
Abstract
A high fat diet (HFD) is considered to be a risk factor for the development of dementia because it increases oxidative stress in the brain; thus, diets rich in antioxidants, such as polyphenols, may protect against oxidative damage. In this study, we explored the antioxidant activity and neural protective function of millet on high fat diet-induced oxidative stress in rat brains. Our results suggested that the intake of millet could alleviate oxidative stress in the hippocampus and downregulate the expression of the Alzheimer’s disease (AD)-related genes amyloid precursor protein (App), tau, and γ-secretase. Furthermore, we extracted millet polyphenols and verified whether they play important roles during this process. The results showed that millet polyphenols significantly prevented H2O2-induced cell death of SH-SY5Y cells and decreased oxidative stress levels in cells. In addition, the expression levels of pro-inflammatory factors and AD-related genes were also downregulated by treatment with millet polyphenols. The above results indicated that millet and millet polyphenols could exert neural protective effects under high fat diet-induced oxidative stress by upregulating the expression of antioxidant enzymes and downregulating the expression of AD-related genes.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Excessive intake of fat in the human diet presents high risks to human health; thus, it has become a global public health concern. In recent years, epidemiological studies have shown that a high-fat and high-calorie diet increases the risks of dementia [1] because a HFD has been demonstrated to induce oxidative stress in the brain [2], which can lead to brain dysfunction [3]. Oxidative stress is also reported to be an accelerator and exacerbator of neurodegenerative diseases, such as AD [4]. The brain is more vulnerable to oxidative stress than any other organ because it is rich in polyunsaturated fatty acids and lipid peroxidation causes severe damage to neural cells. Elevated oxidative stress also activates the expression of pro-inflammatory factors, causing inflammation in the brain [5], which can lead to dementia. Amyloid plaques and tau tangles formed by the aggregation of Aβ42 or tau found in the cerebral cortex and hippocampus are typical pathological characteristics of AD [6]. Studies have revealed that oxidative damage to Aβ could promote its aggregation and the subsequent development of AD [7]. It is predictable that the amelioration of elevated oxidative stress could possibly prevent or limit the extent of brain dysfunction.
Millet is an old traditional cereal that has been consumed as a staple food for a long time in northwest China [8]. Millet contained high levels of polyphenols showed excellent oxidation resistance [9]. However, few studies have mentioned the in vivo antioxidant function of millet polyphenols. A growing body of evidence suggests that polyphenols and their metabolites play important roles in regulating oxidative stress levels in the brain [10]. Dietary polyphenols may exert antioxidant and neuroprotective effects either by directly crossing the blood-brain barrier into the brain [11] or by being converted into small metabolic derivates with higher biological activity through the metabolism of intestinal microbiota [12].
In this paper, we examined the effect of millet on high fat diet-induced oxidative stress in the brain by detecting oxidative stress levels and the expression of both inflammatory factors and AD-related genes in the hippocampus. We also evaluated the function of millet polyphenols in an in vitro neural cell model to verify their neural protective effects.
Materials and Methods
Materials, Animals and Cell Lines
Foxtail millet was purchased from Shanxi Dongfangliang Life Technology Co. Ltd. Sprague Dawley rats were obtained from Shanghai Jiesijie Experimental Animal Co. LTD. Human neuroblastoma cells (SH-SY5Y) were obtained from the Chinese Academy of Sciences. Rat feed was processed and produced by Jiangsu Xietong Organism Engineering Co. Ltd., and formula of HFD was provided in supplementary files.
Set up of HFD Animal Model and Isolation of Hippocampus
A total of 24 healthy male Sprague Dawley rats (200 ± 5 g, aged 6 weeks) were randomly classified into three groups: a basic diet (BC) group, a HFD group, and a HFD with 50% millet (HFD + M) group. Food and water were available ad libitum in temperature- and humidity-controlled rooms with a 12 h light-dark cycle. Rats were continuously fed a basal diet (BC group) or high-fat diet (HFD and HFD + M groups) for 3 weeks, and then, the diet of the HFD + M group was changed to the 50% millet diet for another 3 weeks, while the diets of the BC and HFD groups were kept the same as before. The hippocampus was isolated and immediately stored in liquid nitrogen after the rats were anesthetized with 2% pentobarbital.
