
Abstract
Study was conducted to recover proteins from pangas (Pangasius pangasius) processing waste (fillet frames) using pH shift method and to characterize the recovered isolates. pH 2.0 from acidic range and pH 13.0 from alkaline range were found to have maximum protein recovery (p < 0.05). During the recovery process, acidic pH (pH 2.0) was found to have minimal effect on proteins resulting in more stable isolates and strong protein gels. Alkaline pH (pH 13.0) caused protein denaturation resulting in less stable proteins and poor gel network. Both acidic and alkaline-aided processing caused significant (p < 0.05) reductions in total lipid, myoglobin, and pigment content thus by resulting in whiter protein isolates and gels. The content of total essential amino acids increased during pH shift processing, indicating the enrichment of essential amino acids. No microbial counts were detected in any of the isolates prepared using acid and alkaline extraction methods. pH shift processing was found to be promising in the utilization of fish processing waste for the recovery of functional proteins from pangas processing waste thus by reducing the supply demand gap as well pollution problems.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Pangas (Pangasius pangasius) is one of the important cultured and marketed fish species in India. Processing of pangas generates high amount of waste, which sometimes goes up to 50% of the total body weight, polluting local water bodies and lands. During the filleting, which is the most commonly used process for pangas, considerable amount of meat will be retained with the fillet frames. This leftover meat contains valuable components which can be used for edible and non-edible purposes. In view of addressing pollution problems associated with pangas processing waste, to minimize supply-demand gap and to meet increasing demand for nutritious food, these valuable compounds from the processing waste should be recovered and used for edible purpose.
Initial efforts by a few researchers (Niki et al. 1985; Huang et al. 1997, etc.) to recover valuable and nutritional components from surimi wash water led to unsatisfactory results due to the loss of sarcoplasmic proteins and associated poor recovery yields. After a few years, Hultin and Kelleher (1999, 2000) patented acid and alkaline solubilization process as a most promising method to isolate fish muscle proteins with improved recovery yields, functionality, and stability. Thereafter, an appreciable amount of work was done on recovery of proteins from fish, fish portions, fish processing waste, and by-products using acid and alkaline solubilization process (Cortes-Ruis et al. 2001; Kristinsson and Demir 2003; Kristinsson and Ingadottir 2006; Fatin et al. 2015; Surasani et al. 2017). Proteins recovered using different methods from different sources were found to be different in process yields and functionality.
Fish processing in India generates high amount of wastes, which can be a potential source of valuable components (Surasani et al. 2017). Utilization of these processing wastes for human consumption will reduce pollution problems as well as demand supply gap and product cost (Surasani 2017). Pangas fish being one of the most important commercial species in India, the objectives of the study were set to recover proteins from pangas fillet frames using acid and alkaline solubilization method. Protein isolates obtained through acid and alkaline processing were also studied for their characteristics and functionality.
Materials and methods
Raw material
Pangas processing waste (fillet frames) were purchased from local market (Sherpur, Ludhiana, India) and transported to the laboratory on ice followed by grinding using a laboratory mincer to a fine and thick paste. This thick paste was used as raw material and was transferred to zipped plastic bags followed by storage at − 20 °C until further use. All raw material processing and sample preparation steps were performed in a cold room at 4 °C or on ice to maintain temperature below 5 °C.
Preparation of homogenate
Ground pangas fillet frames were mixed with cold deionized water (4 °C, 1:6, w:w) followed by homogenization (Ultra-Turrax, T 25, Janke & Kunkel GmbH & Co. KG Staufen, Germany) for 60 s (2 × 30 s). The homogenate was transferred into a plastic beaker and was adjusted to the desirable pH using 2 M HCl or 2 M NaOH in continuous stirring condition (Surasani et al. 2017). The pH of the homogenate was kept constant throughout the process by adding desired diluted acid or alkali.
Protein solubility
Solubility curve
A protein solubility curve was drawn using the solubility of proteins at each pH from pH 2.0 to 13.0 with 1.0 unit interval. Homogenates from the same raw material were divided into sets to enable the pH adjustment of acidic and alkaline range. Following the pH adjustment, 30 g of homogenate at each pH was taken into 50 ml centrifuge tubes and centrifuged at 5,000×g for 20 min using refrigerated centrifuge (Sorvall ST 16R, Thermo Fisher Scientific, Germany) to separate insoluble material from soluble proteins. Sample (2 g) was drawn at each pH before and after the centrifugation to calculate the total protein content. Biuret Method (Robinson and Hogden 1940) was used for analyzing protein content after homogenizing with a hand-held Tissue Tearor (Biospec Products, Inc., Bartlesville, UK) for 20 s on speed 7. For analyzing protein content, sample was diluted fivefold with cold deionized water adjusted to pH 11.0, wherever applicable.
Solubility after isoelectric precipitation
Following the protein solubility curve, homogenate was adjusted to the pH with maximum protein recovery in acidic and alkaline side (pH 2.0 and pH 13.0) in a slow and continuous stirring condition using a magnetic stirrer. pH-adjusted homogenates were kept at 4 °C for 60 min followed by centrifugation to separate insoluble matter. Supernatants obtained after first centrifugation were adjusted to precipitation pH of 5.5 followed by precipitation for 30 min at 4 °C and centrifugation at 5,000×g for 20 min. The amount of soluble protein after the 2nd centrifugation cycle was determined by taking a 2-g sample before and after centrifugation and analyzing by Biuret method.
