
Abstract
We studied the production of rhamnolipids by Pseudomonas aeruginosa UFPEDA 614 in submerged culture, using glycerol as the carbon source. A rhamnolipid yield of 15.9 g/L was obtained with 40 g/L glycerol and 5 g/L sodium nitrate as nitrogen source after 7 days of cultivation. Structural analysis carried out at different cultivation periods showed that the four major mono-rhamnolipid homologues are present in higher proportion in the first 48 h. Over time, the corresponding four major di-rhamnolipid homologues predominated, representing about 75 % of the total rhamnolipids after 96 h. Physicochemical analysis of the rhamnolipid mixtures obtained at different cultivation periods showed that the sample obtained from the first day of cultivation had the lower critical micelle concentration (15.6 mg/L), which is probably related to the higher proportion of mono-rhamnolipids. The results presented here show that the composition of the mixture of rhamnolipid homologues produced by P. aeruginosa UFPEDA 614 varies over time and that this variation influences the physicochemical properties of the mixture. These findings can be used in order to produce rhamnolipid mixtures that have suitable properties for different applications.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Surfactants reduce the surface tension at air-liquid interfaces and the interfacial tension at liquid-liquid interfaces [1]. Their amphipathic nature enables them to form microemulsions, which allow the solubilization of hydrophobic compounds in water. These properties make them suitable to be used in many sectors of modern industry [2]. For example, they are used as emulsifiers in the food and cosmetics industries [3], as components in both domestic and industrial cleaning products and as additives in the construction and oil industries. Almost all surfactants currently used in industry are synthesized from petroleum, which is a nonrenewable resource [2]. Additionally, these surfactants are often toxic to microorganisms, which limit their suitability for bioremediation processes [4].
Biosurfactants are surfactants produced by living organisms. They can be used in all of the industrial segments mentioned above, but their ecological compatibility makes them especially suitable for processes that involve the dispersion of surfactants in the environment, such as bioremediation of soils and aquifers contaminated by recalcitrant hydrophobic compounds [5], microbially enhanced oil recovery (MEOR) and biological control of diseases and pests [6].
One of the most important biosurfactants is the mixture of rhamnolipid homologues produced by Pseudomonas strains [7–10]. These strains are known to produce two main classes of homologues: the mono-rhamnolipids, which contain one unit of rhamnose and two of β-hydroxydecanoic acid (Rha-C10-C10), and the di-rhamnolipids, which contain two units of rhamnose linked to two units of hydroxydecanoic acid (Rha-Rha-C10-C10). Pseudomonas strains have been reported to produce as many as 28 different homologues, with saturated and unsaturated acyl chains, varying from C8 to C14, and with branched sugar moieties [7, 11–13]. The composition of the rhamnolipid mixture influences its surface properties and varies with the strain, the culture media composition, the age of the culture and the fermentation conditions [14].
The use of rhamnolipids has been limited by their high production costs relative to those of chemical surfactants [15]. Various workers have attempted to reduce costs by improving the rhamnolipid production levels, through manipulation of nutrients in submerged fermentation [16] and even by using solid-state fermentation [16–18]. However, many processes involve relatively long cultivations, from 12 to 30 days [19].
In the current work, we explored the possibility of achieving high productivities in shorter cultivation periods, while maintaining a desirable set of surface properties of the rhamnolipids produced by Pseudomonas aeruginosa UFPEDA 614. In order to do so, we evaluated the effect of the cultivation time by analyzing the composition of the rhamnolipid mixture by electrospray ionization–mass spectrometry (ESI-MS) and by determining its surface properties such as critical micelle concentration (CMC) and emulsifying capacity.
Materials and Methods
Microorganism and Cultivation
P. aeruginosa UFPEDA 614, originally isolated from a petrochemical plant located in Canto do Amaro, Rio Grande do Norte, Brazil, was provided by the Department of Antibiotics of the University of Pernambuco, Recife, Brazil. Stock cultures were stored in Luria-Bertani (LB) broth with 40 % (w/v) glycerol at −18 °C. A seed culture was prepared by inoculating 1 mL of stock culture into 50 mL of LB broth, in a 250-mL conical flask. The seed culture was incubated at 30 °C and 200 rpm in an orbital shaker until the optical density (at 600 nm) reached 0.6 to 0.8, indicating mid-exponential phase. One millilitre of this broth was then inoculated into a 250-mL Erlenmeyer flask containing 50 mL of a mineral salt medium containing (in g/L) the following: KH2PO4, 3.0; K2HPO4, 7.0; MgSO4, 0.2; glycerol, 40 and NaNO3, 5. Flasks were incubated at 30 °C and 200 rpm on an orbital shaker for 12 days.