Detection of MDA, SOD and GSH
The levels of lipid peroxidation products, namely, malondialdehyde (MDA), superoxide dismutase (SOD), and glutathione (GSH), in the hippocampus or in SH-SY5Y cells were determined by using commercial kits (Beyotime Biotechnology Co. Ltd) following the manufacturer’s instructions.
RNA Extraction and QPCR Analysis
Total RNA was extracted by using Magzol reagent (Vazyme Biotech Co., Ltd) in accordance with the manufacturer’s instructions. The RNA was then converted to cDNA by using the Hiscript II Q RT Supermix for qPCR (+gDNA wiper) (Vazyme Biotech Co., Ltd) following the manufacturer’s instructions. Real-time PCR analysis was performed on the Thermo Life Tech ABI Quant Studio 3. β-actin was used as the internal control gene, and ΔΔCt was used to calculate the relative expression of genes. All of the primer sequences are listed in Supplementary Table S1 and S2.
Millet Polyphenol Extraction
Millet was cooked with a millet to water ratio of 1:1.2 using a rice cooker (MB-WFS 5017TM, Midea Group Finance Co., Ltd). The phenolic compounds were extracted by previously reported method with slight modification [13]. The specific method was described in Supplementary Method 2. The concentration of polyphenols was determined by the Folin-Ciocalteu method, and gallic acid was employed as a standard substance. The concentration of millet polyphenols is presented as mg gallic acid equivalents (GAE)/g.
Cell Culture and Chemical Treatment
SH-SY5Y cells were cultured in DMEM/F12 medium (Shanghai Lirui Co.) supplemented with 10% fetal bovine serum and maintained in a humidified atmosphere of 5% CO2 and 95% air at 37 °C for 2–3 days before experimentation. The cells were induced with 10 μM all-trans-retinoic acid (ATRA, Lirui Co.) for 48 h and then treated with 300 μM H2O2 or different concentrations of millet polyphenols (MP), which were diluted with culture medium to working concentrations of 100, 200, 300, 400, and 500 μg/mL, for 12 h.
In Vitro Antioxidant Activity Determination and Cell Viability Analysis
The radical scavenging activity and reducing power of 2,2-diphenyl-1-picrylhydrazyl radical (DPPH) were assessed by using a previously described method [14]. The 2,2′-azinobis (3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt (ABTS) scavenging assay was performed following a previously described method [15]. The anti-lipid peroxidation capacity was examined according to the method described by reseachers [16]. The cytoprotective activity of millet polyphenols on H2O2-induced cell injury was assessed with a 3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide (MTT) assay. The specific method is based on the manufacturer’s instructions from the MTT Cell Proliferation and Cytotoxicity Assay Kit (Beyotime Co., Ltd).
Western Blotting Analysis
Western blotting analysis was performed as described previously [17]. Cells were lysed by RIPA total protein lysate (AS1004, ASPEN) after treatment with millet polyphenols as described in section 2.6. Protein concentrations were determined by BCA assay (Beyotime Biotechnology Co., Ltd). The cell lysate of each sample was analyzed by using SDS-PAGE. The specific primary antibodies for western blot analysis were listed in Supplementary Method 3.
Statistical Analysis
All experiments were carried out in at least triplicate, and the data are presented as the mean ± SD. Statistical data were analyzed with GraphPad Prism. The significance of the viability of SH-SY5Y cells after treatment with MPs at different concentrations was analyzed with one-way ANOVA, while others were analyzed with Student’s t test.
