
Abstract
The effect of forest management on biodiversity is a crucial issue for sustainable forestry and nature conservation. However, the ways in which management affects macrofungal and plant communities and diversity of mountain temperate forests still remain poorly understood. We performed a random sampling stratified by stand age and stand type on the sites of temperate montane fir–beech forests. Diversity of macrofungi and the vascular plant understorey in beech- and spruce-dominated managed stands was investigated and compared to primeval forests located in the Poľana Biosphere Reserve, Western Carpathians. Both the vascular plant and the macrofungal communities were altered by management, and the response of the macrofungal species (especially wood-inhabiting fungi) was more pronounced in terms of species composition change. Species turnover evaluation seems to be an important tool of forest natural status assessment, because alpha diversity did not change as much as species composition. Certain species of Carpathian primeval forests were confirmed as good indicators for natural forest change; others were proposed. Species pool and mean number of species per plot were the highest in unmanaged fir–beech forests, and species diversity significantly decreased in spruce plantations. The number of species decreased significantly due to the change of canopy tree species composition only in the macrofungal communities. As an outcome for forest management, we recommend keeping mixed forests involving all natural tree species and providing at least a minimal amount of dead wood necessary for wood-inhabiting organisms and leaving some area of unmanaged natural forests within complexes of managed stands.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Direct anthropogenic influence on forests is realized through the changes in stand structure and tree species composition. Altered forest stand structure and tree species composition affect forest understorey (Barbier et al. 2008; Máliš et al. 2012; Hofmeister et al. 2014). The influence of particular tree species on the understorey organisms and soil is realized by the change of litter quality and quantity (Barbier et al. 2008), change of microclimate and soil moisture (Chavez and Macdonald 2010) and chemical and physical properties (Schoenholtz et al. 2000; Zhang et al. 2010). Despite the fact that a majority of forests in the temperate zone is managed, we know little about long-term management effects on different forest organisms (Thomas et al. 1999) and the functioning of the entire ecosystem. Therefore, an increasing number of studies is aimed at comparing the diversity of managed and unmanaged stands (Abs et al. 2005; Rudolf and Rasmussen 2013) and search for bioindicators of environmental changes and natural forests (Lindenmayer et al. 2000; Liira and Kohv 2010; Blasi et al. 2010).
Effects of forest management on plant species pool (γ-diversity) and species richness (α-diversity) have been referred to by many authors (e.g. Paillet et al. 2010; Durak 2012; Schulze et al. 2015). The clear-cut system was reported to affect diversity the most, while more close-to-nature techniques such as selective cutting may not significantly alter species composition of the undergrowth (Battles et al. 2001). Moreover, diversity of understorey plants varies over time in even-aged stands (Gilliam and Roberts 1995; Ujházy et al. 2017). It usually increases from young stands towards the oldest (Halpern and Spies 1995). The most species-rich stages are those of open clearings (Swanson et al. 2011), while it is the lowest in the young pole stage as well as in the growing stage of primeval forest (Ujházy et al. 2009). On the other hand, several case studies have shown that cessation of management leads to a decrease in species diversity due to a decline of the heliophilous plant species of more open forests (Ujházyová and Ujházy 2007; Hédl et al. 2010).
Effects on other organisms including fungi are less known (Paillet et al. 2010). In contrast to autotrophic vascular plants responding especially to abiotic site properties, heterotrophic fungi are linked mainly to dead organic matter—terrestrial saprotrophs, and/or to particular tree species—ectomycorrhizal species (Dix and Webster 2012). The anthropogenic influence on fungal diversity varies according to the duration and intensity of management (Brunet et al. 2010; Bässler et al. 2014), and macrofungal species richness in both managed and unmanaged forests also depends on forest age (Hofmeister et al. 2014). Increasing intensity of management causes macrofungal diversity decrease and shifts in species composition (Bässler et al. 2014). Many macrofungal species are threatened by the reduction of natural and close-to-natural forests (Holec and Beran 2006). Dahlberg and Croneborg (2003) estimated that up to 20% of European species are threatened by the reduction of habitats and by the degradation of the environment. Therefore, macrofungi can serve as indicators of ecological continuity and the natural character of forest ecosystems (Nordén and Appelqvist 2001; Christensen et al. 2004; Holec 2008; Dvořák et al. 2017). However, individual trophic groups of macrofungi respond differently to forest management (Dvořák et al. 2017). In particular, wood-inhabiting fungi, whose diversity is significantly dependent on the amount of dead wood (Persiani et al. 2015), have frequently been proposed as good indicators of natural forests (Christensen et al. 2004; Küffer et al. 2008; Blaser et al. 2013; Dvořák et al. 2017). Therefore, diversity of wood-inhabiting fungi has frequently been recently studied (Ódor et al. 2006; Brunet et al. 2010; Saitta et al. 2011; Granito and Lunghini 2011; Granito et al. 2015; Dvořák et al. 2017).
Understorey plant and fungal communities may show contradictory responses to the same management influence. It is still unclear if their communities respond independently or if they are mutually related. The relationship between macrofungal and plant communities (richness and species composition) was studied with contradictory results, showing both positive and negative relationships (Sæetersdal et al. 2003; Chiarucci et al. 2005; Brown et al. 2006; McMullen-Fisher et al. 2010; Hofmeister et al. 2014). According to some authors, the relationship between plant and fungal diversity is doubtful, and diversity of plants cannot be used as a proxy for macrofungal diversity in general (Berglund and Jonsson 2001; Kinga et al. 2013). However, positive correlations were sometimes found depending on forest stand types studied, similarity of site conditions and intensity of management (Sæetersdal et al. 2003; Schmit and Lodge 2005; McMullen-Fisher et al. 2010; Hofmeister et al. 2014).
To elucidate the above-mentioned problems, we performed a study of the two important components of forest ecosystems representing a major part of the biomass—plants and macrofungi on the community level. To avoid the ambiguity of previous studies, we realized representative sampling of both groups within a single series of plots of the same size under homogeneous site conditions. The aim is to identify the response of both macrofungal and plant communities under the same site conditions to man-induced changes in the overstorey structure and to assess the real impact of forest management on the species diversity of the two taxonomic groups of understorey organisms. We studied two most widespread types of managed age-class stands of the Central European mountain ranges in contrast to one of the largest and best preserved remnants of Carpathian primeval forests. In particular, we addressed the following questions:
-
(1)
does overstorey change affect species composition of vascular plants and macrofungi?
-
(2)
which species of vascular plants and macrofungi are indicators of primeval forest change?
-
(3)
does species diversity of both taxonomic groups differ among different stand types?
-
(4)
is there a congruence in response of plant and macrofungal communities to the forest management?
Methods
Study area and sampling design
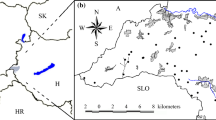
The study area is located in the Poľana Mts.—a tertiary volcanic massif in Central Slovakia, the Western Carpathians (Fig. 1). Research plots are situated both in unmanaged primeval forests of the Zadná Poľana National Nature Reserve and in the neighbouring managed age-class forests. Mean annual air temperature under a montane climate is about 3.7 °C, and mean annual precipitation ranges between 900 and 1200 mm (Šťastný et al. 2002). Eutric Cambisols with transitions to Andosols are developed on the andesite bedrock here. A series of 65 circular plots with an area of 314 m2 (10 m radius) were set and sampled in 2013 and 2014 under comparable site conditions using stratified random sampling within the above-mentioned area. Environmental conditions were restricted to the site type of montane mesotrophic fir–beech forests on south-eastern aspect, 10°–35° slopes (excluding ridges, valleys, wet and rocky sites) and the elevation zone of 950–1250 m a.s.l.

