
Abstract
Planting of spruce on the sites of natural beech forests is very common across Europe. Its effect on ground vegetation is assumed to be strong on poorly buffered soils. We investigate associated patterns of herb layer composition and diversity on crystalline and carbonate parent rock in the Western Carpathians. We analysed a series of vegetation plots with respect to the partial relationship of spruce cover with vascular understorey species (ordination), to the affinity of plant species to sample groups with different cover of spruce and to plant diversity and species turnover within these groups. In our data set spruce and beech were the most important tree species with the closest association with understorey composition and their covers explained similar proportions of herb layer variability. Species composition was significantly different under spruce canopies, but species richness was not lower (crystalline region) or even higher (carbonate region), and overall diversity of vascular plants was only slightly affected. Most species negatively or positively associated with spruce differed between bedrock types, but those with the same positive relationship are diagnostic for natural spruce forests (Vaccinio-Piceetea). The species turnover between beech- and spruce-dominated stands was much higher on crystalline bedrock. The higher amounts of light and acid litter in spruce stands, limited growth of spruce on dolomite bedrock and the competitive pressure of beech seem to be the most important drivers of the herb layer changes.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The relationships between canopy trees and ground vegetation in the forest understorey have been studied frequently in various forest ecosystems, including the temperate forests of Central Europe (cf. the review paper of Barbier et al. 2008). Trees dominating forest communities are long-lived organisms producing the major part of biomass and consequently affecting the other components of an ecosystem. Apart from the abiotic site properties, plant species composition of forest ecosystems is largely affected by the canopy of tree species (Kuuluvainen and Pukkala 1989; van Oijen et al. 2005; Wulf and Naaf 2009). These effects can be observed in natural forests (Saetre 1999) and even more markedly in planted stands of non-native or allochtonous tree species (Quine and Humphrey 2010). As the species diversity is highest in the herb layer, biodiversity of forest community is largely a function of the herb layer community. According to Gilliam (2007), the ratio between herb and tree species at the broad scale of North American temperate forests ranges from 2 to 10. Therefore, changes in the tree layer and their impact on the herb layer should have serious consequences for the biodiversity of the whole ecosystem, and an understanding of tree–herb interactions is essential for maintaining biodiversity of forests.
A proper evaluation of the effect of changes in the tree species composition on biodiversity is also required to meet the commitment of European states to halting the loss of biodiversity (EEA 2007) and for the purpose of forest ecosystems status assessment (EEA 2010). As the pressure on forests grows and the proportion of natural forests decreases, the need for bioindicators in forest monitoring to track environmental changes and identify remnants of original natural forests increases (Liira and Kohv 2010; Blasi et al. 2010). The failure to achieve the 2010 biodiversity target (CBD 2011) calls for a new biodiversity strategy with progress in biodiversity data measuring and monitoring (EC 2011; EEA 2010). Diversity indices based on the ground layer plant species composition are considered important indicators of forest status assessments (Pitkänen 1998; MCPFE 2002; Schmidt 2005; Schmidt et al. 2006; EEA 2007; Petriccione et al. 2007). As one aspect of tree-herb interactions, the values of diversity measures should be influenced by tree canopy.
We approach the problem using the example of secondary spruce stands. Planting of spruce (Picea abies (L.) Karst.) in the place of natural broadleaved stands is one of the most common forms of human influence on forest ecosystems in Central Europe. Forest managers’ preference for spruce has induced changes in nutrient cycling, litter accumulation and soil acidification in large areas (Fanta 1997; Jonášová and Prach 2004). Emmer et al. (1998, 2000) have described this process as “anthropogenic borealization” with serious impacts on the whole landscape and its biodiversity.
P. abies is a natural species in the higher mountains of Slovakia and in many other mountain ranges in Europe but it was planted on extended areas beyond the limits of natural occurence, especially on sites originally occupied by beech forests. Its influence on the herb layer has been studied by several authors. Thus, Teuscher (1985) and Simmons and Buckley (1992) reported an inhibition of vascular plants by spruce, resulting in lower species richness under coniferous canopies. On the other hand Bürger (1991), Lücke and Schmidt (1997) and Ewald (2000a) found spruce plantations to be even richer than deciduous stands. It was reported that beech stands tend to have low understory diversity in comparison with pure stands of other tree species (Tárrega et al. 2011) and mixed forests, which was attributed to the properties of beech litter (Mölder et al. 2008). Barbier et al. (2008) concluded in a review paper that broadleaved forests generally provide more diversified understorey vegetation than coniferous and that maximum diversity is observed in pure stands, not in mixed ones. However, due to varying management and site attributes, it is difficult to generalize the results.
Fajmonová (1974) and Hadač and Sofron (1980) reported that the changes in herb layer composition depend on the number of planted generations of P. abies on the same locality, and several authors use the intensity of these changes as one of the basic criteria for classifying coniferous forests (Hornstein 1951; Hadač and Sofron 1980). The most important drivers of spruce influence on herb layer are the changes of topsoil conditions caused by the needle litter; such as acidification, decrease in base saturation and C/N ratio, accumulation of litter and inhibition of nutrient release (Fajmonová 1977; Ewald 2000a; Augusto et al. 2002; Fabiánek et al. 2009). This primarily results in an increase in acidophilous species, especially shallow-rooted ones, and decrease in nitrophytes (Fajmonová 1974; Ewald 2000b; Poleno 2001; Šimurdová 2001; Chytrý et al. 2002; Šomšák and Balkovič 2002; Šomšák 2003; Máliš et al. 2010). Soils under spruce lock up nitrogen and release it in flushes after disturbance (Ewald 2009a). All of the above-mentioned effects are generally observed regardless of parent rock materials, even in case of base rocks as different as calcareous and siliceous rocks that are characterized by unique composition and diversity of plant communities (Moravec et al. 2000; Chytrý and Tichý 2003; Ewald 2003; Willner and Grabherr 2007; Jarolímek and Šibík 2008). Crystalline and carbonate rocks represent a substantial proportion of sites with Fagus sylvatica in Central Europe (Pott 2000; Asch 2005; Bohn et al. 2003), however, the effect of bedrock type on diversity change in secondary spruce stands is not yet known.
The aim of this paper is to find out whether (1) species composition of the herb layer is significantly changed in stands with artificial spruce (P. abies) admixture or dominance, (2) species groups positively or negatively responding to spruce introduction can be defined, (3) species diversity of herb layer is higher in stands dominated by beech or spruce, or in mixed stands, (4) mechanisms behind the effects of spruce planting differ between crystalline and carbonate bedrocks.
Materials and methods
Study area
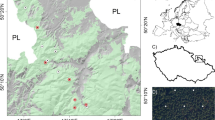
The field research was carried out in two study areas in the central part of Slovakia and belonging to the central West Carpathians, the Veporské Vrchy Mts. and the Veľká Fatra Mts. (Fig. 1). In both areas, the research was restricted to the altitudinal zone with natural dominance of beech.

