
Abstract
Purpose
The present study compared isometric, concentric and eccentric contractions at the same torque-time integral for changes in neuromuscular fatigue and muscle damage parameters.
Method
Healthy men (18–24 years) were placed to either isometric (ISO), concentric (CONC), or eccentric (ECC) group (n = 11/group) that performed corresponding contractions of the knee extensors to exert the same amount of torque-time integral (24,427 ± 291 Nm·s). Changes in maximal voluntary contraction (MVC) torque, voluntary activation, evoked torque at 10 Hz and 100 Hz and its ratio, M-wave amplitude, and muscle soreness were assessed immediately before and after, 1 h, 1 day and 2 days after each exercise, and were compared among the groups.
Results
MVC torque decreased immediately after ISO (− 17.0 ± 8.3%), CONC (− 21.7 ± 11.5%) and ECC (− 26.2 ± 15.6%) similarly (p = 0.35), but the decrease sustained longer (p < 0.05) for ECC (2 days post-exercise: − 12.9 ± 14.8%) and ISO (− 5.5 ± 7.9%) than CONC (+ 5.0 ± 11.0%). Muscle soreness developed after ECC (25.1 ± 19.8 mm) and ISO (17.5 ± 21.0 mm) similarly (p = 0.15). Voluntary activation decreased immediately (− 3.7 ± 6.6%) and 1 h post-exercise (− 4.7 ± 7.6%) for all groups similarly. Electrically evoked forces decreased greater immediately (− 30.1 ± 15.6%) and 1 h post-exercise (− 35.0 ± 12.8%) for ECC than others, and the decrease in 10/100 Hz ratio was also greater immediately (− 30.5 ± 12.6%) and 1 h after ECC (− 23.8 ± 10.3%) than others.
Conclusion
ISO, CONC and ECC with the same torque-time integral produced similar neuromuscular fatigue at immediately post-exercise, but the force loss was longer-lasting after ISO and ECC than CONC, and the changes in peripheral fatigue parameters were the greatest after ECC, suggesting greater muscle damage.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
A decrease in performance with an increase in perception of effort is observed during and/or after exercise (Ansdell and Dekerle 2020). This is generally described as “fatigue” that is defined as a reduction of the capacity to produce a power or strength (Bigland-Ritchie and Woods 1984) and is generally determined by a decrease in maximal voluntary contraction (MVC) strength (Place et al. 2006). Neuromuscular fatigue is caused by a combination of central (cortical and/or spinal-motoneuronal circuits) and peripheral (beyond the neuromuscular junction) mechanisms. When a loss of MVC strength persists for several days following a bout of exercise, it indicates the presence of muscle damage (Paulsen et al. 2012).
Neuromuscular fatigue and muscle damage are largely task dependent (Enoka and Stuart 1992), particularly affected by muscle contraction types (Ochi et al. 2016). It is well documented that eccentric (lengthening) contractions produce a greater decrease of MVC force than isometric or concentric (shortening) contractions, and the loss of force generation capability persists longer following eccentric contractions due to a greater extent of muscle damage (Byrne et al. 2004). The greater force production during eccentric contractions leads to greater increases in muscle strength and muscle hypertrophy in resistance training (Roig et al. 2009). Pasquet et al. (2000) compared the neuromuscular changes after 150 maximal voluntary concentric (CONC) and eccentric contractions (ECC) of the elbow flexors at the same angular velocity and range of motion. The authors reported that CONC resulted in greater decreases in elbow flexor torque and biceps brachii electromyogram (EMG) activity at the end of the exercise when compared with ECC that did not change voluntary activation, suggesting that the central fatigue was greater after CONC than ECC. Kay et al. (2000) compared maximal ECC, CONC and isometric contractions (ISO) with the same contraction time (100 s), and showed that only CONC and ISO induced a decrease in MVC at the end of the exercise. However, in these studies (Kay et al. 2000; Pasquet et al. 2000), no measurements were taken beyond immediately post-exercise, and importantly the fatigue at the end of each exercise was not equal among different muscle contraction types.
To avoid this limitation, Babault et al. (2006) induced a similar MVC torque reduction (~ − 60%) immediately after repeated muscle contractions of the knee extensors to compare ISO and CONC for changes in central and peripheral parameters from pre- to post-exercise. They reported that voluntary activation and EMG activity were lower in ISO than CONC, indicating a greater central fatigue for ISO than CONC, but contractile properties represented by doublet twitch amplitude and relaxation were more impaired after CONC than ISO. However, they did not include ECC in the comparison. This approach could provide a better picture for the neuromuscular fatigue and muscle damage profiles among CONC, ISO and ECC.
It is well documented that ECC induces greater muscle damage than CONC or ISO (Kroon and Naeije 1991). Ochi et al. (2016) reported that 5 sets of 6 maximal contractions of the elbow flexors produced greater muscle damage after ECC than CONC, demonstrated by more prolonged decreases in elbow flexors torque, greater delayed onset muscle soreness (DOMS) and increase in creatine kinase (CK) activity in the blood. It has also been reported that ISO at a long muscle length induces greater MVC torque decreases and DOMS than that at a short muscle length (Allen et al. 2018), indicating that ISO contractions at a long muscle length could also induce muscle damage. The same number of maximal muscle contractions produces an unequal level of fatigue given that ECC produces greater torque than ISO or CONC. Souron et al. (2018) matched the magnitude of decrease in MVC torque immediately post-exercise (− 40%) between CONC and ECC, and found that MVC torque decreased greater for 5 days after ECC than CONC, and DOMS was greater for ECC than CONC. However, the work produced during ECC (16,000 ± 6500 kJ) was much greater than that of CONC (8100 ± 2700 kJ) in the study.
To the best of our knowledge, no previous studies have compared between ISO, CONC, and ECC with the same torque-time integral for changes in neuromuscular parameters after exercise. The aim of this study was therefore to compare ISO, CONC and ECC of the knee extensors with the same amount of torque-time integral for changes in neuromuscular indices (central and peripheral factors) before and up to 2 days post-exercise. It was hypothesized that the loss of MVC torque immediately after CONC exercise would be greater than others, but muscle damage would be greater after ECC than CONC or ISO.