Quantification of Biomass
Two millilitres of the culture were centrifuged at 8000×g for 20 min; the resulting pellet was washed with saline solution and then dried at 60–80 °C until constant weight. The biomass was expressed as dry weight of bacterial cells per litre of culture.
Extraction of Rhamnolipids
Each culture broth was centrifuged for 10 min at 12,500×g and the supernatant was extracted at least three times, each time with 15 mL chloroform–methanol (3:1, v/v). The organic phase was concentrated at reduced pressure at 40 °C, giving rise to a crude extract containing the rhamnolipid mixture.
Quantification of Rhamnolipids
Rhamnolipids were quantified by measuring the amount of rhamnose using the phenol–sulfuric acid method [20] with rhamnose as the standard. A control was done with a sample prepared from uncultured medium in order to check for interference from medium components. Since the rhamnose moiety represents only part of the rhamnolipid molecule, it was necessary to multiply the mass of rhamnose by a correction factor. This factor has been calculated as ranging between 3.0 and 3.4 [21, 22]. An average value of 3.2 was used, in accordance with Camilios-Neto et al. [17].
Quantification of Glycerol
Glycerol was quantified in the culture supernatant according to the method of Soloni [23], with the modifications proposed by Sassaki et al. [24]. In the assays, due to the high glycerol content of the samples, a sample volume of 1 mL was mixed with 1 mL of the periodate reagent. Then, 2.5 mL of the acetylacetone solution (1 % (v/v) in isopropanol) was added and the solution kept at 56 °C for 20 min. The absorbance was read at 410 nm and the glycerol content of the samples was calculated from a calibration curve prepared from a stock solution of glycerol.
Electrospray Ionisation–Mass Spectrometry
ESI-MS analyses were performed with a triple Quadrupole Quattro LC (Micromass, Manchester, UK) in negative-ion mode. The rhamnolipids (2 mg of each fraction) were dissolved in 1 mL chloroform–methanol (1:1, v/v), and aliquots of 0.1 mL were removed and diluted in 1.9 mL of acetonitrile–water (7:3, v/v), which was introduced by direct infusion with a syringe pump at a flow rate of 10 μL/min. ESI tandem mass spectra were acquired by mass-selecting the target ion using a quadrupole mass analyzer. The conditions of the analyses were as follows: F1—capillary 2.03 kV, cone 15 V and collision energy 18 V for collision-induced dissociation (CID); F2—capillary 2.03 kV, cone 27 V and collision energy 45 V for CID. The relative abundances of the rhamnolipid homologues in the mixture were obtained from the relative intensities of their corresponding pseudomolecular ions or fragments [9].
Surface Properties of the Crude Organic Extract
The surface properties of the rhamnolipid mixtures obtained at different culture periods were evaluated. The surface tension was determined using a Du-Nouy Tensiometer (Kruss K12, Germany) at 25 °C using the ring correction mode of the instrument. The CMC in water was determined by plotting the surface tension as a function of surfactant concentration and considering the CMC as the point where the slope of the curve abruptly changes [25]. To determine the emulsifying properties, 6 mL aliquots of different compounds (commercial kerosene, gasoline or diesel) were added to 4 mL aqueous samples containing 1.0 g/L of rhamnolipids (except for the sample from the first day, which, due to its higher mono-rhamnolipid content, could only be solubilized at 0.3 g/L of rhamnolipids). The amount of rhamnolipid in the aqueous phase was determined by the phenol–sulphuric method, as described above. Each mixture was vortexed for 2 min and then left to stand at room temperature [26]. The value of the emulsification index (E 24) was determined as the height of the emulsified layer, expressed as a percentage of the total height of the liquid column, after 24 h.
Results
Kinetics of Rhamnolipid Production
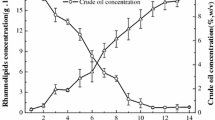
In previous studies of rhamnolipid production in submerged cultivation with P. aeruginosa UFPEDA 614, we obtained 12.4 g/L of rhamnolipids in 9 days [9] and 4.2 g/L of rhamnolipids in 4 days [26]. The culture medium contained 30 g/L glycerol and 1 g/L ammonium sulphate, giving a C/N ratio of 55. In the current work, the glycerol concentration was increased to 40 g/L and ammonium sulphate was replaced with 5 g/L sodium nitrate, giving a C/N ratio of 20. The rhamnolipid concentration initially increased rapidly, reaching 12 g/L in only 4 days (Fig. 1). After this, the production of rhamnolipids was slower, with a maximum concentration of 15.9 g/L being reached at 7 days. The biomass concentration reached 3.8 g/L on day 4, at which time the glycerol was depleted from the medium.