Study locations
The Veporské Vrchy Mts. cover approximately 800 km2. The most widespread parent rock material is granodiorite (Bezák et al. 1999). The soils are mostly classified as Eutric and Dystric Cambisols, less frequently as Skeletic Cambisols (IUSS Working Group WRB 2006; Máliš et al. 2005). Elevation ranges from 300 to 1,439 m. Mean annual temperatures vary between 3.5 and 7.0 °C (Št’astný et al. 2002), and mean annual precipitation totals between 700 and 950 mm (Faško and Št’astný 2002). Relevés were sampled between 490 m and 1,195 m a.s.l., in which zone beech forests dominated originally. The upper boundary of beech-dominated stands under subalpine spruce forests is approximately in 1,200 m a.s.l. in Slovakia (Blattný and Št’astný 1959). According to present observation, it is even higher than 1,300 m (depending on relief and bedrock type). In the zone above ca. 800 m, natural beech forests are mainly mixed with Abies alba, Fraxinus excelsior, Acer pseudoplatanus and above 1,200 m only very rarely with P. abies.
The second area is situated in the south-western part of the Veľká Fatra range and covers 160 km2. Parent rock consists predominantly of dolomites, limestones and marly limestones; therefore, the Rendzic Leptosols and Eutric Cambisols (IUSS Working Group WRB 2006) are the most common soil types. Calcium carbonate content is high on dolomites within the whole soil profile; on limestones, it may increase with depth, and on the marly limestones, usually only the lower parts of soil profile contain calcium carbonate. Elevation range is between 400 m and 1,595 m. Mean annual temperatures vary between 2.5 and 6.5 °C (Št’astný et al. 2002), and mean annual precipitation totals between 750 and 1,250 mm (Faško and Št’astný 2002). The elevation of relevés ranges between 550 and 1,300 m a.s.l. Beech forests are mostly present in this area; however, due to the more diversified geology with different topsoil properties, the vegetation cover is also more variable. In the zone above ca. 800 m, beech stands are again mixed with A. alba, F. excelsior, A. pseudoplatanus as in the Veporské Vrchy Mts.; however, F. sylvatica still prevails as the dominant species. Especially, the natural admixture of A. alba is very common in this altitudinal zone of the West Carpathians, and consequently the mountain beech forests are called fir-beech. The overview of both areas is presented in Table 1.
Vegetation sampling
In order to reduce site variability as much as possible and to increase the proportion of variability caused by tree species, the stratified sampling was applied. The selection of sites for vegetation sampling was based on specification of following criteria.
Terrain characteristics, landforms and spatial distribution
Samples were positioned on slopes between 10° and 40°, and all other landforms such as ridges and valley bottoms occupied by specific communities were excluded. Suitable locations were identified by overlaying of geomorphological and geological information. Extraction of landforms was based on Wood (1996) and identified from a DEM (30 m resolution) using the module r.param.scale in GRASS GIS (Neteler and Mitasova 2008). Although systematic stratification by elevation was not applied, observers made an effort in subjective selection of relevés position to keep the sampling balanced along the altitudinal gradient and more or less regular within spatial distribution.
Geological substratum
Granodiorite in the Veporské Vrchy Mts. (representing crystalline and siliceous parent rocks), dolomites, limestone and marly limestones in the Veľká Fatra range (representing carbonate bedrock types) were considered as geological substrata for sampling and extracted from geological maps (Polák et al. 1997; Bezák et al. 1999).
Potential vegetation
Only the sites of potential vegetation units Eu-Fagenion or Cephalanthero-Fagenion (only in the calcareous region Veľká Fatra Mts.) suballiance (Jarolímek and Šibík 2008) with potential dominance of F. sylvatica were sampled. Occurrence of spruce in sampled stands is not natural in crystalline region (Vladovič 2003) with the exception of a certain admixture in the coldest sites above 1,200 m a.s.l. In the Cephalanthero-Fagenion communities of calcareous region P. abies is regularly admixed with increasing abundance with altitude (Vladovič 2003); however, in the majority of sites it occurs with a natural proportion of less than 10 %. Sites of the vegetation units were identified according to potential vegetation maps provided by National Forest Centre (NFC).
Tree species composition and structure
Plots were situated in natural stands with beech dominance and in secondary stands with various proportions of spruce. Both managed and unmanaged stands with a similar structure (closed stands dominated by upper tree layer) were sampled. Management followed uniform patterns under centralized socialist planning schemes during the second half of the twentieth century. Stands were managed by more or less regular thinning. Some of the stands in the Veľká Fatra Mts. (calcareous region), both beech- and spruce-dominated, were left without thinning in the last few decades as they were incorporated into protected areas. Nevertheless, there was no strong difference in stand structure between managed and unmanaged stands within our data set, as several structural characteristics (canopy cover, tree density, age) were used as selection criteria of plot placement. The sampling plots were located only in the closed stands (canopy cover over 70 %) aged 70–130 years to avoid inhibition in high-density young stands on the one hand, and luxuriance of ground vegetation in low density senescing stands on the other. Stands affected by insect-induced mortality were not included. Information on tree composition, age and crop density of stands was extracted from forest stand inventory data. Selection criteria were verified directly in the field during sampling.
Forest stand persistency
Stands planted on former grasslands were not included and continuity of forest cover was checked on historical maps from the nineteenth century (Timár et al. 2006).
Altogether 123 phytosociological relevés from the crystalline region and 153 relevés from the calcareous region were analysed. Data set sampled by the authors was completed from literature and database of NFC by relevés, which meet the same selection criteria as sampling procedure. Plots were square or rectangular, with an area of ca. 400 m2 (in several cases up to 500 m2). Species abundance was estimated using Braun-Blanquet’s scale adjusted by Barkman et al. (1964) and Zlatník’s scale (Randuška et al. 1986) and transformed to percentage values for the purpose of analysis. According to Zlatník (1976), the following vegetation layers were distinguished: trees above the main storey (tree layer 1), trees of the main storey (tree layer 2), trees under the main storey and higher than half-height of the main storey (tree layer 3), trees and shrubs lower than half-height of the main storey (tree layer 4), trees and shrubs lower than 130 cm (tree layer 5), and herbs (herb layer). In analyses, species data were aggregated to the three basic layers (tree, shrub, herb); the moss layer was not included. The plant species names follow the checklist of Slovakia (Marhold and Hindák 1998), and those of syntaxa, the list of vegetation units of Slovakia (Jarolímek and Šibík 2008).
Data analyses
Data analyses follow two analytical approaches. The first is based on constrained ordination using the matrix of the canopy tree species as environmental variables. The association of individual herb species with the increasing spruce cover was investigated. This approach implies the cover of herbs as a measure of their response and provides the possibility to exclude the influence of other environmental factors. The second approach compared the plant species composition between three groups of relevés with different covers of spruce. The association of herbs with spruce is evaluated using their presence expressed by the frequency within relevé groups and the fidelity value as well. Measuring fidelity (Chytrý et al. 2002) allows to statistically define the species that are diagnostic for the differentiation of vegetation units (Bruelheide 2000), in this case defined by different cover of spruce. This approach eliminates the effect of species abundance, which could be affected by various hidden effects. In this matter, the impact of spruce is considered as negative only when species is excluded from the plant community. The second approach does not imply the correction for covariables, and many confounding variables remain hidden. However, there is another advantage coming from the possibility of standardization of relevé group size, which eliminates the unevenness in the number of relevés with different cover of spruce.
Cover of tree species sampled in more than one tree layer was calculated using the JUICE programme (Tichý 2002) as follows, under the assumption that canopies overlap randomly:
Calculation of cover in case of one tree species occurrence in two layers:
-
A + (1–A) × B = cumulative cover of tree
-
A—cover value of tree species in the first layer
-
B—cover value of tree species in the second layer
Calculation of cover in case of one tree species occurrence in three layers:
-
A + (1 − A) × B + (1 − [A + (1 − A) × B]) × C = cumulative cover of tree species
-
A—cover value of tree species in the first layer
-
B—cover value of tree species in the second layer
-
C—cover value of tree species in the third layer
Only the upper three tree layers (tree layer 1, 2 and 3) were used as constraining variables in ordination. The matrices of these cumulative cover values were used in analyses.