Methods
Participants
Male sport science students (n = 33, average age: 21.1 ± 1.9 y; body mass: 73.4 ± 8.9 kg; height: 178.7 ± 6.6 cm) who did not have an injury or musculoskeletal disorder of the right leg participated in this study. They were not engaged in resistance training in the last 6 months prior to the study. All participants were informed about the risks and discomfort associated with the experimental procedure before giving their written consent to participate. This study was conducted in conformity with the declaration of Helsinki and approved by the local ethics committee of the University of Nantes. To quantify their level of physical activity, participants answered the International Physical Activity Questionnaire (IPAQ). The sample-size was calculated using G*Power (version 3.1.9.2; Kiel University, Kiel, Germany), based on an expected “medium” effect size (f2 = 0.25) for a difference in MVC torque changes at 1 day post-exercise among ISO, CONC and ECC, with α level of 0.05, power (1 − β) of 0.8 and correlation among repeated measures of 0.85. No significant differences among the groups were observed for age, body mass, height, MVC torque and IPAQ (Table 1).
Experimental design
All participants attended a familiarization session in which anthropometric measures were taken and isometric MVC torque was measured at two different angles (90 ° and 110 ° knee flexion). Based on these measurements, the participants were placed in one of the three groups; isometric (ISO), concentric (CONC) or eccentric (ECC) group by considering anthropometric characteristics and the MVC torque to be similar among the groups as much as possible. Each participant performed only one session of either isometric, concentric, or eccentric contractions of the knee extensors. This study design was chosen to limit the bias of the influence of the repeated bout effect, which could attenuate the MVC torque loss after exercise (McHugh 2003). Neuromuscular measures including MVC torque, voluntary activation and evoked torque, M-wave and ratio 10 Hz/100 Hz were performed before (PRE), immediately after (POST), 1 h (POST1), 24 h (POST24) and 48 h (POST48) after each exercise (ISO, CONC, ECC) performed at the same amount of torque-time integral. Muscle soreness was also assessed at the same time points except POST.
Exercise protocol
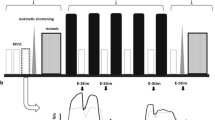
In ISO, CONC, and ECC sessions, each participant performed several sets of 10 contractions. To match the total work among the groups, the number of sets varied among the exercises, based on the amount of work performed by the first participant of a group of three participants. Souron et al. (2018) showed that the work was approximately twice in ECC than CONC for the knee extensors, and maximal torque produced by ECC was 30–50% greater than ISO (Duchateau and Enoka 2016). Thus, when the CONC condition was performed first in a group of three participants, 15 sets were performed, however, when ECC and ISO were performed first in a group, 8 and 10 sets were performed, respectively. The torque-time integral during the first exercise (e.g., 15 sets of 10 concentric contractions) was analyzed in real-time with a MatLab script for a participant, and the number of sets was modulated for a participant in ISO, and another participant in ECC group for the torque-time integral to be similar for the three participants in the group. Thus, the torque-time integral was matched for a group of three participants who performed either ISO, CONC or ECC, and this was repeated for ten more groups. For example, when a first participant performed 15 sets for the CONC (the total torque-time integral: 27,084 Nm·s), the second participant performed 9 sets for the ECC (26,150 Nm·s) for the torque-time integral to be close to that of the first participant, and the third participant performed 10 sets for the ISO (26,286 Nm·s) for the torque-time integral to be close to those of the first two participants.
CONC and ECC contractions were performed at 60°/s over a range of motion of 100 ° from 120 ° to 20 ° of knee joint angle (0 ° = full extension) and from 20 ° to 120 ° of knee joint angle, respectively, with a 90-s rest period between sets. This angular velocity was reported to be highly reproducible (Pincivero et al. 1997). ISO contractions were performed at 90 ° of knee joint angle with the same contraction time (i.e., 1.6 s) and rest period (90-s) as those of CONC and ECC using audio feedback.
Neuromuscular tests
Each participant sat on an isokinetic dynamometer (CMV-AG, Dübendorf, Switzerland) with 85 ° hip flexion (full hip extension = 0 °) and 90 ° knee flexion (full knee extension = 0 °). The right knee axis was aligned with the dynamometer rotation axis. The participant was firmly fastened with belts to avoid the trunk movements and was asked to keep his arms crossed during all contractions. Torque and angular position were digitized by digital converter module (Delsys® Inc., Boston, MA, USA), sampled in real-time at 2000 Hz in a data acquisition software (NeuroEval©, University of Nantes, Nantes, France). All contractions (neuromuscular tests and exercise protocol) were performed with this position.
Surface electromyography
After skin preparation, a pair of circular electrodes (diameter = 1 cm, ADInstruments Pty Ltd) were placed over vastus lateralis (VL) and vastus medialis (VM) according to the SENIAM recommendation, and the ground electrode (diameter = 1 cm, ADInstruments Pty Ltd) was placed on the patella. EMG signal was amplified via a biosignal amplifier (g.BSamp 0201a, Guger Technologies, Schiedlberg, Austria; bandwidth = 2–1000 Hz, gain = 1000), sampled at 5000 Hz in a data acquisition software (NeuroEval©, University of Nantes, Nantes, France). The location of electrodes was marked on the skin by using black marker for the following sessions.
Electrical stimulation
The femoral nerve was stimulated using a circular cathode (diameter = 1 cm, ADInstruments Pty Ltd) placed on the femoral triangle, and the anode (5 × 9 cm; Chattanooga Medical Supply Inc. USA) placed under the gluteal fold. Electrical impulses of single and paired (10 Hz and 100 Hz) were software-triggered using a constant-current stimulator (Digitimer DS7A, Digitimer Ltd, Hertforshire, UK). Rectangular pulses with 1-ms duration and 400 V maximal output voltage were applied. Constant pressure was applied on the cathode during stimulations (Cattagni et al. 2018). Ramp of electrical nerve stimulation of 10 mA increment were evoked until the optimal intensity of stimulation inducing maximal M-wave and twitch torque amplitudes was found. This optimal intensity of stimulation was then increased by 20% to obtain supramaximal intensity.
Maximal isometric voluntary contraction
After a standardized warm-up protocol consisting of submaximal ISO and CONC contractions at 30–80% of MVC, each participant performed 3 isometric MVC at 90 ° for 4 s interspaced by a 90-s rest period. This angle (90 °) was chosen because it represents the angle where the torque produced during isokinetic (CONC or ECC) contractions was close to the maximal. Strong verbal encouragements were given to each participant, and the participants were asked to reach their maximal force as fast as possible. The first MVC was performed without electrical stimulations whereas electrical stimulations were applied in the following two MVC measures. MVC with the best peak torque was used for further analyses.
During the second and third MVC, stimulations were evoked during and after the MVC. When the MVC torque plateaued, paired stimulations (100 Hz) were superimposed to quantify voluntary activation (see below). At the end of the MVC (4 s after), potentiated paired stimulation (100 Hz) was evoked following by 2 s after a second potentiated paired stimulation (10 Hz) to assess low-frequency fatigue. Then, single stimulation was evoked (2 s after) to obtain M-wave.