Protein measurements
Biuret method was used to determine the protein content of homogenate as well as supernatants. The cloudiness due to lipids in the solution was reduced by the addition of 10% deoxycholic acid. Absorbance was read at 540 nm using a T90+ UV/VIS spectrometer (PG Instruments Ltd., India), and protein concentrations were calculated from a standard curve prepared with bovine serum albumin (BSA) with a concentration of 1–10 mg/ml.
Proteins recovery
Protein recovery was expressed as theoretical and actual recovery (%). Both theoretical and actual recoveries were categorized into (a) recovery after 1st centrifugation, (b) recovery after 2nd centrifugation, and (c) process recovery.
Theoretical recoveries were calculated by the formulas given below:
where A is the soluble protein in initial homogenate, B is the soluble protein in supernatant after first centrifugation, and C is the soluble protein in supernatant after second centrifugation.
Actual recoveries were calculated by the formulas given below:
where A is the soluble protein in initial homogenate, WA is the weight of the homogenate, B is the soluble protein in supernatant after first centrifugation, WB is the weight of the supernatant after first centrifugation, C is the soluble protein in supernatant after second centrifugation, WC is the weight of the supernatant after second centrifugation.
Lipid content and its removal
Lipid content of the homogenate and isolate was determined using a Soxhlet as per the method of AOAC 920.39B (2000). Lipid content (dry weight basis) was calculated using the following equation:
The reduction in lipid content was calculated using the equation:
Total myoglobin content and its removal
Total pigment content of homogenate and isolate was determined as per the methodology used by Chaijan et al. (2006). Two grams of sample was taken in a 50-ml centrifuge tube and was added with 20 ml of 40 mM, pH 6.8 phosphate buffer. It was followed by homogenization (3000×g for 30 min at 4 °C) and filtration using Whatman no. 1 paper. Supernatant was added with 0.2 ml of 1% (w/v) sodium dithionite followed by absorbance measurement at 555 nm using phosphate buffer (pH 6.8) as blank. Myoglobin content was calculated as below:
where A = absorbance, F = dilution factor, WS = weight of sample in g, 7.6 = millimolar extinction coefficient, 16.111 = molecular weight.
Myoglobin removal (%) was calculated as:
Total pigment content and its removal
Total pigment content was determined as per the method described by Rawdkuen et al. (2009). One gram of sample was mixed with 9 ml of acetone acid (9% acetone, 8% distilled water, and 2% hydrochloric acid) followed by incubation for 1 h. The sample was filtered using Whatman no. 1 filter paper followed by absorbance reading at 640 nm by using acetone as blank. Using the ratio of 680 and the weight of the test portion, the overall concentration of heme pigments (hematin) in mg/100 g muscle tissue was calculated.
(680 is the conversion factor to convert the total pigments and the extracted nitric acid pigments into parts per million of hematin).
Total pigment removal (%) was calculated as:
Color analysis
Color of the fillet frame homogenate, protein isolates, and protein gel was determined using HunterLab (Virginia, USA) as described by Lubana et al. (2016). Whiteness of the raw material as well as isolates and gels was calculated after obtaining lightness (L*), Red/Greenness (a*) and Yellow/Blueness (b*) values.
Foaming properties
Foaming capacity and foaming stability of protein isolates obtained at pH 2.0 and 13.0 were measured as described by Sathe et al. (1982). About 1 g of protein sample was added into 50-ml distilled water followed by heating at 60 °C to dissolve the proteins. Foam was prepared by homogenizing the protein solution at 9000 g for 5 min using homogenizer. Following homogenization solution was poured gently into a measuring cylinder. Foaming capacity of isolates was calculated as the volume ratio of foam to the original liquid volume and expressed as percentage. The foam stability was calculated as the ratio of the initial volume of foam to the volume of foam after 30 min.
SDS-PAGE analysis
Proteins in the initial homogenate as well as supernatants after 1st and 2nd centrifugation cycle were analyzed by SDS-PAGE (Laemmli 1970). Harlow and Lane (1988) methodology was used for preparing samples, electrophoresis reagents and assembly with minor modifications. Protein samples were mixed with appropriate volumes of × 5 sample loading buffer (SDFCL, India), followed by incubation at 95 °C for 5 min and loading into 5% stacking gel. Resolving of proteins was done on 12% gel with constant current of 20 mA for 3–4 h. Four micrograms of broad range marker (SDFCL, India) was also loaded on each gel. After completion of electrophoresis, overnight staining with Coomassie Brilliant Blue R250 and destaining were carried out as per standard protocols.
Aminoacid analysis
Amino acid composition of pangas frame homogenate and protein isolate was determined by reverse phase HPLC on Water Picotag TM amino acids analysis system, after hydrolyzing the protein with 6 N HCl to yield free amino acid (Bidlingmeyer et al. 1984). The amino acid composition of carp protein, including the starting material, was determined and expressed as g/100 g protein.
