
Abstract
Due to the rapid global expansion of the aquaculture industry, access to key feedstuffs (fishmeal and fish oil) is becoming increasingly limited because of the finite resources available for wild fish harvesting. This has resulted in other sources of feedstuffs being investigated, namely plant origin substitutes for fishmeal and fish oil for aquafeed. Conventional land-based crops have been favored for some applications as substitutes for a portion of the fishmeal, but they can result in changes in the nutritional quality of the fish produced. Microalgae can be regarded as a promising alternative that can replace fishmeal and fish oil and ensure sustainability standards in aquaculture. They have a potential for use in aquaculture as they are sources of protein, lipid, vitamins, minerals, pigments, etc. This comprehensive review summarizes the most important and recent developments of microalgae use as supplement or feed additive to replace fishmeal and fish oil for use in aquaculture. It also reflects the microalgal nutritional quality and digestibility of microalgae-based aquafeed. Simultaneously, safety and regulatory aspects of microalgae feed applications, major challenges on the use microalgae in aquafeed in commercial production, and future research and development perspective are also presented in a critical manner. This review will serve as a useful guide to present current status of knowledge and highlight key areas for future development of a microalgae-based aquafeed industry and overall development of a sustainable aquaculture industry.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The aquaculture industry is growing three times faster than the terrestrial animal production industry as to compensate the loss of or stagnated production from natural or wild fisheries for the last 30 years (FAO 2014). In 2014, the total aquaculture food fish and crustacean production was 73.8 million tonnes valued at US$ 160.2 billion (FAO 2016). Approximately 70% of all aquaculture production is direct-fed species, which is roughly equivalent to 35.7 million tonnes of product. Of these direct-fed species, 68% are dependent on commercially produced feed products and consumed an estimated 39.6 million tonnes of aquafeed in 2012 (Tacon and Metian 2015). The aquafeed market has grown exponentially in the last decade, and this growth is expected to continue. Fishmeal and fish oil are commonly used in fish feed due to their respective content of high-quality proteins and beneficial omega-3 long-chain polyunsaturated fatty acids (LC-PUFA). Since fishmeal is produced from wild-harvested fish populations, both production limits and consequences for when those limits are surpassed exist. Many fishmeal fisheries stocks, such as the anchoveta (Engraulis ringens) stocks in the Southeast Pacific and the herring stocks in the North Atlantic, are considered fully exploited (FAO 2007). The FAO suggests that our reliance on fishmeal for aquaculture is undermining both marine biodiversity and human food security (FAO 2012). The proportion of fish processed into fishmeal is not likely to grow due to an increasing demand for fish products in emerging economies such as China. As fish farming has increased worldwide, the demand and the limits in wild fish catch have created an uncertain market for fishmeal with prices rising by almost 300% in the past 10 years (Origin Oil 2014). Trimmings from edible fish and some wild fish—typically small boney and oily fish such as capelin, sand eel, anchovy, horse mackerel, pilchard, and menhaden which are unappetizing for direct human consumption are being utilized for fish oil production. Global production of fish oil from these sources is estimated at one million tons annually. It is unlikely that this could change. Moreover, oily fish are expected to be processed for direct human use, thus reducing that available for fish oil production (Pike and Jackson 2010). Aquaculture is the biggest consumer of fishmeal and fish oil. On a global basis, in 2012, the aquaculture sector consumed 70% of global fishmeal production and 73% of global fish oil production (IFFO 2016). Within the next decade, fish oil production may not meet the required quantities for aquaculture, meaning that food-grade fisheries which provide fish oil and fishmeal have reached their maximum sustainable yield (Nasopoulou and Zabetakis 2012). The expansion of the aquafeed industry requires finding substitute ingredients because over-reliance of aquaculture feeds on fishmeal and fish oil may be unsustainable (Kiron et al. 2016; Sarker et al. 2016a; b). Plant protein-based substitutes have been proven to support good growth in fish, but at the same time, there are some disadvantages. Some of the plant protein-based alternatives include having low digestibility, are deficient in certain essential amino acids such as lysine, methionine, threonine, and tryptophan, and may result in significant changes in the nutritional quality of the final fish products (Li et al. 2009b). Some plant oil-based alternatives such as soybean oil, vegetable oil, cottonseed oil, and sunflower oil are rich in omega-6 contents but poor in omega-3 long-chain PUFAs which are very important for good growth and production of high consumer-quality fish products (Turchini et al. 2009). Moreover, most of these alternative sources usually have imbalanced nutrient profiles and may contain some antinutrients affecting performance of the fish and final product quality.
Microalgae have the potential to reduce the dependence on conventional raw materials in aquafeed. The use of microalgae could have significant beneficial effects and could potentially replace or reduce common feed stuff because of their nutritional quality and positive effect on the rate of growth of aquatic species due to increased triglyceride and protein deposition in muscle, improved resistance to disease, decreased nitrogen output into the environment, omega-3 fatty acid content, physiological activity, and carcass quality (Becker 2004). There are several other advantages such as microalgae can grow in a wide range of habitats, some species have several-fold higher biomass production than plants, can divide fast with simple nutritional requirements, can accumulate useful metabolites, and its availability is not dependent on wild fish harvesting for fishmeal (Hemaiswarya et al. 2011). There are some drawbacks and challenges of using microalgae as replacement of fishmeal and fish oil in the aquaculture industry, such as the high production costs of microalgae (Becker 2007; Sarker et al., 2016a), some microalgae may have poorly digestible cell walls (Skrede et al. 2011), while the availability of a large amount of biomass could be affected by contamination (Hannon et al. 2010), etc. Until recently, technological developments and commercial applications have mainly focused on microalgae as a microfeed ingredient, focusing on specific beneficial properties rather than gross nutrients to the recipient animal. Studies on the use of microalgae as a possible ingredient for aquafeed for different fish species have been increasing exponentially. Several recent reviews (Hemaiswarya et al. 2011; Priyadarshani and Rath 2012; Benemann 2013; Roy and Pal 2014; Yaakob et al. 2014) have indicated high potential for microalgae as a bulk feedstuff for aquaculture feeds. However, there is still limited availability of upgraded knowledge and recent progress on utilization of microalgal biomass to replace fishmeal and fish oil for development of sustainable aquaculture industry. This article reviews the current level of comprehensive knowledge on general attributes of microalgal species used in aquaculture and their nutritional properties and the use of microalgae in formulated aquafeeds as replacement of fishmeal and fish oil. Safety and regulatory aspects of microalgae feed applications and directions for future research and potential areas of industrial development are identified.
Microalgal nutritional quality as fishmeal and fish oil supplement or feed additive
Microalgae have great potential to provide protein, lipids, vitamins, carotenoids, and energy in feed. Generally, microalgae contain 30–40% protein, 10–20% lipid, and 5–15% carbohydrate in the late logarithmic growth phase (Brown et al. 1997). Chemical composition of some commercially important microalgal species is comparable with the available feed ingredients which are used in the aquafeed industry (Table 1). The chemical-nutrient composition of microalgae including proximate analysis, amino acid and fatty acid compositions, and vitamin and mineral contents has been well documented in various published reports (Brown 2002; Becker 2004, 2007; Shields and Lupatsch 2012; Yaakob et al. 2014; Kent et al. 2015; Tibbetts et al. 2015c). The nutritional value of microalgae is determined mainly by the protein content and, secondly, by the polyunsaturated fatty acid (e.g., eicosapentaenoic acid (EPA), arachidonic acid (ARA), and docosahexaenoic acid (DHA)) content (Reitan et al. 1997). Various auto- and heterotrophic microalgae species from different classes produce EPA and DHA, whereas AA is generally found in scarce amounts (Thompson 1996; Bigogno et al. 2002). According to recent reviews of total lipid extracts, Bacillariophyceae (diatoms) and Chrysophyceae species may be rich sources of EPA and DHA; Cryptophyceae, Prasinophyceae, Rhodophyceae, Xanthophyceae, Glaucophyceae, and Eustigmatophyceae can represent interesting EPA sources, whereas DHA is found in significant amounts mostly in Dinophyceae, Prymnesiophyceae, and Euglenophyceae (Hu et al. 2008; Lang et al. 2011). Some microalgae such as Arthrospira (Spirulina), Dunaliella, and Chlorella are commercially available, their nutritional profiles are well documented, and they have a long history of use as sources of animal feed. There are several recent studies on the utilization of microalgal biomass as replacement of fishmeal or fish oil or as supplement which is documented in Table 2. In the following sections, we will discuss in more details the specific nutritional properties of microalgae and their applications in aquafeed.
Microalgal protein
Microalgal protein could be a plausible alternative to fishmeal protein because of their good quality and amino acid profiles comparable with that of other reference protein sources (Becker 2007). For example, dried Arthrospira biomass contains all essential amino acids and ca. 68% of its biomass is protein (Blazencic 2007). Analyses of microalgae have found similar high contents of essential amino acids, as exemplified by a comprehensive study of 40 species of microalgae from seven algal classes that found that “All species had similar amino acid composition and were rich in essential amino acids” (Brown et al. 1997). Some recent studies also evaluated the biochemical composition, amino acid profiles, fatty acid, carotenoid and elemental composition of freshwater, and marine microalgal biomass for animal feed applications (Tibbetts et al., 2015a, b, d), and these studies indicated the potentiality of incorporating microalgal protein in feed. There are several studies on the utilization of biomass of Arthrospira sp., Chlorella sp., Scenedesmus sp., Nanofrustulum sp., and Tetraselmis suecica as valuable supplementary protein sources or partial substitutes for fishmeal protein in the diet of various omnivorous and carnivorous fish species (Badwy et al. 2008). Studies have indicated the ability of carnivorous fish to make use of proteins from microalgae (Table 2). The previous studies concerning low levels of fishmeal replaced with Arthrospira are uniformly positive. Between 0.5 and 10%, Arthrospira inclusion in diet has led to positive growth results. For example, Parrotfish (Oplegnathus fasciatus) fed with 5% Arthrospira in formulated feed experienced significantly higher weight gain, protein efficiency ratios, feed intake, and lower feed conversion ratios than the fishmeal control (Kim et al. 2013). Similarly, Rainbow trout (Oncorhynchus mykiss) experienced the highest weight gain when fishmeal was replaced with 7.5% Arthrospira (Teimouri et al. 2013). The consequences of using Arthrospira as a main protein source or in excess of 10% when replacing fishmeal are less clear. In many cases, growth results for diets containing over 10% Arthrospira protein have been encouraging. Golden barb (Puntius gelius) displayed significantly increased growth rates when 20% of the fishmeal in their diets was substituted for Spirulina (Hajiahmadian et al. 2012). Similarly, Silver seabream (Rhabdosargus sarba) fed a diet containing up to 50% Arthrospira did not suffer negative growth effects compared with a fishmeal control and had a similar feed conversion ratio (FCR) (El-Sayed 1994). The fishmeal could be substituted with up to 30% Arthrospira maxima meal without impacting growth performance in red tilapia fingerlings (Oreochromis sp.) (Rincón et al. 2012). Tilapia (larvae/juveniles) have been fed diets containing 43% Arthrospira without growth or feed intake being negatively impacted and had a better FCR than a corn-gluten meal control (Hussein et al. 2013). Similar results were obtained by Velasquez et al. (2016). Therefore, growth results for both omnivorous and carnivorous fish fed diets containing Arthrospira as a protein source have been significantly higher or comparable with those of the commercial or control diets. Other studies, however, have found that high Arthrospira inclusion results in depressed growth rates. Sharma and Panta (2012) and El-Sayed (1994) suggest that Arthrospira inclusion above 30% is detrimental to fish growth. Similarly, Olvera-Novoa et al. (1998) reported elevated growth rates in diets containing 10 and 20% Arthrospira protein but halved growth rates in diets containing more than 40% Arthrospira compared with a fishmeal control.