Results and Discussion
Millet Ameliorated HFD-Induced Oxidative Stress in the Hippocampus
To verify whether millet affects HFD-induced oxidative stress in the rat hippocampus, we established HFD rat models and fed the rats millet-supplemented diets. The expression levels of oxidative stress-related genes in the hippocampus showed that the expression of Sod1, Sod2, Cat and GPx2 was downregulated in the HFD group, whereas GPx1 was not affected. By contrast, when rats were fed with millet, the expression of these genes was upregulated (Fig. 1a). In addition, SOD activity was also significantly increased in the hippocampus of the HFD + M group, while MDA and GSH levels were not affected in either the HFD or HFD + M groups (Fig. 1b–d). These results indicated that a HFD weakened the antioxidant system in the brain by decreasing the expression or activity of antioxidant enzymes, while millet intake could reverse this effect. In other words, millet could suppress HFD-induced oxidative stress by increasing the expression and activities of antioxidant enzymes in the hippocampus.

The effect of millet on HFD-induced oxidative stress in the rat hippocampus. a The relative expression levels of Sod1, Sod2, Cat, GPx1 and GPx2; b SOD activity; c MDA levels; d GSH levels; e The relative expression levels of Tnf-α, Il-1β and NF-κB; f the relative expression levels of App, tau, γ-secretase, PS1, Bace1 and Ide. * represents the significance between the BC and HFD groups; # represents the significance between the HFD and HFD + M groups. *p < 0.05, **p < 0.01, ****p < 0.0001; #p < 0.05, ##p < 0.01, ###p < 0.001, ####p < 0.0001
Millet Intake Alleviated Neural Inflammation in the Hippocampus and Decreased the Expression of AD-Related Genes
Elevated oxidative stress levels often lead to inflammation, which is also an important cause of AD. To determine whether the intake of millet could inhibit neuroinflammation and decrease the risk of AD, we examined the expression of inflammatory factors and AD-related genes in rat hippocampus. Our results indicated that HFD feeding significantly increased the expression of the inflammatory factors Tnf-α and Il-1β in rat hippocampus, whereas millet feeding completely abolished this effect, resulting in significantly lower gene expression levels (Fig. 1e). The key transcription factor of both Tnf-α and Il-1β, the expression level of NF-κB, were also upregulated in the HFD group and downregulated in the HFD + M group (Fig. 1e). Studies of kodo millet also proved the anti-inflammatory effect of millet intake. It has been found that millet whole grain and bran supplementation decreases serum Il-6 and expression of NF-κB in the liver [18].
Oxidative stress is reported to be related to the hyperphosphorylation of tau and the aggregation of Aβ [19]. The release of the toxic Aβ42 peptide from App requires cleavage by γ-secretases [20]. In this study, we found that the expression levels of App, tau and γ-secretase were markedly increased in the HFD group compared with the BC group, and millet intake significantly lowered their expression (Fig. 1f). PS1, Bace1and Ide are also typical AD-related genes, and a HFD did not alter the expression of these genes, while millet intake downregulated the expression of Ide (Fig. 1f). The above results indicated that millet intake could suppress the inflammation caused by a HFD and reduce the risk of AD by downregulating the expression of App and tau, although the protein level needs to be further determined.
Millet Polyphenols Showed Antioxidant Activity In Vitro and Promoted the Viability of SH-SY5Y Cells under Oxidative Stress
Polyphenols are important antioxidants in plant food. In previous studies, the antioxidant activity of polyphenols from cereal grains towards HFD-induced oxidative stress has been discussed. It has been reported that barley polyphenols show strong free radical scavenging activity and high ferric reducing antioxidant ability and could downregulate the expression of oxidative stress-related genes in HFD mouse livers, such as Sod, Cat, and GPx [21]. Here, we extracted millet polyphenols and detected their antioxidant activity in vitro. The total phenolic concentration of the alcohol extract was 3.308 mg GAE/g. The free radical scavenging ability, reducing power and anti-lipid peroxidation capacity increased accompanying the increased concentration of millet polyphenols (Fig. S1). These results suggested that millet polyphenols have high antioxidant activities in vitro.
Next, we established an H2O2-induced oxidative stress cell model to detect the protective effect of millet polyphenols on neural cells under oxidative stress. MTT results showed that treatment of millet polyphenols (100 μg/mL, 200, 300, 400, and 500 μg/mL) did not affect the viability of SH-SY5Y cells (Fig. S2a). When cells were treated with 300 μM H2O2, the viability of the SH-SY5Y cells was decreased significantly, whereas the administration of millet polyphenols markedly and significantly improved cell viability (Fig. S2b). Our findings indicated that millet polyphenols showed a protective effect against oxidative stress-induced damage in neural cells.