Ordination analyses
Ordination analyses of the two data sets were carried out in Canoco for Windows 4.5 (Ter Braak and Šmilauer 2002) using logarithmic transformation of herb cover values and biplot scaling on inter-species distances. The adequacy of unimodal versus linear response models in ordination was assessed by detrended correspondence analysis (DCA) and detrended canonical correspondence analysis (DCCA), both with detrending by segments and SD scaling of axes (Økland 1990). Impact of tree species on the understorey was evaluated by direct ordination techniques, once using all dominant tree species as environmental variables and once using spruce cover as the only environmental variable.
Stratified selection of sampling sites excluded undesirable landforms; however, the effect of other abiotic factors such as altitude still remained in vegetation data. Therefore, significance of altitude, total canopy cover, slope inclination and orientation (southness) influence was tested by Monte Carlo permutation test with forward manual selection, unrestricted permutation and 999 runs in CCA (canonical correspondence analysis). CCA was suggested by lengths of gradient in DCA (3.7, 3.4 SD) in the cases of both areas. Southness was derived from slope orientation and used in order to obtain an ecologically meaningful evaluation of slope aspect from the thermal point of view. Transformed aspect (a trans) was calculated following the approach of Reger et al. (2011) and Beers et al. (1966) (see also Ewald 2009b) as
where a is the slope aspect in degrees, and a max the thermally most favoured aspect. A SSE (202.5°) orientation was assumed to have the highest heat exposure. Slope aspect in degrees was converted to radians by multiplying by pi/180, resulting in values that vary between −1.0 (thermally least favoured aspect) and 1.0 (thermally most favoured aspect). Finally, all tested abiotic factors showed significant influences on the herb layer and were included in further ordinations as covariables in order to eliminate their influence and to extract the partial effect of the tree layer. The impact of tree species on the understorey was subsequently evaluated by constrained ordination methods and according to the length of constrained gradients in DCCA (all dominant tree species as constraining variables 1.6, 1.4 SD; spruce cover as constraining variable 1.2, 1.1 SD), the linear method redundancy analysis (RDA) was applied. RDA was run twice, once using all dominant tree species as environmental variables and once using spruce cover as the only environmental variable. The purpose of the first case was to investigate the joint influence of tree species on herb layer and their interactions, using RDA biplots (Fig. 2) and explained variance of species data (Table 2). The isolated influence of each tree species was tested again by the Monte Carlo test. The purpose of the second case was to extract the direct relationship between P. abies and herb species through the RDA score on the first ordination axis (Figs. 3, 4). Additionally to this analysis, the significance of the herb species response to spruce was tested by partial correlation analysis (Statistica 7.1, StatSoft 2005). To obtain the same model, the cover values of understorey species were also logarithmically transformed, and calculation was done with controlling for the same variables that were used as covariables in RDA.

Tree species influence on the herb layer in the crystalline (left) and calcareous region (right) according to RDA (log-transformation of herb covers; covariables: elevation, slope, southness, total canopy cover) with cover of dominant tree species as environmental variables. Herb species with highest fit range are plotted (5–100 % in both cases)

Comparison of plant species diversity between relevé groups with increasing spruce cover in both mountain ranges. Box plots present median, 25 and 75 % quantiles, min and max values. The significance of differences of the mean values between groups within region are marked by capital (crystalline region) or small (calcareous region) letters

Changes of mean Ellenberg value for soil reaction and light in relevé groups with different spruce cover. Box plots present median, 25 and 75 % quantiles, min and max values. The significance of differences of the mean values between groups within region are marked by capital (crystalline region) or small (calcareous region) letters
Analyses of relevé groups with different proportions of spruce
Comparison of three relevé groups with a different proportion of spruce in the tree layer was based on the same relevé data as in ordination. The first group represents the natural beech-dominated stands without P. abies in the crystalline region and with the proportion of spruce under 10 % in the calcareous region. These zero and 10 % limits were derived from the natural proportion of tree species in forest types according to Vladovič (2003). The second group involves mixed stands with a spruce proportion of under 50 %. The relevés with a proportion of spruce exceeding 50 % are classified into the third group. Groups were compared in terms of floristic composition, diversity and similarity. The floristic comparison of groups was carried out using the JUICE software (Tichý 2002). Fidelity (phi coefficient based on presence/absence data) with respect to relevé groups was calculated for each species in each data set. The size of all relevé groups was standardized to equal size and Fisher’s exact test was carried out using significance level p < 0.05. Juvenile shrubs and trees present in the herb layer were excluded.
The influence of P. abies on herb layer diversity was assessed by comparing common diversity indices between relevé groups. Shannon–Wiener index, Shannon`s equitability (evenness) as proposed by Pielou (1975), and species richness were calculated in JUICE. Results are presented as box plots (Statistica 7.1, StatSoft 2005). Mean values of diversity measures in relevé groups were compared by Tukey’s post hoc test in analysis of variance.
To evaluate the overall extent of compositional change caused by increasing proportions of spruce, pairwise similarity (Sørensen index based on presence/absence) of relevés was calculated in JUICE (Tichý 2002) and compared between groups by a Mann–Whitney U test. Relevé groups with varying proportions of P. abies were also compared with respect to indicator values for soil reaction and light (Ellenberg et al. 1992) to assess the specific effect of soil acidification and changes in the light conditions caused by spruce. The calculation was done without the weighting by species cover.
Results
Ordinations
In total 275 taxa were recorded, 121 common to both regions, 17 unique for the crystalline and 137 for the calcareous range. The number of taxa was almost twice as high in the calcareous area. Testing of the abiotic factors showed their significant influence on the herb layer. Together, they explained 9.5 % of the species composition variance in the crystalline area and 8.3 % in the calcareous region. Altitude was the most significant variable in the both regions (Table 2). Consequently, all factors were included into both types of ordinations as the covaribles.
The first type of a constrained ordination (using all dominant tree species as explanatory variables) showed that all dominant trees (except F. excelsior in calcareous area) have significant correlation with the herb layer vegetation (Table 2). In the case of the crystalline area, the correlation was almost twice higher. The cover of beech and spruce in the tree layers explained similar proportions of variability, and their influence was more or less the most relevant. P. abies cover explained a relatively small proportion of overall variability of species composition (crystalline region, 4.75 %; calcareous region, 1.5 %), nevertheless its influence was highly significant in both mountain ranges. RDA biplots (Fig. 2) show that the influence of conifers and especially of spruce has an opposite effect to that of broadleaved trees.
The second type of the constrained ordination used the spruce cover in the tree layers as the only one explanatory variable. The species score on the first RDA axis (the only constrained axis in this analysis) was used to express a positive or negative affinity of the herb species to spruce canopies. The values as well as the diagnostic status of species (those with significant fidelity in one of the relevé groups with different proportion of spruce) are presented in the Tables 3 and 4. Negative affinity to the spruce cover usually implies a positive affinity to beech and/or other broadleaved trees.