Plot distribution within managed and unmanaged forests in the Poľana Mts. (black circles unmanaged stands, white circles managed beech stands, grey circles managed spruce stands). Boundary of the Zadná Poľana Reserve is depicted by the solid line
Similar numbers of plots were established in three stand types using pre-selection of plot positions in GIS according to actual and historical orthophotomaps, digital terrain model, forest inventory maps and preliminary field mapping. Twenty-one plots were randomly situated in unmanaged primeval forests of the Zadná Poľana National Nature Reserve, formed by mixed, uneven-aged and highly structured stands of European beech (Fagus sylvatica), silver fir (Abies alba), sycamore maple (Acer pseudoplatanus), Norway spruce (Picea abies), with an admixture of common ash (Fraxinus excelsior) and wych elm (Ulmus glabra; Table 1). Large gaps without tree canopy were omitted from the selection. High amount of dead wood of all present tree species is typical of all developmental stages in the reserve.
In the managed area, 24 beech-dominated secondary stands originating from natural regeneration were randomly selected, whereas 20 plots were situated within planted spruce stands. After the regeneration phase, both stand types were managed under the same silvicultural system. To represent a stand chronosequence and to compare similar stages of stand development, random selection was performed within four age classes of forest inventory (40–59, 60–79, 80–99 and 100+), and similar number of plots was selected within each age class. The length of the chronosequence was limited by common rotation time (about 100 years). The managed stands formed a spatial patchy mosaic of beech, spruce and mixed stands of various sizes. Frequent admixture of other tree species apart from dominants was present directly in the plots or in their close neighbourhood. A large amount of thin dead wood was frequently left in the thinning phase in contrast to older stands where a majority of dead wood was logged.
Sampling of overstorey structure and environmental conditions
Trees with DBH ≥ 7 cm were recorded. Overstorey age of the managed age-class stands was determined according to wood cores taken from the biggest canopy tree. Uneven-aged primeval stands were not cored. Hemispherical photographs were taken from the centres of plots. Leaf area index was calculated using GLA 2.0 (Frazer et al. 1999). Actual soil moisture was measured at all plots on the same day by the ThetaProbe ML2x soil moisture sensors. Ten measurements were performed within each plot area, and average values were used for analyses. Soil samples were taken from three positions within each plot to determine the properties of the humus (O) and topsoil (A) horizons (0–10 cm). Soil reaction of the O and A horizons (pH KCl) was determined using the WTW inoLab pH 720 digital pH meter, and total C and N content by the CNS analyser. Percentage cover of dead wood (diameter > 3 cm) was estimated. A complete list of dendrometric and environmental characteristics recorded in plots and the resulting variables is shown in Online Resource 1.
Sampling of understorey plants and macrofungi
Vascular plants were recorded in 314 m2 plots twice, in the late spring and the summer using a semi-quantitative cover-abundance scale. Phytosociological relevés were saved in the Turboveg database (Hennekens and Schaminee 2001). In the analyses, all plant species recorded at both dates were used. Percentage cover values of vascular plants were weighted as follows: 0.7 summer cover values + 0.3 spring cover values. The nomenclature of vascular plants was unified according to Marhold and Hindák (1998).
Fungal sampling was carried out six times within 2 years—from April 2013 to November 2014, to identify as many macrofungal species as possible. The research was focused on above-ground macrofungal fruiting bodies collected mainly from the forest floor, litter and living and dead wood. Most species were identified directly in the field, and some species were collected for latter identification, archived and identified by mycologists specializing in the respective genera. Only 29 species of Pyrenomycetes were determined on the species level, and many other species of this group and resupinate fungi were not determined. The maximum abundance of species within a plot during the monitoring period was used as a numeric variable in analyses. The nomenclature of macrofungi was unified according to Index Fungorum database (http://www.indexfungorum.org) with three exceptions, where author’s name is provided. Taxa were classified within trophic groups according to substrate or host type as proposed by Agerer (2001), Holec and Beran (2006) and Tedersoo et al. (2010). Red list species were identified according to Lizoň (2001).
As the observed macrofungal diversity (especially ectomycorrhizal) strongly depends on sampling frequency and size, we performed a sample size-based rarefaction and extrapolation of macrofungal species diversity to assess the completeness of the sampling effort (Online Resource 2). Moreover, the completeness of the macrofungal sampling was quantified by sampling coverage (Chao et al. 2014). Sampling coverage of macrofungal trophic groups for pooled forest stands ranged between 93.7% (ectomycorrhizal fungi) and 94.9% (wood-inhabiting fungi). However, within forest stand types, e.g. for ectomycorrhizal fungi in unmanaged forest stands, sampling coverage decreased to 82.6%.
The number of species in a particular plot was calculated as a sum of species recorded in all sampling dates for both macrofungi and vascular plants.
Statistical analyses
Relevés of both macrofungal and plant communities were processed by the JUICE program (Tichý 2002), where diversity measures were calculated. Alpha diversity was expressed by a number of species per plot (sample species richness) and by Shannon’s diversity index (H′) and calculated for 20 plots within the stand type. Gamma diversity was expressed as a total number of species (species pool) recorded in all plots within a stand type. To make results comparable among stand types differing in the number of sites, we standardized the species pools to a common number of sites (i.e. 20 plots) using a sample-based rarefaction. For analysing gamma diversity, the “iNEXT” package (Hsieh et al. 2016) was used in the R environment (R Core Team 2016).
Differences in average numbers of plant and macrofungal species per plot as well as Shannon’s diversity index between the three stand types were tested using one-way analysis of variance (ANOVA). The homogeneity of variances between groups and the normality within groups were checked graphically using diagnostic plots. Pairwise comparisons among groups were performed using the Tukey’s HSD test after the global F test of stand type effect was statistically significant (p < 0.05).
Multivariate analogue of ANOVA with permutations—PERMANOVA (Anderson 2001)—was applied to test for the differences in species composition of plant and macrofungal communities among stand types. The Bray–Curtis index was used as a measure of dissimilarity, and the calculation of p values was based on 9999 permutations (McArdle and Anderson 2001). Both species matrices were log-transformed prior to the analysis to reduce the effect of dominant species. The effect of stand type on plant and macrofungal community composition was visualized by nonmetric multidimensional scaling (NMDS, Kruskal 1964). The same procedure (Bray–Curtis index, log-transformed species matrix) was applied to both species datasets. The analyses were performed in the R environment (R Core Team 2016) using the “vegan” package (Oksanen et al. 2016).
To express floristic differentiation among the stand types, the phi coefficient (Chytrý et al. 2002) was computed using the JUICE 7.0 program (Tichý 2002). Additionally, Fisher’s exact test was calculated to obtain significances. Species significantly associated with stand type with phi > 0.15 were considered as specific (differential) species of the categories.
The Mantel test (Mantel 1967) was performed to evaluate the congruence between (dis)similarities of plant and macrofungal communities in all stand types together and within each stand type separately. Both species matrices were transformed into matrices of the Bray–Curtis pairwise dissimilarities, and the Mantel statistic was calculated as the Spearman correlation coefficient between the two matrices. The permutation test with 10,000 permutations was used to assess the statistical significance of the test statistic, and the 95% bootstrapped confidence interval for the Mantel statistic was obtained from 10,000 iterations. The Mantel test was conducted in the R environment (R Core Team 2016) using the “ecodist” package (Goslee and Urban 2007).
We assessed the relationships between the number of plant and macrofungal species per plot and environmental variables using Spearman’s rank correlations for the three stand types separately. Correlations were also calculated between macrofungal trophic groups (wood-inhabiting fungi, terricolous, ectomycorrhizal and parasitic species) and environmental variables.
Results
Species composition
Species composition of both plant and macrofungal communities differed significantly among the three stand types (Table 2). Species differentiation of the communities within the stand types is illustrated by ordination diagrams (Fig. 2). Species composition of both communities in the managed stands was different compared to the unmanaged primeval forests; moreover, understorey of the spruce plantations deviated from the secondary beech forests. The macrofungal communities were apparently more differentiated compared to the plant communities (Fig. 2). Nevertheless, the response of particular macrofungal trophic groups was different (Fig. 3), and species composition shift in macrofungal communities was caused mainly by the changes of wood-inhabiting fungi.