Millet Polyphenols Exerted Antioxidant Effects on H2O2-Treated SH-SY5Y Cells
To verify whether millet polyphenols exert a protective function through their antioxidant activity, we examined oxidative stress levels in H2O2-induced SH-SY5Y cells. QPCR analysis of Sod, Cat, and GPx genes revealed that Sod expression was not influenced, while Cat and GPx2 expression levels were decreased when the cells were treated with H2O2; millet polyphenol treatment increased their expression significantly (Fig. 2a). Measurement of SOD activity in SH-SY5Y cells suggested that millet polyphenol treatment significantly increased SOD activity in cells compared to the H2O2 group (Fig. 2b). In addition, the MDA level was increased when the cells were treated with H2O2, and millet polyphenol treatment significantly reduced it (Fig. 2c). We also observed that the GSH level was not altered in either the H2O2 group or in the H2O2 + MP group (Fig. 2d), which is in accordance with the in vivo results. These results indicated that millet polyphenol treatment elevated the antioxidant capacity of neural cells to confront oxidative stress.

Millet polyphenol treatment altered H2O2-induced oxidative stress levels in SH-SY5Y cells. a The relative expression levels of Sod1, Sod2, Cat, GPx1 and GPx2 analyzed by QPCR; b SOD activity; c MDA levels; d GSH levels. * represents the significance between the control and H2O2 treated group; # represents the significance between the H2O2 group and H2O2 + MP group. *p < 0.05, **p < 0.01, ****p < 0.0001; #p < 0.05, ##p < 0.01, ####p < 0.0001
Millet Polyphenol Treatment Decreased the Expression of Inflammatory Factors and AD-Related Genes in SH-SY5Y Cells
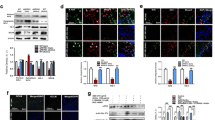
The expression of inflammatory factors and AD-related genes in H2O2-induced SH-SY5Y cell oxidative stress models was examined. From the results, we observed that millet polyphenols significantly suppressed the upregulation of the H2O2-induced inflammatory factors Tnf-α and Il-1β and the transcription factor NF-κB (Fig. 3a). Meanwhile, millet polyphenol treatment also suppressed the upregulation of App, tau, γ-secretase, PS1 and Bace1 induced by the H2O2 treatment (Fig. 3b). These results were consistent with the in vivo data, indicating that millet may exert a neural protective effect through the function of millet polyphenols. The protein levels of Tnf-α, Il-1β, NF-κB, tau and Aβ were also detected and found to be in accordance with their mRNA (Fig. 3c). The potential benefits of polyphenols protecting against the neuropathology of AD have been reported by many studies. For instance, grape seed extracted polyphenols are reported to interfere with the assembly of A1–42 peptide into neurotoxic Aβ aggregates [22]. In this study, our results indicated that millet polyphenols not only reduced the relative expression of App, the precursor of Aβ, but also influenced the expression of γ-secretase, PS1and Bace1, which was related to the cleavage of App. However, further studies need to be performed to verify the exact functional mechanism of how millet polyphenols affect the formation of Aβ aggregates (Figure 3).

Millet polyphenol treatment suppressed the H2O2-induced upregulation of pro-inflammatory factors and AD-related genes in SH-SY5Y cells. a the relative expression levels of Tnf-α, Il-1β and NF-κB; b the relative expression levels of App, tau, γ-secretase, PS1 and Bace1. c representative western blot for Tnf-α, Il-1β, tau, NF-κB, Aβ and GAPDH. * represents the significance between the control and H2O2 treated groups; # represents the significance between the H2O2 group and H2O2 + MP group. *p < 0.05, **p < 0.01, #p < 0.05, ##p < 0.01, ####p < 0.0001
Conclusion
In this study, we investigated the neural protective effect of millet towards high fat diet-induced oxidative stress. Examinations of oxidative stress-related indexes in the hippocampus suggested that millet exerted antioxidant activity by increasing the expression and activities of antioxidant enzymes, such as SOD, CAT, and GPx. Moreover, millet had anti-inflammatory and neural protective effects by downregulating the expression of the Tnf-α, Il-1β, App, PS1 and tau genes. Further examinations of cells indicated that millet polyphenols may exert dominating roles in this process. Our studies provide evidence of the antioxidant and neural protective activities of millet and millet polyphenols towards HFD-induced oxidative stress.