Comparison of relevé groups with different cover of spruce
The first group representing original beech-dominated forests had the lowest number of diagnostic species in both study areas. Only 6 out of 82 diagnostic species (7 %) showed a clear affinity to the first relevé group without P. abies on calcareous bedrock, while on the crystalline bedrock of the Veporské Vrchy Mts. it was 7 out of 24 (21 %). On the other hand, in the Veporské Vrchy range, all groups were similar in size, whereas in the calcareous Veľká Fatra range, there were many more species associated with spruce. The majority of species in the calcareous bedrock region showed an affinity to the mixed or spruce-dominated forests—spruce appeared to have a very positive effect on species richness of vegetation on limestones or dolomites. When abundance of species was taken into account (RDA), the reaction of communities was similar. Of taxa from calciphilous vegetation, 64.7 % had positive RDA scores. In the case of vegetation on crystalline bedrock, the proportions of species with different scores were very balanced (49.6 % positive score/50.4 % negative score). It can be concluded that spruce promotes the occurrence of a large number of new species (especially on calcareous bedrock) and reduces the presence of only a few species. Many species of the natural community persist in spruce forests. Consequently, species richness is higher in relevés with elevated cover of P. abies (Fig. 3).
Species diversity in the secondary spruce-dominated forests was not lower, but species composition was significantly different from the beech forests (Table 5). In general, the relationship between the spruce cover and the species diversity expressed by the Shannon–Wiener index was weak (Fig. 3). Only the mixed forests with beech dominance in the crystalline region (2nd relevé group) had significantly increased diversity compared with the 1st group. Non-significant differences of Shannon–Wiener index values in the calcareous region are a consequence of a contradictory change of the two components of the index: evenness decreased and species richness increased with spruce cover. Lower evenness of calciphytic communities under mixed and spruce canopies is caused by the occurrence of new spruce-related species with low cover values as well as by increased abundance of species such as Sesleria varia, Carex alba, Oxalis acetosella and Melampyrum sylvaticum, which increased in cover two to four times. Spruce stands in the calcareous range had 5–6 (10 %) more taxa on average compared with the pure beech stands, whereas on the crystallines were richer only by two species on average.
Species association with spruce across regions
A large proportion of taxa (59.5 %) showed consistent association with spruce cover across the studied areas. Taking into account only species with a significant correlation with spruce according to partial correlation analysis, the number was reduced to 25 species (20.7 %). Finally, only O. acetosella, Hieracium murorum, Luzula luzuloides and Vaccinium myrtillus were significantly and positively associated with spruce in both regions. A significant and negative correlation of the same species in both regions was not found; however, according to fidelity measuring within relevé groups, Mercurialis perennis showed significant affinity to beech stands without spruce (1st relevé group) regardless of bedrock. Veronica officinalis, Anemone nemorosa and Epilobium montanum were diagnostic for stands with spruce admixture (2nd and 3rd relevé group) in both the Veľká Fatra and the Veporské Vrchy ranges.
Many characteristic species of beech forests such as Dentaria enneaphyllos, Dentaria bulbifera, Galium odoratum, Tithymalus amygdaloides, Bromus benekenii and Hedera helix showed negative correlation with P. abies cover or affinity to beech stands at least in one of the regions, confirming their tight relation to F. sylvatica. On the other hand, some of the beech-related species (Asarum europaeum, Paris quadrifolia, Glechoma hederacea agg., Viola reichenbachiana) showed significant affinity to spruce stands (3rd relevé group) on calcareous bedrock. Contradictory affinity to relevé groups was found in case of Galeobdolon luteum agg., Actaea spicata, Geranium robertianum, Urtica dioica, Aegopodium podagraria, Stachys sylvatica and Milium effusum. A large number of species had a significant correlation with spruce cover or affinity to some relevé group in one area, but not in the other.
Several common species such as Athyrium filix-femina, Polygonatum verticillatum, Fragaria vesca, Sanicula europaea and Ajuga reptans showed neither a significant difference of frequency between groups, nor any significant correlation with cover of spruce.
Of course, there were also many species with a clear association with beech or spruce that were specific to one mountain range and therefore not comparable. Thus, acidophilous species such as Avenella flexuosa and Festuca altissima were associated with spruce in the crystalline region and missing, or having a very low percentage frequency on carbonate bedrock. Conversely, calcicolous species such as Cephalanthera rubra, Epipactis atrorubens and Hacquetia epipactis were typical for beech forests in calcareous range and missing on crystalline bedrock. The increasing proportion of acidophytes in spruce stands of the crystalline range is also indicated by Ellenberg indicator values (Fig. 4).
The strength of the relationship with spruce on species turnover differed between bedrock types (Table 5). Whereas it appeared to increase continuously with spruce cover on crystallines, mixed stands with 10–50 % spruce on carbonate rocks were composed of the greatest number of unique species. Calciphytic communities of almost pure beech or spruce stands were more similar to each other than to mixed stands, and many species occurred in both types of pure stands rather than in mixed ones. Most of these species, for example, Rubus idaeus, A. europaeum, G. robertianum, E. montanum, P. quadrifolia, O. acetosella, Brachypodium sylvaticum, Mycelis muralis, Dryopteris filix-mas and A. spicata showed an overall positive association to spruce on calcareous bedrock due to their significant fidelity to the 3rd group. This kind of discontinuous reaction was not observed in the crystalline Veporské Vrchy Mts.
Discussion
The results demonstrate that variation in P. abies cover is significantly associated with species composition and diversity on crystalline as well as calcareous bedrocks. Spruce cover is associated with higher richness, especially on calcareous rocks, whereas diversity peaked in mixed stands of the crystalline area.
Only a few species seem to be inhibited by spruce, more species were positively associated with the cover and the presence of P. abies. However, many more positive correlations were found on the calcareous bedrock. This partly conflicts with the results from the calcareous Alps, where no systematic differences in herb species richness due to various spruce proportions (Ewald 2000a, b) or lower richness caused by anthropogenic promotion of P. abies were found (Teuscher 1985). In comparison with our work, studies of Ewald (2000a, b) used more precise stratified sampling design, which strictly balanced sites and canopy types and was corrected for mineral soil properties. Ewald (2000b) reported that indicators of acidity were significantly favoured by spruce canopies (as in our case), whereas the decrease in species characteristics of deciduous forests and nitrogen indicators was not significant. Contrary to this, spruce appears to significantly inhibit several species in the calcareous region of the Veľká Fatra range, especially those which are considered to be characteristic of broadleaved forests of the Querco-Fagetea class (D. bulbifera, D. enneaphyllos, Bromus bennekenii, H. epipactis and M. perennis). Species of the Dentaria genus as well as M. perennis are shade tolerant spring geophytes with relatively high demands for pH and nutrients (Ellenberg et al. 1992). H. epipactis has similar properties, but it is a hemicryptophyte (Jurko 1990, Klotz et al. 2002). Spring geophytes are adapted to environmental conditions of various broadleaved and especially beech forests (Ellenberg 2009), particularly the high annual production of litter (Packham et al. 1992) and high light availability in the understorey in early summer. D. enneaphyllos is diagnostic especially for fir-beech mountain forests (Moravec et al. 2000; Willner and Grabherr 2007), as is D. bulbifera for the submontane beech forests of the Western Carpathians (Jarolímek and Šibík 2008).
