
Abstract
Purpose
This study examined the mechanisms for force and power reduction during and up to 48 h after maximal eccentric contractions of the knee extensor muscles in young men and women.
Methods
13 men (22.8 ± 2.6 years) and 13 women (21.6 ± 2.2 years) performed 150 maximal effort eccentric contractions (5 sets of 30) with the knee extensor muscles at 60° s−1. Maximal voluntary isometric contractions (MVIC) and maximal voluntary concentric contractions (MVCC) were performed before and after the 150 eccentric contractions. The MVCCs involved a set of two isokinetic contractions at 60° s−1 and sets of isotonic contractions performed at seven different resistance loads (1 N m, 10, 20, 30, 40, 50, and 60% MVIC). Electrical stimulation was used during the MVICs and at rest to determine changes in voluntary activation and contractile properties.
Results
At baseline, men were stronger than women (MVIC: 276 ± 48 vs. 133 ± 37 N m) and more powerful (MVCC: 649 ± 77 vs. 346 ± 78 W). At termination of the eccentric contractions, voluntary activation, resting twitch amplitude, and peak power during concentric contractions at the seven loads and at 60° s−1 decreased (P < 0.05) similarly in the men and women. At 48 h post-exercise, the MVIC torque, power (for loads ≥20–60% MVIC), and voluntary activation remained depressed (P < 0.05), but the resting twitch had returned to baseline (P > 0.05) with no sex differences.
Conclusion
Central mechanisms were primarily responsible for the depressed maximal force production up to 48 h after repeated eccentric contractions of the knee extensors and these mechanisms were similar in men and women.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Eccentric or lengthening muscle contractions occur when an external force applied to the muscle exceeds the force produced, resulting in the muscle fibers lengthening while being activated (Lindstedt et al. 2001). Eccentric contractions help promote neuromuscular adaptations during training and rehabilitation to increase muscle mass, strength, balance, and power. These adaptations are possible because of the large forces obtained during eccentric contractions that exceed isometric or concentric (shortening) maximal force and at a relatively low metabolic cost (LaStayo et al. 2003, 2014). When a muscle is actively lengthened, the cross bridges develop more tension as they operate higher on the force-tension curve. The actin–myosin bonds may be broken largely by mechanical means, so in comparison to concentric activation, less adenosine triphosphate (ATP) is required for cross bridge cycling (Nishikawa 2016). However, the high forces can lead to myofibril damage when muscles are required to perform repetitive unaccustomed lengthening contractions (Nishikawa 2016). Symptoms of muscle damage include disruption of the sarcomeres, sarcolemma, and extracellular matrix, as well as delayed-onset muscle soreness and inflammation that peaks 24–72 h after the eccentric contractions (Byrne et al. 2004).
Fatigability (often referred to a muscle fatigue) is the exercise induced reduction in force or power of muscle (Enoka and Duchateau 2008). In contrast to muscle damage, it is reversible usually within several hours (Allen 2001). While motor output is ultimately reduced at the muscle, force reductions can originate from various neuromuscular sites including inadequate activation of the motor units by the central nervous system and processes at the contractile proteins of the exercising muscles (Gandevia 2001). Thus, immediately after eccentric contractions, the reduction in muscle function could be due to processes associated with muscle fatigue, muscle damage, or both. For eccentric exercise, it has been suggested that impairment in excitation–contraction coupling is largely responsible for acute force reduction (muscle fatigue), whereas muscle damage may account for reduction in muscle function observed up to 72 h days following eccentric contraction tasks (Martin et al. 2004).
Isometric contractions and repeated concentric contractions do not generally cause muscle damage, and women are usually less fatigable than men during fatiguing contractions with these contraction types (Hunter 2014, 2016a). For repeated eccentric contractions, however, there is a poor understanding of whether men and women differ in their recovery of muscle function. Several studies indicate that there were no sex differences immediately after or up to 72 h after eccentric contractions for maximal voluntary isometric contraction (MVIC) force, work rate, and rate of torque development (Hubal et al. 2008; Sayers and Clarkson 2001; Rinard et al. 2000; Hicks et al. 2016). However, Sewright and colleagues (2008) reported that women had a greater loss of strength in the elbow flexor muscles immediately following maximal eccentric contractions compared with men, with no sex differences during the 10-day recovery period of maximal strength nor in the time-course of muscle soreness (Sewright et al. 2008). For a lower limb muscle, the ankle dorsiflexors, Power and colleagues (2013) showed that MVIC was depressed more so in women than men during recovery at 10-min–24-h post-task (Power et al. 2013). Furthermore, they also found that women had a greater loss of power at higher force loads (≥60% MVIC) during maximal voluntary shortening contractions, with a delayed recovery for up to 48 h following the eccentric task, despite similar levels of self-reported soreness for men and women (Power et al. 2013). The mechanisms for the sex differences at the high-force contractions were attributed to muscular mechanisms involving excitation–contraction coupling and reduced rates of torque development (Power et al. 2013).
Given the paucity of data on other muscle groups such as the knee extensor muscle that are important for locomotion and during rehabilitation and sports performance, the purpose of this study was to compare the reduction in muscle function and indices of neural and muscular mechanisms following repeated eccentric contractions of the knee extensor muscles in men and women. We hypothesized that after 150 maximal effort eccentric contractions of the knee extensors, women would have greater reductions than men in maximal strength and power immediately after and up to 48 h of recovery. We also hypothesized that both neural and muscular mechanisms would contribute to the reductions in strength and power of the knee extensor muscles in both sexes, but any sex difference would be primarily due to deficits in contractile function in the women. Finally, to determine the long-term effects of the repeated eccentric contractions, we quantified muscle function and indices of neural and muscular mechanisms 14 days after the eccentric contractions in a subset of participants.
Methods
Thirteen young men (22.8 ± 2.6 years) and thirteen young women (21.5 ± 2.2 years) volunteered to participate in the study. Participants were recreationally active college students, who did not participate in regular exercise training and were not college athletes. They were asked to refrain from strenuous exercise 48 h prior to the first testing session, and to avoid additional exercise throughout the 3-day testing period. All participants provided informed consent prior to participation. This study was approved by the Michigan Technological University’s Institutional Review Board for the Protection of Human Subjects.
