
Abstract
The need to boost agricultural production in the coming decades in a climate change scenario requires new approaches for the development of new crop varieties that are more resilient and more efficient in the use of resources. Crop wild relatives (CWRs) are a source of variation for many traits of interest in breeding, in particular tolerance to abiotic and biotic stresses. However, their potential in plant breeding has largely remained unexploited. CWRs can make an effective contribution to broadening the genetic base of crops and to introgressing traits of interest, but their direct use by breeders in breeding programs is usually not feasible due to the presence of undesirable traits in CWRs (linkage drag) and frequent breeding barriers with the crop. Here we call for a new approach, which we tentatively call ‘introgressiomics’, which consists of mass scale development of plant materials and populations with introgressions from CWRs into the genetic background of crops. Introgressiomics is a form of pre-emptive breeding and can be focused, when looking for specific phenotypes, or un-focused, when it is aimed at creating highly diverse introgressed populations. Exploring germplasm collections and identifying adequate species and accessions from different genepools encompassing a high diversity, using different strategies like the creation of germplasm diversity sets, Focused identification of germplasm strategy (FIGS) or gap analysis, is a first step in introgressiomics. Interspecific hybridization and backcrossing is often a major barrier for introgressiomics, but a number of techniques can be used to potentially overcome these and produce introgression populations. The generation of chromosome substitution lines (CSLs), introgression lines (ILs), or multi-parent advanced inter-cross (MAGIC) populations by means of marker-assisted selection allows not only the genetic analysis of traits present in CWRs, but also developing genetically characterized elite materials that can be easily incorporated in breeding programs. Genomic tools, in particular high-throughput molecular markers, facilitate the characterization and development of introgressiomics populations, while new plant breeding techniques (NPBTs) can enhance the introgression and use of genes from CWRs in the genetic background of crops. An efficient use of introgressiomics populations requires moving the materials into breeding pipelines. In this respect public–private partnerships (PPPs) can contribute to an increased use of introgressed materials by breeders. We hope that the introgressiomics approach will contribute to the development of a new generation of cultivars with dramatically improved yield and performance that may allow coping with the environmental changes caused by climate change while at the same time contributing to a more efficient and sustainable agriculture.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The expected increasing demand of plant products in the coming decades, with an estimated need of the doubling in relation to 2005 levels of agricultural production by 2050 (Tilman et al. 2011) in a climate change scenario, represents a formidable challenge for plant breeders. The current yearly rate of yield increase for major crops is clearly insufficient to meet this goal (Ray et al. 2013). Forecasts of agricultural productivity indicate that agriculture will be one of the sectors most affected by climate change (Rosenzweig et al. 2014). The negative effects of climate change on agricultural productivity will probably be greatest in tropical and subtropical areas (Knox et al. 2012; Rosenzweig et al. 2014), where a large part of the human population lives in marginal conditions in developing countries. Climate change will undoubtedly increase both permanent and occasional abiotic stresses (drought, high temperatures, salinity) in a large part of the global agricultural land and has the potential to drive abandonment of cultivation and desertification in some regions of the world. In addition, climate change is expected to result in increased biotic stress, as many pests and diseases will potentially migrate to areas where they were not present due to environmental limitations (Bebber et al. 2013).
Although many improvements have been made in increasing the efficiency of inputs used in agriculture (energy, water, agrochemicals) in the near future higher yields will have to be obtained with less input (Godfray et al. 2010; Ray et al. 2013). Also, many agricultural lands have become salinized or have lost fertility due to mismanagement (Fita et al. 2015). The availability of new arable land is not only limited but it is undesirable to increase the agricultural land area on a global scale given that conservation of biodiversity is important for maintenance of functioning ecosystems and genetic input to agriculture in the longer term.
Meeting the challenges of increasing agricultural production in the face of climate change will require new strategies to develop new crop varieties with increased resilience to climate change-related stresses (Fita et al. 2015). Significant advances have been made in developing varieties tolerant to abiotic or biotic stresses but mostly for monogenic or oligogenic traits. The genetic variation available in cultivated species for adaptation to climate change is generally limited and it may be impossible to achieve major advances by resorting only to alleles present in the cultivated genepool (Hajjar and Hodgkin 2007; Vincent et al. 2013; Dempewolf et al. 2014; Warschefsky et al. 2014).
Breeders have long recognized the value of crop wild relatives (CWRs) as sources of novel beneficial variation for resistance or tolerance to stresses (Bessey 1906; Harlan and de Wet 1971; Hajjar and Hodgkin 2007). In this respect, most of the modern varieties of several important crops, like wheat or tomato, carry introgressions from wild relatives (Menda et al. 2014; Wulff and Moscou 2014). CWRs are wild species that are able to exchange genes with the cultivated taxa through sexual or somatic hybridization (Harlan and de Wet 1971; Maxted et al. 2006). CWRs are phylogenetically closely related to the crop and often encompass great genetic diversity. Some CWRs develop well under marginal conditions (low rainfall, high temperatures, high pressure of pests and diseases) that would be highly stressful for elite varieties of crops (Dwivedi et al. 2008; Dempewolf et al. 2014; Warschefsky et al. 2014). The enormous potential of crop wild relatives for breeding new varieties adapted to climate change has largely remained unexploited, however, and the use of CWRs in breeding has largely focused on introgressing loci for disease resistance, while tolerance to abiotic stress has been little explored (Hajjar and Hodgkin 2007; Maxted and Kell 2009).
Recently an initiative has been launched aiming at adapting agriculture to climate change through the use of crop wild relatives (CWRs) for improving the cultivated gene pool of 29 crops included in the Annex 1 of the International Treaty on Plant Genetic Resources for Food and Agriculture (ITPGRFA) (Dempewolf et al. 2014). This project is aiming at collecting, preserving, evaluating, and preparing the use of CWRs with the ultimate aim to broaden the genetic base and thereby adapt our crops to climate change. Other initiatives, like Diversity Seek (DivSeek) are aimed at unlocking the potential of the diversity of crops and wild relatives present in genebanks (Meyer 2015).
In this paper we present a novel strategy, tentatively named ‘introgressiomics’, for the systematic exploitation of the native variation present in CWRs for an efficient adaptation of our crops to climate change.
Crop wild relatives for broadening the genetic base of crops
Plant domestication is a unique and complex evolutionary process in which natural and artificial selection resulted in new plants adapted to human needs (Meyer and Purugannan 2013). Domesticated plants generally present a syndrome of distinctive morphological, physiological and reproductive features compared to their wild ancestors (Hammer 1984). For example in cereals, non-shattering phenotypes, with larger seeds, synchronized ripening, change in the plant architecture and loss of seed dormancy are characteristic traits of domesticates (Salamini et al. 2002). In other crops, such as those used for their fruits, tubers or leaves, domestication involved loss of bitterness and anti-nutritional or toxic compounds, removal or reduction of physical defence mechanisms like prickles, and gigantism of the organs used by humans (Meyer et al. 2012). One extreme example is that of giant pumpkins (Cucurbita maxima) with record weights of over a ton for a single fruit (Savage et al. 2015). Many crops also experienced changes in the reproductive system that isolated them from the CWRs (Meyer and Purugannan 2013). In other cases crossability has been maintained within the genepool (Jarvis and Hodgkin 1999; Meyer and Purugganan 2013). Autogamy, which allows fixation and maintenance of selected characteristics, has been favoured in a number of crops (Meyer et al. 2012). Increased yield potential is also a general feature of domestication.
On many occasions domestication has involved genetic bottlenecks, resulting from a founder effect (domestication from a limited number of individuals), reproductive isolation and/or from strong selection pressures during domestication or crop improvement (Dempewolf et al. 2012; Meyer and Purugannan 2013). This resulted in a narrow genetic base for most crops as compared to their closest wild progenitors and CWRs (Dwivedi et al. 2008; Hajjar and Hodgkin 2007; Meyer and Purugannan 2013; Dempewolf et al. 2014). Modern plant breeding has generally led to a further reduction of genetic diversity in comparison to modern elite varieties and landraces (Tanksley and Nelson 1996; de Wouw et al. 2009). Notable exceptions to this trend are crops where modern varieties carry introgressions from CWRs. For example, in tomato, modern varieties are genetically more diverse than local landraces due to the multiple introgressions, usually for resistance to diseases, from CWRs (Sim et al. 2012). Nonetheless, the diversity of modern tomato is much lower than that present in its CWRs (Aflitos et al. 2014; Dodsworth et al. 2016).
A narrow toolbox of alleles in the elite varieties limits options for to better adapt crops to climate change. CWRs are often adapted to environments that are more stressful than artificial agricultural conditions. The large genetic diversity present in CWRs thus may constitute a major source of variation for improving crops with higher resilience (Warschefsy et al. 2014).
