eggplant. This pest causes direct damage on leaves that result in reduced leaf photosynthetic efficiency, as well as on the fruits which increases the number of unmarketable fruit (Schuster et al. 1996; Rakha et al. 2017; Taher et al. 2020). Through indirect damage, whitefly transmitsmany species of plant viruses as well. The control of whitefly is highlydifficult due to high reproductive capacity and it can quickly develop resistance against insecticides (Rakha et al. 2017). Eggplant farmers apply a lot of insecticides, particularly in developing countries, which is often costly. Biological control by natural enemies is not sufficient in the open fieldconditions and no eggplant cultivars are resistant to whitefly (Taher et al. 2020).
4.3 Genetic Resources of Resistance Genes
Crop wild relatives of eggplant are rich sources of variation for pre-breeding and breeding programs, particularlyfor traits related to adaptation to climate change (Taher et al. 2017, 2020). These wild relatives mostly produce small, bitter, multi-seeded fruits, almost always inedible and with prickly calyx. Some of the eggplant wild relatives contain high levels of chlorogenic acid and other bioactive compounds, which may have potential interest for human health (Meyer et al. 2015). The wild species were classified into three genepools based on crossability relationships and following the biological concept of species (Harlan and de Wet 1971). For instance, in eggplant, S.insanum can be crossed easily and produce normal fertile hybrids whichis considered underprimary genepool (GP1) of eggplant (Plazas et al. 2016; Syfert et al. 2016). More than 40 African and Southeast Asian species are classified assecondary genepool (GP2) ofeggplant based on crossability relationships and phylogenetic studies. However, some interspecific hybrids derived from GP2 were partly sterile or weak due to reproductive barriers such as S. dasyphyllum, S. linnaeanum or S. tomentosum (Rotino et al. 2014; Kouassi et al. 2016). The tertiary genepool (GP3) includes more distantly related species and the crosses with cultivated eggplant generally failed or need specific breeding techniques such as embryo rescue to succeed (e.g., Solanum torvum and S. elaeagnifolium) (Kouassi et al. 2016; Plazas et al. 2016; García-Fortea et al. 2019).
Resistance to biotic stress has been reported in crop wild relatives of eggplant (Table 4.1). At World Vegetable Center, about 200 accessions were evaluated for resistance to bacterial wilt (Ralstonia solanacearum) under greenhouse using root wounding and soil drenching inoculation methods and 38 accessions were identified with high levels of resistance (AVRDC 1999).
Resistance to whitefly was detected in the eggplant wild relatives Solanum dasyphyllum, S. campylacanthum, S. tomentosum and S. pyracanthos (Taher et al. 2020).In addition, resistance to spider mite was detected in African eggplant S. macrocarpon as well as wild relatives such as S. sisymbriifolium, S. dasyphyllum and S. torvum (Taher et al. 2019). Resistance to leafhopper and aphids was found in eggplant accessions VI034971, VI035822, and VI035835. These results show that crop wild relatives of eggplant are very promising materials for breeding pest tolerant and resistant varieties can be developed.
4.4 Glimpses on Traditional Breeding
4.4.1 Focus of Traditional Breeding
The classical breeding of this crop has focused on the development of high yielding varieties with larger and more uniform fruits with less flesh browning, a characteristic that greatly reduces the quality of the fruit (Hurtado et al. 2013). Eggplant is highly variable in shape, color and size, and this has allowed the selection of a broad array of improved cultivars adapted to local preferences for fruit size, shape, and color; however, the genetic diversity of the cultivated eggplant genepool is narrow (Muñoz-Falcón et al. 2009a, b). The focus aimedatbreeding for tolerance to stresses has been much less intense than in other major Solanaceae crops, like tomato, potato or pepper. In this way, although tolerance to abiotic stresses have also been sought (Plazas et al. 2019), more works have been performed on finding resistance to biotic stresses, like root-knot nematodes, bacteria, fungus and some insects (Kalloo 1993; Rotino et al. 2014; Miyatake et al. 2016). Other recent works have focused on the study of the functional and nutraceutical characteristics attributed to this crop, rich in antioxidant compounds (Plazas et al. 2013; Kausik et al. 2016). The development of seedless varieties has also been a major focus of eggplant breeding, and some highly parthenocarpic materials have been obtained (Miyatake et al. 2012; Du et al. 2016; Li et al. 2012).
The feasibility of obtaining hybrids with many wild relatives (Daunay et al. 2012; Rotino et al. 2014; Kouassi et al. 2016; Plazas et al. 2016) holds an immense potential to genetically improve eggplant. This makes possible the transfer of genes of interest from wild relatives into the genetic background of eggplant. In the last decades some examples of genetic transfer of interest from wild species to the genetic background of the eggplant have been established. These include the development of introgression lines with several wild species such as S. incanum (Gramazio et al. 2017a, b) for adaptation to climate change, as well as backcross generations with S. linnaeanum and S. tomentosum for resistance to Fusarium, Verticillium and nematodes (Toppino et al. 2018).
4.4.2 Limitations of Classical Endeavors and Utility of Molecular Breeding
The increasing population in the world, climate change and the high incidence of new diseases in crops has posed a challenge for modern agriculture and breeding (Ray et al. 2013; Brooks and Blandford 2019). In eggplant, as in most major crops, classical breeding methods, with a selection of traits of interest, and the subsequent hybridization, selection and fixation in the cultured materials has been very efficient in delivering improved cultivars; however, nowadays other approaches must be used to accelerate the processes. The speed at which cultivars become obsolete has led breeders to develop breeding methods in a shorter time, where marker-assisted selection is essential. Access to the eggplant genome (Barchi et al. 2019a) and the syntenic relation with the tomato genome (Wei et al. 2020), together with the large number of wild species with which the eggplant can obtain fertile hybrids, is making the speeding of the breeding process possible.
The application of modern molecular marker based approaches carried out have accelerated the process of introduction and localization of genes of interest, including genes from related wild species, in the eggplant genepool. In this way, Wei et al. (2020) found 210 markers associated with around 71 traits of interest in eggplant. Furthermore, the new methodology suggested by Prohens et al. (2017) called “introgressiomics”, proposes the massive introduction of genes from wild species in the genetic background of the crop of interest, where the generated materials, with wild introgressions, will be tested and genotyped when necessary to solve the problem that arises. This approach greatly benefits from the use of marker assisted selection for forward and background selection.
The potential of molecular markers for the development of a new generation of eggplant cultivars comes through its application for genetic mapping and detection of quantitative trait loci (QTLs) in natural (germplasm collections) and experimental (biparental, multiparental, advanced backcrosses and introgression lines sets) populations. The use of genome-wide association studies (GWAS) in germplasm populations as well as in biparental populations has allowed the identification of severalQTLs for morphological and agronomic traits of interest (Ge et al. 2013a; Cericola et al. 2014; Portis et al., 2015; Toppino et al. 2016). One of the potentially most useful experimental populations in eggplant will be a multiparent advanced generation intercross (MAGIC) population which is under development at the Universitat Politècnica de València. In these populations, eight eggplant parents (including a S. incanum wild accession) have been intercrossed and recombinant inbred lines (RILs) which are an admixture of the eight genomes are under development (Arrones et al. 2020). The use of experimental populations in eggplant can go beyond the identification of QTLs, and thanks to the fine mapping of a segregating eggplant population, a deletion has been found in the genome of eggplant that could be related to the prickliness trait (Miyatake et al. 2020). Also, Gramazio et al. (2014) found some interesting genes related with polyphenol oxidases involved in chlorogenic acid biosynthesis pathway in a first backcross population.
4.4.3 Positive and Negative Selection in Eggplant Breeding
Traditionally, the breeding objectives in eggplant have been to improve the size and uniformity of the fruit, an increase in yield, as well as to eliminate the bitter taste that characterizes the fruit of primitive varieties and wild ancestors of eggplant. In this way, some traits, such as yield and fruit size have been under positive selection, while others have been under negative selection. Regarding traits under negative selection, eggplant, as other Solanum species, contains solasonine and solamargine, two glycoalkaloids with a toxic effect on humans and that give this crop a bitter taste, limiting its use at a commercial level (Cham 2012; Ranil et al. 2017). The presence of saponins also contributes to the bitter taste (Toppino et al. 2016) and has also been selected against its presence. Another undesirable trait is the oxidation that occurs when cutting the fruit and that depreciates its quality, known as browning (Plazas et al. 2013). Selection against this trait has indirectly led to the negative selection of phenolic acids content, and therefore accessions with low browning and at the same time with low antioxidant content. Also, in the case of eggplant, the reduction in the number of seeds results in decreased browning (Maestrelli et al. 2003). Another characteristic that has been under strong negative selection is the presence of prickles both on the plant and on the fruit calyx, as this makes the management difficultof the crop and the marketing of the fruits (Miyatake et al. 2020). Another important trait under negative selection has been the presence of dormancy, very common in eggplant wild relatives, and that is clearly detrimental in the cultivated eggplant.
Some traits that have been under positive selection include the high yield and fruit set. Other traits in which a great emphasis has been paidis the intense dark color, resulting from the combination of chlorophylls and anthocyanins in the eggplant peel. Some recent breeding programs have also been aimed at improving the tolerance to biotic and abiotic stresses, although up to now the results have been less impressive, as well to improving the bioactive properties of the eggplant fruit by improving the content in bioactive phenolic acids of interest for human health.
4.4.4 Classical Breeding Achievements and Limitations
Many of the efforts made to improve the cultivation of eggplant have been carried out using classical breeding methods, where much importance has been given to yield and the external characteristics of the fruit, as well as to tolerance to some biotic and abiotic stresses (Kalloo 1993).
Taking advantage of the wide phenotypic diversity that this crop has, today it is possible to find eggplants of different sizes, from a few g to more than 1 kg, colors, such as white, black, purple, or green, with different patterns of colorations, and shapes including long and narrow (even serpentiform), long and wide, round, oval, and flattened. Depending on the target market, some types are more demanded than others. A wide variability can also be found in terms of texture and taste, as well as on response to biotic and abiotic stresses, which are one of the most important sources of economic losses.
Most of the modern eggplant cultivars have been developed using only intraspecific diversity. Given the potential of wild species as sources of variation for many traits and the limitations that have been found in the use of traditional breeding, multiple strategies have been developed to foster the use of wild species in eggplant breeding. In the "introgressiomics" approach (Prohens et al. 2017) the objective is to develop materials and populations that contain a large number of fragments of wild species, distributed throughout the genome of cultivated species. The application of this approach in eggplant is generating a large array of materials that can be used to address present and future challenges in this crop. For this new approach we will need to have identified the introgressions need to be identified taking advantage of the high throughput molecular markers.
4.5 Brief on Diversity Analysis
4.5.1 Phenotype-Based Diversity Analysis
A main challenge with phenotype-based characterization is to avoid environmental bias. Standardized sets of descriptors and methods are developed for eggplants (IBPGR 1990; van der Weerden and Barendse 2007). In addition, the phenomic tool developed for tomato (Rodríguez et al. 2010) has been applied for eggplants with good results (Prohens et al. 2012; Hurtado et al. 2013; Kaushik et al. 2016). Coming to how to express phenotypic diversity and relationships, different measures are used, from Coefficient of variation and Shannon diversity index to more multivariate tools (Everitt 1998; Spellerberg and Fedor 2003).
Brinjal eggplant, and especially its fruits, exhibit large phenotypic variation, as people have been selecting for traits as color, shape, texture and size. This adaptive evolution has produced many cultivars with a seemingly high diversity, but based on a limited number of traits. Vegetative traits have been less emphasized. Nevertheless, they are of major importance for photosynthesis and plant arrangement, and for producing high yielding and robust plants. Regarding phenotypic diversity within a population or an accession, modern varieties are per definition uniform and stable, in contrast to traditional cultivars (landraces) that are more diverse, and wild eggplants that are even more diverse. A study in black-fruited brinjal eggplant clearly showed such a pattern (Muñoz-Falcón et al. 2009a, b). Furthermore, round-fruited cultivars have been found to be more diverse than semi-long or long-fruited cultivars (Prohens et al. 2005; Tümbilen et al. 2011).
Crop diversity is safeguarded in public seed banks (gene banks), in breeders' collections and in-situ in farmers’ fields and in the wild. Gene banks provide information on phenotypic characters, and this on accession (seed sample, cultivar) level. Data include plant growth habit, plant height, branching and leaf- flower- and fruit trait characters (e.g. Boyaci et al. 2015; Taher et al. 2017). The largest ex-situ collection of eggplants is maintained at World Vegetable Center, with more than 2,700 accessions (Taher et al. 2017). This collection houses large phenotypic diversity. Regarding abiotic and biotic stress resistance, less data are available. Nevertheless, the information is increasingly important. An old study at the World Vegetable Center examined two hundred brinjal eggplant accessions for bacterial wilt (Ralstonia solanacearum) resistance and 38 were identified as promising (AVRDC 1999). Other trials have examined resistance for eggplant fruit and shoot borer (Leucinodes orbonalis), two-spotted spider mite (Tetranychus urticae), leafhopper (Amrasca devastans), and aphids (Aphis gossypii). Some brinjal eggplant accessions showed high level of resistance (Ramasamy 2009). For two-spotted spider mite, resistance was detected in accessions of scarlet- and gboma eggplants (Taher et al. 2019).