Nonmetric MDS ordinations of macrofungal (left) and vascular plant (right) communities in relation to different stand types. The size of a centroid is relative to the number of species recorded in a given plot

Nonmetric MDS plots of wood-inhabiting (left), terricolous (middle) and ectomycorrhizal (right) macrofungal communities in relation to different stand types
Species differentiation
There were 12 constant understorey vascular plant species of primeval forests frequent in both managed stand types (Online Resource 3). No unique plant species was identified for primeval forests. The unmanaged stands were differentiated especially by higher frequencies of common beech forest species such as Asarum europaeum, Ajuga reptans, Daphne mezereum, Sanicula europaea and Veronica montana. Canopy trees Abies alba and Ulmus glabra were typical for the unmanaged stands. Juveniles of Abies alba were also common in managed forests, while juveniles of Ulmus glabra were not. On the other hand, eutrophic heliophilous geophytes such as Corydalis cava, Anemone ranunculoides, Gagea lutea and Isophyrum thalictroides had an optimum in the managed beech forests. Influence of spruce was reflected in an increasing frequency of some oligotrophic species (Sorbus aucuparia, Veronica officinalis) and occurrence of acidophilous graminoids (Calamagrostis arundinacea, Luzula luzuloides).
Only four of the most widespread macrofungal species with a frequency of over 40% in all types were found (Online Resource 4). All stand types were well differentiated by a similar number of diagnostic species. Several species preferring old-growth forests and/or primeval forests were identified in the unmanaged stands of Zadná Poľana: Camarops tubulina, Crepidotus applanatus, Inocybe petiginosa, Pseudoplectania melaena (Fr.) Sacc., Tatraea dumbirensis and Tectella patellaris. Conspicuous pyrenomycete Camarops tubulina occurred in the unmanaged stands on large decaying logs of Picea abies, Abies alba and Fagus sylvatica and once in the spruce plantation on a large spruce log. Tatraea dumbirensis was frequently found in both the unmanaged and managed beech-dominated stands, whereas only one locality was registered in the spruce plantation. Similarly, Tectella patellaris occurred at several localities in the unmanaged and managed beech stands and once in the spruce plantation. Pseudoplectania melaena (Fr.) Sacc. was linked to fir logs (found during the spring) in the unmanaged fir–beech forests. Occurrence of other species such as Panellus violaceofulvus (Batsch) Singer or rare species Rutstroemia elatina and Hymenochaete cruenta (Online Resource 4) in the unmanaged forests was also related to silver fir. Among the species preferring spruce plantations, Rutstroemia bulgarioides and Strobilurus esculentus occurred to a massive extent on spruce cones during the spring.
Diversity pattern of vascular plant and macrofungal communities
The total species pool of vascular plants recorded in all plots was 138 species (Online Resource 3) compared to 382 taxa of macrofungi determined on the species level and 29 taxa of macrofungi determined on the genus level (Online Resource 4). Up to 35% of macrofungal species were recorded within a single plot at a single date only. The species pool of macrofungal communities was the highest in the unmanaged primeval stands (Table 3).
Wood-inhabiting fungi represented 53% of total species pool, terricolous saprotrophic fungi 21%, ectomycorrhizal species 20%, parasitic species 5% (Table 3) and macrofungi of other trophic groups 1%. The species pool of wood-inhabiting fungi was the highest in unmanaged stands, but the proportions of trophic groups were similar in all stand types.
The minimum number of plant and macrofungal species per plot was observed in the managed spruce stands, while the highest average number of species of both taxonomic groups was found in the unmanaged forests. However, the only significant difference in the number of plant species was identified when comparing the managed and unmanaged beech stands. On the other hand, the average number of macrofungal species was significantly lower in the spruce plantations (Fig. 4).

Variability in the number of vascular plant (left) and macrofungal (right) species per plot (314 m2) in different stand types (each represented by 20 plots). Different letters indicate heterogeneous groups (Tukey’s post hoc test, p < 0.05). Points in boxes show medians, boxes include 25–75% of values, whiskers show the range of nonoutlying values, and circles represent outliers
Patterns of species diversity in the stand types were similar for vascular plants and macrofungi. The highest average values of Shannon’s diversity index were observed in the unmanaged stands and the lowest in the spruce plantations (Fig. 5), where the minimum values were found as well. Species diversity of the managed beech forests did not differ significantly from the unmanaged ones.

Shannon’s diversity index (H′) of vascular plants (left) and macrofungi (right) in the stand types (each represented by 20 plots). Different letters indicate heterogeneous groups (Tukey’s post hoc test, p < 0.05). Points in boxes show medians, boxes in boxplots include 25–75% of values, whiskers show the range of nonoutlying values, and circles out of boxes represent outliers
Relationships between plant and macrofungal communities
To answer the question whether both studied groups respond similarly to the overstorey change in terms of their species composition (dis)similarities, we performed the Mantel test. The plot-to-plot variation in macrofungal communities was significantly correlated with the variation in plant communities when all plots (regardless of stand type) were analysed together. This means that plots with similar macrofungal species composition also had similar plant species composition and vice versa (Table 4). Within particular stand types, a significant correlation was only found in the spruce forests, in contrast to the unmanaged stands.
Numbers of plant and macrofungal species per plot were not significantly correlated when all plots were included in the analysis (r = −0.16, p = 0.19). However, a negative correlation was significant within the spruce plantations (Fig. 6). Similarly, significant negative correlation was found between numbers of wood-inhabiting fungi and plant species per plot in the beech managed stands. All other relationships between numbers of species in fungal trophic groups and vascular plant understorey were insignificant (Online Resource 5). The negative relationships of plants and fungi in the spruce stands were related to different responses of both communities to their ageing. Whereas the number of plant species significantly increased with the stand age, the number of macrofungal species significantly decreased (Online Resource 6). This was caused especially by a rapid decline in the number of wood-inhabiting species (r = −0.51, p = 0.017) with the stand age.

Relationships between the number of vascular plant and macrofungal species per plot in different stand types. Significant relationship was found and indicated by a linear regression only in the spruce stands
Discussion
Species composition change in the managed stand types
Composition of vascular plant understorey was significantly changed in both types of managed stands as was shown also in earlier studies (Barbier et al. 2008; Molder et al. 2008; Durak 2012; Ujházy et al. 2017). We documented a significant effect of stand type on fungal communities as well. However, the response was most pronounced in the wood-inhabiting fungi in contrast to the group of ectomycorrhizal species. It is consistent with previously published results from Central European forests (Dvořák et al. 2017). Shift in species composition was the highest in spruce plantations compared to primeval mixed and managed beech forests. Similar effect of spruce plantations on epigeous fungal communities was referred to by Buée et al. (2011), and significant changes in the group of wood-inhabiting fungi as a consequence of management forms in spruce forests by Bässler et al. (2012). As a majority of macrofungal species are strongly associated with tree species, the effect of overstorey change in managed stands on their communities can be particularly explained by tree species replacement.
On the other hand, the observed changes in the species composition of macrofungi corresponded especially to the wood-inhabiting fungi with the highest proportion of species in the studied communities. Their occurrence is significantly affected by resource quality and availability in the European beech forests (Abrego et al. 2016), and therefore, changes of tree species and dead wood amount and also size of woody debris (Bässler et al. 2012) can significantly change the entire species composition of fungal communities (Lonsdale et al. 2008). A high amount of large-diameter dead wood—about 180 m3/ha (Christensen et al. 2005; Kucbel et al. 2012) is typical of Carpathian beech-dominated primeval forests. The reduction of dead wood amount and the change of its quality in the managed stands was probably the most important driver of the fungal community change. Consequently, the group of wood-inhabiting fungi was confirmed as an important indicator of the forest management influence (following Berglund and Jonsson 2008; Abrego et al. 2016; Dvořák et al. 2017).
Indicator species of primeval forests
In beech forest sites, some vascular plant species can indicate a change of overstorey composition or management interventions in forests (Ewald 2000; Šamonil and Vrška 2007; Máliš et al. 2012; Ujházy et al. 2013; Verstraeten et al. 2013). However, particular plant species show weak indication value of overstorey change in the studied area, and no species were found to be restricted to primeval forest environment. All species of primeval forests can also grow in managed stands with changed frequencies. Managed beech stands were differentiated from the primeval unmanaged forests by a significantly higher frequency of eutrophic spring geophytes. It is apparently an effect of higher beech cover in homogenous managed stands combined with the ability of the geophytes to utilize favourable light conditions and available nutrients after a rapid litter decomposition in the early spring in broadleaved forests (Schulze et al. 2005).