From a breeder’s point of view, the utilization of CWRs present some significant challenges (Hajjar and Hodgkin 2007; Dwivedi et al. 2008; Meyer and Purugganan 2013). For example, crossing barriers and low hybrid fertility or sterility represent major barriers for the use of some species of CWRs in breeding programs. Although well-adapted to their natural environment, CWRs often contain a range of undesirable traits to agricultural conditions (low yield, undesirable physical and chemical defence systems, unpleasant flavour, lack of adaptation to cultivated conditions, etc.) (Meyer and Purugganan 2013). In many cases, these “wild” traits are dominant and polygenic, and thus challenging to select against in the breeding programs. Linkage drag due to reduced recombination is another issue that is frequently observed in introgression programs. Once introgressed into the cultivated genetic background, the CWR chromosomal fragments are challenging to break into smaller components. These fragments often contain genes that confer undesirable phenotypes—and often these are linked to the gene/s controlling the traits of interest (Tanksley and Nelson 1996; Wendler et al. 2015). In addition, phenotypes of interest in a CWR may not be expressed in a cultivated genetic background. Nonetheless, use of CWRs in breeding has allowed significant improvements in a number of crops (Hajjar and Hodgkin 2007). Maxted and Kell (2009) list 183 CWR taxa of a total of 29 crops that have been used for the transfer of traits of interest to the crop, while Dempewolf et al. (2017) lists 4175 potential or confirmed uses of CWR taxa in crop improvement research, spread across 127 crops and 970 CWR taxa. In several cases the use of CWRs in breeding research has not resulted in their use in the development of cultivated varieties (Kilian et al. 2011). Maxted and Kell (2009) list only 39 CWR taxa that have been utilized for the development of advanced cultivars in nine major cereal and legume crops. Furthermore, most uses of CWRs for improvement of these nine crops are related to resistance to diseases and pests (61%), while their use for the improvement of tolerance to abiotic stresses (16%) or yield (7%) has been much lower. Other uses (16%) mostly involve quality traits. In tomato, most of the introgressions from wild taxa have been aimed at disease resistance, with 18 resistances having been introduced from seven different species (Díez and Nuez 2008). In sum, the utilization of CWRs has mostly been restricted to resistance or tolerance to pests and diseases, while other potential uses, like adaptation to abiotic stresses, have been largely neglected. With their adaptations to challenging environments, the utilization of CWRs represents a largely untapped opportunity for breeders to improve the adaptation of crops to abiotic stresses such as drought, salinity and high temperatures (Dwivedi et al. 2008; Dempewolf et al. 2014; Fita et al. 2015).
The ‘genepool’ concept was established by Harlan and de Wet (1971) to denote differences between CWR taxa with regards to the ease of exchanging genes with the crop. Typically, the primary genepool includes CWRs that can be easily crossed with the crop and the offspring is fertile. Secondary genepool CWRs are those that can be crossed with the cultivated species, although sometimes the degree of success is low, and/or the offspring may present low fertility. Finally, the tertiary genepool is composed of CWRs which have strong reproductive barriers with the crop and obtaining hybrids may require specific techniques such as embryo rescue, use of bridge species, stigma excision, or the use of pollen mixtures. Furthermore, hybrids between the crop and tertiary genepool species are often sterile and polyploidization may be needed to restore fertility in some cases. Thus the use of tertiary genepool CWRs for crop improvement is more challenging (Harlan and de Wet 1971; Khush and Brar 1992; Dwivedi et al. 2008). Given that crossabilities have not been sufficiently studied for numerous CWRs, phylogenetic relationships are sometimes used to assign CWRs to the respective genepool (Maxted et al. 2016).
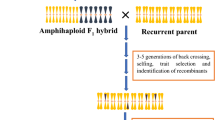
CWRs are not only of interest for their use in breeding, but can also be used directly. For example, CWRs with high vigour, resistance to diseases or with higher tolerance to abiotic stresses can be used as rootstocks in fruit and vegetable crops (Schwarz et al. 2010). The wild eggplant Solanum torvum is commonly used for eggplant grafting due to its resistance to multiple diseases and high vigour (Ranil et al. 2015). CWRs may also be of direct use for creating new cultivated species, such as for tritordeum, which is an amphiploid hybrid between durum wheat and the wild Hordeum chilense (Martín et al. 1999). Tritordeum has good tolerance to drought and high temperatures (Villegas et al. 2010).
Although many CWRs are of interest for adapting our crops to climate change, many CWRs are also threatened by climate change and human impact, as demonstrated for CWRs of cowpea, peanut and potato, among others (Jarvis et al. 2008). The collection and conservation of the CWR diversity continues to be an urgent priority (Maxted and Kell 2009; Dempewolf et al. 2014).
The introgressiomics approach
As CWRs cannot be directly incorporated into commercial breeding programs, pre-breeding activities have to be initiated as a first step for the utilization of CWRs (Longin and Reif 2014). Most pre-breeding works use CWRs as donors of novel genetic diversity and is based on the following rationale:
-
1
Ad hoc, whenever an urgent breeding need appears (most commonly the occurrence of a new disease or pest), sources of useful variation are being explored. Screening usually starts within the primary genepool, mostly among the cultivars and landraces, sometimes including the closest CWRs. If unsuccessful, then screening is expanded to secondary and tertiary genepools.
-
2
Once a suitable source of variation is detected, a crossing (and backcrossing) program is initiated to introgress the gene(s) conferring the desirable trait into the crop. Typically a single CWR donor species and most commonly just a single accession (Hajjar and Hodgkin 2007) is considered.
Depending on the trait and introgressed fragment, this conventional approach can be time consuming and can require several cycles of backcrossing and selection, followed by selfing and then again further cycles of selection. Examples can be found in tomato, where most modern commercial varieties of tomato harbor resistances to nematodes or viral diseases caused by Tomato mosaic virus (ToMV) or Tomato yellow leaf curl virus (TYLCV) (Díez and Nuez 2008). Resistance to nematodes conferred by the Mi locus was introduced in the 1940s from Solanum peruvianum accession PI128657 (Smith 1944), while resistance to ToMV conferred by Tm2 2 was introduced in the 1960s from S. peruvianum accession number PI128650 (Alexander 1963), and later the resistance to TYLCV conferred by Ty-1 was provided by S. chilense accession LA1969 in the 1990s (Zamir et al. 1994). Another successful example of alien gene introgression was the introduction of the Lr19 leaf rust resistance allele, from Thinopyrum ponticum to wheat (Sharma and Knott 1966). This gene not only confers rust resistance to wheat, but also increases yield, biomass and grain number (Bedő and Láng 2015).
Alternative approaches have been proposed. McIntosh (1992), for example, suggested the development of ‘pre-emptive’ breeding populations to introgress resistance loci to wheat rusts with the aim of having promising materials ready for the rapid generation of resistant cultivars—in case the predominant rust races changed or a major resistance locus break down. In this way breeders could quickly develop new resistant varieties. However, this procedure bears significant risks and is expensive. Breeders need a clear strategy, a long time horizon and significant human and financial resources. A modification to this original pre-emptive breeding approach is the creation of ‘pre-breeding populations’ by crossing the crop with one or several CWRs. Valkoun et al. (2001) created pre-breeding populations of wheat containing introgressions from five different CWRs from genera Triticum and Aegilops. Some of these carry beneficial traits under certain environments, such as earliness, short plant stature, drought tolerance or resistance to several rusts. Introgression lines (ILs) contain one or a few introgressed genome fragments from a CWR (Zamir 2001; Gur and Zamir 2004). Sets of ILs with overlapping fragments of different size can be exploited for different purposes, including for the analysis of the genetic basis for traits of (Zamir 2001; Alseekh et al. 2013; Guerrero et al. 2016).
Here we suggest a novel approach for the development of pre-breeding materials, tentatively called ‘introgressiomics’ (Fig. 1). ‘Introgressiomics’ consists of a mass scale systematic development of plant materials and populations carrying introgressions of genome fragments obtained from (mostly wild) crop relatives into the genetic background of crops that may allow developing new generations of cultivars with improved properties. ‘Introgressiomics’ is aiming at the massive generation of introgression materials for future (foreseen and unforeseen) needs and therefore may be considered as an advanced form of pre-emptive breeding. The ultimate aim of introgressiomics is to provide breeders with a significantly enlarged genetic pool from which new generations of cultivars adapted to future challenges in crop production can be rapidly obtained. Importantly, the materials developed through an introgressiomics approach are ready to be directly incorporated into breeding pipelines. Such materials will facilitate the work of breeders, especially with respect to the challenge of breeding for complex traits (Cattivelli et al. 2008; Dempewolf et al. 2014).

Outline of the introgressiomics pre-breeding approach workflow process, from the use of genetic resources to the development of elite materials with introgressions from crop wild relatives (CWRs)
The first steps introgressiomics program are as follows: (1) identification of CWRs to be used in the program; (2) hybridization and backcrossing of the crop with a number (as large as possible) of CWRs from different genepools using special techniques when needed; (3) development of multiple special introgression populations containing introgressed fragments from one or several CWRs using genomic tools; (4) creating repositories of the introgressiomics populations and materials and databases with phenotypic and genomic information; and lastly, (5) moving the materials into breeding pipelines (Fig. 1).
An important aspect of introgressiomics is that populations may be created consisting of multiple genomic fragments obtained from different CWR sources. This approach of ‘pyramiding of genomic regions of interest’ would be beneficial for some breeding programs aiming at improving complex traits such as yield-related parameters.
Introgressiomics is similar, although more ambitious, than the approach proposed by Warschefsky et al. (2014), who pointed to the “need for systematic efforts to introgress broad subsets of wild relative diversity to incorporate the range of useful adaptations for disease resistance, abiotic stress tolerance, and other agronomic challenges that are required in order to increase the resiliency and productivity of agriculture in the 21st century”. Warschefsky et al. (2014) suggested a five-step approach similar to that of introgressiomics consisting of: (1) build comprehensive collections of CWRs, (2) sequence them, (3) create sets of purpose-driven hybrid populations and characterize them phenotypically, (4) develop a predictive network of genotype-phenotype associations, and (5) deploy the identified phenotypes into crop breeding pipelines. Their proposal presents substantial differences in its conception than the introgressiomics approach, as can be observed by comparing our Fig. 1 with the Fig. 1 of Warschefsky et al. (2014). The SCREAMing (Systematic Creation of Really Exotic Abnormal Material) approach has been proposed by Bert Vandenberg (pers. comm.) to develop pre-breeding populations using CWRs, but a comprehensive description of the approach has not been published yet.
Introgressiomics, as proposed here can be: (i) focused and directed at specific phenotypes, which will determine what CWRs and types of populations are required, or (ii) un-focused, in which the objective is the generation of introgression materials encompassing the maximum genetic diversity possible.