4.5.2 Genotype-Based Diversity Analysis
Over the years, different marker systems have come. In the early 1990s, restriction fragment length polymorphism (RFLP) markers were used in eggplant research but they were not so reliable (Sakata and Lester 1997). Later, random amplified polymorphic DNA (RAPD) and amplified fragment length polymorphism (AFLP) markers for eggplant were used (e.g.,Furini and Wunder 2004; Sing et al. 2006), as well as simple sequence repeats (SSRs) or microsatellites (Stàgel et al. 2008; Li et al. 2010; Tümbilen et al. 2011) and most recently came the single nucleotide polymorphism (SNP) markers (Gramazio et al., 2017a, b). The latter, with a panel of 5 k SNPs for coding regions and introns/UTRs has now been applied in single primer enrichment technology (SPET) for high-throughput genotyping in tomato and eggplant (Barchi et al. 2019a, b). A set of 422 eggplant accessions (including wild relatives and cultivated material) generated 30,731 high confidence SNPs, respectively. The authors concluded that this representsa robust; high-throughput technology for genetic fingerprinting that can be used to study genetic relationships among accessions and species, and that also could be useful in identification of mislabelled accessions and duplicates in genebanks. Many studies have been conducted of genotype-based diversity analysis. Details on a few studies are presented in the sections that follow. Here we restrict us to emphasizethat major efforts have been based on identifying and introducing resistance genes and on developing molecular markers for breeders. For Fusarium wilt, a resistance gene (Rfo-sa1), identified in scarlet eggplant, was introduced to brinjal eggplant (Toppino et al. 2008). Cleaved amplified polymorphic sequence (CAPS) markers could link the resistance to one major QTL (Miyatake et al. 2016; Barchi et al. 2018). Two other minor QTLs have been reported but for other Fusarium resistance genes (Miyatake et al. 2016; Mutlu et al. 2008). Verticillium vilt resistance has been introduced to brinjal eggplant from the wild species S. linnaeanum (Liu et al. 2015). Three QTLs were identified for resistance to this disease (Barchi et al. 2018). For bacteria wilt, a linkage map based on SNP markers has been developed, and one major specific and two broad-spectrum QTLs have been identified associated with this resistance (Salgon et al. 2017). Coming to other traits, markers have been developed for parthenocarpy (Daunay and Hazra 2012). Markers have also been associated to fruit contents of glycoalkaloids, sugars and organic acids (Toppino et al. (2016).
4.5.3 Relationship with Other Cultivated Species and Wild Relatives
Wild relatives are important gene sources for new diversity, and especially for pest- and disease resistance (Toppino et al. 2008; Daunay and Hazra 2012; Rotino et al. 2014; Liu et al. 2015). In addition, many of the wild species grow in extreme environment and can be valuable for climate adaptation (Knapp et al. 2013; Rotino et al. 2014; Syfert et al. 2016). Brinjal eggplant closest relative is S. insanum that grows wild in South- and Southeast Asia and is very prickly and weedy (Lester and Hasan 1991; Knapp et al. 2013; Ranil et al. 2017). A genetic similarity as high as 0.947 between the species was found in an old study using 52 accessions and RAPD markers (Karihaloo et al. 1995). The authors concluded that even despite morphologically different, it is not appropriate to distinguish S. melongena and S. insanum into different species. However, other studies do not agree (e.g. Iwata et al. 2008) and different relationships between the different eggplant species have been presented (Mace et al. 1999; Meyer et al. 2012). Iwata et al. (2008) compared brinjal eggplant to eight related species using inter-simple sequence repeat (ISSR) markers to evaluate the phylogenetic relationship, and identified seven groups: (i) S. melongena; (ii) S. aethiopicum and S. anguivi; (iii) S. incanum; (iv) S. violaceum and S. kurzii; (v) S. macrocarpon; (vi) S. virginianum and (vii) S. torvum.
With regardsto wild relatives that are close to brinjal eggplant, one also finds S. incanum L. and S. linnaeanum Hepper & P-M. L. Jaeger, as well as S. lichtensteinii Willd (Vorontsova et al. 2013; Acquadro et al. 2017). These species can be intercrossed with brinjal eggplants. In addition, the cultivated scarlet eggplant (S. aethiopicum L.) and gboma eggplant (S. macrocarpon L.) can also be hybridized with eggplant. These, in addition to their wild relatives S. anguivi Lam., S. dasyphyllum Schumach. & Thonn. and to some extent also S. tomentosum L., can be valuable gene sources for brinjal eggplant but are not closely related (Kouassi et al. 2016; Plazas et al. 2016). Intercrosses can however be made with intermediate fertility. Especially scarlet eggplant has served as a gene source for disease resistance in brinjal eggplant breeding (Prohens et al. 2012). More distant are the American species, S. sisymbriifolium Lam. and S. torvum Sw. (Vorontsova et al. 2013; Acquadro et al. 2017) but they could harbor potential genes of interest.
4.5.4 Relationship with Geographical Distribution
Brinjal eggplant can be divided into an oriental group traditionally grown in southern and eastern Asia and an occidental group traditionally grown in the Mediterranean basin including the Middle East, Europe and northern Africa. Several studies have pointed to geographical differentiation based on morphology (Chadha 1993; Daunay and Janick 2007) and genetics (Hurtado et al. 2012; Vilanova et al. 2012). For example, Vilanova et al. (2012), by using UPGMA procedures on SSR data, showed that 15 out of 16 accessions from the Mediterranean, Central Europe and Africa clustered together and another cluster had 5 out of 6 accessions from Eastern and Southeastern Asia. On a more detailed level, differentiation within regions is seen. For example, Gramazio et al. (2019a, b) examined landraces from Greece and found differentiation between accessions from the island and from the mainland. They concluded that Greece was part of a Mediterranean secondary center of diversity. We should say that the above-mentioned studies were in landraces, which are local varieties developed over time by farmers. What seems logical is that, as cultivars are becoming broader and with the use of genetic resources from different sources, geographical distribution patterns will be less clear. Liu et al. (2018) exemplifies this by showing that the accessions only partly clustered according to geographic origin. They examined 287 accessions and from around the world and included both inbred lines, cultivars and landraces.
4.5.5 Extent of Genetic Diversity
Genetic diversity can be expressed as Nei's genetic diversity index, expected heterozygosity (He) and/or Shannon's Information index (I). In addition, number of alleles per polymorphic locus, polymorphism information content (PIC) and observed heterozygosity (Ho) are measures used in genetic diversity studies.
Although morphologically diverse in fruits, cultivated eggplants have a much more narrow genetic background than its wild ancestors (Tümbilen et al. 2011; Vorontsova et al. 2013). This is a typical “bottleneck” effect of domestication (Meyer et al. 2012). To illustrate this, Kaushik et al. (2016) characterized 21 accessions from 12 different wild relatives to brinjal eggplant. In addition, they included interspecific hybrids and cultivars. The result clearly demonstrated that wild specieswere more variable than cultivars and the interspecific hybrids were in between.
Regarding genetic diversity within different types of brinjal eggplant accessions, Hurtado et al. (2012) examined 52 landraces (or selections within landraces) from three recognized secondary centers of diversity, China, Sri Lanka and Spain. They applied 12 highly polymorphic SSR markers that resulted in average PIC of 0.574 and totally 110 alleles were identified with 4.3, 5.3 and 4.2 alleles per polymorphic locus for accessions from China, Sri Lanka and Spain, respectively. The calculated genetic diversity was relatively high, with He of 0.494–0.540 for the Chinese and Sri Lankan landraces and somewhat lower for the Spanish landraces. Muñoz-Falcón et al. (2011) examined 42 landraces of brinjal eggplant from Spain, which included 25 striped accessions most of them of the popular Listada type but also non-striped landraces. They applied 17 SSR and 32 EST-SSR markers and the SSRs had greater polymorphism and polymorphic information content (PIC) than EST-SSRs. A considerable level of diversity was found, with a mean Nei's genetic diversity (He) value of 0.323 (varied from 0.195 to 0.441) and a Shannon's information index (I) value of 0.570.
Ge et al. (2013a) examined 92 cultivars (pure lines) of brinjal eggplant collected from 21 provinces in China. They applied a set of 100 SSR markers with a mean PIC value of 0.285, and found 311 polymorphic alleles. The Nei’s genetic diversity (He) was 0.323 and the average Shannon's Information index (I) was 0.570, but ranged from 0.060 to 1.341. The levels of eggplant genetic diversity decreased from south to north. Overall, the genetic diversity was lower in these cultivars than reported for landraces from China by Hurtado et al. (2012). Vilanova et al. (2012) examined 22 brinjal eggplant cultivars from around the world. They used 55 SSR markers with average PIC at 0.47. In total 203 alleles were detected, with an average of 4.7 per locus. The mean expected heterozygosity was 0.52 but the observed heterozygosity was as low as 0.06 and ranged from 0.00 for 16of the markers to 0.24 for one of the markers. Overall, the lowest diversity was found in non-hybrid cultivars compared to hybrid cultivars. The results demonstrate that inbreeding seems to be common in non-hybrid cultivars and that a very narrow genepool is applied in breeding programs. Liu et al. (2018) examined 287 of brinjal eggplant accessions and included inbred lines, cultivars and landraces from around the world. They applied 45 SSR markers and resolved 242 alleles and that ranged from 2 to 14 alleles per locus with an average of 5.38. Shannon information index ranged from 0.276 to 1.903 with an average of 1.055. Observed heterozygosity varied from 0.104 to 0.832 with a mean value at 0.558. There were big differences among all the 45 markers in polymorphism detection. The PIC value ranged from 0.102 to 0.815 with a mean value of 0.507.
4.6 Association Mapping Studies
4.6.1 Extent of Linkage Disequilibrium
Linkage disequilibrium (LD) is defined as allelic (neighbor variants) association at various genomic regions within the investigated population (Ramakrishnan 2013). In case of linkage disequilibrium, haplotypes do not happen at the expected rates when the alleles were independent (Goode 2011). On the contrary, LD takes place when haplotype frequencies are equal to the product of their corresponding allele frequencies. A haplotype is known to be the linked set of genes connected with one haploid genome. It is usually used to describe the linked genes of the major histocompatibility complex, where one haplotype is inherited from each parent. Linkage disequilibrium can be formed as a cause of selection, admixture, or bottlenecks. Studies of linkage disequilibrium patterns can contribute in branches like plant breeding, identification of genes responsible for a disease, and population history (Ramakrishnan 2013). There are plentiful ways to estimate deviation of LD which is referred to as D which was introduced in 1918 and is defined as comparing the observed and expected frequency of one haplotype, then the deviation is the difference between these two. LD can be positive or negative depending on when two alleles occur together on the same haplotype more or less often than expected (Goode 2011). The perception of LD is very important in genome-wide association studies (GWAS) where it helps in the identification of genetic markers that label the actual causal variants. A single-nucleotide polymorphism (SNP) is dissimilarity in a single nucleotide that arises at a specific position in the genome. In GWAS, markers for known complex traits, including some diseases, can be distinguished using SNPs across the genome. In population model LD is highly dependable for the success of GWAS using SNPs as genetic markers. LD is different from gametic phase disequilibrium which explains the non-random connotation of alleles within gametes (even for physically unconnected loci on different chromosomes) (Joiret et al. 2019). In eggplant, LD for some agronomic traits like anthocyanin pigmentation and fruit color was investigated with pair-wise r2, rs2 and r2sv where global LDlevel was 3.4 cM (Cericola et al. 2014). Additionally, Ge et al. (2013b) reported that analysis of LD revealed an extensive long-range LD of 11 cM within 141 accessions of eggplant using 105 microsatellite markers.
4.6.2 Target Gene Based LD Studies
Solanaceae species are considered a well-defined family regarding identification of quantitative trait loci (QTLs) as well as other genetic and genomic research (Gebhardt 2016). High-quality genetic linkage map and LD of eggplant genome aresubstantial approaches to identify target genes associated with biotic and abiotic stresses that are quantitatively inherited or qualitative among various germplasm collections. Target genes research based on LD concept is an advantageous pathway in molecular plant breeding programs and analysis of biotechnological processes for the development a valuable methods for crop improvement (Collonnier et al. 2001). Moreover, genetic linkage map construction representing quantitative genetic and genomics dataset that allows identifyingnew markers tightly related to the desirable horticultural traitsplays a key role in crop improvement and sustainable development. In eggplant, RAPD and AFLP markers have been used for a linkage map construction of fruit shape and color (Nunome et al. 2001). On the other hand, many linkage maps have been utilizedregarding a lot of horticultural traits and improvement of the level of resistance to various plant pathogens and insects that restrict production of eggplant (Lebeau et al. 2013; Portis et al. 2015; Barchi et al. 2016; Gramazio et al. 2017a, b). Genomic research strategies played a key role in smart plant breeding programs to detect particular trait through marker-assisted selection approach. The advantages of these DNA-based markers provide better chance for genetic variability assessment and mapping of QTLs among tested germplasm due to their cost, speed and reproducibility (Thapa et al. 2015). At present, there are many DNA-based markers for instance AFLP, RAPD, RFLP, microsatellite, SCoT (start codon targeted), etc. that can be applied in genetic and genomics studies including characterization, taxonomy, genetic diversity of wild and domesticated crop species.