Microbial quality analysis
Microbial quality of the initial homogenate as well as isolates was determined using total plate count technique as described in AOAC (2000). The total microbial count was expressed as CFU/ml or CFU/g.
Gel preparation
Protein isolate obtained by acid and alkaline processing was cut into small pieces of 1-cm thickness and minced using a mincer. The moisture content of isolate was adjusted to 80% (w/w) followed by addition of salt (2.5%, w/w). The mixture was finely chopped and stuffed into polyvinylidine chloride casing with 2.5-cm diameter. Both ends were sealed tightly followed by incubation at 40 °C for 30 min and then heating at 90 °C for 20 min (Kristinsson and Ingadottir 2006). Then, the gel was kept in chilled water for 30 min followed by storage at 4 °C overnight prior to analysis.
Gel quality analysis
Folding test
Gel prepared using protein isolate has been tested as per the method of Kudo et al. (1973). Approximately 3-mm slices were cut from the gel of isolates and folded by hand at room temperature. The ability of gels to fold was assessed using a five-point system. 5: No crack occurred even after folding into four, 4: no crack occurred when folded in to two but cracked when folded into four, 3: no crack occurred when folded into two but broken into two when folded into four, 2: cracked when folded into two, 1: broken into two when folded into two.
Texture profile analysis (TPA)
Texture profile analysis (two-cycle compression test) of gels made of protein isolates obtained at pH 2.0 and pH 13.0 were performed with a slight modification of the method described by Surasani (2017). Gels with uniform size of 2.5 cm × 2.5 cm (diameter × height) were used for TPA analysis using a Taxt-plus Texture Analyzer (Stable Micro Systems Ltd., Surrey, UK), with a 5-kg load cell. A 75-mm diameter compression platen was used with a pre- and post-speed of 1.0 mm/s and test speed of 0.5 mm/s. Strain mode was used for compressing the gels to 50% of their original height at 30 °C temperature and 10-g force. A 10-s time gap was given between first and second bites. Every lot was tested to obtain six measurements and the average value was reported for each parameter. A data acquisition rate of 200 pps was used to generate force-time graph, and textural parameters were calculated using taxt-plus software provided with the instrument.
Cutting test
Cutting test for the gels made of acid- and alkali-processed isolates was performed with a slight modification of the method described by Reddy (2016). A blade set (HDP/BSW) was used with a speed of 2 mm/s throughout the process. The gels were compressed to a distance of 10 mm at 30 °C and 20-g force. Every lot was tested to obtain six measurements, and the average value was reported for each parameter. A data acquisition rate of 200 pps was used to generate force-time graph and textural parameters were calculated using taxt-plus software provided with the instrument.
Expressible moisture
Expressible moisture was determined as per the method given by Feng and Hultin (2001). Gel sample of 3-mm thickness was placed between five layers of Whatman filter papers and was pressed by a metal weight of 3000 g standard weight for 1 min. The weight of the gel before and after pressing was recorded, and the expressible moisture content was calculated using;
Statistics analysis
Results are expressed as mean ± SD. Analysis of variance (ANOVA) and t test were used to determine significant differences (P < 0.05) between treatments. All results were analyzed using the SPSS Version 20 software.
Results and discussion
Effect of pH on pangas protein solubility and recovery
Solubility
Solubility and recovery yields of pangas proteins at different pHs (pH 1.0–pH 13.0) are given in Table 1. Protein solubility was maximum at extreme pHs in both acidic and alkaline sides, while it decreased significantly towards neutral pH being maximum at pH 2.0 (16.87 mg/ml) in acidic range and pH 12.0 (16.21 mg/ml) in alkaline range (p < 0.05). The sudden shift in solubility was observed at pH 4.0 to pH 3.0 and pH 8.0 to pH 9.0. pH 2.0 from acidic range and pH 13.0 from alkaline range had highest theoretical as well as actual protein recoveries. The minimum solubility of fish proteins at pH 5 to 5.5 might be due to myosin, which is a major myofibrillar protein with isoelectric pH between 5.0 and 5.5. Above and below this pH range, the proteins will get more positive or more negative charge, thus by increasing the solubilization due to electrostatic repulsions (Hamm 1994; Kelleher and Hultin 2000). The sudden increase in solubility might be because of more ionizable groups with pKa values between those pHs (Undeland et al. 2002). Similar results were reported by Chen et al. (2007) and Surasani et al. (2017).
Process recovery
Viability of any extraction process depends on process yields. Theoretical and actual process recoveries during first and second centrifugation of acid and alkali processed isolates are given in Table 2. Acid- and alkaline-aided processing of proteins from pangas fillet frames resulted in a total process yield of 59.16% and 68.52%, respectively. No significant difference was observed between the acid- and alkaline-processed isolates during first centrifugation step (p > 0.05), while recovery during second centrifugation step and total process yield were significantly different (p < 0.05). Tian et al. (2016) found protein recoveries of 87.6 and 76.3% at pH 12.5 and pH 2.5 during pH shift processing of common carp proteins. Batista et al. (2007) obtained 73.0 and 77.0% yields during acid and alkaline processing of sardine proteins. Shabanpour et al. (2015) reported protein recoveries of 70.2 and 64.5% for common carp and 75.1 and 80.8% for of silver carp during acid and alkaline processing. Similar findings were also reported by Kristinsson and Ingadottir (2006) during acid and alkaline processing of tilapia proteins.