There are several studies which summarize the results of the inclusion of other microalgae species in aquafeeds (Table 2). Three aquaculture species (i.e., Atlantic salmon, common carp, and Pacific white shrimp) have been fed with algal meal (i.e., Nanofrustulum and Tetraselmis) as 5 or 10% replacement of fishmeal without significant changes in growth performance and feed utilization compared with the fishmeal-based feed (Kiron et al. 2012). The diatom Phaeodactylum tricornutum can replace up to 6% of the fishmeal without adverse effects on nutrient digestibility, utilization of the feed, and growth performance of Atlantic salmon, Salmo salar (Sørensen et al. (2016). Feed intake and growth improved in the carnivorous fish (juvenile Atlantic cod, Gadus morhua) fed the diet with 15% fishmeal protein replacement by algal meal (combination of dried Nannochloropsis sp. and Isochrysis sp.) (Walker and Berlinsky 2011). A feeding study on rainbow trout fry indicated a significant reduction in the growth of the fish that received 25 and 50% algae (a consortium of algal biomass consisting of Chlorophyceae and Cyanobacteria) in their diets, but at 12.5%, there was no negative effects on growth, body lipid, and energy content of the fish (Dallaire et al. 2007). Dried biomass of T. suecica was shown to be able to replace up to 20% fishmeal protein without hampering the growth performance of European sea bass (Tulli et al. 2012). The gilthead sea bream (Sparus aurata) juveniles were not affected by feeding with 38% microalgae (Scenedesmus almeriensis) (Vizcaíno et al. 2014). Vizcaíno et al. (2016) also showed that up to 5% T. suecica and Tisochrysis lutea could be added to the diets of gilthead sea bream fry.
Deoiled microalgae are rich in protein and essential amino acids that could be utilized as aquaculture feeds or as a dietary supplement. Partial replacement of fishmeal protein by a defatted microalgae meal based on Haematococcus pluvialis on Pacific white shrimp diet showed that it can be considered a valuable alternative protein and pigmentation in shrimp feed due to the presence of carotenoids in the microalgae meal (Ju et al. 2012). In their study, after an 8-week feeding trial, shrimp fed the diet with 12.5% of the fishmeal protein replaced showed a significantly higher growth rate and lower feed conversion ratio than the shrimp fed the control diet. In another study, the replacement of up to 10% of crude protein from fishmeal and soy protein concentrate with lipid-extracted algae (Navicula sp. and Nannochloropsis salina) was possible without causing significant reductions in growth performance of juvenile Red drum fish (Sciaenops ocellatus) (Patterson et al. 2013) (Table 2). Microalgal “green water meal” has also been shown to be a suitable fish meal replacement in the diets of juvenile Pacific white shrimp, Litopenaeus vannamei (Basri et al. 2015). Similarly, Arthrospira platensis at 8 to 10% of the diet improved growth and reproductive performance in the red cherry shrimp, Neocaridina davidi (Namaei Kohal et al. 2017). Given the nutritional composition of microalgae and its relative digestibility, it should be a viable alternative protein source for use in aquafeeds provided that feed intake is maintained. In addition, the processing of microalgal biomass during the elaboration of aquafeeds might also affect its digestibility which could be taken into consideration. For example, in a recent study, the defatted microalga Nannochloropsis sp. was more digestible than Desmodesmus sp., when fed to Atlantic salmon, S. salar and extrusion processing of microalgae helped to improve digestibility (Gong et al. 2017). The results in the literature suggest that the percentage of microalgal meal inclusion in the aquafeed could be varied with microalgae and aquaculture species. More research to evaluate the grow-out potential of microalgae and pinpointing factors affecting its effectiveness should be undertaken.
Microalgal lipid and fatty acids
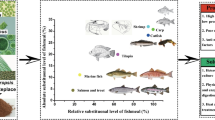
Lipids have roles as both energy storage molecules and in the formation of biological membranes. Oil content in microalgae can exceed 60% by weight of dry biomass, while levels of 20–50% are quite common (Guschina and Harwood 2013). Some microalgal lipids are often contain polyunsaturated fatty acids such as DHA (Crypthecodinium, Schizochytrium), EPA (Nannochloropsis, Phaeodactylum, Nitzschia, Isochrysis, Diacronema), and ARA (Porphyridium) (Brown 2002) (Fig. 1). These long-chain (LC)-PUFA are usually present in high concentrations. For example, content of EPA in EPA-producing microalgal species range from 7 to 34% of total fatty acids (Brown 2002) (Fig. 1). These fatty acids are highly sought after as they cannot be easily synthesized in a laboratory. They are currently obtained from fish oil and are considered a limiting factor in the use of vegetable oils such as palm, soybean, and rapeseed/canola oils in aquafeed. The fatty acid composition is associated with light intensity, culture media, temperature, and pH (Daroch et al. 2013). Appropriate measures and control, along with the suitable selection of a species, are necessary to produce microalgae with desired lipid level and composition. Several autotrophically and heterotrophically grown species of microalgae have been evaluated for their high EPA and/or DHA contents, such as Monodus sp., Nannochloropsis sp., Phaeodactylum sp., Schizochytrium sp. Thraustochytrium sp., and Aurantiochytrium sp. (Chauton et al. 2015; Sharma and Schenk 2015; Liu et al. 2014). The EPA and/or DHA contents in these species are reported to be from 2.2 to 3.9% in P. tricornutum (Ceron-Garcia et al. 2013; Qiao et al. 2016), 3% in Monodus subterraneus (Jo and Hur 2015), and 2.8–4.3% in Nannochloropsis spp. (Chen et al. 2015; Sharma and Schenk 2015). Microalgae oils from Isochrysis, Nannochloropsis, Phaeodactylum, Pavlova, and Thalassiosira contain sufficient omega-3 LC-PUFA to serve as an alternative for fish oil (Ryckebosch et al. 2014).

Average percentage composition of the long-chain PUFA docosahexaenoic acid (DHA), eicosapentaenoic acid (EPA), and arachidonic acid (ARA) of microalgae commonly used in aquaculture (adopted from Brown 2002)
EPA and DHA are essential for growth and survival of marine fish larvae (Kitajima 1983). Both polyunsaturated fatty acids can be synthesized by some of microalgae such as N. oculata and P. tricornutum among others. The use of ARA-rich microalga Parietochloris incisa as a dietary supplement during the first month of life of guppy fry (Poecilia reticulata) enhanced their survival and stress resistance (Nath et al. 2012). One of the microalgae used as a source of PUFA for the cultivation of salmon is represented by the thraustochytrid Schizochytrium sp. It is characterized by high lipid content (55–75% in dry matter) and up to 49% DHA of total lipids and commonly heterotrophically cultivated for large-scale production (Ren et al. 2010). Its exploitability as an alternative lipid source has been investigated in aquaculture to enrich zooplankton for feeding to finfish larvae (Barclay and Zeller 1996) or to replace fish oils in diets for Atlantic salmon (Carter et al. 2003; Miller et al. 2007; Kousoulaki et al. 2015; Sprague et al. 2016) and sea bream (Ganuza et al. 2008). The addition of dried microalgae (Schizochytrium sp.) in the diet, albeit at low concentrations of 1.0–1.5%, resulted in increased weight gain of the channel catfish (Ictalurus punctatus) along with improving the feed efficiency ratio and levels of PUFA (Li et al., 2009a). Spray-dried microalgae biomass (Schizochytrium sp.) that included up to 5% extruded feeds for Atlantic salmon (S. salar) can successfully replace fish oil as a source of omega-3 LC-PUFA without compromising fish growth rate and food conversion ratio (FCR), dietary protein, energy digestibility, and flesh quality. Dietary Schizochytrium sp. improved the retention efficiency of EPA + DHA and monounsaturated fatty acids and improved the technical quality of salmon fillets in terms of gaping. In particular, DHA-rich Schizochytrium sp. microalgae whole biomass did not reveal any signs of toxicity, stress, inflammation, or any other negative effects of supplementation in diets for Atlantic salmon (S. salar) (Kousoulaki et al. 2015) (Table 2). Whole cell Schizochytrium sp. significantly improved weight gain, feed conversion ratio, and protein efficiency ratio when fish oil was fully replaced by Schizochytrium sp. in tilapia diet. Tilapia fed with Schizochytrium sp. that fully replaced fish oil had higher content of DHA in the fillet lipids, and this reflected the higher DHA supplied by this diet (Sarker et al., 2016b). A feeding trial was conducted to assess the effects of dietary Schizochytrium meal supplementation on survival, growth performance, activities of digestive enzymes, and fatty acid composition in Pacific white shrimp (L. vannamei) larvae. It was concluded that Schizochytrium (4%) meal in microdiets of shrimps can improve growth performance and may be a valuable additive in the microdiets of shrimps (Wang et al. 2016). Mixtures of Nannochloropsis sp. and Schizochytrium sp. could replace 100% fish oil in diets for olive flounder (Paralichthys olivaceus) without negative effects on growth, feed efficiency, or nutritive quality (Qiao et al. 2014). Schizochytrium sp. and Crypthecodinium cohnii biomasses, either homogenized or non-homogenized, showed their potential as alternative DHA sources for fish oil in microdiets for gilthead seabream but also point out the necessity of EPA sources to completely replace fisheries-derived oils (Ganuza et al. 2008). Fish oil substitution by the dinoflagelate C. cohnii in early weaning diets for gilthead seabream (S. aurata) improved fish survival as well as growth performance, in agreement with a higher proportion of DHA in diets and in total lipids of fish (Atalah et al. 2007). A recent investigation has shown that replacing up to 36% fish lipid by the dried Isochrysis biomass in a diet with reduced level of fish oil, did not adversely affect feed intake or growth performance relative to controls for European sea bass (Dicentrarchus labrax) (Tibaldi et al. 2015). In another study, the potential of microalga Pavlova viridis (Diacronema viridis) as an omega-3 PUFA source was evaluated and compared with Nannochloropsis sp. in diets for juvenile European sea bass (D. labrax). Based on the basal diet (40% fish oil and 60% plant oil—in equal parts rapeseed, sunflower, and linseed oil), a 50% fish oil replacement by Nannochloropsis sp. meal and a total replacement by P. viridis meal were possible without negative effects on the growth performance and nutrient utilization of juvenile sea bass (Haas et al. 2016).
