
Abstract
Biophysical signaling, an integral regulator of long-term cell behavior in both excitable and non-excitable cell types, offers enormous potential for modulation of important cell functions. Of particular interest to current regenerative medicine efforts, we review several examples that support the functional role of transmembrane potential (Vmem) in the regulation of proliferation and differentiation. Interestingly, distinct Vmem controls are found in many cancer cell and precursor cell systems, which are known for their proliferative and differentiation capacities, respectively. Collectively, the data demonstrate that bioelectric properties can serve as markers for cell characterization and can control cell mitotic activity, cell cycle progression, and differentiation. The ability to control cell functions by modulating bioelectric properties such as Vmem would be an invaluable tool for directing stem cell behavior toward therapeutic goals. Biophysical properties of stem cells have only recently begun to be studied and are thus in need of further characterization. Understanding the molecular and mechanistic basis of biophysical regulation will point the way toward novel ways to rationally direct cell functions, allowing us to capitalize upon the potential of biophysical signaling for regenerative medicine and tissue engineering.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
It has long been known that in addition to the chemical determinants exchanged by cells during growth and development, bioelectrical signals represent a rich and interesting system for intracellular communication and cellular control [1–3]. These signals function also in the process of regeneration [4–6], a cornerstone aspect of modern biomedicine. This field is enjoying a resurgence [7, 8], as the powerful techniques of molecular physiology are being merged with developmental biology and biophysics to reveal novel mechanisms by which bioelectricity controls morphogenesis and can be harnessed to control it [9, 10]. In parallel with the growing importance of stem cell biology in cancer, in addition to the fields of embryogenesis and regeneration, a variety of channelopathies have drawn attention to the role of specific ion transport in neoplasm [11–13]. Here, we review exciting data implicating bioelectrical signals in the control of stem cell behavior, focusing on transmembrane voltage as a cell-autonomous signal (as distinct from exogenous electric fields).
Transmembrane potential (Vmem) refers to the voltage difference across a cell’s bilayer membrane that is established by the balance of intracellular and extracellular ionic concentrations. Such a balance is maintained via passive and active ion transport through various ion channels and transporters located within the membrane. According to traditional membrane potential theory, the resting membrane potential of a cell is achieved when the electrochemical forces driving ion movement are equalized and ionic equilibrium is maintained. Although maintenance of ionic homeostasis is a critical feature of cell viability and metabolism [14, 15], surprising specificity has been uncovered in the relationship between changes in Vmem levels and alteration of cell function. Furthermore, increasing evidence has pointed toward not only a correlation, but a functional relationship between Vmem and cell functions such as proliferation and differentiation. This relationship can be seen in many cell types, several of which will be reviewed here. That this biophysical relationship is conserved in a wide range of cell types (precursor and mature cells; proliferative and quiescent cells; normal and cancerous cells) suggests that Vmem regulation is a fundamental control mechanism. Better characterization of Vmem-regulating and Vmem-regulated pathways will uncover novel ways to control cell behavior. Such knowledge may significantly advance regenerative medicine applications, including stem cell-related tissue engineering efforts, where the potential of bioelectrical regulation is largely unexplored.
Membrane Potential Measurements
Several techniques are currently used to measure Vmem. They fall in two main categories: electrophysiological recordings and dye imaging.
Traditionally, electrophysiological recordings are obtained either by intracellular “sharp” microelectrode recording or by patch clamping. To obtain intracellular recordings, a glass microelectrode impales a cell to make direct contact with the cytoplasm, while another electrode is immersed in the bath solution surrounding the cell [16, 17]. The potential difference between the bath electrode and the penetrating electrode is the Vmem. Sharp microelectrode tips are small in diameter, on the order of tens of nanometers, to minimize damage to the membrane during insertion [16, 18].
In patch clamping, a patch electrode is brought in contact with the cell membrane but does not penetrate the membrane. Instead, the electrode is positioned against the membrane, allowing the glass to form a tight seal (gigaseal) with the membrane. There are several modes of patch clamping; however, only current clamping in the whole-cell configuration allows for direct membrane potential measurements [18]. In the whole-cell configuration, the patch of membrane sealed by the electrode is ruptured by a suction pulse or a large current pulse. Like intracellular recordings, the patch electrode is electrically connected to the cytoplasm of the cell, and when no current is injected, the endogenous Vmem can be recorded relative to a reference electrode in the bath solution. The patch electrode has a larger tip than a sharp microelectrode, has less resistance to allow for current injection, and is typically filled with a cytoplasm-like solution to measure endogenous membrane potential [18].
While both electrophysiological techniques have been widely and successfully used to record membrane potential, there are several inherent limitations of the electrophysiological recording setup. Most systems are designed to record from only single cells at a time and are therefore laborious and low-throughput [17, 19, 20]. Single-cell recordings are also unable to provide information about the spatial dynamics of Vmem change in a cell population and are unable to reflect the degree to which electrical changes in one cell affects neighboring cells [19]. As with spatial resolution in a multicellular system, spatial resolution across the surface of a single cell also cannot be resolved with these techniques [20]. Electrophysiological methods also generally do not provide information about long-term temporal changes in Vmem, since recordings are typically conducted over only minutes or hours. Some of these limitations have begun to be addressed with microchip-based patch clamping. For example, several chip-based devices use a planar patch clamp approach, where microchips are fabricated with apertures that serve as inverted patch electrode tips, allowing parallel processing of many cell recordings simultaneously [21].
Another approach to membrane potential measurements is the use of voltage-sensitive fluorescent dyes. Several of these dyes are thought to operate by an electrochromic effect, where the dye spectra are altered due to the coupling of molecular electronic states with the electric field present in the membrane, or an electrophoretic effect, where distribution of the dye across the membrane is voltage-sensitive [20, 22, 23]. These dyes typically respond to membrane potential with sensitivities of 10% per 100 mV [20, 22]. In addition to changes in fluorescence, second harmonic generation signals from some dyes also exhibit voltage sensitivities of up to 43% [22]. Advantages of optical detection of Vmem changes include ease of use, simultaneous monitoring of many cells over many different regions, and the ability to resolve spatial differences over the surface of a single cell [17, 19, 20]. Voltage-sensitive dyes also facilitate Vmem measurements in small cells or structures (such as the thin dendritic processes of neurons) that are traditionally difficult to impale or patch with electrodes [17]. One major disadvantage to optical methods, however, is the difficulty of dye calibration, and thus the difficulty of obtaining absolute values for membrane potential [17]. Most data are reported as percentage changes in fluorescence over a basal fluorescence value and are sometimes converted into an estimated membrane potential value based on reported dye sensitivities [17]. Ratiometric imaging using fluorescence resonance energy transfer (FRET) between a mobile voltage-sensing dye and a membrane-bound fluorophore can improve voltage sensitivity, reduce experimental error, and provide information about the magnitude of the voltage change [19].
Proliferation
It has long been observed that Vmem levels are tightly correlated with cell proliferation-related events such as mitosis, DNA synthesis, and overall cell cycle progression. Resting potentials of various cell types fall within a wide range (generally −10 mV to −90 mV), and cells’ positions along such a Vmem scale generally correspond to their proliferative potential [24]. Somatic cells that have a high degree of polarization (a hyperpolarized Vmem) tend to be quiescent and do not typically undergo mitosis. Conversely, developing cells and cancerous cells tend to have a smaller degree of polarization (a depolarized Vmem) and are mitotically active [24, 25]. In addition, cells transferred to in vitro culture from an in vivo environment tend to undergo spontaneous proliferation, which is accompanied by Vmem depolarization [25]. Similarly, proliferation induced by malignant transformation of somatic cells is also accompanied by depolarization [25].
Cone (1971) theorized that this correlation is indicative of a functional relationship between Vmem and mitotic level: transmembrane potential in non-proliferative cells could act as an inhibitory signal for mitosis (or the preparative events associated with mitosis), which, upon stimulation, could be reversibly altered to a level that is permissive for proliferation [25]. In cell cycle progression, a plausible scenario is that a highly polarized Vmem level blocks quiescent somatic cells residing in the G1 phase of the cell cycle from entering the S phase of DNA synthesis, thus inhibiting mitosis [25]. From the observation that most non-proliferative cells have relatively hyperpolarized (more negative) Vmem, while proliferative and cancerous cells have relatively depolarized (less negative) Vmem, Binggeli and Weinstein (1986) further hypothesized that there may be a boundary Vmem level that serves as a threshold or trigger for DNA synthesis [24].
Several studies have confirmed that Vmem modulation can stimulate or inhibit proliferation in a predictable way. Cone and Tongier (1973) investigated the effects of different Vmem levels on mitotic activity of Chinese hamster ovary cells [26]. Vmem levels were varied by changing the ionic composition of the medium to simulate a range of Vmem normally seen in vivo (−10 mV to −90 mV). Complete mitotic arrest was achieved by hyperpolarizing the Vmem to −75 mV but could be reversed by returning to a normal Vmem of −10 mV [26].
Since these initial studies, ionic regulation of cellular behavior has been increasingly studied and has been found to play a critical role in proliferation. It has become clear that the relationship between Vmem and proliferation is not a simple one. Since Vmem is a parameter that reflects the cumulative activity of many ion channels and currents, Vmem-induced cell behavior could be the result of one or many ion-related events. In dissecting control pathways, it is important to determine whether downstream events are controlled by the pure voltage, or by the flow (or concentration) of individual ions [27].
Cell proliferation is a multi-step event regulated by a system of checkpoints at different phases of the cell cycle. Such complexity has been addressed in more recent work on the role of Vmem in proliferation, resulting in a better understanding of the major ion channels and currents involved, as well as stage-specific regulation of the cell cycle. Many of these studies have implicated K+ currents as protagonists of proliferation and cell cycle progression [28, 29]. Correlations between K+ channel inhibition and inhibition of proliferation have been shown in a variety of cell types, including lymphocytes, peripheral blood mononuclear cells (PBMCs), lymphoma, brown fat, melanoma, breast cancer, Schwann cells, astrocytes, oligodendrocytes, neuroblastoma, lung cancer, bladder cancer, and melanoma (reviewed in [28, 30]). Several model systems will be reviewed below. In most systems, K+ flux changes resulting in depolarization favor proliferation, although there are cases where depolarization inhibits proliferation.
Activation of Proliferation
To understand endogenous regulation of proliferation, it is particularly useful to study systems in which cells endogenously switch from quiescent to proliferative phenotypes, or vice versa, or systems in which proliferative activity can be switched on by well-characterized stimuli (e.g., in response to injury or in response to mitogen exposure). Particularly impressive is the initiation of mitosis in normally post-mitotic cells, such as in the CNS; although the molecular details remain to be worked out, even mature neurons can be coaxed to re-enter the cell cycle by long-term depolarization, raising the possibility that a degree of stem cell-like plasticity could be induced in terminally-differentiated somatic cells by bioelectric signals [31–33].