Species favoured by spruce differ in requirements of nutrients or soil reaction, and they are not characteristic of a single class. The results from the crystalline Veporské Vrchy Mts. showed more markedly that spruce favours acidophytes such as A. flexuosa, V. myrtillus, O. acetosella, L. luzuloides, H. murorum and Maianthemum bifolium. These species are considered as diagnostic of spruce forests (Jarolímek and Šibík 2008; Chytrý et al. 2002; Husová et al. 2002; Ewald 2000b). However, they are widespread also in other acidophilous forest communities, such as Luzulo-Fagion, Genisto germanicae-Quercion or Abietion albae, as well. Stands on carbonate bedrock are less affected by acidification, and the proportion of acidophytes within a species favoured by spruce was markedly lower. This may be a consequence of the higher buffer capacity of soils with high calcium carbonate content (Šály 1978; Ulrich 1983), which reduces the acidic influence of spruce litter. Furthermore, spruce admixture in a crystalline area inhibits nitrophytes (Alliaria petiolata, G. robertianum, U. dioica, S. sylvatica), and partly mesotrophic plants (T. amygdaloides, G. odoratum), many of which are characteristic for F. sylvatica forests. These results are in accordance with other studies from acid soils (Šomšák 2003; Fajmonová 1974).
M. perennis, the only common species with significant affinity to beech stands in both areas, is considered to be diagnostic for beech (Fagion) and scree forests (Tilio-Acerion) (Jarolímek and Šibík 2008; Chytrý and Tichý 2003). On the other hand, there were four species positively associated with spruce cover regardless of bedrock (evaluated by both methodic approaches): V. myrtillus, O. acetosella, L. luzuloides and H. murorum. All of them are diagnostic species for the Vaccinio-Piceetea class in Slovakia (Jarolímek and Šibík 2008) and constant and diagnostic for spruce forests in the Eastern Alps according to Chytrý et al. (2002). The group comprises mesophilous species indifferent to temperature, partly oligotrophic (H. murorum) and acidophilous (V. myrtillus, L. luzuloides) (Ellenberg et al. 1992). O. acetosella and V. myrtillus are shallow-rooted plants as reported by Ewald (2000b), which are able to spread roots in the raw humus layer (Oh horizon) produced by coniferous trees (Emmer 1995).
Several authors reported a negative effect of P. abies on the diversity and cover of vascular plants (e.g. Simmons and Buckley 1992; Augusto et al. 2002), while our results and those of Mölder et al. (2008) demonstrate the negative effect of beech-dominated canopy on herb layer diversity. The most important mechanisms of beech influence on the understorey are the reduction in light, forming of thick litter layer and root competition for water. Beech produces a large amount of litter annually. The rate of leaf-litter biomass on the total biomass of broadleaves is much higher than in the conifers (Packham et al. 1992). Mölder et al. (2008) surmised that herbaceous understorey diversity is influenced also by litter layer thickness. On the other hand, Slavíková (1986) demonstrated by field experiment that the phenomenon of “nudal” or “herbless” beech forests is caused primarily by the root competition for water and soluble nutrients, not only by the direct effect of thick litter layer. Intensive transpiration ability and dense root system of beech result into the effective depletion of soil water (Čermák et al. 1993; Pichler et al. 2006; Schume et al. 2004). Beech stands show high transpiration rates both under the humid and under partly dry summer climates in the Central Europe (Granier et al. 2000). Therefore, beech is the superior competitor for water (Schume et al. 2004). Its transpiration rate in the Slovak mountain forests was found one-third higher than the rate of spruce (Střelcová et al. 2006). According to Jennings et al. (1999), F. sylvatica is a strong shade-creating species, and richness is inhibited by the lack of light. Extraordinary crown growth plasticity of beech enables this tree species to fill the tree-layer canopy completely (Otto 1994) and to cover the canopy gaps more quickly then spruce. Understorey light availability in the vegetation period can reach only 1.5–2.5 % of total solar radiation in the closed beech stands (van Eimern 1984), while the solar transmissivity through conifer canopy of uniform stands can be about 25–30 % (Hardy et al. 2004). On the other hand, there is opposite situation in the beginning of vegetation period when broadleaves are not foliated. It is reflected in higher proportion of early-spring species (most often geophytes) in beech-dominated stands, such as M. perennis, D. enneaphyllos, D. bulbifera or H. epipactis. The light transmissivity is closely related to the canopy cover that is controlled by thinning in managed forests. Therefore, less difference in the similarly managed stands are expected.
Ewald (2008) hypothesized that richness is controlled chiefly by herb layer cover (biomass), which could be predicted by a linear combination of tree layer cover, elevation, C/N ratio and moisture. Also, Mölder et al. (2008) found that herb layer biomass in deciduous forest is positively correlated with the herb layer diversity. Herb cover, and consequently species richness, is negatively correlated with tree canopy cover that controls light availability in the forest understorey. Light availability increases with P. abies proportion in canopy and spruce, and mixed spruce-beech stands have more heliophytes than pure beech stands (Lücke and Schmidt 1997). This offers an explanation for the increased richness in spruce stands. Spruce stands in our study had several light-requiring species such as Senecio nemorensis agg., R. idaeus and E. montanum (Ujházy et al. 2005; Šamonil and Vrška 2008) or Luzula pilosa, V. officinalis and F. vesca which are considered as species with a positive correlation to light by Tinya et al. (2009). However, mean indicator values for light within relevé groups support the idea only in the case of crystalline area, where the significant increase in values were found (Fig. 4). Only the mixed stands showed higher values for light in the calcareous region. This is probably due to the fact that many unique species of the mixed stands are rather light-requiring, such as Calamagrostis varia, Lotus corniculatus, Carduus defloratus, Valeriana tripteris, Leontodon incanus, Laserpitium latifolium, Sesleria albicans and others. The highest number of unique species together with the prevalence of the calciphilous grasses indicates special site conditions. The comparison of bedrock types among three groups of stands showed that while in pure beech and spruce stands prevail limestones and marly limestones, the most frequent bedrock of mixed stands is dolomite. The results of Fajmonová (1971, 1973, 1978) showed, that communities with the dominance of calciphilous grasses occur very often on sites with the dolomite bedrock, high proportion of rock outcrop and shallow soils, with higher calcium carbonate content in topsoil in comparison with the sites on limestones. The silvicultural effort consisted of planting pure spruce stands regardless of the parent rock type. Higher frequency of mixed stands on dolomites can be explained by the inhibition of spruce growth by the calcareous substrates and rock outcrops (Seynave et al. 2005). Less vigorous growth of spruce in the carbonate areas could be also the reason, why number of herb species in the pure spruce stands compared with pure beech increased more on carbonates than on crystallines. Beech and the other tree species are limited by the lower soil fertility on dolomites, as well. Therefore, prevalence of a single-tree species is less frequent than on the other bedrock types and tree influence on herb layer is limited.
More favourable light conditions under the pure spruce canopies could lead also into the increasing frequency of several species (e.g. V. reichenbachiana, S. europaea, G. hederacea agg.); however, they are very frequent also in the pure beech stands. More light at the forest floor causes changes in the top-soil properties due to the faster decomposition of litter (Mladenoff 1987; Zhang and Liang 1995; Ritter 2005; Berg and McClaugherty 2008). Thus, on the well-buffered soils (especially on the calcareous bedrocks), active humus forms may develop from spruce litter resulting in the conditions similar to the beech-dominated communities.
Results can be affected by local differences in the management form, of course. Unfortunately, detailed long-term historical evidence of management events within both areas is not available. Planting of conifers was widely adopted to the forestry practise in the eighteenth century in Slovakia, in general. Higher mountain ranges (including studied areas) were affected later than the other regions with better accessibility. Therefore, we consider spruce stands to be from the 1st to 3rd generation. More likely, 1st and 2nd generation prevails as a majority of original species of natural communities still occur in the studied spruce-dominated stands. It was found, that eutrophic beech forest species (Querco-Fagetea) frequently persist on the flysch bedrock in the Western Carpathians within the first two generations and disappear in the 3rd generation of spruce plantations (Fajmonová 1974); however, the evidence of this effect from the studied regions is still missing.