Characteristics of isolates
Color
Color values of protein isolates obtained by acid and alkaline processing and gels made of them are given in Table 3. Isolates made by acid and alkaline processing were found to have high low redness (a) and yellowness (b) than the raw material used. Acid processed isolates had more lightness and whiteness compared to raw material used (p < 0.05). Protein gels had high L and whiteness values (p < 0.05) than the raw material as well as isolates they are made of. The order of whiteness values was acid processed isolate gel> acid processed isolate> alkali processed isolate gel> alkali processed isolate> homogenate (p < 0.05). Panpipat and Chaijan (2016) reported that pH shift processing resulted in improved whiteness of isolates by increasing lightness (L) and reducing redness (a) of isolates. They found higher whiteness in acid-processed isolates than alkaline-processed isolates. Higher lightness in acid-processed isolates might be due to a greater removal of pigments, hemoglobin, myoglobin, and melanin. Acid process results in dissociation of hemoglobin forming colorless heme and globin thus by increasing the whiteness (Panpipat and Chaijan 2016). Alkaline process possible resulted in Maillard reaction, causing increased b (yellowness discoloration) value of isolates (Panpipat and Chaijan 2016). Similar findings were reported by Fatin et al. (2015) during pH shift processing of Japanese scad proteins. Color of isolates and their gels in the present study followed the same trend as reported by earlier studies. Rawdkuen et al. (2009) reported higher whiteness values for acid processed kamaboko and modori gels compared to alkaline processed gels from minced meat of tilapia. Nolsøe et al. (2011) found isolates having higher whiteness than starting mince and surimi gels with higher whiteness than the isolates.
Total lipid, myoglobin and pigment content
pH shift processing showed significant effect on total lipid, myoglobin, and pigment content of acid- and alkali-processed isolates (Table 4.). Acid-aided processing caused higher reductions in total lipid and pigment content than alkaline processed isolates (p < 0.05).
Acid and alkaline processes resulted in a significant reductions (p < 0.05) of 97.11 and 90.33% in lipid content. The lower lipid removing ability in alkaline processing might be due to reaction between alkali and lipids to form soap, which could co-precipitate during precipitation process (Panpipat and Chaijan 2016). Batista et al. (2007) reported lipid reductions of 51.0 and 65.3% during acid and alkaline solubilization of sardine mince. Rawdkuen et al. (2009) reported lipid reductions of 85.2% for tilapia muscle concentrate, and Kristinsson et al. (2005) reported lipid reduction of 85.4% for catfish concentrate. Lipid reductions in the present study were higher than some of the previous reports. Batista et al. (2007) and Freitas et al. (2015) obtained lipid reductions of 95.3, 95.31, and 95.12% for sardine concentrates, anchovy muscle, and whitemouth croaker muscle, respectively, which are similar to those of concentrates obtained in this study.
Total myoglobin content was reduced significantly by 93.77 and 93.48% during acid and alkaline processing of pangas proteins (p < 0.05). Similar findings were reported by Shabanpour et al. (2015) during protein recovery from common carp and silver carp. Myoglobin could be removed efficiently during alkaline-aided processing of sardine and mackerel muscles (Chaijan et al. 2006). Myoglobin contents were decreased efficiently in pH shift processing when compared to conventional surimi processing (Rawdkuen et al. 2009). In general, myoglobin extracting efficiency depends on species, muscle type, storage time, and washing process (Chaijan et al. 2006).
Acid and alkaline processes resulted in a significant total pigment reduction of 68.84 and 58.27% (p < 0.05). Similar findings were reported by Panpipat and Chaijan (2016) during protein recovery from big eye snapper head byproduct. High pigment removal in acid-aided processing might be due to the degradation of haem pigments at extreme acid condition rather than removal of them from the isolate (Panpipat and Chaijan 2016). At acidic pHs, the proton-catalyzed displacement process and a protoporphyrin IX ring destruction may be responsible for promoting haem loss which limited total pigment detection (Chaijan and Undeland 2015). Similar findings were also reported by Jafarpour et al. (2013) and Shabanpour et al. (2015).
Foaming capacity and foaming stability
Foaming capacity and foaming stability of isolates produced from acid and alkaline processing is given in Table 5. In the present study, foaming capacity of acid-processed isolates was found to be higher compared to that of alkali-processed isolates (p < 0.05). Only a few reports are available on foaming properties of pH shift processed isolates. Foaming capacity of proteins can be improved by increasing the exposure of hydrophobic residues at air-water interface and reducing the surface tension (Mutilangi et al. 1996). The additional hydrophobic residues at air-water interface could form a large hydrophobic sphere on surface resulting in excess foaming (Jongjareonrak et al. 2010). The differences in foaming capacity between acid- and alkali-processed isolates might be due to differences in hydrophobic residue content (Surasani et al. 2017).
Foam stability indicates the extent of protein-protein interaction within the matrix (Mutilangi et al. 1996; Jongjareonrak et al. 2010). In the present study, foam stability of alkali-processed isolates was found to be higher than that of acid-processed isolates (p < 0.05). Differences in foaming stability of isolates obtained by acid- and alkaline-aided processing might be due to protein aggregation, interfering the interactions between the protein and water needed for foam formation (Kinsella 1976).