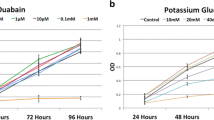
Astrocyte cells display such behavior and have consequently been well studied. In several models of astrocyte injury (scarring of confluent spinal cord astrocytes; cortical freeze-lesions in rat brain), only astrocytes with relatively depolarized resting Vmem and lacking functional inward rectifier K+ (Kir) channels displayed active proliferation in response to injury [34, 35]. In astrocytes from developing rat spinal cord, hyperpolarization of the resting membrane potential (approximately −50 mV to −80 mV) was accompanied by decreased cell proliferation and expression of Kir channels [36, 37]. Conversely, depolarization of quiescent astrocytes with ouabain or extracellular K+ increased proliferation and DNA synthesis [28]. The correlation between ionic activity and proliferation was examined in further detail by studying the effects of cell cycle arrest on ion channel currents and the effects of exogenous current inhibition on cell cycle progression. In proliferating astrocytes, cell arrest in G1/G0 induced premature up-regulation of an inwardly rectifying K+ current (IKIR), resulting in a relatively hyperpolarized phenotype, while arrest in S phase induced downregulation of IKIR with a concomitant increase in a delayed outwardly rectifying current (IKDR), resulting in a relatively depolarized phenotype [28]. Pharmacological inhibition of IKDR in normally proliferating astrocytes resulted in G0/G1 arrest, while inhibition of IKIR in quiescent astrocytes resulted in increased proliferation and DNA synthesis [28]. These data imply that there is a G1/S transition checkpoint where increased IKDR and decreased IKIR currents and the corresponding changes in Vmem are prerequisites for cell cycle progression.
Vascular smooth muscle cells (VSMCs) retain much plasticity even in the adult, and can undergo significant changes in phenotype (phenotypic switching, or modulation) in response to environmental stimuli. During vascular development and in response to vascular injury, VSMC modulation is characterized by a loss of contractile phenotype accompanied by an increase in proliferative and migratory ability [38]. One feature of modulation is a significant change in ion transport mechanisms between contractile and proliferative phenotypes [39]. Contractile VSMCs express an abundance of large-conductance calcium-activated K+ channels (BKCa, also called MaxiK, KCa1.1), which modulate Ca2+ influx through L-type voltage-gated Ca2+ channels (CaV1.2), which are also highly expressed. However, during VSMC modulation, these ion channels are downregulated [40, 41], while an intermediate-conductance calcium-activated K+ channel (IKCa, also called KCa3.1) is activated [42]. BKCa is activated by depolarization, while IKCa is not depolarization-dependent and is thus able to open at more hyperpolarized Vmem, driving Ca2+ entry through voltage-independent, and possibly lipid-sensing, channels [39]. Proliferation-inducing switch from BKCa to IKCa may therefore hint at different voltage sensitivities driving Ca2+ transport during the two cell states.
Activation of quiescent human T lymphocytes and PBMCs by mitogens also involves ionic regulation of cell cycle progression. Upon activation by the mitogen phytohemagluttinin, lymphocytes undergo a transition from G0 to G1 and express the cytokine interleukin-2, which further stimulates a transition from G1 to S [30]. T lymphocyte and PBMC proliferation can be blocked with peptide toxins with high affinity to K+ channels [43–45], suggesting that mitogen-stimulated proliferation is mediated by K+ channel activity. Similarly, activation of murine B lymphocytes and murine noncytolytic T lymphocytes can be blocked by K+ channel inhibitors, which inhibit their progression through the G1 phase of the cell cycle [46, 47]. Molecular studies targeting K+ channels have implicated particularly the voltage-gated K+ channel Kv1.3 in G1 progression and lymphocyte activation. Ca2+ signaling, which is required for activation, is modulated by the activity of Kv1.3 and IKCa channels, which together regulate resting Vmem levels and thereby modulate Ca2+ entry [48]. The relative abundance of these channels changes during lymphocyte activation, and may account for the changes in Ca2+ signaling [49–51]. In quiescent cells, Kv1.3 expression is greater than IKCa expression and is therefore thought to control Vmem. However, upon mitogen stimulation, IKCa is upregulated and may play a greater role in modulating Vmem for further regulation of Ca2+ signaling [50, 52]. Thus, differential K+ channel expression may be responsible for modulating Vmem, which regulates the Ca2+ signaling necessary for a downstream immune response [50].
Proliferation of Cancer Cells
Cancer cells, which show an abnormally high propensity to proliferate, are useful models in which to study ionic regulation, or mis-regulation, of proliferation and cell cycle progression [53, 54].
For example, MCF-7 human breast cancer cell proliferation has been shown to require a characteristic Vmem hyperpolarization during the G0/G1 phase transition [55, 56]. Hyperpolarization occurs via an ATP-sensitive, hyperpolarizing K+ current, comprised of several K+ currents including human ether à go-go (hEAG) and IKCa currents [57–61]. Inhibition of hEAG and IKCa channels induces membrane depolarization and a decrease in intracellular Ca2+, resulting in early G1 phase arrest [62]. K+ channel inhibition also results in accumulation of cyclin-dependent kinase inhibitor p21, which is known to block the G1/S transition [62]. A current model for cell cycle regulation by hyperpolarizing K+ channels is that hEAG is activated during early G1, when Vmem is depolarized to about −20 mV. hEAG expression is then further upregulated during late G1, causing Vmem hyperpolarization and increasing the driving force for Ca2+ entry. Ca2+ entry triggers activation of hIKCa channels, resulting in further hyperpolarization that drives the G1/S transition [62].
A glioma cell model has also been used to demonstrate the functional importance of the inwardly-rectifying Kir4.1 channel in glial cell proliferation. Kir4.1 is not expressed in immature, proliferating glial cells [36, 63, 64], but is widely expressed in glial-differentiated astrocytes [28, 65], and its expression is associated with a hyperpolarized phenotype and an exit from the cell cycle [36, 37]. Functional Kir4.1 channels are also absent in glial-derived tumor cells, and the resulting depolarized phenotype has been suggested to contribute to uncontrolled glioma tumor growth [66]. When Kir4.1 channels were selectively overexpressed in astrocyte-derived gliomas, glioma cells exhibited a differentiated astrocyte-like phenotype, including membrane hyperpolarization and cell growth inhibition by a transition from the G2/M to the G0/G1 phase of the cell cycle [66]. This study demonstrated that Kir4.1 expression was sufficient to induce cell maturation characterized by changes in Vmem (hyperpolarization) and proliferative capacity.
Proliferation of Precursor and Stem Cells and Proliferation in Regenerating Systems
Vmem-associated changes have also been shown to regulate proliferation in precursor cells, stem cells, and regenerating systems. In neural precursor cells (NPCs) isolated from neurospheres derived from adult mice, IKIR and IKDR channels were responsible for establishing a hyperpolarized resting Vmem of approximately −80 mV [67]. Depolarization by extracellular Ba2+ or K+ accelerated mitosis in NPCs, resulting in an increase in cell number and neurosphere size. It is hypothesized that Vmem depolarization via modulated Kir channel activity is responsible for NPC proliferation and cell cycle progression [67]. Interestingly, the effect was maximal at 100 μM but declined at 1 mM. This biphasic effect reveals the presence of an optimal membrane potential range—a window effect which will have to be taken into account when designing modulation techniques for biomedical applications.
In human (hESCs) and mouse (mESCs) embryonic stem cells, IKDR currents are present and are permissive for proliferation, as application of K+ channel blockers inhibited DNA synthesis [68]. In Xenopus embryos, the K+ channel KCNQ1 (also called Kv7.1) contributes significantly to the membrane potential [69]. When its regulatory subunit KCNE1 (also called minK, Isk) was misexpressed in the embryo, KCNQ1 currents were suppressed, resulting in Vmem depolarization and ectopic induction of the neural crest regulator genes Sox10 and Slug. Consistent with these two genes’ known roles in neoplastic progression [70–73], reduction of KCNQ1-dependent currents induced overproliferation of melanocytes and conferred upon them a highly invasive, migratory phenotype resembling metastasis [69]. Thus, a functional role was identified for the channel KCNQ1, whose Vmem-controlling activity regulates the mitotic and invasive activity of the melanocyte neural crest lineage through known signaling pathways. A functional role was also found for the vacuolar ATPase H+ pump (V-ATPase) in Xenopus tadpole tail regeneration, which requires the H+ pumping activity of endogenously expressed V-ATPases [74]. Loss of V-ATPase function, and the resulting depolarization in the tail bud region, decreased the number of proliferating cells in the bud and abolished regeneration. Conversely, expression of a heterologous H+ pump repolarized the bud and induced a significant degree of regeneration in normally non-regenerative conditions [74]. These studies demonstrate the importance of Vmem modulation by specific molecular species in regulating cell growth during embryogenesis and regeneration.
Differentiation
Regulation of proliferation and cell cycle progression is closely associated with differentiation, since cells must coordinate their exit from the cell cycle with the initiation of their differentiation programs [75]. Thus, since Vmem regulates proliferation in many cell types, Vmem-related signals may also act as triggers for differentiation. A thorough understanding of these signals would be an invaluable resource for characterizing and controlling cell development and maturation in many systems. Two key questions must be answered in order to extract therapeutically relevant information about Vmem in differentiation: (1) what are the electrophysiological differences between the differentiated and undifferentiated states, and (2) are these differences instructive for differentiation? Following the identification of functionally significant changes in bioelectric state during differentiation, we may be able to manipulate the parameters so as to control differentiation outcomes for therapeutic applications.