During exercise protocol, paired stimulations (100 Hz) were also evoked at the fifth contraction of each set. For CONC and ECC, stimulations were delivered at 75 ° degrees. At the end of each set, a single stimulation was evoked to assess electrical and contractile properties. To evaluate the recovery rate of the MVC torque, the changes in the MVC torque from immediately to 1 h post-exercise as well as immediately to 1 day post-exercise were calculated.
Voluntary activation
Voluntary activation level was calculated with the following formula for neuromuscular measurements (Allen et al. 1995):
where VA is the voluntary activation level; A represents the amplitude of the superimposed doublet torque, B represents the amplitude of the potentiated doublet torque. If the electrical stimulation was not delivered at peak torque, Strojnik and Komi (1998) correction was used to calculate voluntary activation with the following formula:
where Tstim represents torque at stimulation time. MVC represents maximal voluntary peak torque.
During the CONC and ECC protocols, the superimposed torque was calculated by subtracting evoked torque and torque if there was not a stimulation at the angle of evoked torque. This extrapolated torque could be estimated via torque data linear extrapolation intervening 50-ms before the stimulation (Babault et al. 2001). Voluntary activation during exercise (ECC, CONC, ISO) was calculated using the central Activation Ratio (CAR):
where Textrap represents an extrapolated torque and Tsurimp represents a superimposed torque.
Electrical and contractile properties
Single stimulation at rest was delivered to obtain M-wave properties (first phase amplitude and duration). Potentiated doublet stimulation was used to investigate contractile properties (peak twitch torque and time to peak) of knee extensors. In addition, 10/100 Hz ratio was assessed to evaluate low-frequency fatigue (Edwards et al. 1977). All the evoked responses were chosen from the stimulated MVCs with the best peak value for different testing sessions, except for POST and POSTH1 where the first stimulated MVC was selected to minimize recovery effects.
Muscle soreness
Muscle soreness was assessed with a 100-mm visual analog scale (VAS), where 0 corresponds to “no pain” and 100 to “extremely painful”. The participants reported their subjective soreness on the VAS after being palpated. Palpation was always made by the same investigator with identical pressure of 2-s being applied successively on rectus femoris (RF), vastus medialis and vastus lateralis. All of these measures were carried out at pre- and post-exercise (POSTH1, POSTD1, POSTD2).
Statistical analyses
Analyses were performed with Statistica® 8.0 (Statsoft Inc, Tulsa, OK, USA). The normality of the data was tested by Kolmogorov–Smirnov. The participants’ characteristics were compared among the groups using one-way measure ANOVA. Since the number of sets that were performed in the exercise were different among participants and conditions (7–15 sets), the number of sets were divided into 4 sections (every 25% of the total number of sets). To compare changes in mechanical and EMG parameters during exercise among the three contraction types, a two-way ANOVA was used for the condition (ISO, CONC, ECC) × section (baseline: 0, 25%, 50%, 75%, 100%) interaction effect. Changes in the neuromuscular fatigue and indirect markers of muscle damage parameters (MVC torque, VA, Potentiated doublet properties, 10/100 Hz ratio, M-wave properties, muscle soreness) before (PRE), immediately after (POST), 1 h (POSTH1) and 1 (POSTD1) and 2 days after exercise (POSTD2) were compared among the three contraction types (ISO, CONC, ECC) using two-way repeated measures ANOVA. One-way ANOVA was also used to compare the recovery rate of MVC torque from immediately to 1 h as well as 1 day post-exercise among ISO, CONC and ECC. When a significant time or interaction effect was found, post-hoc analysis was made by Fisher LSD test. Partial eta square was calculated to assess effect size, with 0.07 and 0.14 for medium and large effect size, respectively (Cohen 1988). Cohen’s dz effect sizes are also reported with dz = 0.2, dz = 0.5 et dz = 0.8 for small, moderate and large effect size, respectively (Cohen 1988). For all tests, significance level was set at p < 0.05. Data are presented as mean values ± standard deviation (SD).
Results
Exercise
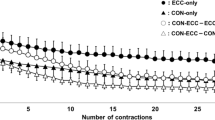
No significant difference in the torque-time integral (p = 0.97, pƞ2 = 0.4) was evident among ISO (24,474 ± 6553 Nm.s), CONC (24,692 ± 6568 Nm.s) and ECC (24,117 ± 6338 Nm.s) groups as planned, but the total number of contractions was greater for CONC (135.5 ± 22.5) than ECC (96.4 ± 15.0) (p < 0.01, dz = 2.08) or ISO (100.0 ± 15.5) group (p < 0.01, dz = 1.86). The two-way ANOVA showed a significant time effect (F4,120 = 2.96, p < 0.05, pƞ2 = 0.09) for changes in torque over time (Fig. 1A), but no group (contraction type) × time interaction effect (F8,120 = 1.98, p = 0.54, pƞ2 = 0.12). Compared to the first set, torque was significantly decreased at the end of the exercise for all types of contractions (p < 0.05, dz = 0.79). The rate of decrease from baseline to the end of exercise was significantly different between contraction types (F2,30 = 3.48, p < 0.05, pƞ2 = 0.20). Post-hoc analysis revealed a significantly greater rate of decrease in voluntary torque for CONC than ECC (p < 0.05, dz = 0.97) without a significant difference between CONC and ISO (p = 0.06, dz = 0.36). A contraction effect (F2,30 = 3.85, p < 0.05, pƞ2 = 0.20) showed that the torque produced during ECC was greater than that of CONC (p < 0.05, dz = 0.74), but was not different from that during ISO (p = 0.07, dz = 0.68). Peak twitch torque was significantly reduced after 25% of the total number of contractions (− 5.9 ± 14.3%; p < 0.05, dz = 0.81) as well as for other sections (50%, 75%, 100%) throughout the exercise (− 14.7 ± 16.3%, − 21.2 ± 17.3%, and − 25.0 ± 18.0%, respectively, p < 0.001, dz ranged from 1.80 to 2.78) (Fig. 1B). No significant differences among ISO, CONC and ECC were observed for voluntary activation during exercise (Fig. 1C).