Identification of CWRs for introgressiomics
The identification of CWRs to be utilized for introgression in the genetic background of the crop is the first step in introgressiomics (Fig. 1). This largely depends on the strategy to be used (“focused” vs. “un-focused”) and also on the availability of CWRs (Vincent et al. 2013; Castañeda-Álvarez et al. 2016). In general, maximizing genetic diversity to the extent possible is a priority, as this increases the likelihood of capturing beneficial allelic diversity for breeding (Tanksley and McCouch 1997; Porch et al. 2013). The number of CWR accessions to be used for introgressiomics depends on the resources available, the scale and time horizon, and objectives of the introgressiomics program. In order to select and identify CWRs, the ‘Harlan and de Wet’ Crop Wild Relatives Inventory (http://www.cwrdiversity.org/checklist/; Vincent et al. 2013) is a helpful resource. Currently, 173 crop genepools are described. Previous and potential use cases of CWRs are also presented (Dempewolf et al. 2017).
One option for the identification of CWRs for ‘un-focused’ introgressiomics is the creation of germplasm diversity sets. This type of germplasm sets allows for a more rational use of accessions, while adjusting the number of accessions that can be realistically used (McKhann et al. 2004). Ideally, these germplasm subsets should include CWR representatives of all genepools. However, germplasm diversity sets of CWRs merely based on diversity (genetic, phenotypic, origins, environmental, or a combination of them) might result in underrepresentation of CWRs from the primary genepool, which generally are typically less variable than CWRs from the secondary or tertiary genepools. Adjustments may be made in the germplasm diversity sets to ensure a good representation of primary genepool CWRs, which are the easiest to use for introgression breeding (Harlan and de Wet 1971).
For ‘focused’ introgressiomics, germplasm sets based only on diversity are not appropriate, as for this approach it is important that the germplasm sets are enriched for material which is harbouring alleles that may contribute to improving the target trait/s. For example, in creating introgressiomics populations aimed at improving drought tolerance, emphasis should be given to include CWRs known to be tolerant to drought. In this case, strategies, like the Focused Identification of Germplasm Strategy (FIGS) could help selecting potentially beneficial material. FIGS is based on the assumption that accessions carry adaptive traits that reflect the selection pressures to which these were subjected in the environment during evolution (Street et al. 2016). Therefore, the retrieval of climatic and environmental data from databases such as WorldClim (http://www.worldclim.org; Hijmans et al. 2005), WorldGrids (http://worldgrids.org), or the CGIAR Consortium for Spatial Information (http://www.cgiar-sci.org), using the geographical coordinates may help in identifying promising CWR accessions. The FIGS strategy has been successfully applied for crops like wheat and beans (Bari et al. 2012; Khazaei et al. 2013). However, the potential of FIGS for selecting CWRs for introgression breeding is yet to be fully exploited (Street et al. 2016). A similar approach consisting of the use of eco-geographical data and expert assessment has allowed the identification of CWRs of sunflower to improve tolerance to various abiotic stresses (Kantar et al. 2015). Also, the utilization of historic characterization and evaluation data obtained ex situ may be appropriate to identify CWR accessions of potential interest for breeding.
The creation of CWR sets requires the exploration of publicly available ex situ germplasm repositories as well as private working collections, when possible. Unlike accessions of the cultivated species, CWRs are often underrepresented in ex situ genebanks (Maxted and Kell 2009; Castañeda-Álvarez et al. 2016), which means that relevant diversity may not be immediately available for introgressiomics. This clearly reduces the potential for introgressing genes of interest from CWRs. The detection of potential collection gaps for CWRs is a first order step in order to fully exploit CWRs (Dempewolf et al. 2014). Gap analysis of germplasm collections is a strategy that was proposed by Maxted et al. (2008) and consists of identifying the taxa and geographical areas underrepresented in ex situ collections using taxonomic, genetic, eco-geographical, and threatened status data and other assessments. Additionally, comparison of georeferenced herbarium specimen data with the geographical coordinates of collection of the accessions conserved in ex situ collections allows identification of priority CWR taxa for collection as well as conservation areas as has been demonstrated in beans (Ramírez-Villegas et al. 2010) or eggplant (Syfert et al. 2016). In this respect, the Crop Wild Relative Occurrence Database (Centro Internacional de Agricultura Tropical 2017) contains over five million records of germplasm accessions and herbarium databases on CWR and is of great interest for identification of gaps in the present germplasm collections.
Combining genomics and phenomics information might further assist the selection of CWRs for introgressiomics, as shown by Vosman et al. (2016) for CWRs of cabbage resistant to the cabbage aphid. The combination of phenotypic and genomics data from introgressiomics populations can be used for ‘introgressiomics by design’, which is an extension of the Breeding by Design™ approach (Peleman and van der Voort 2003). The ‘introgressiomics by design’ approach would consist in mapping loci of agronomic interest from different donor CWR and using crossing and pyramiding schemes (Gur and Zamir 2015) to introduce several favourable introgressions from different CWRs in a single line with the crop genetic background (Peleman and van der Voort 2003).
Interspecific hybridization and backcrossing
A central step for introgressiomics is the hybridization between the crop and the CWR to create introgression populations. Theoretically, the crossability between the crop and all taxa within the primary genepool, which often comprise also the genome donors of the crop, should not present more difficulties than for intra-specific hybridizations (Harlan and de Wet 1971; Jones 2003; Maxted et al. 2006; Vincent et al. 2013). Although occasionally and depending on the crop, hybridization can be challenging for several reasons, such as non-synchronous flowering, cross-incompatibilities or fertility issues (Rieseberg and Carney 1998; Dwivedi et al. 2008; Jones 2003; Vincent et al. 2013).
Interspecific hybridization between the crop and CWR taxa from the secondary and tertiary genepools (wide or distant crosses) are often more challenging and hampered by pre-zygotic and post-zygotic reproductive barriers (Zenkteler 1990; Khush and Brar 1992) (Fig. 2). In addition, although it may be possible to obtain hybrids, sterility issues may limite the development of introgressiomics populations. One important pre-zygotic reproductive barrier is pollen-style incompatibility (Dwivedi et al. 2008). As a consequence pollen does not germinate. When ‘unilateral’ incompatibility exists, hybrids can be obtained using the female parent in which the pollen is able to germinate (Fig. 2). Unilateral incompatibility can frequently be observed when crossing self-compatible with self-incompatible species. The cross is successful when using the self-compatible species as a female parent (Dhaliwal 1992). For example when crossing the cultivated tomato (Solanum lycopersicum) with its wild relative S. peruvianum the cross is unsuccessful when using the self-incompatible S. peruvianum species as female parent, but possible when using it as male parent. In the former case the barrier is pre-zygotic and the pollen tube can not penetrate the style. The reciprocal cross is possible, although the embryo must be rescued and cultured in vitro to prevent the embryo abortion (Hogenboom 1984). When ‘bilateral’ incompatibility exists, several techniques have proven useful to overcome it (Rieseberg and Carney 1998; Dwivedi et al. 2008). Among others, pollen mixtures of compatible pollen, inactivated or not, stigma exertion, physical or chemical treatments to the stigmas, or in vitro fertilization can be applied (Zenkteler 1990; Khush and Brar 1992; Dwivedi et al. 2008). Pollen mixture and treatment of stigmas with H3BO3 and GA3 were used by Picó et al. (2002) to obtain offspring from crosses between the cultivated tomato and the distant wild relatives S. chilense and S. peruvianum.

Pre- and post-zygotic barriers difficulting interspecific hybridization and introgression breeding with crop wild relatives (CWRs) and strategies to overcome them
If the male gamete is able to reach the ovary and to fertilize the female gamete in the ovule, post-zygotic barriers may prevent obtaining a viable hybrid seed. Most important post-zygotic barriers are caused by differences in number of chromosomes, ploidy levels, chromosomal alterations (rearrangements, duplications, inversions or translocations), embryo-endosperm incompatibility and hybrid lethality (Khush and Brar 1992). These post-zygotic barriers (Fig. 2) may result in embryo abortion, preventing the development of viable seed. However, different techniques can be applied to try to avoid post-zygotic barriers. For example, when crossing individuals of different ploidy levels, the use of technologies that modify the ploidy level can contribute to the development of viable hybrids. Most frequently used techniques to obtain parental lines with the same level of ploidy are anther culture to produce haploid individuals (chromosome complement reduced to a half), or the duplication of the genome with colchicine (Khush and Brar 1992; Rieseberg and Carney 1998; Dwivedi et al. 2008), Also, embryo rescue, at an early stage of development (i.e. before it aborts due to either embryo-endosperm incompatibility or due to abnormal development resulting from genetic imbalance or other alterations), has been a successful tool in producing interspecific hybrids between crops and CWRs (Khush and Brar 1992; Sharma et al. 1996).
When pre-zygotic and post-zygotic barriers cannot be overcome to obtain hybrid plants, the use of bridge species has to be considered (Shivanna and Bahadur 2015). These species are compatible with one or both target taxa for interspecific hybridization. Once the interspecific hybrid between one of the taxa and the bridge species has been obtained, F1 plants can then been crossed with the other species—or with the other interspecific hybrid between the bridge species and the other parent. In this way, it has been possible to transfer genes from some CWRs to cultivated species. For example, the wild strawberry Fragaria vesca is a potential a bridge species for introgression breeding of strawberry (Fragaria x ananassa) from the wild F. nilgerriensis, F. nubicola, F. pentaphylla and F. viridis (Bors and Sullivan 2005). Somatic hybridization may be an alternative to obtain interspecific hybrids, as long as regeneration of plants is possible from hybrid somatic cells (Johnson and Veilleux 2000). In this case the hybrid plant may be a polyploid having the full genome complements of both parental species. In some cases, like bread wheat, where crossing between elite varieties and CWR is frequently unsuccessful, inhibition of crossing is under the control of a few genes, so that wheat varieties or stocks homozygous for the crossability alleles kr1 and kr2 have increased crossability with CWR (Alfares et al. 2009).
