
Abstract
Well-established studies have shown an elevated level of reactive oxygen species (ROS) that induces oxidative stress in the Alzheimer’s disease (AD) patient’s brain and an animal model of AD. Herein, we investigated the underlying anti-oxidant neuroprotective mechanism of natural dietary supplementation of anthocyanins extracted from Korean black beans in the amyloid precursor protein/presenilin-1 (APP/PS1) mouse model of AD. Both in vivo (APP/PS1 mice) and in vitro (mouse hippocampal HT22 cells) results demonstrated that anthocyanins regulate the phosphorylated-phosphatidylinositol 3-kinase-Akt-glycogen synthase kinase 3 beta (p-PI3K/Akt/GSK3β) pathways and consequently attenuate amyloid beta oligomer (AβO)-induced elevations in ROS level and oxidative stress via stimulating the master endogenous anti-oxidant system of nuclear factor erythroid 2-related factor 2 (Nrf2) and heme oxygenase-1 (Nrf2/HO-1) pathways and prevent apoptosis and neurodegeneration by suppressing the apoptotic and neurodegenerative markers such as activation of caspase-3 and PARP-1 expression as well as the TUNEL and Fluoro-Jade B-positive neuronal cells in the APP/PS1 mice. In vitro ApoTox-Glo™ Triplex assay results also showed that anthocyanins act as a potent anti-oxidant neuroprotective agent and reduce AβO-induced neurotoxicity in the HT22 cells via PI3K/Akt/Nrf2 signaling. Importantly, anthocyanins improve memory-related pre- and postsynaptic protein markers and memory functions in the APP/PS1 mice. In conclusion, our data suggested that consumption and supplementation of natural-derived anti-oxidant neuroprotective agent such as anthocyanins may be beneficial and suggest new dietary-supplement strategies for intervention in and prevention of progressive neurodegenerative diseases, such as AD.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Alzheimer’s disease (AD) is the most common age-related neurodegenerative disorder and is characterized by progressive learning and memory deficits. The neuropathological hallmarks of AD include senile plaques, neurofibrillary tangles, synaptic deficits, and neurodegeneration of the brain. Senile plaques are extracellular accumulations of amyloid beta (Aβ) peptide, and neurofibrillary tangles are composed of hyperphosphorylated tau protein [1]. Aβ peptides are produced by cleavage of the transmembrane glycoprotein amyloid precursor protein (APP) by the β-secretase and presenilin-dependent γ-secretase enzymes. According to Hardy and Selko and the Aβ cascade hypothesis, Aβ is the major cause of neurotoxic insults in AD; it activates a number of biochemical pathogenic mediators, which lead to AD pathology, including oxidative stress and synaptic dysfunction [2,3,4]. However, the exact underlying molecular mechanisms associated with dementia remain unknown. Importantly, reactive oxygen species (ROS)-induced oxidative stress triggers AD pathology mediators because the brain, as a result of its increase consumption of oxygen and limited anti-oxidant enzymes, is the most sensitive region to oxidative stress [5]. Nuclear factor erythroid 2-related factor 2 (Nrf2) is a key redox-regulated gene that has a critical role against oxidative stress, and the level of Nrf2 in the nuclei is decreased in the neurological disorders such as AD [6]. Recently, accumulating studies have reported that activated nuclear Nrf2 further regulates several endogenous redox-regulated enzymes such as heme oxygenase-1 (HO-1) and glutathione cysteine ligase modulatory subunit (GCLM) via the phosphorylated phosphatidylinositol 3-kinase, phosphorylated Akt, and phosphorylated glycogen synthase kinase 3 beta (p-PI3K/Akt/GSK3β) pathway. The PI3K/Akt/GSK3β signaling is the most important pathways, having a key and pivotal role in various survival signaling and in other functions. Impaired PI3K/Akt/GSK3β signaling has been investigated in the AD brain and AD mouse models [7,8,9,10,11,12,13,14,15]. Notably, HO-1 is beneficial in various diseases, and its activation improves learning and memory functions [6, 16]. Recently, it was reported that nuclear translocation of Nrf2 increased the expression of HO-1 [17]. Elevated expression of Nrf2 in both in vitro and in vivo AD models decreased Aβ-induced neurodegeneration and oxidative stress [18]. Similarly, Kanninen et al. revealed that overexpression of hippocampal Nrf2 improves spatial learning and memory in a mouse model of AD [19].
Natural plant-derived polyphenolic flavonoids have beneficial effects on health and possess versatile biological activity, such as anti-oxidant, anti-neuroinflammatory, anti-cancer, and anti-neurodegenerative actions. The development and use of natural substances, particularly polyphenolic flavonoids, may prevent various metabolic disorders, such as diabetes mellitus and obesity; cardiovascular disease; and chronic neurodegenerative disorders, such as AD [20,21,22,23,24,25,26]. Previously, we evaluated polyphenolic anthocyanins from Korean black consists of cyanidine-3-glucosides (C3G) (67%), delphinidin-3-glucosides (D3G) (25%), and petunidin-3-glucosides (P3G) (5%), and showed that these compounds exert neuroprotective effects, via attenuation of oxidative stress, against D-galactose- and ethanol-induced neurodegeneration in the adult, neonatal and developing rat brain respectively; and reduce kainic acid-induced excitotoxicity and apoptosis in hippocampal neuronal cells [27,28,29,30,31]. It has been reported that anthocyanins and C3G protect against cerebral ischemia and Aβ-induced mitochondrial degeneration [32, 33]. Dietary flavonoids such as anthocyanins improved spatial working memory in aged and LPS-treated rodents [34, 35]. Recently, we also found that Korean black bean anthocyanins exert neuroprotective effects against Aβ1–42 in both in vitro and in vivo Aβ1–42 models [36, 37]. In the present study, we hypothesized that Korean black bean anthocyanins, a natural potent anti-oxidant neuroprotective agent, would reduce synaptic and memory dysfunction and neurodegeneration by attenuating Aβ-induced ROS-mediated oxidative stress via regulation of the anti-oxidant neuroprotective mechanism of the PI3K/Akt/Nrf2 pathways in the in vitro AD model and APP/PS1 transgenic mouse model of AD.
Materials and Methods
Materials
Anthocyanins were extracted from Korean black bean, as previously described [27]. Aβ1–42 peptide, 2′7′-dichlorodihydrofluorescein diacetate (DCFH-DA), 2-(4-Morpholinyl)-8-phenyl-1(4H)-benzopyran-4-one hydrochloride (LY294002), propidium iodide (PI), 4′,6′-diamidino-2-phenylindole (DAPI) and dimethyl sulfoxide (DMSO), diaminobenzidine tetrahydrochloride (DAB) were purchased from Sigma Chemical Co. (St. Louis, MO, USA).
Animals Grouping and Drug Treatments
Male C57BL/6N wild type (WT) mice were purchased from Samtako Bio (Osan, Republic of Korea) and double transgenic B6.Cg-Tg (APPswe, PSENdE9)85Dbo/Mmjax (APP/PS1) AD-model mice were purchased from Jackson Laboratory (Bar Harbor, ME, USA). The double transgenic mice express a chemiric mouse-human amyloid precursor protein bearing Swedish mutation (Mo/HuAPP695swe) as well as a mutant human presenilin 1 protein (PS1-dE9) in the brain. The mice were kept in the university animal house under a 12-h/12-h light/dark cycle at 23 °C, 60 ± 10% humidity with food and water ad libitum. The mice when reached to 10 month of age were brought to injection and behavioral room to acclimatize for 1 week.
The animals were grouped as following: (1) WT vehicle (Veh)-treated, (2) APP/PS1 (Veh)-treated, (3) APP/PS1 anthocyanins-treated, and (4) WT anthocyanins-treated.
Anthocyanins were dissolved in DMSO and make the final administered amount in the normal saline. Anthocyanins were intraperitoneally (i.p.) administered at dose of 12 mg/kg/day body weight for 30 days to the APP/PS1 and WT mice. Similarly, the same amount of DMSO and volume of normal saline as a Veh injected to the WT and APP/PS1 mice.
After the treatment and behavioral analysis, the experimental animals were sacrificed for further biochemical and immunohistochemical analysis.
Behavioral Study
The behavioral study was performed after the completion of the treatment for all the experimental mice using a Morris water maze (MWM) task and Y-maze task.
MWW test was performed with some modification as we described previously [38]. Briefly, each mouse received four training periods per day for six consecutive days. Latency to escape from the water maze (finding the submerged escape platform) was calculated for each trial. On day 7, the probe test was performed by removing the platform and allowing each mouse to swim freely for 60 s. The time that the mice spent swimming in the target quadrant and the number of platform crossings (where the platform was located during hidden platform training) was measured. All data were recorded using video-tracking software (SMART, Panlab Harvard Apparatus, Bioscience Company, USA).
Y-maze test was performed as we described previously [39]. Briefly, each mouse was placed at the center of the apparatus and allowed to move freely through the maze for three 8-min sessions. The series of arm entries was noted. Spontaneous alternation was defined as the successive entry of the mouse into the three arms in overlapping triplet sets. Alternation behavior (%) was calculated as [successive triplet sets (entries into three different arms consecutively)/total number of arm entries-2] × 100.
Protein Extraction from Mouse Brain
After behavioral analysis the mice were sacrificed, brains were immediately removed, hippocampus was dissected carefully and the tissue were frozen on dry ice and stored at − 80°C. The hippocampus tissue was homogenized in pro-prep™ protein extraction solution according to the provider instruction (iNtRON, Biotechnology, Inc) using homogenizer (DAIHAN, Scientific Co, Ltd., Republic of Korea) having specifications e.g., 230 V, 50/60 Hz; Watts 280 W, Fuse 250 V, T3AL. We calculated the amount of homogenized proteins in pro-prep™ protein extraction solution and further some of the amount were separated into the cytosolic and nuclear fractions using a nuclear and cytoplasmic protein extraction kit, according the manufacturer’s instruction (catalog # K266-25) Biovision Incorporated, A 95035 USA. The cytosolic and nuclear fractions protein were collected and stored at − 80°C. The remaining protein of the hippocampus homogenate in pro-prep™ protein extraction solution were then centrifuged at 10000×g at 4°C for 25 min (min). The supernatants were collected and stored at − 80°C.