All participants attended a familiarization session involving a physical activity questionnaire (Kriska and Bennett 1992), and the collection of physical characteristics (height, weight, and age). Body fat percentage of each participant was estimated from electrical impedance with a Tanita body composition analyzer (BC-418, Tanita Corp., Tokyo, Japan). All participants were habituated to the electrical stimulation of the quadriceps muscles and practiced MVICs, MVCCs, and eccentric contractions during a submaximal effort (<20% MVIC) for ten repetitions only to familiarize the participant without eliciting considerable fatigue or muscle damage. Subjects used their non-dominant leg for the familiarization session, while the dominant leg was used for the testing protocol. One exception occurred where a female participant used her non-dominant leg for both the familiarization session and testing procedure, due to a knee injury on her dominant leg. Hand dominance was estimated using the Edinburgh Handedness Inventory (Oldfield 1971). Dominant leg was based on a combination of both the Edinburgh Handedness Questionnaire as well as the subject self-reporting which leg they favor in sports (i.e., kicking a soccer ball, snowboarding, jumping, etc.).
Experimental set up and mechanical recordings
Experimental sessions were performed with the participant seated in a Biodex multi-joint dynamometer (System 4 Pro; Biodex Medical System, Shirley, NY, USA). Each participant was seated in a slightly reclined position with the hip and knee angles at 95° and 90°, respectively. Participant’s shank was strapped to the distal end of the Biodex lever, with the lateral epicondyle of the femur aligned with the axis of rotation of the dynamometer.
All voluntary and evoked isometric contractions were performed at 90° of flexion (0° being where the knee joint is fully extended and the lower limb horizontal). This angle was kept consistent to compare relative changes in angle and force production throughout the testing sessions (De Ruiter et al. 2004). Shortening (concentric) contractions moved through a 60° range of motion from 90° to 30° of flexion and lengthening (eccentric) contractions moved from 30° back to 90° of flexion. Torque, angle, and angular velocity data were sampled at a rate of 2000 Hz using a micro 1401 AD converter and Spike 2 software [Version 8, Cambridge Electronics Design (CED), Cambridge, UK]. Torque signal was displayed on a 70-in TV monitor (Sharp Electronics, New Jersey, USA) located 2.5 m in front of the subject.
Electrical recordings
Surface electromyography (EMG) (Bagnoli 16; Delsys, Natick, MA, USA) was used to record muscle activity of the knee extensors, including the rectus femoris (RF), vastus lateralis (VL), and vastus medialis (VM). Electrode placement was determined according to recommendations by Surface Electromyography for the Noninvasive Assessment of Muscles (SENIAM Project) (Hermens et al. 2000). The ground electrode was positioned over the patella. The skin was scrubbed with alcohol soaked cleansing cloths before electrode placement, and location was marked via a permanent pen to ensure that placement was consistent for the entirety of the testing sessions. The EMG signal was amplified (100×), bandpass filtered (10–450 Hz), and sampled at a rate of 2000 Hz using a micro 1401 AD converter and Spike 2 software (CED, Cambridge, UK).
Electrical stimulation
All four quadriceps muscles were electrically stimulated to determine contractile properties from a single maximal stimulus at rest and an additional stimulus was also applied during MVICs to determine voluntary activation. A single electrical pulse (square wave, 100-μs duration) was elicited via a pair of self-adhesive surface electrodes placed over the muscle (6.98 × 12.7 cm, Dura-Stick plus DJO Brands) and with a computer-controlled stimulator (D185; Digitimer, Welwyn Garden City, UK). Although multiple stimulations can decrease variability in the interpolated twitch torque (Suter and Herzog 2001), a single pulse was used to minimize discomfort and is sensitive to changes with fatigue (Bampouras et al. 2006; Kennedy et al. 2015; Yoon et al. 2007). The exact electrode positions were marked with a permanent pen, allowing the investigator to replicate positioning of electrode pads for subsequent trials. The superior aspect of the proximal electrode (anode) was positioned in line with the greater femoral trochanter with the midline of the electrode aligned with the anterior–superior iliac spine (Pietrosimone et al. 2011). The distal electrode (cathode) was positioned, so that the inferior aspect of the electrode sat ~3 cm superior to the patella, with the medial border of the electrode aligned with the midline of the patella (Pietrosimone et al. 2011). At the start of each testing session, the stimulation voltage was increased until the twitch force response plateaued. Voltage was further increased by an additional 20% to ensure full activation of all motor units by supramaximal stimulation. This supramaximal voltage intensity (120%) was used for all electrically evoked contractions for the remainder of the testing session for each individual.
Muscle soreness and perceived exertion
Muscle soreness was assessed using a 10-cm visual analog scale, self-reported by the subject (Lau et al. 2015). The left anchor of the scale was titled “no pain”, and the right anchor titled “worst pain ever”. While performing a 10-s sustained isometric contraction at 20% of MVIC of baseline, participants indicated their level of soreness of their quadriceps muscles. This measure was performed at baseline, after warm-up and following each set of task measures during recovery.
Rating of perceived exertion (RPE) was assessed with the modified Borg ten-point scale (Borg 1982). The participants were instructed to focus the assessment on the effort of their knee extensor muscles while performing the eccentric contraction fatiguing task. The scale was anchored, so that 0 represented “resting state” and 10 represented “maximal exertion” that the knee extensors could perform. The RPE was measured at regular intervals occurring once during each of the five sets of the eccentric fatigue task, at approximately the 25th repetition.
Experimental protocol
Baseline measures
Maximal voluntary isometric contractions (MVIC)
Three MVICs were performed by each participant, for 3–5 s followed by 2-min rest between trials. If two of the three MVICs values were not within 5% of each other, a fourth trial was performed. Visual feedback of the torque-time tracing was given to the participants on a 70-in TV monitor, as well as verbal encouragement to ensure maximal effort during all MVICs. During each MVIC, electrical stimulation was elicited to evoke a superimposed twitch. An additional twitch was evoked at rest (resting twitch) ~1 s following the MVIC.
Maximal voluntary concentric contractions (MVCCs)
There were two types of shortening contraction modes used—isokinetic and isotonic contractions. It should be noted that due to mechanical limitations of the dynamometer, the isotonic contractions cannot be considered truly isotonic because of the dynamometer’s inability to maintain an internal constant load throughout the complete range of motion (Power et al. 2011). For both contraction modes, the dynamometer was programmed to allow the active range of motion at the knee joint to extend from 90° to 30° and the dynamometer passively returned the shank back to the 90° starting position. Familiarization and warm-up for these shortening contractions consisted of a total of ten submaximal repetitions (two sets of five reps, with 30-s rest period between sets) of isotonic contractions at moderate load (35 N m for women, 50 N m for men).