4.6.3 Genome-Wide LD Studies
Despitethe economic importance of eggplant, omics research approaches including phenomic, genomics, transcriptomic, metabolomics and proteomic are still limited compared to other species of the Solanaceae family. Interestingly, the strategy of GWAS is considered a powerful substitutional strategy for investigating the genetic background of various agronomic traits. Furthermore, GWAS can be applied with many samples and accessions to represent the association between phenotypic and genotypic data as well as acceleration of plant breeding programs and crop improvement. Recently, GWAS approach was involved in eggplant research to identify multiple agri-horticultural traits such as fruit color, size and shape; serious biotic stress resistance, and productivity under difficult environmental conditions (Rotino et al. 2014). The genome-wide association (GWA) analysisis efficient in supporting and confirming QTLs and targeted genomic regions with different unique DNA markers. Concerning germplasm sets used for GWAS, LD may be specified by plant hybridization system, frequency of recombination, level of mutations, structure of tested populations, genetic linkage, alteration of gene sequence and natural selection and domestication (Rafalski and Morgante 2004). Globally, the genome-wide association mapping approach has been accomplished on eggplant populations with plenty of traits of interest (Fig. 4.1). Additionally, 191 accessions of eggplant including cultivars, lines and landraces were genotyped for association of fruit color and anthocyanin pigmentation where 338 SNPs were identified (Cericola et al. 2014).

Overview of association mapping in eggplant species to exploiting the available genetic resources for single nucleotide polymorphisms identification and gene annotation
4.6.4 Future Potential for the Application of Association Studies for Germplasm Enhancement
Association mapping is a very useful tool in modern agricultural and breeding programs to identify genomic regions that are responsible for traits of interest. Genomic association research in crop species are still ongoing where convenient trait measurements methodology and genotyping platforms with powerful analysis software are still the great challenges regarding crop improvement and molecular breeding. To maximize the future benefitof association mapping studies, all historical information about the population structure, size and marker density have to be known and informed (Álvarez et al. 2015). Moreover, advanced computational tools and bioinformatics analysis can play a crucial role in genetic resources and germplasm enhancement in the next decades. Furthermore, the next-generation sequencing technologies and whole-genome resequencing approaches are a great opportunity to improve association studies not only in model crops but also in non-model plant species. In the future, network of artificial intelligence can be applied in association research by large scale to minimize the genome-wide error rate and get more accuracy dataset. On the other hand, the future approaches of association mapping can be selected upon LDhistory in the tested germplasm, targeted traits complication, the provided historical knowledge of pedigree, availability of the reference genome of the target crops and structure of population under the study.
Analysis of GWA of 191 eggplant accessions demonstrated that 79 SNPs were mapped on 39 genomic regions and scattered over all 12 chromosomes of eggplant (Portis et al. 2015). Specific length amplified fragment sequencing (SLAF-Seq) strategy was applied for construction of high-throughput SNP markersin the genome of eggplant. Subsequently, GWA and QTL analysis were conducted within F2 population of 121progynies where 2,122 SNPs, 12 linkage groups and 19 QTLs were generated for leaf morphology, height of stem and fruits (Wei et al. 2020). Moreover, Toppino et al. (2020) genotyped 163 RILs using GBS strategy where 7249 SNP markers were assigned to the 12 eggplant chromosomes and spanning 2169.23 cM. This association revealed the molecular bases of seven horticultural traits associated with anthocyanin and seed vigor where from 7 to 17 QTLs for each trait were detected as well as development of MAS for further research.
4.7 Brief Account of Molecular Mapping of Resistance Genes and QTLs
4.7.1 Brief History of Mapping Efforts
Genome mapping is a widely uasedmeansto investigate the genetic information of an organism for the genomic regions/genes that are associated with a desirable trait. Two main groups of genome mapping have been obviously reported which are genetic map and physical map. Actually, genetic map refers to the Mendelian rules of segregation and recombination to decide the distance between varied genomic sites within a gene or between different genes on the same chromosome with cM (centi-Morgan) units, where the linkage idea is important in genetic map construction. The connotation of genetic linkage mapping has been reported earlerin 1913 where five sex genes on the chromosome Y of Drosophila melanogaster were linked (Sturtevant 1913). On the other hand, physical map is a molecular biology strategy to determine the order of DNA fragments at the level of chromosome comprising of the whole genome or a specific genomic region that is responsible for trait of interest and can be a count as a real map expressed in million base pairs (Mbps). It is noticeable that great efforts have been successfully done regarding genomic and genetic research in eggplant. Many QTLs for horticulturally desirable traits such as fruit weight (fw), fruit shape (fs), fruit calyx prickliness (fcpri), number of seed locules (slon), plant growth habit (hab), leaf prickliness (lepri), etc. have been effectively identified in eggplant (Portis et al. 2015; Toppino et al. 2020; Wei et al. 2020). GWAS hasobviousadvantages than other approaches for QTL mapping and SNPs discovery. Furthermore, GWAS enables phenotypic/genotypic variation within and between an array of accessions to identify targeted genes and for crop improvement (Portis et al. 2015). The high-quality reference genome of eggplant (Barchi et al. 2019a) will provide new insights with relevance to resequencing research, domestication and evolutionarly mechanisms of eggplant population. Addtionally, other genomic approaches and bioinformatics tools play substantial role in association studies, smart breeding programs, ecology and geographical origin, and climate-resilient cultivars in order to face the potential challenges (Gramazio et al. 2019b).
4.8 Marker-Assisted Breeding for Resistance Traits
4.8.1 Germplasm Characterization
Identification of sources of tolerance or resistance to stresses is a first step for conventional or marker-assisted breeding of these traits. Throughout the last decades multiple screenings for some of the most important stresses, mostly biotic (diseases and pests), involving both intraspecific and interspecific genetic resources have been performed (Toppino et al. 2021).
The most damaging soil-borne pathogens of eggplant are Ralstonia solanacearum, Fusarium oxysporum f. sp. melongenae and Verticillium dahliae, which are the respective causal agents of bacterial wilt, Fusarium wilt, and Verticillium wilt. Screening of collections of genetic resources against the different phylotypes of R. solanacearum have allowed the identification of several sources of resistance in the cultivated eggplant and in eggplant relatives (Lebeau et al. 2011; Namisy et al. 2019). Within S. melongena some resistant accessions have been discovered and so far the most studied are EG203 and AG91-25 (Lebeau et al. 2013; Salgon et al. 2018), although many others have been identified (Barik et al. 2020). Interestingly, AG91-25 derives from the hybridization between S. melongena and S. aethiopicum (Ano et al. 1991; Salgon et al. 2018). Amongst the eggplant relatives that can be successfully hybridized with eggplant via sexual crosses (Daunay et al. 2019), resistances have been found in several species, such as S. aethiopicum, S. anguivi, S. incanum, S. insanum, and S. torvum (Namisy et al. 2019; Barik et al. 2020). In addition, high levels of resistance have been found in other Solanum wild species. However, given the high genetic variability of R. solanacearum, resistance levels often depend on the strain used (Lebeau et al. 2011; Namisy et al. 2019). Resistance to Fusarium wilt has been found both in the cultivated species and in wild and cultivated eggplant relatives such as S. aethiopicum, S. incanum, S. linnaeanum, S. sisymbrifolium, S. torvum, S. viarum or S. violaceum (Cappelli et al. 1995; Boyaci et al. 2012; Altinok et al. 2014). Some of the resistances found are stable against a wide range of isolates (Altinok et al. 2014) providing sources of resistance of great value for the breeders. In particular, the resistance derived from S. aethiopicum has proved of great interest for eggplant breeding (Toppino et al. 2008). Different levels of resistance and tolerance to Verticillium wilt have been identified in the eggplant germplasm genepools. However, reports of high levels of tolerance within the cultivated species are scarce. In this respect, the best sources of resistance have been found in wild species such as S. anguivi, S. incanum, S. linnaeanum, S. tomentosum, S. torvum, S. sisymbriifolium, or S. viarum.
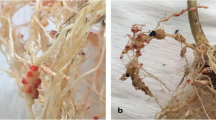
Resistance to diseases other than the three major ones mentioned above has been found both in the cultivated and wild eggplant genepools (Toppino et al. 2021). In this way, resistance has been found either in cultivated eggplant and/or in eggplant relatives to fungal pathogens that may affect eggplant such as Alternaria melongenae, Cercospora solani, Cercospora solani-melongenae, Colletorichum coccodes, Colletotrichum gloesporioides f. sp. melongenae, Fusarium solani, Leveillula taurica, Phomopsis vexans, Phytophthora parasitica, Phytophthora capsici, Sclerotinia sclerotiorum, Sclerotinia rolfsii, or Verticillium albo-atrum (Daunay and Hazra 2012; Rotino et al. 2014; Toppino et al. 2021). A number of studies have also screened cultivated and wild germplasm for resistance to root-knot nematodes (Meloidogyne spp.), mostly M. incognita. Although most of the cultivated eggplant materials have been found to be susceptible, some lines have found to be tolerant or partially resistant to root-knot nematodes (Colak-Ates et al. 2018), or in the case of line A-264-A from the Philippines, fully resistant to M. javanica (Boiteux and Charchar 1996). The most promising materials have been found in the related species genepool. In this way, high levels of resistance to nematodes have been found in accessions of S. torvum, S. viarum and S. stramonifolium (García-Mendivil et al. 2019).
Regarding viruses and phytoplasma, a large screening for resistance to tobamoviruses (Rast 1991) resulted in the identification of sources of resistance against Bell pepper mottle virus (BPMV), Tobacco mosaic virus (TMV) and Tomato mosaic virus (ToMV). Resistance to Pepper mild mottle virus (PMMV) has also been found in some accessions of S. aethiopicum (Tzortzakakis et al. 2006). However, no resistance against Potato virus Y (PVY) was found in a screening of 77 eggplant accessions (Colak-Ates et al. 2018). Screening for little leaf disease, caused by phytoplasma, has been found within eggplant germplasm, as well as in S. aethiopicum.
Compared to disease resistance, the screening of sources of resistance against pests has been less intense. One of the major pests in Southeast Asia is the fruit and shoot borer (Leucinodes orbonalis). Few materials of potential interest have been found in the cultivated eggplant genepool against this insect, although it has been suggested that varieties with high contents of phenolics, glycoalkaloids, dietary fiber, ash, starch and polyphenol oxidase activity are less susceptible (Doshi 2004; Prasad et al. 2014). Some resistances to L. orbonalis have been found in some eggplant relatives, such as S. aethiopicum, S. incanum, S. macrocarpon, and S. violaceum. A few research works have been performed on the identification of sources of resistance to other eggplant pests. Recently, Taher et al. (2020) have identified several sources of resistance to the whitefly Bemisia tabaci in one eggplant accession (MEL2) as well as in the wild species S. campylacanthum, S. dasyphyllum, S. pyracanthos, and S. tomentosum. Hasanuzzaman et al. (2018) found that eggplant varieties with lower contents of nitrogen, glucose and aminoacids and higher contents of phenolics were less susceptible to B. tabaci. Leaf hopper (Amrasca devastans) resistance has been identified both in the cultivated and wild germplasm, associated to higher leaf hairiness and thin leaf lamina, as well as to higher content in phenolics and sugars (Ali et al. 2016). Also, sources of resistance to A. devastans have been identified in S. aethiopicum, S. insanum and S. violaceum (Warade et al. 2004). Certain levels of resistance or tolerance have been found against other insect pests affecting eggplant, such as thrips, aphids, or spotted beetle has been identified. Regarding spider mites, differences among accessions in susceptibility have been described, and high levels of resistance against the two-spotted spider mite (Tetranychus urticae) have been identified in S. dasyphyllum, S. macrocarpon, S. sisymbriifolium, and S. torvum (Schaff et al. 1982; Taher et al. 2020).
A few germplasm characterizations have been performed for the identification of sources of tolerance to abiotic stresses such as drought, salinity, or extreme temperatures. Although no large screenings have been performed, some differences among eggplant cultivars have been found for tolerance to drought or salinity (Hanachi et al. 2009; Saracanlao et al. 2016; Tani et al. 2018; Kiran et al. 2019; Plazas et al. 2019). Some wild species have proved to be more tolerant than the cultivated species. In this way, S. insanum and S. torvum have been found to be more tolerant to salinity than S. melongena and this may be associated to a higher accumulation of proline as well as of the ions Na+ and Cl− (Brenes et al. 2020a, b). García-Fortea et al. (2019) also been found S. elaeagnifolium, a particularly drought tolerant species, had a root system that explored a larger volumen than that of S. melongena. Although data on tolerance to stresses is lacking for many wild eggplant relatives, some of them grow in highly stressful environments indicating that they are highly tolerant to the stresses they suffer (Vorontsova and Knapp 2016). In this way, S. incanum, which grows in desertic and semi-desertic areas, has been identified as highly promising for breeding for tolerance to drought (Gramazio et al. 2017a, b). Kouassi et al. (2021) found that S. sisymbriifolium and the interspecific hybrids of S. melongena with S. anguivi, S. dasyphyllum and S. insanum were tolerant to drought, being the interspecific hybrids heterotic for the tolerance to this abiotic stress. Differences in eggplant cultivars have been observed for tolerance to low temperatures (Boyaci et al. 2009; Yang et al. 2020), and some wild species such as S. aculeatissimum, S. grandiflorum and S. mammosum have been reported as cold tolerant (Toppino et al. 2021). Differences among eggplant cultivars have been observed for tolerance to high temperatures, with several promising varieties having been identified (Santhiya et al. 2019).