Changes (mean ± SEM) in torque (A), peak twitch (B), voluntary activation (C) assessed during isometric (ISO), concentric (CONC), and eccentric (ECC) exercise at 25, 50, 75, and 100% of the total contractions performed in each exercise. *Significantly different from 0% (p < 0.05). ϕSignificantly different from 25% (p < 0.05). αSignificantly different from 50% (p < 0.05). #Significantly different from ISO (p < 0.05)
Changes in neuromuscular fatigue and indirect markers of muscle damage parameters
MVC torque
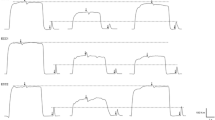
As shown in Fig. 2 and 3A, a significant contraction type × time interaction (F8,120 = 3.44, p < 0.05, pƞ2 = 0.18) was found. MVC torque decreased at immediately after ISO (− 17.0 ± 8.3%, dz = 4.08, p < 0.001), CONC (− 21.7 ± 11.5%, dz = 3.77, p < 0.001) and ECC (− 26.2 ± 15.6%, dz = 3.37, p < 0.001), without a significant difference among the groups. However, MVC torque was significantly lower at POSTH1 for ECC (− 23.9% ± 10.2%) than both ISO (− 14.2 ± 6.7%) and CONC (− 9.9 ± 9.4%) (dz = 0.90, p < 0.05 vs ISO; dz = 1.16, p < 0.05 vs CONC). MVC torque was lower for ECC and ISO than CONC at POSTD1 (dz = 1.41, p < 0.001, dz = 1.61, p < 0.05) and POSTD2 (dz = 1.12, p < 0.001, dz = 1.11, p < 0.05). However, no significant difference was found between ECC and ISO condition at POSTD1 (p = 0.09) and POSTD2 (p = 0.12) (Fig. 3A). No significant difference among ISO, CONC and ECC was observed for the recovery rate of MVC torque from immediately to 1 h post-exercise (F2,30 = 2.26, p = 0.12, pƞ2 = 0.13). However, the recovery rate from immediately to 1 day post-exercise was significantly different between ISO, CONC and ECC (F2,30 = 4.85, p = 0.01, pƞ2 = 0.24). Post hoc analysis revealed a significant faster recovery rate for CONC compared to ISO (p < 0.01, dz = 1.2) or ECC (p < 0.05, dz = 1.00).

Typical traces of a representative participant for the changes of the maximal voluntary contraction (MVC) torque and evoked torque before (PRE), immediately (POST) after and 1 day (D1) after isometric (ISO), concentric (CONC), and eccentric (ECC) exercise

Changes (mean ± SEM) in maximal voluntary contraction (MVC) torque (A), potentiated peak doublet (B), 10/100 Hz ratio (C), muscle soreness assessed by visual analogic scale (VAS) (D) before (PRE), immediately after (POST), 1 h (POSTH1), 1 (POSTD1) and 2 days (POSTD2) after isometric (ISO), concentric (CONC), and eccentric (ECC) exercise. *Significantly different from PRE (p < 0.05). #Significantly different from CONC (p < 0.05). †Significantly different from ISO (p < 0.05)
Voluntary activation
No significant type x time interaction effect was found for voluntary activation, but a significant time effect (F4,120 = 11.72, p < 0.001, pƞ2 = 0.28) was observed. When compared to PRE, voluntary activation was significantly lower at POST (− 3.7 ± 6.6%, p < 0.001, dz = 0.61) and POSTH1 (− 4.7 ± 7.6% p < 0.001, dz = 0.70) for all groups, without significant differences among groups. No significant difference was evident for POSTD1 and POSTD2 from PRE.
Potentiated doublet properties
A significant contraction type × time interaction effect (F8,120 = 3.62, p < 0.001, pƞ2 = 0.19) was found for changes in the doublet peak torque amplitude (Fig. 3B). Doublet peak torque amplitude was significantly lower for ECC than ISO at POST (p < 0.05, dz = 0.97) and POSTH1 (p < 0.05, dz = 1.52). Furthermore, ECC resulted in a significantly greater decrease in doublet peak torque amplitude than CONC at POSTH1 (p < 0.001, dz = 1.71), POSTD1 (p < 0.001, dz = 1.35) and POSTD2 (p < 0.001, dz = 0.74). Doublet peak torque amplitude was significantly decreased for ISO and CONC at POST (− 18.4% ± 8.3%, p < 0.001, dz = 4.44 for ISO and − 22.1% ± 16.4%, p < 0.001, dz = 2.68 for CONC) and POSTH1 (− 35.0% ± 12.8%, p < 0.001, dz = 4.03 ECC; − 18.2% ± 3.5%, p < 0.001, dz = 7.51 ISO) from PRE, but the decrease was significantly greater for ISO than CONC (dz = 1.75, p < 0.05) at POSTD1 (Fig. 3B).
For the time to peak torque, a contraction type × time interaction (F8,120 = 5.40, p < 0.05, pƞ2 = 0.26) was observed. Post-hoc analysis revealed a significant decrease for all contraction types at POST (ISO: − 11.0 ± 3.4%, CONC: − 8.3 ± 6.3%, ECC: − 17.2 ± 3.1%, p < 0.001, dz ranged from 2.64 to 11.23) and POSTH1 (ISO: -5.3 ± 4.0%, CONC: − 3.4 ± 7.1%, ECC: − 12.4 ± 3.6%, p < 0.05, dz ranged from 0.95 to 6.83) with greater decrease for ECC at POST (all p < 0.001, dz ranged from 1.89 to 1.90) and POSTH1 (all p < 0.001, dz ranged from 1.69 to 1.86). At POSTD1, ECC was significantly lower than CONC (p < 0.05, dz = 1.14) but not different from ISO (p = 0.21).
10/100 Hz ratio
A significant contraction type × time interaction (F8,120 = 2.33, p < 0.05, pƞ2 = 0.13) was evident for changes in 10/100 Hz ratio (Fig. 3C). 10/100 Hz ratio significantly decreased at POST and POSTH1 for ISO (− 21.4 ± 9.3%, − 16.5 ± 9.7%, p < 0.001, dz ranged from 3.39 to 4.58), CONC (− 28.1 ± 14.6% et − 16.3 ± 12.8%, p < 0.001, dz ranged from 2.55 to 3.84) and ECC (− 30.5 ± 12.6% et − 23.8 ± 10.3% p < 0.001, dz ranged from 4.62 to 4.82) with a significant difference between ISO and ECC at POST. However, 10/100 Hz ratio was significantly lower after ECC than both ISO and CONC at POSTH1 (p < 0.05, dz ranged from 0.65 to 0.72), but no significant differences were found among the contraction types at POSTD1 (Fig. 3C).