After 5 min of rest following the warm-up, two repetitions of isokinetic shortening contractions at 60° s−1 were performed for baseline values to assess changes in muscle series compliance. Participants were asked to maximally extend the dynamometer arm throughout the full 60° range of motion of knee extension. Participants were provided with verbal encouragement and real-time torque feedback was displayed on a TV monitor (Campenella et al. 2000).
After 5 min of rest following the isokinetic shortening contractions, seven sets of isotonic contractions at various predetermined resistance loads were performed, including 1 N m, 10, 20, 30, 40, 50, and 60% of MVIC. These loads were performed by each participant in randomized order, and for each participant, this order remained constant for the duration of the testing procedures. Participants were instructed to move the resistance load as “fast and hard as possible” throughout the full 60° range of motion. Participants were provided with verbal encouragement and real-time torque feedback displayed on a TV monitor to encourage a maximal effort (maximal velocity) (Campenella et al. 2000). Two consecutive repetitions at each resistance were performed to improve the chances that true maximal velocity was reached. Thirty seconds of rest separated the sets of contractions. The peak velocity reached at each resistance load was used to establish baseline values for angular velocity at each of the seven resistance loads, with the peak angular velocity (obtained at 1 N m) considered the maximal shortening velocity. Power was calculated across each of the seven resistance loads, with the peak power being the highest product of torque and velocity at any given time-point during the contraction.
Repeated lengthening contractions
Participants performed 5 sets of 30 repetitions of maximal effort eccentric contractions at 60° s−1, with 10 s between each set. Participants were asked to maximally resist the lowering of the shank by the dynamometer to forced flexion throughout the full range of motion (30–90°). The shank was then passively returned to the start position of 30° of flexion. Verbal encouragement and visual feedback of the real-time torque curve on the TV screen were provided to the participant to encourage maximal effort (Kellis and Baltzopoulos 1996).
Recovery measures
MVIC and MVCCs (both isokinetic and isotonic contractions) measures were performed during recovery at the following times: immediately upon completion of eccentric fatiguing task, and then at 10, 20, 30 min, 24, and 48 h following the task (see Fig. 1). In addition, 9 of 26 men and women (4 men, 5 women) in this study were tested after 14 days following 150 maximal eccentric contractions of the knee extensors.

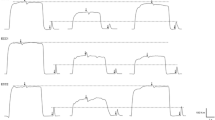
Schematic experimental protocol (a) and representative data (b). White solid rectangles indicate MVICs. Gray solid triangles indicate maximal isokinetic dynamic shortening contractions. Vertical striped gray rectangle indicates isotonic loaded dynamic shortening contractions. Black rectangles indicate a set of 30 maximal eccentric contractions. Recovery time-points were immediately upon task termination, 10, 20, 30 min, 24, and 48 h. Representative MVIC recordings in a man and woman (left and right, respectively). Solid line baseline, gray line post fatigue
Data analysis
Data analysis of angle, torque, velocity, power, and EMG signals was performed offline with Spike 2 software. The torque signal was low-pass filtered with a 10-Hz cut-off frequency. MIVC torque was quantified as the average torque of 100-ms duration prior to electrical stimulation during the MVIC.
Voluntary activation was assessed by measuring the torque response in knee extensor electrical stimulation. Both the peak amplitude of the interpolated twitch (IT) and the resting twitch (RT) produced by electrical stimulation in a relaxed but potentiated muscle (1 s after the MVIC), were used to calculate voluntary activation with the following formula: voluntary activation (%) = 100 × (1 − IT/RT) (Gandevia 2001).
Optimal angle of torque production during the 60° s−1 isokinetic contraction was identified as the angle at which the maximal torque was produced during each contraction. The change in optimal angle of torque production was calculated as the difference between the optimal angle of torque production before and after the eccentric task.
Knee extension power (watt) was calculated as the product of torque (N m) and angular velocity (rad s−1). Peak power was determined for each of the seven predetermined loads used in the isotonic contractions.
EMG activity of the knee extensor muscles was determined as the root-mean-squared (RMS) value over 100-ms interval, which was time interval used to determine the MVIC torque. All subsequent MVIC RMS values were normalized to the level obtained during the baseline MVIC.
Statistical analysis
Data are reported as means ± SD within the text and displayed as means ± SEM in the figures. Normality and homogeneity were tested using the Shapiro–Wilk and Levene tests, respectively. Because voluntary activation data were not normally distributed, Friedman’s ANOVA and Mann–Whitney U test were performed. Univariate analyses were used to compare subject characteristics, physical activity levels, baseline variables including MVIC torque, resting twitch amplitude, maximal velocity during maximal isokinetic contraction; peak power during maximal isotonic contraction in men and women. Two-way analysis of variance (ANOVA) with repeated measures was used to compare variables in men and women across time-points (baseline, post, 10, 20, 30 min, 24, and 48 h). For the nine participants who completed the testing at 14-day post-exercise, same two-way ANOVA with repeated measures was performed to compare variables. When an interaction (sex × time) was found, independent t tests were used to compare groups at each time interval. The variables include MVIC torque, voluntary activation, resting twitch, peak power, and optimal angle of torque production. The strength of an association between fatigue-related variables is reported as the Pearson product-moment correlation coefficient (r). To test the difference in coefficients between men and women, Fisher r-to-z transformation tests were also performed. Significant differences were noted at P < 0.05.
Results
Baseline measures
Table 1 shows the subject characteristics and baseline data. Self-reported physical activity levels were similar for men and women (P = 0.97); however, men were taller, leaner, and had more body mass than women (P < 0.05). Men were stronger and more powerful, and had a greater resting twitch torque amplitude compared with women (P < 0.05). There was no differences between men and women in their optimal angle for torque development during a 60° s−1 maximal isokinetic shortening contraction, the relative load in which peak power occurred during maximal isotonic contractions, nor the level of voluntary activation during MVICs at baseline (Table 1).