Electrophysiological Changes During Cell Development and Differentiation
Currently, most work in this area has focused on comparing the electrophysiological profiles of differentiated cells and undifferentiated cells. In addition to providing clues about potential control points for differentiation, these profiles can be used to better characterize the maturation of stem and progenitor cells, as well as to identify and distinguish between subpopulations that may not show other differentiating phenotypes. The majority of work has been done in neural and muscular systems, as the acquisition of electrophysiological features contributes to the excitability of the mature cell. During early stages of development, maturing neural crest (NC) cells express human ether à go-go related gene encoded K+ currents (IHERG) and IKDR currents, while during later stages, NC cells exhibit Vmem hyperpolarization and expression of IKDR, IKIR, and Na+ (INa) currents [76–79]. Based upon these data, it was suggested that the ordered expression of ion channels defines NC cell developmental stages [78]. Similarly, the NC-derived SY5Y neuroblastoma cell line exhibits specific electrophysiological profiles (relative ratios of IHERG, IKDR, INa) depending on differentiated state and specific subtype, N- or S-type [80]. N-type cells display an immature, nonexcitable neural phenotype and are characterized by IHERG and IKDR currents and a depolarized Vmem. Upon stimulation, they differentiate into excitable neural cells and express IKIR [80]. S-type cells display negligible IHERG, IKDR, and INa currents, but upon differentiation along a smooth muscle pathway or a neural abortive pathway, display characteristic (and different) levels of these currents [80]. Characteristic changes in Na+ and K+ channel expression and ionic currents have also been found to accompany neural differentiation of other stem-like cell types, such as neural stem-like cells from human umbilical cord blood [81], immortalized human neural stem cells [82], and mESCs [83]. These studies suggest that electrophysiological profiles can be coupled with traditional immunocytochemical techniques to describe the maturation state of cells and to distinguish between different cell populations originating from common precursors.
Similarly, the electrophysiology of myocyte differentiation has also been characterized. Differentiation of mouse embryonic stem cells [84] and of embryonic carcinoma P19 cells [85] into cardiomyocytes correlates with upregulation of cardiac-related ion channels in specific temporal patterns. P19 cells express L-type Ca2+ and transient outward channels early during differentiation, and Na+ and delayed and inward rectifier channels later during differentiation [85]. Skeletal myoblasts also exhibit different current and ion channel profiles in their proliferating and differentiating states. Murine C2C12 myoblasts undergoing active proliferation express an ATP-induced K+ current, a swelling-activated Cl- current, and an IKCa current [86–88]. Upon initiation of differentiation, these currents are replaced with a tetrodotoxin-sensitive Na+ current, an IKDR current, an IKIR current, and an L-type Ca2+ current [87–90]. In muscle satellite cell-derived human myoblasts, voltage-gated Na+ and Ca2+-activated K+ channels are expressed during proliferation [91], while hEAG, IKDR, IKIR, T-type Ca2+, and L-type Ca2+ channels are expressed in differentiated fusion-competent myoblasts [92–96].
Functional Role of Vmem Signaling During Differentiation
Beyond their role as markers of the maturation process, electophysiological changes play functional and instructive roles in the differentiation process. In such a role, they could provide more than just a passive readout of developmental stage or of linage commitment, but could actively contribute to transcriptional and other activity leading to expression of a differentiated phenotype. Observations made over 30 years ago showed that neural differentiation depended on the function of specific ion transporters such as the Na,K-ATPase [97, 98]. By uncovering the molecular and mechanistic basis of the underlying signaling pathways, we may find novel ways to direct stem cell behavior by modulating Vmem and related ion channel expression.
Several recent studies have demonstrated that endogenous Vmem modulation does indeed have an instructive role during cell differentiation and maturation. For example, Vmem hyperpolarization not only precedes human myoblast differentiation, but is also required for differentiation, as myocyte fusion and transcription factor activity are blocked when hyperpolarization is blocked [96, 99]. It is the earliest detectable event in the differentiation process, and is therefore thought to be a trigger for myoblast differentiation [99]. Hyperpolarization, and thus differentiation, is thought to be initiated by tyrosine dephosphorylation of the Kir2.1 channel and results in Ca2+ influx through T channels, calcineurin (CaN) activation, and expression of two myocyte transcription factors, myogenin and myocyte enhancing factor 2 [93, 95, 99–101]. Similarly, expression of the chloride channel ClC-3 and its corresponding Cl- current is required for fibroblast-to-myofibroblast differentiation [102], and expression and function of two inward K+ rectifier channels is essential for the differentiation of human hematopoietic progenitor cells [103, 104]. Taken together, these results support a relationship between ion channel modulation and the intracellular signaling pathways involved in the differentiation process. That the endogenous hyperpolarization happens upstream of known conventional biochemical signaling events also hints at the possibility of using a single control point (e.g., Vmem, or Kir2.1 channel phosphorylation) to modulate the differentiation-related signaling pathways that diverge from that point.
Hyperpolarization also plays a role in the development and maturation of mammalian cerebellar granule cells. Developing granule cells hyperpolarize from −25 mV to −55 mV, and it is hypothesized that these Vmem changes alter Ca2+ signaling via CaN and Ca2+-calmodulin-dependent protein kinase to control stage-specific gene expression [105, 106]. Vmem- and CaN-mediated changes in granule cell gene expression were found after treatment with depolarization agents and/or a CaN inhibitor FK506 [107]. Interestingly, ~80% of developmentally-relevant genes corresponded to depolarization-regulated genes, and the correlation was such that developmentally-upregulated genes were downregulated with depolarization, while developmentally-downregulated genes were upregulated with depolarization [107]. Furthermore, there was a large overlap and inverse relation seen between depolarization- and FK506-regulated genes [107]. These data suggest that endogenous regulation of Vmem level controls genes associated with maturation of granule cells, and that this regulation may be mediated by CaN.
More recently, we have shown a similar connection between Vmem and differentiation propensity in bone marrow-derived human mesenchymal stem cells (hMSCs). Similar to what was found for human myoblast and cerebellar granule cell differentiation, and also in line with Binggeli and Weinstein’s hypothesis [24] about Vmem levels in developing vs. quiescent cells, hMSCs undergo hyperpolarization during both osteogenic (OS) and adipogenic (AD) differentiation [108]. More importantly, hyperpolarization was found to be necessary for differentiation. When normal Vmem progression was disrupted by depolarization with high K+ or ouabain, OS and AD differentiation markers decreased significantly, suggesting suppression or delay of differentiation under depolarized Vmem conditions [108]. Conversely, during OS differentiation, treatment with hyperpolarizing agents pinacidil or diazoxide induced upregulation of bone-related gene expression [108]. These depolarizing and hyperpolarizing experiments demonstrate that hMSCs are sensitive to bidirectional changes in Vmem and provide compelling evidence for an instructive role of Vmem in differentiating hMSCs. The discovery that Vmem can regulate long-term cell behavior in a non-excitable cell type is exciting because it highlights the fact that ion flows are important for a broad range of cell functions, of which excitability is only a small part. Rational modulation of ion currents and ion channel expression may therefore be potential control mechanisms for a variety of cell signaling pathways. Possibilities include maintenance of a renewable stem cell population in vitro and acceleration or augmentation of stem cell differentiation for therapeutic purposes or for tissue engineering.
It should also be noted that bioelectric signals may function in de-differentiation, a process of considerable importance for understanding regeneration of some structures [109, 110]. This work [111–115] is largely pre-molecular, and it remains to be seen what ion transporters and mechanisms can be capitalized upon in order to de-differentiate somatic cells for biomedical applications.
Electrophysiological Characterization and Profiling of Stem Cells
Realizing the potential of bioelectric control for stem cell therapies and for general understanding of stem cell biology requires thorough characterization of ion channel and current expression during proliferation and differentiation of stem cells [83, 116–120]. Alongside transmembrane voltage gradient, cells’ dielectric properties reveal surface charge, membrane conductivity, nucleic acid content, cell size, and presence and conductivity of internal membrane-bound vesicles; this can be used to distinguish stem cells and their differentiated progeny [121].
A number of studies have begun to profile ion current and channel expression in undifferentiated stem cells [80, 122]. For example, hMSCs derived from bone marrow express IKCa, IKDR, transient outward K+ currents, and slow-activating currents [123, 124]. They display high expression levels for several channel subunits, including Kv4.2, Kv4.3, MaxiK, α1c subunit of L-type Ca2+ channels, and hyperpolarization-activated cyclic nucleotide-gated ion channel isoform 2 [123]. Similar to bone marrow-derived stem cells, undifferentiated human adipose tissue-derived stem cells express IKDR and IKCa currents [125]. Human and mouse ESCs exhibit IKDR currents at different levels of homogeneity, and it is hypothesized that differential ion channel gene expression is responsible for current expression in the two cell types [68]. Rat embryonic neural stem cells can be characterized by a signature ion channel profile, including several Na+, Ca2+, and K+ channels [126]. Bioelectrical components (e.g., Kir current) are differentially present in mesenchymal stem cells, and can perhaps be used to identify distinct sub-populations [127]. Human vascular endothelial cells consist of 3 discrete sub-populations based on their ion channel properties [128].
We believe that obtaining a full profile of stem cell bioelectric state will be a key step in understanding the major players in ionic regulation and will help identify potential targets for manipulation. Such characterization may also supplement current immunohistochemical techniques for identifying stem cell sub-populations, and the development of sensitive fluorescent dyes that report Vmem, pH, and individual ion content will allow cells in unique physiological states to be segregated non-invasively via FACS. Profiling of stem cell bioelectric state may prove to be a novel technique to identify stem cells that are difficult to phenotype by traditional methods (such as hMSCs, whose set of identifying markers is still unclear), or a technique to distinguish between a heterogenous stem cell population (again, such as an hMSC population, which is thought to be composed of cells with varying differentiation propensities) [129, 130].
Migration
The function of stem cells in vivo is often determined by their position within tissues and organs; this is crucial because microenvironment controls stem cell behaviors [131–133], and because targeting stem cells towards areas of injury is a key goal of regenerative medicine. Moreover, stem cells are now an increasingly promising vector for cancer therapies, because some types, such as neural stem cells, appear to preferentially target aggressive tumors such as gliomas [134, 135]. While the cues that guide stem cells’ homing in mammals have mainly been studied from the perspective of chemical gradients and ECM molecules [136–140], there is a model in which physiological signals have begun to be investigated.
Planaria, flatworms with impressive regenerative abilities [141–143], possess a resident adult stem cell population. These neoblasts appear to migrate to wounds and recreate the necessary tissues [144, 145]. Importantly, this requires the stem cells to be informed as to where the damage occurred, what cell types are missing, what morphogenetic structures must be recreated by a combination of coordinated proliferation and differentiation, and when the target morphology is complete (regenerative stem cell activity can cease); the process involves molecules like PTEN [146], which are powerful regulators of stem cell activity in mammals. Recent data show that gap junctions, direct channels between adjacent cells that allow transfer of ions and small signaling molecules, are centrally involved in this process in planaria [147, 148] and many other systems [149]; gap junctional communication is an ideal way for stem cells to rapidly communicate with their niche. Gap junctions are both gated by Vmem [150, 151] and establish iso-potential cell fields (define physiological compartments) [152, 153]. Moreover, recent molecular data implicated KCNQ1 potassium channels [154] and NaV sodium channels [12, 155, 156] in the regulation of migration and invasiveness of several stem-like cell types. These findings implicate the cell-autonomous property Vmem in migration control, and complement the long-known ability of stem cells to migrate in physiological-strength electric gradients in their environment [157–159]. Thus, an investigation of voltage in the guidance of stem cell position during regeneration and morphostasis is a crucial area for future work.