M-wave properties
A significant time effect (F4,120 = 2.87, p < 0.05, pƞ2 = 0.01) was found for the VM first phase amplitude. A significant decrease was observed at POST (p < 0.001, dz = 1.13) and POSTH1 (p < 0.05, dz = 0.93) from PRE. For VM M-wave duration, a significant contraction type × time interaction effect was evident (F8,120 = 2.41, p < 0.05, pƞ2 = 0.14) showing that VM M-wave duration significantly decreased only for ECC at POST (− 10.3 ± 6.7%, p < 0.05, dz = 3.08). However, M-wave duration increased after ECC (+ 16.4 ± 6.1%, p < 0.001, dz = 5.33) and ISO (+ 9.7 ± 9.9%, p < 0.05, d = 1.95) at POSTH1. A significant time effect was observed with a significant increase for VL M-wave duration at POSTH1 (+ 11.0 ± 24.9%, p < 0.05, d = 0.91) (Table 2).
Muscle soreness
A significant contraction type × time interaction (F6,90 = 5.65, p < 0.05, pƞ2 = 0.27) was evident for changes in muscle soreness (Fig. 3D). Muscle soreness increased only for ECC at POST (10.4 ± 7.4 mm p < 0.05, dz = 1.92) and peaked at POSTD1 (26.2 ± 12.6 mm, p < 0.001, dz = 3.28). For ISO condition, muscle soreness peaked at POSTD2 (17.5 ± 21.0 mm, p < 0.05, dz = 1.24). Muscle soreness were significantly greater after ECC than CONC at POSTD1 (p < 0.001, dz = 1.43) and POSTD2 (p < 0.001, dz = 1.72) while ISO was only significantly higher than CONC at POSTD2 (p < 0.05, dz = 1.06).
Discussion
The present study compared neuromuscular fatigue and muscle damage induced by isometric (ISO), concentric (CONC), and eccentric (ECC) contractions at the same amount of torque-time integral. We hypothesized that the loss of MVC torque immediately after CONC exercise would be greater than others, but muscle damage would be greater after ECC than others. Contrary to the hypothesis, the results showed that (i) the decreases in MVC torque immediately after exercise were similar among ISO, CONC and ECC, (ii) MVC torque recovered slower for ECC (POSTD2) and ISO (POSTD1) than CONC (POSTH1), and (iii) DOMS was greater for ECC and ISO than CONC. These results did not support the hypotheses especially for the expected differences between ECC and ISO, but showed distinct differences between CONC and ECC for neuromuscular fatigue and muscle damage profiles.
The exercise protocol was aimed to induce the same torque-time integral for the ISO, CONC and ECC exercises at maximal intensity. To achieve this, the number of contractions was smaller for ISO and ECC than CONC (Table 1), since ISO and ECC contractions produced greater torque than CONC (Fig. 1A). This was in line with previous studies showing that ECC produced higher torque-time integral than CONC, when the level of fatigue was matched at the end of the exercise (Dundon et al. 2008; Souron et al. 2018). It should be noted that number of contractions performed in ECC was approximately 30% smaller than that of CONC, but similar to that of ISO (Table 1). This was due to approximately 30% greater torque production during ECC and ISO when compared with CONC (Fig. 1A). During ISO, CONC and ECC, voluntary activation was not changed over time (Fig. 1C), but evoked torque was progressively reduced for all exercises without any differences among the three conditions (Fig. 1B). Previous studies have also observed similar peak twitch torque reductions among ISO, CONC and ECC contractions during fatiguing exercise (Babault et al. 2006; Doguet et al. 2016). This suggests that for the same torque-time integral, the influence of contraction types on the evoked torque during repeated maximal contractions is similar. This was in contrast to the findings by Babault et al (2006) who reported a greater evoked torque loss during ISO than CONC exercise. This difference may be due to the design of the protocol such that the rest period between sets (60 s) in the study of Babault et al (2006) was shorter than that of the present study (90 s). It should be noted that the similar changes in evoked torque in the CONC and ECC exercises were induced by a smaller number of contractions in ECC than CONC. Thus, if the number of contractions had been matched between ECC and CONC, the decrease in evoked torque would have been greater at the end of ECC than CONC exercise. It is also important to note that voluntary torque was maintained better during ECC than CON (Fig. 1A), but this was not shown by evoked torque (Fig. 1B). ECC appears to be less fatigable than CON and ISO at least for the voluntary torque.
The magnitude of the decrease in MVC torque immediately post-exercise was similar among ECC (− 26%), ISO (− 17%) and CONC (− 22%) as shown in Fig. 3A. In contrast to our results, previous studies reported a greater decrease in MVC torque after CONC than ECC or ISO exercise for the same time under contractions or the same number of contractions (Kay et al. 2000; Pasquet et al. 2000). Souron et al. (2018) and Dundon et al. (2008) showed that the torque-time integral in ECC was greater than that of CONC for the same fatigue level (− 40%) after exercise of the knee extensors and the elbow flexors, respectively. It should be noted that the rest time between sets was shorter during the protocol used by Souron et al. (2018; 1 sets of 50 contractions and X sets of 25 contractions/5-s rest between sets) and Dundon et al (2008; X sets of 10 contractions /20-s rest between sets) when compared with the present study (X sets of 10 contractions /90-s rest between sets). In addition, Pasquet et al. (2000) reported that the torque recovery during rest period between sets was faster after CONC than ECC contractions. Therefore, the longer rest between sets could have minimized the loss of torque in CONC in the present study. In fact, it was demonstrated that a longer recovery duration between sets of CONC contractions minimized the loss of force during exercise (Celes et al. 2010). However, it is interesting that the voluntary torque decreased greater in CONC than ECC as shown in Fig. 1. Thus, the similar loss of MVC torque at immediately post-exercise among contraction types was probably due to a similar effect of the combination between fatigue and muscle damage induced during the exercise as discussed below.
Although the recovery rate of MVC torque from immediately to 1-h post-exercise between contraction types was similar, a faster recovery rate was found for CONC than ECC and ISO (Fig. 3A). Chen et al. (2020) reported that the recovery rate of MVC torque from immediately post- to 1 day post-eccentric exercise was highly correlated with the MVC torque levels in the following days and changes in other indirect muscle damage markers. Thus, the larger recovery of MVC torque from immediately to 1 day post-exercise shown after CONC suggests less muscle damage after CONC than ECC and ISO. It seems likely that CONC produced mainly acute neuromuscular fatigue, while ECC or ISO induced greater excitation–contraction coupling failure and physical damage to muscle contractile proteins (Warren et al 2002).