Fatigue and recovery from dynamic lengthening fatiguing session
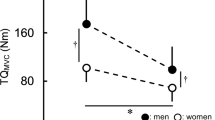
MVIC torque decreased following the eccentric fatiguing contractions from baseline by 39.4 ± 21.5% (fatigue effect, F 1,24 = 84.0, P < 0.001, η 2 p = 0.778). The relative decline was similar in men and women (40.7 ± 21.1 vs. 38.1 ± 22.8% respectively; sex × time; F 1,24 = 0.098, P = 0.757, η 2 p = 0.004). During recovery, MVIC torque increased (recovery effect; F 5,24 = 3.88, P = 0.012, η 2 p = 0.139), and the relative increase was similar in men and women (recovery × sex; F 5,24 = 1.373, P = 0.258, η 2 p = 0.054). MVIC had not reached baseline levels by 48 h of recovery with both sexes showing reduced MVIC torque at 48-h recovery compared with baseline (recovery, F 1,24 = 41.7, P < 0.001, η 2 p = 0.635). See Fig. 2.

Maximal voluntary isometric contraction torque before and in recovery from the eccentric fatiguing task. Men were stronger than women at baseline (P < 0.05). MVIC decreased following the eccentric contraction task for both men and women (P < 0.05), and did not recover after 48 h of rest (P < 0.05). No sex differences in the relative fatigue or recovery pattern for MVIC were seen. Neither sex fully recovered MVIC strength by 48 h. *Time effect, P < 0.05; †sex effect, P < 0.05
Resting twitch torque decreased following the eccentric fatiguing contractions from control values by 26.4 ± 28.7% (fatigue effect, F 1,24 = 21.2, P < 0.001, η 2 p = 0.469). The relative decrease was similar for men and women (sex × fatigue, F 1,24 = 0 0.012, P = 0.913, η 2 p = 0 0.001). During recovery, twitch amplitude was depressed for 30 min and increased after 24 h of recovery (recovery effect; F 5,24 = 49.8, P < 0.001, η 2 p = 0.675). The relative increase was similar for men and women (sex × recovery; F 5,24 = 0.633, P = 0.533, η 2 p = 0.026). For both sexes, resting twitch recovered by 48 h in comparison to baseline (recovery effect, F 1,24 = 2.591, P = 0.121, η 2 p = 0 0.097). See Fig. 3a.

Resting twitch (a) and voluntary activation (b) before and in recovery from the eccentric fatiguing task. RT decreased following the eccentric contraction task for both men and women (P < 0.05), and recovered after 24 and 48 h of rest (P > 0.05). VA decreased following the eccentric contraction task for both men and women (P < 0.05), and did not recover after 48 h of rest. No sex differences in the relative fatigue and recovery pattern between the sexes were seen in either RT or VA. *Time effect, P < 0.05; †sex effect, P < 0.05
Voluntary activation during MVICs decreased 13.7 ± 22.1% following the eccentric fatiguing contractions from control values (fatigue effect, P < 0.001). The relative decrease was similar for men and women (sex effect, P = 0.840). Voluntary activation values did not return to baseline levels by 48 h after the task (P = 0.049). See Fig. 3b.
Peak power for all loads with the exception of the 1 N m condition (F 1,23 = 9.36, P = 0.059, η 2 p = 0.147) significantly decreased following the eccentric fatiguing contractions (0 < P < 0.05). The relative decline was similar in men and women for all resistance loads (sex × fatigue, 0.169 < P < 0.807). During recovery, peak power at 10% MVIC increased (recovery effect, F 5,24 = 2.847, P = 0.040, η 2 p = 0.106), and this increase was similar for both men and women (sex × recovery, F 1,24 = 1.433, P = 0.238, η 2 p = 0.056). Peak power at 20–50% MVIC did not recover to baseline levels for either men or women (recovery effect, 0.053 < P < 0.456). However, peak power at 60% MVIC increased (recovery effect, F 5,22 = 4.124, P = 0.013, η 2 p = 0.158) and the relative increase was different for men and women (sex × recovery, F 1,22 = 3.99, P = 0.015, η 2 p = 0.154). This was because men increased power between 30 min and 24 h, but women showed a decrease in power during this time period. See Fig. 4.

Peak power during isotonic MVCCs across multiple torque loads. Each graph represents a time-point during recovery from the eccentric fatiguing task. Peak power decreased following the eccentric contraction task for both men and women for every intensity condition (P < 0.05), with the exception of the 1-N m condition (P > 0.05). The magnitude of reduction in peak power following the eccentric contractions increased as contraction intensity increased. The higher intensity loads also took longer to recover. The moderate intensity loads (20–60% MVIC) did not fully recovery by 48 h (P < 0.05)
EMG activity during MVIC decreased from baseline following the eccentric fatiguing task for rectus femoris (fatigue effect, F 1,23 = 7.519, P = 0.012, η 2 p = 0.246), vastus medialis (fatigue effect, F 1,23 = 23.7, P = 0.001, η 2 p = 0.507), and vastus lateralis (fatigue effect, F 1,23 = 12.2, P = 0.002, η 2 p = 0.347). The change in RMS EMG during the MVIC immediately following the fatiguing task was similar between men and women for the rectus femoris (fatigue × sex, F 1,23 = 2.643, P = 0.118, η 2 p = 0.103), vastus medialis (fatigue × sex, F 1,23 = 0.002, P = 0.964, η 2 p = 0.001), and vastus lateralis (F 1,23 = 0.324, P = 0.576, η 2 p = 0.14). Furthermore, during recovery, the EMG activity during the MVIC changed similarly for men and women up to 30-min post-eccentric contractions for the rectus femoris (recovery × sex, F 3,23 = 2.192, P = 0.114, η 2 p = 0.087), vastus medialis (recovery × sex, F 3,23 = 0.434, P = 0.658, η 2 p = 0.019), and vastus lateralis (recovery × sex, F 3,22 = 0.325, P = 0.774, η 2 p = 0.015). The RMS EMG during the MVIC decreased following the eccentric contractions did not recover to baseline after 30 min of recovery in men and women for the rectus femoris (79.0 ± 9.5 vs. 84.7 ± 7.5%, respectively), vastus medialis (68.2 ± 6.7 vs. 78.6 ± 13.9%, respectively), and vastus lateralis (66.6 ± 9.7 vs. 74.3 ± 12.8%, respectively).