Mechanisms: How is Vmem Transduced into Cellular Behaviors?
Since Vmem signals may be control points for guiding stem cell behavior in biomedical settings, it is necessary to identify the Vmem-sensing pathways that link bioelectric signals to cell behavior such as proliferation and differentiation. In order to mechanistically dissect bioelectrical signals, it is important to distinguish which aspect of ion flow bears the instructive signals for cell behavior: membrane potential change, long-range electric field, or flow of individual ions. In many cases, this can be very difficult to untangle. However, it has been accomplished in some developmental studies by using mutants of ion transporters that allow separation of individual biophysical events [160]. It is now possible to use molecular-genetic reagents in gain- and loss-of-function approaches to specifically modulate different aspects of ion flux [154], controlling corneal healing [10], inducing tail regeneration [9] at non-regenerative stages, and drastically altering the positioning and proliferation of neural crest cells [154]. For example, misexpression of electroneutral transporters can differentiate between the importance of voltage changes vs. that of flux of specific ions. Pore mutants can distinguish between ion conductance roles vs. possible functions of channels/pumps as scaffolds or binding partners (non-electrical signaling); for example, in the Na+/H+ exchanger, both ion-dependent and ion-independent functions control cell directionality and Golgi apparatus localization to wound edge [161]. Gating channel mutants and pumps with altered kinetics can, respectively, be used to reveal upstream signals controlling the bioelectric events, and the temporal properties of the signal. Heterologous transporters, combined with blockade of endogenous channels or pumps, can be used in elegant rescue experiments.
The question of which aspect of ion flow is relevant in any instance of cell behavior is intimately tied to transduction mechanism: how does the cell (or a neighboring cell) know the membrane voltage has changed? Mechanisms that transduce electrical signal into second-messenger cascades [162] include those outlined in Table 1. While many questions remain, the molecular details of at least some such transduction pathways have recently been revealed (Fig. 1). One major candidate for Vmem-sensing mechanisms is Ca2+ signaling [163, 164]. Calcium signaling is crucially important to many cell behaviors, including proliferation [165–167], differentiation [168, 169], and galvanotaxis [170]. It also functions as a patterning signal in large-scale morphogenesis [171–175]. It is not possible to do justice here to the enormous literature on calcium signaling, and this has been well-reviewed elsewhere [176–179]. Most of these signaling events take place through specialized receptors such as calmodulin or calcineurin [180], since calcium signals largely by virtue of its unique chemical properties—it is not a true electrical signal. However, one area where Ca2+ signaling is integral to bioelectrical cues is in the transduction of membrane voltage to downstream cellular effector mechanisms. This often occurs through voltage-gated calcium channels [181–184], although in some instances of K+-dependent signaling, Ca2+ fluxes were not affected by K+ channel activity, showing that proliferative effect is not always due to modulation of intracellular Ca2+. Other Vmem-transducing mechanisms may include integrin-linked signaling involving hERG1 channels [185–189]; voltage-sensitive phosphatases operating through the phosphoinositide kinase pathway [190–193]; voltage-dependent changes in the function of intracellular transporters of signaling molecules such as serotonin [162]; and others.

Integration of bioelectric events with canonical biochemical and genetic pathways occurs through a number of sequential phases. Such signals can be initiated at the cell membrane of individual cells (function of ion transporters), can arrive through gap junctional connections to their neighbors, or be imposed through breaks in an epithelium that carries a transepithelial potential. Physically, such signals are carried by changes in transmembrane potential, pH gradients, flows of specific ions, or long-range electric fields. A number of mechanisms serve as biophysical receptors for these signals, including voltage-sensing domains within proteins, changes of intracellular ion content, electro-osmosis, changes in the gating of transporters for signaling molecules, calcium influx, and electrophoresis of morphogens through gap junctional paths between cells. A number of early response genes have been identified immediately downstream, including integrins, Slug/Sox10, Notch, NF-kB, and PTEN. Because these transcriptional cascades can control all aspects of cell behavior, including proliferation, differentiation, and migration, transduction into these secondary pathways allow bioelectrical signals to control cell number and type during complex morphogenetic events such as tissue regeneration
A key issue for future work concerns specificity. How much information can be encoded in a number such as Vmem (do cells interpret it as a binary depolarized vs. hyperpolarized switch, or a larger number of discrete levels)? Do cells have a single Vmem value, or more likely, is the cell membrane a manifold containing a huge number of local microdomains expressing different transporters and thus presenting a very rich amount of information to neighboring cells as well as intracellular processes within the same cell? Do individual ion channels provide different signals to cells even when their effect on Vmem is similar? What are the time-dependent kinetics of Vmem changes in non-excitable cells (slow changes in transmembrane potential)? Such mechanistic understanding will be critical for pinpointing the most effective and specific molecular targets for pharmacological and molecular-genetic interventions [130].
Conclusions
Ionic regulation is a rich yet largely untapped toolbox for rational manipulation of cell behavior [194]. From existing studies, it is clear that ion flows contribute to much more than cell excitability, playing a functional role in proliferation, cell cycle progression, and cell maturation and differentiation. To capitalize upon the potential of bioelectric regulation for regenerative medicine, the fields of electrophysiology and stem cell biology must converge to uncover the molecular and mechanistic basis of ion channel and current contributions to cell behavior, with a view to using Vmem and other biophysical properties as (1) a profiling tool with which to characterize cell populations as they undergo changes in behavior and (2) as a control point with which to modulate their behavior. With the availability of pharmacological agents and molecular genetics tools to regulate ion channel activity and expression, we have many tools at our disposal for probing biophysical parameters and learning how to exploit them to direct cell functions for therapeutic applications.
References
Robinson, K. R., & Messerli, M. A. (1996). Electric embryos: the embryonic epithelium as a generator of developmental information. In C. D. McCaig (Ed.), Nerve growth and guidance (pp. 131–150). London: Portland.
Jaffe, L. F., & Nuccitelli, R. (1977). Electrical controls of development. Annual Review of Biophysics and Bioengineering, 6, 445–476.
Lund, E. (1947). Bioelectric fields and growth. Austin: University of Texas Press.
Borgens, R. B. (1982). What is the role of naturally produced electric current in vertebrate regeneration and healing. International Review of Cytology, 76, 245–298.
Borgens, R. B., Vanable, J. W., Jr., & Jaffe, L. F. (1977). Bioelectricity and regeneration. I. Initiation of frog limb regeneration by minute currents. Journal of Experimental Zoology, 200, 403–416.
Mathews, A. P. (1903). Electrical polarity in the hydroids. American Journal of Physiology, 8, 294–299.
McCaig, C. D., Rajnicek, A. M., Song, B., & Zhao, M. (2005). Controlling cell behavior electrically: Current views and future potential. Physiological Reviews, 85, 943–978.
Levin, M. (2007). Large-scale biophysics: Ion flows and regeneration. Trends in Cell Biology, 17, 262–271.
Adams, D. S., Masi, A., & Levin, M. (2007). H+ pump-dependent changes in membrane voltage are an early mechanism necessary and sufficient to induce Xenopus tail regeneration. Development, 134, 1323–1335.
Zhao, M., Song, B., Pu, J., et al. (2006). Electrical signals control wound healing through phosphatidylinositol-3-OH kinase-gamma and PTEN. Nature, 442, 457–460.
Arcangeli, A. (2005). Expression and role of hERG channels in cancer cells. Novartis Foundation Symposium, 266, 225–232. discussion 32–4.
Mycielska, M. E., & Djamgoz, M. B. (2004). Cellular mechanisms of direct-current electric field effects: Galvanotaxis and metastatic disease. Journal of Cell Science, 117, 1631–1639.
Wang, Z. (2004). Roles of K+ channels in regulating tumour cell proliferation and apoptosis. Pflugers Archiv, 448, 274–286.
Bortner, C. D., & Cidlowski, J. A. (2004). The role of apoptotic volume decrease and ionic homeostasis in the activation and repression of apoptosis. Pflugers Archiv, 448, 313–318.
Franco, R., Bortner, C. D., & Cidlowski, J. A. (2006). Potential roles of electrogenic ion transport and plasma membrane depolarization in apoptosis. Journal of Membrane Biology, 209, 43–58.
Ling, G., & Gerard, R. W. (1949). The normal membrane potential of frog sartorius fibers. Journal of Cellular and Comparative Physiology, 34, 383–396.
Stuart, G. J., & Palmer, L. M. (2006). Imaging membrane potential in dendrites and axons of single neurons. Pflugers Archiv, 453, 403–410.
Molleman, A. (2003). Patch clamping: an introductory guide to patch clamp electrophysiology. Chichester, England: Wiley.
González, J. E., & Tsien, R. Y. (1997). Improved indicators of cell membrane potential that use fluorescence resonance energy transfer. Chemistry & Biology, 4, 269–277.
Loew, L. M. (1992). Voltage-sensitive dyes: Measurement of membrane potentials induced by DC and AC electric fields. Bioelectromagnetics, (Suppl 1):179–89.
Brüggemann, A., Stoelzle, S., George, M., Behrends, J. C., & Fertig, N. (2006). Microchip technology for automated and parallel patch-clamp recording. Small, 2, 840–846.
Millard, A. C., Jin, L., Wei, M. D., Wuskell, J. P., Lewis, A., & Loew, L. M. (2004). Sensitivity of second harmonic generation from styryl dyes to transmembrane potential. Biophysical Journal, 86, 1169–1176.
Plášek, J., & Sigler, K. (1996). Slow fluorescent indicators of membrane potential: A survey of different approaches to probe response analysis. Journal of Photochemistry and Photobiology. B, Biology, 33, 101–124.
Binggeli, R., & Weinstein, R. C. (1986). Membrane potentials and sodium channels: Hypotheses for growth regulation and cancer formation based on changes in sodium channels and gap junctions. Journal of Theoretical Biology, 123, 377–401.
Cone, C. D., Jr. (1971). Unified theory on the basic mechanism of normal mitotic control and oncogenesis. Journal of Theoretical Biology, 30, 151–181.
Cone, C. D., Jr., & Tongier, M., Jr. (1973). Contact inhibition of division: Involvement of the electrical transmembrane potential. Journal of Cellular Physiology, 82, 373–386.