4.8.2 Marker-Assisted Gene Introgression
The success of marker-assisted gene introgression of the resistance or tolerance to abiotic stresses depends on the availability of markers closely linked to the gene/s that have to be introgressed. The success of introgression of the trait also depends on the genetic control and the expression of the gene/s in the recipient genetic background. Genetic analysis of resistance to diseases has revealed different patterns of inheritance to the major eggplant diseases. In this way, for bacterial wilt resistance, different mechanisms have been found including, monogenic and polygenic inheritance, with different gene action mechanisms and interactions (Barik et al. 2020). Through the use of segregating generations some molecular markers have been identified associated to resistance to R. solanacearum. In this way, Bi-hao et al. developed a SCAR marker associated to a dominant resistance to bacterial wilt from resistant accession E-31. Lebeau et al. (2013) also found a major dominant gene (Ers1, subsequenlty renamed EBRW9) for resistance against three strains of R. solanacearum from accession AG91-25, which has been positioned in LG9 (Salgon et al. 2017). More recently, Salgon et al. (2018) detected several QTLs associated to resistance to phylotypes I and III coming from accession EG203, although the expression of the QTLs was highly influenced by environmental conditions. Despite the availability of these markers, there are no reports of introgression of resistance to bacterial wilt in eggplant élite genetic backgrounds. In the case of Fusarium wilt resistance, a major dominant resistance gene derived from S. aethiopicum (Rfo-sa1) has been introgressed by Toppino et al. (2008) in the genetic background of cultivated eggplant. Several molecular markers, such as CAPS (Toppino et al. 2008), associated to Rfo-sfa1, which maps in chromosome 2 (Barchi et al. 2018), are available. Using a recombinant inbred line (RIL) population using a parent with resistance introgressed from S. aethiopicum, Barchi et al. identified a major QTL cosegregating with Rfo-sfa1. Apart from this major QTL, these authors also identified a minor QTL, accounting for 11% of the variation, in chromosome 11. By using a bulked segregant analysis (BSA) strategy, Mutlu et al. (2008) also developed SCAR markers linked to a dominant gene of resistance derived from the eggplant resistant line LS2436. This resistance has been introgressed into eggplant (Boyaci et al. 2020). In another study (Miyatake et al. 2016), using the two eggplant resistant varieties, one of which is LS2436 (as in Mutlu et al. 2008), found two resistance alleles (Fm1L and Fm1E) that mapped in the same genomic region asRfo-sa1. In addition, Miyatake et al. (2016) also found an additional QTL in chromosome 4 derived from LS2436. Regarding Verticillium wilt, the high levels of resistance to this disease found in S. linnaeanum have been introgressed into the eggplant genetic background by Acciarri et al. (2004) and Liu (2015). Sunseri et al. (2003) using AFLP markers found two tentative QTLs for resistance to Verticillium wilt in segregating populations using the same source of resistance than Acciarri et al. (2004). Liu et al. (2015) found that the resistance to Verticillium of the S. linnaeanum accession PI388846 could be selected with a molecular marker for the homolog of the tomato Ve resistance gene. In addition to the resistance introgressed from S. linnaeanum, Barchi et al. (2018) identifyied three QTLs for tolerance to Verticillium wilt in chromosomes 5, 8 and 9 in a RIL population. Interestingly, Barbierato et al. (2016) found that after inoculation with Fusarium oxysporum f. sp. melongenae, materials carrying the Rfo-sfa1 gene expressed improved tolerance to Verticillium wilt. Marker-assisted breeding to other biotic, either diseases or pests, and abiotic stresses is still in its infancy in eggplant and introgression of resistance into the eggplant genetic background has not been reported yet. However, the availability of introgression lines with wild species that may harbor genes for tolerance of stresses, such as the one with S. incanum (Gramazio et al. 2017a, b) may facilitate the identification of lines with tolerance to stresses introgressed from the donor wild species. Also, some genes involved in tolerance to stresses have been identified. For example, Zhou et al. (2018a) found a gene from the eggplant wild relative S. aculeatissimum potentially involved in resistance to the root-knot nematode M. incognita. Also, Li et al. (2019) found that the SmAKT1 K+ transporter gene contributed to higher tolerance to salinity in eggplant, while Zhou et al. (2018b) found that three C-repeat binding factor genes (SmCBF1, SmCBF2 and SmCBF3) were involved in tolerance to cold, drought and salinity in eggplant.
To our knowledge, no gene pyramidimg works have been performed aimed at developing eggplant varieties with tolerance to several eggplant biotic and abiotic stresses. However, the availability of markers for genes and QTLs associated to some of these traits would facilitate this task. Similarly, hybrids resistant or tolerant to several stresses could be easily obtained in eggplant by crossing complementary parents for the resistance or tolerance (Sidhu et al. 2005).
4.9 Map-Based Cloning of Resistance Genes
Several cultivated accessions and varieties and wild relatives have traits useful for breeding new and robust eggplant varieties. Traditionally this is done by hybridization and backcrossing methods. This works as long as the species are cross compatible and time allows. Gene editing and cloning can speed up the process, and allow introduction of genes from species further away in the taxonomy system. Still there are limitations, especially when it comes to risks and public acceptance. Several genes have been identified, characterized and cloned, and these are especially genes encoding for disease- and insect pest resistance (Table 4.2).
For root-knot nematode resistance, the gene SacMi was recently cloned (Zhou et al. (2018a, b). The full-length DNA is 4,014 bp and enhances the production for a protein of 1,338 amino acids. The gene has been cloned into S. aculeatissimum, where tobacco rattle virus was used as a vector, and from where the plasmids were transformed into Agrobacterium tumefaciens. Interspecific hybridization between S. aculeatissimum and S. melongena gives opportunity to utilize this, and other resistance genes, from S. aculeatissimum, through traditional breeding as well as further gene editing research. Another nematode resistance gene, but from tomato (Mi-1.2), has also been transferred to eggplant (Goggin et al. 2006). Furthermore, a modified rice cystatin gene, OC-IΔD86, which is controlling nematodes, has been introduced to eggplants (Papolu et al. 2016) but further research is needed before it can be taken into fields.
A Verticillium wilt resistant gene, Ve, has been isolated in the wild relative S. linnaeanum (accession number PI388846). Through hybridization and backcrossing it was introduced into eggplant (Liu et al. 2014). Another resistance mechanism is ove-expression of a yeast desaturase gene done through transgenic introduction (Xing and Chin 2000). The gene increases the production of 16:1 and 16:3 fatty acids that again inhibit the Verticillium wilt pathogen. Resistance genes to Verticillium wilt and Fusarium has also been introduced by expressing a glucanase gene from of alfalfa and a chitinase gene from rice, respectively (Singh et al. 2014). Furthermore, the gene Dm-AMP1 from a Dahlia species has been introduced to eggplant to increase the resistance to Botritys cinerea and some other fungi through a protein release via the root exudates (Turrini et al. 2004). Resistance to Alternaria solani has been introduced to eggplant by expressing a defensingene from wasabi.
Coming to insect pest resistance, a Bacillus thuringiensis (Bt) cry1Ac gene, which has been introduced into several important crops, has also been introduced into eggplant to reduce the damage from eggplant fruit and shoot borer (Leucinodes orbonalis) and other insect pests (Shelton et al. 2018). Modified eggplant varieties with the gene are now used in South Asia since 2014 (ISAAA 2019).
4.10 Genomics-Aided Breeding for Resistance Traits
4.10.1 Structural and Functional Genomic Resources Developed and Applications
The fact that eggplant is one of the most cultivated Solanaceae crop, which has received little attention open a huge range of possibilities for the scientific community. It has been observed that at least 40% of the most important agronomic traits of this crop are controlled by the same genes that have been previously described in other Solanaceaecrops such as potato, tomato and pepper (Doganlar et al. 2002b). This feature will be very useful for genotyping characters on genetic maps and for generating comparative maps between species.
The first interspecific linkage map developed in eggplant was constructed using an interspecific cross S. linneanum × S. melongena (Doganlar et al. 2002a). For the development of this map, markers previously described and located in tomato and potato was used. Several years later this map was improved by Wu et al. (2009). Later on, a new linkage map based on a different interspecific cross (S. melongena × S. incanum) was developed by Gramazio et al. (2014). In this map, the prickliness trait as well as the genes involved in the chlorogenic acid pathway and genes of the polyphenol oxidases were located in different chromosomes. A new intraspecific map was developed and used to anchor the genome sequence (Hirakawa et al. 2014). More recently, another linkage map developed using 114 RILs between the “Ramnagar Giant” eggplant variety and the wild species S. incanum “W-4”, locating 1443 polymorphic markers between the two species of diverse molecular nature (Mishra et al. 2020).
The first intraspecific map of eggplant was constructed by Nunome et al. (2001) and improved by Nunome et al. (2009). Subsequently, Barchi et al. (2010) developed two intraspecific mapping populations from the cross between the breeding lines ‘305E40’ and ‘67/3’. This map was improved using RAD-seq approach and is composed mainly of SNPs (Barchi et al. 2012). Finally, two high-density intraspecific genetic maps were developed using a cross between S. melongena MM738 and S. melongena AG91-25 (Salgon et al. 2017) and a double haploid population from the cross EG203 × MM738 (Salgon et al. 2018).
The genome-wide association mapping approach (GWA) was also implemented in eggplant. The first attempt was carried out by Ge et al. (2013a), whom analysed the association of several fruit traits. Taking advantage of the high-throughput SNP technologies, new association studies were performed by Cericola et al. (2014) and Portis et al. (2015). The authors were able to associate SNPs to several traits related to fruit, plant, and leaf morphology.
Some works have attempted to unravel the relationship between eggplant and some of its related species, seeking to find the reason for the resistance or tolerance present in wild species and related cultivated species (Lebeau et al. 2013; Toppino et al. 2008; Yang et al. 2014). QTLs related to fungal resistance have been found (Barchi et al. 2018) and also some rootstock cultivars were developed to be resistant to bacterial wilt (Rakha et al. 2020). The mechanisms involved in the development of the eggplant ovary when it is crossed with other cultivated species (S. aethiopicum) and with different wild species have been studied. Performing a comparative analysis of the transcriptome, more than 1600 transcription factors involved in interspecific hybridization have been located (Li et al. 2020). In addition, the mechanism that regulates anther dehiscence has been studied in depth. Using fertile and sterile eggplant accessions, relevant information has been obtained that makes it possible to clarify what reactions are taking place in the flower (Yuang et al. 2021).
4.10.2 Details of Genome Sequencing
The first version of the genome was published in 2014 by Hirakawa et al. (http://eggplant.kazusa.or.jp/) and it was a giant step in the improvement of the crop, since the improvement processes were going to accelerate, for the development of this genome an Asian type eggplant was used, elongated and purple. Despite this, this genome was just a first draft and only reached scaffold assembly level and only covers 70% of the genome.
The availability of the genome of the eggplant has allowed carrying out synteny studies with respect to other nightshades (Portis et al. 2015; Gramazio et al. 2016) of interest, such as tomato and potato. This has made possible to annotate a multitude of genes (more than 800 in the work by Barchi et al. 2019a), some of them previously described in otherrelated species, and to locate multiple translocations. In this sense, the potential of using wild species in breeding was observed (Acquadro et al. 2017). Wild relatives of eggplant separated into four clusters species that have been used successfully in breeding programs (Plazas et al. 2016).
The new version of the eggplant genome developed and published by Barchi et al. (2019b). This new genome was sequenced using a combination of Illumina and single molecule optical mappingreaching chromosome-anchored genome assembly. The annotation of the genome detected 34,916 high-quality protein-coding genes; whichis similar to the previously annotated Solanum genomes (http://www.eggplantmicrosatellite.org/). This genome information was used for resequencing seven eggplants and its wild relative S. incanum to develop useful breeding tools (Gramazio et al. 2019a, b).
A new genome is currently available, developed by Wei et al. (2020). Its coverage is around 91% compared to 74% of the genome developed in 2004 (Hirawaka et al. 2014). The HQ-1315 accession was used, obtaining a genome of 1.17 GB in size with more than 36,582 protein-coding genes. This study has made it possible to locate genes that control important crop traits. In addition, a QTL has been located on chromosome 3 involved in the length of the fruit, and more specifically, a gene from the SUN family (Smechr0301963) that regulates the length of the fruit in eggplant. In addition, more than 200 markers have been identified associated with 71 morphological and physiological characters (size and color of the fruit, leaf morphology and some nutritional components). Of these genes, at least 1009 are listed. Comparison of this new genome with the previous ones has made it possible to locate some variations and confirm that the eggplant is phylogenetically speaking, closer to the potato and tomato than to the pepper.
The currentgenome of the eggplant developed by Li et al. (2021) has found slightly fewer high-quality protein-coding genes. Despite this, one of its potentialities is that, within the specific genes of the family (646 genes), the genes related to the bacterial spot of the eggplant and the genes involved in the synthesis pathway of chlorogenic acid have been located. This feature makes this new genome a very useful tool for the scientific community.