Muscle soreness reported on an analog scale of 1–10 by each participant while performing a 10 s sustained isometric contraction at 20% of MVIC, increased from baseline after completion of the eccentric contraction fatiguing task, and continued to increase up until 30 min after the task (time effect, F 5,24 = 6.07, P = 0.001, η 2 p = 0.202) for both men and women (from baseline to 30 min: 0.01 ± 0.1–2.2 ± 2.2 vs. 0.03 ± 0.03–2.1 ± 2.7 respectively). By 48 h, muscle soreness remained elevated (recovery effect, F 1,24 = 17.1, P < 0.001, η 2 p = 0.416) for men and women (1.7 ± 2.1 vs. 1.7 ± 2.0, respectively). There were no sex differences in muscle soreness values at baseline or at any time-points following the eccentric contractions (time × sex, F 5,24 = 0.176, P = 0.971, η 2 p = 0. 007).
Rating of perceived exertion (RPE) increased (time effect, F = 31.5, P < 0.001, η 2 p = 0.022) during the eccentric fatiguing task from 4.2 ± 1.7 during the first set of 30 contractions to 7.3 ± 2.3 during the last (5th) set of contractions. The relative increase was similar for both men and women (time × sex, F = 0.543, P = 0.619, η 2 p = 0.022).
Recovery at 14-day post-exercise for a subgroup (4 men and 5 women)
The MVIC torque and voluntary activation increased from 48-h post-eccentric contractions to 14-day post-contractions. However, recovery was not complete, so that by day 14, MVIC torque was at 77.9 ± 37% of baseline (P = 0.007, η 2 p = 0.673) and voluntary activation had recovered to 86.5 ± 18% (P = 0.008, η 2 p = 0.658). In contrast, resting twitch amplitude had recovered to baseline levels by day 14 (P = 0.658, η 2 p = 0.030, compared to baseline).
Associations with reductions in strength and power
The reduction in the MVIC torque performed immediately after the 150 eccentric contractions was associated with the reduction of voluntary activation (r = 0.626, r 2 = 0.392, P = 0.001) and also with the reduction of the resting twitch amplitude (r = 0.490, r 2 = 0.240, P = 0.011) at the same time-point in recovery (see Fig. 5). Thus, those participants who had a greater reduction in MVIC also had a greater decrease in their level of voluntary activation and amplitude of their resting twitch immediately after the eccentric contractions. When the correlations were performed in men and women separately, reduction in MVIC in men was associated with decline in voluntary activation only (r = 0.691, r 2 = 0.477, P = 0.009). The reduction in MVIC in women was associated with both the loss of voluntary activation (r = 0.579, r 2 = 0.335, P = 0.038) as well as decline in the resting twitch (r = 0.709, r 2 = 0.503, P = 0.007). Despite the differences in correlations, the correlation coefficients between the reduction in MVIC and the loss of voluntary activation in men and women were not different when tested with the Fisher r-to-z transformation (P = 0.285).

Associations with reductions in MVIC. Reduction in MVIC immediately after the fatigue task was associated with the a reduction in RT (P < 0.05) and b reduction in VA immediately after the eccentric fatiguing task (P < 0.05) in men (filled circle) and women (open circle)
During the period immediately after the fatigue task and to 48-h recovery, the change in MVIC was associated with recovery of voluntary activation (r = 0.764, P < 0.001) and recovery of resting twitch (r = 0.714, P < 0.001) for both men and women over the same time-course.
The association between power reduction immediately measured after the eccentric contraction task and the reduction in resting twitch was r = 0.396 (P = 0.050).
Discussion
This study compared reductions of strength and power of the knee extensor muscles in young men and women after repeated maximal eccentric contractions performed at 60° s−1. The new findings from this study were that (1) both reductions in voluntary activation and contractile function (resting twitch torque) were associated with the reductions in MVIC torque immediately after the termination of the eccentric contraction exercise; (2) there were no sex-related differences in either muscle soreness, the reduction of maximal isometric strength, or recovery of peak power up to 48 h after repeated maximal eccentric contractions; and (3) the loss in MVIC torque at 48 h was primarily due to central mechanisms, because voluntary activation was reduced and the resting twitch amplitude had recovered to baseline levels. Thus, while DOMS persisted to 48 h after repeated eccentric contractions in the knee extensor muscles, the mechanisms for the reduction in maximal force were largely neural in origin. Finally, in a subgroup of nine men and women, maximal isometric strength and voluntary activation remained depressed 14 days after the eccentric contractions, although the resting twitch torque had fully recovered. Thus, functional recovery was not complete in men or women 14 days after exercise primarily due to neural mechanisms despite full recovery of muscle soreness.
Acute reductions is muscle function after eccentric exercise
Neither sex showed a reduction of power at the 1-N m condition immediately following the eccentric task. For the higher intensity contractions (10–60% MVIC), fatigue did occur and the magnitude of fatigue increased as the intensity of contraction increased, although the relative decline was similar in men and women at each intensity. Because power is the product of torque and the angular velocity, both groups experienced a similar decrease in angular velocity given the similar decrease in strength. These results are similar to the findings of the previous study (Power et al. 2013) and consistent with the changes in MVIC torque for both sexes as discussed below.
Immediately after the eccentric contractions, the reduction in MVIC torque of the knee extensor muscles (~39%) was large but also similar in the men and women. This loss of strength was consistent with the previous findings tested with the elbow flexors (Borsa and Sauers 2000; Hubal et al. 2008; Sayers et al. 1999; Rinard et al. 2000), dorsiflexors (Power et al. 2013), and knee extensors (Hicks et al. 2016). In contrast to our findings, there was a greater decrease in strength of the elbow flexor muscles in women compared with men (57.8 vs 50.4%, respectively) after eccentric exercise (Sewright et al. 2008), and men had a greater reduction in isometric strength than women after 240 maximal eccentric contractions with knee extensors (bilaterally) (Minahan et al. 2015).
In general, the lack of sex difference in the decline in torque and power immediately after the eccentric contractions are in contrast to the large sex difference fatigability observed after isometric and or slow-dynamic concentric fatiguing contractions. Typically, women are less fatigable for isometric and slow-dynamic shortening contractions (Avin et al. 2010; Wust et al. 2008; Hunter et al. 2004; Clark et al. 2005) (see Hunter 2016a, b for reviews), although isometric and concentric contractions do not typically elicit muscle soreness. Furthermore, there was no association between baseline strength and the decline in torque as has been shown for isometric and concentric fatiguing tasks in young adults (Pincivero et al. 2000; Hunter and Enoka 2001; Billaut and Bishop 2012; Hunter 2016a, b). These results indicate that immediately after exercise, fatigability assessed with maximal isometric strength was dependent on the fatiguing task and contraction type. After repeated eccentric contractions, the reduction in torque may be due to fatiguing processes that will recover within hours, or muscle damage that can last up to 72 h (Byrne et al. 2004). Determining whether the reduction in MVIC torque immediately post-eccentric exercise is due to fatigability or muscle damage, however, was not able to be distinguished in this study.