Adams, D. S., & Levin, M. (2006). Strategies and techniques for investigation of biophysical signals in patterning. In M. Whitman & A. K. Sater (Eds.), Analysis of growth factor signaling in embryos: Taylor and Francis books (pp. 177–262).
MacFarlane, S. N., & Sontheimer, H. (2000). Changes in ion channel expression accompany cell cycle progression of spinal cord astrocytes. GLIA, 30, 39–48.
Dubois, J. M., & Rouzaire-Dubois, B. (1993). Role of potassium channels in mitogenesis. Progress in Biophysics and Molecular Biology, 59, 1–21.
Wonderlin, W. F., & Strobl, J. S. (1996). Potassium channels, proliferation and G1 progression. Journal of Membrane Biology, 154, 91–107.
Cone, C. D., & Cone, C. M. (1976). Induction of mitosis in mature neurons in central nervous system by sustained depolarization. Science, 192, 155–158.
Stillwell, E. F., Cone, C. M., & Cone, C. D. (1973). Stimulation of DNA synthesis in CNS neurones by sustained depolarisation. Nature: New Biology, 246, 110–111.
Cone, C. D., & Tongier, M. (1971). Control of somatic cell mitosis by simulated changes in the transmembrane potential level. Oncology, 25, 168–182.
Bordey, A., Lyons, S. A., Hablitz, J. J., & Sontheimer, H. (2001). Electrophysiological characteristics of reactive astrocytes in experimental cortical dysplasia. Journal of Neurophysiology, 85, 1719–1731.
MacFarlane, S. N., & Sontheimer, H. (1997). Electrophysiological changes that accompany reactive gliosis in vitro. Journal of Neuroscience, 17, 7316–7329.
Bordey, A., & Sontheimer, H. (1997). Postnatal development of ionic currents in rat hippocampal astrocytes in situ. Journal of Neurophysiology, 78, 461–477.
Ransom, C. B., & Sontheimer, H. (1995). Biophysical and pharmacological characterization of inwardly rectifying K+ currents in rat spinal cord astrocytes. Journal of Neurophysiology, 73, 333–346.
Owens, G. K., Kumar, M. S., & Wamhoff, B. R. (2004). Molecular regulation of vascular smooth muscle cell differentiation in development and disease. Physiological Reviews, 84, 767–801.
Beech, D. J. (2007). Ion channel switching and activation in smooth-muscle cells of occlusive vascular diseases. Biochemical Society Transactions, 35, 890–894.
Gollasch, M., Haase, H., Ried, C., et al. (1998). L-type calcium channel expression depends on the differentiated state of vascular smooth muscle cells. FASEB Journal, 12, 593–601.
Richard, S., Neveu, D., Carnac, G., Bodin, P., Travo, P., & Nargeot, J. (1992). Differential expression of voltage-gated Ca2+-currents in cultivated aortic myocytes. Biochimica et Biophysica Acta—Protein Structure and Molecular Enzymology, 1160, 95–104.
Neylon, C. B., Lang, R. J., Fu, Y., Bobik, A., & Reinhart, P. H. (1999). Molecular cloning and characterization of the intermediate-conductance Ca(2+)-activated K(+) channel in vascular smooth muscle: relationship between K(Ca) channel diversity and smooth muscle cell function. Circulation Research, 85, e33–e43.
Freedman, B. D., Price, M. A., & Deutsch, C. J. (1992). Evidence for voltage modulation of IL-2 production in mitogen-stimulated human peripheral blood lymphocytes. Journal of Immunology, 149, 3784–3794.
Lin, C. S., Boltz, R. C., Blake, J. T., et al. (1993). Voltage-gated potassium channels regulate calcium-dependent pathways involved in human T lymphocyte activation. The Journal of Experimental Medicine, 177, 637–645.
Price, M., Lee, S. C., & Deutsch, C. (1989). Charybdotoxin inhibits proliferation and interleukin 2 production in human peripheral blood lymphocytes. Proceedings of the National Academy of Sciences of the United States of America, 86, 10171–10175.
Amigorena, S., Choquet, D., Teillaud, J. L., Korn, H., & Fridman, W. H. (1990). Ion channel blockers inhibit B cell activation at a precise stage of the G1 phase of the cell cycle. Possible involvement of K+ channels. Journal of Immunology, 144, 2038–2045.
Lee, S. C., Sabath, D. E., Deutsch, C., & Prystowsky, M. B. (1986). Increased voltage-gated potassium conductance during interleukin 2-stimulated proliferation of a mouse helper T lymphocyte clone. Journal of Cell Biology, 102, 1200–1208.
Cahalan, M. D., & Chandy, K. G. (1997). Ion channels in the immune system as targets for immunosuppression. Current Opinion in Biotechnology, 8, 749–756.
Deutsch, C., Krause, D., & Lee, S. C. (1986). Voltage-gated potassium conductance in human T-lymphocytes stimulated with phorbol ester. Journal of Physiology, 372, 405–423.
Ghanshani, S., Wulff, H., Miller, M. J., et al. (2000). Up-regulation of the IKCa1 potassium channel during T-cell activation: Molecular mechanism and functional consequences. Journal of Biological Chemistry, 275, 37137–37149.
Grissmer, S., Nguyen, A. N., & Cahalan, M. D. (1993). Calcium-activated potassium channels in resting and activated human T lymphocytes: Expression levels, calcium dependence, ion selectivity, and pharmacology. Journal of General Physiology, 102, 601–630.
Khanna, R., Change, M. C., Joiner, W. J., Kaczmarek, L. K., & Schlichter, L. C. (1999). hSK4/hIK1, a calmodulin-binding K(Ca) channel in human T lymphocytes. Roles in proliferation and volume regulation. Journal of Biological Chemistry, 274, 14838–14849.
Kim, C. F., & Dirks, P. B. (2008). Cancer and stem cell biology: How tightly intertwined? Cell Stem Cell, 3, 147–150.
Normile, D. (2002). Cell proliferation. Common control for cancer, stem cells. Science, 298, 1869.
Wonderlin, W. F., Woodfork, K. A., & Strobl, J. S. (1995). Changes in membrane potential during the progression of MCF-7 human mammary tumor cell through the cell cycle. Journal of Cellular Physiology, 165, 177–185.
Woodfork, K. A., Wonderlin, W. F., Peterson, V. A., & Strobl, J. S. (1995). Inhibition of ATP-sensitive potassium channels causes reversible cell-cycle arrest of human breast cancer cells in tissue culture. Journal of Cellular Physiology, 162, 163–171.
Klimatcheva, E., & Wonderlin, W. F. (1999). An ATP-sensitive K+ current that regulates progression through early G1 phase of the cell cycle in MCF-7 human breast cancer cells. Journal of Membrane Biology, 171, 35–46.
Ouadid-Ahidouch, H., Chaussade, F., Roudbaraki, M., et al. (2000). Kv1.1 K+ channels identification in human breast carcinoma cells: Involvement in cell proliferation. Biochemical and Biophysical Research Communications, 278, 272–277.
Ouadid-Ahidouch, H., Le Bourhis, X., Roudbaraki, M., Toillon, R. A., Delcourt, P., & Prevarskaya, N. (2001). Changes in the K+ current-density of MCF-7 cells during progression through the cell cycle: Possible Involvement of a h-ether.a-gogo K+ channel. Receptors and Channels, 7, 345–356.
Ouadid-Ahidouch, H., Roudbaraki, M., Ahidouch, A., Delcourt, P., & Prevarskaya, N. (2004). Cell-cycle-dependent expression of the large Ca2+-activated K+ channels in breast cancer cells. Biochemical and Biophysical Research Communications, 316, 244–251.
Ouadid-Ahidouch, H., Roudbaraki, M., Delcourt, P., Ahidouch, A., Joury, N., & Prevarskaya, N. (2004). Functional and molecular identification of intermediate-conductance Ca 2+-activated K+ channels in breast cancer cells: Association with cell cycle progression. American Journal of Physiology. Cell Physiology, 287, C125–C134.
Ouadid-Ahidouch, H., & Ahidouch, A. (2008). K+ channel expression in human breast cancer cells: Involvement in cell cycle regulation and carcinogenesis. Journal of Membrane Biology, 221, 1–6.
MacFarlane, S. N., & Sontheimer, H. (2000). Modulation of Kv1.5 currents by Src tyrosine phosphorylation: Potential role in the differentiation of astrocytes. Journal of Neuroscience, 20, 5245–5253.
Sontheimer, H. (1994). Voltage-dependent ion channels in glial cells. GLIA, 11, 156–172.
Li, L., Head, V., & Timpe, L. C. (2001). Identification of an inward rectifier potassium channel gene expressed in mouse cortical astrocytes. GLIA, 33, 57–71.
Higashimori, H., & Sontheimer, H. (2007). Role of Kir4.1 channels in growth control of glia. GLIA, 55, 1668–1679.
Yasuda, T., Bartlett, P. F., & Adams, D. J. (2008). Kir and Kv channels regulate electrical properties and proliferation of adult neural precursor cells. Molecular and Cellular Neurosciences, 37, 284–297.
Wang, K., Xue, T., Tsang, S. Y., et al. (2005). Electrophysiological properties of pluripotent human and mouse embryonic stem cells. Stem Cells, 23, 1526–1534.
Morokuma, J., Blackiston, D., Adams, D. S., Seebohm, G., Trimmer, B., & Levin, M. (2008). Modulation of potassium channel function confers a hyperproliferative invasive phenotype on embryonic stem cells. Proceedings of the National Academy of Sciences of the United States of America, 105, 16608–16613.
Ferletta, M., Uhrbom, L., Olofsson, T., Ponten, F., & Westermark, B. (2007). Sox10 has a broad expression pattern in gliomas and enhances platelet-derived growth factor-B-induced gliomagenesis. Molecular Cancer Research, 5, 891–897.
Bannykh, S. I., Stolt, C. C., Kim, J., Perry, A., & Wegner, M. (2006). Oligodendroglial-specific transcriptional factor SOX10 is ubiquitously expressed in human gliomas. Journal of Neuro-oncology, 76, 115–127.
Martin, T. A., Goyal, A., Watkins, G., & Jiang, W. G. (2005). Expression of the transcription factors snail, slug, and twist and their clinical significance in human breast cancer. Annals of Surgical Oncology, 12, 488–496.
Kurrey, N. K., Amit, K., & Bapat, S. A. (2005). Snail and Slug are major determinants of ovarian cancer invasiveness at the transcription level. Gynecologic Oncology, 97, 155–165.