Western Blot Analysis
The protein concentration from tissue homogenates and cell lysates were measured (BioRad protein assay kit, BioRad Laboratories, CA, USA). Equal amounts of protein (15–25 μg) were electrophoresed under same experimental condition using 4–12% Bolt™ Mini Gels and MES SDS running buffer 1× (Novex, Life Technologies, Kiryat Shmona, Israel), broad range prestained protein marker (GangNam stain™, Intron Biotechnology) was used as molecular size control. The membranes were blocked in 5% (w/v) skim milk to reduce non-specific binding and incubated with primary antibodies (detailed information provided in the Table 1) overnight at 4°C. After reaction with a horseradish peroxidase-conjugated secondary antibody, as appropriate. Secondary antibodies HRP-conjugated IgG used were goat-anti-mouse (1: 3000) was purchased from Promega, Madison, WI, USA, goat-anti-rabbit (SC: 2004) (1: 2000–2500), mouse-anti-rabbit (SC: 2354) (1: 2000–2500), mouse-anti-goat (SC: 2354) (1: 4000–5000) were purchased from Santa Cruz Biotechnology, USA. After incubation with secondary antibodies, the membranes were washed three time with 1X TBST. After washing, the proteins were detected using an ECL detection reagent according to the manufacturer’s instructions (Amersham Pharmecia Biotech, Uppsala, Sweden). The X-ray films were scanned, and the optical densities of the bands were analyzed through densitometry using the computer-based Sigma Gel program, version 1.0 (SPSS, Chicago, IL, USA).
Aβ1–42 Oligomer (AβO) Preparation for In Vitro Experiments
For in vitro experiments, AβO was prepared as we described previously [39].
ROS Assay both in Mouse Hippocampus and In Vitro HT22 Cells
ROS assay was performed to determine the elevated ROS in the hippocampus (n = 8 mice per group) homogenates. The assay was based on the oxidation of DCFH-DA to 2′7′-dichlorofluorescein (DCF) as we described previously [38]. The conversion of DCFH-DA to the DCF was analyzed by spectrofluorimeter with excitation at 484 nm and emission at 530 nm. For background fluorescence (conversion of DCFH-DA to DCF in the absence of homogenate), was run a parallel blanks and used as a standard DCF. We quantified ROS production from a DCF-standard curve and expresses as relative DCF pmol/mg protein.
Further we also analyzed intracellular ROS via DCF fluorescence intensity through confocal microscopy as we described previously [29] with some modification in vitro using mouse hippocampal (HT22) neuronal cells, a kind gift from Prof. Koh (Gyeongsang National University, S. Korea). HT-22 cells (2 × 104/ml) were grown in chamber slides (Thermofisher Scientific 75 Panorama Creek Drive Rochester, NY14625–2385, USA) in Dulbecco’s modified Eagle medium (DMEM) (Gibco by life technologies, Grand Island, NY, USA) supplemented with 10% fetal bovine serum (FBS) and 1% antibiotics (penicillin-streptomycin) at 37 °C in humidified air containing 5% CO2. After 70–80% confluence, cells were co-treated with AβO (5 μM) and anthocyanins (100 μg/ml) for 24 h, besides the control group (cells treated with DMSO (0.01%)). After 24 h, cells were washed with PBS, followed by adding 50 μM DCFH-DA in DMSO and incubated at 37 °C in humidified air containing 5% CO2 for 30 min covered by aluminum foil. After this the cells were washed with PBS and fixed in 4% paraformaldehyde. The slides were mounted with DAPI and Prolong Antifade Reagent (Molecular Probe, Eugene, OR, USA) and analyzed by a laser confocal microscopy (Fluoview FV 1000, Olympus, Japan) with excitation and emission wavelengths of 484 and 530 nm, respectively. Fluorescence intensity was measured using image J software and expressed as a relative fluorescence of the samples to control cells.
Lipid Peroxidation (LPO) Analysis in Mouse Hippocampus
The LPO levels determined in the hippocampus (n = 8 mice per group) homogenates through analyzing the malondialdehyde (MDA) level, a biomarker of LPO, by using the commercial lipid peroxidation kit (catalog # K739-100) from Biovision Incorporated, A 95035 USA. The assay was performed according to the provided protocol.
Glutathione (GSH) Analysis in Mouse Hippocampus
The GSH levels in the hippocampus (n = 8 mice per group) homogenates were assessed by using the commercially available glutathione Assay kit (BioVision’s Catalog #K264-100) according to the provided protocol.
Tissue Collection and Sample Preparation from Mouse Brain
For the tissue analysis, the mice (n = 5 mice per group) were perfused transcardially with 4% ice-cold paraformaldehyde, and the brains were post-fixed for 72 h in 4% paraformaldehyde and transferred to 20% sucrose for 72 h. The brains were frozen in O.C.T compound (A.O, USA), and 14-μm coronal sections were cut using a CM 3050C cryostat (Leica, Germany). The sections were thaw-mounted on probe-on plus charged slides (Fisher, Rockford, IL, USA).
Immunofluorescence Staining and Confocal Microscopy
For immunofluorescence staining the slides were washed twice for 10 min in 0.01 M PBS, followed by incubation for 1 h in blocking solution containing 2% normal serum according to the antibody treatment and 0.3% Triton X-100 in PBS. After blocking, the slides were incubated overnight at 4°C in the primary antibodies e.g., p-Akt, HO-1, Nrf-2, SNAP-23, PSD-95,GCLM and 8-Oxoguanine (8-OxoG) (detailed information provided in Table 1) diluted 1:100 in blocking solution. Following incubation with primary antibody, the sections were incubated for 2 h in the secondary tetramethyl rhodamine isothiocyanate (TRITC)/fluorescein isothiocyanate (FITC)-labeled antibodies (1:50) (Santa Cruz Biotechnology). Subsequently, after the incubation of the TRITC/FITC-labeled, sections were incubated overnight in another primary antibody, following incubation with primary antibody, the sections were incubated for 2 h FITC/TRITC-labeled antibody (1:50) (Santa Cruz Biotechnology) under the same conditions. The slides were mounted with DAPI and Prolong Antifade Reagent (Molecular Probe, Eugene, OR, USA). Staining images of the double immunofluorescence and single immunofluorescence were examined using a confocal laser-scanning microscope (Flouview FV 1000, Olympus, Japan). Three to five original immunofluorescence images per sample were captured from each group and the images were converted to tagged image file format (TIF). The immunofluorescence intensity in the same region of hippocampus/total area of TIF images for all groups was measured using imageJ software. By using the ImageJ software, the TIF image background was optimized according to the threshold intensity and the immunofluorescence intensity was analyzed at the same threshold intensity for all groups and were expressed as the relative integrated density of the samples relative to the control which is normalized as one.
Terminal Deoxynucleotidyl Transferase dUTP Nick End Labeling (TUNEL) Immunohistochemical Assay
TUNEL immunohistochemical staining was performed using the DNA Fragmentation Detection Kit (Oncogene Research Products, Cambrige, MA, USA). Briefly, sections were two times washed for 5 min each with PBS. After washing the sections were incubated in equilibration buffer and terminal deoxynucleotidyl transferase (TdT) enzyme for minimum of 60 min at room temperature (R.T). The reaction was terminated by incubation in stop buffer for 5 min at R.T. Sections were labeled with streptavidin HRP conjugate and visualized with DAB substrate. TUNEL-positive cells were analyzed using light microscopy. The number of TUNEL-positive nuclei cells is analyzed and counted through image J program.
Fluoro-Jade B (FJB) Staining
FJB staining was performed according to the provider protocols (Millipore, USA, cat# AG310, lot# 2159662). The slides of the brain tissue were air-dried for at least 4 h. Slides were washed twice for 5 min. Following washing the slides were immersed in a 1% sodium hydroxide and 80% ethanol solution for 5 min and then in 70% alcohol for 2 min followed by 2 min in distilled water. Next, the slides were transferred to a solution of 0.06% potassium permanganate for 10 min over a slow shaker, rinsed with distilled water and then immersed in a 0.1% acetic acid and 0.01% FJB solution for 20 min. The slides were then rinsed with distilled water and dried for at least 10 min. The slides were mounted with PI and glass cover slips using DPX non-fluorescent mounting medium. The representative images were captured using a FITC filter on a confocal laser-scanning microscope (FV 1000, Olympus, Japan). The number of FJB-positive cells is analyzed and counted through image J program.
In Vitro HT22 Cells Culture for Western Blotting
The HT-22 neuronal cells (2 × 104/ml) were cultured (in 35 mm Petri dish (Nunc A/S, Kamstrupvej 90.P.O.Box 280 DK-4000 Rosklide, Denmark) for western blots in DMEM containing 10% FBS and 1% antibiotics at 37 °C in humidified air containing 5% CO2. Nrf2 (SC: 37,049, Santa Cruz Biotechnology, Inc) genes silencing small interference RNA (siRNA) with concentration of 10 μM per transfection for 36 h according to manufacturer protocol. The negative siRNA (Ambion, USA) was used as control. The transfection were performed when culture was 75–80% confluent with lipofectamine™ reagent 2000 (Invitrogen). After 36 h of cells transfection the cells were co-treated with AβO (5 μM), LY294002 (10 μM) and anthocyanins (100 μg/ml) for 24 h. The cell lysates were collected in in pro-prep™ protein extraction solution according to the provider instruction (iNtRON, Biotechnology, Inc). The cell lysates were mixed properly and incubated on ice for at least 30 min to enhance cell lysis. After incubation the cell lysates were centrifuged at 10000×g at 4 °C for 25 min. The supernatants were collected and stored at − 80°C until use for the Western blotting.