Consistent with the MVIC torque, there was no difference between men and women in the average reduction in the resting twitch (indicative of muscular mechanisms) and the decline in voluntary activation from baseline (neural mechanisms) immediately post-eccentric exercise. These findings are inconsistent with other studies that reported no change in voluntary activation following lengthening contractions of ankle dorsiflexors (Power et al. 2010, 2013), but is consistent with other studies (Doguet et al. 2016; Prasartwuth et al. 2005; Racinais et al. 2008). We also found that there were significant associations between the reduction in MVIC post-exercise with the decline in the resting twitch and the reduction in voluntary activation. However, when correlations with each sex were considered individually, the findings indicate some sex-specificity. The reduction in MVIC for men was only significantly associated with reduction in voluntary activation, whereas reduction in MVIC for women was associated with both reduction in voluntary activation as well as reduction in resting twitch. During lower limb fatiguing contractions, men have been more susceptible to reductions in maximal torque due to reduction in voluntary activation (Russ and Kent-Braun 2003; Martin and Rattey 2007). It is plausible that a slightly different contribution of mechanisms for men and women contributed to their reductions in MVIC torque post-eccentric contractions.
RMS EMG of knee extensor muscles during MVIC contraction decreased immediately following the eccentric task, and this was similar between men and women. These results contrast with the previous findings of other studies where there was no change in EMG activity following lengthening contractions of the ankle dorsiflexors (Power et al. 2010, 2013). The decline in RMS EMG during the maximal contraction may result from a decline in the drive from the central nervous system (Bigland-Ritchie et al. 1983), although there are many other factors that can influence the global EMG activity (Farina et al. 2004, 2014). The decreased RMS EMG was not recovered to baseline after 30 min of recovery in both men and women, which is consistent with the declines in voluntary activation. Collectively, our results support that neural mechanism contributed to decrements in torque immediately after the repeated eccentric contractions similarly in men and women.
Reductions is muscle function 24–48-h post-eccentric exercise
At 24 h after exercise, fatiguing processes are primarily recovered, so any changes in function post 24 h of eccentric contractions are likely due to muscle damage (Byrne et al. 2004). The recovery pattern for MVIC torque at the 24- and 48-h post-exercise was similar in men and women, which is consistent with the previous studies that compared the recovery up to a 10-day post-exercise (Hicks et al. 2016; Sewright et al. 2008; Rinard et al. 2000). Others, however, observed a faster recovery in men compared with women (Power et al. 2013) and in contrast a faster recovery in women than men (Minahan et al. 2015). The reason for the differing results between the recovery of men and women across studies is not clear but might be in part be attributed to the prior exercise conditioning of men and women and the involved muscle group.
Power recovered by 48 h at the lower resistance loads (≤10% MVIC), but had not recovered for the moderate and high resistance loads (≥20% MVIC). The intensity at which the absolute peak power occurred (greatest power output across all intensity conditions) also shifted to lower intensities. Furthermore, for both sexes, MVIC strength did not recover, but power at the high velocity, low intensity contractions did recover. These results suggest that after eccentric contractions, the force rather than velocity is more impaired when power production is required.
Both central and peripheral factors contributed to MVIC recovery for both sexes up to 24-h post-eccentric contractions, because the recovery pattern of resting twitch and voluntary activation was similar. However, the incomplete recovery of voluntary activation at 48-h post-exercise suggests that the impaired torque and power production were primarily due to central impairments in both sexes. These findings are inconsistent with other studies that have shown voluntary activation was well maintained after task and 48 recovery (Power et al. 2013) or recovered fully within 2–24 h after the eccentric task (Doguet et al. 2016; Prasartwuth et al. 2005). However, a previous study also found the incomplete recovery in voluntary activation up to 48 h after a-30-min downhill walking exercise (Racinais et al. 2008) and suggested that this long-lasting decrement in voluntary activation could be due to long-lasting muscle pain after eccentric exercise (Le Pera et al. 2001).
In general, there were minimal sex differences recovery pattern of power, although for the 60% MVIC load condition, there were sex differences between 30-min and 24-h recovery (indicated by a sex × time interaction) in that the men had a faster recovery than the women. However, when each time-point was assessed for relative sex differences using independent t tests, no sex differences in any time-points were found. In addition, because a similar pattern was not seen with the 50% MVIC condition, we concluded that this one data point did not provide enough evidence for sex differences in overall recovery pattern of power after eccentric exercise.
After the eccentric contractions, we expected to see a shift of optimal length (angle) to longer muscle lengths, indicating changes in muscle series compliance and muscle damage. However, there was no change in the optimal angle for torque production as measured by an isokinetic MVCC at 60° s−1, despite the reported muscle soreness. Interestingly, recent work (Coratella et al. 2015) found that after a lower limb fatigue task, the knee flexors showed a shift in optimal torque/angle relationship, whereas this did not occur for the knee extensor muscles. The knee extensors are often involved in eccentric contractions in sports and even in daily living (e.g., descending stairs), possibly explaining the lack of change in the optimal angle for force production that we observed.
Reductions is muscle function 14-day post-eccentric exercise
Fourteen days following 150 maximal eccentric contractions of the knee extensors, maximal strength (MVIC) had recovered similarly in the men and women, although both groups did not fully recover to the baseline levels. Recovery periods after repeated eccentric contractions can range from 24 h (Power et al. 2013) and up to 89 days (Sayers et al. 1999) before full recovery to baseline levels of function. The extent of recovery likely depends on the different protocols (50–240 contractions), muscle groups (elbow flexors, dorsiflexors, and knee extensors), and intra-individual variability (0–76% strength loss immediately after the task of this study) (Sewright et al. 2008). Our study, however, showed that the resting twitch amplitude for both sexes was fully recovered, but voluntary activation values were still depressed suggesting that the reduced strength at 14 days of recovery was due primarily to central mechanisms and independent of pain, because soreness had recovered.