Adams, D. S., Masi, A., & Levin, M. (2007). H+ pump-dependent changes in membrane voltage are an early mechanism necessary and sufficient to induce Xenopus tail regeneration. Development, 134, 1323–1335.
Miller, J. P., Yeh, N., Vidal, A., & Koff, A. (2007). Interweaving the cell cycle machinery with cell differentiation. Cell Cycle, 6, 2932–2938.
Arcangeli, A., Bianchi, L., Becchetti, A., et al. (1995). A novel inward-rectifying K+ current with a cell-cycle dependence governs the resting potential of mammalian neuroblastoma cells. Journal of Physiology, 489, 455–471.
Arcangeli, A., Rosati, B., Cherubini, A., et al. (1998). Long term exposure to retinoic acid induces the expression of IRK1 channels in HERG channel-endowed neuroblastoma cells. Biochemical and Biophysical Research Communications, 244, 706–711.
Arcangeli, A., Rosati, B., Cherubini, A., et al. (1997). HERG- and IRK-like inward rectifier currents are sequentially expressed during neuronal development of neural crest cells and their derivatives. European Journal of Neuroscience, 9, 2596–2604.
Arcangeli, A., Rosati, B., Crociani, O., et al. (1999). Modulation of HERG current and herg gene expression during retinoic acid treatment of human neuroblastoma cells: Potentiating effects of BDNF. Journal of Neurobiology, 40, 214–225.
Biagiotti, T., D’Amico, M., Marzi, I., et al. (2006). Cell renewing in neuroblastoma: Electrophysiological and immunocytochemical characterization of stem cells and derivatives. Stem Cells, 24, 443–453.
Sun, W., Buzanska, L., Domanska-Janik, K., Salvi, R. J., & Stachowiak, M. K. (2005). Voltage-sensitive and ligand-gated channels in differentiating neural stem-like cells derived from the nonhematopoietic fraction of human umbilical cord blood. Stem Cells, 23, 931–945.
Cho, T., Bae, J. H., Choi, H. B., et al. (2002). Human neural stem cells: Electrophysiological properties of voltage-gated ion channels. NeuroReport, 13, 1447–1452.
Chafai, M., Louiset, E., Basille, M., et al. (2006). PACAP and VIP promote initiation of electrophysiological activity in differentiating embryonic stem cells. Annals of the New York Academy of Sciences, 1070, 185–189.
Van Kempen, M. J. A., Van Ginneken, A., De Grijs, I., et al. (2003). Expression of the electrophysiological system during murine embryonic stem cell cardiac differentiation. Cellular Physiology and Biochemistry, 13, 263–270.
Van Der Heyden, M. A. G., Van Kempen, M. J. A., Tsuji, Y., Rook, M. B., Jongsma, H. J., & Opthof, T. (2003). P19 embryonal carcinoma cells: A suitable model system for cardiac electrophysiological differentiation at the molecular and functional level. Cardiovascular Research, 58, 410–422.
Fioretti, B., Pietrangelo, T., Catacuzzeno, L., & Franciolini, F. (2005). Intermediate-conductance Ca2+-activated K+ channel is expressed in C2C12 myoblasts and is downregulated during myogenesis. American Journal of Physiology. Cell Physiology, 289, C89–C96.
Kubo, Y. (1991). Comparison of initial stages of muscle differentiation in rat and mouse myoblastic and mouse mesodermal stem cell lines. Journal of Physiology, 442, 743–759.
Voets, T., Wei, L., De Smet, P., et al. (1997). Downregulation of volume-activated Cl- currents during muscle differentiation. American Journal of Physiology. Cell Physiology, 272, C667–C674.
Lesage, F., Attali, B., Lazdunski, M., & Barhanin, J. (1992). Developmental expression of voltage-sensitive K+ channels in mouse skeletal muscle and C2C12 cells. FEBS Letters, 310, 162–166.
Wieland, S. J., & Gong, Q. H. (1995). Modulation of a potassium conductance in developing skeletal muscle. American Journal of Physiology. Cell Physiology, 268, C490–C495.
Hamann, M., Widmer, H., Baroffio, A., et al. (1994). Sodium and potassium currents in freshly isolated and in proliferating human muscle satellite cells. Journal of Physiology, 475, 305–317.
Bernheim, L., Liu, J. H., Hamann, M., Haenggeli, C. A., Fischer-Lougheed, J., & Bader, C. R. (1996). Contribution of a non-inactivating potassium current to the resting membrane potential of fusion-competent human myoblasts. Journal of Physiology, 493, 129–141.
Bijlenga, P., Liu, J. H., Espinos, E., et al. (2000). T-type α1H Ca2+ channels are involved in Ca2+ signaling during terminal differentiation (fusion) of human myoblasts. Proceedings of the National Academy of Sciences of the United States of America, 97, 7627–7632.
Bijlenga, P., Occhiodoro, T., Liu, J. H., Bader, C. R., Bernheim, L., & Fischer-Lougheed, J. (1998). An ether-a-go-go K+ current, I(h-eag), contributes to the hyperpolarization of human fusion-competent myoblasts. Journal of Physiology, 512, 317–323.
Fischer-Lougheed, J., Liu, J. H., Espinos, E., et al. (2001). Human myoblast fusion requires expression of functional inward rectifier Kir2.1 channels. Journal of Cell Biology, 153, 677–685.
Liu, J. H., Bijlenga, P., Fischer-Lougheed, J., et al. (1998). Role of an inward rectifier K+ current and of hyperpolarization in human myoblast fusion. Journal of Physiology, 510, 467–476.
Messenger, E. A., & Warner, A. E. (1979). The function of the sodium pump during differentiation of amphibian embryonic neurones. Journal of Physiology, 292, 85–105.
Messenger, E. A., & Warner, A. E. (1976). The effect of inhibiting the sodium pump on the differentiation of nerve cells [proceedings]. Journal of Physiology, 263, 211P–212P.
Konig, S., Hinard, V., Arnaudeau, S., et al. (2004). Membrane hyperpolarization triggers myogenin and myocyte enhancer factor-2 expression during human myoblast differentiation. Journal of Biological Chemistry, 279, 28187–28196.
Hinard, V., Belin, D., Konig, S., Bader, C. R., & Bernheim, L. (2008). Initiation of human myoblast differentiation via dephosphorylation of Kir2.1 K+ channels at tyrosine 242. Development, 135, 859–867.
Konig, S., Béguet, A., Bader, C. R., & Bernheim, L. (2006). The calcineurin pathway links hyperpolarization (Kir2.1)-induced Ca2+ signals to human myoblast differentiation and fusion. Development, 133, 3107–3114.
Yin, Z., Tong, Y., Zhu, H., & Watsky, M. A. (2008). ClC-3 is required for LPA-activated Cl- current activity and fibroblast-to-myofibroblast differentiation. American Journal of Physiology. Cell Physiology, 294, C535–C542.
Shirihai, O., Attali, B., Dagan, D., & Merchav, S. (1998). Expression of two inward rectifier potassium channels is essential for differentiation of primitive human hematopoietic progenitor cells. Journal of Cellular Physiology, 177, 197–205.
Shirihai, O., Merchav, S., Attali, B., & Dagan, D. (1996). K+ channel antisense oligodeoxynucleotides inhibit cytokine-induced expansion of human hemopoietic progenitors. Pflugers Archiv, 431, 632–638.
Nakanishi, S., & Okazawa, M. (2006). Membrane potential-regulated Ca2+ signalling in development and maturation of mammalian cerebellar granule cells. Journal of Physiology, 575, 389–395.
Rossi, P., D’Angelo, E., Magistretti, J., Toselli, M., & Taglietti, V. (1994). Age dependent expression of high-voltage activated calcium currents during cerebellar granule cell development in situ. Pflugers Archiv, 429, 107–116.
Sato, M., Suzuki, K., Yamazaki, H., & Nakanishi, S. (2005). A pivotal role of calcineurin signaling in development and maturation of postnatal cerebellar granule cells. Proceedings of the National Academy of Sciences of the United States of America, 102, 5874–5879.
Sundelacruz, S., Levin, M., & Kaplan, D. L. (2008). Membrane potential controls adipogenic and osteogenic differentiation of mesenchymal stem cells. PLoS ONE, 3, e3737.
Echeverri, K., & Tanaka, E. M. (2002). Mechanisms of muscle dedifferentiation during regeneration. Seminars in Cell & Developmental Biology, 13, 353–360.
Odelberg, S. J. (2002). Inducing cellular dedifferentiation: A potential method for enhancing endogenous regeneration in mammals. Seminars in Cell & Developmental Biology, 13, 335–343.
Chiabrera, A., Hinsenkamp, M., Pilla, A. A., et al. (1979). Cytofluorometry of electromagnetically controlled cell dedifferentiation. Journal of Histochemistry and Cytochemistry, 27, 375–381.
Chiabrera, A., Viviani, R., Parodi, G., et al. (1980). Automated absorption image cytometry of electromagnetically exposed frog erythrocytes. Cytometry, 1, 42–48.
Harrington, D. B. (1972). Electrical stimulation of RNA and protein-synthesis in frog erythrocyte. Anatomical Record, 172, 325.
Harrington, D. B., & Becker, R. O. (1973). Electrical stimulation of RNA and protein synthesis in the frog erythrocyte. Experimental Cell Research, 76, 95–98.
Hinsenkamp, M., Chiabrera, A., Ryaby, J., Pilla, A. A., & Bassett, C. A. (1978). Cell behaviour and DNA modification in pulsing electromagnetic fields. Acta Orthopaedica Belgica, 44, 636–650.
Balana, B., Nicoletti, C., Zahanich, I., et al. (2006). 5-Azacytidine induces changes in electrophysiological properties of human mesenchymal stem cells. Cell Research, 16, 949–960.
Ravens, U. (2006). Electrophysiological properties of stem cells. Herz, 31, 123–126.
Wenisch, S., Trinkaus, K., Hild, A., et al. (2006). Immunochemical, ultrastructural and electrophysiological investigations of bone-derived stem cells in the course of neuronal differentiation. Bone, 38, 911–921.
Biagiotti, T., D’Amico, M., Marzi, I., et al. (2006). Cell renewing in neuroblastoma: electrophysiological and immunocytochemical characterization of stem cells and derivatives. Stem Cells, 24, 443–453.
Wang, K., Xue, T., Tsang, S. Y., et al. (2005). Electrophysiological properties of pluripotent human and mouse embryonic stem cells. Stem Cells, 23, 1526–1534.
Flanagan, L. A., Lu, J., Wang, L., et al. (2007). Unique dielectric properties distinguish stem cells and their differentiated progeny. Stem Cells.