All the phenomic and genomic information available has allowed a more efficient improvement in horticuoral traits. Shortening the development and adaptation times of new cultivars is a primary objective, since mainly due to climate change and the disproportionate increase in the world population, agriculture has had to accelerate its objectives. It took more than sevn years to develop markers associated with resistance to Fusarium (Toppino et al. 2008), however, it is possible to reduce these times with new information and technologies (Barchi et al. 2018).
4.10.3 Impact on Germplasm Characterization and Gene Discovery
At this time, having the phenotypic and genotypic information of the cultivated species in the same database would save a lot of time and work for the breeders (Raubach et al. 2020). Focusing on this objective, the “Germinate” database (https://ics.hutton.ac.uk/cwr/eggplant/#/) was developed, which is a repository that collects both the passport data of the material, genotypic data and phenotypic characterization information, among others. One of its most important uses is to be able to search for sources of resistance in related species, and thus improve the species more efficiently. Many times, that variability is present, but that information is unknown, so for germplasm banks to be more useful and accessible, they must have that information collected and easily accessed.
Throughout the process of developing eggplant materials with crop wild relative (CWR) introgressions, different evaluations have been made for biotic and abiotic stresses (Barchi et al. 2018; Brenes et al. 2020a, b; Kaushik et al. 2016; Kouassi et al. 2020). Some interspecific hybrids were found to be very vigorous and exhibited a powerful root system, which may explain greater tolerance to drought, as well as greater stem vigor when eggplant is grafted onto vigorous interspecific aubergine hybrids (Mangino et al., 2020). Therefore, the direct use of interspecific eggplant hybrids with wild relatives as rootstocks is promising (Somvanshi et al. 2020). Global warming, in addition to the multiple consequences it has had on crops, has accelerated the search for sources of resistance or tolerance. Trying to find eggplant accessions that are better adapted to very high temperatures, 315 upregulated genes and 342 downregulated genes have been located when different accessions of this cropwere subjected to stress due to high temperatures.
Selection that has traditionally been made towards the shape of the fruit has had a considerable impact on the genetic structure and diversity of the eggplant (Liu et al. 2019; Stagel et al. 2008), an impact that has been detected with unique changes distributed throughout the genome. The variability found in the Asian center of diversity (Miyatake et al. 2019) has been represented using almost 900 molecular markers and 893 eggplant accessions, from which a nuclear collection of 100 accessions was developed. The increase in cystatin represented a considerable advance in the development of rootstocks tolerant to nematodes (Papolu et al. 2016), as well as the silencing of the Mi-msp-1 gene (Chaudhary et al. 2019). These advances have occurred thanks to having the genome of the eggplant available (Chapman 2020).
4.11 Genetic Engineering
4.11.1 Brief on Genetic Engineering for Resistance Traits
Gene silencing (GS) is the switching off a certain gene expression in the plant cell through transcription or translation process that possibly developed as a molecular defense system against biotic stress (Waterhouse et al. 2001). Gene activity suppression in plant species can be controlled by various molecular and biochemical mechanisms such astranscriptional gene silencing (TGS), virus-induced gene silencing (VIGS), RNA interference (RNAi) and micro RNAs. Transgenic plants may be released as a result to manipulation of gene silencing approach that switch off genes of interest. These indispensable strategieshaveopened new insights in front of plant scientists (breeders, pathologist, entomologist, agronomist, etc.) forcrop improvement as well as to accelerate and facilitate plant breeding programs in model and non-model species. Moreover, GS technology has been significantly applied in different research objectives like resistance to biotic stress (Scorza et al. 2001), food quality (Ogita et al. 2003), protein value in tomato and maize (Segal et al. 2003), quantity and quality of forest trees. In eggplant, VIGS tool has been successfully used as an authoritative technology regarding gene function investigation. To accomplish VIGS tool, a lot of vectors have been widely utilized for functional genomics analysis like Tomato golden mosaic virus (TGMV), Tobacco mosaicvirus (TMV), Potato virus X (PVX) and Tobacco rattle virus (TRV) where TRV is particularly excessively applied in Solanaceae species (Fu et al. 2005). Interestingly, four genes PDS, Chl H, Su (Sulfur), CLA1 were silenced in eggplant by VIGS (Liu et al. 2012). Wang and Fu (2018) applied VIGS approach to chalcone synthase gene (SmCHS) expression through fruit ripening process of eggplant where CHS gene expression was related to a negative gravitropic response.Unfortunately, the gene silencing research on eggplant is lack, thus many investigations are needed in the future regarding an array of horticultural traits.
4.11.2 Bt Eggplant
As indicated above, the eggplant fruit and shoot borer (Leucinodes orbonalis) is a major constraint to eggplant production in Southeast Asia. An alternative to conventional plant breeding against this pest is the development of transgenic plants expressing the Bacillus thuringiensiscry1Ac gene (Shelton et al. 2018). This strategy has proved as very efficient in other crops, such as cotton or maize, where other Lepidoptera borers are major pests (Hautea et al. 2016). The Indian company Maharashtra Hybrid Seeds Co. Pvt. Ltd. (Mahyco) transformed eggplant with the cry1Ac gene under the control of the constitutive 35S CaMV promoter and the transformation event called EE-1, which proved successful to control the eggplant fruit and shoot borer, has been introduced in several lines and hybrids (Shelton et al. 2018). While in India the cultivation of Bt eggplant was cleared for commercial cultivation in 2009, since then it is under a moratorium (Hautea et al. 2016). However, in Bangladesh commercial cultivation of several Bt eggplant varieties were approved and nowadays there are over 1,000 ha cultivated of this transgenic eggplant with excellent results in terms of yield and revenue, which was 19.7% and 21.7% higher compared to non-Bt varieties (Shelton et al. 2020), demonstrating the high potential of transgenic eggplant for dealing with pests such as the eggplant fruit and shoot borer.
4.12 Gene Stacking
Gene stacking is the process of combining two or more desirable genes (multigene) into a single crop plant. The newly produced traits from this strategy can be called as stacked traits and the crop species is known as biotech stacked (Singh et al. 2018). Genetic variability plays a crucial role in crop improvement, breeding programs and gene pyramiding. The source of variation may be derived from landraces, wild species, RILs, etc. that support plant breeders to generate new cultivars and hybrids with genes of interest through interspecific or intergeneric hybridization. Biotech or stacked cultivars have advantages more than mono-trait varieties and theyallow farmers to increase their crop production and overcoming the biotic (insects, diseases, weeds) and abiotic (high temperature, salinity, drought) problems. Gene stacking can be accomplished via two main approaches: hybrid stacking and molecular stacking. Hybrid stacking aaproachcan be conducted by hybridization between the two parental lines that possess multi-agronomic traits of interestin order to introduce new populations with desirable traits. Although, the aforementioned tool is simplest and earliest method of developing stacks, however there are some drawbacks like high cost, time consuming, long process, selectable markers requiredand difficult to gain homozygous crop plants for all transgenes. In molecular stacking approach, all genes of interest can be inserted into the target plant jointly or sequentially using Agrobacterium mediated transformation as a standard transfer technique or biolistic method (Que et al. 2010).In eggplant, gene pyramiding strategy can be applied for crop improvement where transgenic plants with a modified rice cystatin and root-knot nematodes resistance were developed (Papolu et al. 2016). Interestingly, many future advanced studies that associated with gene stacking to improve quality, quantity, and biotic and abiotic stress resistance of eggplant particularly in omics era are indispensable.
4.13 Protection of Eggplant Plant Material
Usually, plant breeders exert their intelectual property rights by protecting the eggplant varieties they developed according to the International Union for the Protection of New Varieties of Plants (UPOV) requirements (Sanderson 2017). This is usually done by filing an individual application for each variety with the authorities of UPOV members granting breeders’ rights. New varieties have to conform to the Disctinctness, Uniformity, and Stability (DUS) criteria according to the descriptors used for evaluation of eggplant, which in the case of the UPOV guidelines for eggplant include 43 morphological descriptors (UPOV 2002). New eggplant varieties can have different genetic constitutions, typically being lines or hybrids and nowadays, only in the European Union there are 405 registered eggplant varieties (European Commission 2020). An special case of protection in the European Union for landraces with distinctive features, is including them in the register of conservation varieties. In the case of eggplant, there is one eggplant conservation variety, which is the ‘Almagro’ eggplant, grown in a limited geographical area in the center of Spain, and used for pickling (Hurtado et al. 2014).
4.14 Future Perspectives
Eggplant is a major vegetable crop with an increasing global cultivation. However, eggplant is exposed to many abiotic and biotic stresses. Abiotic stress includes salinity, drought and high tempratuers, and biotic stress includes soil-borne diseases and many pests such as nematode, spider mite, whiteflies, aphids, eggplant fruit and shoot borer, leafhopper, and thrips, in particularly tropics and subtrobics (Taher et al. 2017, 2020). Eggplant wild relatives are richer than cultivated eggplant in genetic variations and could provide a great source for resistance to biotic and abiotic stresses (Daunay and Hazra 2012). Commercial varieties containing wild relative introgressions are not yet available and more research is required on using wild relatives in eggplant breeding. In the past, researchers had limited molecular markers to work with and could not easily introgress resistance genes into cultivated genetic backgrounds because of wild characteristics linked at other resistance loci. Now with functional genomics we have saturated markers, and such crossovers can be detected in each region. From that point forward, these genes could be easily deployed in pest resistant cultivars in the future. Of course if disease and insect resistance genes are combined in a single vaieity it would likely be a durable resistance package for biotic stress especially under climate change.
References
Acciarri N, Rotino GL, Sabatini E, Valentino D, Sunseri F et al (2004) Improvement of eggplants for resistance to Verticillium. In: Proceedings of 12th EUCARPIA meeting on genetics and breeding of capsicum and eggplant, 17–19 May 2004, Noordwijkerhout, The Netherlands, p 178
Acquadro A, Barchi L, Gramazio P, Portis E, Vilanova Set al (2017) Coding SNPs analysis highlights genetic relationships and evolution pattern in eggplant complexes. PLoS ONE12(7):e0180774
Alam SN, Rashid MA, Rouf FMA, Jhala RC, Patel JR et al (2003) Development of an integrated pest management strategy for eggplant fruit and shoot borer in South Asia. AVRDC—the World Vegetable Center, Tainan, Taiwan
Ali M, Ashfaq M, Ghaffar A, Bahtti AU, Ali A, Mubashar U (2016) Role of physio-morphic characters of different genotypes of eggplant, Solanum melongena L. and its association with the fluctuation of jassid, Amrasea biguttula biguttula (Ishida) population. Pak J Zool 48:1511–1515
Altinok HH, Can C, Boyaci HF, Topku V (2014) Genetic variability among breeding lines and cultivars of eggplant against Fusarium oxysporum f. sp melongenae from Turkey. Phytoparasitica 42:75–84