The loss of MVC torque could result from alterations of central mechanisms. We observed a reduction of voluntary activation at POST (− 3.7 ± 6.6%) and POSTH1 (− 4.7 ± 7.6%) without difference among contraction types, indicating similar central fatigue. The decrease in voluntary activation in the present study was smaller than that observed in the previous studies by Babault et al. (2006) (27% in CONC and 36% in ISO) and Souron et al. (2018) (22% in ECC). Clos et al. (2020) showed that voluntary activation was reduced after ECC but not CONC exercise of the knee extensors with the same amount of work. The small number of contractions in a set (n = 10) and the long rest between sets (90 s) could have contributed to the voluntary activation preservation found in the present study (Carroll et al. 2017). It appears that central alterations are not the main cause of the loss of MVC torque, thus it is more likely due to peripheral alterations.
The ECC protocol induced a greater decrease of evoked torque and the time to peak twitch than ISO and CONC at POST and POSTH1 (Fig. 3B). Moreover, the 10/100 Hz ratio remained lower for ECC than CONC and ISO at POSTH1 (− 23.8%). There was an increase in M-wave duration for the VM and VL at POSTH1 after ECC and ISO (Table 2). Previous studies (Babault et al. 2006; Souron et al. 2018) also reported differences in the alteration in twitch properties following exercise with different contraction types. The reduction of peak twitch torque, 10/100 Hz ratio (Fig. 3) indicates impairment of the excitation–contraction coupling, which is due to disturbances in calcium homeostasis such as a reduction of calcium released by sarcoplasmic reticulum, reduction of myofilament calcium affinity (Fitts 2008) or a calcium accumulation in the sarcoplasm (Armstrong 1984). It has been documented that ECC induces structural damage to sarcolemma and reticulum sarcoplasmic, and alters permeability of the cell membrane, which increases intracellular calcium (Armstrong et al. 1991). This is shown by an increase in the low-frequency fatigue, which is thought to reflect an alteration in calcium homeostasis within excitation–contraction coupling (Hill et al. 2001). Disturbance of excitation–contraction coupling can be also represented by a decrease in M-wave amplitude (Doguet et al. 2016; Pasquet et al. 2000), highlighting an alteration of neuromuscular propagation or a reduction of sarcolemma excitability (Bigland-Ritchie and Woods 1984). Furthermore, damage to the sarcolemma and subsequent activation of stretch-activated ion channels could change sarcolemma excitability, which may explain the differences observed in M-wave properties after ECC (Piitulainen et al. 2008). Thus, it is likely that the ECC induced greater and prolonged impairment of the excitation–contraction coupling than other contraction types.
At POSTH1, MVC torque showed a greater decrease after ECC than other contraction types. According to Paulsen et al. (2012), MVC torque reduction of around − 20% at 2 days after ECC exercise indicates moderate muscle damage. For the same torque-time integral, the prolonged MVC torque loss after ECC was similar to that reported by previous studies (Babault et al. 2006; Dundon et al. 2008; Ochi et al. 2016). It may be that mechanical and neural specificities of ECC contractions such as lower activations of the motor unit or the high external torque (Guilhem et al. 2010) were the main reasons for the greater and longer-lasting decreases in MVC torque after ECC than ISO and CONC. Mean torque during ECC exercise showed that the muscle was producing high tension during lengthening actions, which would likely to provoke more alteration and damage in contractile proteins (Guilhem et al. 2016) than ISO or CONC. The loss of torque after ECC and ISO exercise was greater at POSTD1 and POSTD2 than CONC exercise (Fig. 2). The prolonged loss of torque in the days following ECC and possibly ISO exercise was likely to be attributed to failure of the excitation–contraction coupling as discussed above followed by a progressive degradation of contractile proteins (Warren et al. 2002). Moreover, we observed a greater increase of muscle soreness after ECC and ISO at POSTD1 and POSTD2 than CONC. It seems that ECC as well as ISO to a smaller extent, developed damage within muscular or conjunctive tissues, and triggered an inflammatory response that stimulated group III and IV nociceptive afferents and caused a sensation of pain (Cheung et al. 2003).
It is interesting that ISO contractions induced a prolonged torque loss until POSTD1 and muscle soreness at POSTD2. This is in contrast to the findings of previous studies (Kroon and Naeije 1991; Talag 1973) showing low muscle soreness after ISO exercise, except when isometric contractions were performed at long muscle lengths (Allen et al. 2018). This study confirmed that ISO contractions at long muscle lengths could induce muscle damage as previous studies reported (Allen et al. 2018; Jones et al. 1989; Philippou et al. 2004). Over-stretched sarcomeres are more susceptible to physical disruption due to the lack of interactions between actin and myosin. In turn, this would produce an unequal distribution of constraint into muscle fiber, and even if the constraint is lower in ISO than in ECC, this could provoke structural lesions (Proske and Morgan 2001). Therefore, over-stretched sarcomeres may appear similarly within muscle fibers during ECC and ISO contractions at long muscle lengths, which would explain greater muscle damage for these two contractions types compared to CONC. The torque produced during maximal isometric contractions at a long muscle length may also explain the similar changes in the variables to those shown after ECC. Previous studies showed that ISO contraction at a long muscle length could produce similar (Webber and Kriellaars 1997) or slightly lower torque than ECC (Doguet et al. 2016). To obtain MVC, large motor units are likely to be recruited, and it has been shown that fast-twitch fibers are more susceptible to damage induced by eccentric contractions (Vijayan et al. 2001). It may be that the high force applied to over-stretched fast-twitch fibers in ISO contractions induces similar stress and strain to those during ECC contractions.
The present study found that the evoked torque recovery was slower after ECC (− 15.4% at POSTD1) than CONC or ISO (Fig. 3B). As discussed above, ECC exercise leads to perturbations such as physical damage or excitation–contraction coupling alteration within the muscle (Lieber et al. 1991). Thus, it seems that changes in peripheral function account for a major part of the strength loss after ECC exercise (Hubal et al. 2007). Thus, evoked torque could be more suitable than MVC torque to reveal muscle damage, and the greater and more prolonged decreases in the evoked torque (Fig. 3B) indicate a greater muscle damage after ECC. It seems likely that much greater decreases in the evoked torque would have been observed if the number of eccentric contractions had been similar to that of CONC.
One of the main limitations of the present study was the use of isometric MVC to assess neuromuscular fatigue for all contraction types. Using contraction-mode-specific MVCs might have provided a more sensitive assessment of neuromuscular fatigue. Thus, the isometric MVC measures used in the present study may have induced bias towards the relatively greater decreases in MVC torque after the ISO protocol, despite the magnitude of MVC loss being similar between the three contraction modes. However, whether using contraction-mode specific MVCs provides a more sensitive means of detecting muscle fatigue is unknown, and this presents an interesting area for future research. The second limitation of the present study was the use of the stimulation during exercise for CONC and ECC in a different angle than the angle used during neuromuscular measurement. Although the neuromuscular measurements were performed in isometric conditions, the use of the same knee angle during isokinetic condition could have potentially improved the interpretation of the electrophysiological parameters.