SDS-PAGE pattern
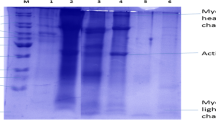
SDS-PAGE pattern of pangas proteins showed that the protein bands ranged from 10 kDa to > 200 kDa with little background smear (Fig. 1). In supernatant samples, a new protein band with approximately 190 kDa appeared, indicating the partial dissociation of heavy meromyosin. All the samples showed an intense band near to 43 kDa, which is actin. These findings were in agreement with the previous findings reported by Kristinsson and Liang (2006). The number of bands in supernatant after first centrifugation was more, indicating the proteolysis during acid- and alkaline-aided extraction. Kelleher and Hultin (2000) reported that, activation of proteolytic enzymes leads to the hydrolysis of heavy meromyosin chain resulting in the formation of low molecular weight bands. In both supernatants obtained by acid and alkaline solubilization, the partial hydrolysis of heavy myosin was observed. A band with molecular weight of 10 kDa was observed in both the supernatants, which are identified as myosin light chains. The findings were in agreement with the results reported by Azadian et al. (2012). Most of the protein bands, except three were disappeared in the supernatant after second centrifugation, indicating the maximum protein recovery after second centrifugation.

SDS-PAGE protein pattern of homogenate and supernatants from pangas fillet frame during acid (pH 2.0) and alkaline (pH 13.0) solubilization process
Amino acid profile
Aminoacid profile of pangas fillet frame homogenate and isolates obtained through acid- and alkaline-aided processing is given in Table 6. Results indicated that pH shift processing did not affect the amino acid profiling much and all the amino acids present in the homogenate were recovered efficiently by alkaline processing. The major amino acids found in homogenate were Glutamic acid (GLU), glycine (GLY), alanine (ALA) and aspartic acid (ASP), while glutamic acid (GLU), aspartic acid (ASP), leucine (LEU) and lysine (LYS) were dominant in pH shift processed isolates. The content of proline and glycine was less in acid-processed isolates followed by alkaline-processed isolates compared to homogenate, which might be due to nonextractability of connective tissues in alkaline processing (Batista 1999). Quality of protein is determined by the presence and quantity of essential amino acids. The content of total essential amino acids increased during pH shift processing, indicating the enrichment of essential amino acids. Similar findings were reported by Batista (1999). Isolates obtained using alkaline processing had higher amount of amino acids than isolates obtained by acid processing. High amino acid content of alkaline processed isolates might be due to less pH-induced proteolysis in alkaline processing (Gehring et al. 2010). The overall amino acid content in pangas protein isolates obtained by pH shift processing were well above the recommended levels for adults by WHO/FAO/UNU, 2007. Similar findings were reported by Marmon and Undeland (2010) during their studies on protein isolates obtained from herring through pH shift processing.
Microbial stability
Microbial stability of food products is an important parameter during their production and storage (Nolsøe and Undeland 2009). Live or fresh fish immediately after harvesting contain a high number of microorganisms, mainly psychrotrophic Gram-negative bacteria (Huss 1983). The question has been however raised as it was expected that highly acidic and alkaline processing have a killing effect on microbes associated with the raw material (Jay 1986). In this study, no microbial counts were detected in isolates prepared using acid and alkaline extraction methods, whereas homogenate had a total plate count of 5.35 × 103 cfu/ml. Similar findings were reported by Surasani et al. (2017) during their studies on rohu protein isolates obtained through pH shift processing. Kristinsson and Demir (2003) found a reduction in aerobic plate count numbers directly after acid, alkaline, and washing processes, but the largest reduction was achieved through alkaline processing.
Gel quality
Texture profile analysis (TPA)
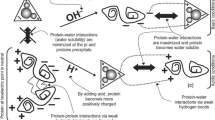
Texture is a complex sensory attribute that includes hardness, springiness, cohesiveness, gumminess, chewiness, and resilience that cumulatively contribute to the mouth feel/texture experienced by humans (Haard 1992; Szczesniak 2002). Cohesiveness indicates the strength of the internal bonds and springiness is the rate at which a material goes back to its previous condition after the removal of force (Szczesniak 2002). TPA values of gels made of acid- and alkaline-processed isolates are given in Table 7. Except for springiness and gumminess, all the values were higher for acid processed isolates than alkali processed isolates (p < 0.05). Higher TPA values of acid processed isolate gels indicate the integrity and strength of gel. Lower TPA values of alkaline-processed protein gels indicate poor gel network causing poor gel strength. Similar findings were reported by Fatin et al. (2015) during protein recovery from Japanese scad. They reported that acid-processed surimi gel had high gel strength, hardness, springiness, cohesiveness, and chewiness compared to alkaline-processed surimi gel. The more hardness of acid-processed protein gels might be due to excess protein-protein interactions resulting in hard gels, while excess protein-water interactions results in weak fragile gels in alkaline processing (Nolsøe and Undeland 2009).