Gersdorff Korsgaard, M. P., Christophersen, P., Ahring, P. K., & Olesen, S. P. (2001). Identification of a novel voltage-gated Na+ channel rNa(v)1.5a in the rat hippocampal progenitor stem cell line HiB5. Pflugers Archiv, 443, 18–30.
Heubach, J. F., Graf, E. M., Leutheuser, J., et al. (2004). Electrophysiological properties of human mesenchymal stem cells. Journal of Physiology, 554, 659–672.
Li, G. R., Sun, H., Deng, X., & Lau, C. P. (2005). Characterization of ionic currents in human mesenchymal stem cells from bone marrow. Stem Cells, 23, 371–382.
Bai, X., Ma, J., Pan, Z., et al. (2007). Electrophysiological properties of human adipose tissue-derived stem cells. American Journal of Physiology. Cell Physiology, 293(5), C1539–C1550.
Cai, J., Cheng, A., Luo, Y., et al. (2004). Membrane properties of rat embryonic multipotent neural stem cells. Journal of Neurochemistry, 88, 212–226.
Park, K. S., Jung, K. H., Kim, S. H., et al. (2007). Functional expression of ion channels in mesenchymal stem cells derived from umbilical cord vein. Stem Cells, 25, 2044–2052.
Yu, K., Ruan, D. Y., & Ge, S. Y. (2002). Three electrophysiological phenotypes of cultured human umbilical vein endothelial cells. General Physiology and Biophysics, 21, 315–326.
Baksh, D., Song, L., & Tuan, R. S. (2004). Adult mesenchymal stem cells: Characterization, differentiation, and application in cell and gene therapy. Journal of Cellular and Molecular Medicine, 8, 301–316.
Levin, M. (2007). Large-scale biophysics: Ion flows and regeneration. Trends in Cell Biology, 17, 261–270.
Constantinescu, S. N. (2000). Stem cell generation and choice of fate: Role of cytokines and cellular microenvironment. Journal of Cellular and Molecular Medicine, 4, 233–248.
Bianchi, G., Muraglia, A., Daga, A., Corte, G., Cancedda, R., & Quarto, R. (2001). Microenvironment and stem properties of bone marrow-derived mesenchymal cells. Wound Repair Regen, 9, 460–466.
Kasemeier-Kulesa, J. C., Teddy, J. M., Postovit, L. M., et al. (2008). Reprogramming multipotent tumor cells with the embryonic neural crest microenvironment. Developmental Dynamics, 237, 2657–2666.
Heese, O., Disko, A., Zirkel, D., Westphal, M., & Lamszus, K. (2005). Neural stem cell migration toward gliomas in vitro. Neuro-oncology, 7, 476–484.
Jeon, J. Y., An, J. H., Kim, S. U., Park, H. G., & Lee, M. A. (2008). Migration of human neural stem cells toward an intracranial glioma. Experimental & Molecular Medicine, 40, 84–91.
Quesenberry, P. J., & Becker, P. S. (1998). Stem cell homing: Rolling, crawling, and nesting. Proceedings of the National Academy of Sciences of the United States of America, 95, 15155–15157.
Whetton, A. D., & Graham, G. J. (1999). Homing and mobilization in the stem cell niche. Trends in Cell Biology, 9, 233–238.
Krause, D. S., Theise, N. D., Collector, M. I., et al. (2001). Multi-organ, multi-lineage engraftment by a single bone marrow-derived stem cell. Cell, 105, 369–377.
Penn, M. S., Zhang, M., Deglurkar, I., & Topol, E. J. (2004). Role of stem cell homing in myocardial regeneration. International Journal of Cardiology, 95(Suppl 1), S23–S25.
Chute, J. P. (2006). Stem cell homing. Current Opinion in Hematology, 13, 399–406.
Sanchez Alvarado, A. (2004). Planarians. Current Biology, 14, R737–R738.
Reddien, P. W., & Sanchez Alvarado, A. (2004). Fundamentals of planarian regeneration. Annual Review of Cell and Developmental Biology, 20, 725–757.
Oviedo, N., & Levin, M. (2008). The planarian regeneration model as a context for the study of drug effects and mechanisms. In R. B. Raffa & S. M. Rawls (Eds.), Planaria: A model for drug action and abuse. Austin: RG Landes Co.
Sanchez Alvarado, A. (2003). The freshwater planarian Schmidtea mediterranea: Embryogenesis, stem cells and regeneration. Current Opinion in Genetics and Development, 13, 438–444.
Salo, E., & Baguna, J. (1985). Cell movement in intact and regenerating planarians. Quantitation using chromosomal, nuclear and cytoplasmic markers. Journal of Embryology and Experimental Morphology, 89, 57–70.
Oviedo, N. J., Pearson, B. J., Levin, M., & Sanchez Alvarado, A. (2008). Planarian PTEN homologs regulate stem cells and regeneration through TOR signaling. Disease Models & Mechanisms, 1, 131–143.
Nogi, T., & Levin, M. (2005). Characterization of innexin gene expression and functional roles of gap-junctional communication in planarian regeneration. Developmental Biology, 287, 314–335.
Oviedo, N. J., & Levin, M. (2007). smedinx-11 is a planarian stem cell gap junction gene required for regeneration and homeostasis. Development, 134, 3121–3131.
Wong, R. C., Pera, M. F., & Pebay, A. (2008). Role of gap junctions in embryonic and somatic stem cells. Stem Cell Reviews, 4, 283–292.
Spray, D., Harris, A., & Bennett, M. (1981). Equilibrium properties of a voltage-dependent junctional conductance. Journal of General Physiology, 77, 77–93.
Harris, A., Spray, D., & Bennett, M. (1983). Control of intercellular communication by voltage dependence of gap junctional conductance. Journal of Neuroscience, 3, 79–100.
Menichella, D. M., Majdan, M., Awatramani, R., et al. (2006). Genetic and physiological evidence that oligodendrocyte gap junctions contribute to spatial buffering of potassium released during neuronal activity. Journal of Neuroscience, 26, 10984–10991.
Verselis, V., Trexler, E., Bargiello, T., & Bennett, M. (1997). Studies of voltage gating of gap junctions and hemichannels formed by connexin proteins. In R. Latorre, J. Saez (Eds.), From ion channels to cell-to-cell conversations (pp. 323–347). New York.
Morokuma, J., Blackiston, D., Adams, D. S., Seebohm, G., Trimmer, B., & Levin, M. (2008). Modulation of potassium channel function confers a hyperproliferative invasive phenotype on embryonic stem cells. Proceedings of the National Academy of Sciences of the United States of America, 105, 16608–16613.
Djamgoz, M. B. A., Mycielska, M., Madeja, Z., Fraser, S. P., & Korohoda, W. (2001). Directional movement of rat prostate cancer cells in direct-current electric field: Involvement of voltage-gated Na+ channel activity. Journal of Cell Science, 114, 2697–2705.
Brackenbury, W. J., & Djamgoz, M. B. (2006). Activity-dependent regulation of voltage-gated Na+ channel expression in Mat-LyLu rat prostate cancer cell line. Journal of Physiology, 573, 343–356.
Gruler, H., & Nuccitelli, R. (1991). Neural crest cell galvanotaxis: new data and a novel approach to the analysis of both galvanotaxis and chemotaxis. Cell Motility and the Cytoskeleton, 19, 121–133.
Nuccitelli, R., & Erickson, C. A. (1983). Embryonic cell motility can be guided by physiological electric fields. Experimental Cell Research, 147, 195–201.
Nuccitelli, R., & Smart, T. (1989). Extracellular calcium levels strongly influence neural crest cell galvanotaxis. Biological Bulletin, 176, 130–135.
Adams, D. S., Robinson, K. R., Fukumoto, T., et al. (2006). Early, H+-V-ATPase-dependent proton flux is necessary for consistent left-right patterning of non-mammalian vertebrates. Development, 133, 1657–1671.
Denker, S. P., & Barber, D. L. (2002). Cell migration requires both ion translocation and cytoskeletal anchoring by the Na-H exchanger NHE1. Journal of Cell Biology, 159, 1087–1096.
Levin, M., Buznikov, G. A., & Lauder, J. M. (2006). Of minds and embryos: Left-right asymmetry and the serotonergic controls of pre-neural morphogenesis. Developmental Neuroscience, 28, 171–185.
Shi, H., Halvorsen, Y. D., Ellis, P. N., Wilkison, W. O., & Zemel, M. B. (2000). Role of intracellular calcium in human adipocyte differentiation. Physiological Genomics, 2000, 75–82.
Zayzafoon, M. (2006). Calcium/calmodulin signaling controls osteoblast growth and differentiation. Journal of Cellular Biochemistry, 97, 56–70.
Munaron, L., Antoniotti, S., & Lovisolo, D. (2004). Intracellular calcium signals and control of cell proliferation: How many mechanisms? Journal of Cellular and Molecular Medicine, 8, 161–168.
Whitaker, M. (2006). Calcium microdomains and cell cycle control. Cell Calcium, 40, 585–592.
Soliman, E. M., Rodrigues, M. A., Gomes, D. A., et al. (2009). Intracellular calcium signals regulate growth of hepatic stellate cells via specific effects on cell cycle progression. Cell Calcium, 45, 284–292.
Palma, V., Kukuljan, M., & Mayor, R. (2001). Calcium mediates dorsoventral patterning of mesoderm in Xenopus. Current Biology, 11, 1606–1610.
Sun, S., Liu, Y., Lipsky, S., & Cho, M. (2007). Physical manipulation of calcium oscillations facilitates osteodifferentiation of human mesenchymal stem cells. FASEB Journal, 21, 1472–1480.
Trollinger, D. R., Isseroff, R. R., & Nuccitelli, R. (2002). Calcium channel blockers inhibit galvanotaxis in human keratinocytes. Journal of Cellular Physiology, 193, 1–9.
Albrieux, M., & Villaz, M. (2000). Bilateral asymmetry of the inositol trisphosphate-mediated calcium signaling in two-cell ascidian embryos. Biology of the Cell, 92, 277–284.
Linask, K. K., Han, M. D., Artman, M., & Ludwig, C. A. (2001). Sodium-calcium exchanger (NCX-1) and calcium modulation: NCX protein expression patterns and regulation of early heart development. Developmental Dynamics, 221, 249–264.
McGrath, J., Somlo, S., Makova, S., Tian, X., & Brueckner, M. (2003). Two populations of node monocilia initiate left-right asymmetry in the mouse. Cell, 114, 61–73.
Raya, A., Kawakami, Y., Rodriguez-Esteban, C., et al. (2004). Notch activity acts as a sensor for extracellular calcium during vertebrate left-right determination. Nature, 427, 121–128.