In Vitro ROS Assay in HT-22 Cells with Nrf2 siRNA
The HT-22 neuronal cells were cultured in 75 cm2 flasks (Thermo scientific, Nunc™ EasYFlask™ 75 cm2 Nunclon™ Delta surface, thermo fisher scientific A/S, Kamstrupvej 90.P.O.Box 280 DK-4000 Rosklide, Denmark) containing DMEM supplemented with 10% FBS and 1% antibiotics at 37 °C in humidified air containing 5% CO2, and the number of HT22 cells were counted using a disposable hemocytometer as described in the above section. The cells (2 × 104/ml) were further cultured in 96-well plate (Thermofisher Scientific 75 Panorama Creek Drive Rochester, NY14625-2385, USA) containing DMEM supplemented with 10% FBS and 1% antibiotics at 37 °C in humidified air containing 5% CO2. After the cells reached 70% confluence, they were transfected with 10 μM Nrf2 siRNA with lipofectamine™ 2000 reagent for 36 h according to the manufacturer’s protocol. Negative siRNA was used as a control. After 36 h of transfection, the cells were co-treated with AβO (5 μM), LY294002 (10 μM) and anthocyanins (100 μg/ml) for 24 h. After 24 h of treatment, the cells were exposed to 50 μM DCFH-DA and incubated for 30 min. The relative absorbance of the ROS-positive cells treated with DCFH-DA was measured at 484/530 nm with a multidetector (Promega).
In Vitro ApoTox-Glo™ Triplex Assay
ApoTox-Glo™ Triplex assay (Promega Corporation, 2800 Woods Hollow Road Madison, WI53711–5399, USA) was performed to assess viability, cytotoxicity and caspase-3/7 activation using HT22 cells. HT22 cells (2 × 104/ml) were cultured in 96-well plate (Thermofisher Scientific 75 Panorama Creek Drive Rochester, NY14625-2385, USA) containing DMEM supplemented with 10% FBS and 1% antibiotics at 37 °C in humidified air containing 5% CO2. After 70% confluences the cells were treated with AβO (5 μM) and anthocyanins at different three concentrations of 25 μg/ml, 50 μg/ml and 100 μg/ml for 24 h, besides the control group (cells treated with DMSO (0.01%)). In addition the HT22 cells were transfected with Nrf2 siRNA (m) (SC-37049) and treated with AβO (5 μM), LY294002 (10 μM) and anthocyanins 100 μg/ml for 24 h; the control group subjected to negative siRNA (Ambion, ThermoFisher Scientific) and treated with 0.01% DMSO. The assay was performed according to our previous described protocol [39].
Statistics
The Western blot bands were scanned and analyzed through densitometry using the Sigma Gel System (SPSS Inc., Chicago, IL). One-way analysis of variance (ANOVA) followed by a two-tailed independent Student’s t test and Tukey’s multiple comparison test were used for comparisons among the treated groups and the control. The ImageJ software was used for the relative immunofluorescence quantitative analysis. The density values of the data were expressed as the means ± SEM of three independent experiments. Behavioral data for escape latency were analyzed using a two-way ANOVA with training days as repeated measurement. For the probe trials, statistical comparisons among groups for the time over quadrants were done using one-way ANOVA. P values less than 0.05 were considered to be statistically significant. *Significantly different from the WT Veh treated; # significantly different from the Veh treated APP/PS1 mice. Similarly, for in vitro represented by bars. *p < 0.05, **p < 0.01 and ***p < 0.001; and # p < 0.05, ## p < 0.01 and ### p < 0.001.
Results
Anthocyanins Treatment Upregulated p-PI3K/Akt/GSK3β (Ser9) Signaling, Nuclear Translocation of Nrf2, and Expression of its Target Genes (HO-1 and GCLM) in APP/PS1 Mice and In Vitro AβO-Exposed HT22 Cells
Impairment of the p-PI3K/Akt/GSK3β pathway has been associated with various neurodegenerative disorders [7, 8, 13, 15, 30, 39]. Here, our Western blot results showed reduced p-PI3K, p-Akt (Ser473) and p-GSK3β (Ser9) in the Veh-treated APP/PS1 mice compared with the Veh-treated WT mice. Anthocyanins treatment (12 mg/kg i.p. for 30 days) significantly upregulated the expression of p-PI3K, p-Akt and p-GSK3β (Ser9) in the APP/PS1 mice compared with the Veh-treated APP/PS1 mice (Fig. 1a). Immunofluorescence results showed decreased immunofluorescence reactivity of p-Akt (Ser473) in the CA1 region of the hippocampi in Veh-treated APP/PS1 mice compared with the Veh-treated WT mice. Anthocyanins treatment significantly increased p-Akt (Ser473) immunofluorescence reactivity in the CA1 region of the hippocampi in APP/PS1 compared with the Veh-treated APP/PS1 group (Fig. 1b).

Anthocyanins treatment upregulated p-PI3K/Akt/GSK3β (Ser9) signaling, nuclear translocation of Nrf2, and expression of its target genes (HO-1 and GCLM) in APP/PS1 mice and in vitro AβO-exposed HT22 cells. a The Western blot results for p-PI3K/Total-PI3K, p-Akt (Ser473)/Total Akt and p-GSK3β (Ser9)/Total GSK3β in the hippocampus of the experimental mice. The bands were quantified using Sigma Gel software, and the differences are represented by a histogram. β-Actin was used as a loading control. Data are expressed as the means ± SEM for the indicated proteins (n = 8 mice/group), n = 3 experiments. b A representative image of the immunofluorescence reactivity of p-Akt (Ser473) (red, TRITC; blue, DAPI) in the CA1 region of the hippocampus. N = 5 mice/group, n = 3 experiments. Magnification ×10. Scale bar = 100 μm. c The Western blot results of nuclear/cytosolic Nrf2 expression using Nrf2, HO-1 and GCLM in the hippocampus of the experimental mice. The bands were quantified using Sigma Gel software, and the differences are represented by a histogram. β-Actin was used as a loading control. Data are expressed as the means ± SEM for the indicated proteins (n = 8 mice/group). d, e Images of co-localized immunofluorescence reactivity of Nrf2 (green, FITC; blue, DAPI) and HO-1 (red, TRITC; blue, DAPI) in the DG and CA1 regions of the hippocampus. N = 5 mice/group. Magnification ×40. Scale bar = 50 μm. f A representative image of the immunofluorescence reactivity of GCLM (green, FITC; blue, DAPI) in the CA1 and CA3 region of the hippocampus. N = 5 mice/group, n = 3 experiments. Magnification ×10. Scale bar = 100 μm. g The Western blot results for p-PI3K, p-Akt (Ser473), p-GSK3β (Ser9), Nrf2, HO-1 and GCLM in the HT22 cells subjected to Nrf2 siRNA and exposed to anthocyanins (100 μg/ml), AβO (5 μΜ) and LY294002 (10 μM). The bands were quantified using Sigma Gel software, and the differences are represented by a histogram. Data are expressed as the means ± SEM for the indicated proteins. The presented data are relative to the control. Significance = *P < 0.05, **P < 0.01, ***P < 0.001, # P < 0.05, ## P < 0.01, ### P < 0.001
Recently, several studies reported that Nrf2 is a downstream target of the PI3K/Akt/GSK3β pathway [9,10,11, 13]. It has been shown that Nrf2 and its target anti-oxidant genes such as HO-1 have a critical role in the mechanism that mitigates oxidative stress and that they activate an anti-oxidant mechanism that might be beneficial in AD and other neurological disorders [40]. Nuclear translocation of Nrf2 increased the expression of its target genes HO-1 and GCLM [19]. Our Western blot results revealed that anthocyanins supplementation significantly increased the nuclear translocation of Nrf2 and decreased the cytoplasmic expression of Nrf2 in the APP/PS1 mice compared with the Veh-treated APP/PS1 mice (Fig. 1c). Consistently, Western blots results indicated that anthocyanins significantly increased the expression of HO-1 and GCLM in the hippocampi of APP/PS1 mice compared with the Veh-treated APP/PS1 mice (Fig. 1c).
Next, we analyzed Nrf2/HO-1 co-localization through double immunofluorescence staining. Figure 1d, e shows decreased expression of Nrf2/HO-1 immunofluorescence reactivity in the hippocampi of Veh-treated APP/PS1 mice compared with that of Veh-treated WT mice. Anthocyanins supplementation (12 mg/kg i.p. for 30 days) significantly increased the immunofluorescence reactivity of Nrf2/HO-1 in the DG and CA1 regions of the hippocampi in the APP/PS1 mice compared with the Veh-treated APP/PS1 mice (Fig. 1d, e). In addition we analyzed the immunofluorescence reactivity of GCLM by using confocal microscopy. The confocal microscopy results reveal that anthocyanins significantly increased the immunofluorescence reactivity of GCLM in the CA1 and CA3 region of the hippocampi of APP/PS1 mice as compared to the Veh-treated APP/PS1 mice (Fig. 1f).
Further, we sought to determine the underlying neuroprotective anti-oxidant mechanism of anthocyanins, investigating whether anthocyanins regulate the PI3K/Akt/GSK3β pathway and whether they activate the key endogenous anti-oxidant Nrf2 and its target genes. We observed reduced expression of p-PI3K/Akt/GSK3β (Ser 9) as well the Nrf2 and its targets genes HO-1 and GCLM in the AβO (5 μM)-exposed HT22 cells, which was reversed by anthocyanins (100 μg/ml) and significantly increased in its expression. However, anthocyanins were not able to activate and increase the expression of p-PI3K/Akt/GSK3β (Ser 9) and Nrf2/HO-1/GCLM in the AβO-exposed HT22 cells when they were exposed to a PI3K inhibitor (LY294002 (10 μM)) and Nrf2 siRNA (Fig. 1g). These results indicated that anthocyanins regulated PI3K/Akt/GSK3β pathway and activated the endogenous anti-oxidant Nrf2/HO-1 pathway which might be associated with the anti-oxidant neuroprotective activity of anthocyanins.