Shear test
Shear strength and work of shear values for protein gels made of acid- and alkaline-processed isolates are given in Table 7. Gels made of acid-processed isolates were found to have high shear strength and work of shear values compared to alkaline-processed protein gels (p < 0.05), indicating the strong gel network. Lower shear test values of alkaline processed protein gels indicate poor gel network resulting in poor gel strength. Freitas et al. (2015) reported that acid-processed isolate gels from whitemouth croaker and anchovy had good cutting strength values. Kristinsson and Liang (2006) found greater gel strength in gels of acid-processed tilapia proteins than alkaline-processed protein gels. Hultin and Kelleher (1999) also reported that acid process showed good gels from cod and mackerel proteins. Differences in gel forming ability may be due to differences in protein integrity and the bond formation during heating process (Chaijan et al. 2006). Ability to form good protein gels depends on species, methods used, and the extraction time, which causes differences in results (Freitas et al. 2015).
Expressible moisture content
Expressible moisture of gel content indicates the integrity of protein gel to hold water. Gels made of acid- and alkaline-processed isolates had an expressible moisture content of 17.50 ± 0.64% and 26.42 ± 0.86%, respectively. Gels made from alkaline-processed isolate had significantly higher expressible moisture, i.e., low water holding capacity, compared to gels made of acid-processed isolates indicating the poor gel network (p < 0.05). Similar findings were reported by Freitas et al. (2011) during protein recovery from Argentine anchovy residue. They found higher water holding capacity for acid-processed protein gels compared to alkaline-processed protein gels. Fatin et al. (2015) reported that gel produced by acid-aided processing of Japanese scad proteins had low expressible moisture than alkali-aided processed protein gels indicating the high water holding capacity of acid processed protein gels.
Folding test
Gels made of acid- and alkaline-processed isolates had an average scores of 5 and 1 during the folding test. Gels from acid-processed isolates showed excellent gelling properties by scoring 5 during folding test (p < 0.05). Alkaline-processed isolate gels scored 1, indicating poor gel network. Marmon and Undeland (2010) during their study on herring protein isolates found that both acid and alkaline processed isolates produced gels that are strong and scored 5 during folding test. In the present study, alkaline-processed isolate gels showed poor gelling ability which might be due to more lipid, pigment, and myoglobin content than acid-processed isolates or might be due to denaturation during the alkaline extraction process.
Conclusions
Both acidic (pH 2.0) and alkaline (pH 13.0) solubilization processes could recover proteins from pangas fillet frames with good recovery yield. Acid solubilization process found to have minimal effect on protein functionality during processing resulting in the recovery of more stable and functional proteins. Further studies are needed to find out the reasons behind denaturation of proteins during alkaline solubilization and associated loss of functionality. pH shift processing found to be promising in the utilization of fish processing waste for the recovery of functional proteins thus by reducing the supply demand gap as well pollution problems.
References
AOAC (2000) Association of official analytical chemists, 16th edn. Washington, DC
Azadian M, Moosavi-Nasab M, Abedi E (2012) Comparison of functional properties and SDS-PAGE patterns between fish protein isolate and surimi produced from silver carp. Eur Food Res Technol 235(1):83–90. https://doi.org/10.1007/s00217-012-1721-z
Batista I (1999) Recovery of proteins from fish waste products by alkaline extraction. Eur Food Res Technol 210(2):84–89. https://doi.org/10.1007/s002170050539
Batista I, Pires C, Nelhas R (2007) Extraction of sardine proteins by acidic and alkaline solubilisation. Food Sci Technol Int 13(3):189–194. https://doi.org/10.1177/1082013207079619
Bidlingmeyer BA, Cohen SA, Tarvin TL (1984) Rapid analysis of amino acids using precolumn derivatisation. J Chromatogr 336(1):93–104. https://doi.org/10.1016/S0378-4347(00)85133-6
Chaijan M, Undeland I (2015) Development of a new method for determination of total haem protein in fish muscle. Food Chem 173:1133–1141. https://doi.org/10.1016/j.foodchem.2014.11.010
Chaijan M, Benjakul S, Visessanguan W, Faustman C (2006) Physicochemical properties, gel forming ability and myoglobin content of sardine (Sardinnella gibbosa) and mackerel (Rastrelliger kanagurta) surimi produced by conventional method and alkaline solubilization process. Eur Food Res Technol 222(1-2):58–63. https://doi.org/10.1007/s00217-005-0091-1
Chen YC, Tou JC, Jaczynski J (2007) Protein recovery from rainbow trout (Oncorhynchus mykiss) processing byproducts via isoelectric solubilization/precipitation and its gelation properties as affected by functional additives. J Agric Food Chem 55(22):9079–9088. https://doi.org/10.1021/jf071992w