Conclusion
In conclusion, the present study showed that the magnitude of decrease in MVC torque immediately after exercise was similar between ISO, CONC and ECC, but the decrease was prolonged after ECC and ISO that also induced greater muscle soreness when compared with CONC, when the torque-time integral was matched. Recovery in MVC torque were not different between ECC and ISO, but changes in peripheral parameters (M-Wave and twitch properties) were greater after ECC than ISO, suggesting that excitation–contraction coupling was affected more after ECC than ISO. Therefore, ECC and ISO at long muscle length induce greater muscle damage than CONC, and this should be taken into considerations when using these contractions in a practical or clinical context.
Abbreviations
- ISO:
-
Isometric
- CONC:
-
Concentric
- ECC:
-
Eccentric
- MVC:
-
Maximal voluntary contraction
- EMG:
-
Electromyography
- DOMS:
-
Delayed onset muscle soreness
- CK:
-
Creatine kinase
- KE:
-
Knee extensors
- IPAQ:
-
International physical activity questionnaire
- VA:
-
Voluntary activation
- CAR:
-
Central activation ratio
- VAS:
-
Visual analogic scale
- VM:
-
Vastus medialis
- VL:
-
Vastus lateralis
References
Allen GM, Gandevia SC, McKenzie DK (1995) Reliability of measurements of muscle strength and voluntary activation using twitch interpolation. Muscle Nerve 18:593–600. https://doi.org/10.1002/mus.880180605
Allen TJ, Jones T, Tsay A, Morgan DL, Proske U (2018) Muscle damage produced by isometric contractions in human elbow flexors. J Appl Physiol 124:388–399. https://doi.org/10.1152/japplphysiol.00535.2017
Ansdell P, Dekerle J (2020) Sodium bicarbonate supplementation delays neuromuscular fatigue without changes in performance outcomes during a basketball match simulation protocol. J Strength Cond Res 34:1369–1375. https://doi.org/10.1519/JSC.0000000000002233
Armstrong RB (1984) Mechanisms of exercise-induced delayed onset muscular soreness: a brief review. Med Sci Sports Exerc 16:529–538
Armstrong R, Warren G, Warren J (1991) Mechanisms of exercise-induced muscle fibre injury. Sports Med 12:184–207. https://doi.org/10.2165/00007256-199112030-00004
Babault N, Pousson M, Ballay Y, Van Hoecke J (2001) Activation of human quadriceps femoris during isometric, concentric, and eccentric contractions. J Appl Physiol 91:2628–2634. https://doi.org/10.1152/jappl.2001.91.6.2628
Babault N, Desbrosses K, Fabre MS, Michaut A, Pousson M (2006) Neuromuscular fatigue development during maximal concentric and isometric knee extensions. J Appl Physiol 100:780–785. https://doi.org/10.1152/japplphysiol.00737.2005
Bigland-Ritchie B, Woods JJ (1984) Changes in muscle contractile properties and neural control during human muscular fatigue. Muscle Nerve 7:691–699. https://doi.org/10.1002/mus.880070902
Byrne C, Twist C, Eston R (2004) Neuromuscular function after exercise-induced muscle damage: theoretical and applied implications. Sports Med 34:49–69. https://doi.org/10.2165/00007256-200434010-00005
Carroll TJ, Taylor JL, Gandevia SC (2017) Recovery of central and peripheral neuromuscular fatigue after exercise. J Appl Physiol 122:1068–1076. https://doi.org/10.1152/japplphysiol.00775.2016
Cattagni T, Merlet AN, Cornu C, Jubeau M (2018) H‐reflex and M‐wave recordings: effect of pressure application to the stimulation electrode on the assessment of evoked potentials and subject's discomfort. Clin Physiol Funct Imaging 38(3):416–424
Celes R, Brown LE, Pereira MC, Schwartz FP, Junior VA, Bottaro M (2010) Gender muscle recovery during isokinetic exercise. Int J Sports Med 31:866–869. https://doi.org/10.1055/s-0030-1254156
Chen TC, Huang GL, Hsieh CC, Tseng KW, Tseng WC, Chou TY, Nosaka K (2020) Comparison among three different intensities of eccentric contractions of the elbow flexors resulting in the same strength loss at one day post-exercise for changes in indirect muscle damage markers. Eur J Appl Physiol 120(1):267–279
Cheung K, Hume PA, Maxwell L (2003) Delayed onset muscle soreness. Sports Med 33:145–164. https://doi.org/10.2165/00007256-200333020-00005
Clos P, Garnier Y, Martin A, Lepers R (2020) Corticospinal excitability is altered similarly following concentric and eccentric maximal contractions. Eur J Appl Physiol 120:1457–1469. https://doi.org/10.1007/s00421-020-04377-7
Cohen J (1988) Statistical Power Analysis for the Behavioral Sciences (2nd edn), 411 Lawrence Erlbaum Associates Hillsdale, NJ 412
Doguet V, Nosaka K, Plautard M, Gross R, Guilhem G, Guevel A, Jubeau M (2016) Neuromuscular changes and damage after isoload versus isokinetic eccentric exercise. Med Sci Sports Exerc 48:2526–2535. https://doi.org/10.1249/MSS.0000000000001042
Duchateau J, Enoka RM (2016) Neural control of lengthening contractions. J Exp Biol 219:197–204. https://doi.org/10.1242/jeb.123158
Dundon JM, Cirillo J, Semmler JG (2008) Low-frequency fatigue and neuromuscular performance after exercise-induced damage to elbow flexor muscles. J Appl Physiol 105:1146–1155. https://doi.org/10.1152/japplphysiol.01339.2007
Edwards RH, Hill DK, Jones DA, Merton PA (1977) Fatigue of long duration in human skeletal muscle after exercise. J Physiol 272:769–778. https://doi.org/10.1113/jphysiol.1977.sp012072
Enoka RM, Stuart DG (1992) Neurobiology of muscle fatigue. J Appl Physiol 72:1631–1648. https://doi.org/10.1152/jappl.1992.72.5.1631
Fitts RH (2008) The cross-bridge cycle and skeletal muscle fatigue. J Appl Physiol 104:551–558. https://doi.org/10.1152/japplphysiol.01200.2007
Guilhem G, Cornu C, Guevel A (2010) Neuromuscular and muscle-tendon system adaptations to isotonic and isokinetic eccentric exercise. Ann Phys Rehabil Med 53:319–341. https://doi.org/10.1016/j.rehab.2010.04.003