As opposed to fungi, there were no vascular plant species growing exclusively on dead wood and exclusively in the studied primeval forests. However, we registered many European ancient forest plant species identified in a meta-analysis of Hermy et al. (1999). Some of them (Veronica montana, Sanicula europaea and Daphne mezereum) with optima in the unmanaged stands of Zadná Poľana also showed a common incidence with the red list fungal species in the Czech forest reserves (Hofmeister et al. 2014). Nevertheless, ancient forest plant species prevailed in the species composition in both types of managed stands, and moreover, some of them showed their optima in the managed stands. This can be explained by a continuity of forest environment in the studied area and by the ability of such species to survive disturbances connected with timber harvesting.
Multiple macrofungal indicators of old-growth forests and rare forest species were found in the unmanaged stands of the studied area. Camarops tubulina and Hericium coralloides recorded only in the primeval stands of Zadná Poľana Reserve were approved as the indicator species of the best preserved beech forests in Europe (Christensen et al. 2004) and also confirmed later in other forest reserves (Adamčík et al. 2007, 2016; Holec 2005; Abrego et al. 2016). Among other species of old-growth forests (Bredesen et al. 1997; Ripková and Blanár 2004; Holec et al. 2015; Dvořák et al. 2017), Mycena pelianthina, Xylaria longipes, Ascotremella faginea, Phlebia centrifuga, Pseudoplectania melaena (Fr.) Sacc. and Crepidotus applanatus were confirmed by our study.
Eutypa spinosa and Hymenogaster hessei found in the beech-dominated forests of the Poľana Mts. were recorded in the Oravské Beskydy Mts. for the first time and later in the Bukovské vrchy Mts. and Stolické vrchy Mts. (Tomáň and Kabát 2004; Adamčík et al. 2007; Glejdura 2013). Eutypa spinosa with a high frequency and fidelity in the unmanaged forests of Zadná Poľana seems to be rather common species linked to large dead wood in general (Heilmann-Clausen and Walleyn 2007), and therefore, it cannot be considered a good indicator of primeval forests.
Overall, nine red list macrofungi species were found in the studied area: Ascotremella faginea, Camarops tubulina, Chrysomphalina grossula, Entoloma byssisedum, Phylloporus rhodoxanthus, Pseudoplectania melaena (Fr.) Sacc., Stereum subtomemtosum, Tatraea dumbierensis and Tubaria confragosa. They were found most frequently in the primeval stands, but Ascotremella faginea and Camarops tubulina occurred also in the managed beech stands. Another species—Gyromitra fastigiata—was recorded only outside of plots in the unmanaged forests of the reserve. These recently found red list species completed the list of fungi of the Poľana Mts. (Glejdura and Kunca 2010), and along with other indicators of old-growth forests confirm a primeval character of the mixed forests in the Zadná Poľana Reserve.
The rare species Tatraea dumbirensis is considered an old-growth forest and forest continuity indicator (Baral et al. 1999; Holec et al. 2015; Adamčík et al. 2016). While it prefers unmanaged stands in the area, it occurred also in the beech-dominated managed forests, as was also found by Dvořák et al. (2017). This can be explained by the fact that a substantial part of recently managed forests of the studied part of the Poľana Mts. represents the first forest generation after primeval forest clearing according to historical orthophotomaps (© Geodis Slovakia, mapy.tuzvo.sk/HOFM/), and some amount of dead wood regularly occurs there. Panellus violaceofulvus (Batsch) Singer, also considered an indicator species of old-growth forests, is in fact a species fixed especially to silver fir (Gminder and Krieglsteiner 2001), and apart from the primeval forests, it was also found on dead fir wood in various managed forest stands in Slovakia (Jančovičová et al. 2017).
Diversity in different stand types
The average species richness of vascular plants in the unmanaged stands corresponds to the average of Western Carpathian beech forests (Ujházyová et al. 2016). It was significantly higher compared to the even-aged managed beech forests, most likely due to lower competition by overstorey beech trees in a multilayered canopy with frequent gaps providing more heterogeneous light conditions, what is typical for beech-dominated old-growth forests of the Carpathians (Standovár and Kenderes 2003). The negative effect of homogeneous beech-dominated canopies (typical for the managed stands of the area) on understorey diversity was observed earlier by Molder et al. (2008). This can be explained by the extraordinary crown growth plasticity of beech (Schröter et al. 2012), resulting to its ability to completely close a canopy, and by a high annual production of litter which limits herb layer development (Sydes and Grime 1981). Beech competition on the understorey is also limited by the admixture of the other canopy tree species in the primeval stands. A relatively higher richness in the spruce plantations of the area is probably caused by a lower canopy cover and by limited overstorey tree competition in the oldest spruce stands (Ewald 2000). These results partly contradict to a meta-analysis of Paillet et al. (2010), in which the positive response of species richness to clear-cut management was found. This can be explained by a heterogeneity of the compared material in the above-mentioned meta-analysis, especially concerning tree stand age, structure and dominant tree species (Ujházy et al. 2017). We found that the number of plant species per plot is negatively correlated to the tree density and also to the cover of spruce canopy trees (Online Resource 7) in the 40–100-year-old even-aged stands produced by the clear-cut system. High alpha diversity in managed forests is, therefore, linked only to the oldest stands and also to the ephemeral clear-cut communities (not included in this study).
Negative effect of forest management for whole fungal community richness (Paillet et al. 2010) and especially for saproxylic fungi (Brunet et al. 2010) was expected. However, material compared in the meta-analyses and reviews varies in stand age, intensity of management and site types. We confirmed a lower number of macrofungal species per plot in the managed forests using a stratified sampling design within a single site type. The negative effect of management on the species richness of macrofungi was also confirmed by similar recent regional studies (Hofmeister et al. 2014; Dvořák et al. 2017). The number of macrofungal species per plot reflected the impact of all groups of environmental factors related to management interventions and consequent stand structure changes. We proved that altered stand properties significantly affect species diversity as well as the proportion of fungal trophic groups. Dead wood amount significantly correlated with the number of wood-inhabiting fungal species in the managed stand types as has already been seen for spruce forests (Bässler et al. 2010). On the other hand, the relationship was weak in the unmanaged mixed stands where dead wood of multiple tree species was present in sufficient amount in all plots. The number of wood-inhabiting fungal species was related strongly to the dendrometric variables especially in the spruce plantations, while the number of ectomycorrhizal species showed opposite relationships with the same variables in the beech stands (Online Resource 7). Changes in the tree canopy structure and composition affect the topsoil properties (Augusto et al. 2003), and they were important especially for ectomycorrhizal species in the spruce plantations with significantly lower average values of diversity indices, which corresponds to results of Bueé et al. (2011) and Dvořák et al. (2017). Moreover, the number of fungal species in the spruce plantations significantly decreased with the stand age. This relationship is opposite to vascular plants, and this finding has serious implications for the interpretation of other comparative studies using the space-for-time substitution approach. Such studies should be designed, and their results interpreted with respect to the stand age.
A major part of the macrofungal species pool was formed by wood-inhabiting species with significantly limited species pool in the spruce plantations. It means that the negative effect of forest management resulted from differential resource availability for these species, and it was more pronounced in more intensively managed spruce plantations with a limited admixture of original climax tree species. A decrease in the number of wood-inhabiting fungal species as management intensity increases was also reported by Bässler et al. (2012), Mihál (2012) and Dvořák et al. (2017). The proportion of macrofungal trophic groups was similar in all stand types, with general prevalence of wood-inhabiting species (about 53%). Ectomycorrhizal species accounted only for 19% on average, with the highest values in the spruce stands. An increased rate of ectomycorrhizal/wood-inhabiting species was also found by Dvořák et al. (2017) in Czech secondary spruce forests. The situation in beech-dominated managed and primeval stands is consistent with long-term research results from a number of primeval forests of the Western Carpathians (Kuthan et al. 1999; Glejdura 2002; Ripková et al. 2007; Čadek and Gáper 2008; Mihál 2011, 2013; Bučinová et al. 2012), where the proportion of wood-inhabiting species varies from 37 to 54% and the proportion of ectomycorrhizal species from 20 to 30%. Our results are fully comparable with the other studies based on fruiting body sampling. However, higher numbers of species and different proportions of trophic groups can be identified using DNA analyses of soil samples (Schall et al. 2017).