Kinetics of the production of rhamnolipids by Pseudomonas aeruginosa UFPEDA 614 with 40 g/L glycerol and 5 g/L sodium nitrate. Filled diamonds indicate rhamnolipid (g/L); filled squares indicate biomass (g/L); filled triangles indicate glycerol (g/L). All determinations were performed in triplicate and data are given as average values ± standard error of the mean. Where error bars are not visible, they are smaller than the size of the symbol
Evaluation of the Production of Rhamnolipid Homologues
Table 1 shows the temporal profiles for the relative abundances and concentrations of the various homologues produced by P. aeruginosa UFPEDA 614. On day 1, Rha-C20 accounted for 47 % of the total rhamnolipids, while Rha2-C20 accounted for only 24.5 %. By day 2, these two homologues each accounted for between 35 and 40 % of the total. For longer cultivation periods, the di-rhamnolipid homologues accounted for 70 to 75 % of the total, with Rha2-C20 being by far the major di-rhamnolipid homologue.
At all culture periods the mono-rhamnolipid homologue Rha-C20 and the di-rhamnolipid homologue Rha2-C20 accounted for at least 60 % of the total rhamnolipids, which is in agreement with previous reports [9, 12, 14, 27]. Six other homologues were present in smaller quantities, namely Rha-C18, Rha-C22:1, Rha-C22, Rha2-C18, Rha2-C22:1 and Rha2-C22. They were detected at all cultivation periods except for the absence of Rha2-C18 on the first day (Table 1).
Surface Properties of the Rhamnolipid Mixture
The critical micelle concentration (CMC) and the emulsification index (E 24) were determined for aqueous suspensions of the organic extract of the culture broth, containing 0.3 g/L rhamnolipids for the sample from the first day of culture and 1 g/L rhamnolipids for the samples from days 2, 4, 7 and 9. The CMC of the rhamnolipid mixture corresponding to the first day of cultivation (15.6 mg/L) was about a half of the others (Table 2), which might be due to the predominance of mono-rhamnolipids in this sample (Tables 1 and 2). The presence of only one rhamnose unit in the mono-rhamnolipids would make them less soluble in water than the di-rhamnolipids, which have two rhamnose units. As a consequence, the CMC would be reached at a lower concentration of mono-rhamnolipids [26, 28].
Additionally, the sample from the first day of cultivation was unable to emulsify any of the three hydrocarbons tested (Table 2). The sample from day 2 gave a significant E 24 value only for diesel (about 50 % of emulsification).
Discussion
The current work shows that it is possible to produce a high concentration of rhamnolipids (12 g/L in a cultivation time of only 4–7 days with P. aeruginosa DAUPE 614, a strain isolated from a petrochemical plant in Northeast Brazil). In a previous work [9], with a culture medium containing 3 % (w/v) glycerol and ammonium sulphate as the nitrogen source, this strain produced a concentration of only 3.9 g/L after 9 days of culture. The relatively short process time that we achieved in the present work brings two main advantages: First, it decreases the cost of production of rhamnolipid and, secondly, it reduces the possibility of the contamination of the culture. Contamination is more of a problem for the production of biosurfactants than for other microbial products [29, 30]. If efforts are not made to break the foam, it can fill the headspace, coming into contact with seals in the headplate and facilitating the entry of contaminants.
The production of 12 g/L of rhamnolipids in 4 days that was obtained in the current work is the highest value obtained in such a short time using a medium based on glycerol and inorganic nitrogen sources. With these nutrients, other authors have obtained rhamnolipid levels from 7.0 to 9.5 g/L in 4 days of cultivation [31, 32]. It is possible to obtain higher levels using fatty acids or oils as the carbon source. For example, a production of as much as 18.7 g/L in 4 days was reported for a Pseudomonas strain cultivated in soap stock, which contains around 50 % of oil and 50 % of free fatty acids, as the sole carbon source [22, 33]. However, the presence of residual fatty acids in the final culture broth has disadvantages: First, the recovery and purification of the rhamnolipids is more difficult [17, 18, 26]. Secondly, if the culture broth is to be used in an environmental application, the fatty acids can increase the load of insoluble organic compounds in the soil or water to be treated [26]. The use of glycerol avoids these problems. It has an additional advantage: since it is a by-product of the production of biodiesel by transesterification of oils, it is cheap and abundant in some countries, particularly in Brazil.