Álvarez MF, Mosquera T, Blair MW (2015) The use of association genetics approaches. Plant Breed Rev 38:17–68
Ano G, Hebert Y, Prior P, Messiaen C (1991) A new source of resistance to bacterial wilt of eggplants obtained from a cross: Solanum aethiopicum L. × Solanum melongena L. Agronomie 11:555–560
Arrones A, Vilanova S, Plazas M, Mangino G, Pascual L et al (2020) The dawn of the age of multi-parent MAGIC populations in plant breeding: novel powerful next-generation resources for genetic analysis and selection of recombinant elite material. Biology 9(8):229
AVRDC (1999) AVRDC 1998 Report. Asian Vegetable Research and Development Center, Tainan, Taiwan, pp 32–36
Barbierato V, Toppino L, Rinaldi P, Sala T, Bassolino L et al (2016) Phenotype and gene expression analyses of the Rfo-sa1 resistant aubergine interaction with Fusarium oxysporum f. sp. melongenae and Verticillium dahliae. Plant Pathol 5:1297–1309
Barchi L, Lanteri S, Portis E, Stàgel A, Valè G et al (2010) Segregation distortion and linkage analysis in eggplant (Solanum melongena L.). Genome 53(10):805–815
Barchi L, Lanteri S, Portis E, Valè G, Volante A et al (2012) A RAD tag derived marker based eggplant linkage map and the location of QTLs determining anthocyanin pigmentation. PLoS ONE 7(8):e43740
Barchi L, Rotino GL, Toppino L, Vale G, Acciarri N et al (2016) SNP mapping and identification of QTL for horticultural key breeding traits in eggplant (Solanum melongena L.). Acta Hortic 1145:9–16
Barchi L, Toppino L, Valentino D, Bassolino L, Portis E et al (2018) QTL analysis reveals new eggplant loci involved in resistance to fungal wilts. Euphytica 214(2):20
Barchi L, Pietrella M, Venturini L, Minio A, Toppino L et al (2019a) A chromosome-anchored eggplant genome sequence reveals key events in Solanaceae evolution. Sci Rep 9:11769
Barchi L, Acquadro A, Alonso D, Aprea G, Bassolino L et al (2019b) Single primer enrichment technology (SPET) for high-throughput genotyping in tomato and eggplant germplasm. Front Plant Sci 10:1005
Barik S, Reddy AC, Ponnam M, Kumari M, Acharya GC et al (2020) Breeding for bacterial wilt resistance in eggplant (Solanum melongena L.): Progress and prospects. Crop Protec 137:105270
Boiteux LS, Charchar JM (1996) Genetic resistant to root-knot nematode (Meloidogyne javanica) in eggplant (Solanum melongena). Plant Breed 115:198–200
Boshou L (2005) A broad review and perspective on breeding for resistance to bacterial wilt. In: Allen C, Prior P, Hayward AC (eds) Bacterial wilt disease and the Ralstonia solanacearum species complex. American Phytopathological Society, St. Paul, MN, USA, pp 225–238
Boyaci F, Oğuz A, Ünlü M, Denizer B, Abak K (2009) Growth, pollen quantity and quality and fruit characteristics of some parthenocarpic and non-parthenocarpic eggplants in unheated greenhouse. Acta Hort 807:239–244
Boyaci F, Unlu A, Abak K (2012) Screening for resistance to Fusarium wilt of some cultivated eggplants and wild Solanum accessions. Acta Hort 935:23–27
Boyaci HF, Topcu V, Tepe A, Yildirim IK, Oten M, Aktas A (2015) Morphological and molecular characterization and relationships of Turkish local eggplant heirlooms. Not Bot Hort Agrobot Cluj Napoca 43:100–107
Boyaci HF, Prohens J, Unlu A, Gumrukcu E, Oten M, Plazas M (2020) Association of heterotic groups with morphological relationships and general combining ability in eggplant. Agriculture 10:203
Brenes M, Pérez J, González-Orenga S, Solana A, Boscaiu M et al (2020a)Comparative studies on the physiological and biochemical responses to salt stress of eggplant (Solanum melongena) and its rootstock S. torvum. Agriculture 10:328
Brenes M, Solana A, Boscaiu M, Fita A, Vicente O et al(2020b) Physiological and biochemical responses to salt stress in cultivated eggplant (Solanum melongena) and in S. insanum L., a close wild relative. Agronomy 10:651
Brooks J, Blandford D (2019) Policies for global food security. Global Challenges Future Food Agric Policies 1:5
Cappelli C, Stravato VM, Rotino GL, Buonaurio R(1995) Sources of resistance among Solanum spp. to an Italian isolate of Fusarium oxysporum f. sp. melongenae. In: Proceedings of 9th EUCARPIA meeting on genetics and breeding of capsicum and eggplant, budapest 21–25 Aug 1995, Hungary, pp 221–224
Cericola F, Portis E, Lanteri S, Toppino L, Barchi L et al (2014) Linkage disequilibrium and genome-wide association analysis for anthocyanin pigmentation and fruit color in eggplant. BMC Genomics 15:896
Chadha ML (1993) Improvement of brinjal. In: Chadha KL, Kalloo G (eds) Advances in horticulture, vol 5. Vegetable crops: Part 1. Malholtra Publishing House, New Delhi, pp 105–135
Chakrabarti AK, Choudhury B (1975) Breeding brinjal resistant to little leaf disease. Proc Ind Natl Sci Acad B 41:379–385
Cham BE (2012) Intralesion and curaderm BEC5 topical combination therapies of solasodine rhamnosyl glycosides derived from the eggplant or Devil’s Apple result in rapid removal of large skin cancers. Methods of treatment compared. Int J Clin Med 3(2):115–124
Chapman MA (2020) Eggplant breeding and improvement for future climates. In: Kole C (ed) Genomic designing of climate-smart vegetable vrops. Springer, Cham, Switzerland, pp 257–276
Chaudhary S, Dutta TK, Tyagi N, ShivakumaraTN PPK (2019) Host-induced silencing of Mi-msp-1 confers resistance to root-knot nematode Meloidogyne incognita in eggplant. Transgen Res 28(3–4):327–340
Colak-Ates A, Fidan H, Ozarslandan A, Ata A (2018) Determination of the resistance of certain eggplant lines against Fusarium wilt, Potato Y potyvirusand root-knot nematode using molecular and classic markers. Fresenius Environ Bull 27:7446–7453
Collonnier C, Fock I, Kashyap V, Rotino G, Daunay M et al (2001) Applications of biotechnology in eggplant. Plant Cell Tiss Org Cult 65(2):91–107
Daunay MC, Janick J (2007) History and iconography of eggplant. Chron Hort 47(3):16–22
Daunay MC, Hazra P (2012) Eggplant. In: Peter KV, Hazra P (eds) Handbook of vegetables. Studium Press, Houston, TX, USA, pp 257–322
Daunay MC, Salinier J, Aubriot X (2019) Crossability and diversity of eggplants and their wild relatives. In: Chapman M (ed) The Eggplant genome. Springer, New York, NY, USA, pp 135–191
Doganlar S, Frary A, Daunay MC, Lester RN, Tanksley SD (2002a) A comparative genetic linkage map of eggplant (Solanum melongena) and its implications for genome evolution in the Solanaceae. Genetics 161(4):1697–1711
Doganlar S, Frary A, Daunay MC, Lester RN, Tanksley SD (2002b) Conservation of gene function in the Solanaceae as revealed by comparative mapping of domestication traits in eggplant. Genetics 161(4):1713–1726
Doshi KM (2004) Influence of biochemical factors on the incidence of shoot and fruit borer infestation in eggplant. Capsicum Eggplant Newsl 23:145–148
Du L, Bao C, Hu T, Zhu Q, Hu H et al (2016) SmARF8, a transcription factor involved in parthenocarpy in eggplant. Mol Genet Genom 291(1):93–105
European Commission (2020) EU plant variety database. https://ec.europa.eu/food/plant/plant_propagation_material/plant_variety_catalogues_databases/search/public/index.cfm. Accessed 8 November 2020
Everitt B (1998) The Cambridge dictionary of statistics. Cambridge University Press, Cambridge, UK
FAO (2017) FAOSTAT Production databases. Available online at: http://www.faostat.fao.org
Fu DQ, Zhu BZ, Zhu HL, Jiang WB, Luo YB (2005) Virus-induced gene silencing in tomato fruit. Plant J43:299–308
Furini A, Wunder J (2004) Analysis of eggplant (Solanum melongena)-related germplasm: morphological and AFLP data contribute to phylogenetic interpretations and germplasm utilization. Theor Appl Genet 108:197–208
García-Fortea E, Gramazio P, Vilanova S, Fita A, Mangino G et al (2019) First successful backcrossing towards eggplant (Solanum melongena) of a New World species, the silverleaf nightshade (S. elaeagnifolium) and characterization of interspecific hybrids and backcrosses. Sci Hort 246:563–573
García-Mendivil HA, Escudero N, Sorribas FJ (2019) Host suitability of Solanum torvum cultivars to Meloidogyne incognita and M. javanica and population dynamics. Plant Pathol 68:1215–1224
Ge H, Liu Y, Jiang M, Zhang J, Han H, Chen H (2013a) Analysis of genetic diversity and structure of eggplant populations (Solanum melongena L.) in China using simple sequence repeat markers. Sci Hort 162:71–75
Ge HY, Liu Y, Zhang J, Han HQ, Li HZ et al (2013b) Simple sequence repeat-based association analysis of fruit traits in eggplant (Solanum melongena). Genet Mol Res 12:5651–5663
Gebhardt C (2016) The historical role of species from the Solanaceae plant family in genetic research. Theor Appl Genet 129:2281–2294
Genin S, Denny TP (2012) Pathogenomics of the Ralstonia solanacearum Species Complex. Annu Rev Phytopathol 50:67–89
Goggin FL, Jia L, Shah G, Hebert S, Williamson VM, Ullman DE (2006) Heterologous expression of the Mi-1.2 gene from tomato confers resistance against nematodes but not Aphids in Eggplant. Mol Plant-Microbe Interact 19(4):383–388
Goode EL (2011) Linkage disequilibrium. In: Schwab M (ed) Encyclopedia of cancer. Springer, Berlin/Heidelberg, Germany, pp 2043–2048
Gramazio P, Prohens J, Plazas M, Andùjar I, Herraiz FJ et al (2014) Location of chlorogenic acid biosynthesis pathway and polyphenol oxidase genes in a new interspecific anchored linkage map of eggplant. BMC Plant Biol 14:350
Gramazio P, Blanca J, Ziarsolo P, Herraiz FJ, Plazas M et al (2016) Transcriptome analysis and molecular marker discovery in Solanum incanum and S. aethiopicum, two close relatives of the common eggplant (Solanum meleongena) with interest for breeding. BMC Genomics 17:300
Gramazio P, Prohens J, Plazas M, Mangino G, Herraiz FJ, Vilanova S (2017a) Development and genetic characterization of advanced backcross materials and an introgression line population of Solanum incanum in a S. melongena background. Front Plant Sci 8:1477
Gramazio P, Prohens J, Borràs D, Plazas M, Herraiz FJ, Vilanova S (2017b) Comparison of transcriptome-derived simple sequence repeat (SSR) and single nucleotide polymorphism (SNP) markers for genetic fingerprinting, diversity evaluation, and establishment of relationships in eggplants. Euphytica 213:264
Gramazio P, Chatziefstratiou E, Petropoulos C, Chioti V, Mylona P et al (2019a) Multi-level characterization of eggplant accessions from Greek islands and the mainland contributes to the enhancement and conservation of this germplasm and reveals a large diversity and signatures of differentiation between both origins. Agronomy 9:887
Gramazio P, Yan H, Hasing T, Vilanova S, Prohens J, Bombarely A (2019b) Whole-genome resequencing of seven eggplant (Solanum melongena) and one wild relative (S. incanum) accessions provides new insights and breeding tools for eggplant enhancement. Front Plant Sci 10:1220
Hanachi S, van Labeke MC, Mehouachi T (2009) Application of chlorophyll fluorescence to screen eggplant (Solanum melongena L.) cultivars for salt tolerance. Photosynthetica 52:57–62
Harlan JR, de Wet JMJ (1971) Toward a rational classification of cultivated plants. Taxon 20:509–517
Hasanuzzaman ATM, Islam MN, Liu FH, Cao HH, Liu TX (2018) Leaf chemical composition of different eggplant varieties affect performance of Bemisia tabaci (Hemiptera: Aleyrodidae) nymphs and adults. J Econ Entomol 111:445–453
Hautea DM, Taylo LD, Masanga APL, Sison MLJ, Narciso JO et al (2016) Field performance of Bt eggplants (Solanum melongena L.) in the Philippines: Cry1Ac expression and control of the eggplant fruit and shoot borer (Leucionodes orbonalis Guenée). PLoS One 11:e0157498
Hirakawa H, Shirasawa K, Miyatake K, Nunome T, Negoro S et al (2014) Draft genome sequence of eggplant (Solanum melongena L.): the representative Solanum species indigenous to the old world. DNA Res 21(6):649–660
Huet G (2014) Breeding for resistances to Ralstonia solanacearum. Front Plant Sci 5:715
Hurtado M, Vilanova S, Plazas M, Gramazio P, Fonseka HH et al (2012) Diversity and relationships of eggplants from three geographically distant secondary centers of diversity. PLoS ONE 7:e41748
Hurtado M, Vilanova S, Plazas M, Gramazio P, Herraiz FJ et al (2013) Phenomics of fruit shape in eggplant (Solanum melongena L.) using tomato analyzer software. Sci Hort 164:625–632
Hurtado M, Vilanova S, Plazas M, Gramazio P, Andújar I et al (2014) Enhancing conservation and use of local vegetables: the Almagro eggplant (Solanum melongena) case study. Genet Resour Crop Evol 61:787–795
International Board for Plant Genetic Resources (IBPGR) (1990) Descriptors for eggplant. Italy, Rome
ISAAA (2019) ISAAA's GM Approval Database. Available online at: www.isaaa.org/gmapprovaldatabase (Accessed February 5, 2019)
Iwata S, Isshiki N, Khan MMR (2008) ISSR variations in eggplant (Solanum melongena L.) and related Solanum species. Sci Hort 117:186–190