Abbreviations
- AD:
-
Alzheimer’s disease
- ABTS:
-
2,2′-Azinobis(3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt
- App:
-
Amyloid precursor protein
- ATRA:
-
All-trans-retinoic acid
- Bace1:
-
Beta-site App-cleaving enzyme I
- BC:
-
Basal diet control
- CAT:
-
Catalase
- DMEM:
-
Dulbecco’s minimum essential medium
- DPPH:
-
2,2-diphenyl-1-picrylhydrazyl radical
- GSH:
-
Glutathione
- HFD:
-
High fat diet
- MDA:
-
Malondialdehyde
- MP:
-
Millet polyphenols
- MTT:
-
3-(4,5-Dimethylthiazol-2-yl)-2,5 -diphenyltetrazolium bromide
- NF-κB:
-
nuclear factor kappa-B
- PS1:
-
Presenilin-1
- SOD:
-
Superoxide dismutase
References
Freeman LR, Haley-Zitlin V, Rosenberger DS, Granholm AC (2014) Damaging effects of a high-fat diet to the brain and cognition: a review of proposed mechanisms. Nutr Neurosci 17(6):241–251
Chung A, Gurtu S, Chakravarthi S, Moorthy M, Palanisamy UD (2018) Geraniin protects high-fat diet-induced oxidative stress in Sprague Dawley rats. Front Nutr 5:17. https://doi.org/10.3389/fnut.2018.00017
Kumar KH, Khanum F (2013) Hydroalcoholic extract of Cyperus rotundus ameliorates H2O2-induced human neuronal cell damage via its anti-oxidative and anti-apoptotic machinery. Cell Mol Neurobiol 33(1):5–17
Busquets O, Ettcheto M, Pallàs M, Beas-Zarate C, Verdaguer E, Auladell C, Folch J, Camins A (2017) Long-term exposition to a high fat diet favors the appearance of β-amyloid depositions in the brain of C57BL_6J mice. A potential model of sporadic Alzheimer’s disease. Mech Ageing Dev 162:38–45
Yeo ETY, Wong KWL, Seea ML, Wong KY, Gan SY, Chanb EWL (2018) Piper sarmentosum Roxb. confers neuroprotection on beta-amyloid (Aβ)-induced microglia-mediated neuroinflammation and attenuates tau hyperphosphorylation in SH-SY5Y cells. J Ethnopharmacol 217. https://doi.org/10.1016/j.jep.2018.02.025
Nagpure BV, Bian J-S (2014) Hydrogen sulfide inhibits A2A adenosine receptor agonist induced β-amyloid production in SH-SY5Y neuroblastoma cells via a cAMP dependent pathway. PLoS One 9:1–12
González-Reyes RE, Ariza-Salamanca D, Nava-Mesa MO, Mora-Muñoz L, Vargas-Sánchez K (2017) Involvement of astrocytes in Alzheimer’s disease from a neuroinflammatory and oxidative stress perspective. Front Mol Neurosci 10(427):1–20
Sharma S, Sharma N, Handa S, Pathania S (2017) Evaluation of health potential of nutritionally enriched Kodo millet (Eleusine coracana) grown in Himachal Pradesh, India. Food Chem 214:162–168
Kumari D, Madhujith T, Chandrasekara A (2017) Comparison of phenolic content and antioxidant activities of millet varieties grown in different locations in Sri Lanka. Food Sci Nutr 5:474–485
Filosa S, Di Meo F, Crispi S (2018) Polyphenols-gut microbiota interplay and brain neuromodulation. Neural Regen Res 13(12):2055–2059
Figueira I, Garcia G, Pimpao RC, Terrasso AP, Costa I, Almeida AF, Tavares L, Pais TF, Pinto P, Ventura MR, Filipe A, McDougall GJ, Stewart D, Kim KS, Palmela I, Brites D, Brito MA, Brito C, Santos CN (2017) Polyphenols journey through blood-brain barrier towards neuronal protection. Sci Rep-UK 7(1):11456. https://doi.org/10.1038/s41598-017-11512-6