Anthocyanins Treatment Mitigated ROS and Oxidative Stress in APP/PS1 Mice and In Vitro AβO-Exposed HT22 Cells
Further, to assess the potent anti-oxidant activity of natural anthocyanins in the APP/PS1 mice, we conducted ROS assay, LPO assay and GSH assay. The ROS assay results showed that ROS levels [based on the oxidation of DCFH-DA to DCF significantly increased in the APP/PS1 mice compared with the Veh-treated WT mice. Anthocyanins supplementation (12 mg/kg i.p. for 30 days) overcame ROS activation and significantly reduced DCF levels in the APP/PS1 mice compared with those of Veh-treated APP/PS1 mice (Fig. 2a).

Anthocyanins treatment mitigated ROS and oxidative stress in APP/PS1 mice and in vitro AβO-exposed HT22 cells. a A representative histogram of ROS levels in the hippocampus homogenates of the experimental mice. The graphs were expressed as the means ± SEM (n = 8 mice/group) and the number of experiments = 3. The presented data are relative to the control. b A representative histogram of MDA levels in the hippocampus homogenates of experimental mice. The graphs were expressed as the means ± SEM (n = 8 mice/group) and the number of experiments = 3. The presented data are relative to the control. c A representative histogram of GSH levels in the hippocampus homogenates of the experimental mice. The graphs were expressed as the means ± SEM (n = 8 mice/group) and the number of experiments = 3. The presented data are relative to the control. d Image of immunofluorescence staining of 8-oxoG (red, TRITC; blue, DAPI) in the CA1, DG, and CA3 regions of the hippocampus. N = 5 mice/group. Magnification ×10. Scale bar = 100 μm. e Images of the DCF immunofluorescence intensity (green, FITC; blue, DAPI) in the HT22 cells exposed to AβO (5 μΜ) and anthocyanins (100 μg/ml) for 24 h. N = 3 experiments. Magnification ×10. Scale bar = 100 μm. The presented data are relative to the control. f A representative histogram of the relative absorbance of ROS-positive HT22 cells that were subjected to Nrf2 siRNA and treated with AβO (5 μΜ) and anthocyanins (100 μg/ml) for 24 h. The number of experiments = 3. The data are expressed as the mean ± SEM. The presented data are relative to the control. Significance = *P < 0.05, **P < 0.01, ***P < 0.001, # P < 0.05, ## P < 0.01, ### P < 0.001
An increased level of MDA has been observed in the transgenic mice [41]. Similarly, we also observed an elevated level of MDA in the Veh-treated APP/PS1 mice compared with the Veh-treated WT mice. Anthocyanins supplementation prevents elevated MDA levels and significantly reduced the MDA levels of the APP/PS1 mice compared with those of Veh-treated APP/PS1 mice (Fig. 2b). Next, we analyzed the GSH levels using a commercial GSH kit. The results revealed that anthocyanins significantly increased GSH levels in the brain homogenates of APP/PS1 mice compared with those of the Veh-treated APP/PS1 mice (Fig. 2c).
The 8-OxoG is a key biomarker of ROS-induced oxidative stress, increased in the AD brain and the APPswe/PS1deltaE9 transgenic mouse model of AD [42]. To assess ROS-induced oxidative stress, we analyzed the oxidative stress marker 8-OxoG through immunofluorescence staining. The results showed increased expression of 8-OxoG in the Veh-treated APP/PS1 mice compared with the Veh-treated WT mice. Anthocyanins supplementation overcame oxidative stress and significantly reduced the expression of 8-OxoG in the APP/PS1 mice compared with the Veh-treated APP/PS1 mice (Fig. 2d). Furthermore, we found in vitro that 100 μg/ml anthocyanins significantly reduced the elevated ROS production, as indicated by decreased DCF fluorescence intensity, in the AβO (5 μM)-exposed HT22 cells (Fig. 2e). Additionally, the in vitro ROS assay results manifested that anthocyanins (100 μg/ml) significantly mitigated the ROS-positive cells in the AβO-(5 μM)-exposed HT22 cells. Interestingly, when the HT22 cells were treated with a LY294002 (10 μM) and subjected to Nrf2 siRNA, anthocyanins were unable to prevent the formation and accumulation of ROS-positive cells in the AβO-exposed HT22 cells (Fig. 2f).
Anthocyanins via PI3K/Akt/Nrf2 Pathways Prevented Neurodegeneration in APP/PS1 Mice and In Vitro AβO-Exposed HT22 Cells
Recently, we reported that Korean black bean anthocyanins exert neuroprotective effects against Aβ1–42 in both in vitro and in vivo Aβ1–42 models [36, 37]. Therefore, we assessed the apoptotic and neurodegenerative markers such as activated caspase-3 and activated poly (ADP-ribose) polymerase 1 (PARP-1) expression level; and TUNEL and FJB positive neuronal cells in the APP/PS1 mice. Our western blot analysis indicated increases in activated caspase-3 and PARP-1 in the Veh-treated APP/PS1 mice compared with the Veh-treated WT mice. Anthocyanin (12 mg/kg i.p. for 30 days) supplementation significantly downregulated activated caspase-3 and activated PARP-1 levels in the APP/PS1 mice compared with Veh-treated APP/PS1 (Fig. 3a). The immunohistochemical TUNEL results revealed that TUNEL-positive neuronal cells were significantly increased in the hippocampi of Veh-treated APP/PS1 mice compared with the Veh-treated WT group. Anthocyanins supplementation significantly reduced the number of these TUNEL-positive neuronal cells in the CA3, CA1 and DG regions of hippocampi, showing that anthocyanins reduced neurodegeneration in the APP/PS1 mice compared with the Veh-treated APP/PS1 group (Fig. 3b). Similarly, the number of FJB-positive neuronal cells was significantly increased in the hippocampi of Veh-treated APP/PS1 mice compared with the Veh-treated WT group. Anthocyanins significantly reduced neurodegeneration in the CA1 and CA3 region of the hippocampi, which showed decreased numbers of FJB-positive neurons in APP/PS1 mice compared with the Veh-treated APP/PS1 group (Fig. 3c).

Anthocyanins prevented neurodegeneration in APP/PS1 mice. a Western blot analysis of mouse hippocampi using antibodies against activated caspase-3 and PARP-1. The bands were quantified using Sigma Gel software, and the differences are represented by a histogram. The density values are expressed in arbitrary units (A.U.) as the means ± SEM for the indicated hippocampal proteins (n = 8 mice/group), n = 3 experiments. b The TUNEL-positive neuronal cells are shown (n = 5 animals/group), n = 3 experiments. Magnification ×20. Scale bar = 50 μm. c Representative images of FJB staining (green, FITC; red, PI). (N = 5 animals/group), n = 3 experiments. Magnification ×10. Scale bar = 100 μm. The presented data are relative to the control. Significance = *P < 0.05, **P < 0.01, ***P < 0.001, # P < 0.05, ## P < 0.01, ### P < 0.001
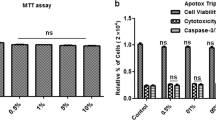
Further, here again we sought to determine in vitro whether anthocyanins ameliorate AβO-induced neurotoxicity. The results of an ApoTox-Glo™ Triplex assay in neuronal HT22 cells showed that cell viability was reduced, whereas the cytotoxicity and activation of caspase3/7 increased, after treatment with AβO (5 μM) compared with the control [cells exposed to DMSO (0.01%)]. Anthocyanins treatment at three different concentrations (25, 50, and 100 μg/ml) reduced the effects of AβO (5 μM); however, the 100 μg/ml dose significantly reduced the effects of AβO (5 μM), increasing cell viability and decreasing cytotoxicity and caspase3/7 activation (Fig. 4a–c). Furthermore, to assess the role of the cytoprotective and anti-oxidant PI3K/Akt/Nrf2 pathways in AβO-induced neurotoxicity, we performed an ApoTox Glo™ Triplex assay in neuronal HT22 cells by using a PI3K inhibitor (LY294002 (10 μM)) and Nrf2 siRNA. Figure 4d–f demonstrates that 100 μg/ml anthocyanins reversed the effect of AβO, significantly increased cell viability and reduced cytotoxicity and caspase3/7 activation in the AβO-exposed HT22 cells. Nevertheless, when a PI3K inhibitor (LY294002 (10 μM)) and Nrf2 siRNA were applied, the anthocyanins were ineffective against AβO-induced neurotoxicity and could not increase cell viability and reduce cytotoxicity and caspase3/7 activation in the HT22 cells (Fig. 4d–f). These in vitro results suggest that anthocyanins act as a potent neuroprotective anti-oxidant and reduced AβO-induced neurotoxicity in a PI3K/Akt/Nrf2-dependent manner.

Anthocyanins via PI3K/Akt/Nrf2 pathway prevented AβO-induced neurotoxicity in HT22 neuronal cells. a–c ApotoxGlo Triplex assays of the cell viability, cytotoxicity and activated caspase3/7 in HT22 cells treated with AβO (5 μM) and anthocyanins at three different concentrations (25, 50, and 100 μg/ml). d–f ApotoxGlo Triplex assays of the cell viability, cytotoxicity and activated caspase3/7 in HT22 cells subjected to Nrf2 siRNA and treated with AβO (5 μM), LY294002 (10 μM) and anthocyanins (100 μg/ml). N = 3 independent experiments. The data are expressed as the means ± SEM. Significance = ***P < 0.001, ### P < 0.001
Anthocyanins Improved Pre- and Postsynaptic Protein Markers in APP/PS1 Transgenic AD Mice
After the antioxidant and neuroprotective activity of anthocyanins in the APP/PS1 mice, we determined the memory-associated protein markers in the APP/PS1 mice. Western blot analysis revealed that memory-associated presynaptic [synaptophysin, synaptosomal associated protein 25 (SNAP-25)] and postsynaptic protein markers [postsynaptic density protein 95 (PSD95) and synaptosomal-associated protein 102 (SAP-102)] were decreased in the hippocampi of Veh-treated APP/PS1 mice compared with the Veh-treated WT mice. Anthocyanins supplementation (12 mg/kg i.p. for 30 days) significantly increased the levels of synaptophysin, SNAP-25, PSD95, and SAP-102 in the hippocampi of APP/PS1 mice (Fig. 5a).