The results of the previous studies clearly indicate that microalgae can be a potential candidate to partially/completely replace the fish oil in aquafeed. It has been observed that some heterotrophic microalgae such as Schizochytrium and C. cohnii could be more promising than the other autotrophic microalgae discussed in this section, probably due to their nutritional quality and ability to be efficiently utilized by the desired aquaculture species. Since the mass cultivation technology of Schizochytrium is already well developed, it is expected that this microalga can be produced commercially in large quantity to reduce the use of fish oil in aquafeed in the future. More research should be focused on the utilization of promising microalgae species to replace fish oil at sustainable level in aquafeed.
Microalgal pigments
Pigments of microalgae contribute to their nutritional performance in aquaculture. Beside chlorophylls, there are other types of pigments found in microalgae such as carotenoids and phycobiliproteins. Among the microalgae, Nannochloropsis sp. is a well-known source of different valuable pigments, such as chlorophyll a, zeaxanthin, canthaxanthin, and astaxanthin. Another microalga, Phaeodactylum sp. is rich in fucoxanthin, carotene, chlorophyll a, and chlorophyll c (Brown et al. 1997), while Tetraselmis sp. contains chlorophyll a and b, β-carotene, zeaxanthin, and violaxanthin (Brown and Jeffry 1992), Chlorella vulgaris contains fucoxanthin, zeaxanthin, and lutein (Tibbetts et al., 2015c), and Scenedesmus sp. contains lutein (Tibbetts et al., 2015d). A leading role is undoubtedly played by H. pluvialis which is the main source of astaxanthin, responsible for the pink color of the muscles of salmon (Ambati et al. 2014; Shah et al. 2016). H. pluvialis has been shown to be successful in enhancing the reddish skin coloration of red porgy, Pagrus pagrus (Chatzifotis et al. 2005), rainbow trout, O. mykiss (Sommer et al. 1991, 1992), and also of the penaeid shrimp, L. vannamei (Parisenti et al. 2011). Another source of astaxanthin is represented by the microalgae Chlorella zofingiensis. The effect of the microalgae H. pluvialis and C. zofingiensis on the gonad of the sea urchin Anthocidaris crassispina was studied by Peng et al. (2012) based on the fact that their sexual organs are a highly valued seafood and are considered a prized delicacy in countries such as China. Their results showed that sea urchins fed diets containing astaxanthin pigments revealed higher incorporation of these known antioxidant constituents, with the resultant seafood products therefore being of potential higher nutritive value. Studies have shown that an astaxanthin-enriched diet provided to the bivalve mollusk, lion’s paw scallop (Nodipecten nodosus), and brood stock during conditioning can affect larval survival (Suhnel 2008). A microalgal diet based on a mix of two different microalgae, Isochrysis galbana and Pavlova lutheri, produced an optimization of the growth as well as acceleration in the larval settlement when broodstocks were fed the astaxanthin-enriched diet (Carvalho et al. 2013).
Carotenoids are used as natural feed colorants, additive for feeds, vitamin supplements, and health food products. Dunaliella salina is the most commonly used species for large-scale production because of the high accumulation of carotenoids, up to 14% of dry weight (Borowitzka 2013b). A recent study was assessed on evaluating the physiological condition of L. vannamei shrimp fed diets enriched with Dunaliella sp. containing high levels of carotenes (Medina-Félix et al. 2014). A basal diet containing 35% crude protein and experimental diets which included 1 and 2% microalgae meal, respectively, were evaluated. The results of the study suggest a positive effect of the dietary inclusion (1–2%) of Dunaliella meal on shrimp survival (80%) and an unclear effect on hemolymph metabolites. Similar results were obtained by Boonyaratpalin et al. (2001) with the shrimp Penaeus monodon using D. salina.
Lutein, among the most important carotenoids in foods and human serum, is used as food colorant and especially as feed additives in aquaculture (Lorenz and Cysewski 2000). Arthrospira is used as a source of zeaxanthin and carotene (Miki et al. 1986), and its inclusion can increase fillet color and firmness. Red tilapia fed 30% A. platensis contained more carotenoids than individuals fed less Arthrospira, demonstrating that A. platensis can be exploited as a pigment source for incorporation in Red tilapia diets to produce coloration and gain market acceptance (Ruangsomboon et al. 2010). Watanabe et al. (1990) reported improved taste and texture of Striped jack (Pseudocaranx dentex) related to depressed lipid content in muscle, when supplied with 5% dietary Arthrospira. When Arthrospira was included at 2%, tilapia displayed increased muscle quality, increased firmness, and fibrousness of raw meat (Mustafa and Nakagawa 1995). This effect has been reported for ornamental carp where a test demonstrated that its pigmentation can be modified by supplementing the diet with 7.5% A. platensis (Xiangjun et al. 2012). A similar brightly colored pigmentation has been obtained supplementing Arthrospira as a source of zeaxanthin and myxoxanthophyll in the diet of Fancy carp (Matsuno et al. 1979), Striped jack (Okada et al., 1991), as a source of lutein, rhodoxanthin for Red tilapia (Matsuno et al. 1980), Ayu (Mori et al. 1987), and Black tiger prawn (Liao et al. 1993). Addition of P. tricornutum to the finishing diet for gilthead seabream (S. aurata) enhanced external pigmentation (Ribeiro et al. 2017). Microalgal “green water meal” has also been shown to enhance pigmentation in L. vannamei (Basri et al. 2015).
Overall, these studies suggest that the color and taste of many fish species including tilapia can be improved by supplementing low levels of Arthrospira or other microalgae in the diet. However, further research should be continued to evaluate performances of the various microalgal pigments on commercial aquaculture.
Microalgae as immunostimulants
In recent years, the role of microalgae in aquaculture has extended further with its use as a potential immunostimulant to commercially important aquaculture species. Microalgae can also help to improve survival rate and immune system is shrimp larvae. There are several studies which highlighted the immunostimulating properties of various microalgae in fish and crustaceans. Chlorella could be used as a good additive and could promote the growth performance and physiological parameters and can be involved in regulating adaptive and innate immunity of gibel carp (Carassius auratus gibelio) (Xu et al. 2014; Zhang et al. 2014). Usage of C. vulgaris in the diet (6–8% replacement of fishmeal) of giant freshwater prawn Macrobrachium rosenbergii postlarvae has effectively increased specific growth rate, enhanced immune response (total haemocyte count and prophenol oxidase activity), and enhanced resistance of M. rosenbergii postlarvae against Aeromonas hydrophila infection (Maliwat et al. 2016). Orally administered microalgae (Nannochloropsis gaditana, Tetraselmis chuii, and P. tricornutum) could enhance gilthead seabream (S. aurata) defense activity (Cerezuela et al. 2012). The consumption of Euglena viridis has immunostimulatory effects on freshwater fish Rohu (Labeo rohita) against pathogenic bacterium like A. hydrophila. Fish fed with Euglena had an increased level of lysozyme, serum bactericidal activity, and superoxide anion production (Das et al. 2009). A dietary combination of Navicula sp. and Lactobacillus sakei had considerable effectiveness on intestinal morphology, humoral immune parameters, and weight gain of pacific red snapper (Lutjanus peru) (Reyes-Becerril et al. 2013). A diet of microalgae P. incisa in Guppy fish (Poecilia reticulata) revealed an increase in stress resistance and the survival rate associated with the increase of lysozyme level (Nath et al. 2012). Dunaliella salina strongly enhanced the immunological and antioxidant factors (superoxide dismutase and catalase) and increased the surviving rate of white spot syndrome virus-infected shrimp P. monodon (Madhumathi and Rengasamy 2011). Arthrospira (5–10%) has been reported to increase non-specific immune activity and increase both juvenile and grow-out survival of many species when presented with specific pathogens (Watanuki et al. 2006; Ibrahem et al. 2013).
The inclusion of 10% A. platensis can significantly increase red blood count (RBC), white blood count (WBC), hemoglobin, total protein, and albumin levels of rainbow trout (O. mykiss) so it can be introduced as an immunostimulant in diets (Yeganeh et al. 2015). These previous reports suggest that utilization of microalgal biomass in combination with diet can help to improve the immune system and physiological condition of larval, juvenile, and adult fish and crustacean species. Utilization of microalgae-derived immunostimulants in aquafeed can be beneficial for the development of aquafeed industry throughout the world.