Once interspecific hybrids have been obtained, backcross generations are produced (Zamir 2001; Gur and Zamir 2004). Although interspecific hybrids may be viable, they may be partially or completely sterile due to irregular chromosome pairing due to different ploidy levels or a low degree of synteny that results in non-viable gametes (De Storme and Mason 2014). Some approaches can be used to increase the success rate of obtaining viable progeny from hybrids with reduced or low sterility. One strategy is to use the low-fertility hybrid as a female parent, since for the fertilization the pollen must have a high vigour to germinate and reach the ovule—a requirement not needed for the female gamete. This applies also to other plants from subsequent backcross generations. One way to recover fertility of the hybrid is duplicating its genome (Khush and Brar 1992; Rieseberg and Carney 1998; Shivanna and Bahadur 2015). In this way normal chromosome pairing can be restored, although the hybrid plant will be polyploid, which may be an obstacle for obtaining subsequent generations due to different ploidy levels. To prevent this, a common technique is to duplicate also the genome of the recurrent cultivated parent to obtain backcross progeny and to restore the diploid status at a later stage. For example, Toppino et al. (2008) introgressed of resistance to Fusarium wilt from S. aethiopicum into the genetic background of eggplant (S. melongena) by backcrossing the tetraploid somatic hybrid to tetraploid eggplant to obtain the first backcross generation, which was subsequently returned to the diploid stage by anther culture. Generally, the loss of fertility in interspecific hybrids can be recovered in backcross generations, with increasing levels of fertility as the genome of the recurrent cultivated parent is being recovered (Wall 1970). However, occasionally the phenomenon of selective chromosome elimination of the donor parent may occur, complicating the introgression of fragments of CWR (Dwivedi et al. 2008). In other cases, recombination in the hybrids between chromosomes of the cultivated species and the CWR is supressed or reduced, which makes introgression more difficult (Bedő and Láng 2015). In this way, in wheat the gene Ph1 suppresses pairing and recombination of wheat and alien chromosomes; however, in plants that are nullisomic for the Ph1 gene, or in ph1b mutant stocks, homoelogous wheat and alien chromosomes can pair and recombine, which facilitates introgression from CWR in the cultivated wheat (Friebe et al. 2012).
Introgressiomics generally is conceived as the introgression of nuclear genes from wild species into a cultivated genetic background. However, introgression of cytoplasmic genes is also possible to produce alloplasmic hybrids with the nuclear genome of the cultivated species and the cytoplasm of the wild species (Khan et al. 2015). In this case, the cultivated species has to be used as male parent to ensure that the wild cytoplasm is maintained during the process. Nevertheless, it has to be taken into account that the interaction between the nuclear genome of the recurrent parent (cultivated species) and the cytoplasm of the donor (wild species) may result in cytoplasmic sterility (Prakash et al. 2001; Dwivedi et al. 2008; Khan et al. 2015), which may not be desirable if fertile plants of the cultivated species are needed. In that case, the cultivated species should be used as female parent to recover their cytoplasm, at some point during the backcross program.
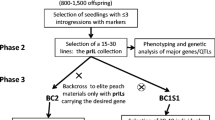
Development of introgressiomics populations
Once interspecific hybrids have been obtained, introgressiomics populations (Fig. 3), have to be developed. Some of the most commonly used populations, which contain genome fragments from CWR, are chromosome substitution lines (CSLs) and introgression lines (ILs). These are considered ‘immortal’ populations as they can be maintained by selfing, based on backcrossing the hybrid to the recurrent cultivated parent for several generations (Zamir 2001; Lippman et al. 2007). CSLs consist of lines harboring the full genome complement of the crop except for one chromosome pair that corresponds to the wild donor parent (Cavanagh et al. 2008). CSLs have been extensively used in wheat breeding (Kilian et al. 2011; Khlestkina 2014) and allow ascribing genes and traits to specific chromosomes. Although deleterious genes may be present in the introgressed pair of chromosomes, in contrast to CSLs ILs harbour the full genome of the crop, except for a small chromosomal segment of a donor parent, typically a CWR (Zamir 2001). The development of collections of ILs, can be of great utility for breeders, because given that the introgression represents only a part of a chromosome, it may contain less deleterious alleles than CSLs (Gur and Zamir 2004; Lippman et al. 2007). Both CSLs and ILs can be obtained through repeated backcrossing of the hybrid to the recurrent parent. Molecular markers and/or complementary cytogenetic techniques such as genome in situ hybridization (GISH) help tracking the introgressed fragments and thus support the selection of beneficial materials for subsequent backcross cycles (Gupta et al. 2016). A final step in obtaining ILs is selfing or obtaining doubled haploids to fix the introgressed fragment in a homozygous state (Herzog et al. 2014). Also, ILs can be obtained from CSLs by crossing with the recurrent parent and subsequent selection of individuals in which recombination has taken place (Cavanagh et al. 2008). Similarly sub-ILs (Fig. 3) can be obtained from ILs to shorten the introgressed fragment to reduce linkage drag (Alkeesh et al. 2013). A further advantage of ILs is the ability to intercross favourable traits that are present in different ILs for pyramiding favourable alleles (Gur and Zamir 2015). Advanced backcross populations, in which no marker assisted selection has been carried out during population development, may also be of interest as introgressiomics populations (Tanksley and Nelson 1996; Cowling et al. 2009). In these populations the percentage of the donor genome will be reduced on average by half in each backcross cycle. Typically, these advanced backcross populations contain several fragments of the donor in several parts of the genome.

Scheme for the development of several types of introgressiomics populations: chromosome substitution lines (CSLs, left); introgression lines (ILs, center); and, multi-parent advanced inter-cross (MAGIC) lines (right)
Other types of populations can be utilized to obtain ‘introgressiomics populations’. For example, recombinant inbred lines (RILs) obtained after crossing one cultivated species and a CWR followed by several generations of selfing have been very useful to dissect traits of interest present in CWR (Peleg et al. 2009; Salinas et al. 2013). However, RILs carry on average 50% of the wild parent, which in most cases are unsuitable for commercial breeding programs without a pre-breeding pipeline in place. An alternative to the backcross method is to develop multi-parental populations, such as multi-parent advanced generation inter-cross (MAGIC), which consist of multi-parent RIL populations (Cavanagh et al. 2008; Pascual et al. 2016).
A more simplistic approach for the development of introgressiomics populations involves the development of several populations, each of which contains introgressions from a single donor CWR parent. Introgressiomics populations may contain introgressions from several CWRs. For example, interspecific hybrids between two CWRs or double hybrids between four CWRs can be used as starting point to develop advanced backcross generations that may contain genome fragments from several CWRs. Also, more than one CWR parent can be included in MAGIC populations to generate RILs that have genomic fragments of several wild species.
One of the main challenges to the development and use of introgressiomics populations is linkage drag, based on reduced recombination at introgressed fragments (Tanksley and Nelson 1996; Wendler et al. 2015). The use of molecular markers allows selecting individuals carrying recombined introgressed fragments (Alkeesh et al. 2013). Special mating designs, such as sib-mating, can be considered in the backcross scheme to enhance recombination (Wall 1970; Liu et al. 1996; Rieseberg et al. 1996). Also, the use of congruency backcrossing, where backcrosses towards the recurrent parent are alternating to backcrosses to the donor parent to obtain fertile materials can be used to increase recombination (Haghighi and Ascher 1998; Muñoz et al. 2004). Recently, Wendler et al. (2015) proposed crossing ILs with overlapping fragments originating from different donors to obtain progenies exhibiting higher levels of recombination within the introgressed fragment. However, in some occasions, due to the lack of synteny, recombination is prevented (Tanksley and Nelson 1996). For example, in tomato, repeated attempts to reduce the introgression size associated to Tomato yellow leaf curl virus (TYLCV) resistance gene Ty-1 introgressed from S. chilense failed due to lack of recombination caused by an chromosomal translocation (inversion) (Verlaan et al. 2011). In these cases reducing the linkage drag is not possible by using standard crossing schemes. One alternative to no or low levels of recombination due to the lack of crossing-over is the development of addition lines, containing an extra chromosome from a donor. In this way it may be possible to recover some plants in which a fragment of the donor is introgressed in the genome of the recipient parent (Friebe et al. 1996; Jacobsen and Schouten 2007).
In polyploid crops, like bread wheat, an alternative to obtain introgressiomics populations using the diversity of CWR is the artificial synthesis of the allopolyploid using genetically diverse materials of the ancestor species as parental lines. For example, hexaploid wheats have been resynthesized through the hybridization of tetraploid wheats and Aegilops species (Bedő and Lang 2015). Some of these materials were backcrossed to breeding lines and were found to be a source of variation for adaptation to several abiotic and biotic stresses (Trethowan and Mujeeb-Kazi 2008). These synthetic introgression materials revealed a significant increase in genetic diversity (Warburton et al. 2006).
Genomic tools and new plant breeding techniques for introgressiomics
Tremendous advances in next generation sequencing (NGS) technologies have provided breeders with a wide array of genomic tools that facilitate the introgression of CWR fragments into crops (Baute et al. 2015; Kole et al. 2015; Abberton et al. 2016; Brozynska et al. 2016). Some of the most important tools from genomics are the development of molecular markers distributed throughout the genome, the availability of dense genetic maps, the availability of reference genome sequences, transcriptome sequences, the discovery of regulatory elements, as well as gene annotations (Pérez-de-Castro et al. 2012). The use of these tools facilitates the identification of sources of variation and the development and characterization of introgressiomics populations.
The availability of molecular markers that can be adapted to high-throughput genotyping platforms is of great utility for identifying those CWRs that may contribute beneficial diversity to the introgressiomics approach. For example, the development of introgressiomics populations in eggplant using wild relatives (Plazas et al. 2016) benefited from molecular marker studies that evaluated the genetic diversity and relationships between eggplant and a wide number of CWRs (Furini and Wunder 2004; Vorontsova et al. 2013). This facilitated the selection of 15 eggplant CWR accessions from all genepools for the initiation of an introgression breeding program (Plazas et al. 2016).