Anthocyanins improved pre- and postsynaptic protein markers in APP/PS1 transgenic mice. a The Western blot results for synaptophysin, SNAP25, PSD95, SAP102, SNAP23, p-GluR1 (Ser845) and p-CREB (Ser133) in the hippocampi of the mice. The bands were quantified using Sigma Gel software, and the differences are represented by a histogram. Data are expressed as the means ± SEM for the indicated proteins (n = 8 mice/group), n = 3 experiments. b Images of co-localized immunofluorescence reactivity of SNAP23 (green, FITC; blue, DAPI) and PSD95 (red, TRITC; blue, DAPI) in the CA1, CA3 and DG regions of the hippocampus. N = 5 mice/group. Magnification ×40. Scale bar = 50 μm. The presented data are relative to the control. Significance = *P < 0.05, **P < 0.01, ***P < 0.001, # P < 0.05, ## P < 0.01, ### P < 0.001
Aβ-induced synaptic impairment is associated with the reduction of α-amino-3-hydroxy-5-methylisoxazole-4-propionic acid (AMPA) receptors (AMPARs). Notably, phosphorylation of the AMPA receptor (GluR1) at Ser845 plays a pivotal role in the trafficking of postsynaptic glutamate receptors and is related to spatial working memory [15, 37, 39]. Our immunoblot results showed decreased levels of these post-synaptic markers in the Veh-treated APP/PS1 mice compared with the Veh-treated WT mice. Anthocyanin treatment significantly increased SNAP23 and p-GluR1 (Ser845) in the APP/PS1 mice compared with the Veh-treated APP/PS1 mice (Fig. 5a).
It has been reported that activating phosphorylated cAMP response element binding protein (p-CREB) (Ser133) in the rodent’s brain improved spatial working memory [37, 43, 44]. Our black bean anthocyanins supplementation activated p-CREB (Ser133) and significantly increased the level of p-CREB (Ser133) in the APP/PS1 mice compared with the Veh-treated APP/PS1 mice (Fig. 5a).
The immunofluorescence results of PSD95 and SNAP23 revealed that anthocyanins treatment enhanced the immunofluorescence reactivity of PSD95 and SNAP23 in the CA1, CA3 and DG region of hippocampi in the APP/PS1 mice compared with the Veh-treated APP/PS1 mice (Fig. 5b).
Anthocyanins Improved Memory Functions in APP/PS1 Transgenic AD Mice
The beneficial effects of polyphenolic flavonoid anthocyanins (12 mg/kg i.p. for 30 days) on spatial learning and memory in APP/PS1 mice were investigated using the MWM and Y-maze tasks. MWM training tests were performed for 6 consecutive days, and the latency to escape to the hidden platform and the trajectories to reach the hidden platform were recorded. As shown in Fig. 6a, the mean escape latency was significantly increased in the APP/PS1 mice compared with the WT Veh-treated mice. Anthocyanin (12 mg/kg i.p. for 30 days) treatment significantly improved memory functions and reduced the mean escape latency compared with the Veh-treated APP/PS1 mice. Moreover, we measured the final escape latency on day seven before the probe test. Regarding the final latency, the Veh-treated WT, anthocyanin-treated APP/PS1 and anthocyanin-treated WT mice took less time than the Veh-treated APP/PS1 mice to reach the hidden platform (Fig.6b). We also observed the mean swimming speeds during the training days and on day 7 before the probe test. The swimming speed measurements revealed no difference among the Veh-treated WT, anthocyanin-treated WT, Veh-treated APP/PS1 and anthocyanin-treated APP/PS1 mice, indicating that any differences in ability to reach the hidden platform were unrelated to the motor functions of the mice (Fig. 6c, d).

Anthocyanins improved memory functions in APP/PS1 transgenic mice. The MWM and the Y-maze test were used to evaluate spatial working memory. Thirteen mice per group were used for the behavioral analysis. a Mean escape latency (sec) to the hidden platform during training. b Final escape latency on the final day of the probe test. c, d Histograms represent swimming speed (cm/s) of the mice during their search for the hidden platform during the training sessions and on day 7 before the probe test, respectively. To calculate swimming speed (cm/s), we used the equation v = distance/time. No significant difference was observed among the swimming speeds of the mouse groups. e Number of crossings over the previous platform location during the probe test. f Time spent in the target quadrant (where the platform was located during the hidden platform training session) during the probe test. g The spontaneous alteration behavior percentage. The graphs express the means ± SEM (n = 13 mice/group). Significance = *P < 0.05, **P < 0.01, ***P < 0.001, # P < 0.05, ## P < 0.01, ### P < 0.001. ns = no significant
On day 7, after the escape latency test, we removed the hidden platform and conducted a probe test. The number of crossings over the previous site of the platform was significantly greater in the anthocyanins-treated APP/PS1 mice than in the Veh-treated APP/PS1 mice (Fig. 1e). Additionally, anthocyanin-treated APP/PS1 mice spent more time in the target quadrant than Veh-treated APP/PS1 mice (Fig. 6f), demonstrating that anthocyanins protected against spatial memory deficits in the APP/PS1 mice.
Next, we performed a Y-maze task to evaluate spatial working memory using spontaneous alternation behavior percentage. The observation showed that the APP/PS1 mice had a lower percentage of alternation than the Veh-treated WT mice, indicating weaker working memory. Anthocyanins treatment significantly increased spontaneous alteration percentage in the APP/PS1 mice compared with the Veh-treated APP/PS1 mice (Fig. 6g), indicating that anthocyanins alleviated short-term memory dysfunction in APP/PS1 AD mice.
Discussion
The objective of the current study was to investigate the underlying anti-oxidant neuroprotective mechanism of dietary Korean black bean anthocyanins in the APP/PS1 mouse model of AD. Several well-established studies have investigated elevated ROS and oxidative stress in this AD model and it has been shown that in the AD brain, Aβ accumulation induces increased ROS production and oxidative stress, which subsequently leads to neurodegeneration and memory impairment [45,46,47,48]. Attenuation of ROS and oxidative stress reduced the memory impairment in the transgenic mouse model of AD [48]. Similarly, the present study noted elevated ROS, enhanced LPO, as determined by increased MDA levels, reduced GSH level and activated immunofluorescence reactivity of 8-OxoG in the APP/PS1 mouse model of AD. Interestingly, anthocyanins supplementation attenuated the elevated ROS levels and oxidative stress in the AD mouse model.
Studies have reported translocation of Nrf2 from the cytosol to the nucleus, where it activates anti-oxidative enzymes, such as HO-1 and NAPDH quinine dehydrogenase-1 (NQO1), and produces a notable anti-oxidative response to counteract escalated ROS-induced oxidative stress and protect against oxidative stress in different neurodegenerative models such as ischemia. Moreover, it has been shown that Nrf2 knockout mice possess elevated oxidative stress and ROS levels [49,50,51,52]. Recently, Zhang et al. found that activation of Nrf2 and its target anti-oxidant genes is protective against oxidative stress [53]. Both in vitro and in vivo numerous studies have proposed that endogenous anti-oxidant genes such as Nrf2 and HO-1 activation are possibly involved in neuroprotective mechanisms against AD [54, 55]. Notably, it has also been reported that the Nrf2/HO-1 pathway was dysregulated in an AD transgenic mouse model and that overexpression of these Nrf2/HO-1 pathways improves spatial learning in an AD mouse model [19, 40, 41]. Consistently, in our AD model (APP/PS1 mice), the expression levels of Nrf2 and HO-1 were decreased. Both in vitro and in vivo, we confirmed that anthocyanin treatment reduced the ROS-induced elevation of oxidative stress via the activation of Nrf2 and its target genes such as HO-1 and GCLM, which might be associated with improved spatial working memory in APP/PS1 mice.
PI3K/Akt/GSK-3β signaling is one of the most important pathways, having a key and pivotal role in various signaling and in other functions. Impaired PI3K/Akt/GSK-3β signaling has been investigated in the AD brain and AD mouse models [7, 8, 13,14,15, 39]. Accumulation of ROS induces oxidative stress that mediates the downregulation of p-PI3K/p-Akt, which triggers the activation of GSK-3β, resulting in Nrf2 translocation from the nucleus to cytosol and producing a low level anti-oxidant activity [56, 57]. ROS accumulation induces oxidative stress that mediates GSK-3β activation, while GSK-3β inhibition is implicated in the attenuation of oxidative stress in hippocampal cell lines [58]. It has been reported that activated GSK-3β induces the impairment and degradation of Nrf2 [59, 60]. Moreover, activation of GSK-3β is prevented by activation of p-GSK-3β (Ser9) via the activation of PI3K/Akt [15]. Kitagishi et al. reviewed the evidence that dietary supplementation of some natural pleiotropic neuroprotective agents, especially anti-oxidants, provides neuroprotection by regulating the PI3K/Akt/GSK-3β pathway [61]. Other studies also reported that nuclear translocation and activation of Nrf2 via upregulation of p-PI3K/p-Akt/GSK3β pathway provides protection against oxidative stress-induced cell injury [61, 62]. Recently, we found that natural anti-oxidant anthocyanins induced neuroprotection against ethanol in the developing rat brain by attenuating oxidative stress through regulation of the PI3K/Akt/GSK-3β pathway [27]. Here, our results also described and elaborated on the anti-oxidant protective mechanism in which chronic anthocyanins treatment regulates the PI3K/Akt/GSK3β signaling cascade and activated the Nrf2/HO-1 pathway and its target anti-oxidant genes, ultimately providing effective anti-oxidant neuroprotection in APP/PS1 mice.
