
Abstract
The aims of the present study were to examine (1) endurance time and (2) activation pattern of vastus lateralis (VL), vastus medialis (VM) and rectus femoris (RF) muscles during fatiguing isometric knee extensions performed with different EMG biofeedbacks. Thirteen men (27 ± 5 year) volunteered to participate in three experimental sessions. Each session involved a submaximal isometric contraction held until failure at an EMG level corresponding to 40% maximal voluntary contraction torque (MVC), with visual EMG biofeedback provided for either (1) RF muscle (RF task), (2) VL and VM muscles (Vasti task) or (3) the sum of the VL, VM and RF muscles (Quadriceps task). EMG activity of VL, VM and RF muscles was recorded during each of the three tasks and further analyzed. Time to task failures and MVC loss (P < 0.001) after exercises were similar (P > 0.05) between the three sessions (182 s and ∼28%, respectively) (P > 0.05). Moreover, the magnitude of central and peripheral fatigue was not different at failure of the three tasks. Activation pattern was similar for knee extensors at the beginning of each task (P > 0.05). However, RF EMG activity decreased (P < 0.05) during the Vasti and the Quadriceps tasks (from ∼33 to ∼25% maximal EMG), whereas vasti EMG activity remained constant during the RF task (∼41% maximal EMG). These findings suggest that (1) task failure occurs when sustaining a submaximal level of EMG activity for as long as possible and (2) CNS is not able to differentiate descending drive to the different heads of the quadriceps at the beginning of a sustained contraction, despite a different activation pattern for the bi-articular RF muscle compared to the mono-articular vasti muscles during fatigue.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Sustaining a submaximal level of force for as long as possible leads ultimately to task failure. In the last 30 years, numerous studies have used electromyography combined to such tasks to monitor the change in voluntary drive to working muscles (e.g. Lloyd 1971; Bigland-Ritchie et al. 1986; Hunter and Enoka 2003a). The increase in electromyographic (EMG) activity classically reported is attributed to the recruitment of new unfatigued motor units concomitant with a rise in the firing rate (de Ruiter et al. 2004), even if it is not a consistent finding (Garland et al. 1994), to a higher degree of discharge synchronization between motor units (Kleine et al. 2001) and/or to an increased duration of motor unit action potential (Fallentin et al. 1994). Besides, it appears from recent works that the decrease in maximal voluntary contraction (MVC) at failure of a submaximal isometric contraction is quite similar (∼30–40%) independently of age (Bilodeau et al. 2001), gender (Hunter and Enoka 2001; Hunter et al. 2003b) or load type (Hunter et al. 2005; Maluf et al. 2005).
In a recent study, we reversed the task by providing subjects with feedback of the summed RMS EMG activity of vastus lateralis (VL), vastus medialis (VM) and rectus femoris (RF) muscles during performance of a fatiguing isometric task involving the knee extensor muscles (Place et al. 2006). Although the subjects were stopped by the investigators, we found a significant amount of muscle fatigue after the task, as revealed by a loss in MVC. Therefore we can reasonably imagine that sustaining a submaximal level of EMG for as long as possible will ultimately lead to failure to perform the task. The first purpose of the present study was to examine endurance time for three tasks requiring the maintenance of constant (1) RF muscle EMG (RF task), (2) VL and VM muscles EMG (Vasti task), or (3) the sum of RF, VL and VM muscles EMG (Quadriceps task). We hypothesized that task failure would occur when the maximal torque generating capacity attained a threshold value, and that this MVC reduction would be the same whatever the task. To delve deeper into the understanding of neuromuscular fatigue induced by the three tasks considered, we measured the amount of central versus peripheral component of fatigue by means of electrically-evoked contractions applied transcutaneously on the femoral nerve after the sustained contraction.
Activation strategy of the central nervous system (CNS), i.e. the distribution of voluntary drive among the different synergistic muscles performing a task, has been extensively studied and notably for the knee extensors. For instance, Housh et al. (1996) reported that RF muscle is not activated to the same degree as VL and VM muscles during cycling at submaximal levels of power output. Furthermore, de Ruiter et al. (2004) recently showed using near-infrared spectroscopy that RF muscle oxygen consumption was lower compared to vasti muscles at submaximal intensities, and thus that RF was recruited to a lesser extent than vasti muscles, despite similar oxidative capacity during MVC. Finally, in the last stages of a task where a submaximal level of force has to be sustained until failure, it appears that EMG activity of the bi-articular RF muscle is lower than that of mono-articular vasti muscles when expressed as a percentage of the maximal EMG measured during a MVC (e.g. Place et al. 2005; Clark et al. 2005). In addition to these considerations, RF muscle has been shown to be more susceptible to fatigue than vasti muscles during repeated contractions of the knee extensors (Shinohara et al. 1998; Akima et al. 2000). Nevertheless, none of the above cited tasks, where subjects were instructed to contract their whole quadriceps muscle in order to keep the constant force or power output, provide evidence of a different activation strategy among knee extensor muscles by the CNS.