Molecular markers greatly facilitate the creation and characterization of CSL and IL populations. Marker-assisted foreground and background selection strategies—for those individuals that contain the target chromosomal segments, while at the same time selecting for beneficial alleles in the recipient parental genome—are being applied in several crops (Zamir 2001; Gur and Zamir 2004; Pérez-de-Castro et al. 2012). In this way, collections of CSLs and ILs containing introgressed genome fragments of CWRs have been obtained in many crops (Gur and Zamir 2004; Dwivedi et al. 2008; Khlestkina 2014). The increased availability of high-density and mapped markers allows the fine mapping of the introgressed fragments, their extent as well as the breaking of ILs into sub-ILs by detecting recombinants (Lippman et al. 2007; Wendler et al. 2015; King et al. 2016).
Apart from providing molecular markers and identifying allelic variants, CWR transcriptome and genome sequences as well as resequencing studies can contribute other relevant information for introgressiomics. For example, synteny studies derived from the comparison of sequences of the crop and its CWRs may help identifying those genomic regions for which major chromosomal rearrangements have taken place during crop evolution and for which recombination will certainly be supressed (Verlaan et al. 2011). Furthermore, sequence information facilitates allele mining in CWR collections in order to detect allelic variants of candidate genes controlling traits of interest (Ramkumar et al. 2016).
The use of the so-called ‘new plant breeding techniques’ can be useful for introgressiomics (Lusser et al. 2011; Hartung and Schiemann 2014) as well. In this respect, cis-genesis (Jacobsen and Schouten 2007) and genome editing, in particular based on CRISPR/Cas9 (Belhaj et al. 2013), are promising. Cis-genesis consists of the genetic transformation of a recipient parent, in the case of introgressiomics the crop, with isolated genes and their promoters from a crossable donor (CWR), without the introduction of reporter or selectable markers from other organisms (Jacobsen and Schouten 2007). In this way, by using cis-genesis, genes isolated from CWRs can be transferred in the genetic background of the crop without linkage drag (Tardi 2016). Theoretically, by introducing genes isolated from CWRs from different genepools it would be possible to produce collections of isogenic lines with genes from different donors, as well as to pyramid or to ‘stack’ genes from different species in a single genotype (Jo et al. 2014). This is particularly interesting in the case of secondary and tertiary genepool species, with strong hybridization barriers (Khush and Brar 1992; Dwivedi et al. 2008). For example, Jo et al. (2014) introduced two genes for resistance to late blight from the potato CWRs S. stoloniferum and S. venturii in several potato varieties. Several techniques exist for genome editing in plants (Cardi 2016), but among them currently the most promising is the CRISPR/Cas9 system (Kole et al. 2015). In this way, it will potentially be possible to introgress traits whose expression depends on one or a few functional SNPs.
Cis-genic materials are genetically indistinguishable from those obtained by induced translocation breeding (Jacobsen and Schouten 2007) and CRISPR/Cas edited genes cannot be distinguished from a natural mutation at the same locus (Belhaj et al. 2013). However, legal uncertainty and the risk of non-acceptance of these technologies by consumers restrict their current usefulness (Lusser et al. 2011; Hartung and Schiemann 2014).
Moving the introgressed material into the breeding pipeline
The development of introgressiomics materials may require several years depending on the crop and trait, among others. In most occasions, the development of these types of materials cannot been achieved within the time span of a single research project (which is on average 3–5 years). Therefore, on many occasions the public introgressiomics programs may remain unfinished and the materials remain unused by the breeding sector. An important reason could be the lack of characterization and evaluation data, which is essential for breeders. In some cases, materials obtained in the public sector are available. For example, over 300 ILs and subIL of S. pennellii introgressed into cultivated tomato are available (Alkeesh et al. 2013). Also, the Tomato Genetic Resources Center (http://tgrc.ucdavis.edu/) maintains a large stock of pre-breeding materials readily usable by breeders.
We argue that long-term public–private pre-breeding partnerships (PPPPs) could bridge the gap between the development of introgressiomics materials (pre-breeding) and their utilization in subsequent breeding programs (Lusser 2014; Dempewolf et al. 2017). These PPPPs, which would involve scientists and breeders, could focus on characterising and evaluating the pre-bred materials in more detail, before breeders are willing to include them in their programs (Lusser 2014; Warschefsky et al. 2014). Currently, phenotyping is the limiting factor in using introgressiomics populations (Gur and Zamir 2004) and PPPPs could make a great contribution to an increased utilization by conducting more phenotyping experiments in diverse environments. Several successful examples of PPPPs are presented in Lusser (2014) or Moore (2015). For an efficient utilization in breeding, plant materials and related information resulting as an outcome of public–private pre-breeding partnership programs should be publically accessible as a community resource, so that they can contribute to the development of new cultivars (Campi and Nuvolari 2015). In this respect, global gateways such as Genesys (https://www.genesys-pgr.org) or international inititatives, like DivSeek, can make an important contribution to the sharing of phenotypic and genotypic characterization data of materials stored in genebanks (Meyer 2015).
We propose that introgressiomics materials that clearly belong in the ‘pre-competitive domain’ should be made available under the terms of the International Treaty for Plant Genetic Resources for Food and Agriculture (ITPGRTA). In this respect, one possibility could be to store introgressiomics materials in germplasm banks for long term conservation (Khoury et al. 2010). Clearly, the development of conservation strategies for pre-breeding materials including mapping populations, cytogenetic stocks and mutant collections would enhance the utilization of CWR and other sources of native diversity.
Conclusions
Introgression breeding from CWRs can make a major contribution to adapting our crops to climate change as well as to the development of new generations of crops with new and improved properties. The new approach we propose, ‘introgressiomics’, calls for an increased and most efficient use of CWRs in breeding by using the available genetic resources, introgression and population development schemes, phenotyping and genotyping tools and integration of data for obtaining elite materials that can be readily incorporated by breeders in their breeding programs. We hope that by using this holistic approach the practical use of CWRs in developing new commercial varieties with better characteristics will be enhanced. This may result in having more diverse, resilient and resource efficient crops that can contribute to a more sustainable and productive agriculture under the environmental changes resulting from climate change.
References
Abberton M, Batley J, Bentley A, Bryant J, Cai H, Cockram J, Costa de Oliveira A, Cseke LJ, Dempewolf H, De Pace C, Edwards D, Gepts P, Greenland A, Hall A, Henry J, Hori K, Howe GT, Hughes S, Humphreys M, Lightfoot D, Marshall A, Mayes S, Nguyen HT, Ogbonnaya FC, Ortiz R, Paterson AH, Tuberosa R, Valliyodan B, Varshney RK, Yano M (2016) Global agricultural intensification during climate change: a role for genomics. Plant Biotechnol J 14:1095–1098. doi:10.1111/pbi.12467
Aflitos S, Schiljen E, de Jong H, de Ridder D, Smit S, Finkers R, Wang J, Zhang G, Li N, Mao L, Bakker F, Dirks R, Breit T, Gravendeel B, Huits H, Struss D, Swanson-Wagner R, van Leeuwen H, van Ham RCHJ, Fito L, Guignier L, Sevilla M, Ellul P, Ganko E, Kapur A, Reclus M, de Geus B, van de Geest H, te Lintel Hekkert B, van Haarst J, Smits L, Koops A, Sanchez-Perez G, van Heusden AW, Visser R, Quan Z, Min J, Liao L, Wang X, Wang G, Yue Z, Yang X, Xu N, Schranz E, Smets E, Vos R, Rauwerda J, Ursem R, Schuit C, Kerns M, van den Berg J, Vriezen W, Janssen A, Datema E, Jahrman T, Moquet F, Bonnet J, Peters S (2014) Exploring genetic variation in the tomato (Solanum section Lycopersicon) clade by whole-genome sequencing. Plant J 80:136–148. doi:10.1111/tpj.12616
Alexander LJ (1963) Transfer of a dominant type of resistance to the four known Ohio pathogenic strains of tobacco mosaic virus (TMV) from Lycopersicon peruvianum to L. esculentum. Phytopathology 53:869