Cortes-Ruis J, Pachero-Aguilar R, Garcia-Sanchez G, Lugo-Sanches ME (2001) Functional characterization of a protein concentrate from bristly sardine made under acidic conditions. J Aquat Food Prod Technol 10(4):5–23. https://doi.org/10.1300/J030v10n04_02
Fatin NS, Huda N, David W (2015) Physicochemical properties of Japanese scad (Decapterus Maruadsi) surimi prepared using the acid and alkaline solubilization methods. Int J Sci Eng Res 6(4):141–147
Feng YM, Hultin HO (2001) Effect of pH on the rheological and structural properties of gels of water-washed chicken-breast muscle at physiological ionic strength. J Agric Food Chem 49(8):3927–3935. https://doi.org/10.1021/jf001021f
Freitas IR, Gauterio GV, Rios DG, Prentice C (2011) Functionality of protein isolates from argentine anchovy (Engraulis anchoita) residue using pH shift processing. J Food Sci Eng 1:374–378
Freitas IR, Cortez-Wega WR, Prentice C (2015) Evaluation of properties of protein recovered from fish muscle by acid solubilization process. Int Food Res J 22(3):1067–1073
Gehring CK, Gigliotti JC, Moritz JS, Tou JC, Jaczynski J (2010) Functional and nutritional characteristics of proteins and lipids recovered by isoelectric processing of fish by-products and low-value fish: a review. Food Chem 124:422–431
Haard NF (1992) Control of chemical composition and food quality attributes of cultured fish. Food Res Int 25(4):289–307. https://doi.org/10.1016/0963-9969(92)90126-P
Hamm R (1994) The influence of pH on the protein net charge in the myofibrillar system. Reciprocal Meat Conf Proc 47:5–9
Harlow E, Lane D (1988) Antibodies: a laboratory manual. Cold Spring Harbor Laboratory Press, New York
Huang L, Chen Y, Morrissey MT (1997) Coagulation of fish proteins from frozen fish mince wash water by ohmic heating. J Food Process Eng 20(4):285–300. https://doi.org/10.1111/j.1745-4530.1997.tb00423.x
Hultin HO, Kelleher SD (1999) Process for isolating a protein composition from a muscle source and protein composition. US Patent 6,005,073
Hultin HO, Kelleher SD, Feng YM, Kristinsson HG, Richards MP, Undenand IA (2000) High efficiency alkaline protein extraction. US Patent 6,136,959
Huss HH (1983) Fresk Fisk. K valitet Og Holdbarhed. [Fresh fish. Quality and quality changes]. Ministry of Fisheries, Technological laboratory, Lyngby
Jafarpour SA, Shabanpour B, Filabadi SS (2013) Biochemical properties of fish protein isolate (FPI) from silver carp (Hypophthalmychthis molitrix) by application of acid-alkali process compared to traditional prepared surimi. Ecopersia 1(3):315–327
Jay JM (1986) Modern food microbiology. Van Nostrand Reinhold Company, New York
Jongjareonrak A, Rawdkuen S, Chaijan M, Benjakul S, Osako K, Tanaka M (2010) Chemical compositions and characterization of skin gelatin from farmed giant catfish (Pangasianodon gigas). LWT Food Sci Technol 43(1):161–165. https://doi.org/10.1016/j.lwt.2009.06.012
Kelleher SD, Hultin HO (2000) Functional chicken muscle protein isolates prepared using low ionic strength, acid solubilisation/precipitation. Reciprocal Meat Conf Proc 3:76–81
Kinsella JE (1976) Functional properties of proteins in foods, a survey. CRC Crit Rev Food Sci Nutr 7(3):219–280. https://doi.org/10.1080/10408397609527208
Kristinsson H, Demir N (2003) Functional fish protein ingredients from fish species of warm and temperate waters: comparison of acid- and alkali-aided processing vs. conventional surimi processing. In: Betchel PJ (ed) Advances in seafood byproducts 2002 Conference Proceedings, Alaska Sea Grant College Program University of Alaska, pp. 277–295
Kristinsson H, Ingadottir B (2006) Recovery and properties of muscle proteins extracted from tilapia (Oreochromis niloticus) light muscle by pH shift processing. J Food Sci 1(3):E132–E141
Kristinsson HG, Liang Y (2006) Effect of pH-shift processing and surimi processing on Atlantic croaker (Micropogonias undulates) muscle proteins. J Food Sci 71(5):C304–C312. https://doi.org/10.1111/j.1750-3841.2006.00046.x
Kristinsson H, Theodore AE, Demir N, Ingadottir B (2005) A comparative study between acid- and alkali-aided processing and surimi processing for the recovery of proteins from channel catfish muscle. J Food Sci 70(4):C298–C306
Kudo G, Okada M, Miyauchi D (1973) Gel-forming capacity of washed and unwashed flesh of some Pacific coast species of fish. Mar Fish Rev 32:10–15
Laemmli UK (1970) Cleavage of structural proteins during assembly of the head of bacteriophage T4. Nature 227(5259):680–685. https://doi.org/10.1038/227680a0
Lubana GK, Kaur B, Surasani VKR (2016) Quality changes in fresh rohu (Labeo rohita) cutlets added with fibers from ragi, oat and jowar. Nutr Food Sci 46(4):571–582. https://doi.org/10.1108/NFS-02-2016-0023
Marmon SK, Undeland I (2010) Protein isolation from gutted herring (Clupea harengus) using pH-shift processes. J Agric Food Chem 58(19):10480–10486. https://doi.org/10.1021/jf101057q
Mutilangi WAM, Panyam D, Kilara A (1996) Functional properties of properties of hydrolysates from proteolysis of heat-denatured whey protein isolate. J Food Sci 61(2):270–274. https://doi.org/10.1111/j.1365-2621.1996.tb14174.x