Phenolic and polyphenolic flavonoids act as neuroprotective agents via activation of the PI3K/Akt/GSK3β/Nrf2 pathway, which suppresses the apoptotic caspases and neurodegeneration [63,64,65,66,67,68]. Previously, we demonstrated that anthocyanins prevents the apoptotic neurodegeneration both in vivo and in vitro studies [69]. Similarly, our Western blot results indicated that anthocyanins administration prevented apoptosis via the suppression of apoptotic markers, e.g., activated caspase-3 and PARP-1 levels, in the hippocampi of APP/PS1 mice. Moreover, the increased numbers of TUNEL- and FJB-positive neuronal cells in the hippocampi of APP/PS1 mice were reduced by anthocyanins treatment, which indicated that polyphenolic flavonoids such as anthocyanins may prevent neurodegeneration via activation of the endogenous Nrf2/HO-1 anti-oxidant mechanism. Additionally, it has also been determined that anthocyanins dose-dependently (25, 50,100 μg/ml) prevented Aβ1–42-induced neurotoxicity in vitro in HT22 cells. Furthermore, we exposed HT22 cells to LY294002 (10 μM) and knocked down Nrf2 with Nrf2 siRNA in the presence of Aβ1–42-induced neurotoxicity, against which anthocyanins (100 μg/ml) were ineffective. This provided further evidence that activation of the endogenous neuroprotective anti-oxidant PI3K/Akt/Nrf2 pathway by anthocyanins has a promising role in the prevention of apoptosis and neurodegeneration in the AD.
In AD synaptic deficit and loss is implicated in the memory and cognitive dysfunction and has been considered a primary target in the AD therapy [70]. Synaptic impairment is associated with the decreased expression of presynaptic proteins such as synaptophysin, and the postsynaptic proteins PSD95 and SAP102 were also decreased in the brains of AD patients and animal models of AD [71]. An Aβ-induced deficit of AMPARs (Ser845) has been associated with early synapse and memory dysfunction [15]. P-CREB (Ser133) is an important biochemical marker for memory functions. High expression of p-CREB in the hippocampus increases memory functions and promotes synapse formation. Hippocampus-dependent memory formation is impaired through attenuation of p-CREB [72]. In agreement with previous reported studies, the present study observed a decreased level of pre- (synaptophysin) and postsynaptic proteins (PSD95, SAP102, SNAP23, p-GluR1 at Ser845) as well as p-CREB (Ser133) in the APP/PS1 mouse model of AD. Interestingly, chronic anthocyanins treatment produces beneficial effects and significantly restores these memory-related pre-and postsynaptic markers as well the expression of p-CREB (Ser133) in APP/PS1 mice.
Several studies reported that natural dietary supplements, e.g., phenolic and polyphenolic flavonoids have potent anti-oxidant neuroprotective activities which induce beneficial effects and have been reported to improve learning/memory and cognitive functions in the rodent animal models [63,64,65,66,67,68,69, 73,74,75]. The APP/PS1 mouse is a well-known animal model of memory deficits in the animal AD model [70, 76]. Mounting evidences indicated that PI3K/Akt/GSK3β/Nrf2/HO-1 pathways have a key role in the behavioral performance as well in the learning and memory function of the rodents [75, 77, 78]. Here, we investigated memory impairment via the MWM and Y-maze tasks in 10-month-old male APP/PS1 mice. Notably, in our study, anthocyanins treatment at 12 mg/kg/day i.p. for 30 days improves learning and memory functions as measured by escape latency in the trial session as well as on the final day of the probe test. No significant differences were recorded in the swimming speed among the groups, indicating that spatial learning and memory functions were not associated with motor function. Anthocyanins also improved the number of platform crossings and the time spent in the target quadrant during the probe test on the final day. Anthocyanins also rescued the spatial working memory deficit, as indicated by increasing spontaneous alteration behavior (%) in the APP/PS1 mice compared with Veh-treated APP/PS1 mice (Fig. 6).
In summary, this study demonstrated the underlying anti-oxidant neuroprotective mechanism of natural dietary Korean black bean anthocyanins in the APP/PS1 mouse model of AD. Our results established that anthocyanins regulate the PI3K/Akt/GSK3 pathway and consequently activate the downstream endogenous anti-oxidant Nrf2/HO-1 pathway and its target genes, reducing the AβO-induced elevation of ROS-mediated oxidative stress and preventing neurodegeneration via a PI3K/Akt/Nrf2-dependent pathway. Importantly, anthocyanins improve memory-related pre- and postsynaptic markers as well cognitive functions in APP/PS1 mice. Taken together, these data suggest that anthocyanins derived from natural sources act as potent anti-oxidant; are a promising potential dietary component and could be beneficial as a dietary supplement for the prevention of age-related neurological disorders, such as AD.
References
Roychaudhuri R, Yang M, Hoshi MM, Teplow DB (2009) Amyloid β-protein assembly and Alzheimer disease. J Biol Chem 284(8):4749–4753. https://doi.org/10.1074/jbc.R800036200
Hardy J, Selkoe DJ (2002) The amyloid hypothesis of Alzheimer’s disease: progress and problems on the road to therapeutics. Science 297(5580):353–356. https://doi.org/10.1126/science.1072994
De Felice FG, Velasco TP, Lambert MP, Viola K, Fernandez SJ, Ferreira ST, Kleivin WL (2007) Abeta oligomers induce neuronal oxidative stress through an N-methyl-D-aspartate receptor-dependent mechanism that is blocked by the Alzheimer drug memantine. J Biol Chem 282(15):11590–11601. https://doi.org/10.1074/jbc.M607483200
Lambert MP, Barlow AK, Chromy BA, Edwards C, Freed R, Liosatos M, Morgan TE, Rozovsky I et al (1998) Diffusible, nonfibrillar ligands derived from Abeta1-42 are potent central nervous system neurotoxins. Proc Natl Acad Sci U S A 282(11):33305–33312
Perluigi M, Sultana R, Cenini G, Di Domenico F, Memo M, Pierce WM, Coccia R, Butterfield DA (2009) Redox proteomics identification of 4-hydroxynonenal-modified brain proteins in Alzheimer’s disease: role of lipid peroxidation in Alzheimer’s disease pathogenesis. Proteomics Clin Appl 3(6):682–693. https://doi.org/10.1002/prca.200800161
Niedzielska E, Smaga I, Gawlik M, Moniczewski A, Stankowicz P, Pera J, Filip M (2016) Oxidative stress in neurodegenerative diseases. Mol Neurobiol 53(6):4094–4125. https://doi.org/10.1007/s12035-015-9337-5
Singh AK, Kashyap MP, Tripathi VK, Singh S, Garg G, Rizvi SI (2016) Neuroprotection through rapamycin-induced activation of autophagy and PI3K/Akt1/mTOR/CREB signaling against amyloid-β-induced oxidative stress, synaptic/neurotransmission dysfunction, and neurodegeneration in adult rats. Mol Neurobiol. https://doi.org/10.1007/S12035-016-0129-3
Luo J (2009) GSK3β in ethanol neurotoxicity. Mol Neurobiol 40(2):108–121. https://doi.org/10.1007/s12035-8075-y
Nakaso K, Yano H, Fukuhara Y, Takeshima T, Wada- Isoe K, Nakashima K (2003) PI3K is a key molecule in the Nrf2- mediated regulation of antioxidative proteins by hemin in human neuroblastoma cells. FEBS Lett 546(2–3):181–184
Wang L, Chen Y, Sternberg P, Cai J (2008) Essential roles of the PI3 kinase/Akt pathway in regulating Nrf2-dependent antioxidant functions in the RPE. Invest Ophthalmol Vis Sci 49(4):1671–1678. https://doi.org/10.1167/iovs.07-1099
Martin D, Rojo AI, Salinas M, Diaz R, Gallardo G, Alam J, DeGalarreta CM, Cuadrado A (2004) Regulation of heme oxygenase-1 expression through the phosphatidylinositol 3-kinase/Akt pathway and the Nrf2 transcription factor in response to the antioxidant phytochemical carnosol. J Biol Chem 279(10):8919–8929. https://doi.org/10.1074/jbc.M309660200
Surh YJ, Kundu JK, Na HK (2008) Nrf2 as a master redox switch in turning on the cellular signaling involved in the induction of cytoprotective genes by some chemopreventive phytochemicals. Planta Med 74(13):1526–1539. https://doi.org/10.1055/s-0028-1088302
Le Belle JE, Orozco NM, Paucar AA, Saxe JP, Mottahedeh J, Pyle AD, Wu H, Kornblum HI (2011) Proliferative neural stem cells have high endogenous ROS levels that regulate self-renewal and neurogenesis in a PI3K/Akt-dependant manner. Cell Stem Cell 8(1):59–71. https://doi.org/10.1016/j.stem.2010.11.028
Erdogdu O, Nathanson D, Sjoholm A, Nyström T, Zhang Q (2010) Exendin-4 stimulates proliferation of human coronary artery endothelial cells through eNOS-, PKA- and PI3K/Aktdependent pathways and requires GLP-1 receptor. Mol Cell Endocrinol 325(1–2):26–35. https://doi.org/10.1016/j.mce.2010.04.022