The results of our work support the hypothesis that mono-rhamnolipids are precursors for di-rhamnolipids. As mono-rhamnolipids become more available inside the cell, rhamnosyltransferase 2 (RhlC), the enzyme responsible for the addition of the second rhamnosyl group to a mono-rhamnolipid to form a di-rhamnolipid [34], starts to catalyze the synthesis of di-rhamnolipids with a higher rate. The basal mono-rhamnolipid concentration that remains in the culture medium suggests that some of the mono-rhamnolipids that are produced are transported out of the cell, becoming unavailable to be converted to di-rhamnolipids by RhlC [34, 35].
Although previous authors have already shown that the composition of the rhamnolipid mixture changes during the cultivation [7, 32], the goal of our study was to investigate how these changes affect the surface properties of the biosurfactant mixture. The rhamnolipid mixture produced at 4 days has surface active and emulsifying properties similar to those of mixtures produced in longer cultivation periods (12 to 30 days) [26].
Regarding the differences found in the CMC and E 24 determinations, lower values of CMC are found for mixtures rich in mono-rhamnolipids [36–38]. However, in the presence of hydrophobic solvents, the nonpolar molecules, such as mono-rhamnolipids with larger β-hydroxy fatty acid chains, tend to dissolve in the form of reverse micelles, with the nonpolar tail pointing outside the micelle, whereas the polar portions containing rhamnose interact with each other and with the water inside the reverse micelle [36]. This behaviour means that mixtures that are rich in mono-rhamnolipids have relatively poor ability to produce an aqueous emulsion. However, if the aim is to obtain rhamnolipids with lower CMC values, for applications that do not involve emulsification, the cultivation time needs to be shorter, as shown by the results presented in this work.
Conclusions
The composition of the mixture of rhamnolipid homologues produced by P. aeruginosa UFPEDA 614 varies over time and this variation influences the physicochemical properties of the mixture. These findings can be used in order to produce rhamnolipid mixtures that have suitable properties for specific applications, such as biodegradation and other environmental processes.
References
Bodour, A., & Miller-Maier, R. (1998). Journal of Microbiological Methods, 32, 273–280.
Cameotra, S. S., & Makkar, R. S. (1998). Applied Microbiology and Biotechnology, 50, 520–529.
Kim, H., Yoon, B., Lee, C., Suh, H., Oh, H., Katsuragi, T., & Tani, Y. (1997). Journal of Fermentation and Bioengineering, 84, 41–46.
Christofi, N., & Ivshina, I. B. (2002). Journal of Applied Microbiology, 93, 915–929.
Bonilla, M., Olivaro, C., Corona, M., Vazquez, A., & Soubes, M. (2005). Journal of Applied Microbiology, 98, 456–463.
Nielsen, T. H., Christophersen, C., Anthoni, U., & Sørensen, J. (1999). Journal of Applied Microbiology, 87, 80–90.
Déziel, E., Lépine, F., Milot, S., & Villemur, R. (2000). Biochimica et Biophysica Acta, 1485, 145–152.
Benincasa, M., Abalos, A., Oliveira, I., & Manresa, A. (2004). Antonie Van Leeuwenhoek, 85, 1–8.
Monteiro, S. A., Sassaki, G. L., de Souza, L. M., Meira, J. A., de Araújo, J. M., Mitchell, D. A., Ramos, L. P., & Krieger, N. (2007). Chemistry and Physics of Lipids, 147, 1–13.
Chrzanowski, L., Lawniczak, L., & Czaczyk, K. (2012). World Journal of Microbiology Biotechnology, 28, 401–419.
Nitschke, M., & Pastore, G. M. (2006). Bioresource Technology, 97, 336–341.
Soberon-Chavez, G., Lepine, F., & Deziel, E. (2005). Applied Microbiology and Biotechnology, 68, 718–725.
Mulligan, C. N. (2005). Environmental Pollution, 133, 183–198.
Mata-Sandoval, J. C., Karns, J., & Torrents, A. (1999). Journal of Chromatography. A, 864, 211–220.
Banat, I. M., Franzetti, A., Gandolfi, I., Bestetti, G., Martinotti, M. G., Fracchia, L., Smyth, T. J., & Marchant, R. (2010). Applied Microbiology and Biotechnology, 87, 427–444.