Alfares W, Bouguennec A, Balfourier F, Gay G, Bergès H, Vautrin S, Sourdille P, Bernard M, Feuillet C (2009) Fine mapping and marker development for the crossability gene SKr on chromosome 5BS of hexaploid wheat (Triticum aestivum L.). Genetics 183:469–481. doi:10.1534/genetics.109.107706
Alseekh S, Ofner I, Pleban T, Tripodi P, Di Dato F, Cammareri M, Mohammad A, Grandillo S, Fernie AR, Zamir D (2013) Resolution by recombination: breaking up Solanum pennellii introgressions. Trends Plant Sci 18:536–538. doi:10.1016/j.tplants.2013.08.003
Bari A, Street K, Mackay M, Endresen DTF, De Pauw E, Amri A (2012) Focused Identification of Germplasm Strategy (FIGS) detects wheat stem rust resistance linked to environmental variables. Genet Resour Crop Evol 59:1465–1481. doi:10.1007/s10722-011-9775-5
Baute GJ, Dempewolf H, Rieseberg L (2015) Using genomic approaches to unlock the potential of CWR for crop adaptation to climate change. In: Redden R, Yadav S, Maxted N, Dulloo ME, Guarino L, Smith P (eds) Crop wild relatives and climate change. Wiley, Hoboken, pp 268–280. doi:10.1002/9781118854396.ch15
Bebber DP, Ramotowski MAT, Gurr SJ (2013) Crop pests and pathogens move polewards in a warming world. Nat Clim Change 3:985–988. doi:10.1038/nclimate1990
Bedő Z, Láng L (2015) Wheat breeding: current status and bottlenecks. In: Molnár-Láng M, Ceoloni C, Doležel J (eds) Alien introgression in wheat. Springer, Berlin Heidelberg, pp 77–101. doi:10.1007/978-3-319-23494-6_3
Belhaj K, Chaparro-Garcia A, Kamoun S, Nekrasov V (2013) Plant genome editing made easy: targeted mutagenesis in model and crop plants using the CRISPR/Cas system. Plant Methods 9:39. doi:10.1186/1746-4811-9-39
Bessey CE (1906) Crop improvement by utilizing wild species. J Hered os-2:112–118. doi:10.1093/jhered/os-2.1.112
Brozynska M, Furtado A, Henry RJ (2016) Genomics of crop wild relatives: expanding the gene pool for crop improvement. Plant Biotech J 14:1070–1085. doi:10.1111/pbi.12454
Campi M, Nuvolari A (2015) Intellectual property protection in plant varieties: a worldwide index (1961-2011). Res Policy 44:951–964. doi:10.1016/j.respol.2014.11.003
Cardi T (2016) Cisgenesis and genome editing: combining concepts and efforts for a smarter use of genetic resources in crop breeding. Plant Breed 135:139–147. doi:10.1111/pbr.12345
Castañeda-Alvarez NP, Khoury C, Achicanoy HA, Bernau V, Dempewolf H, Eastwood RJ, Guarino L, Harker RH, Jarvis A, Maxted N, Müller JV, Ramirez-Villegas J, Sosa CC, Struik PC, Vincent H, Toll J (2016) Global conservation priorities for crop wild relatives. Nat Plants 2:16022. doi:10.1038/nplants.2016.22
Cattivelli L, Rizza F, Badeck FW, Mazzucotelli E, Mastrangelo AM, Francia E, Marè C, Tondelli A, Stanca AM (2008) Drought tolerance improvement in crop plants: an integrated view from breeding to genomics. Field Crops Res 105:1–14. doi:10.1016/j.fcr.2007.07.004
Cavanagh C, Morell M, Mackay I, Powell W (2008) From mutations to MAGIC: resources for gene discovery, validation and delivery in crop plants. Curr Opinion Plant Biol 11:215–221. doi:10.1016/j.pbi.2008.01.002
Centro Internacional de Agricultura Tropical (2017) A global database for the distributions of crop wild relatives. doi: 10.15468/jyrthk. Accessed via http://www.gbif.org/dataset/07044577-bd82-4089-9f3a-f4a9d2170b2e on 2017-03-03
Cowling WA, Buirchell BJ, Falk DE (2009) A model for incorporating novel alleles from the primary gene pool into elite crop breeding programs while reselecting major genes for domestication or adaptation. Crop Pasture Sci 60:1009–1015. doi:10.1071/CP08223
De Storme N, Mason A (2014) Plant speciation through chromosome instability and ploidy change: cellular mechanisms, molecular factors and evolutionary relevance. Curr Plant Biol 1:10–33. doi:10.1016/j.cpb.2014.09.002
de Van Wouw M, Kik C, van Hintum T, van Treuren R, Visser B (2009) Genetic erosion in crops: concept, research results and challenges. Plant Genet Resour C 8:1–15. doi:10.1017/S1479262109990062
Dempewolf H, Hodgkins KA, Rummell SE, Ellstrand NC, Rieseberg LH (2012) Reproductive isolation during domestication. Plant Cell 24:2710–2717. doi:10.1105/tpc.112.100115
Dempewolf H, Eastwood RJ, Guarino L, Khoury C, Müller JV, Toll J (2014) Adapting agriculture to climate change: a global initiative to collect, conserve, and use crop wild relatives. Agrocecol Sust Food Syst 38:369–377. doi:10.1080/21683565.870629
Dempewolf H, Baute G, Anderson J, Kilian B, Smith C, Guarino L (2017) Past and future use of wild relatives in crop breeding. Crop Sci. doi:10.2135/cropsci2016.10.0885
Dhaliwal HS (1992) Unilateral incompatibility. In: Kalloo G, Chowdhury JB (eds) Distant hybridization of crop plants. Springer, Berlin, pp 32–46. doi:10.1007/978-3-642-84306-8_3
Díez MJ, Nuez F (2008) Tomato. In: Prohens J, Nuez F (eds) Vegetables II: Fabaceae, Liliaceae, Solanaceae, and Umbelliferae. Springer, New York, pp 249–323. doi:10.1007/978-0-387-74110-9
Dodsworth S, Chase MW, Särkinen T, Knapp S, Leitch AR (2016) Using genomic repeats for phylogenomics: a case study in wild tomatoes (Solanum section Lycopersicon: Solanaceae). Bot J Linn Soc 117:96–105. doi:10.1111/bij.12612
Dwivedi SL, Upadhyaya HD, Stalker HT, Blair MW, Bertioli DJ, Nielen S, Ortiz R (2008) Enhancing crop gene pools with beneficial traits using wild relatives. Plant Breed Rev 30:179–230. doi:10.1002/9780470380130.ch3
Fita A, Rodríguez-Burruezo A, Boscaiu M, Prohens J, Vicente O (2015) Breeding and domesticating crops adapted to drought and salinity: a new paradigm for increasing food production. Front Plant Sci 6:978. doi:10.3389/fpls.2015.00978
Friebe B, Jiang J, Raupp WJ, McIntosh RA, Gill BS (1996) Characterization of wheat-alien translocations conferring resistance to diseases and pests. Euphytica 91:59–87. doi:10.1007/BF00035277
Friebe B, Qi L, Liu C, Liu W, Gill BS (2012) Registration of a hard red winter wheat genetic stock homozygous for ph1b for facilitating alien introgression for crop improvement. J Plant Regist 6:121–123. doi:10.3198/jpr2011.05.0273crgs
Furini A, Wunder J (2004) Analysis of eggplant (Solanum melongena)-related germplasm: morphological and AFLP data contribute to phylogenetic interpretations and germplasm utilization. Theor Appl Genet 108:197–208. doi:10.1007/s00122-003-1439-1
Gerstetter C, Görlach B, Neumann K, Schaffrin D (2007) The International Treaty on Plant Genetic Resources for Food and Agriculture within the current legal regime complex on plant genetic resources. J World Intellect Prop 10:259–283. doi:10.1111/j.1747-1796.2007.00323.x
Godfray HCJ, Beddington JR, Crute IR, Haddad L, Lawrence D, Muir JF, Pretty J, Robinson S, Thomas SM, Toulmin C (2010) Food security: the challenge of feeding 9 billion people. Science 327:812–817. doi:10.1126/science.1185383
Guerrero RF, Posto AL, Moyle LC, Hahn MW (2016) Genome-wide patterns of regulatory divergence revealed by introgression lines. Evolution 70:696–706. doi:10.1111/evo.12875
Gupta M, Mason AS, Batley J, Bharti S, Banga S, Banga SS (2016) Molecular-cytogenetic characterization of C-genome chromosome substitution lines in Brassica juncea (L.) Czern and Coss. Theor Appl Genet 129:1153–1166. doi:10.1007/s00122-016-2692-4
Gur A, Zamir D (2004) Unused natural variation can lift yield barriers in plant breeding. PLoS Biol 2:e245. doi:10.1371/journal.pbio.0020245
Gur A, Zamir D (2015) Mendelizing all components of a pyramid of three yield QTL in tomato. Front Plant Sci 6:1096. doi:10.3389/fpls.2015.01096
Haghighi KR, Ascher PD (1998) Fertile, intermediate hybrids between Phaseolus vulgaris and P. acutifolius hybrids from congruity backcrossing. Sex Plant Reprod 1:51–58. doi:10.1007/BF00227023
Hajjar R, Hodgkin T (2007) The use of crop wild relatives in crop improvement: a survey of developments over the last 20 years. Euphytica 156:1–13. doi:10.1007/s10681-007-9363-0
Hammer K (1984) Das Domestikationssyndrom. Kulturpfl 32:11–34. doi:10.1007/BF02098682
Harlan JR, de Wet JMJ (1971) Toward a rational classification of cultivated plants. Taxon 20:509–517. doi:10.2307/1218252
Hartung F, Schiemann J (2014) Precise plant breeding using new genome editing techniques: opportunities, safety and regulation in the EU. Plant J 78:742–752. doi:10.1111/tpj.12413
Herzog E, Falke KC, Presteri T, Scheuermann Ouzunova M, Frisch M (2014) Selection strategies for the development of maize introgression populations. PLoS ONE 9:e92429. doi:10.1371/journal.pone.0092429
Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A (2005) Very high resolution interpolated climate surfaces for global land areas. Int J Clim 25:1965–1978. doi:10.1002/joc.1276
Jacobsen E, Schouten HJ (2007) Cisgenesis strongly improves introgression breeding and induced translocation breeding of plants. Trends Biotech 25:219–223. doi:10.1016/j.tibtech.2007.03.008