Joiret M, John JMM, Gusareva ES, van Steen K (2019) Confounding of linkage disequilibrium patterns in large scale DNA based gene-gene interaction studies. BioData Min 12(1):11
Kalloo G (1993) Eggplant (Solanum melongena). In: Kalloo G (ed) Genetic improvement of vegetable crops. Pergamon Press, Oxford, UK, pp 587–604
Karihaloo JL, Brauner S, Gottlieb LD (1995) Random amplified polymorphic DNA variation in the eggplant, Solanum melongena L. (Solanaceae). Theor Appl Genet 90:767–770
Kaushik P, Prohens J, Vilanova S, Gramazio P, Plazas M (2016) Phenotyping of eggplant wild relatives and interspecific hybrids with conventional and phenomics descriptors provides insight for their potential utilization in breeding. Front Plant Sci 7:677
Keatinge JDH, Lin LJ, Ebert AW, Chen WY, Hughes JDA et al (2014) Overcoming biotic and abiotic stresses in the solanaceae through grafting: current status and future perspectives. Biol Agri Hort 30:272–287
Kiran S, Kuşvuran S, Özkay F, Ellįaltioğlu SS (2019) Change in physiological and biochemical parameters under drought stress in salt-tolerant and salt-susceptible eggplant gentoypes. Turk J Agri Forest 43:593–602
Knapp S, Vorontsova MS, Prohens J (2013) Wild relatives of the eggplant (Solanum melongena L.: Solanaceae): new understanding of species names in a complex group. PLoS ONE 8:e57039
Kouassi B, Prohens J, Gramazio P, Kouassi AB, Vilanova S et al (2016) Development of backcross generations and new interspecific hybrid combinations for introgression breeding in eggplant (Solanum melongena). Sci Hort 213:199–207
Kouassi AB, Kouassi KBA, Sylla Z, Plazas M, Fonseka RM et al (2021) Genetic parameters of drought tolerance for agromorphological traits in eggplant, wild relatives, and interspecific hybrids. Crop Sci 61:55–68
Kumar M, Madhupriya RGP (2017) Molecular characterization, vector identification and sources of phytoplasmas associated with brinjal little leaf disease in India. Biotechnology 7:7. https://doi.org/10.1007/s13205-017-0616-x
Kumari S, Nagendran K, Rai AB, Singh B, Rao GP, Bertaccini A (2019) Global status of phytoplasma diseases in vegetable crops. Front Microbiol 10:1349
Lebeau A, Daunay MC, Frary A, Palloix A, Wang JF et al (2011) Bacterial wilt resistance in tomato, pepper, and eggplant: genetic resources respond to diverse strains in the Ralstonia solanacearum complex. Phytopathology 101:154–165
Lebeau A, Gouy M, Daunay MC, Wicker E, Chiroleu F et al (2013) Genetic mapping of a major dominant gene for resistance to Ralstonia solanacearum in eggplant. Theor Appl Genet 126(1):143–158
Lester RN, Niakan L (1986) Origin and domestication of the scarlet eggplant, Solanum aethiopicum, from S. anguivi in Africa. In: D’Arcy WG (ed) Solanaceae: biologyand systematics. Columbia University Press, New York, USA, pp 433–456
Lester RN, Hasan SMZ (1991) Origin and domestication of the brinjal eggplant, Solanum melongena, from S. incanum, in Africa and Asia. In: Hawkes JG, Lester RN, Nee M, Estrada N (eds) Solanaceae III: taxonomy, chemistry, evolution. Royal Botanic Gardens Kew, Richmond, UK, pp 369–387
Lester RN, Daunay MC (2003) Diversity of African vegetable Solanum species and its implications for a better understanding of plant domestication. Schriften Genetischen Ressour 22:137–152
Lester RN, Jaeger PM, Child A (2011) Solanum in Africa. Celia Lester, Birmingham, UK
Li D, Li S, Li W, Liu A, Jiang Y et al (2020) Comparative transcriptome analysis provides insights into the molecular mechanism underlying double fertilization between self-crossed Solanum melongena and that hybridized with Solanum aethiopicum. PLoS ONE 15(8):e0235962
Li D, Qian J, Li W, Yu N, Gan G et al (2021) A high-quality genome assembly of the eggplant provides insights into the molecular basis of disease resistance and chlorogenic acid synthesis. Mol Ecol Resour 21(4):1274–1286
Li H, Chen H, Zhuang T, Chen J (2010) Analysis of genetic variation in eggplant and related Solanum species using sequence-related amplified polymorphism markers. Sci Hort 125:19–24
Li J, Gao Z, Zhou L, Li LZ, Zhang JH et al (2019) Comparative transcriptome analysis reveals K+ transporter gene contributing to salt tolerance in eggplant. BMC Plant Biol 19:67
Li X, Liu F, Zhang Y, Li Y, Chen Y, Lian Y (2012) Research progress on eggplant parthenocarpy. China Vegetab 6:8–14
Liang C, Jayawardena RS, Zhang W, Wang X, Liu M et al (2016) Identification and characterization of Pseudocercospora species causing grapevine leaf spot in China. J Phytopathol 164:75–85
Liu H, Fu D, Zhu B, Yan H, Shen X et al (2012) Virus-induced gene silencing in eggplant (Solanum melongena). J Integr Plant Biol 54(6):422–429
Liu J, Zhou XH, Feng C, Wu W, Zhuang Y (2014) The function confirmation of Ve gene homolog in S. linnaeanum related to Verticillium wilt resistance by VIGS. Acta Agri Boreali-Sin 29:217–221
Liu J, Zheng Z, Zhou X, Feng C, Zhuang Y (2015) Improving the resistance of eggplant Solanum melongena to Verticillium wilt using wild species Solanum linnaeanum. Euphytica 2013:463–469
Liu J, Yang Y, Zhou X, Bao S, Zhuang Y (2018) Genetic diversity and population structure of worldwide eggplant (Solanum melongena L.) germplasm using SSR markers. Genet Resour Crop Evol 65:1663–1670
Liu N, Zhou B, Zhao X, Lu B, Li Y, Hao J (2009) Grafting eggplant onto tomato rootstock to suppress Verticillium dahliae infection: the effect of root exudates. Hort Sci 44(7):2058–2062
Liu W, Qian Z, Zhang J, Yang J, Wu M et al (2019) Impact of fruit shape selection on genetic structure and diversity uncovered from genome-wide perfect SNPs genotyping in eggplant. Mol Breed 39(11):140
Mace ES, Lester RN, Gebhardt GG (1999) AFLP analysis of genetic relationships among the cultivated eggplant, Solanum melongena L., and wild relatives (Solanaceae). Theor Appl Genet 99:626–633
Maestrelli A, Scalzo RL, Rotino GL, Acciarri N, SpenaA et al (2003) Freezing effect on some quality parameters of transgenic parthenocarpic eggplants. J Food Eng 56(2–3):285–287
Mangino G, Plazas M, Vilanova S, Prohens J, Gramazio P (2020) Performance of a set of eggplant (Solanum melongena) lines with introgressions from its wild relative S. incanum under open field and screenhouse conditions and detection of QTLs. Agronomy 10(4):467
Meyer RS, KarolKG LDP, Nee MH, Litt A (2012) Phylogeographic relationships among Asian eggplants and new perspectives on eggplant domestication. Mol Phylogenet Evol 63:685–701
Meyer RS, Whitaker BD, Little DP, Wu SB, Kennelly EJ, Long CL (2015) Parallel reductions in phenolic constituents resulting from the domestication of eggplant. Phytochemistry 115:194–206
Mishra P, Tiwari SK, Kashyap SP, Tiwari KN, Singh M, Singh B (2020) High-density genetic linkage map based on arbitrary and microsatellite markers using inter-specific recombinant inbred lines in eggplant (Solanum melongena L.). J Plant Biochem Biotechnol 29(3):427–438.
Miyatake K, Saito T, Negoro S, Yamaguchi H, Nunome T et al (2012) Development of selective markers linked to a major QTL for parthenocarpy in eggplant (Solanum melongena L.). Theor Appl Genet 124(8):1403–1413
Miyatake K, Saito T, Negoro S, Yamaguchi H, Nunome T et al (2016) Detailed mapping of a resistance locus against Fusarium wilt in cultivated eggplant (Solanum melongena). Theor Appl Genet 129(2):357–367
Miyatake K, Shinmura Y, Matsunaga H, Fukuoka H, Saito T (2019) Construction of a core collection of eggplant (Solanum melongena L.) based on genome-wide SNP and SSR genotypes. Breed Sci 69(3):498–502
Miyatake K, Saito T, Nunome T, Yamaguchi H, Negoro S et al (2020) Fine mapping of a major locus representing the lack of prickles in eggplant revealed the availability of a 0.5-kb insertion/deletion for marker-assisted selection. Breed Sci 70(4):438–448
Muñoz-Falcón JE, Vilanova S, Prohens J, Nuez F (2009a) Comparison of morphological, AFLP and SSR markers for the protection of eggplant germplasm. In: International symposium on seed, transplant and stand establishment of horticultural crops 898:123–131
Muñoz-Falcón JE, Prohens J, Vilanova S, Nuez F (2009b) Diversity in commercial varieties and landraces of black eggplants and implications for broadening the breeders’ gene pool. Ann Appl Biol 154:453–465
Muñoz-Falcón JE, Vilanova S, Plazas M, Prohens J (2011) Diversity, relationships, and genetic fingerprinting of the Listada de Gandía eggplant landrace using genomic SSRs and EST-SSRs. Sci Hort 129:238–246
Mutlu N, Boyacı FH, Göçmen M, Abak K (2008) Development of SRAP, SRAP-RGA, RAPD and SCAR markers linked with a Fusarium wilt resistance gene in eggplant. Theor Appl Genet 117(8):1303
Namisy A, Chen JR, Prohens J, MetwallyE EM, Rakha M (2019) Screening cultivated eggplant and wild relatives for resistance to bacterial wilt (Ralstonia solanacearum). Agriculture 9:157
Nunome T, Ishiguro K, Yoshida T, Hirai M (2001) Mapping of fruit shape and color development traits in eggplant (Solanum melongena L.) based on RAPD and AFLP markers. Breed Sci 51(1):19–26
Nunome T, Negoro S, Kono I, Kanamori H, Miyatake K et al (2009) Development of SSR markers derived from SSR-enriched genomic library of eggplant (Solanum melongena L.). Theor Appl Genet 119:1143–1153
Oboh G, Ekperigin MM, Kazeem MI (2005) Nutritional and haemolytic properties of eggplants (Solanum macrocarpon) leaves. J Food Compos Anal 18:153–160
Ogita S, Uefuji H, Yamaguchi Y, Koizumi N, Sano H (2003) RNA interference: producing decaffeinated coffee plants. Nature 423:823
Papolu PK, Dutta TK, Tyagi N, Urwin PE, Lilley CJ, Rao U (2016) Expression of a cystatin transgene in eggplant provides resistance to root-knot nematode. Meloido Gyneincognita. Front Plant Sci 7:1122
Parker BL, Talekar NS, Skinner M (1995) Field guide: Insect pests of selected vegetables in tropical and subtropical Asia. Asian Vegetable Research and Development Center, Shanhua, Tainan, Taiwan
Plazas M, Lopez Gresa MP, Vilanova S, Torres C, Hurtado M et al (2013) Diversity and relationships in key traits for functional and apparent quality in a collection of eggplant: fruit phenolics content, antioxidant activity, polyphenol oxidase activity, and browning. J Agric Food Chem 61:8871–8879
Plazas M, Vilanova S, Gramazio P, Rodríguez-Burruezo A, Fita A et al (2016) Interspecific hybridization between eggplant and wild relatives from different genepools. J Am Soc Hortic Sci 141:34–44
Plazas M, Nguyen HT, González-Orenga S, Fita A, Vicente O et al (2019) Comparative analysis of the responses to water stress in eggplant (Solanum melongena) cultivars. Plant Physiol Biochem 143:72–82
Portis E, Cericola F, Barchi L, Toppino L, Acciarri N et al (2015) Association mapping for fruit, plant and leaf morphology traits in eggplant. PLoS ONE 10:e0135200
Prasad TV, Bhardwaj R, Gangopadhyay KK, Arivalagan M, Bag MK et al (2014) Biophysical and biochemical basis of resistance to fruit and shoot borer (Leucionodes orbonalis Guennee) in eggplant. Ind J Hort 71:67–71
Prohens J, Blanca JM, Nuez F (2005) Morphological and molecular variation in a collection of eggplants from a secondary center of diversity: implications for conservation and breeding. J Amer Soc HortSci 130:54–63
Prohens J, Plazas M, Raigón MD, Seguí-Simarro JM, Stommel JR, Vilanova S (2012) Characterization of interspecific hybrids and first backcross generations from crosses between two cultivated eggplants (Solanum melongena and S. aethiopicum Kumba group) and implications for eggplant breeding. Euphytica 186:517–538
Prohens J, Gramazio P, Plazas M, Dempewolf H, Kilian B et al (2017) Introgressiomics: a new approach for using crop wild relatives in breeding for adaptation to climate change. Euphytica 213:158
Que Q, Chilton M, de Fontes C, He C, Nuccio M et al (2010) Trait stacking in transgenic plants—challenges and opportunities. GM Crops 1(4):220–229
Rafalski A, Morgante M (2004) Corn and humans: recombination and linkage disequilibrium in two genomes of similar size. Trends Genet 20(2):103–111
Rakha M, Namisy A, Chen JR, El-Mahrouk ME, Metwally E (2020) Development of interspecific hybrids between a cultivated eggplant resistant to bacterial wilt (Ralstonia solanacearum) and eggplant wild relatives for the development of rootstocks. Plants 9(10):1405
Rakha M, Hanson P, Ramasamy S (2017) Identification of resistance to Bemisia tabaci (Genn.) in closely related wild relatives of cultivated tomato based on trichome type analysis and choice and no-choice assays. Genet Resour Crop Evol 64:247–260
Ramakrishnan AP (2013) Linkage disequilibrium. In: Maloy S, Hughes K (eds) Brenner’s encyclopedia of genetics (Second Edition). Academic Press, pp 252–253