Digestibility of microalgae-based aquafeed
Knowledge on the digestibility of feed ingredients is essential to assess the nutritional value. Determination of the nutrient digestibility of microalgae is the first step to evaluate the feasibility of the utilization of the microalgal product in aquafeeds (Allan et al. 2000; Tibbetts et al. 2006; Guedes and Malcata 2012). Different studies have reported the effects of feeding microalgae to omnivorous fish. Recently, Sarker et al. (2016a) determined apparent digestibility coefficients (ADCs) of macronutrients, amino acids, and fatty acids for freshwater (Arthrospira, Chlorella) and marine (Schizochytrium) microalgal ingredients in Nile tilapia. ADCs of crude protein and all essential amino acids were significantly higher in Arthrospira (86.1%) than in Chlorella (80.0%), compared well with literature values for fishmeal and plant feedstuffs. Schizochytrium had five to ten times richest lipid and omega-3 PUFA contents, significantly highest ADCs for lipid (total PUFA 97.5%), omega-3 (97.2%), and omega-6 PUFA (92.4%) and highest content and digestibility of DHA. They concluded Spirulina as a good alternative protein and Schizochytrium a quality fish oil substitute or LC-PUFA supplement for tilapia diets. Crude protein digestibility for all three microalgae (Arthrospira, Chlorella, and Schizochytrium) (equal or >80%) was similar to the protein digestibility of Anabaena sp. by tilapia (Ekpo and Bender 1989). Ekpo and Bender (1989) also found that tilapia digest blue-green algae more efficiently than silver carp, which are strict phytoplankton feeders. They reported that the protein ADC in tilapia varied between 90.0 and 94.5% in diets with 20–40% Arthrospira inclusion levels. The capacity to digest microalgal protein may be due to the low pH in herbivorous fish stomachs, which allows them to leach nutrients from the cell without breaking its cell wall (Horn and Messer 1992). This process would be very efficient for Nile tilapia considering that its stomach has the lowest pH level (1.2) (Bowen 1976). The ADC of diets containing microalgae has been evaluated for farmed fish. A study on European sea bass (D. labrax) reported that the ADC values of protein, lipid and energy in the diets containing freeze-dried Isochrysis sp. (14%) were 92.6, 87.6, and 85.0, respectively and microalgal diet did not alter the SGR, FCR, and PER of the fish (Tibaldi et al. 2015). The digestibility of protein in fingerling mirror carp and rainbow trout feeds that contained A. maxima (68%) were 87.1 and 83.1, respectively (Atack et al. 1979). In Atlantic salmon, the digestibility of protein and lipid of feeds containing diatom Entomoneis spp. (5%) were 83 and 87, respectively (Norambuena et al. 2015). The protein, lipid, and energy digestibility values of an algal product made from Schizochytrium sp. (20%) in the feeds of rainbow trout were 87.3, 53, and 60, respectively (Zhang et al. 2013). The protein, lipid, and energy digestibility values in the 20% alga-containing (defatted biomass from the marine microalga, Desmodesmus sp.) feeds were 84, 94, and 80, respectively, for Atlantic salmon (S. salar) smolts (Kiron et al. 2016). The energy digestibility of the feeds containing 10% alga was significantly different from that of the control feed. However, the higher inclusion of the alga did not cause a significant reduction in energy digestibility. The results indicate that 20% inclusion level in the feeds of Atlantic salmon does not have any adverse effects on the specific growth rate, condition factor, protein efficiency ratio, and whole body proximate composition of fish. Nutrient digestibility of P. tricornutum in feeding experiment (diets with 0, 3%, and 6% inclusion of algae) with Atlantic salmon (S. salar) showed a linear reduction in apparent digestibility of dry matter, lipid, and protein for the fish (Sørensen et al. 2016). Protein digestibility was estimated to be 90.1, 89.3, and 89.7 for the three diets, respectively. Lipid digestibility was also high, ranging from 96.0, 95.9, and 95.7, respectively, for the three experimental diets. The inclusion rates of P. tricornutum in their study had no effect on nutrient digestibility, growth, and feed conversion of up to 6% inclusion rate. The digestibility of the lipids of the diatom Chaetoceros muelleri in the larvae of L. vannamei has also been studied (Pacheco-Vega et al. 2016). On the contrary, greater ADC was reported for the cyanobacterium Arthrospira fed to Atlantic salmon and Arctic charr at 30% inclusion level in the feed (Burr et al. 2012). Digestibility of protein in Arthrospira was estimated to range between 82 and 84.7% for the two fish species, respectively. These relatively high digestibility coefficients compare favorably with terrestrial plant ingredients, confirming the high potential of Arthrospira as a protein source for farmed fish.
These studies provide evidence that microalgae are suitable sources of protein and lipid for aquaculture species. However, the inclusion of microalgae in feeds should be evaluated at a species level, including a deeper understanding of their digestibility and bioavailability.
Safety and regulatory aspects of microalgae feed applications
The most common microalgae used in aquaculture or for production of food supplements, such as Isochrysis, Chaetoceros gracilis, T. suecica, P. lutheri, Skeletonema costatum, Dunaliella tertiolecta, Nannochloropsis sp., P. tricornutum, and Chlorella sp., do not produce toxins (Enzing et al. 2012). However, it is considered that, even within the same species, large differences exist between toxic and non-toxic microalgae strains. It is widely accepted that dinoflagellates, diatoms, and cyanobacteria can affect humans by producing toxins. Therefore, in view of application of algae for food or feed, it is very important to know their safety at strain level. Even if there are microalgae group (i.e., some dinoflagellates) categorized as potentially harmful, some strains are indeed used as food and feed supplements. For instance, the dinoflagellate C. cohnii has a generally recognized as safe (GRAS) status and is used for commercial production of the omega-3 fatty acid DHA (Mendes et al. 2009; Ryan et al. 2010).
The safety aspect of common microalgae used in aquaculture has been analyzed and reviewed by the two most important authorities, the Food and Drug Administration (FDA) and the European Food Safety Authority (EFSA). On the matter of regulation on marketing of microalgae for food and feed, some differences are found in Europe and the USA. In general, three main European regulations apply to the marketing of microalgae or its components: on food safety (EC regulation 178/2002), on novel food and novel food ingredients (EC regulation 258/97), and on nutrition and health claims for food (EC regulation 1924/2006). Several substances derived from algae are used as food and feed additives. In this case, Regulation (EC) 1333/2008 applies. In the past, prior to market access, a food and feed business operator (FBO) was obliged to apply for authorization for the use of proteins derived from algae in feed under Directive 82/471/EEC. Assessment of safety and nutritional value was done according to the guidelines in Directive 83/228/EEC. To assist applicants, a “Guidance for the assessment of biomasses for use in animal nutrition” was published by the European Food Safety Authority (EFSA) in 2011. Broadly speaking, microalgae are likely to belong to the product group “algae and prokaryotes organism” in the Annex I of Regulation EC 752/2014. Algae or their products used as feed should comply with all the legal maximum levels mentioned in Directive 2002/32/EC. The regulation EU 68/2013 established the Catalog of Feed Material and includes microalgae within the group 7 of the list of feed material, specifically in “other plant, algae, and products derived thereof.” In Europe, the European Food Safety Authority (EFSA) is requested to assess the safety of any new food and feed compound before they are authorized for production and commercialization. EFSA’s scientific panels maintain a list of biological agents to which the concept of qualified presumption of safety (QPS) can be applied. Although no algae are included in the QPS assessment list of 2012, β-carotene from D. salina and DHA from C. cohnii are already approved as food ingredients by EFSA (Melis 2009) and Chlorella and Arthrospira (Spirulina) are commonly sold as food supplements in the EU and elsewhere.
In the USA, two main laws are applicable on microalgae-based food and feed products once they are sold on the consumer market: The Federal Food, Drug, and Cosmetic Act (FDC) introduced in 1938 (FDA 2010), which regulates all foods and food additives, and the Dietary Supplement Health and Education Act (DSHEA) introduced in 1994, which amended the FDC Act to cover dietary ingredients and supplements. In particular, according to FDC, the legal status of a food substance depends on whether it is used in a conventional food, a dietary supplement, or as ingredient in a dietary supplement product. This substance is subject to premarket review and approval, unless the substance is generally recognized, among qualified experts, as safe (GRAS) under the conditions of its intended use. To date, there are seven algae-based GRAS food ingredients that have been reviewed by FDC: microalgal oil derived from Ulkenia sp. SAM2179; H. pluvialis extract containing astaxanthin esters; algal oil from Schizochytrium sp.; Spirulina: the dried biomass of A. platensis; Laminaria japonica broth and extract powder; DHASCO as derived-product from the microalgal species C. cohnii; and “Calcified seaweed” derived from Phymatolithon calcareum or Lithothamnium corrallioides (Enzing et al. 2014).
Current challenges to use microalgae in aquafeed
The high production costs of microalgae remain a constraint to the aquaculture industry. Using microalgae can be economically feasible by reducing its sales price and large-scale production (Becker 2007; Sarker et al. 2016a). The collection, drying and pelletization of microalgae require considerable time and effort. Inappropriate drying may adversely affect their nutritional and physical properties and greatly reduce their value as feeds. The thick cell walls of some microalgae (e.g., Chlorella) can prevent absorption of the nutritional value of the cell contents. The type and quantity of extracellular polysaccharides of certain microalgae (e.g., D. tertiolecta) can also interfere with nutrient absorption. The microalgae which are used as fish feed sometimes may create a problem due to their low digestibility and high amount of salt accumulation in marine microalgal species. Only about 10–15% of dietary protein requirement can be met by algae in test diets without compromising growth and food utilization. Compared with fishmeal, some microalgae may have lower protein content and higher content of carbohydrates, mainly in the poorly digestible cell walls which also impair digestibility of nutrients such as protein and lipid (Skrede et al. 2011).
Depending on growth and processing conditions, microalgae biomass may contain high concentrations of trace elements and toxins that may constrain their incorporation in aquafeeds. It has often been proposed that the protein portion remaining after lipid extraction for biofuel production might be a useful input for animal feeds (Chen et al. 2010). However, the microalgae chosen for biofuel production may not be optimal for use as a feed input, and the economic pressure for the lowest-cost methods of fuel production is likely to result in protein residues with contamination that makes them unfit for use as feed (Hussein et al. 2013). When considering such utilization of algal biomass, it would be better to focus on higher-value product, i.e., aquafeed first, and use any residual compounds for biofuel production, not the other way around. In other words, high-value product first principle should apply (Li et al. 2015). The availability of large amount of microalgal biomass could be an obstacle to the development of aquafeed industry. Due to contamination by grazers, pests, and pathogens, large-scale algal cultivation sustainability can be effected (Hannon et al. 2010), but it is obligatory to achieve successful large-scale cultivation of commercially important microalgae species to supply large quantity of microalgal biomass for aquafeed industry.