Conclusions
Spruce significantly affects the herb layer and causes quite large changes in its species composition. A species group positively associated with increasing spruce cover regardless of bedrock type was identified. Increased cover of P. abies very often entails species turnover without concurrent reduction in overall vegetation diversity. A negative effect on species diversity was not observed.
Measures of diversity do not reflect the fact of species turnover which is detected by the analysis of floristic similarity. Therefore, changes in vegetation diversity must be addressed by evaluating species turnover and not only diversity measures. They should be related to natural reference stands in more or less untouched forests.
Our results show not only that species composition is specific for different bedrock types but also that the reaction of particular species to increasing spruce cover is highly specific to bedrock. Species diversity is higher on calcareous bedrock, and the original herb layer composition is less changed here compared with crystalline bedrock. Despite the fact that species richness usually increased and the majority of species was common to natural as well as modified stands, a shift from natural species diversity of beech forests towards artificial spruce forests is evident. Species turnover induced by spruce planting in both mountain ranges and bedrock types shows a significant shift in the species composition from the Querco-Fagetea towards Vaccinio-Piceetea class, which is followed by a decrease in Ellenberg’s indicator value of soil reaction in crystalline region. The risk of losses in original species diversity is particularly high when monodominant spruce stands are planted on the crystalline bedrock, but typical beech-forest species are also seriously affected on the calcareous bedrock. We expect that the effect will be amplified after repeated spruce planting.
On the other hand, minor spruce admixture may have a positive effect on herb diversity. The effects of limited spruce admixtures on soil properties (Rothe et al. 2002) and ground vegetation are mild. In order to preserve natural plant diversity and minimize other risks associated with soil acidification, spruce cover should be limited at sites of original beech and fir-beech montane forests. To quantify the feasible limits of spruce admixture, further research effort oriented to the “tree-herb” interactions is needed, especially regarding its temporal dynamics with respect to the stand rotation time and the number of planted generations of spruce.
References
Asch K (2005) The 1:5 Million International Geological Map of Europe and Adjacent Areas. BGR (Hannover), 1 map
Augusto L, Ranger J, Binkley D, Rothe A (2002) Impact of several common tree species of European temperate forests on soil fertility. Ann For Sci 59:233–253
Barbier S, Gosselin F, Balandier P (2008) Influence of tree species on understory vegetation diversity and mechanisms involved—a critical review for temperate and boreal forests. For Ecol Manage 254(1):1–15
Barkman JJ, Doing H, Segal S (1964) Kritische Bemerkungen und Vorschläge zur quantitativen Vegetationsanalyse. Acta Botanica Neerlandica, Amsterdam 13:394–419
Beers TW, Dress PE, Wensel LC (1966) Aspect transformation in site productivity research. J For 64:691–692
Berg B, McClaugherty C (2008) Plant litter: decomposition, humus formation, carbon sequestration, 2nd edn. Springer, Berlin
Bezák V, Hraško L’, Kováčik M, Madarás J, Siman P, Pristaš J, Dublan L, Konečný V, Plašienka D, Vozárová A, Kubeš P, Švasta J, Slavkay M, Liščák P (1999) Geological map of Slovenské rudohorie Mts.—western part. Regional geological maps of Slovakia, scale 1:50 000. State Geological Institute of Dionýz Štúr. Bratislava
Blasi C, Marchetti M, Chiavetta U, Aleffi M, Audisio P, Azzella MM, Brunialti G, Capotorti G, Del Vico E, Lattanzi E, Persiani AM, Ravera S, Tilia A, Burrascano S (2010) Multi-taxon and forest structure sampling for identification of indicators and monitoring of old-growth forest. Plant Biosyst 144(1):160–170
Blattný T, Št’astný T (1959) Prirodzené rozšírenie lesných drevín na Slovensku (Natural distribution of tree species in Slovakia), Slovenské vydavatel’stvo pôdohospodarskej literatúry, Bratislava
Bohn U, Gollub G, Hettwer C, Neuhäuslová Z, Schlüter H, Weber H (2003) Map of the natural vegetation of Europe. Bundesamt für Naturschutz, Bonn
Bruelheide H (2000) A new measure of fidelity and its application to defining species groups. J Veg Sci 11:167–178
Bürger R (1991) Immissionen und Kroenverlichtung als Ursachen für Veränderungen der Waldbodenvegetation im Schwarzwald. Tüexenia 11:407–424
CBD-Convention on Biological Diversity (2011) Report of the tenth meeting of the conference of the parties to the convention on biological diversity, 341 http://www.cbd.int/doc/meetings/cop/cop-10/official/cop-10-27-en.doc
Čermák J, Matyssek R, Kučera J (1993) A cause of a rapid decline of large beech trees on heavy-textured soils after substantial opening of the canopy. Lesnictví 39:175–183
Chytrý M, Tichý L (2003) Diagnostic, constant and dominant species of vegetation classes and alliances of the Czech Republic: a statistical revision. Masaryk University, Brno
Chytrý M, Exner A, Hrivnák R, Ujházy K, Valachovič M, Willner W (2002) Context-dependence of diagnostic species: a case study of the central European spruce forests. Folia Geobotanica 37:403–417
EC-European Commission (2011) EU biodiversity strategy to 2020, COM(2011) 244 final, 16 http://ec.europa.eu/environment/nature/biodiversity/comm2006/pdf/2020/1_EN_ACT_part1_v7%5B1%5D.pdf
EEA-European Environment Agency (2007) Halting the loss of biodiversity by 2010: proposal for a first set of indicators to monitor progress in Europe. Technical report No. 11/2007, EEA, Copenhagen
EEA-European Environment Agency (2010) Assessing biodiversity in Europe—the 2010 report. Report No. 5/2010, EEA, Copenhagen
Ellenberg H, Weber HE, Düll R, Wirth W, Werner W, Pauliben D (1992) Zeigenwerte von Pflanzen in Mitteleuropa, Ed. 2., Scripta Geobotanica, Göttingen 18:1–258
Emmer IM (1995) Humus form development and succession of dwarf shrub vegetation in grass dominated primary Pinus sylvestris forest. Ann For Sci 52:561–571
Emmer IM, Fanta J, Kobus AT, Kooijman A, Sevink J (1998) Reversing borealization as a means to restore biodiversity in Central-European mountain forests—an example from the Krkonose Mountains, Czech Republic. Biodivers Conserv 7(2):229–247
Emmer IM, Wessel WW, Kooijman A, Sevink J, Fanta J (2000) Restoration of degraded Central-European mountain forest soils under changing environmental circumstances. In: Klimo E, Hager H, Kulhavy J (eds) Spruce monocultures in Central Europe—problems and prospects. EFI Proceedings 33:81–92
Ewald J (2000a) The influence of coniferous canopies on understorey vegetation and soils in mountain forests of the northern Calcareous Alps. Appl Veg Sci 3:123–134