Relationship between vascular plant and macrofungal communities
High fungus–plant ratio (ca 3:1) at the total species pool level is expected in general (Hawksworth 2001). Nevertheless, mean numbers of species per plot of the two taxonomic groups were comparable in all stand types. Alpha diversity of both taxonomic groups seemed also to be similar; however, abundances of plant and fungal populations are hardly comparable. Multiple studies comparing diversity of macrofungi and plants in forests were published mostly with negative results (Berglund and Jonsson 2001; Chiarucci et al. 2005; Santi et al. 2010; Kinga et al. 2013) and conclude that the diversity of plants cannot be used as a proxy for macrofungal diversity prediction. On the contrary, several authors found positive correlations (Sæetersdal et al. 2003; Schmit and Lodge 2005; McMullen-Fisher et al. 2010) and Hofmeister et al. (2014) found a positive correlation with ancient forest plant species richness. We confirmed significant negative correlation between vascular plants and all fungal species in the spruce plantations (similarly to Bučinová et al. 2012) and between vascular plants and the wood-inhabiting fungi in the managed beech forests. In both managed stand types, the amount of dead wood and the tree density decreased in time along with improving light conditions and increasing vascular plant cover and species richness in the understorey. Some terrestrial fungi could be outcompeted by plant roots in the plots with the most dense plant understoreys. On the other hand, an insignificant relationship between fungal and plant diversity was found in the unmanaged primeval forests. It can be explained by a lower variation in stand structure and plant understorey and by the largest proportion and the largest species pool of wood-inhabiting fungi linked to the persistently present dead wood in this stand type.
Congruence between the species composition similarities of plant and fungal communities in forests was frequently identified (Sæetersdal et al. 2003; Chiarucci et al. 2005; Brown et al. 2006; McMullen-Fisher et al. 2010; Kinga et al. 2013). Hofmeister et al. (2014) referred to a positive correlation in species composition similarity of macrofungi and plants in unmanaged forest stands with a decreasing power of correlation in intensively managed stands. However, when a single site type was analysed in this study, a significant relationship between macrofungal and plant community similarity was found only in the spruce plantations. The spruce plantations resulting in homogeneous age-class forests showed the highest variability in the species composition of both taxonomical groups, which is similar to the findings of the above-mentioned studies, where a more heterogeneous dataset was compared. Apart from the site heterogeneity, these contradictory results could arise when stands with different overstorey dominants (Chiarucci et al. 2005) or different management forms (Hofmeister et al. 2014) are compared.
Conclusions
Vascular plant and macrofungal communities responded to stand structure altered by management in a rather similar manner; however, the response of macrofungal species (especially wood-inhabiting) showed to be more pronounced in terms of species composition change. We found that macrofungi can serve as good bioindicators of forest quality under comparable site conditions. Some species of the Carpathian primeval forests were confirmed as indicators for natural beech forests, and others were proposed, and several species considered previously as indicators were also found in the managed forests.
Species diversity did not change as much as species composition (in both taxonomical groups). Therefore, species turnover evaluation seems to be a more important tool of forest natural status assessment than diversity measures. Moreover, multiple taxonomical groups should be involved in the comparisons. The importance of homogeneity of the compared material was emphasized in the study as well. To obtain relevant results, similar sites, stands of similar age and comparable overstorey tree composition should be compared.
As an outcome for the forest management, we recommend keeping mixed forests involving all natural tree species and providing at least a minimal amount of dead wood necessary for wood-inhabiting organisms. A positive example from the Poľana Mts. also underlines the importance of leaving some area of natural forests unmanaged.
References
Abrego N, Christensen M, Bässler C, Ainsworth AM, Heilmann-Clausen J (2016) Understanding the distribution of wood-inhabiting fungi in European beech reserves from species-specific habitat models. Fungal Ecol. https://doi.org/10.1016/j.funeco.2016.07.006
Abs C, Fischer A, Heinrich J, Kugler F, Schöffmann C, Mayer P (2005) Pattern of plant diversity in natural and managed mountain forest. In: Commarmot B, Hamor FD (eds) Natural forests in the temperate zone of Europe—values and utilisation (proceedings, conference 13–17 October 2003 Mukachevo, Ukraine). Swiss Federal Research Institute WSL, Birmensdorf, Carpathian Biosphere Reserve, Rakhiv, pp 273–279
Adamčík S, Christensen M, Heilmann-Clausen J, Walleyn R (2007) Fungal diversity in the Poloniny National Park with emphasis on indicator species of conservation value of beech forests in Europe. Czech Mycol 59:67–81
Adamčík S, Aude E, Bässler C, Christensen M, van Dort K, Ódor P (2016) Fungi and lichens recorded during the Cryptogam Symposium on Natural Beech Forests, Slovakia 2011. Czech Mycol 68:1–40
Agerer R (2001) Exploration types of ectomycorrhizae. Mycorrhiza 11:107–114
Anderson MJ (2001) A new method for non-parametric multivariate analysis of variance. Aust Ecol 26:32–46. https://doi.org/10.1111/j.1442-9993.2001.01070
Augusto L, Dupouey JL, Ranger J (2003) Effects of tree species on understory vegetation and environmental conditions in temperate forests. Ann Forest Sci 60:823–831. https://doi.org/10.1051/forest:2003077
Baral HO, Galan-Marquez R, Krisai-Greilhuber I, Matočec N, Palmer JT (1999) Tatraea dumbirensis, new records of a rare leotialean discomycete in Europe. Österr Z Pilzk 8:71–82
Barbier S, Gosselin F, Balandier P (2008) Influence of tree species on understory vegetation diversity and mechanisms involved—a critical review for temperate and boreal forests. Forest Ecol Manag 254:1–15. https://doi.org/10.1016/j.foreco.2007.09.038
Bässler C, Müller J, Dziock F, Brandl R (2010) Effects of resource availability and climate on the diversity of wood-decaying fungi. J Ecol 98:822–832
Bässler C, Müller J, Svoboda M, Lepšová A, Hahn C, Holzer H, Pouska V (2012) Diversity of wood-decaying fungi under different disturbance regimes—a case study from spruce mountain forests. Biodivers Conserv 21:33–49. https://doi.org/10.1007/s10531-011-0159-0
Bässler C, Ernst R, Cadotte M, Heibl Ch, Müller J (2014) Near-to-nature logging influences fungal community assembly processes in a temperate forest. J Appl Ecol 51:939–948. https://doi.org/10.1111/1365-2664.12267
Battles JJ, Shlisky AJ, Barrett RH, Heald RC, Allen-Diaz BH (2001) The effects of forest management on plant species diversity in a Sierran conifer forest. Forest Ecol Manag 146:211–222. https://doi.org/10.1016/S0378-1127(00)00463-1