Jarvis DI, Hodgkin T (1999) Wild relatives and crop cultivars: detecting natural introgression and farmer selection of new genetic combinations in agroecosystems. Mol Ecol 8:S159–S173. doi:10.1046/j.1365-294X.1999.00799.x
Jarvis A, Lane A, Hijmans RI (2008) The effect of climate change on crop wild relatives. Agric Ecosyst Environ 126:13–23. doi:10.1016/j.agee.2008.01.013
Jo KR, Kim CJ, Kim SJ, Kim TY, Bergervoet M, Jongsma MA, Visser RGF, Jacobsen E, Vossen JH (2014) Development of late blight resistant potatoes by cisgene stacking. BMC Biotech 14:50. doi:10.1186/1472-6750-14-50
Johnson AAT, Veilleux RE (2000) Somatic hybridization and applications in plant breeding. Plant Breed Rev 20:167–225. doi:10.1002/9780470650189.ch6
Jones TA (2003) The restoration gene pool concept: beyond the native versus non-native debate. Restor Ecol 11:281–290. doi:10.1046/j.1526-100X.2003.00064.x
Kantar MB, Sosa CS, Khoury CK, Castañeda-Álvarez NP, Achicanoy HA, Bernau V, Kane NC, Marek L, Seiler G, Rieseberg LH (2015) Ecogeography and utility to plant breeding of the crop wild relatives of sunflower (Helianthus annuus L.). Front. Plant Sci 6:841. doi:10.3389/fpls.2015.00841
Khan MMR, Hasnunnahar M, Iwayoshi M, Ogura-Tsujira Y, Isshiki S (2015) Pollen degeneration in three functional male-sterile lines of eggplant with the wild Solanum cytoplasms. Hortic Environ Biotech 56:350–357. doi:10.1007/s13580-015-0015-3
Khazaei H, Street K, Bari A, Mackay M, Stoddard FL (2013) The FIGS (Focused Identification of Germplasm Strategy) approach identifies traits related to drought adaptation in Vicia faba genetic resources. PLoS ONE 8:e63107. doi:10.1371/journal.pone.0063107
Khlestkina EK (2014) Current applications of wheat and wheat-alien precise genetic stocks. Mol Breed 34:273–281. doi:10.1007/s11032-014-0049-8
Khoury C, Laliberté B, Guarino L (2010) Trends in ex situ conservation of plant genetic resources: a review of global crop and regional conservation strategies. Genet Resour Crop Evol 57:625–639. doi:10.1007/s10722-010-9534-z
Khush GS, Brar DS (1992) Overcoming the barriers in hybridization. In: Kalloo G, Chowdhury JB (eds) Distant hybridization of crop plants. Springer, Berlin, pp 47–61. doi:10.1007/978-3-642-84306-8_4
Kilian B, Mammen K, Millet E, Sharma R, Graner A, Salamini F, Hammer K, Özkan H (2011) Aegilops. In: Kole C (ed) Wild Crop Relatives: genomic and breeding resources, cereals. Springer, Berlin Heidelberg, pp 1–76. doi:10.1007/978-3-642-14228-4_1
King J, Grewal S, Yang C, Hubbart S, Scholefield D, Ashling S, Edwards KJ, Allen AM, Burridge A, Bloor C, Davassi A, da Silva GJ, Chalmers K, King IP (2016) A step change in the transfer of interspecific variation into wheat from Amblyopyrum muticum. Plant Biotech. doi:10.1111/pbi.12606
Knox J, Hess T, Daccache A, Wheeler T (2012) Climate change impacts on crop productivity in Africa and South Asia. Environ Res Lett 7:034032. doi:10.1088/1748-9326/7/3/034032
Kole C, Muthamilarasan M, Henry R, Edwards D, Sharma R, Abberton M, Batley J, Bentley A, Blakeney M, Bryant J, Cai H, Cakir M, Cseke LJ, Cockram J, de Oliveira AC, De Pace C, Dempewolf H, Ellison S, Gepts P, Greenland A, Hall A, Hori K, Highes S, Humphreys MW, Iorizzo M, Ismail AB, Marshall A, Mayes S, Nguyen HT, Ogbonnaya FC, Ortiz R, Paterson AH, Simon PW, Tohme J, Tuberosa R, Valliyodan B, Varshney RK, Wullschleger SD, Yano M, Prasad M (2015) Application of genomics-assisted breeding for generation of climate resilient crops: progress and prospects. Front Plant Sci 6:563. doi:10.3389/fpls.2015.00563
Lippman ZB, Semel Y, Zamir D (2007) An integrated view of quantitative trait variation using tomato interspecific introgression lines. Curr Opin Genet Dev 17:545–552. doi:10.1016/j.gde.2007.07.007
Longin CFH, Reif JC (2014) Redesigning the exploitation of wheat genetic resources. Trends Plant Sci 19:631–636. doi:10.1016/j.tplants.2014.06.012
Lusser M (2014) Workshop on public-private partnerships in plant breeding. Science and Policy Report by the Joint Research Centre of the European Commission. Publications Office of the European Union, Luxembourg. doi:10.2791/80891
Lusser M, Parisi C, Plan D, Rodríguez-Cerezo E (2011) New plant breeding techniques: state-of-the –art and prospects for commercial development. Reference Report by the Joint Research Centre of the European Commission. Publications Office of the European Union, Luxembourg. doi:10.2791/60346
Martín A, Alvarez JB, Martín LM, Barro F, Ballesteros J (1999) The development of tritordeum: a novel cereal for food processing. J Cereal Sci 30:85–95. doi:10.1006/jcrs.1998.0235
Maxted N, Kell S (2009) Establishment of a global network for the in situ conservation of crop wild relatives: Status and needs. Commission on Genetic Resources for Food and Agriculture. FAO, Rome
Maxted N, Ford-Lloyd BV, Jury S, Kell S, Scholten M (2006) Towards a definition of a crop wild relative. Biodivers Conserv 15:2673–2685. doi:10.1007/s10531-005-5409-6
Maxted N, Dulloo E, Ford-Lloyd BV, Iriondo JM, Jarvis A (2008) Gap analysis: a tool for complementary genetic conservation assessment. Divers Distrib 14:1018–1030. doi:10.1111/j.1472-4642.2008.00512.x
McIntosh RA (1992) Pre-emptive breeding to control wheat rusts. Euphytica 63:103–113. doi:10.1007/BF00023916
McKhann HI, Camilleri C, Bérard A, Bataillon T, David JL, Reboud X, Le Corre V, Caloustian C, Gut IG, Brunel D (2004) Nested core collections maximizing genetic diversity in Arabidopsis thaliana. Plant J 38:193–202. doi:10.1011/j.1365-313X.2004.02034.x
Menda N, Strickler SR, Edwards JD, Bombarely A, Dunham DM, Martin GB, Mejia L, Hutton SF, Havey MJ, Maxwell DP, Mueller LA (2014) Analysis of wild-species introgressions in tomato inbreds uncover ancestrals origins. BMC Plant Biol 14:287. doi:10.1186/s12870-014-0287-2
Meyer RS (2015) Encouraging metadata curation in the diversity seek initiative. Nat Plants 1:15099. doi:10.1038/nplants.2015.99
Meyer RS, Purugganan MD (2013) Evolution of crop species: genetics of domestication and diversification. Nature Rev Genet 14:840–852. doi:10.1038/nrg3605
Meyer RS, DuVal AE, Jensen HR (2012) Patterns and processes in crop domestication: an historical review and quantitative analysis of 203 global food crops. New Phytol 196:29–48. doi:10.1111/j.1469-8137.2012.04253.x
Moore G (2015) Strategic pre-breeding for wheat improvement. Nature Plants 1:15018. doi:10.1038/nplants.2015.18
Muñoz LC, Blair MW, Duque MC, Tohme J, Roca W (2004) Introgression in common bean × tepary bean interspecific congruity-backcross lines as measured by AFLP markers. Crop Sci 44:637–645. doi:10.2135/cropsci2004.6370
Pascual L, Albert E, Sauvage C, Duangjit J, Bouchet JP, Bitton F, Desplat N, Brunel D, Le Paslier MC, Ranc N, Bruguier L, Chauchard B, Verschave P, Causse M (2016) Dissecting quantitative trait variation in the resequencing era: complementarity of bi-parental, multi-parental and association panels. Plant Sci 242:120–130. doi:10.1016/j.plantsci.2015.06.017
Peleg Z, Fahima T, Krugman T, Abbo S, Yakir D, Korol AB, Saranga Y (2009) Genomic dissection of drought resistance in durum wheat × wild emmer wheat recombinant inbreed line population. Plant Cell Environ 32:758–779. doi:10.1111/j.1365-3040.2009.01956.x
Peleman JD, van der Voort JR (2003) Breeding by design. Trends Plant Sci 8:330–334. doi:10.1016/S1360-1385(03)00134-1
Pérez-de-Castro AM, Vilanova S, Cañizares J, Pascual L, Blanca JM, Díez MJ, Prohens J, Picó B (2012) Application of genomic tools in plant breeding. Curr Genom 13:179–195. doi:10.2174/138920212800543084
Plazas M, Vilanova S, Gramazio P, Rodríguez-Burruezo A, Fita A, Herraiz FJ, Ranil R, Fonseka R, Niran L, Fonseka H, Kouassi B, Kouassi A, Kouassi A, Prohens J (2016) Interspecific hybridization between eggplant and wild relatives from different genepools. J Am Soc Hortic Sci 141:34–44
Porch TG, Beaver JS, Debouck DG, Jackson SA, Kelly JD, Dempewolf H (2013) Use of wild relatives and closely related species to adapt common bean to climate change. Agronomy 3:433–461. doi:10.3390/agronomy3020433
Prakash S, Ahuja I, Upreti HC, Kumar VD, Bhat SR, Kirti PB, Chopra VL (2001) Expression of male sterility in alloplasmic Brassica juncea with Erucastrum canariense cytoplasm and the development of a fertility restoration system. Plant Breed 120:479–482. doi:10.1046/j.1439-0523.2001.00627_x
Ramírez-Villegas J, Khoury C, Jarvis A, Debouck DG, Guarino L (2010) A gap analysis methodology for collecting crop genepools: a case study with Phaseolus beans. PLoS ONE 5:e13497. doi:10.1371/journal.pone.0013497
Ramkumar G, Madhav MS, Rama Devi SJS, Umakanth B, Pandey MK, Prasad MS, Sundaram RM, Viraktamath BC, Ravindra Babu V (2016) Identification and validation of novel alleles of rice blast resistant gene Pi54, and analysis of their nucleotide diversity in landraces and wild Oryza species. Euphytica 209:725–737. doi:10.1007/s10681-016-1666-6