Ramasamy S (2009) Insect and mite pests on eggplant: A field guide for identification and management. AVRDC—The World Vegetable Center, Shanhua, Taiwan. AVRDC, pp 09–729
Ranil RHG, Prohens J, Aubriot X, Niran HML, Plazas M (2017)Solanum insanum L. (subgenus Leptostemonum Bitter, Solanaceae), the neglected wild progenitor of eggplant (S. melongena L.): a review of taxonomy, characteristics and uses aimed at its enhancement for improved eggplant breeding. Genet Resour Crop Evol 64(7):1707–1722
Rao GP, Mall S, Raj SK, Snehi SK (2010) Phytoplasma diseases affecting various plant species in India. Acta Phytopathol Entomol Hung 46:59–99
Rast ATB (1991) Screening germplasm of Solanum melongena for resistance to the eggplant strain of bell pepper mottle virus (BPMV) and other tobamoviruses. Capsicum Eggplant Newsl 10:26–32
Raubach S, Kilian B, Dreher K, Amri A, Bassi FM (2020) From bits to bites: Advancement of the germinate platform to support prebreeding informatics for crop wild relatives. Crop Sci 1–29
Ray D, Muelle N, West P, Foley J (2013) Yield trends are insufficient to double global crop production by 2050. PLoS ONE 8:e66428
Rodríguez GR, Moyseenko JB, Robbins MD, Huarachi Morejón N, Francis DM, van der Knaap E (2010) Tomato analyzer: a useful software application to collect accurate and detailed morphological and colorimetric data from two-dimensional objects. J vis Exp 37:e1856
Rotino GL, Sala T, Toppino L (2014) Eggplant. In: Pratap A, Kumar J (eds) Alien gene transfer in crop plants. Springer, New York, NY, pp 381–409
Sakata Y, Lester RN (1997)Chloroplast DNA diversity in brinjal eggplant (Solanummelongena L.) and related species. Euphytica 97:295
Salgon S, Raynal M, Lebon S, Baptiste JM, Daunay MC et al (2018) Genotyping by sequencing highlights a polygenic resistance to Ralstonia pseudosolanacearum in eggplant (Solanum melongena L.). Intl J Mol Sci 19:357
Salgon S, Jourda C, Sauvage C, Daunay M, Reynaud B (2017) Eggplant resistance to the Ralstonia solanacearum species complex involves both broad-spectrum and strain-specific quantitative trait loci. Front Plant Sci 8:828
Sanderson J (2017) Plants, people and practices—the nature and history of the UPOV convention. Cambridge University Press, Cambridge, UK
Santhiya S, Saha P, Tomar BS, Jaiswal S, Gopala KS (2019) Heat stress tolerance study in eggplant based on morphological and yield traits. Ind J Hort 76:691–700
Saracanlao RJR, Ocampo ETM, Canama AO, Manaday SJB, Maghirang RG, Delfin EF (2016) SSR-based genetic relationship in eggplant (Solanum melongena) genotypes with varying morphological response to drought. Philipp J Crop Sci Forest 41:57–64
Schaff DA, Jelenkovic G, Boyer CD, Pollack BL (1982) Hybridization and hybrid fertility of hybrid derivatives of Solanum melongena L. and Solanum macrocarpon L. Theor Appl Genet 62:149–153
Schuster DJ, Stansly PA, Polston JE (1996) Expressions of plant damage by Bemisia. In: Gerling D, Mayer RT (eds) Bemisia (1995) Taxonomy, biology, damage control and management. Intercept Andover, Hants, pp 153–165
Scorza R, Callahn A, Levy L, Damsteegt V, Webb K, Ravelonandro M (2001) Post-transcriptional gene silencing in plum pox virus resistant European plum containing the plum pox potyvirus coat protein gene. Transgen Res 10:201–209
Segal G, Song RT, Messing JA (2003) new opaque variant of maize by a single dominant RNA-interference-inducing transgene. Genetics 165:387–397
Shelton AM, Hossain MJ, Paranjape V, Azad AK, Rahman ML (2018) Bt eggplant Project in Bangladesh: history, present status, and future direction. Front Plant Sci 6:106
Shelton AM, Sarwer SH, Hossain MJ, Brookes G, Paranjape V (2020) Impact of Bt brinjal cultivation in the market value chain in five districts of Bangladesh. Front Bioengineer Biotechnol 8:498
Shivalingaswamy TM, Satpathy S (2007) Integrated pest management in vegetable crops. In: Jain PC, Bhargava MC (eds) Entomology: novel approaches. New India Publishing Agency, New Delhi, India, pp 353–375
Sidhu AS, Bal SS, Behera TK, Rani M (2005) An outlook in eggplant hybrid breeding. J New Seeds 6(2/3):15–29
Singh A, Singh M, Singh R, Kumar S, Kalloo G (2006) Genetic diversity within the genus Solanum (Solanaceae) as revealed by RAPD markers. Curr Sci 90:711–716
Singh D, Ambroise A, Haicour R, Sihachakr D, Rajam MV (2014) Increased resistance to fungal wilts in transgenic eggplant expressing alfalfa glucanase gene. Physiol Mol Biol Plants 20(2):143–150
Singh SK, Sahoo JP, Swain E (2018) A review on gene stacking in crop plant. J Pharmacog Phytochem 7(4):1862–1865
Somvanshi VS, Phani V, Banakar P, Chatterjee M, Budhwar Ret al (2020) Transcriptomic changes in the pre-parasitic juveniles of Meloidogyneincognita induced by silencing of effectors Mi-msp-1 and Mi-msp-20. 3 Biotech 10(8):360
Spellerberg IF, Fedor PJ (2003)A tribute to Claude Shannon (1916–2001) and a plea for more rigorous use of species richness, species diversity and the ‘Shannon–Wiener’Index. Glob Ecol Biogeogr 12 (3):177–179.
Stàgel A, Portis E, Toppino L, Rotino GL, Lanteri S (2008) Gene-based microsatellite development for mapping and phylogeny studies in eggplant. BMC Genomics 9(1):357
Sturtevant AH (1913) The linear arrangement of six sex-linked factors in Drosophila, as shown by their mode of association. J Exp Zool 14:43–59
Sunseri F, Sciancalepore A, Martelli G, Rotino GL, Acciarri N et al (2003) Development of RAPD-AFLP map of eggplant and improvement of tolerance to Verticillium Wilt. Acta Hort 625:107–115
Syfert MM, Castañeda-Álvarez NP, Khoury CK, Särkinen T, Sosa CC et al (2016) Crop wild relatives of the brinjal eggplant (Solanum melongena): poorly represented in genebanks and many species at risk of extinction. Amer J. Bot 103:635–651
Taher D, Rakha M, Ramasamy S, Solberg SØ, Schafleitner R (2019) Sources of Resistance for Two-spotted spider mite (Tetranychus urticae) in scarlet (Solanumaethiopicum L.) and gboma (S. macrocarpon L.) eggplant germplasms. Hort Sci 54:240–245
Taher D, Solberg SØ, Prohens J, Chou Y, Rakha M, Wu T (2017) World Vegetable Center ggplant collection: origin, composition, seed dissemination and utilization in breeding. Front Plant Sci 8:1484
Taher D, Ramasamy S, Prohens J, Rakha M (2020) Screening cultivated eggplant and wild relatives for resistance to sweetpotato whitefly (Bemisia tabaci) and to two-spotted spider mite (Tetranychus urticae). Euphytica 216:157
Talekar NS(2003) Development of an integrated pest management strategy for eggplant fruit and shoot borer in South Asia, Technical Bulletin TB28, AVRDC – The World Vegetable Center, Shanhua, Taiwan, p 66
Tani E, Kizis D, Markellou E, Papadakis I, Tsamadia D (2018) Cultivar-dependent responses of eggplant (Solanum melongena) to simultaneous Verticillium dahliae infection and drought. Front Plant Sci 9:1181
Thapa SP, Miyao EM, Michael Davis R, Coaker G (2015) Identification of QTLs controlling resistance to Pseudomonas syringaepv. tomato race 1 strains from the wild tomato, Solanum habrochaites LA1777. Theor Appl Genet 128(4):681–692
Toppino L, Barchi L, Lo Scalzo R, Palazzolo E, Francese G (2016) Mapping quantitative trait loci affecting biochemical and morphological fruit properties in eggplant (Solanum melongena L.). Front Plant Sci 7:256
Toppino L, Barchi L, Mercati F, Acciarri N, Perrone D et al (2020) A new intra-specific and high-resolution genetic map of eggplant based on a RIL population, and location of QTLs related to plant anthocyanin pigmentation and seed vigour. Genes 11:745
Toppino L, Prohens J, Rotino GL, Plazas M, Parisi M et al (2021) Pepper and eggplant genetic resources. In: Aversano R, Ercolano MR (eds) The wild solanums genomes; Carputo D. Springer, Cham, Switzerland, pp 119–154
Toppino L, Ribolzi S, Shaaf S, Bassolino L, Carletti G et al (2018) Development of an introgression lines population and genetic mapping of novel traits linked to key breeding traits in eggplant. In: Proceedings of the 62th SIGA congress Verona, Italy, 25–28 September, 1.07
Toppino L, Valè G, Rotino GL (2008) Inheritance of Fusarium wilt resistance introgressed from Solanum aethiopicum Gilo and Aculeatum groups into cultivated eggplant (S. melongena) and development of associated PCR-based markers. Mol Breed 22(2):237–250
Tümbilen Y, Frary A, Mutlu S, Doganlar S (2011) Genetic diversity in Turkish eggplant (Solanummelongena) varieties as determined by morphological and molecular analyses. Intl Res J Biotechnol 2:16–25
Turrini A, Sbrana C, Pitto L, Ruffini CM, Giorgetti L (2004) The antifungal Dm-AMP1 protein from Dahlia merckii expressed in Solanum melongena is released in root exudates and differentially affects pathogenic fungi and mycorrhizal symbiosis. The New Phytol 163(2):393–403
Tzortzakakis EA, Bletsos FA, Avgelis AD (2006) Evaluation of Solanum rootstock accessions for control of root-knot nematodes and tobamoviruses. J Plant Dis Protec 113:188–189
UPOV (2002) Guidelines for the conduct of tests for distinctness, uniformity and stability—eggplant (Solanum melongena L.). International Union for the Protection of New Varieties of Plants: Geneva, Switzerland
Vaghefi N, Pethybridge SJ, Shivas RG, Nelson SC (2016) Confirmation of Paracercospora egenula causing leaf spot of eggplant in Hawaii. Australas Plant Dis Notes 11:35
van der Weerden GM, Barendse GWM (2007) A web-searchable database developed for the EGGNET project and applied to the Radboud University Solanaceae database. Acta Hort 745 503–506
Vilanova S, Manzur JP, Prohens J (2012) Development and characterization of genomic simple sequence repeat markers in eggplant and their application to the study of diversity and relationships in a collection of different cultivar types and origins. Mol Breed 30:647–660
Vorontsova MS, Stern S, Bohs L, Knapp S (2013) African spiny Solanum subgenus Leptostemonum, Solanaceae): a thorny phylogenetic tangle. Bot J Linn Soc 173:176–193
Vorontsova MS, Knapp S (2016) A revision of the “spiny solanums”, Solanum subgenus Leptostemonum (Solanaceae) in Africa and Madagascar. Syst Bot Monogr 99:1–436
Wang C, Fu D (2018) Virus-induced gene silencing of the eggplant chalcone synthase gene during fruit ripening modifies epidermal cells and gravitropism. J Agric Food Chem 66(11):2623–2629
Warade SD, Sanap PB, Sawant DM, Gupta NS (2004) Studies on interspecific hybridization in eggplant (Solanum melongena L.) for little leaf disease resistance. In: Proceedings of 12th EUCARPIA meeting on genetics and breeding of capsicum and eggplant, pp 79–83
Waterhouse PM, Wang MB, Lough T (2001) Gene silencing as an adaptive defense against viruses. Nature 411:834–842
Wei Q, Wang J, Wang W, Hu T, Hu H, Bao C (2020) A high-quality chromosome-level genome assembly reveals genetics for important traits in eggplant. Hortic Res 7(1):1–15
Wu FN, Eannetta NT, Xu YM, Tanksley SD (2009)A detailed synteny map of the eggplant genome based on conserved ortholog set II (COSII) markers. Theor Appl Genet 118:927–935
Xing J, Chin CK (2000) Modification of fatty acids in eggplant affects its resistance to Verticillium dahliae. Physiol Mol Plant Pathol 56(5):217–225
Yang X, Cheng YF, Deng C, Ma Y., Wang ZW et al (2014) Comparative transcriptome analysis of eggplant (Solanum melongena L.) and turkey berry (Solanum torvum Sw.): phylogenomics and disease resistance analysis. BMC Genomics 15(1):412
Yang Y, Liu J, Zhou XH, Liu SY, Zhuang Y (2020) Transcriptomics analysis unravels the response to low temperature in sensitive and tolerant eggplants. Sci Hort 271:109468
Yuan C, Zhang S, Hu R, Wei D, Tang Q et al (2021) Comparative transcriptome analysis provides insight into the molecular mechanisms of anther dehiscence in eggplant (Solanum melongena L.). Genomics 113(2):497–506
Zhou L, Li J, He YJ, Liu Y, Chen HY (2018a) Functional characterization of SmCBF genes involved in abiotic stress response in eggplant (Solanum melongena). Sci Hort 233:14–21
Zhou X, Liu J, Bao S, Yang Y, Zhuang Y (2018b) Molecular cloning and characterization of a wild eggplant Solanum aculeatissimum NBS-LRR gene, involved in plant resistance to Meloidogyne incognita. Intl J Mol Sci 19(2):583
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2022 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

Cite this chapter
Arafa, R.A., Prohens, J., Solberg, S.Ø., Plazas, M., Rakh, M. (2022). Breeding and Genome Mapping for Resistance to Biotic Stress in Eggplant. In: Kole, C. (eds) Genomic Designing for Biotic Stress Resistant Vegetable Crops. Springer, Cham. https://doi.org/10.1007/978-3-030-97785-6_4
Download citation
DOI: https://doi.org/10.1007/978-3-030-97785-6_4
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-97784-9
Online ISBN: 978-3-030-97785-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)