Berglund H, Jonsson BG (2001) Predictability of plant and fungal species richness of old-growth boreal forest islands. J Veg Sci 12:857–866. https://doi.org/10.2307/3236874
Berglund H, Jonsson BG (2008) Assessing the extinction vulnerability of wood-inhabiting fungal species in fragmented northern Swedish boreal forests. Biol Conserv 141:3029–3039. https://doi.org/10.1016/j.biocon.2008.09.007
Blaser S, Prati D, Senn-Irlet B, Fischer M (2013) Effects of forest management on the diversity of deadwood-inhabiting fungi in Central European forests. Forest Ecol Manag 304:42–48. https://doi.org/10.1016/j.foreco.2013.04.043
Blasi C, Marchetti M, Chiavetta U, Aleffi M, Audisio P, Azzella MM, Brunialti G, Capotorti G, Del Vico E, Lattanzi E, Persiani AM, Ravera S, Tilia A, Burrascano S (2010) Multi-taxon and forest structure sampling for identification of indicators and monitoring of old-growth forest. Plant Biosyst 144:160–170. https://doi.org/10.1080/11263500903560538
Bredesen B, Haugan R, Aanderaa R, Lindblad I, Økland B, Røsok Ø (1997) Wood-inhabiting fungi as indicators of continuity in spruce forests in eastern Norway. Blyttia 54:131–140
Brown N, Bhagwat S, Watkinson S (2006) Macrofungal diversity in fragmented and disturbed forests of the Western Ghats of India. J Appl Ecol 43:11–17. https://doi.org/10.1111/j.1365-2664.2005.01107.x
Brunet J, Fritz Ö, Richnau G (2010) Biodiversity in European beech forests–a review with recommendations for sustainable forest management. Ecol Bull 53:77–94
Bučinová K, Ujházy K, Glejdura S, Mihál I, Križová E (2012) Druhová diverzita trofických skupín makromycétov a cievnatých rastlín vo východnej časti Národnej prírodnej rezervácie Dobročský prales v porovnaní s priľahlými hospodárskymi lesmi. [Species diversity of trophic groups of macrofungi and vascular plants in the eastern part of Dobročský prales National Natural Reserve in comparison with the neighbouring managed forests]. Acta Facultatis Forestalis Zvolen 53:65–93
Buée M, Maurice JP, Zeller B, Andrianarisoa S, Ranger J, Courtecuisse R, Marçais B, Le Tacon F (2011) Influence of tree species on richness and diversity of epigeous fungal communities in a French temperate forest stand. Fungal Ecol 4:22–31. https://doi.org/10.1016/j.funeco.2010.07.003
Čadek P, Gáper J (2008) Diversity of macrofungi in the Plavno and Badínsky prales national nature reserves. Matthias Belivs Univ Proc 4:27–37
Chao A, Gotelli NJ, Hsieh TC, Sander EL, Ma KH, Colwell RK, Ellison AM (2014) Rarefaction and extrapolation with hill numbers: a framework for sampling and estimation in species diversity studies. Ecol Monogr 84:45–67
Chavez V, Macdonald SE (2010) The influence of canopy patch mosaics on understory plant community composition in boreal mixedwood forest. Forest Ecol Manag 259:1067–1075. https://doi.org/10.1016/j.foreco.2009.12.013
Chiarucci A, D’auria F, De Dominicis V, Laganà A, Perini C, Salerni E (2005) Using vascular plants as a surrogate taxon to maximize fungal species richness in reserve design. Conserv Biol 19:1644–1652. https://doi.org/10.1111/j.1523-1739.2005.00202.x
Christensen M, Heilmann-Clausen J, Walleyn R, Adamčík S (2004) Wood-inhabiting fungi as indicators of nature value in European beech forests. In: Marchetti M (ed) Monitoring and indicators of forest biodiversity in Europe—from ideas to operationality. European Forest Institute, Joensuu, pp 229–237
Christensen M, Hahn K, Mountford EP, Odor P, Standovár T, Rozenbergar D, Diaci J, Wijdeven S, Meyer P, Winter S, Vrška T (2005) Dead wood in European beech (Fagus sylvatica) forest reserves. Forest Ecol Manag 210:267–282
Chytrý M, Tichý L, Holt J, Botta-Dukát Z (2002) Determination of diagnostic species with statistical fidelity measures. J Veg Sci 13:79–90
Dahlberg A, Croneborg H (2003) 33 threatened fungi in Europe. Complementary and revised information on candidates for listing in Appendix I of the Bern Convention, Swedish Species Information Centre, Swedish Environmental Protection Agency and European Council for Conservation of Fungi, Uppsala
Dix NJ, Webster J (2012) Fungal ecology. Springer, Berlin
Durak T (2012) Changes in diversity of the mountain beech forest herb layer as a function of the forest management method. Forest Ecol Manag 276:154–164. https://doi.org/10.1016/j.foreco.2012.03.027
Dvořák D, Vašutová M, Hofmeister J, Beran M, Hošek J, Běťák J, Burel J, Deckerová H (2017) Macrofungal diversity patterns in central European forests affirm the key importance of oldgrowth forests. Fungal Ecol. https://doi.org/10.1016/j.funeco.2016.12.003
Ewald J (2000) The partial influence of Norway spruce stands on understorey vegetation in montane forests of the Bavarian Alps. Mt Res Dev 20:364–371. https://doi.org/10.1659/0276-4741(2000)020%5B0364:TPIONS%5D2.0.CO;2
Frazer GW, Canham CD, Lertzman KP (1999) Gap Light Analyzer (GLA): Imaging software to extract canopy structure and gap light transmission indices from true-colour fisheye photographs. User’s manual and program documentation. Simon Fraser University, Burnaby, and the Institute of Ecosystem Studies, Millbrook
Gilliam FS, Roberts MR (1995) Impacts of forest management on plant diversity. Ecol Appl 5:911–912. https://doi.org/10.2307/2269342
Glejdura S (2002) Zoznam húb [A list of fungi of Dobroč virgin forest]. In: Slávik D et al. Dobročský prales, Národná prírodná rezervácia. ÚVVP LVH SR, Zvolen, pp 48–55
Glejdura S (2013) New records of basidiomycetes and ascomycetes in the Stolické vrchy Mts. (Slovakia). Mykol Listy 124:15–40
Glejdura S, Kunca V (2010) Nové poznatky o mykoflóre CHKO Poľana. [New knowledge of mycoflora in Poľana]. In: Midriak R (ed) Biosférické rezervácie na Slovensku VIII Zvolen: Technická univerzita vo Zvolene. Zvolen, Slovenský výbor pre Program MAB UNESCO, Bratislava, pp 71–77
Gminder A, Krieglsteiner GJ (2001) Die Großpilze Baden-Württembergs. 3. Ständerpilze: Blätterpilze. Ulme
Goslee SC, Urban DL (2007) The ecodist package for dissimilarity-based analysis of ecological data. J Stat Softw 22:1–19
Granito VM, Lunghini D (2011) Biodiversity of macrofungi in the beech forests and calcareous grasslands of the Simbruini Mountains Regional Park (central Apennines, Italy). Plant Biosyst 145:381–396. https://doi.org/10.1080/11263504.2011.563523
Granito VM, Lunghini D, Maggi O, Persiani AM (2015) Wood-inhabiting fungi in southern Italy forest stands: morphogroups, vegetation types and decay classes. Mycologia 107:1074–1088. https://doi.org/10.3852/13-400
Halpern CB, Spies TA (1995) Plant-species diversity in natural and managed forests of the Pacific-northwest. Ecol Appl 5:913–934. https://doi.org/10.2307/2269343
Hawksworth DL (2001) The magnitude of fungal diversity: the 1.5 million species estimate revisited. Mycol Res 105:1422–1432. https://doi.org/10.1017/S0953756201004725
Hédl R, Kopecký M, Komárek J (2010) Half a century of succession in a temperate oakwood: from species-rich community to mesic forest. Divers Distrib 16:267–276. https://doi.org/10.1111/j.1472-4642.2010.00637.x
Heilmann-Clausen J, Walleyn R (2007) Some records of wood-inhabiting fungi on Fagus sylvatica in Northern Spain. Rev Catalana Micol 29:67–80
Hennekens SM, Schaminee JHJ (2001) TURBOVEG, a comprehensive data base management system for vegetation data. J Veg Sci 12:589–591. https://doi.org/10.2307/3237010