Future perspectives and conclusion
Currently, there is a large discrepancy in the global supply and purchase cost of microalgal biomass versus existing commodity animal feedstuffs. Improved new technologies such as specialized mass culture facilities using heterotrophic methods (Jia et al. 2014) and affordable closed photobioreactors for culturing microalgal strains may add value to the microalgal biotechnology agenda of biofuels and high-value biomass extracts through integrated biorefinery (Li et al. 2015). It is expected that significant benefits will return to the aquaculture sector via current biotechnology investment, in the form of more efficient microalgal production systems and greater availability of high-quality microalgal biomass. Techno-economic analysis suggests that, for an algae-to-biofuel process to be profitable, it will be necessary to combine biofuel production with higher-value product (Davis et al. 2011; Borowitzka 2013a; Lu et al. 2016). Recent classical energy and eco-energy analysis showed that industrial algae co-products in aquafeeds could drastically decrease environmental costs in the future when compared with fishmeal and plant protein sources (Draganovic et al. 2013). Thus, different industries are able to generate algal co-product for use in animal feeds and integrating industrial co-products into aquafeed could lower feed prices and provide an additional revenue stream from algae biofuel and nutraceutical production. In order for innovative ways formulated to sustain aquaculture feed to succeed in the market, researchers have to explore the ways to cut the high production cost of microalgae (Walker 2009; Wan et al. 2015). Towards this end, scientists are exploring ways to reduce production costs and the environmental footprint of microalgae production by using organic waste streams as a partial replacement for expensive inputs of inorganic fertilizer normally used to grow microalgae (Alam et al. 2015; Sarker et al. 2016b). Furthermore, ongoing research on the use to lower costs of producing and processing microalgae as for biofuel and human nutraceutical sources may lead to higher market availability and lower price of microalgae for aquafeed in the near future (Chauton et al. 2015; White and Ryan 2015).
Beside the availability of microalgal biomass at sufficient quantity with suitable price, microalgae and aquafeed manufacturers need to take into consideration about several things such as potentially large variations in proximate composition, digestibility encountered among different algal strains and growing conditions, etc. A good selection of microalgal species is needed to support the aquaculture industries. Microalgal species with improved nutritional quality or growth characteristics could improve their efficiencies. The composition of different microalgae should be studied in light of fish nutrition requirements. If a certain microalgal strain has low digestibility levels, that needs effective treatment to disrupt the cell wall and thus make the algal constituents accessible to digestive enzyme (Becker 2004; Berge et al. 2013; Halim et al. 2012) and improve their digestibility. However, additional processing steps may add further cost. To positively affect the health of fish, prior to commercialization, microalgal material must be analyzed for the presence of toxic compounds to prove their harmlessness. In addition to the nutritional value and digestibility aspects, the technological requirements for processing must also be taken into consideration with regards to the industrial production line for compound feed. A review has recently addressed causes for variation in extruded fish feed (Sørensen et al. 2012) and ingredient composition is known to be one of the most important variables affecting physical quality. To our knowledge, recently a single study (Gong et al. 2017) has been published evaluating effects of microalgae on physical quality of extruded fish feed, and more research focus is clearly needed.
Genetic engineering of microalgae research is scarce for application in aquaculture. The insertion of genes or genetic manipulation of microalgae can improve the nutritional quality of microalgae which could eventually increase the quality of fish fed with aquafeed (supplemented with microalgae) (Li and Tsai 2009; Khatoon and Pal 2015). Li and Tsai (2009) inserted an algae-codon-optimized bovine lactoferin (an antimicrobial peptide) into Nannochloropsis oculata to provide an organism against bacterial pathogenic infection. The average survival rate of the medaka fish fed with the transgenic algae was higher (85%). that of those fed with wild type algae (5%). A combined effort to standardize a genetically modified microalgae coupled to a controlled bioprocess system will lead to an upliftment in the status of aquaculture (Hemaiswarya et al. 2011).
In order to establish scientific evidence and to gain a good understanding of how microalgae can best be used in fish feeds, a large number of well-designed feeding trials are still required to evaluate potential of microalgae as a replacement of fishmeal and fishoil. However, it already seems that microalgae will play an important role in the effort to move the formulation of aquafeed to a more sustainable future. We may conclude further that available microalgal biomass with better composition and cheaper price will allow producing more available and reliable aquafeed and this will free the aquaculture industry to keep growing and meet current and future demands.
References
Alam MA, Wan C, Zhao XQ, Chen LJ, Chang JS, Bai FW (2015) Enhanced removal of Zn2+ or Cd2+ by the flocculating Chlorella vulgaris JSC-7. J Hazard Mater 289:38–45
Allan GL, Parkinson S, Booth MA, Stone DA, Rowland SJ, Frances J, Warner-Smith R (2000) Replacement of fishmeal in diets for Australian silver perch, Bidyanus bidyanus: I. Digestibility of alternative ingredients. Aquaculture 186:293–310
Ambati RR, Phang SM, Ravi S, Aswathanarayana RG (2014) Astaxanthin: sources, extraction, stability, biological activities and its commercial applications—a review. Mar Drugs 12:128–152
Atack T, Jauncey K, Matty A (1979) The utilization of some single cell proteins by fingerling mirror carp (Cyprinus carpio). Aquaculture 18:337–348
Atalah E, Cruz CMH, Izquierdo MS, Rosenlund G, Caballero MJ, Valencia A, Robaina L (2007) Two microalgae Crypthecodinium cohnii and Phaeodactylum tricornutum as alternative source of essential fatty acids in starter feeds for seabream (Sparus aurata). Aquaculture 270:178–185
Badwy TM, Ibrahim EM, Zeinhom MM (2008) Partial replacement of fishmeal with dried microalgae (Chlorella spp. and Scenedesmus spp.) in Nile tilapia (Oreochromis niloticus) diets. In: Elghobashy H, Fitzsimmons K, Diab AS (eds) From the pharaohs to the future: proceedings of the 8th international symposium on tilapia in aquaculture. Egypt Ministry of Agriculture, Cairo, pp 801–810
Barclay W, Zeller S (1996) Nutritional enhancement of n-3 and n-6 fatty acids in rotifers and Artemia nauplii by feeding spray-dried Schizochytrium sp. J World Aquacult Soc 27:314–322
Basri NA, Shaleh SRM, Matanjun P, Noor NM, Shapawi R (2015) The potential of microalgae meal as an ingredient in the diets of early juvenile Pacific white shrimp, Litopenaeus vannamei. J Appl Phycol 27:857–863
Becker E (2007) Micro-algae as a source of protein. Biotechnol Adv 25:207–210
Becker EW (1994) Microalgae: biotechnology and microbiology. Cambridge University Press, Cambridge, 293p
Becker W (2004) Microalgae in human and animal nutrition. In: Richmond A (ed) Handbook of microalgal culture: biotechnology and applied phycology. Blackwell Science Ltd, Cambridge, pp 312–351
Benemann J (2013). Microalgae for biofules and animal feeds. Energies 6:5869–5886
Berge G, Hatlen B, Odom J, Ruyter B (2013) Physical treatment of high EPA Yarrowia lipolytica biomass increases the availability of n- 3 highly unsaturated fatty acids when fed to Atlantic salmon. Aquac Nutr 19:110–121
Bigogno C, Khozin-Goldberg I, Boussiba S, Vonshak A, Cohen Z (2002) Lipid and fatty acid composition of the green oleaginous alga Parietochloris incisa, the richest plant source of arachidonic acid. Phytochemistry 60:497–503
Blazencic J (2007) Sistematika algi. NNK Internacional, Beograd
Boonyaratpalin M, Thongrod S, Supamattaya K, Britton G, Schlipalius LE (2001) Effects of ß-carotene source, Dunaliella salina, and astaxanthin on pigmentation, growth, survival and health of Penaeus monodon. Aquac Res 32(Suppl 1):182–190
Borowitzka MA (2013a) High-value products from microalgae-their development and commercialisation. J Appl Phycol 25:743–756
Borowitzka MA (2013b) Dunaliella: biology, production, and markets. In: Richmond A, Hu Q (eds) Handbook of microalgal culture. John Wiley & Sons, Ltd, pp 359–368
Bowen SH (1976). Feeding ecology of the cichlid fish Sarotherodon mossambicus in Lake Sibaya, KwaZulu. Doctoral dissertation, Rhodes University, Grahamstown, South Africa
Brown M, Jeffrey S, Volkman J, Dunstan G (1997) Nutritional properties of microalgae for mariculture. Aquaculture 151:315–331