Ranil RHG, Niran HML, Plazas M, Fonseka RM, Fonseka HH, Vilanova S, Andújar I, Gramazio P, Fita A, Prohens J (2015) Improving seed germination of the eggplant rootstock Solanum torvum by testing multiple factors using an orthogonal array design. Sci Hort 193:174–181. doi:10.1016/j.scienta.2015.07.030
Ray DK, Mueller ND, West PC, Foley JA (2013) Yield trends are insufficient to double global crop production by 2050. PLoS ONE 8:e66428. doi:10.1371/journal.pone.0066428
Rieseberg LH, Carney SE (1998) Plant hybridization. New Phytol 140:599–624. doi:10.1046/j.1469-8137.1998.00315.x
Rieseberg LH, Arias DM, Ungerer MC, Linder CR, Sinervo B (1996) The effects of mating designs on introgression between chromosomally divergent sunflower species. Theor Appl Genet 93:633–644. doi:10.1007/BF00417959
Rosenzweig C, Elliott J, Deryng D, Ruane AC, Müller C, Arneth A, Boote KJ, Folberth C, Glotter M, Khabarov N, Neumann K, Piontek F, Pugh TAM, Schmid E, Stehfest E, Yang H, Jones JW (2014) Assessing agricultural risks of climate change in the 21st century in a global gridded crop model intercomparison. Proc Natl Acad Sci USA 111:3268–3273. doi:10.1073/pnas.1222463110
Salamini F, Özkan H, Brandolini A, Schäfer-Pregl R, Martin W (2002) Genetics and geography of wild cereal domestication in the Near East. Nat Rev Genet 3:429–441. doi:10.1038/nrg817
Salinas M, Capel C, Alba JM, Mora B, Cuartero J, Fernández-Muñoz R, Lozano R, Capel J (2013) Genetic mapping of two QTL from the wild tomato Solanum pimpinellifolium L. controlling resistance against two-spotted spider mite (Tetranychus urticae Koch). Theor Appl Genet 126:83–92. doi:10.1007/s00122-012-1961-0
Savage JA, Haines DF, Holbrook NM (2015) The making of giant pumpkins: how selective breeding changed the phloem of Cucurbita maxima from source to sink. Plant Cell Environ 38:1543–1554. doi:10.1111/pce.12502
Schwarz D, Rouphael Y, Colla G, Venema JH (2010) Grafting as a tool to improve tolerance of vegetables to abiotic stresses: thermal stress, water stress and organic pollutants. Sci Hort 127:162–171. doi:10.1016/j.scienta.2010.09.016
Sharma D, Knott DR (1966) The transfer of leaf rust resistance from Agropyron to Triticum by irradiation. Can J Genet Cytol 8:137–143. doi:10.1139/g66-018
Sharma DR, Kaur R, Kumar K (1996) Embryo rescue in plants—a review. Euphytica 99:325–337. doi:10.1007/BF00022289
Shivanna KR, Bahadur B (2015) Efficacy of biotechnological approaches to raise wide sexual hybrids. In: Bahadur B, Rajam MV, Sahijram L, Krishnamurthy KV (eds) Plant Biology and biotechnology, vol II. Plant genomics and biotechnology. Springer, New Delhi, pp 347–362. doi:10.1007/978-81-322-2283-5_17
Sim SC, Van Deynze A, Stoffel K, Douches DS, Zarka D, Ganal MW, Chetelat RT, Hutton SF, Scott JW, Gardner RG, Panthee DR, Mutschler M, Myers JR, Francis DM (2012) High density SNP genotyping of tomato (Solanum lycopersicum L.) reveals patterns of genetic variation due to breeding. PLoS ONE 7:e45520. doi:10.1371/journal.pone.0045520
Smith PG (1944) Embryo culture of a tomato species hybrid. Proc Am Soc Hort Sci 44:413–416
Street K, Bari A, Mackay M, Amri A (2016) How the Focused Identification of Germplasm Strategy (FIGS) is used to mine plant genetic resources for adaptive traits. In: Maxted N, Dulloo ME, Ford-Lloyd BV (eds) Enhancing crop genepool use: capturing wild relative and landrace diversity for crop improvement. CABI, Wallingford, pp 54–65. doi:10.1079/9781780646138.0054
Syfert M, Castañeda-Álvarez NP, Khoury C, Särkinen T, Sosa CC, Achicanoy HA, Bernau V, Prohens J, Daunay MC, Knapp S (2016) Crop wild relatives of the brinjal eggplant (Solanum melongena): poorly represented in genebanks and many species at risk of extinction. Am J Bot 103:635–651. doi:10.3732/ajb.1500539
Tanksley SD, McCouch SR (1997) Seed banks and molecular maps: unlocking genetic potential from the wild. Science 277:1063–1066. doi:10.1126/science.277.5329.1063
Tanksley SD, Nelson JC (1996) Advanced backcross QTL analysis: a method for the simultaneous discovery and transfer of valuable QTLs from unadpated germplasm into elite breeding lines. Theor Appl Genet 92:191–203. doi:10.1007/BF00223376
Tilman D, Balzer C, Hill J, Befort BL (2011) Global food demand and the sustainable intensification of agriculture. Proc Natl Acad Sci USA 108:20260–20264. doi:10.1073/pnas.1116437108
Trethowan RM, Mujeeb-Kazi A (2008) Novel germplasm resources for improving environmental stress tolerance of hexaploid wheat. Crop Sci 48:1255–1265. doi:10.2135/cropsci2007.08.0477
Valkoun JJ (2001) Wheat pre-breeding using wild progenitors. Euphytica 119:17–23. doi:10.1023/A:1017562909881
Verlaan MG, Szinay D, Hutton SF, de Jong H, Kormelink R, Visser RGF, Scott JW, Bai Y (2011) Chromosomal rearrangements between tomato and Solanum chilense hamper mapping and breeding of TYLCV resistance gene Ty-1. Plant J 68:1093–1103. doi:10.1111/j.1365-313X.2011.04762.x
Villegas D, Casadesús J, Atienza SG, Martos V, Maalouf F, Karam F, Aranjuelo I, Nogués S (2010) Tritordeum, wheat and triticale yield components under multi-local Mediterranean drought conditions. Field Crops Res 116:68–74. doi:10.1016/j.fcr.2009.11.012
Vincent H, Wiersma J, Kell S, Fielder H, Dobbie S, Castañeda-Álvarez NP, Guarino L, Eastwood R, León B, Maxted N (2013) A prioritized crop wild relative inventory to help underpin global food security. Biol Conserv 167:265–275. doi:10.1016/j.biocon.2013.08.011
Vorontsova MS, Stern S, Bohs L, Knapp S (2013) African spiny Solanum (subgenus Leptostemonum, Solanaceae): a thorny phylogenetic tangle. Bot J Linn Soc 173:176–193. doi:10.1111/boj.12053
Wall JR (1970) Experimental introgression in the genus Phaseolus. I. Effect of mating systems on interspecific gene flow. Evolution 24:356–366
Warburton ML, Crossa J, Franco J, Kazi M, Trethowan R, Rajaram S, Pfeiffer W, Zhang P, Dreisigacker S, van Ginkel M (2006) Bringing wild relatives back into the family: recovering genetic diversity in CIMMYT improved wheat germplasm. Euphytica 149:289–301. doi:10.1007/s10681-005-9077-0
Warschefsky E, Penmetsa RV, Cook DR, von Wettberg EJB (2014) Back to the wilds: tapping evolutionary adaptations for resilient crops through systematic hybridization with crop wild relatives. Am J Bot 101:1791–1800. doi:10.3732/ajb.1400116
Wendler N, Mascher M, Himmelbach A, Johnston P, Pickering R, Stein N (2015) Bulbosum to go: a toolbox to utilize Hordeum vulgare/bulbosum introgressions for breeding and beyond. Mol Plant 8:1507–1519. doi:10.1016/j.molp.2015.05.004
Wulff BBH, Moscou MJ (2014) Strategies for transferring resistance into wheat: from wide crosses to GM cassettes. Front Plant Sci 5:692. doi:10.3389/fpls.2014.00692
Zamir D (2001) Improving plant breeding with exotic genetic libraries. Nature Rev Genet 2:983–989. doi:10.1038/35103590
Zamir D, Ekstein-Michelson I, Zakay Y, Navot N, Zeidan M, Sarfatti M, Eshed Y, Harel E, Pleban T, van-Oss H, Kedar N, Rabinowitch HD, Czosnek H (1994) Mapping and introgression of a tomato yellow leaf curl virus tolerance gene, TY-1. Theor Appl Genet 88:141–146. doi:10.1007/BF00225889
Zenkteler M (1990) In vitro fertilization and wide hybridization in higher plants. Crit Rev Plant Sci 9:267–279. doi:10.1080/07352689009382290
Acknowledgements
This work was undertaken as part of the initiative “Adapting Agriculture to Climate Change: Collecting, Protecting and Preparing Crop Wild Relatives”, which is supported by the Government of Norway. The Project is managed by the Global Crop Diversity Trust with the Millennium Seed Bank of the Royal Botanic Gardens, Kew and implemented in partnership with national and international gene banks and plant breeding institutes around the world. For further information see the project website: http://www.cwrdiversity.org/. This work has also been funded in part by European Union’s Horizon 2020 research and innovation programme under Grant agreement No 677379 (G2P-SOL) and from Spanish Ministerio de Economía y Competitividad and Fondo Europeo de Desarrollo Regional (Grant AGL2015-64755-R from MINECO/FEDER, EU). Pietro Gramazio is grateful to Universitat Politècnica de València for a pre-doctoral (Programa FPI de la UPV-Subprograma 1/2013 call) contract.
Author information
Authors and Affiliations
Corresponding author
Additional information
This article is part of the Topical Collection on Plant Breeding: the Art of Bringing Science to Life. Highlights of the 20th EUCARPIA General Congress, Zurich, Switzerland, 29 August–1 September 2016
Edited by Roland Kölliker, Richard G. F. Visser, Achim Walter & Beat Boller
Rights and permissions
About this article

Cite this article
Prohens, J., Gramazio, P., Plazas, M. et al. Introgressiomics: a new approach for using crop wild relatives in breeding for adaptation to climate change. Euphytica 213, 158 (2017). https://doi.org/10.1007/s10681-017-1938-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10681-017-1938-9