Hermy M, Honnay O, Firbank L, Grashof-Bokdam C, Lawesson JE (1999) An ecological comparison between ancient and other forest plant species of Europe, and the implications for forest conservation. Biol Conserv 91:9–22. https://doi.org/10.1016/S0006-3207(99)00045-2
Hofmeister J, Hošek J, Brabec M, Dvořák D, Beran M, Deckerová H, Burel J, Kříž M, Borovička J, Běťák J, Vašutová M (2014) Richness of ancient forest plant species indicates suitable habitats for macrofungi. Biodivers Conserv 23:2015–2031. https://doi.org/10.1007/s10531-014-0701-y
Holec J (2005) Distribution and ecology of Camarops tubulina (Ascomycetes, Boliniaceae) in the Czech Republic and remarks on its European distribution. Czech Mycol 57:97–115
Holec J (2008) Ecology of the rare fungus Hydropus atramentosus (Basidiomycota, Agaricales) in the Czech Republic and its potential value as a bioindicator of old-growth forests. Czech Mycol 60:125–136
Holec J, Beran M (2006) Červený seznam hub (makromycetů) České republiky. [Red list of fungi (macromycetes) of the Czech Republic]. Příroda, Praha
Holec J, Kříž M, Pouzar Z, Šandová M (2015) Boubínský prales virgin forest, a Central European refugium of boreal-montane and old-growth forest fungi. Czech Mycol 67:157–226
Hsieh TC, Ma KH, Chao A (2016) iNEXT: iNterpolation and EXTrapolation for species diversity. R package version 2.0.12. http://chao.stat.nthu.edu.tw/blog/software-download. Accessed 20 Nov 2016
Jančovičová S, Blanár D, Glejdura S, Kunca V (2017) Panellus ringens and P. violaceofulvus (Agaricales, Mycenaceae) from Slovakia: morphological and ecological aspects. Folia Oecol 43:164–175
Kinga R, Morschhauser T, Pál-Fám F, Botta-Dukát Z (2013) Exploring the relationship between macrofungi diversity, abundance, and vascular plant diversity in semi-natural and managed forests in north-east Hungary. Ecol Res 28:543–552. https://doi.org/10.1007/s11284-013-1044-y
Kruskal JB (1964) Multidimensional scaling by optimizing goodness of fit to a nonmetric hypothesis. Psychometrika 29:1–27. https://doi.org/10.1007/BF02289565
Kucbel S, Saniga M, Jaloviar P, Vencurik J (2012) Stand structure and temporal variability in old-growth beech-dominated forests of the northwestern Carpathians: a 40-years perspective. Forest Ecol Manag 264:125–133. https://doi.org/10.1016/j.foreco.2011.10.011
Küffer N, Gillet F, Senn-Irlet B, Job D, Aragno M (2008) Ecological determinants of fungal diversity on dead wood in European forests. Fungal Divers 30:83–95
Kuthan J, Adamčik S, Terray J, Antonín V (1999) Huby národného parku Poloniny. [Fungi of the national park Poloniny]. National park Poloniny Administration Liptovský Mikuláš, Snina
Liira J, Kohv K (2010) Stand characteristics and biodiversity indicators along the productivity gradient in boreal forests: defining a critical set of indicators for the monitoring of habitat nature quality. Plant biosyst 144:211–220. https://doi.org/10.1080/11263500903560868
Lindenmayer DB, Margules CR, Botkin DB (2000) Indicators of biodiversity for ecologically sustainable forest management. Conserv Biol 14:941–950. https://doi.org/10.1046/j.1523-1739.2000.98533.x
Lizoň P (2001) Red list of Slovak fungi. 3rd edition. Catathelasma 2:25–33
Lonsdale D, Pautasso M, Holdenrieder O (2008) Wood-decaying fungi in the forest: conservation needs and management options. Eur J For Res 127:1–22. https://doi.org/10.1007/s10342-007-0182-6
Máliš F, Ujházy K, Vodálová A, Barka I, Čaboun V, Sitková Z (2012) The impact of Norway spruce planting on herb vegetation in the mountain beech forests on two bedrock types. Eur J For Res 131:1551–1569. https://doi.org/10.1007/s10342-012-0624-7
Mantel N (1967) The detection of disease clustering and a generalized regression approach. Cancer Res 27:209–220
Marhold K, Hindák F (1998) Zoznam nižších a vyšších rastlín Slovenska. [Checklist of non-vascular and vascular plants of Slovakia]. Veda, Bratislava
McArdle BH, Anderson MJ (2001) Fitting multivariate models to community data: a comment on distance-based redundancy analysis. Ecology 82:290–297. https://doi.org/10.1890/0012-9658(2001)082%5B0290:FMMTCD%5D2.0.CO;2
McMullen-Fisher SJM, Kirkpatrick JB, May TW, Pharo EJ (2010) Surrogates for macrofungi and mosses in reservation planning. Conserv Biol 24:730–736. https://doi.org/10.1111/j.1523-1739.2009.01378.x
Mihál I (2011) Príspevok k poznaniu mykoflóry Poľany (Stredné Slovensko). [Contribution to the knowledge of mycoflora of Poľana (Central Slovakia)]. Nat Carp 52:7–16
Mihál I (2012) Species diversity, abundance and dominance of macromycetes in beech forest stands with different intensity of shelterwood cutting interventions. Folia Oecol 39:53–62
Mihál I (2013) K poznaniu mykoflóry (Ascomycota, Basidiomycota, Deuteromycota – Fungi. imperfecti) Národnej prírodnej rezervácie Badínsky prales. [Contribution to the knowledge of mycoflora (Ascomycota, Basidiomycota, Deuteromycota – Fungi imperfecti) of the National nature reserve Badínsky virgin forest]. Nat Carp 54:7–16
Mőlder A, Bernhardt-Rőmermann M, Schmidt W (2008) Herb-layer diversity in deciduous forests: raised by tree richness or beaten by beech? Forest Ecol Manag 256:272–281. https://doi.org/10.1016/j.foreco.2008.04.012
Nordén B, Appelqvist T (2001) Conceptual problems of ecological continuity and its bioindicators. Biodivers Conserv 10:779–791. https://doi.org/10.1023/A:1016675103935
Ódor P, Heilmann-Clausen J, Christensen M, Aude E, van Dort KW, Piltaver A, Siller I, Veerkamp MT, Walleyn R, Standovár T, van Hees AFM, Kosec J, Matočec N, Kraigher H, Grebenc T (2006) Diversity of dead wood inhabiting fungi and bryophytes in semi-natural beech forests in Europe. Biol Conserv 131:58–71. https://doi.org/10.1016/j.biocon.2006.02.004
Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Wagner H (2016) Vegan: community ecology package. R package version 2.3-3. http://CRAN.R-project.org/package=vegan. Accessed 15 Feb 2016
Paillet Y, Bergès L, Hjältén J, Ódor P, Avon C, Bernhardt-Römermann M, Bijlsma RJ, De Bruyn L, Fuhr M, Grandin U, Kanka R, Lundin L, Luque S, Magura T, Matesanz S, Mészáros I, Sebastià MT, Schmidt W, Standovár T, Tóthmérész B, Uotila A, Valladares F, Vellak K, Virtanen R (2010) Biodiversity differences between managed and unmanaged forests: meta-analysis of species richness in Europe. Conserv Biol 24:101–112. https://doi.org/10.1111/j.1523-1739.2009.01399.x
Persiani AM, Lombard F, Lunghin D, Granit VM, Tognett R, Magg O, Pioli S, Marchetti M (2015) Stand structure and deadwood amount influences saproxylic fungal biodiversity in Mediterranean mountain unmanaged forests. iForest Biogeosci For 9:115. https://doi.org/10.3832/ifor1304-008
R Core Team (2016) A language and environment for statistical computing. R Foundation for Statistical Computing. http://www.R-project.org/. Accessed 26 July 2016
Ripková S, Blanár D (2004) Výskyt druhov rodu Crepidotus na Muránskej planine a v priľahlej oblasti Slovenského Rudohoria. [Occurrence of species genus Crepidotus in the Muránska Planina Mts. and in adjacent areas of the Slovenské Rudohorie Mts.]. Reussia 1:49–67
Ripková S, Adamčík S, Kučera V, Palko L (2007) Fungi of the Protected Landscape Area of Vihorlat. Institute of Botany of the Slovak Academy of Sciences, Bratislava