Brown MR (2002) Nutritional value and use of microalgae in aquaculture. In: Avances en Nutrición Acuícola VI. Memorias del VI Simposium Internacional de Nutrición Acuícola, vol 3, pp 281–292
Brown MR, Jeffry SW (1992) Biochemical composition of microalgae from the green algal classes Chlorophyceae and Prasinophyceae. 1. Amino acids, sugars and pigments. J Exp Mar Biol Ecol 161:91–113
Burr GS, Wolters WR, Barrows FT, Hardy RW (2012) Replacing fishmeal with blends of alternative proteins on growth performance of rainbow trout (Oncorhynchus mykiss), and early or late stage juvenile Atlantic salmon (Salmo salar). Aquaculture 334:110–116
Carter CG, Bransden MP, Lewis TE, Nichols PD (2003) Potential of Thraustochytrids to partially replace fish oil in Atlantic salmon feeds. Mar Biotechnol 5:480–492
Carvalho YBM, Ferreira JF, Da Silva FC, Bercht M (2013) Factors influencing larval settlement of the Atlantic lion's paw scallop, Nodipecten nodosus. J Shellfish Res 32:719–723
Cerezuela R, Guardiola FA, Meseguer J, Esteban MA (2012) Enrichment of gilthead seabream (Sparus aurata L.) diet with microalgae: effects on the immune system. Fish Physiol Biochem 38:1729–1739
Ceron-Garcia M, Fernandez-Sevilla J, Sanchez-Miron A, Garcia-Camacho F, Contreras-Gomez A, Molina-Grima E (2013) Mixotrophic growth of Phaeodactylum tricornutum on fructose and glycerol in fed-batch and semi-continuous modes. Bioresour Technol 147:569–576
Chatzifotis S, Pavlidis M, Jimeno CD, Vardanis G, Sterioti A, Divanach P (2005) The effect of different carotenoid sources on skin coloration of cultured red porgy (Pagrus pagrus). Aquac Res 36:1517–1525
Chauton MS, Reitan KI, Norsker NH, Tveterås R, Kleivdal HT (2015) A techno-economic analysis of industrial production of marine microalgae as a source of EPA and DHA-rich raw material for aquafeed: research challenges and possibilities. Aquaculture 436:95–103
Chen CY, Chen YC, Huang HC, Ho SH, Chang JS (2015) Enhancing the production of eicosapentaenoic acid (EPA) from Nannochloropsis oceanica CY2 using innovative photobioreactors with optimal light source arrangements. Bioresour Technol 191:407–413
Chen S, Chi Z, O’Fallon JV, Zheng Y, Chakraborty M, Laskar DD (2010) System integration for producing microalgae as biofuel feedstock. Biofuels 1:889–910
Dallaire V, Lessard P, Vandenberg G, de la Noüe J (2007) Effect of algal incorporation on growth, survival and carcass composition of rainbow trout (Oncorhynchus mykiss) fry. Bioresour Technol 98:1433–1439
Daroch M, Shao C, Liu Y, Geng S, Cheng JJ (2013) Induction of lipids and resultant FAME profiles of microalgae from coastal waters of Pearl River Delta. Bioresour Technol 146:192–199
Das BK, Pradhan J, Sahu S (2009) The effect of Euglena viridis on immune response of rohu, Labeo rohita (Ham.) Fish Shellfish Immunol 26:871–876
Davis R, Aden A, Pienkos PT (2011) Techno-economic analysis of autotrophic microalgae for fuel production. Appl Energ 88:3524–3531
Draganovic V, Jørgensen SE, Boom R, Riese G, van der Goot AJ (2013) Sustainability assessment of salmonid feed using energy, classical exergy and eco-exergy analysis. Ecol Indic 34:277–289
Ekpo I, Bender J (1989) Digestibility of a commercial fish feed, wet algae, and dried algae by Tilapia nilotica and silver carp. Prog Fish Cult 51:83–86
El-Sayed AFM (1994) Evaluation of soybean meal, Spirulina meal and chicken offal meal as protein sources for silver seabream (Rhabdosargus sarba) fingerlings. Aquaculture 127:169–176
Enzing C, Ploeg M, Barbosa M, Sijtsma L (2014) Microalgae-based products for the food and feed sector: an outlook for Europe. In: Vigani M, Parisi C, Rodríguez Cerezo E (eds). JRC Scientific and Policy Reports, EU Publications: Luxembourg. 82p
Enzing, CM, Nooijen A, Eggink G, Springer J, Wijffels R (2012) Algae and genetic modification. Research, production and risks. http://www.cogem.net/index. cfm/nl/publicaties/publicatie/onderzoeksrapport-algae-andgenetic-modification-research-production-and-risks
FAO (2007) The state of world fisheries and aquaculture. FAO, Rome. (http://www.fao.org/docrep/fao/009/a0699e/a0699e.pdf )
FAO (2012) The state of world fisheries and aquaculture. FAO, Rome. (http://www.fao.org/docrep/016/i2727e/i2727e.pdf)
FAO (2014) The state of world fisheries and aquaculture: opportunities and challenges. FAO, Rome. (http://www.fao.org/3/a-i3720e.pdf )
FAO (2016) The state of world fisheries and aquaculture 2016. Contributing to food security and nutrition for all. Rome. 200 pp. (http://www.fao.org/3/a-i5555e.pdf )
FDA (2010) http://www.fda.gov/regulatoryinformation/legislation/federal food drug and cosmetic act fdcact/default. html)
Ganuza E, Benítez-Santana T, Atalah E, Vega-Orellana O, Ganga R, Izquierdo M (2008) Crypthecodinium cohnii and Schizochytrium sp. as potential substitutes to fisheries-derived oils from seabream (Sparus aurata) microdiets. Aquaculture 277:109–116
Gong Y, Guterres HADS, Huntley M, Sørensen M, Kiron V (2017) Digestibility of the defatted microalgae Nannochloropsis sp. and Desmodesmus sp. when fed to Atlantic salmon, Salmo salar. Aquacult Nutr doi:10.1111/anu.12533
Guedes AC, Malcata FX (2012) Nutritional value and uses of microalgae in aquaculture. In: Muchlisin Z (ed) Aquaculture. INTECH, Riejeka, pp 59–78
Guschina IA, Harwood JL (2013) Algal lipids and their metabolism. In: Borowitzka MA, Moheimani NR (eds) Algae for biofuels and energy. Springer, Dordrecht, pp 17–36
Haas S, Bauer JL, Adakli A, Meyer S, Lippemeier S, Schwarz K, Schulz C (2016) Marine microalgae Pavlova viridis and Nannochloropsis sp. as n-3 PUFA source in diets for juvenile European sea bass (Dicentrarchus labrax L.) J Appl Phycol 28: 1011
Hajiahmadian M, Vajargah MF, Farsani HG, Chorchi MM (2012) Effect of Spirulina platensis meal as feed additive on growth performance and survival rate in golden barb fish, Punius gelius (Hamilton, 1822). J Fish Int 7:61–64
Halim R, Danquah MK, Webley PA (2012) Extraction of oil from microalgae for biodiesel production: a review. Biotechnol Adv 30:709–732
Hannon M, Gimpel J, Tran M, Rasala B, Mayfield S (2010) Biofuels from algae: challenges and potential. Biofuels 1:763–784
Hemaiswarya S, Raja R, Kumar RR, Ganesan V, Anbazhagan C (2011) Microalgae: a sustainable feed source for aquaculture. World J Microb Biot 27:1737–1746
Horn MH, Messer KS (1992) Fish guts as a chemical reactor: a model of the alimentary canals of marine herbivorous. Mar Biol 113:527–535
Hu Q, Sommerfeld M, Jarvis E, Ghirardi M, Posewitz M, Seibert M, Darzins A (2008) Microalgal triacylglycerols as feedstocks for biofuel production: perspectives and advances. Plant J 54:621–639
Hussein EES, Dabrowski K, El-Saidy DM, Lee BJ (2013) Enhancing the growth of Nile tilapia larvae/juveniles by replacing plant (gluten) protein with algae protein. Aquaculture Res 44:937–949
Ibrahem M, Mohamed MF, Ibrahim MA (2013) The role of Spirulina platensis (Arthrospira platensis) in growth and immunity of Nile tilapia (Oreochromis niloticus) and its resistance to bacterial infection. J Agr Sci 5:109–117
IFFO (2016) Fishmeal and fish oil statistical yearbook http://www.seafish.org/media/publications/SeafishFishmealandFishOilFactsandFigures_201612.pdf
Jia Z, Liu Y, Daroch M, Geng S, Cheng JJ (2014) Screening, growth medium optimisation and heterotrophic cultivation of microalgae for biodiesel production. Appl Biochem Biotech 173:1667–1679
Jo MJ, Hur SB (2015) Growth and nutritional composition of Eustigmatophyceae Monodus subterraneus and Nannochloropsis oceanica in autotrophic and mixotrophic culture. Ocean Polar Res 37:61–71
Ju ZY, Deng DF, Dominy W (2012) A defatted microalgae (Haematococcus pluvialis) meal as a protein ingredient to partially replace fishmeal in diets of Pacific white shrimp (Litopenaeus vannamei, Boone, 1931). Aquaculture 354:50–55
Kent M, Welladsen HM, Mangott A, Li Y (2015) Nutritional evaluation of Australian microalgae as potential human health supplements. PLoS One 10:e0118985
Khatoon N, Pal R (2015) Microalgae in biotechnological application: a commercial approach. In: Bahadur B, Rajam MV, Sahijram L, Krishanamurthy MV (eds) Plant biology and biotechnology. Springer, New Delhi, pp 27–47
Kim SS, Rahimnejad S, Kim KW, Lee KJ (2013) Partial replacement of fishmeal with Spirulina pacifica in diets for parrot fish (Oplegnathus fasciatus). Turkish J Fish Aquat Sci 13:197–204
Kiron V, Phromkunthong W, Huntley M, Archibald I, Scheemaker GD (2012) Marine microalgae from biorefinery as a potential feed protein source for Atlantic salmon, common carp and whiteleg shrimp. Aquaculture Nutr 18:521–531
Kiron V, Sørensen M, Huntley M, Vasanth GK, Gong Y, Dahle D, Palihawadana AM (2016) Defatted biomass of the microalga, Desmodesmus sp., can replace fishmeal in the feeds for Atlantic salmon. Front Mar Sci 3: 67
Kitajima C (1983) Actual examples of mass cultures. In: Koseikaku K (ed) The rotifer Brachionus plicatilis—biology and mass culture. Japanese Fisheries Society, Tokyo, pp 102–128
Kousoulaki K, Østbye TKK, Krasnov A, Torgersen JS, Mørkøre T, Sweetman J (2015) Metabolism, health and fillet nutritional quality in Atlantic salmon (Salmo salar) fed diets containing n-3-rich microalgae. J Nutr Sci 4:e24