Edwards CA, Havlik J, Cong W, Mullen W, Preston T, Morrison DJ, Combet E (2017) Polyphenols and health: interactions between fibre, plant polyphenols and the gut microbiota. Nutr Bull 42:356–360
Dewanto V, Wu X, LIU RX (2002) Processed sweet corn has higher antioxidant activity. J Agric Food Chem 50:4959–4964
Bamdad F, Wu J, Chen L (2011) Effects of enzymatic hydrolysis on molecular structure and antioxidant activity of barley hordein. J Cereal Sci 54(1):20–28
Shang H, Zhang H, Guo Y, Wu H, Zhang N (2019) Effects of inulin supplementation in laying hens diet on the antioxidant capacity of refrigerated stored eggs. Int J Biol Macromol. https://doi.org/10.1016/j.ijbiomac.2019.10.234
Liu Y, Huang G (2018) The derivatization and antioxidant activities of yeast mannan. Int J Biol Macromol 107:755–761
Fukuyama K, Kakio S, Nakazawa Y, Kobata K, Funakoshi-Tago M, Suzuki T, Tamura H (2018) Roasted coffee reduces β-amyloid production by increasing proteasomal β-secretase degradation in human neuroblastoma SH-SY5Y cells. Mol Nutr Food Res 1800238:1–9. https://doi.org/10.1002/mnfr.201800238
Sarma SM, Khare P, Jagtap S, Singh DP, Baboota RK, Podili K, Boparai RK, Kaur J, Bhutani KK, Bishnoi M, Kondepudi KK (2017) Kodo millet whole grain and bran supplementation prevents high-fat diet induced derangements in a lipid profile, inflammatory status and gut bacteria in mice. Food Funct 8(3):1174–1183
Cheignon C, Tomas M, Bonnefont-Rousselot D, Faller P, Hureau C, Collin F (2018) Oxidative stress and the amyloid beta peptide in Alzheimer's disease. Redox Biol 14:450–464
Chakravarthy B, Gaudet C, Menard M, Atkinson T, Brown L, Laferla FM, Armato U, Whitfield J (2010) Amyloid-beta peptides stimulate the expression of the p75(NTR) neurotrophin receptor in SHSY5Y human neuroblastoma cells and AD transgenic mice. J Alzheimers Dis 19(3):915–925
Shen Y, Zhang H, Cheng L, Wang L, Qian H, Qi X (2016) In vitro and in vivo antioxidant activity of polyphenols extracted from black highland barley. Food Chem 194:1003–1012
Wang D, Ho L, Faith J, Ono K, Janle EM, Lachcik PJ, Cooper BR, Jannasch AH, D'Arcy BR, Williams BA, Ferruzzi MG, Levine S, Zhao W, Dubner L, Pasinetti GM (2015) Role of intestinal microbiota in the generation of polyphenol-derived phenolic acid mediated attenuation of Alzheimer's disease beta-amyloid oligomerization. Mol Nutr Food Res 59(6):1025–1040
Acknowledgements
This work was supported by the National Key Research and Development Program of China [Grant number: 2017YFD0401202] and the National Natural Science Foundation of China [Grant number: 31901609].
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
We do not have conflicts of interest to declare.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Sen Li and Furong Xian are Co-first authors
Rights and permissions
About this article

Cite this article
Li, S., Xian, F., Guan, X. et al. Neural Protective Effects of Millet and Millet Polyphenols on High-Fat Diet-Induced Oxidative Stress in the Brain. Plant Foods Hum Nutr 75, 208–214 (2020). https://doi.org/10.1007/s11130-020-00802-6
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11130-020-00802-6