Ali T, Kim MO (2015) Melatonin ameliorates amyloid beta-induced memory deficits, tau hyperphosphorylation and neurodegeneration via PI3/Akt/GSK3β pathway in the mouse hippocampus. J Pineal Res 59(1):47–59. https://doi.org/10.1111/jpi.12238
Mhilaj E, Catino S, Miceli FM, Santangelo R, Trabace L, Cuomo V, Mancuso C (2017) Ferulic acid improves cognitive skills through the activation of the heme oxygenase system in the rat. Mol Neurobiol. https://doi.org/10.1007/s12035-017-0381-1
Zou Y, Hong B, Fan L, Zhou L, Liu Y, Wu Q, Zhang X, Dong M (2013) Protective effect of puerarin against beta-amyloid induced oxidative stress in neuronal cultures from rat hippocampus: involvement of the GSK-3β/Nrf2 signaling pathway. Free Radic Res 47(1):55–63. https://doi.org/10.3109/10715762.2012.743518
Kanninen K, Malm TM, Jyrkkänen HK, Goldsteins G, Keksa-Goldsteine V, Tanila H, Yamamoto M, Yia-Herttuala S et al (2008) Nuclear factor erythroid 2-related factor 2 protects against beta amyloid. Mol Cell Neurosci 39(3):302–313. https://doi.org/10.1016/j.mcn.2008.07.010
Kanninen K, Heikkinen R, Malm T, Rolova T, Kuhmonen S, Leinonen H, Yia-Herttuala S, Tanila H et al (2009) Intrahippocampal injection of a lentiviral vector expressing Nrf2 improves spatial learning in a mouse model of Alzheimer’s disease. Proc Natl Acad Sci U S A 106(38):16505–16510. https://doi.org/10.1073/pnas.0908397106
XL B, Rao PPN, Wang YJ (2016) Anti-amyloid aggregation activity of natural compounds: Implications for Alzheimer’s drug discovery. Mol Neurobiol 53(6):3565–3575. https://doi.org/10.1007/s12035-015-9301-4
Virman A, Pinto L, Binienda Z, Ali S (2013) Food, nutrigenomics, and neurodegeneration-neuroprotection by what you eat! Mol Neurobiol 48(2):353–362. https://doi.org/10.1007/s12035-013-8498-3
Shih PH, Yeh CT, Yen GC (2007) Anthocyanins induce the activation of phase ii enzymes through the antioxidant response element pathway against oxidative stress-induced apoptosis. J Agric Food Chem 55(23):9427–9435. https://doi.org/10.1021/jf071933i
Lan X, Wang W, Li Q, Wang J (2016) The natural flavonoid pinocembrin: molecular targets and potential therapeutic applications. Mol Neurobiol 53(3):1794–1801. https://doi.org/10.1007/s12035-015-9125-2
Schaffer S, Asseburg H, Kuntz S, Muller WE, Eckert GP (2012) Effects of polyphenols on brain ageing and Alzheimer’s disease: focus on mitochondria. Mol Neurobiol 46(1):161–178. https://doi.org/10.1007/s12035-012-8282-9
Lakey-Beitia J, Berrocal R, Rao KS, Durant AA (2015) Polyphenols as therapeutic molecules in Alzheimer’s disease through modulating amyloid pathways. Mol Neurobiol 51(2):466–479. https://doi.org/10.1007/s12035-014-8722-9
Ahmad A, Ali T, Park HY, Badshah H, Rehman SU, Kim MO (2016) Neuroprotective effect of fisetin against amyloid beta-induced cognitive/synaptic dysfunction, neuroinflammation and neurodegeneration in adult mice. Mol Neurobiol 54(3):2269–2285. https://doi.org/10.1007/s12035-016-9795-4
Shah SA, Yoon GH, Kim MO (2015) Protection of the developing brain with anthocyanins against ethanol-induced oxidative stress and neurodegeneration. Mol Neurobiol 51(3):1278–1291. https://doi.org/10.1007/s12035-014-8805-7
Rehman SU, Shah SA, Ali T, Chung JI, Kim MO (2016) Anthocyanins reversed D-galactose-induced oxidative stress and neuroinflammation mediated cognitive impairment in adult rats. Mol Neurobiol 54(1):255–271. https://doi.org/10.1007/s12035-015-9604-5
Ullah I, Park HY, Kim MO (2013) Anthocyanins protect against kainic acid-induced excitotoxicity and apoptosis via ROS-activated AMPK pathway in hippocampal neurons. CNS Neurosci Ther 20(4):327–338. https://doi.org/10.1111/cns.12218.
Shah SA, Ullah I, Lee HY, Kim MO (2013) Anthocyanins protect against ethanol-induced neuronal apoptosis via GABAB1 receptors intracellular signaling in prenatal rat hippocampal neurons. Mol Neurobiol 48(1):257–269. https://doi.org/10.1007/s12035-013-8458-y
Badshah H, Ali T, Ahmad A, Kim MJ, Abid NB, Shah SA, Yoon GH, Lee HY (2015) Co-treatment with anthocyanins and vitamin C ameliorates ethanol- induced neurodegeneration via modulation of GABAB receptor signaling in the adult rat brain. CNS Neurol Disord Drug Targets 14(6):791–803
Ye J, Meng X, Yan C, Wang C (2010) Effects of purple sweet potato anthocyanins on β-amyloid-mediated PC-12 cells death by inhibition of oxidative stress. Neurochem Res 35 (3):357–365. https://doi.org/10.1007/s11064-009-0063-0
Kang TH, Hur JY, Kim HB, Ryu JH, Kim SY (2006) Neuroprotective effects of the cyanidin-3-O-beta-d-glucopyranoside isolated from mulberry fruit against cerebral ischemia. Neurosci Lett 391(3):122–126. https://doi.org/10.1016/j.neulet.2005.08.053
Catarina R, Vauzour D, Rattray M, Waffo-Teguo P, Merillon JM, Butler LT, Williams CM, Spencer JP (2012) Dietary levels of pure flavonoids improve spatial memory performance and increase hippocampal brain-derived neurotrophic factor. PLoS One 8:e63535. https://doi.org/10.1371/journal.pone.0063535
Carvalho FB, Gutierres JM, Bueno A, Agostinho P, Zago AM, Vieira J, Fruhauf P, Cechella JL et al (2017) Anthocyanins control neuroinflammation and consequent memory dysfunction in mice exposed to lipopolysaccharide. Mol Neurobiol 54(5):3350–3367. https://doi.org/10.1007/s12035-016-9900-8
Badshah H, Kim TH, Kim MO (2015) Protective effects of anthocyanins against amyloid beta-induced neurotoxicity in vivo and in vitro. Neurochem Int 80:51–59. https://doi.org/10.1016/j.neuint.2014.10.009
Ali T, Kim MJ, Rehman SU, Ahmad A, Kim MO (2016) Anthocyanin-loaded PEG-gold nanoparticles enhanced the neuroprotection of anthocyanins in an Aβ1–42 mouse model of Alzheimer’s disease. Mol Neurobiol. https://doi.org/10.1007/s12035-016-0136-4
Ali T, Badshah H, Kim T, Kim MO (2015) Melatonin attenuates D-galactose-induced memory impairment, neuroinflammation and neurodegeneration via RAGE/NF-KB/JNK signaling pathway in aging mouse model. J Pineal Res 58(1):71–85. https://doi.org/10.1111/jpi.12194
Ali T, Yoon GH, Shah SA, Lee HY, Kim MO (2015) Osmotin attenuates amyloid beta-induced memory impairment, tau phosphorylation and neurodegeneration in the mouse hippocampus. Sci Rep 5:11708. https://doi.org/10.1038/srep11708
Li L, Li W, Jung SW, Lee YW, Kim YH (2011) Protective effects of decursin and decursinol angelate against amyloid β-protein-induced oxidative stress in the PC12 cell line: The role of Nrf2 and antioxidant enzymes. Biosci Biotechnol Biochem 75(3):434–442
Zhou Y, Xie N, Li L, Zou Y, Zhang X, Dong M (2014) Puerarin alleviates cognitive impairment and oxidative stress in APP/PS1 transgenic mice. Int J Neuropsychopharmacol 17(4):635–644. https://doi.org/10.1017/s146114571300148x
Hamilton A, Holscher C (2012) The effect of ageing on neurogenesis and oxidative stress in the APPswe/PS1deltaE9 mouse model of Alzheimer’s disease. Brain Res 1449:83–93. https://doi.org/10.1016/j.brainres.2012.02.015
Yan J, Lai B, Xu A, Liu Y, Li X, Zhao Y, Li W, Ji M et al (2015) Maged1 co-interacting with CREB through a hexapeptide repeat domain regulates learning and memory in mice. Mol Neurobiol 51(1):8–18. https://doi.org/10.1007/s12035-014-8677-x
Gao H, Yan P, Zhang S, Huang H, Huang F, Sun T, Deng Q, Huang Q et al (2016) Long-term dietary alpha-linolenic acid supplement alleviates cognitive impairment correlate with activating hippocampal CREB signaling in natural aging rats. Mol Neurobiol 53(7):4772–4786. https://doi.org/10.1007/s12035-015-9393-x
Sultana R, Butterfield DA (2013) Oxidative modification of brain proteins in Alzheimer’s disease: perspective on future studies based on results of redox proteomics studies. J Alzheimers Dis 33(1):243–251. https://doi.org/10.3233/JAD-2012-129018
Wan L, Nie G, Zhang J, Luo Y, Zhang P, Zhang Z, Zhao B (2011) β-Amyloid peptide increases levels of iron content and oxidative stress in human cell and Caenorhabditis elegans models of Alzheimer disease. Free Radic Biol Med 50(1):122–129. https://doi.org/10.1016/j.freeradbiomed.2010.10.707
Lovell M, Markesbery W (2007) Oxidative DNA damage in mild cognitive impairment and late-stage Alzheimer’s disease. Nucleic Acids Res 35(22):7497–7504. https://doi.org/10.1093/nar/gkm821
Lee JY, Choi SI, Park HM, Lee YM, Song KJ, Kim YH, Kim KH, Hwang DY et al (2011) 4-O-Methylhonokiol attenuates memory impairment in presenilin 2 mutant mice through reduction of oxidative damage and inactivation of astrocytes and the ERK pathway. Free Radic Biol Med 50(1):66–77. https://doi.org/10.1016/j.freeradbiomed.2010.10.698