Rudolf VH, Rasmussen NL (2013) Ontogenetic functional diversity: size structure of a keystone predator drives functioning of a complex ecosystem. Ecology 94:1046–1056. https://doi.org/10.1890/12-0378.1
Sæetersdal M, Gjerde I, Blom HH, Ihlen PG, Myrseth EW, Pommeresche R, Skartveit J, Solhøy T, Aas O (2003) Vascular plants as a surrogate species group in complementary site selection for bryophytes, macrolichens, spiders, carabids, staphylinids, snails, and wood living polypore fungi in a northern forest. Biol Conserv 115:21–31. https://doi.org/10.1016/S0006-3207(03)00090-9
Saitta A, Bernicchia A, Gorjón SP, Altobelli E, Granito VM, Losi C, Lunghini D, Maggi O, Medardi G, Padovan F, Pecoraro L, Vizzini A, Persiani AM (2011) Biodiversity of wood-decay fungi in Italy. Plant Biosyst 145:958–968. https://doi.org/10.1080/11263504.2011.633114
Šamonil P, Vrška T (2007) Trends and cyclical changes in natural fir-beech forests at the north-western edge of the Carpathians. Folia Geobot 42:337–361. https://doi.org/10.1007/BF02861699
Santi E, Maccherini S, Rocchini D, Bonini I, Brunialti G, Favilli L, Perini C, Pezzo F, Piazzini S, Rota E, Salerni E, Chiarucci A (2010) Simple to sample: vascular plants as surrogate group in a nature reserve. J Nat Conserv 18:2–11. https://doi.org/10.1016/j.jnc.2009.02.003
Schall P, Gossner MM, Heinrichs S, Fischer M, Boch S, Prati D, Jung K, Baumgartner V, Blaser S, Böhm S, Buscot F, Daniel R, Goldmann K, Kaiser K, Kahl T, Lange M, Müller J, Overmann J, Renner SC, Schulze ED, Sikorski J, Tschapka M, Türke M, Weisser WW, Wemheuer B, Wubet T, Ammer Ch (2017) The impact of even-aged and uneven-aged forest management on regional biodiversity of multiple taxa in European beech forests. J Appl Ecol. https://doi.org/10.1111/1365-2664.12950
Schmit JP, Lodge DJ (2005) Classical Methods and Modern Analysis for Studying Fungal Diversity. In: Dighton J, Oudemans P, White JF Jr (eds) The fungal community: its organization and role in the ecosystem, 3rd edn. CRC Press, London, pp 193–214
Schoenholtz SH, Van Miegroet H, Burger JA (2000) A review of chemical and physical properties as indicators of forest soil quality: challenges and opportunities. Forest Ecol Manag 138:335–356. https://doi.org/10.1016/S0378-1127(00)00423-0
Schröter M, Härdtle W, von Oheimb G (2012) Crown plasticity and neighbourhood interactions of European beech ( Fagus sylvatica L.) in an old-growth forest. Eur J Forest Res 131:787–798. https://doi.org/10.1007/s10342-011-0552-y
Schulze ED, Beck E, Müller-Hohenstein K (2005) Plant ecology. Springer, Berlin
Schulze ED, Aas G, Grimm GW, Gossner MM, Walentowski H, Ammer C, Kühn I, Bouriaud O, von Gadow K (2015) A review on plant diversity and forest management of European beech forests. Eur J Forest Res 135:51–67. https://doi.org/10.1007/s10342-015-0922-y
Standovár T, Kenderes K (2003) A review on natural stand dynamics in beechwoods of East Central Europe. Appl Ecol Environ Res 1:19–46
Šťastný P, Nieplová E, Melo M (2002) Mean annual air temperature. In: Miklós L, Maráky P, Klinda J (eds) Landscape Atlas of the Slovak Republic. Ministry of Environment of the Slovak Republic. Slovak Environmental Agency, Bratislava, p 98
Swanson ME, Franklin JF, Beschta RL, Crisafulli CM, DellaSala DA, Hutto RL, Lindenmayer DB, Swanson FJ (2011) The forgotten stage of forest succession: early-successional ecosystems on forest sites. Front Ecol Environ 9:117–125. https://doi.org/10.1890/090157
Sydes C, Grime JP (1981) Effects of tree leaf litter on herbaceous vegetation in deciduous woodland. J Ecol 69:249–262. https://doi.org/10.2307/2259828
Tedersoo L, May TW, Smith ME (2010) Ectomycorrhizal lifestyle in fungi: global diversity, distribution, and evolution of phylogenetic lineages. Mycorrhiza 20:217–263. https://doi.org/10.1007/s00572-009-0274-x
Thomas SC, Halpern CB, Falk DA, Liguori DA, Austin KA (1999) Plant diversity in managed forests: understory responses to thinning and fertilization. Ecol Appl 9:864–879. https://doi.org/10.1890/1051-0761(1999)009%5B0864:PDIMFU%5D2.0.CO;2
Tichý L (2002) JUICE, software for vegetation classification. J Veg Sci 13:451–453. https://doi.org/10.1658/1100-9233(2002)013%5B0451:JSFVC%5D2.0.CO;2
Tomáň P, Kabát V (2004) Vzácne druhy makromycétov oravskej prírody. [Rare species of macromycetes in Orava]. Zborník Oravského múzea 21:178–184
Ujházy K, Križová E, Murín M, Ondruš M (2009) Krátkodobé zmeny vegetácie jedľových bučín Dobročského pralesa. [Short-term changes in the vegetation of fir-beech forests in the Dobročský prales virgin forest]. Zprávy Čes Bot Společ 44:59–77
Ujházy K, Križová E, Glončák P, Benčaťová B, Nič J (2013) Tree species and management effect on herb layer species composition in mountain fir-beech forests of the Western Carpathians. In: Kozak J, Ostapowicz K, Bytnerowicz A, Wyżga B (eds) The Carpathians: integrating nature and society towards sustainability. Springer, Berlin, pp 239–255
Ujházy K, Hederová L, Máliš F, Ujházyová M, Bošeľa M, Čiliak M (2017) Overstorey dynamics controls plant diversity in age-class temperate forests. Forest Ecol Manag 391:96–105. https://doi.org/10.1016/j.foreco.2017.02.010
Ujházyová M, Ujházy K (2007) Dynamika fytocenóz bukových lesov v Kysuckej vrchovine. [Vegetation dynamics of beech forests of the Kysucká Vrchovina Mts.]. In: Križová E, Ujházy K (eds) Dynamika, stabilita a diverzita lesných ekosystémov. TU vo Zvolene, Zvolen, pp 29–36
Ujházyová M, Ujházy K, Chytrý M, Willner W, Čiliak M, Máliš F, Slezák M (2016) Diversity of beech forest vegetation in the Eastern Alps, Bohemian Massif and the Western Carpathians. Preslia 88:435–457
Verstraeten G, Baeten L, De Frenne P, Vanhellemont M, Thomaes A, Boonen W, Muys B, Verheyen K (2013) Understorey vegetation shifts following the conversion of temperate deciduous forest to spruce plantation. Forest Ecol Manag 289:363–370. https://doi.org/10.1016/j.foreco.2012.10.049
Zhang Ch, Xie G, Fan S, Zhen L (2010) Variation in vegetation structure and soil properties, and the relation between understory plants and environmental variables under different Phyllostachys pubescens forests in Southeastern China. Environ Manage 45:779–792. https://doi.org/10.1007/s00267-010-9429-y
Acknowledgements
This research was supported by the Science Grant Agency of the Ministry of Education of the Slovak Republic and the Slovak Academy of Sciences VEGA, Grant Nos. 2/0039/14, 1/0639/17 and Slovak Research and Development Agency under the project APVV-15-0270 and APVV-15-0176. We also acknowledge the postdoc support of Katarína Bučinová in 2015 at the University of Vienna funded by Action Austria—Slovakia, Cooperation in Science and Education (Ministry of Education, Science, Research and Sport of the Slovak Republic and the Federal Ministry of Science, Research and Economy of the Republic of Austria).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Hans Pretzsch.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Ujházy, K., Ujházyová, M., Bučinová, K. et al. Response of fungal and plant communities to management-induced overstorey changes in montane forests of the Western Carpathians. Eur J Forest Res 137, 169–183 (2018). https://doi.org/10.1007/s10342-017-1096-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-017-1096-6