One potential limitation of the study is that we used a single pulse of percutaneous stimulation to evoke force during the MVC to quantify voluntary activation. Thus, not all muscle fibers in the knee extensor muscles were stimulated as can be done with femoral nerve stimulation. In addition, it is suggested that a doublet rather than a single stimuli will provide greater sensitivity to the measure of voluntary activation in large muscles (Suter and Herzog 2001). Nevertheless, the amplitude of resting twitch was >20% of MVC force with no difference between men and women (25.5 and 26.4%, respectively), and these values are comparable to that achieved during supramaximal nerve stimulation. In addition, we did not quantify the maximal compound muscle action potential (M wave) and so we were not able to normalize the surface global EMG activity to account for any contribution to the EMG from potential changes in neuromuscular propagation.
In conclusion, there were minimal to no sex-related differences in fatigue and recovery following maximal eccentric fatiguing contractions of knee extensors. Neither men nor women had fully recovered their strength or power by 48 h of recovery and this was due to impairment of voluntary activation. Importantly, 14-day post-eccentric exercise, a subgroup of individuals still showed a 28% reduction in maximal strength primarily due to neural mechanisms. Considering the importance of eccentric exercise in training and rehabilitation, the functional recovery from eccentric contractions may require more time than initially thought.
Abbreviations
- ANOVA:
-
Analysis of variance
- EMG:
-
Electromyography
- MVCC:
-
Maximal voluntary concentric contraction
- MVIC:
-
Maximal voluntary isometric contraction
- RT:
-
Resting twitch
- RMS:
-
Root mean square
- VA:
-
Voluntary activation
References
Allen D (2001) Eccentric muscle damage: mechanisms of early reduction of force. Acta Physiol Scand 171(3):311–319
Avin KG, Naughton MR, Ford BW, Moore HE, Monitto-Webber MN, Stark AM, Gentile AJ, Law LA (2010) Sex differences in fatigue resistance are muscle group dependent. Med Sci Sports Exerc 42(10):1943–1950
Bampouras TM, Reeves ND, Baltzopoulos V, Maganaris CN (2006) Muscle activation assessment: effects of method, stimulus number, and joint angle. Muscle Nerve 34(6):740–746. doi:10.1002/mus.20610
Bigland-Ritchie B, Johansson R, Lippold OC, Woods JJ (1983) Contractile speed and EMG changes during fatigue of sustained maximal voluntary contractions. J Neurophysiol 50(1):313–324
Billaut F, Bishop DJ (2012) Mechanical work accounts for sex differences in fatigue during repeated sprints. Eur J Appl Physiol 112(4):1429–1436. doi:10.1007/s00421-011-2110-1
Borg GA (1982) Psychophysical bases of perceived exertion. Med Sci Sports Exerc 14(5):377–381
Borsa PA, Sauers EL (2000) The importance of gender on myokinetic deficits before and after microinjury. Med Sci Sports Exerc 32(5):891–896
Byrne C, Twist C, Eston R (2004) Neuromuscular function after exercise-induced muscle damage. Sports Med 34(1):49–69
Campenella B, Mattacola CG, Kimura IF (2000) Effect of visual feedback and verbal encouragement on concentric quadriceps and hamstrings peak torque of males and females. Isokinet Exerc Sci 8(1):1–6
Clark BC, Collier SR, Manini TM, Ploutz-Snyder LL (2005) Sex differences in muscle fatigability and activation patterns of the human quadriceps femoris. Eur J Appl Physiol 94(1–2):196–206
Coratella G, Bellin G, Beato M, Schena F (2015) Fatigue affects peak joint torque angle in hamstrings but not in quadriceps. J Sports Sci 33(12):1276–1282. doi:10.1080/02640414.2014.986185
De Ruiter C, Kooistra R, Paalman M, De Haan A (2004) Initial phase of maximal voluntary and electrically stimulated knee extension torque development at different knee angles. J Appl Physiol 97(5):1693–1701
Doguet V, Jubeau M, Dorel S, Couturier A, Lacourpaille L, Guevel A, Guilhem G (2016) Time-course of neuromuscular changes during and after maximal eccentric contractions. Front Physiol 7:137. doi:10.3389/fphys.2016.00137
Enoka RM, Duchateau J (2008) Muscle fatigue: what, why and how it influences muscle function. J Physiol 586(1):11–23
Farina D, Merletti R, Enoka RM (2004) The extraction of neural strategies from the surface EMG. J Appl Physiol 96:1486–1496
Farina D, Merletti R, Enoka RM (2014) The extraction of neural strategies from the surface EMG: an update. J Appl Physiol (1985) 117 (11):1215–1230. doi:10.1152/japplphysiol.00162.2014
Gandevia SC (2001) Spinal and supraspinal factors in human muscle fatigue. Physiol Rev 81(4):1725–1789
Hermens HJ, Freriks B, Disselhorst-Klug C, Rau G (2000) Development of recommendations for SEMG sensors and sensor placement procedures. J Electromyogr Kinesiol 10(5):361–374
Hicks KM, Onambele GL, Winwood K, Morse CI (2016) Muscle damage following maximal eccentric knee extensions in males and females. PLoS One 11(3):e0150848. doi:10.1371/journal.pone.0150848
Hubal MJ, Rubinstein SR, Clarkson PM (2008) Muscle function in men and women during maximal eccentric exercise. J Strength Cond Res Natl Strength Cond Assoc 22(4):1332–1338. doi:10.1519/JSC.0b013e31817392ec
Hunter SK (2014) Sex differences in human fatigability: mechanisms and insight to physiological responses. Acta Physiol (Oxf) 210(4):768–789. doi:10.1111/apha.12234
Hunter SK (2016a) The relevance of sex differences in performance fatigability. Med Sci Sports Exerc 48(11):2247–2256. doi:10.1249/mss.0000000000000928
Hunter SK (2016b) Sex differences in fatigability of dynamic contractions. Exp Physiol 101(2):250–255. doi:10.1113/ep085370
Hunter SK, Enoka RM (2001) Sex differences in the fatigability of arm muscles depends on absolute force during isometric contractions. J Appl Physiol 91(6):2686–2694