Ewald J (2000b) The partial influence of Norway spruce stands on understorey vegetation in Montane forests of the Bavarian Alps. Mountain Res Dev 20(4):364–371
Ewald J (2003) The calcareous riddle: why are there so many calciphilous species in the Central European flora? Folia Geobotanica 38:357–366
Ewald J (2008) Plant species richness in mountain forests of the Bavarian Alps. Plant Biosyst 142(3):594–603
Ewald J (2009a) Bimodale Spektren von Nährstoffzeigerwerten in Bayerns Nadelwäldern. Forstarchiv 80:189–194
Ewald J (2009b) Epigeic bryophytes do not improve bioindication by Ellenberg values in mountain forests. Basic Appl Ecol 10:420–426
Fabiánek T, Menšík L, Tomášková I, Kulhavý J (2009) Effects of spruce, beech and mixed commercial stand on humus conditions of forest soils. J For Sci 55(3):119–126
Fajmonová E (1971) Príspevok k fytocenológii vápencových bučín stredného Považia (Contribution to the phytosociology of beech forests on limestons of middle Váh river basin). Biológia 26:517–529
Fajmonová E (1973) Fytocenologické zatriedenie submontánnych bylinných vápencových bučín na strednom Považí (Phytosociological classification of submontane beech forests on limestones in middle Váh river basin). Biológia 28:537–545
Fajmonová E (1974) Some results of the study of the secondary and natural growths of conifers in the Javorníky range. Biológia 29:537–549
Fajmonová E (1977) Some results of studying the influence of the spruce on the sorption complex of the soil. Biológia 32:759–767
Fajmonová E (1978) Waldgesellschaften der Strážov – Berggruppe (nordteil des Gebirges Strážovská hornatina). Acta Facultatis Rerum naturalis universitas comeniae, Botanica 26, Bratislava, pp 87–105
Fanta J (1997) Rehabilitating degraded forests in Central Europe into self-sustaining forest ecosystems. Ecol Eng 8:289–297
Faško P, Št’astný P (2002) Mean annual precipitation totals – In: Landscape Atlas of the Slovak Republic, Ministerstvo životného prostredia SR, Bratislava, Slovenská agentúra životného prostredia, Banská Bystrica
Gilliam FS (2007) The ecological significance of the herbaceous layer in temperate forest ecosystems. Bioscience 57(10):845–858
Granier A, Biron P, Lemoine D (2000) Water balance, transpiration and canopy conductance in two beech stands. Agric For Meteorol 100:291–308
Hadač E, Sofron J (1980) Notes on syntaxonomy of cultural forest communities. Folia Geobotanica et Phytotaxonomica 15:245–258
Hardy JP, Melloh R, Koenig G, Marks D, Winstral A, Pomeroy JW, Link T (2004) Solar radiation transmission through conifer canopies. Agric For Meteorol 126(3–4):257–270
Husová M, Jirásek J, Moravec J (2002) Vegetation survey of the Czech Republic. vol. 3, Coniferous forests. Academia, Praha
IUSS Working Group WRB (2006) World reference base for soil resoruces 2006, 2nd edn. World soil resources report no. 103, FAO, Rome
Jarolímek I, Šibík J (2008) Diagnostic, constant and dominant species of the higher vegetation units of Slovakia. Veda, Bratislava
Jennings SB, Brown ND, Sheil D (1999) Assessing forest canopies and understorey illumination: canopy closure, canopy cover and other measures. Forestry 72:59–73
Jonášová M, Prach K (2004) Central-European mountain spruce (Picea abies (L.) Karst.) forests: regeneration of tree species after a bark beetle outbreak. Ecol Eng 23:15–27
Jurko A (1990) Ekologické a socioekonomické hodnotenie vegetácie (Ökologische und sozioökonomische Bewertung der Vegetation). Príroda, Bratislava
Klotz S, Kühn I, Durka W (eds) (2002) BIOLFLOR—Eine Datenbank zu biologisch-ökologischen Merkmalen der Gefäßpflanzen in Deutschland. Schriftenreihe für Vegetationskunde 38. Bundesamt für Naturschutz, Bonn
Kuuluvainen T, Pukkala T (1989) Effect of Scots pine seed trees on the density of ground vegetation and tree seedlings. Silva Fennica 23:159–167
Liira J, Kohv K (2010) Stand characteristics and biodiversity indicators along the productivity gradient in boreal forests: defining a critical set of indicators for the monitoring of habitat nature quality. Plant Biosyst 144(1):211–220
Lücke K, Schmidt W (1997) Vegetation und Standortsverhältnisse in Buchen-Fichten-Mischbeständen des Sollings. Forstarchiv 68:135–143
Máliš F, Rudaš B, Kontriš J (2005) Soil conditions of forest communities in the area of Bykovo massif. The fourth pedology days in Slovakia. Soil Science and Conservation Research Institute, Bratislava, pp 226–231
Máliš F, Vladovič J, Čaboun V, Vodálová A (2010) The influence of Picea abies on herb vegetation in forest plant communities of Veporské vrchy Mts. J For Sci 56(2):58–67
Marhold K, Hindák F (1998) Checklist of non-vascular and vascular plants of Slovakia. Veda, Bratislava
MCPFE, Ministerial Conference on the Protection of Forests in Europe (2002) Improved Pan-European indicators for sustainable forest management http://www.unece.org/timber/docs/stats-25/supp/WA2-2.pdf
Mladenoff DJ (1987) Dynamics of nitrogen mineralization and nitrification in hemlock and hard wood treefall gaps. Ecology 68:1171–1180
Mölder A, Bernhardt-Römermann M, Schmidt W (2008) Herb-layer diversity in deciduous forests: Raised by tree richness or beaten by beech? For Ecol Manage 256:272–281
Moravec J, Husová M, Chytrý M, Neuhauslová Z (2000) Přehled vegetace České republiky, Svazek 2, Hygrofilní, mezofilní a xerofilní opadavé lesy. Academia, Praha
Neteler M, Mitasova H (2008) Open source GIS: a GRASS GIS approach. 3rd edn., 80 illus., Springer, New York
Økland RH (1990) Vegetation ecology: theory, methods and applications with reference to Fennoscandia. Sommerfeltia Suppl 1:233
Otto HJ (1994) Waldökologie. Verlag Eugen Ulmer, Stuttgart
Packham JR, Harding DJL, Hilton GM, Stuttard RA (1992) Functional ecology of woodlands and forests. Chapman and Hall, Cambridge
Petriccione B, Cindolo C, Cocciufa C, Ferlazzo S, Parisi G (2007) Development and harmonization of a Forest Status Indicator (FSI). Italian Forest Service, Roma
Pichler V, Gregor J, Váľka J, Capuliak J, Homolák M (2006) Forest stand (Fagus sylvatica L.) density reduction as a global climate change adaptive forestry tool. For J 52(1–2):89–97
Pielou EC (1975) Ecological diversity. Wiley, New York
Pitkänen S (1998) The use of diversity indices to assess the diversity of vegetation in managed boreal forests. For Ecol Manage 112:121–137
Polák M, Bujnovský A, Kohút M (1997) Geological map of the Vel’ká Fatra Mts. Regional geological maps of Slovakia, scale 1:50 000. State Geological Institute of Dionýz Štúr. Bratislava
Poleno Z (2001) Vliv přeměny smrkové monokultury na stav a vývoj lesní půdy a bylinné vegetace (Influence of transformation of spruce monocultures on state and development of forest soil and herbal vegetation). Rep For Res 46:6–15
Pott R (2000) Palaeoclimate and vegetation—long-term vegetation dynamics in central Europe with particular reference to beech. Phytocoenologia 30(3–4):285–333