Ding Y, Chen M, Wang M, Li Y, Wen A (2015) Posttreatment with 11-Keto-β-boswellic acid ameliorates cerebral ischemia–reperfusion injury: Nrf2/HO-1 pathway as a potential mechanism. Mol Neurobiol 52(3):1430–1439. https://doi.org/10.1007/S12035-014-8929-9
Jiang S, Deng C, Lv J, Fan C, Hu W, Di S, Yan X, Ma Z et al (2017) Nrf2 weaves an elaborate network of neuroprotection against stroke. Mol Neurobiol 54(2):1440–1455. https://doi.org/10.1007/s12035-016-9707-7
Zhang R, Xu M, Wang Y, Xie F, Zhang G, Qin X (2016) Nrf2-a promising therapeutic target for defensing against oxidative stress in stroke. Mol Neurobiol. https://doi.org/10.1007/s12035-016-0111-0
Freitas AE, Egea J, Buendia I, Navarro E, Rada P, Cuadrado A, Rodrigues AL, Lopez MG (2015) Agmatine induces Nrf2 and protects against corticosterone effects in hippocampal neuronal cell line. Mol Neurobiol 51:1504–1519. https://doi.org/10.1007/s12035-014-8827-1
Zhang H, Liu YY, Jiang Q, Li K, Zhao Y (2014) Salvianolic acid a protects RPE cells against oxidative stress through activation of Nrf2/HO-1 signaling. Free Radic Biol Med 69:219–228. https://doi.org/10.1016/j.freeradbiomed.2014.01.025
Kamalvand G, Pinard G, Ali-Khan Z (2003) Heme-oxygenase-1 response, a marker of oxidative stress, in a mouse model of AA amyloidosis. Amyloid 10(3):151–159
Godoy JA, Lindsay CB, Quintanilla RA, Carvajal FJ, Cerpa W, Inestrosa NC (2016) Quercetin exerts differential neuroprotective effects against H2O2 and Aβ aggregates in hippocampal neurons: the role of mitochondria. Mol Neurobiol. https://doi.org/10.1007/s12035-016-0203-x
Rojo AI, Rada P, Egea J, Rosa AO, Lopez MG, Cuadrado A (2008) Functional interference between glycogen synthase kinase-3 beta and the transcription factor Nrf2 in protection against kainate-induced hippocampal cell death. Mol Cell Neurosci 39(1):125–132. https://doi.org/10.1016/j.mcn.2008.06.007
Rojo AI, Sagarra MR, Cuadrado A (2008) GSK-3beta downregulates the transcription factor Nrf2 after oxidant damage: relevance to exposure of neuronal cells to oxidative stress. J Neurochem 105(1):192–202. https://doi.org/10.1111/j.1471-4159.2007.05124.x
Schafer M, Goodenough S, Moosmann B, Behl C (2004) Inhibition of glycogen synthase kinase 3 beta is involved in the resistance to oxidative stress in neuronal HT22 cells. Brain Res 1005(1–2):84–89. https://doi.org/10.1016/j.brainres.2004.01.037
Rada P, Al R, Evrard-Todeschi N, Innamorato NG, Cotte A, Jaworski T, Tobon-Velasco JC, Devijver H et al (2012) Structural and functional characterization of Nrf2 degradation by the glycogen synthase kinase 3/β-TrCP axis. Mol Cell Biol 32(17):3486–3499. https://doi.org/10.1128/MCB.00180-12
HC S, Ma CT, BC Y, Chien YC, Tsai CC, Huang WC, Lin CF, Chuang YH et al (2012) Glycogen synthase kinase-3β regulates anti-inflammatory property of fluoxetine. Int Immunopharmacol 14(2):150–156. https://doi.org/10.1016/j.intimp.2012.06.015
Kitagishi Y, Nakanishi A, Ogura Y, Matsuda S (2014) Dietary regulation of PI3K/AKT/GSK-3β pathway in Alzheimer’s disease. Alzheimers Res Ther 6(3). https://doi.org/10.1186/alzrt265
Chen HH, Chen YT, Huang YW, Tsai HJ, Kuo CC (2012) 4-Ketopinoresinol, a novel naturally occurring ARE activator, induces the Nrf2/HO-1 axis and protects against oxidative stress-induced cell injury via activation of PI3K/AKT signaling. Free Radic Biol Med 52(6):1054–1066. https://doi.org/10.1016/j.freeradbiomed.2011.12.012
Lou H, Jing X, Wei X, Shi H, Ren D, Zhang X (2014) Naringenin protects against 6-OHDA-induced neurotoxicity via activation of the Nrf2/ARE signaling pathway. Neuropharmacology 79:380–388. https://doi.org/10.1016/j.neuropharm.2013.11.026
Xu J, Wang H, Ding K, Zhang L, Wang C, Li T, Wei W, Lu X (2014) Luteolin provides neuroprotection in models of traumatic brain injury via the Nrf2–ARE pathway. Free Radic Biol Med 71:186–195. https://doi.org/10.1016/j.freeradbiomed.2014.03.009
Zhao B (2005) Natural antioxidants for neurodegenerative diseases. Mol Neurobiol 31(1–3):283–293. https://doi.org/10.1385/MN:31:1-3:283
Prakash D, Sudhandiran G (2015) Dietary flavonoid fisetin regulates aluminium chloride induced neuronal apoptosis in cortex and hippocampus of mice brain. J Nutr Biochem 26(12):1527–1539. https://doi.org/10.1016/j.jnutbio.2015.07.017.
Gopinath K, Prakash D, Sudhandiran G (2011) Neuroprotective effect of naringin, a dietary flavonoid against 3-nitropropionic acid-inducedneuronal apoptosis. Neurochem Int 59(7):1066–1073. https://doi.org/10.1016/j.neuint.2011.08.022
Lan X, Han X, Li Q, Wang J (2016) (−)-Epicatechin, a natural flavonoid compound, protects astrocytes against hemoglobin toxicity via Nrf2 and AP-1 signaling pathways. Mol Neurobiol. https://doi.org/10.1007/s12035-016-027-y
Khan MS, Ali T, Kim MW, Jo MH, Jo MG, Badshah H, Kim MO (2016) Anthocyanins protect against LPS-induced oxidative stress-mediated neuroinflammation and neurodegeneration in the adult mouse cortex. Neurochem Int 100:1–10. https://doi.org/10.1016/j.neuint.2016.08.005.
Nistico R, Pignatelli M, Piccinin S, Mercuri NB, Collingridge G (2012) Targeting synaptic dysfunction in Alzheimer’s disease therapy. Mol Neurobiol 46(3):572–587. https://doi.org/10.1007/s12035-012-8324-3
Proctor DT, Coulson EJ, Dodd PR (2010) Reduction in post-synaptic scaffolding PSD-95 and SAP-102 protein in the Alzheimer inferior temporal cortex is correlated with disease pathology. J Alzheimers Dis 21(3):795–811. https://doi.org/10.3233/JAD-2010-100090
Restivo L, Tafi E, Ammassari-Teule M, Marie H (2009) Viral mediated expression of a constitutively active form of CREB in hippocampal neurons increases memory. Hippocampus 19(3):228–234. https://doi.org/10.1002/hipo.20527
Prakash D, Gopinath K, Sudhandiran G (2013) Fisetin enhances behavioral performances and attenuates reactive gliosis and inflammation during aluminum chloride-induced neurotoxicity. NeuroMolecular Med 15(1):192–208. https://doi.org/10.1007/s12017-8210-1.
Scapagnini G, Vasto S, Abraham NG, Caruso C, Zella D, Fabio G (2011) Modulation of Nrf2/ARE pathway by food polyphenols: a nutritional neuroprotective strategy for cognitive and neurodegenerative disorders. Mol Neurobiol 44(2):192–201. https://doi.org/10.1007/s12035-011-8181-5
Liu M, Chen F, Sha L, Wang S, Tao L, Yao L, He M, Yao Z et al (2014) (−)-Epigallocatechin-3-gallate ameliorates learning and memory deficits by adjusting the balance of TrkA/p75NTR signaling in APP/PS1 transgenic mice. Mol Neurobiol 49(3):1350–1363. https://doi.org/10.1007/s12035-013-8608-2
Cui J, Othishankar B, He P, Staufenbiel M, Shen Y, Li R (2014) Amyloid precursor protein mutation disrupts reproductive experience-enhanced normal cognitive development in a mouse model of Alzheimer’s disease. Mol Neurobiol 49(1):103–112. https://doi.org/10.1007/s12035-013-8503-x
Freits AE, Egea J, Buendia I, Gomez-Rangel V, Parada E, Navarro E, Casas AI, Wojnicz A et al (2016) Agmatine, by improving neuroplasticity markers and inducing Nrf2, prevents corticosterone-induced depressive-like behavior in mice. Mol Neurobiol 53(5):3030–3045. https://doi.org/10.1007/s12035-015-9182-6
Dwivedi S, Rajasekar N, Hanif K, Nath C, Shukla R (2016) Sulforaphane ameliorates okadaic acid-induced memory impairment in rats by activating the Nrf2/HO-1 antioxidant pathway. Mol Neurobiol 53(8):5310–5323. https://doi.org/10.1007/s12035-015-9451-4
Acknowledgements
This research was supported by the Brain Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science and ICT (2016M3C7A1904391).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The animal maintenance, treatments, behavioral studies and surgical procedures were carried out in accordance with the animal ethics committee (IACUC) guidelines issued by the Division of Applied Life Sciences, Department of Biology at Gyeongsang National University, South Korea. The experimental methods with animals were carried out in accordance with the approved guidelines (Approval ID: 125) and all experimental protocol were approved by the animal ethics committee (IACUC) of the Division of Applied Life Sciences, Department of Biology at Gyeongsang National University, South Korea.
Conflict of Interest
The authors declared no conflict of interest.
Rights and permissions
About this article

Cite this article
Ali, T., Kim, T., Rehman, S.U. et al. Natural Dietary Supplementation of Anthocyanins via PI3K/Akt/Nrf2/HO-1 Pathways Mitigate Oxidative Stress, Neurodegeneration, and Memory Impairment in a Mouse Model of Alzheimer’s Disease. Mol Neurobiol 55, 6076–6093 (2018). https://doi.org/10.1007/s12035-017-0798-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12035-017-0798-6