Schneider, I., Houston, D. W., Rebagliati, M. R., & Slusarski, D. C. (2007). Calcium fluxes in dorsal forerunner cells antagonize {beta}-catenin and alter left-right patterning. Development.
Slusarski, D. C., & Pelegri, F. (2007). Calcium signaling in vertebrate embryonic patterning and morphogenesis. Developmental Biology, 307, 1–13.
Webb, S. E., & Miller, A. L. (2000). Calcium signalling during zebrafish embryonic development. Bioessays, 22, 113–123.
Jaffe, L. F. (1999). Organization of early development by calcium patterns. Bioessays, 21, 657–667.
Jaffe, L. (1995). Calcium waves and development. In Calcium waves, gradients and oscillations (pp. 4–17). Chichester: CIBA Foundation.
Konig, S., Beguet, A., Bader, C. R., & Bernheim, L. (2006). The calcineurin pathway links hyperpolarization (Kir2.1)-induced Ca2+ signals to human myoblast differentiation and fusion. Development, 133, 3107–3114.
Nilius, B., Schwarz, G., & Droogmans, G. (1993). Control of intracellular calcium by membrane potential in human melanoma cells. American Journal of Physiology, 265, C1501–C1510.
Nilius, B., & Wohlrab, W. (1992). Potassium channels and regulation of proliferation of human melanoma cells. Journal of Physiology, 445, 537–548.
Sasaki, M., Gonzalez-Zulueta, M., Huang, H., et al. (2000). Dynamic regulation of neuronal NO synthase transcription by calcium influx through a CREB family transcription factor-dependent mechanism. Proceedings of the National Academy of Sciences of the United States of America, 97, 8617–8622.
Bidaud, I., Mezghrani, A., Swayne, L. A., Monteil, A., & Lory, P. (2006). Voltage-gated calcium channels in genetic diseases. Biochimica et Biophysica Acta, 1763, 1169–1174.
Cherubini, A., Hofmann, G., Pillozzi, S., et al. (2005). Human ether-a-go-go-related gene 1 channels are physically linked to beta1 integrins and modulate adhesion-dependent signaling. Molecular Biology of the Cell, 16, 2972–2983.
Arcangeli, A., & Becchetti, A. (2006). Complex functional interaction between integrin receptors and ion channels. Trends in Cell Biology, 16, 631–639.
Liu, J., DeYoung, S. M., Zhang, M., Cheng, A., & Saltiel, A. R. (2005). Changes in integrin expression during adipocyte differentiation. Cell Metabolism, 2, 165–177.
Meyers, V. E., Zayzafoon, M., Gonda, S. R., Gathings, W. E., & McDonald, J. M. (2004). Modeled microgravity disrupts collagen I/integrin signaling during osteoblastic differentiation of human mesenchymal stem cells. Journal of Cellular Biochemistry, 93, 697–707.
Nesti, L. J., Caterson, E. J., Wang, M., et al. (2002). TGF-β1 calcium signaling increases α5 integrin expression in osteoblasts. Journal of Orthopaedic Research, 20, 1042–1049.
Iwasaki, H., Murata, Y., Kim, Y., et al. (2008). A voltage-sensing phosphatase, Ci-VSP, which shares sequence identity with PTEN, dephosphorylates phosphatidylinositol 4, 5-bisphosphate. Proceedings of the National Academy of Sciences of the United States of America, 105, 7970–7975.
Murata, Y., Iwasaki, H., Sasaki, M., Inaba, K., & Okamura, Y. (2005). Phosphoinositide phosphatase activity coupled to an intrinsic voltage sensor. Nature, 435, 1239–1243.
Murata, Y., & Okamura, Y. (2007). Depolarization activates the phosphoinositide phosphatase Ci-VSP, as detected in Xenopus oocytes coexpressing sensors of PIP2. Journal of Physiology, 583, 875–889.
Murata, Y., Iwasaki, H., Sasaki, M., Inaba, K., & Okamura, Y. (2005). Phosphoinositide phosphatase activity coupled to an intrinsic voltage sensor. Nature, 435, 1239–1243.
Adams, D. S. (2008). A new tool for tissue engineers: Ions as regulators of morphogenesis during development and regeneration. Tissue Engineering. Part A, 14, 1461–1468.
Chen, J. G., & Rudnick, G. (2000). Permeation and gating residues in serotonin transporter. Proceedings of the National Academy of Sciences of the United States of America, 97, 1044–1049.
Fukumoto, T., Blakely, R., & Levin, M. (2005). Serotonin transporter function is an early step in left-right patterning in chick and frog embryos. Developmental Neuroscience, 27, 349–363.
Hegle, A. P., Marble, D. D., & Wilson, G. F. (2006). A voltage-driven switch for ion-independent signaling by ether-a-go-go K+ channels. Proceedings of the National Academy of Sciences of the United States of America, 103, 2886–2891.
Yang, S. J., Liang, H. L., Ning, G., & Wong-Riley, M. T. (2004). Ultrastructural study of depolarization-induced translocation of NRF-2 transcription factor in cultured rat visual cortical neurons. European Journal of Neuroscience, 19, 1153–1162.
Li, L., Liu, F., Salmonsen, R. A., et al. (2002). PTEN in neural precursor cells: Regulation of migration, apoptosis, and proliferation. Molecular and Cellular Neurosciences, 20, 21–29.
Poo, M. M., & Robinson, K. R. (1977). Electrophoresis of concanavalin-a receptors along embryonic muscle-cell membrane. Nature, 265, 602–605.
Cooper, M. S., Miller, J. P., & Fraser, S. E. (1989). Electrophoretic repatterning of charged cytoplasmic molecules within tissues coupled by gap junctions by externally applied electric fields. Developmental Biology, 132, 179–188.
Fang, K. S., Ionides, E., Oster, G., Nuccitelli, R., & Isseroff, R. R. (1999). Epidermal growth factor receptor relocalization and kinase activity are necessary for directional migration of keratinocytes in DC electric fields. Journal of Cell Science, 112, 1967–1978.
Giugni, T. D., Braslau, D. L., & Haigler, H. T. (1987). Electric field-induced redistribution and postfield relaxation of epidermal growth factor receptors on A431 cells. Journal of Cell Biology, 104, 1291–1297.
Stollberg, J., & Fraser, S. E. (1988). Acetylcholine receptors and concanavalin A-binding sites on cultured Xenopus muscle cells: electrophoresis, diffusion, and aggregation. Journal of Cell Biology, 107, 1397–1408.
Orida, N., & Poo, M. M. (1978). Electrophoretic movement and localisation of acetylcholine receptors in the embryonic muscle cell membrane. Nature, 275, 31–35.
Fukumoto, T., Kema, I. P., & Levin, M. (2005). Serotonin signaling is a very early step in patterning of the left-right axis in chick and frog embryos. Current Biology, 15, 794–803.
Woodruff, R., & Telfer, W. (1980). Electrophoresis of proteins in intercellular bridges. Nature, 286, 84–86.
Korohoda, W., Mycielska, M., Janda, E., & Madeja, Z. (2000). Immediate and long-term galvanotactic responses of Amoeba proteus to dc electric fields. Cell Motility and the Cytoskeleton, 45, 10–26.
Tao, Y., Yan, D., Yang, Q., Zeng, R., & Wang, Y. (2006). Low K+ promotes NF-kappaB/DNA binding in neuronal apoptosis induced by K+ loss. Molecular and Cellular Biology, 26, 1038–1050.
Gillies, R., Martinez-Zaguilan, R., Peterson, E., & Perona, R. (1992). Role of intracellular pH in mammalian cell proliferation. Cellular Physiology and Biochemistry, 2, 159–179.
Uzman, J. A., Patil, S., Uzgare, A. R., & Sater, A. K. (1998). The role of intracellular alkalinization in the establishment of anterior neural fate in Xenopus. Developmental Biology, 193, 10–20.
Schuldiner, S., & Rozengurt, E. (1982). Na+/H+ antiport in Swiss 3 T3 cells: Mitogenic stimulation leads to cytoplasmic alkalinization. Proceedings of the National Academy of Sciences of the United States of America, 79, 7778–7782.
Zhong, M., Kim, S. J., & Wu, C. (1999). Sensitivity of Drosophila heat shock transcription factor to low pH. Journal of Biological Chemistry, 274, 3135–3140.
Lin, H., Xiao, J., Luo, X., et al. (2007). Overexpression HERG K(+) channel gene mediates cell-growth signals on activation of oncoproteins SP1 and NF-kappaB and inactivation of tumor suppressor Nkx3.1. Journal of Cellular Physiology, 212, 137–147.
Chudotvorova, I., Ivanov, A., Rama, S., et al. (2005). Early expression of KCC2 in rat hippocampal cultures augments expression of functional GABA synapses. Journal of Physiology, 566, 671–679.
Burrone, J., O’Byrne, M., & Murthy, V. N. (2002). Multiple forms of synaptic plasticity triggered by selective suppression of activity in individual neurons. Nature, 420, 414–418.
Beech, J. A. (1997). Bioelectric potential gradients may initiate cell cycling: ELF and zeta potential gradients may mimic this effect. Bioelectromagnetics, 18, 341–348.
Redmann, K., Jenssen, H. L., & Kohler, H. J. (1974). Experimental and functional changes in transmembrane potential and zeta potential of single cultured cells. Experimental Cell Research, 87, 281–289.
Sherbet, G. V., & Lakshmi, M. S. (1974). The surface properties of some human intracranial tumour cell lines in relation to their malignancy. Oncology, 29, 335–347.
James, A. M., Ambrose, E. J., & Lowick, J. H. (1956). Differences between the electrical charge carried by normal and homologous tumour cells. Nature, 177, 576–577.
Weihua, Z., Tsan, R., Schroit, A. J., & Fidler, I. J. (2005). Apoptotic cells initiate endothelial cell sprouting via electrostatic signaling. Cancer Research, 65, 11529–11535.
Acknowledgements
S.S. would like to thank the NSF for funding through the Graduate Research Fellowship Program. D.K. is supported by the NIH through the Tissue Engineering Resource Center (P41 EB002520). M.L. is supported by grants from the NHTSA (DTNH22-06-G-00001) and NIH (GM078484, HD055850-01).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Sundelacruz, S., Levin, M. & Kaplan, D.L. Role of Membrane Potential in the Regulation of Cell Proliferation and Differentiation. Stem Cell Rev and Rep 5, 231–246 (2009). https://doi.org/10.1007/s12015-009-9080-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12015-009-9080-2