Lang I, Hodac L, Friedl T, Feussner I (2011) Fatty acid profiles and their distribution patterns in microalgae: a comprehensive analysis of more than 2000 strains from the SAG culture collection. BMC Plant Biol 11:1–16
Li J, Liu Y, Cheng JJ, Mos M, Daroch M (2015) Biological potential of microalgae in China for biorefinery-based production of biofuels and high value compounds. New Biotech 32:588–596
Li MH, Robinson EH, Tucker CS, Manning BB, Khoo L (2009a) Effects of dried algae Schizochytrium sp., a rich source of docosahexaenoic acid, on growth, fatty acid composition, and sensory quality of channel catfish Ictalurus punctatus. Aquaculture 292:232–236
Li P, Mai K, Trushenski J, Wu G (2009b) New developments in fish amino acid nutrition: towards functional and environmentally oriented aquafeeds. Amino Acids 37:43–53
Li SS, Tsai HJ (2009) Transgenic microalgae as a non-antibiotic bactericide producer to defend against bacterial pathogen infection in the fish digestive tract. Fish Shellfish Immunol 26:316–325
Liao WL, Nur-E-Borhan SA, Okada S, Matsui T, Yamaguchi K (1993) Pigmentation of cultured black tiger prawn by feeding with a Spirulina-supplemented diet. Nippon Suisan Gakk 59:165–169
Liu Y, Tang J, Li J, Daroch M, Cheng JJ (2014) Efficient production of triacylglycerols rich in docosahexaenoic acid (DHA) by osmo-heterotrophic marine protists. Appl Microbiol Biotechnol 98:9643
Lu W, Alam MA, Pan Y, Wu J, Wang ZM, Yuan ZH (2016) A new approach of microalgal biomass pretreatment using deep eutectic solvents for enhanced lipid recovery for biodiesel production. Bioresour Technol 218:123–128
Madhumathi M, Rengasamy R (2011) Antioxidant status of Penaeus monodon fed with Dunaliella salina supplemented diet and resistance against WSSV. Int J Eng Sci Tech 3:7249–7259
Maliwat GC, Velasquez S, Robil JL, Chan M, Traifalgar RF, Tayamen M, Ragaza JA (2017) Growth and immune response of giant freshwater prawn Macrobrachium rosenbergii (De Man) postlarvae fed diets containing Chlorella vulgaris (Beijerinck). Aquac Res 48:1666–1676
Matsuno TKM, Iwahashi M, Koike T, Okada M (1980) Intensification of color of red tilapia with lutein, rhodoxanthin and Spirulina. Bull Chem Soc Jap 46:479–482
Matsuno TNS, Iwahashi M, Koike T, Okada M (1979) Intensification of color of fancy red carp with zeaxanthin and myxoxanthophyll, major carotenoid constituents of Spirulina. Bull Chem Soc Jap 45:627–632
Medina-Félix D, López-Elías JA, Martínez-Córdova LR, López-Torres MA, Hernández-López J, Rivas-Vega ME, Mendoza-Cano F (2014) Evaluation of the productive and physiological responses of Litopenaeus vannamei infected with WSSV and fed diets enriched with Dunaliella sp. J Invertebr Pathol 117:9–12
Melis A (2009) Solar energy conversion efficiencies in photosynthesis: minimizing the chlorophyll antennae to maximize efficiency. Plant Sci 177:272–280
Mendes A, Reis A, Vasconcelos R, Guerra P, Lopes da Silva T (2009) Crypthecodinium cohnii with emphasis on DHA production: a review. J Appl Phycol 21:199–214
Miki W, Yamaguchi K, Konosu S (1986) Carotenoid composition of Spirulina maxima. Nippon Suisan Gakk 52:1225–1227
Miller MR, Nichols PD, Carter CG (2007) Replacement of dietary fish oil for Atlantic salmon parr (Salmo salar L.) with a stearidonic acid containing oil has no effect on omega-3 long-chain polyunsaturated fatty acid concentrations. Comp Biochem Physiol B 146:197–206
Mori TMT, Miki W, Yamaguchi K, Konosu S, Watanabe T (1987) Pigmentation of cultured sweet smelt fed diets supplemented with a blue-green alga Spirulina maxima. Nippon Suisan Gakk 53:433–438
Mustafa MG, Nakagawa H (1995) A review: dietary benefits of algae as an additive in fish feed. Isr J Aquacult 47:155–162
Namaei Kohal M, Esmaeili Fereidouni A, Firouzbakhsh F, Hayati I (2017) Effects of dietary incorporation of Arthrospira (Spirulina) platensis meal on growth, survival, body composition, and reproductive performance of red cherry shrimp Neocaridina davidi (Crustacea, Atyidae) over successive spawnings. J Appl Phycol. doi:10.1007/s10811-017-1220-5
Nasopoulou C, Zabetakis I (2012) Benefits of fish oil replacement by plant originated oils in compounded fish feeds. A review. LWT-Food Sci Technol 47:217–224
Nath P, Khozin-Goldberg I, Cohen Z, Boussiba S, Zilberg D (2012) Dietary supplementation with the microalgae Parietochloris incisa increases survival and stress resistance in guppy (Poecilia reticulata) fry. Aquaculture Nutr 18:167–180
Norambuena F, Hermon K, Skrzypczyk V, Emery JA, Sharon Y, Beard A, Turchini GM (2015) Algae in fish feed: performances and fatty acid metabolism in juvenile Atlantic salmon. PLoS One 10:e0124042
Origin Oil (2014) Origin Oil-White-Paper-Algae-As-Aquafeed (http://www.originclear.com/pdf/OriginOil-White-Paper-Algae-As-Aquafeed.pdf)
Okada SLW, Mori T, Yamaguchi K, Watanabe T (1991) Pigmentation of cultured stripedjack reared on diets supplemented with the blue-green alga Spirulina maxima. Nippon Suisan Gakk 57:1403–1406
Olvera-Novoa MA, Dominguez-Cen LJ, Olivera-Castillo L, Martínez-Palacios CA (1998) Effect of the use of the microalgae Spirulina maxima as fishmeal replacement in diets for tilapia, Oreochromis mossambicus (Peters), fry. Aquac Res 29:709–715
Pacheco-Vega JM, Sánchez-Saavedra MP, Cadena-Roa MA, Tovar-Ramírez D (2016) Lipid digestibility and performance index of Litopenaeus vannamei fed with Chaetoceros muelleri cultured in two different enriched media. J Appl Phycol 28:2379–2385
Parisenti J, Beirão L, Maraschin M, Mourino J, Do Nascimento Vieira F, Bedin L, Rodrigues E (2011) Pigmentation and carotenoid content of shrimp fed with Haematococcus pluvialis and soy lecithin. Aquac Nutr 17:e530–e535
Patterson D, Delbert M, Gatlin DM (2013) Evaluation of whole and lipid-extracted algae meals in te diets of juvenile red drum (Sciaenops ocellatus). Aquaculture 416-417:92–98
Peng J, Yuan JP, Wang JH (2012) Effect of diets supplemented with different sources of astaxanthin on the gonad of the sea urchin Anthocidaris crassispina. Nutrients 4:922–934
Pike IH, Jackson A (2010) Fish oil: production and use now and in the future. Lipid Technol 22:59–61
Priyadarshani I, Rath B (2012) Commercial and industrial application of microalgae—a review. J Algal Biomass Utiln 3:89–100
Qiao H, Cong C, Sun C, Li B, Wang J, Zhang L (2016) Effect of culture conditions on growth, fatty acid composition and DHA/EPA ratio of Phaeodactylum tricornutum. Aquaculture 452:311–317
Qiao H, Wang H, Song Z, Ma J, Li B, Liu X, Zhang S, Wang J, Zhang L (2014) Effects of dietary fish oil replacement by microalgae raw materials on growth performance, body composition and fatty acid profile of juvenile olive flounder, Paralichthys olivaceus. Aquaculture Nutr 20:646–653
Reitan KI, Rainuzzo JR, Øie G, Olsen Y (1997) A review of the nutritional effects of algae in marine fish larvae. Aquaculture 155:207–221
Ren LJ, Ji XJ, Huang H, Qu L, Feng Y, Tong QQ, Ouyang PK (2010) Development of a stepwise aeration control strategy for efficient docosahexaenoic acid production by Schizochytrium sp. Appl Microbiol Biotechnol 87:1649–1656
Reyes-Becerril M, Guardiola F, Rojas M, Ascencio-Valle F, Esteban MÁ (2013) Dietary administration of microalgae Navicula sp. affects immune status and gene expression of gilthead seabream (Sparus aurata). Fish Shellfish Immunol 35:883–889
Ribeiro AR, Gonçalves A, Barbeiro M, Bandarra N, Nunes ML, Carvalho ML, Silva J, Navalho J, Dinis MT, Silva T, Dias J (2017) Phaeodactylum tricornutum in finishing diets for gilthead seabream: effects on skin pigmentation, sensory properties and nutritional value. J Appl Phycol 29:1945–1956
Rincón DD, Velásquez HA, Dávila MJ, Semprun AM, Morales ED, Hernández JL (2012) Substitution levels of fishmeal by Arthrospira (= Spirulina) maxima meal in experimental diets for red tilapia fingerlings (Oreochromis sp.). Rev Colomb Cienc Pec 25:430–437
Roy SS, Pal R (2014) Microalgae in aquaculture: a review with special references to nutritional value and fish dietetics. Proc Zool Soc 68:1–8
Ruangsomboon S, Choochote S, Taveekijakarn P (2010) Growth performance and nutritional composition of red tilapia (Oreochromis niloticus x O. mossambicus) fed diets containing raw Spirulina platensis. The International Conference on Sustainable Community Development 2010. 21–23 January 2010. Khon Kaen University, Nongkhai Campus, Thailand and Vientiane, Lao PDR
Ryan AS, Zeller S, Nelson EB (2010) Safety evaluation of single cell oils and the regulatory requirements for use as a food ingredient. In: Cohen Z, Ratledge C (eds) Single cell oils: microbial and algal oils. AOCS Publishing, Urbana, pp 317–350
Ryckebosch E, Bruneel C, Termote-Verhalle R, Goiris K, Muylaert K, Foubert I (2014) Nutritional evaluation of microalgae oils rich in omega-3 long chain polyunsaturated fatty acids as an alternative for fish oil. Food Chem 160:393–400
Sarker P, Gamble M, Kelson S, Kapuscinski A (2016a) Nile tilapia (Oreochromis niloticus) show high digestibility of lipid and fatty acids from marine Schizochytrium sp. and of protein and essential amino acids from freshwater Spirulina sp. feed ingredients. Aquac Nutr 22:109–119