Hunter SK, Critchlow A, Shin IS, Enoka RM (2004) Fatigability of the elbow flexor muscles for a sustained submaximal contraction is similar in men and women matched for strength. J Appl Physiol 96:195–202
Kellis E, Baltzopoulos V (1996) Resistive eccentric exercise: effects of visual feedback on maximum moment of knee extensors and flexors. J Orthop Sports Phys Therapy 23(2):120–124
Kennedy DS, Fitzpatrick SC, Gandevia SC, Taylor JL (2015) Fatigue-related firing of muscle nociceptors reduces voluntary activation of ipsilateral but not contralateral lower limb muscles. J Appl Physiol (1985) 118 (4):408–418. doi:10.1152/japplphysiol.00375.2014
Kriska AM, Bennett PH (1992) An epidemiological perspective of the relationship between physical activity and NIDDM: from activity assessment to intervention. Diabetes Metab Rev 8(4):355–372
LaStayo PC, Woolf JM, Lewek MD, Snyder-Mackler L, Reich T, Lindstedt SL (2003) Eccentric muscle contractions: their contribution to injury, prevention, rehabilitation, and sport. J Orthop Sports Phys Ther 33(10):557–571. doi:10.2519/jospt.2003.33.10.557
LaStayo P, Marcus R, Dibble L, Frajacomo F, Lindstedt S (2014) Eccentric exercise in rehabilitation: safety, feasibility, and application. J Appl Physiol 116(11):1426–1434
Lau WY, Blazevich AJ, Newton MJ, Wu SSX, Nosaka K (2015) Assessment of muscle pain induced by elbow-flexor eccentric exercise. J Athl Train 50(11):1140–1148
Le Pera D, Graven-Nielsen T, Valeriani M, Oliviero A, Di Lazzaro V, Tonali PA, Arendt-Nielsen L (2001) Inhibition of motor system excitability at cortical and spinal level by tonic muscle pain. Clin Neurophysiol 112(9):1633–1641
Lindstedt SL, LaStayo PC, Reich TE (2001) When active muscles lengthen: properties and consequences of eccentric contractions. Physiology 16(6):256–261
Martin PG, Rattey J (2007) Central fatigue explains sex differences in muscle fatigue and contralateral cross-over effects of maximal contractions. Pflugers Arch 454(6):957–969
Martin V, Millet GY, Lattier G, Perrod L (2004) Effects of recovery modes after knee extensor muscles eccentric contractions. Med Sci Sports Exerc 36(11):1907–1915
Minahan C, Joyce S, Bulmer AC, Cronin N, Sabapathy S (2015) The influence of estradiol on muscle damage and leg strength after intense eccentric exercise. Eur J Appl Physiol 115(7):1493–1500. doi:10.1007/s00421-015-3133-9
Nishikawa K (2016) Eccentric contraction: unraveling mechanisms of force enhancement and energy conservation. J Exp Biol 219(Pt 2):189–196. doi:10.1242/jeb.124057
Oldfield RC (1971) The assessment and analysis of handedness: the Edinburgh Inventory. Neuropsychologia 9:97–113
Pietrosimone BG, Selkow NM, Ingersoll CD, Hart JM, Saliba SA (2011) Electrode type and placement configuration for quadriceps activation evaluation. J Athl Train 46(6):621
Pincivero DM, Gear WS, Sterner RL, Karunakara RG (2000) Gender differences in the relationship between quadriceps work and fatigue during high-intensity exercise. J Strength Cond Res 14(2):202–206
Power GA, Dalton BH, Rice CL, Vandervoort AA (2010) Delayed recovery of velocity-dependent power loss following eccentric actions of the ankle dorsiflexors. J Appl Physiol 109(3):669–676. doi:10.1152/japplphysiol.01254.2009
Power GA, Dalton BH, Rice CL, Vandervoort AA (2011) Reproducibility of velocity-dependent power: before and after lengthening contractions. Appl Physiol Nutr Metab 36(5):626–633
Power GA, Dalton BH, Rice CL, Vandervoort AA (2013) Peak power is reduced following lengthening contractions despite a maintenance of shortening velocity. Appl Physiol Nutr Metab 38 (12):1196–1205. doi:10.1139/apnm-2013-0092
Prasartwuth O, Taylor JL, Gandevia SC (2005) Maximal force, voluntary activation and muscle soreness after eccentric damage to human elbow flexor muscles. J Physiol 567(Pt 1):337–348. doi:10.1113/jphysiol.2005.087767
Racinais S, Bringard A, Puchaux K, Noakes TD, Perrey S (2008) Modulation in voluntary neural drive in relation to muscle soreness. Eur J Appl Physiol 102(4):439–446. doi:10.1007/s00421-007-0604-7
Rinard J, Clarkson PM, Smith LL, Grossman M (2000) Response of males and females to high-force eccentric exercise. J Sports Sci 18(4):229–236. doi:10.1080/026404100364965
Russ DW, Kent-Braun JA (2003) Sex differences in human skeletal muscle fatigue are eliminated under ischemic conditions. J Appl Physiol 94(6):2414–2422
Sayers SP, Clarkson PM (2001) Force recovery after eccentric exercise in males and females. Eur J Appl Physiol 84(1–2):122–126
Sayers SP, Clarkson PM, Rouzier PA, Kamen G (1999) Adverse events associated with eccentric exercise protocols: six case studies. Med Sci Sports Exerc 31(12):1697–1702
Sewright KA, Hubal MJ, Kearns A, Holbrook MT, Clarkson PM (2008) Sex differences in response to maximal eccentric exercise. Med Sci Sports Exerc 40(2):242–251
Suter E, Herzog W (2001) Effect of number of stimuli and timing of twitch application on variability in interpolated twitch torque. J Appl Physiol (1985) 90(3):1036–1040
Wust RC, Morse CI, de Haan A, Jones DA, Degens H (2008) Sex differences in contractile properties and fatigue resistance of human skeletal muscle. Exp Physiol 93(7):843–850
Yoon T, Schlinder Delap B, Griffith EE, Hunter SK (2007) Mechanisms of fatigue differ after low- and high-force fatiguing contractions in men and women. Muscle Nerve 36(4):515–524. doi:10.1002/mus.20844
Acknowledgements
Michigan Technological University start-up fund to TY. Thank you to Dr. Elmer for helping to review the manuscript and provide helpful suggestions.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Nicolas Place.
Rights and permissions
About this article

Cite this article
Lee, A., Baxter, J., Eischer, C. et al. Sex differences in neuromuscular function after repeated eccentric contractions of the knee extensor muscles. Eur J Appl Physiol 117, 1119–1130 (2017). https://doi.org/10.1007/s00421-017-3599-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-017-3599-8