Guilhem G, Doguet V, Hauraix H, Lacourpaille L, Jubeau M, Nordez A, Dorel S (2016) Muscle force loss and soreness subsequent to maximal eccentric contractions depend on the amount of fascicle strain in vivo. Acta Physiol 217:152–163. https://doi.org/10.1111/apha.12654
Hill CA, Thompson MW, Ruell PA, Thom JM, White MJ (2001) Sarcoplasmic reticulum function and muscle contractile character following fatiguing exercise in humans. J Physiol 531(3):871–878
Hubal MJ, Rubinstein SR, Clarkson PM (2007) Mechanisms of variability in strength loss after muscle-lengthening actions. Med Sci Sports Exerc 39:461–468. https://doi.org/10.1249/01.mss.0000247007.19127.da
Jones DA, Newham DJ, Torgan C (1989) Mechanical influences on long-lasting human muscle fatigue and delayed-onset pain. J Physiol 412:415–427. https://doi.org/10.1113/jphysiol.1989.sp017624
Kay D, St Clair Gibson A, Mitchell MJ, Lambert MI, Noakes TD (2000) Different neuromuscular recruitment patterns during eccentric, concentric and isometric contractions. J Electromyogr Kinesiol 10:425–431. https://doi.org/10.1016/s1050-6411(00)00031-6
Kroon G, Naeije M (1991) Recovery of the human biceps electromyogram after heavy eccentric, concentric or isometric exercise. Eur J Appl Physiol 63:444–448
Lieber RL, Woodburn TM, Friden J (1991) Muscle damage induced by eccentric contractions of 25% strain. J Appl Physiol 70:2498–2507. https://doi.org/10.1152/jappl.1991.70.6.2498
McHugh MP (2003) Recent advances in the understanding of the repeated bout effect: the protective effect against muscle damage from a single bout of eccentric exercise. Scand J Med Sci Sports 13:88–97. https://doi.org/10.1034/j.1600-0838.2003.02477.x
Ochi E, Tsuchiya Y, Nosaka K (2016) Differences in post-exercise T2 relaxation time changes between eccentric and concentric contractions of the elbow flexors. Eur J Appl Physiol 116:2145–2154. https://doi.org/10.1007/s00421-016-3462-3
Pasquet B, Carpentier A, Duchateau J, Hainaut K (2000) Muscle fatigue during concentric and eccentric contractions. Muscle Nerve 23:1727–1735. https://doi.org/10.1002/1097-4598(200011)23:11%3c1727::aid-mus9%3e3.0.co;2-y
Paulsen G, Mikkelsen UR, Raastad T, Peake JM (2012) Leucocytes, cytokines and satellite cells: what role do they play in muscle damage and regeneration following eccentric exercise? Exerc Immunol Rev 18:42–97
Philippou A, Bogdanis GC, Nevill AM, Maridaki M (2004) Changes in the angle-force curve of human elbow flexors following eccentric and isometric exercise. Eur J Appl Physiol 93:237–244. https://doi.org/10.1007/s00421-004-1209-z
Piitulainen H, Komi P, Linnamo V, Avela J (2008) Sarcolemmal excitability as investigated with M-waves after eccentric exercise in humans. J Electromyogr Kinesiol 18:672–681. https://doi.org/10.1016/j.jelekin.2007.01.004
Pincivero D, Lephart S, Karunakara R (1997) Reliability and precision of isokinetic strength and muscular endurance for the quadriceps and hamstrings. Int J Sports Med 18:113–117. https://doi.org/10.1055/s-2007-972605
Place N, Matkowski B, Martin A, Lepers R (2006) Synergists activation pattern of the quadriceps muscle differs when performing sustained isometric contractions with different EMG biofeedback. Exp Brain Res 174:595–603. https://doi.org/10.1007/s00221-006-0504-9
Proske U, Morgan DL (2001) Muscle damage from eccentric exercise: mechanism, mechanical signs, adaptation and clinical applications. J Physiol 537:333–345. https://doi.org/10.1111/j.1469-7793.2001.00333.x
Roig M, O’Brien K, Kirk G, Murray R, McKinnon P, Shadgan B, Reid W (2009) The effects of eccentric versus concentric resistance training on muscle strength and mass in healthy adults: a systematic review with meta-analysis. Br J Sports Med 43:556–568. https://doi.org/10.1136/bjsm.2008.051417
Souron R, Nosaka K, Jubeau M (2018) Changes in central and peripheral neuromuscular fatigue indices after concentric versus eccentric contractions of the knee extensors. Eur J Appl Physiol 118:805–816. https://doi.org/10.1007/s00421-018-3816-0
Strojnik V, Komi PV (1998) Neuromuscular fatigue after maximal stretch-shortening cycle exercise. J Appl Physiol 84:344–350. https://doi.org/10.1152/jappl.1998.84.1.344
Talag TS (1973) Residual muscular soreness as influenced by concentric, eccentric, and static contractions Research Quarterly American Association for Health. Phys Educ Recreat 44:458–469
Vijayan K, Thompson JL, Norenberg KM, Fitts R, Riley DA (2001) Fiber-type susceptibility to eccentric contraction-induced damage of hindlimb-unloaded rat AL muscles. J Appl Physiol 90:770–776. https://doi.org/10.1152/jappl.2001.90.3.770
Warren GL, Ingalls CP, Lowe DA, Armstrong R (2002) What mechanisms contribute to the strength loss that occurs during and in the recovery from skeletal muscle injury? J Orthop Sports Phys Ther 32:58–64
Webber S, Kriellaars D (1997) Neuromuscular factors contributing to in vivo eccentric moment generation. J Appl Physiol 83:40–45. https://doi.org/10.1152/jappl.1997.83.1.40
Author information
Authors and Affiliations
Contributions
NR, KN, VD and MJ conceived and designed research. NR conducted the experiments and collected the data. NR analyzed the data. NR, KN, VD and MJ drafted the manuscript. All authors provided critical appraisal to the manuscript and approved the final version. All authors have read and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by Nicolas Place.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Royer, N., Nosaka, K., Doguet, V. et al. Neuromuscular responses to isometric, concentric and eccentric contractions of the knee extensors at the same torque-time integral. Eur J Appl Physiol 122, 127–139 (2022). https://doi.org/10.1007/s00421-021-04817-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-021-04817-y