Sarker PK, Kapuscinski AR, Lanois AJ, Livesey ED, Bernhard KP, Coley ML (2016b) Towards sustainable aquafeeds: complete substitution of fish oil with marine microalga Schizochytrium sp. improves growth and fatty acid deposition in juvenile Nile tilapia (Oreochromis niloticus). PloS One 11:e0156684
Shah MMR, Liang Y, Cheng JJ, Daroch M (2016) Astaxanthin-producing green microalga Haematococcus pluvialis: from single cell to high value commercial products. Front Plant Sci 7:531
Sharma K, Schenk PM (2015) Rapid induction of omega-3 fatty acids (EPA) in Nannochloropsis sp. by UV- C radiation. Biotechnol Bioeng 112:1243–1249
Sharma SM, Panta K (2012) Use of Spirulina cultured in industrial effluents as a feed supplement on Sarotherodon mossambicus. Sonsik J 4:39–44
Shields RJ, Lupatsch I (2012) Algae for aquaculture and animal feeds. Anim Sci 21:23–37
Skrede A, Mydland L, Ahlstrøm Ø, Reitan K, Gislerød H, Øverland M (2011) Evaluation of microalgae as sources of digestible nutrients for monogastric animals. J Anim Feed Sci 20:131–142
Sommer TR, D'Souza FML, Morrissy NM (1992) Pigmentation of adult rainbow trout, Oncorhynchus mykiss, using the green alga Haematococcus pluvialis. Aquaculture 106:63–74
Sommer TR, Potts WT, Morrissy NM (1991) Utilization of microalgal astaxanthin by rainbow trout (Oncorhynchus mykiss). Aquaculture 94:79–88
Sørensen I, Rose JK, Doyle JJ, Domozych DS, Willats WG (2012) The Charophycean green algae as model systems to study plant cell walls and other evolutionary adaptations that gave rise to land plants. Plant Signal Behav 7:1–3
Sørensen M, Berge GM, Reitan KI, Ruyter B (2016) Microalga Phaeodactylum tricornutum in feed for Atlantic salmon (Salmo salar)—effect on nutrient digestibility, growth and utilization of feed. Aquaculture 460:116–123
Sprague M, Dick JR, Tocher DR (2016) Impact of sustainable feeds on omega-3 long-chain fatty acid levels in farmed Atlantic salmon, 2006–2015. Sci Rep 6:21892
Suhnel S (2008) Utilizaçao de diferentes dietas em reprodutores davieira Nodipecten nodosus (L.) em laboratorio e seu efeito na maturaçao, no rendimento larval e na produçao de pre-sementes. PhD thesis, Universidade Federal de Santa Catarina, 155p
Sydney EB, Sturm W, de Carvalho JC, Thomaz-Soccol V, Larroche C, Pandey A, Soccol CR (2010) Potential carbon dioxide fixation by industrially important microalgae. Bioresour Technol 101:5892–5896
Tacon AGJ, Metian M (2015) Feed matters: satisfying the feed demand of aquaculture. Rev. Fish Sci Aqua 23:1–10
Teimouri M, Amirkolaie AK, Yeganeh S (2013) The effects of dietary supplement of Spirulina platensis on blood carotenoid concentration and fillet color stability in rainbow trout (Oncorhynchus mykiss). Aquaculture 415:224–228
Thompson GA (1996) Lipids and membrane function in green algae. Lipids Lipid Metab 1302:17–45
Tibaldi E, Zittelli GC, Parisi G, Bruno M, Giorgi G, Tulli F, Venturini S, Tredici M, Poli B (2015) Growth performance and quality traits of European sea bass (D. labrax) fed diets including increasing levels of freeze-dried Isochrysis sp.(T-ISO) biomass as a source of protein and n-3 long chain PUFA in partial substitution of fish derivatives. Aquaculture 440:60–68
Tibbetts SM, Bjornsson WJ, McGinn PJ (2015a) Biochemical composition and amino acid profiles of Nannochloropsis granulata algal biomass before and after supercritical fluid CO2 extraction at two processing temperatures. Animal Feed Sci Technol 204:62–71
Tibbetts SM, Melanson RJ, Park KC, Banskota AH, Stefanova R, McGinn PJ (2015b) Nutritional evaluation of whole and lipid-extracted biomass of the microalga Scenedesmus sp. AMDD isolated in Saskatchewan, Canada for animal feeds: proximate, amino acid, fatty acid, carotenoid and elemental composition. Current Biotech 4:530–546
Tibbetts SM, Milley JE, Lall SP (2006) Apparent protein and energy digestibility of common and alternative feed ingredients by Atlantic cod, Gadus morhua (Linnaeus, 1758). Aquaculture 261:1314–1327
Tibbetts SM, Milley JE, Lall SP (2015c) Chemical composition and nutritional properties of freshwater and marine microalgal biomass cultured in photobioreactors. J Appl Phycol 27:1109–1119
Tibbetts SM, Whitney CG, MacPherson MJ, Bhatti S, Banskota AH, Stefanova R, McGinn PJ (2015d) Biochemical characterization of microalgal biomass from freshwater species isolated in Alberta, Canada for animal feed applications. Algal Res 11:435–447
Lorenz RT, Cysewski GR (2000) Commercial potential for Haematococcus microalgae as a natural source of astaxanthin. Trends Biotechnol 18:160–167
Tulli F, Chini Zittelli G, Giorgi G, Poli BM, Tibaldi E, Tredici MR (2012) Effect of the inclusion of dried Tetraselmis suecica on growth, feed utilization, and fillet composition of european sea bass juveniles fed organic diets. J Aquat Food Prod T 21:188–197
Turchini GM, Torstensen BE, Ng WK (2009) Fish oil replacement in finfish nutrition. Rev Aquacult 1:10–57
Velasquez SF, Chan MA, Abisado RG, Traifalgar RFM, Tayamen MM, Maliwat GCF, Ragaza JA (2016) Dietary Spirulina (Arthrospira platensis) replacement enhances performance of juvenile Nile tilapia (Oreochromis niloticus). J Appl Phycol 28:1023–1030
Vizcaíno A, López G, Sáez M, Jiménez J, Barros A, Hidalgo L, Camacho-Rodríguez J, Martínez T, Cerón-García M, Alarcón F (2014) Effects of the microalga Scenedesmus almeriensis as fishmeal alternative in diets for gilthead sea bream, Sparus aurata, juveniles. Aquaculture 431:34–43
Vizcaíno AJ, Saéz MI, López G, Arizcun M, Abellán E, Martínez TF, Cerón-García MC, Alarcón FJ (2016) Tetraselmis suecia and Tisochrysis lutea meal as dietary ingredients for gilthead sea bream (Sparus aurata L.) fry. J Appl Phycol 28:2843–2855
Walker AB, Berlinsky DL (2011) Effects of partial replacement of fishmeal protein by microalgae on growth, feed intake, and body composition of Atlantic cod. N Am J Aquacult 73:76–83
Walker DA (2009) Biofuels, facts, fantasy, and feasibility. J Appl Phycol 21:509–517
Wan C, Alam MA, Zhao XQ, Zhang XY, Guo SL, Ho SH, Chang JS, Bai FW (2015) Current progress and future prospect of microalgal biomass harvest using various flocculation technologies. Bioresour Technol 184:251–257
Wang Y, Li M, Filer K, Xue Y, Ai Q, Mai K (2016) Evaluation of Schizochytrium meal in microdiets of Pacific white shrimp (Litopenaeus vannamei) larvae. Aquac Res:1–9. doi:10.1111/are.13068
Watanabe T, Liao W, Takeuchi T, Yamamoto H (1990) Effect of dietary Spirulina supplement on growth performance and flesh lipids of cultured striped jack. J Tokyo Univ Fish 77:231–239
Watanuki H, Ota K, Tassakka AR, Sakai M, Kato T, Sakai M (2006) Immunostimulant effects of dietary Spirulina platensis on carp, Cyprinus carpio. Aquaculture 258:157–163
White RL, Ryan RA (2015) Long-term cultivation of algae in open-raceway ponds: essons from the field. Ind Biotechnol 11:213–220
Xiangjun S, Chang Y, Ye Y, Ma Z, Liang Y, Li T, Jiang N, Xing W, Luo L (2012) The effect of dietary pigments on the coloration of Japanese ornamental carp (koi, Cyprinus carpio L.) Aquac Nutr 342–343:62–68
Xu W, Gao Z, Qi Z, Qiu M, Peng JQ, Shao R (2014) Effect of dietary Chlorella on the growth performance and physiological parameters of gibel carp, Carassius auratus gibelio. Turkish J FishAquat Sci 14:53–57
Yaakob Z, Ali E, Zainal A, Mohamad M, Takriff MS (2014) An overview: biomolecules from microalgae for animal feed and aquaculture. J Biol Res Thessalon 21:6
Yeganeh S, Teimouri M, Amirkolaie AK (2015) Dietary effects of Spirulina platensis on hematological and serum biochemical parameters of rainbow trout (Oncorhynchus mykiss). Res Vet Sci 101:84–88
Zhang CN, Li XF, Xu WN, Jiang GZ, Lu KL, Wang LN, Liu WB (2013) Combined effects of dietary fructooligosaccharide and Bacillus licheniformis on innate immunity, antioxidant capability and disease resistance of triangular bream (Megalobrama terminalis). Fish Shellfish Immunol 35:1380–1386
Zhang Q, Qiu M, Xu W, Gao Z, Shao R, Qi Z (2014) Effects of dietary administration of Chlorella on the immune status of gibel carp, Carassius auratus gibelio. Ital J Anim Sci 13:3168
Acknowledgments
The authors would like to acknowledge the support of the National Natural Science Foundation of China for Young International Scientists Grant No. 31550110497), Shenzhen Municipal Government for Special Innovation Fund for Shenzhen Overseas High-level Personnel KQCX20140521150255300, Shenzhen Knowledge and Innovation Basic Research Grant JCYJ20160122151433832, and State Ocean Administration Grant 201305022.
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Contributions
MS collected data, participated in preparation of the manuscript draft, and participated in assembly and editing of the final manuscript; GAL, MAA, AP, and YML collected data and participated in the preparation of the manuscript draft; PKS and KC participated in assembly and editing of the final manuscript; MD participated in the preparation of the manuscript draft and participated in the assembly and editing of the final manuscript.
Corresponding author
Rights and permissions
About this article

Cite this article
Shah, M.R., Lutzu, G.A., Alam, A. et al. Microalgae in aquafeeds for a sustainable aquaculture industry. J Appl Phycol 30, 197–213 (2018). https://doi.org/10.1007/s10811-017-1234-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10811-017-1234-z