Quine ChP, Humphrey JW (2010) Plantations of exotic tree species in Britain: irrelevant for biodiversity or novel habitat for native species? Biodivers Conserv 19(5):1503–1512
Randuška D, Vorel J, Plíva K (1986) Fytocenológia a lesnícka typológia. Príroda, Bratislava
Reger B, Kölling C, Ewald J (2011) Modelling effective thermal climate for mountain forests in the Bavarian Alps: which is the best model? J Veg Sci 22:677–687
Ritter E (2005) Litter decomposition and nitrogen mineralization in newly formed gaps in a Danish beech (Fagus sylvatica L.) forest. Soil Biol Biochem 37(7):1237–1247
Rothe A, Kreutzer K, Küchenhoff H (2002) Influence of tree species composition on soil and soil solution properties in two mixed spruce-beech stands with contrasting history in Southern Germany. Plant Soil 240:47–56
Saetre P (1999) Spatial patterns of ground vegetation, soil microbial biomass and activity in a mixed spruce-birch stand. Ecography 22:183–192
Šály R (1978) Pôda, základ lesnej produkcie (Soil, the Base of Forest Production). Bratislava, Príroda
Šamonil P, Vrška T (2008) Long-term vegetation dynamics in the Šumava Mts. natural spruce-fir-beech forests. Plant Ecol 196:197–214
Schmidt W (2005) Herb layer species as indicators of biodiversity of managed and unmanaged beech forests. For Snow Landsc Res 79(1/2):111–125
Schmidt I, Zerbe S, Betzin J, Weckesser M (2006) An Approach to the identification of indicators for forest biodiversity—the solling mountains (NW Germany) as an example. Restor Ecol 14(1):123–136
Seynave I, Gegout JC, Herve JC, Dhote JF, Drapier J (2005) Picea abies site index prediction by environmental factors and understory vegetation: a two-scale approach based on survey databases. Can J For Res 35(1):1669–1678
Simmons EA, Buckley GP (1992) Ground vegetation under planted mixtures of trees. In: Cannel MGR, Malcolm DC, Robertson PA (eds) The ecology of mixed-species stands of trees. Blackwell, Oxford, pp 211–232
Šimurdová B (2001) Cultural spruce forests of the Hnilec watershed. Bull Slovak Bot Soc 23:141–147
Slavíková J (1986) Ekologie rostlin (Plant Ecology). SPN, Praha
Šomšák L (2003) Effect of secondary spruce forests on phytoenvironment in the Slovenské rudohorie mountains. Folia oecologica 30:41–59
Šomšák L, Balkovič J (2002) Cyclic succession and plant biodiversity within the secondary spruce forests in the Hnilec river watershed, Bratislava. Phytopedon 1:45–51
Št’astný P, Nieplová E, Melo M (2002) Mean annual air temperature – In: Landscape Atlas of the Slovak Republic (2002), Ministerstvo životného prostredia SR, Bratislava, Slovenská agentúra životného prostredia, Banská Bystrica
StatSoft Inc. (2005) Statistica 7.1, StatSoft Inc., Tulsa
Schume H, Jost G, Hager H (2004) Soil water depletion and recharge patterns in a mixed and pure forest stands of European beech and Norway spruce. J Hydrol 289:258–274
Střelcová K, Mind’áš J, Škvarenina J (2006) Influence of tree transpiration on mass water balance of mixed mountain forests of the West Carpathians, Biológia. Bratislava 61(Suppl. 19):305–310
Tárrega R, Calvo L, Taboada Á, Marcos E, Marcos JA (2011) Do mature pine plantations resemble deciduous natural forests regarding understory plant diversity and canopy structure in historically modified landscapes? Eur J For Res. doi:10.1007/s10342-011-0493-5
Ter Braak CJF, Šmilauer P (2002) CANOCO Reference manual and CanoDraw for Windows User guide: Software for Canonical Community Ordination (version 4.5), Ithaca, Microcomputer Power
Teuscher F (1985) Fichtenforste im Mittelland. Schweizer Zeitschrift für Forstwesen 136:755–761
Tichý L (2002) JUICE, software for vegetation classification. J Veg Sci 13:451–453
Timár G, Molnár G, Székely B, Biszak S, Varga J, Jankó A (2006) Digitized maps of the Habsburg Empire—the map sheets of the second military survey and their georeferenced version. Arcanum, Budapest, 59
Tinya F, Márialigeti S, Király I, Németh B, Ódor P (2009) The effect of light conditions on herbs, bryophytes and seedlings of temperate mixed forests in Őrség, Western Hungary. Plant Ecol 204(1):69–81
Ujházy K, Križová E, Vančo M, Freňáková E, Ondruš M (2005) Herb layer dynamics of primeval fir-beech forests in central Slovakia. In: Commarmot B, Hamor FD (eds) Natural forests in the temperate zone of Europe—values and utilisation. Swiss Federal Research Institute WSL, Birmensdorf & Carpathian Biosphere Reserve, Rakhiv, pp 193–202
Ulrich B (1983) Stabilität von Waldokosystemen unter dem Einfluss des Sauren Regens. Allgemeine Forstzeitschrift 26(27):670–677
Van Eimern J (1984) Variations of the Radiation within and above a beech forest during a phenological year. Geo J 8(3):271–275
Van Oijen D, Feijen M, Hommel P, Den Ouden J, De Waal R (2005) Effects of tree species composition on within-forest distribution of understorey species. Appl Veg Sci 8:155–166
Vladovič J (2003) Oblastné východiská a princípy hodnotenia drevinového zloženia a ekologickej stability lesov Slovenska (Regional basis and principles for assessment of tree species composition and ecological stability of forests in Slovakia). Lesnícke štúdie 57, Príroda, Bratislava
von Hornstein F (1951) Wald und Mensch Waldgeschichte des Alpenvorlandes Deutschland, Österreichs un der Schweiz. Ravensburg, Otto Maier Verlag
Willner W, Grabherr G (2007) Die Wälder und Gebüsche Östererichs. Elsevier, München, 1. Textband, 302, 2. Tabellenband
Wood JD (1996) The geomorphological characterisation of digital elevation models PhD Thesis. University of Leicester, UK, http://www.soi.city.ac.uk/~jwo/phd
Wulf M, Naaf T (2009) Herb layer response to broadleaf tree species with different leaf litter quality and canopy structure in temperate forests. J Veg Sci 20:517–526
Zhang Q, Liang Y (1995) Effects of gap size on nutrient release from plant litter decomposition in a natural forest ecosystem. Can J For Res 25:1627–1638
Zlatník A (1976) Lesnická Fytocenologie (Forest Phytosociology). Státní zemědělské nakladatelství, Praha
Acknowledgments
This paper is dedicated to the memory of Jozef Ištoňa, who helped us with data acquisition and provided many valuable comments. We are grateful to the anonymous reviewers for many helpful comments and suggestions improving results and text and to Dušan Gömöry and Michal Bošel’a for consultations on statistical methods. This work was supported by the Slovak Research and Development Agency under contracts No. APVV-0632-07, No. APVV-0022-07 and No. APVT-27-009304 and by the Slovak Grant Agency VEGA (project No. 1/0831/09).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by C. Ammer.
Rights and permissions
About this article
Cite this article
Máliš, F., Ujházy, K., Vodálová, A. et al. The impact of Norway spruce planting on herb vegetation in the mountain beech forests on two bedrock types. Eur J Forest Res 131, 1551–1569 (2012). https://doi.org/10.1007/s10342-012-0624-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-012-0624-7