Clarke, K., Ballot, F., & Reid, S. (2010). World Journal of Microbiology Biotechnology, 26, 2179–2184.
Camilios Neto, D., Meira, J. A., de Araújo, J. M., Mitchell, D. A., & Krieger, N. (2008). Applied Microbiology and Biotechnology, 81, 441–448.
Camilios-Neto, D., Bugay, C., de Santana-Filho, A. P., Joslin, T., de Souza, L. M., Sassaki, G. L., Mitchell, D. A., & Krieger, N. (2011). Applied Microbiology and Biotechnology, 89, 1395–1403.
Pornsunthorntawee, O., Wongpanit, P., Chavadej, S., Abe, M., & Rujiravanit, R. (2008). Bioresource Technology, 99, 1589–1595.
Dubois, M., Gilles, K., Hamilton, J., Rebers, P., & Smith, F. (1956). Analytical Chemistry, 28, 350–356.
Itoh, S., Honda, H., Tomita, F., & Suzuki, T. (1971). The Journal of Antibiotics, 24, 855–859.
Benincasa, M., Contiero, J., Manresa, M., & Moraes, I. (2002). Journal of Food Engineering, 54, 283–288.
Soloni, F. G. (1971). Clinical Chemistry, 17, 529–534.
Sassaki, G. L., Cruz, L. M., Gorin, P. A., & Lacomini, M. (2001). Lipids, 36, 167–174.
Sheppard, J. D., & Mulligan, C. N. (1987). Applied Microbiology and Biotechnology, 27, 110–116.
Camilios Neto, D., Meira, J. A., Tiburtius, E., Zamora, P. P., Bugay, C., Mitchell, D. A., & Krieger, N. (2009). Biotechnology Journal, 4, 748–755.
Maier, R. M., & Soberón-Chávez, G. (2000). Applied Microbiology and Biotechnology, 54, 625–633.
Abalos, A., Pinazo, A., Infante, M., Casals, M., Garcia, F., & Manresa, A. (2001). Langmuir, 17, 1367–1371.
Krieger, N., Camilios Neto, D., & Mitchell, D. A. (2010). Advances in Experimental Medicine and Biology, 672, 203–210.
Yeh, M., Wei, Y., & Chang, J. (2006). Process Biochemistry, 41, 1799–1805.
Benincasa, M., & Accorsini, F. R. (2008). Bioresource Technology, 99, 3843–3849.
Haba, E., Abalos, A., Jauregui, O., Espuny, M., & Manresa, A. (2003). Journal of Surfactants and Detergents, 6, 155–161.
Abalos, A., Maximo, F., Manresa, M., & Bastida, J. (2002). Journal of Chemical Technology and Biotechnology, 77, 777–784.
Rahim, R., Ochsner, U. A., Olvera, C., Graninger, M., Messner, P., Lam, J. S., & Soberon-Chavez, G. (2001). Molecular Microbiology, 40, 708–718.
Abdel-Mawgoud, A., Hausmann, R., Lépine, F., Muller, M., & Déziel, E. (2011). Rhamnolipids: detection, analysis, biosynthesis, genetic regulation, and bioengineering of production. Berlin: Springer-Verlag.
Sánchez, M., Aranda, F. J., Espuny, M. J., Marqués, A., Teruel, J. A., Manresa, A., & Ortiz, A. (2007). Journal of Colloid and Interface Science, 307, 246–253.
Zhang, Y., Maier, W., & Miller, R. (1997). Environmental Science and Technology, 31, 2211–2217.
Zhang, Y., & Miller, R. M. (1995). Applied and Environmental Microbiology, 61, 2247–2251.
Acknowledgments
The work was supported financially by CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico), a Brazilian government agency for the advancement of science and technology. Research scholarships were granted to Arquimedes Paixão de Santana-Filho, Doumit Camilios-Neto and Lauro de Souza by CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nivel Superior), a Brazilian government agency for the development of scientific personnel and to Guilherme Sassaki, David Mitchell and Nadia Krieger by CNPq.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
de Santana-Filho, A.P., Camilios-Neto, D., de Souza, L.M. et al. Evaluation of the Structural Composition and Surface Properties of Rhamnolipid Mixtures Produced by Pseudomonas aeruginosa UFPEDA 614 in Different Cultivation Periods. Appl Biochem Biotechnol 175, 988–995 (2015). https://doi.org/10.1007/s12010-014-1343-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12010-014-1343-y