Niki H, Kato T, Deya E, Igarashi S (1985) Recovery of protein from effluent of fish meat in producing surimi and utilization of recovered protein. Nippon Suisan Gakkaishi 51(6):959–964. https://doi.org/10.2331/suisan.51.959
Nolsøe H, Undeland I (2009) The acid and alkaline solubilization process for the isolation of muscle proteins: state of art. Food Bioprocess Technol 2(1):1–27. https://doi.org/10.1007/s11947-008-0088-4
Nolsøe H, Marmon SK, Undeland I (2011) Application of filtration to recover solubilized proteins during ph-shift processing of blue whiting (Micromesistius poutassou); effects on protein yield and qualities of protein isolates. Open Food Sci J 5(1):1–9. https://doi.org/10.2174/1874256401105010001
Panpipat W, Chaijan M (2016) Biochemical and physicochemical characteristics of protein isolates from bigeye snapper (Priacanthus Tayenus) head by-product using pH shift method. Turkish. J Fish Aquat Sci 16:41–50
Rawdkuen S, Sai-Ut S, Khamsorn S, Chaijan M, Benjakul S (2009) Biochemical and gelling properties of tilapia surimi and protein recovered using an acid-alkaline process. Food Chem 112(1):112–119. https://doi.org/10.1016/j.foodchem.2008.05.047
Reddy SVK (2016) Effect of formulation and processing methods on the quality and acceptability of cutlets made from minced meat of pangas (Pangasius pangasius). SAARC J Agric 14(1):25–36. https://doi.org/10.3329/sja.v14i1.29573
Robinson HW, Hogden CG (1940) The biuret reaction in the determination of serum proteins. J Biol Chem 135:707–725
Sathe SK, Deshpande SS, Salunkhe DK (1982) Functional properties of lupin seed (Supinus mutabilis) proteins and protein concentrates. J Food Sci 7:191–197
Shabanpour B, Etemadian Y, Taghipour B (2015) Physicochemical and rheological parameters changes for determining the quality of surimi and kamaboko produced by conventional, acid and alkaline solubilization process methods from common kilka (Clupeonella cultriventris caspia). Iran J Fish Sci 14(4):826–845
Surasani VKR (2017) Influence of rohu (Labeo rohita) deboning by-product on composition, physical properties and sensorial acceptability of rohu cutlets. Nutr Food Sci 47(3):398–408. https://doi.org/10.1108/NFS-08-2016-0128
Surasani VKR, Tyagi A, Kudre T (2017) Recovery of proteins from rohu processing waste using ph shift method: characterization of isolates. J Aquat Food Prod Technol 26(3):356–365. https://doi.org/10.1080/10498850.2016.1186130
Szczesniak AS (2002) Texture is a sensory property. Food Qual Prefer 13(4):215–225. https://doi.org/10.1016/S0950-3293(01)00039-8
Tian Y, Wang W, Yuan C, Zhang L, Liu J, Liu J (2016) Nutritional and digestive properties of protein isolates extracted from the muscle of the common carp using pH shift processing. J Food Process Preserv 41(1):e12847. https://doi.org/10.1111/jfpp.12847
Undeland I, Kelleher SD, Hultin HO (2002) Recovery of functional proteins from herring (Clupea harengus) light muscle by an acid or alkaline solubilization process. J Agric Food Chem 50(25):7371–7379. https://doi.org/10.1021/jf020199u
WHO/FAO/UNU (2007) Protein and amino requirements in human nutrition. Report of a Joint WHO/FAO/UNU Expert Consultation, WHO Technical Report Series 935, World Health Organization, Geneva
Acknowledgements
Authors wish to express their sincere thanks to the Dean, College of Fisheries, GADVASU and University Authorities, GADVASU for their constant support during the work period. Authors also wish to express their thanks to Ms. Manvinder Kaur for her technical help during the preparation of manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Philippe Garrigues
Highlights
• Study was conducted to utilize pangas (Pangasius pangasius) processing waste (fillet frames) to recover proteins using pH shift method and to characterize the recovered isolates
• pH 2.0 from acidic range and pH 13.0 from alkaline range were found to have maximum protein solubility (p < 0.05).
• Acidic pH (pH 2.0) was found to have minimal effect on proteins, while alkaline pH (pH 13.0) caused protein denaturation resulting in less stable proteins and poor gel network.
• Both acidic and alkaline-aided processing caused significant (p < 0.05) reductions in total lipid, myoglobin, and pigment content thus by resulting in whiter proteins isolates and gels.
• The content of total essential amino acids increased during pH shift processing, indicating the enrichment of essential amino acids.
• No microbial counts were detected in isolates prepared using acid and alkaline extraction methods.
• pH shift processing was found to be promising in the utilization of fish processing waste for the recovery of functional proteins thus by reducing the supply demand gap as well pollution problems
Rights and permissions
About this article

Cite this article
Surasani, V.K.R., Kudre, T. & Ballari, R.V. Recovery and characterization of proteins from pangas (Pangasius pangasius) processing waste obtained through pH shift processing. Environ Sci Pollut Res 25, 11987–11998 (2018). https://doi.org/10.1007/s11356-018-1456-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-018-1456-x