The assessment of activation strategy appears possible by asking the subject to keep constant a level of muscle activity using EMG feedback of a single synergistic muscle while recording the EMG activity of other quadriceps synergists “free to vary”, i.e. not integrated in the EMG biofeedback. The second purpose of the present study was to examine activation pattern of the VL, VM and RF muscles during RF task, Vasti task and Quadriceps task previously described. We supposed that activation pattern of the bi-articular RF muscle would be similar during Vasti and Quadriceps tasks, as the RF muscle contributes to the hip joint integrity, even if not displayed in the biofeedback. Moreover, if CNS is able to differentiate the voluntary drive to knee extensors, we expected an economical activation and thus a low EMG activity of vasti muscles during the RF task. We also used transcutaneous electrical stimulation to examine if selective neuromuscular fatigue of the different heads of the quadriceps (quantified by changes in M-wave and maximal EMG activity relative to M-wave) would occur during these different tasks.
Methods
Subjects
Thirteen subjects (age: 27.2 ± 5.0 year; height: 177.5 ± 5.9 cm; mass: 72.5 ± 7.7 kg; means ± SD) volunteered to participate to the study. None of the subjects had any known neurological disorder. The procedures were conducted according to the Declaration of Helsinki. Prior to the study, each subject gave written consent and the University of Burgundy Committee on Human Research approved the study protocol.
Mechanical and electromyographic recordings
Mechanical measurements were recorded using a Biodex isokinetic dynamometer (Shirley, NY). The axis of the dynamometer was aligned with the knee extension axis, and the lever arm was attached to the shank with a strap. Extraneous movement of the upper body was limited by two crossover shoulder harnesses and a belt across the abdomen. To allow the recording of the antagonistic biceps femoris (BF) EMG, a board was placed underneath the subject with a hole where the electrodes were positioned so as to avoid any compression of the surface electrodes and wires. Experiments were performed on the right (dominant) leg with a knee joint angle of 75° of flexion (0°: knee fully extended) and a hip angle of 90°. Surface EMG activity of the VL, VM, RF muscles and the antagonistic BF muscle was recorded with pairs of silver chloride circular (recording diameter of 10 mm) surface electrodes (Controle Graphique Medical, Brie-Comte-Robert, France) positioned lengthwise over the middle of the muscle belly with an interelectrode (center-to-center) distance of 20 mm. These recording sites were determined in pilot testing by eliciting the greatest M-wave amplitude for each muscle via femoral nerve stimulation at the beginning of each test. This method enabled the comparison of surface EMG signals between the sessions (Place et al. 2005). The shape of the M-waves was also carefully examined; investigators ensured that M-wave for a given muscle was not affected by signals from nearby muscles to limit cross-talk. Low resistance between the two electrodes ( < 5 kΩ) was obtained by abrading the skin, and oil and dirt were removed from the skin using alcohol. The reference electrode was attached to the patella of the left knee. Myoelectrical signals were amplified with a bandwidth frequency ranging from 15 Hz to 5 kHz (common mode rejection ratio = 90 dB; impedance input = 100 MΩ; gain = 1,000), digitized on-line at a sampling frequency of 2 kHz using a computer (IPC 486) and stored for analysis with commercially available software (Tida, Heka elektronik, Lambrecht/Pfalz, Germany).
The EMG RMS biofeedback was provided by an integrated circuit, true RMS-to-DC converter (model AD536A, Analog Devices, USA; characteristics: maximal error for true RMS-to-DC conversion = 0.5%, bandwidth > 450 kHz). This circuit computed instantaneously the true RMS level of the amplified EMG signal for three channels separately with an integration time of 375 ms; in the present study, the EMG RMS of VL, VM and RF muscles could be calculated and summed by the circuit. RMS (expressed in mV) was calculated as follows:
where T = 0.375 s and t = time variable.
In the present study, the circuit displayed on a screen one of the following: (1) the EMG RMS of RF muscle, (2) the summed EMG RMS of VL and VM muscles or (3) the summed EMG RMS of RF, VL and VM muscles depending on the task the subject had to perform. The EMG RMS value obtained was displayed on a 15-in. monitor that was located 1 m in front of the subject.
Electrical stimulation
Transcutaneous electrically-evoked contractions were induced by using a high-voltage (maximal voltage 400 V) constant-current stimulator (model DS7, Digitimer, Hertfordshire, UK). The femoral nerve was stimulated using a monopolar cathode ball electrode (0.5 cm diameter) pressed into the femoral triangle by the experimenter. The site of stimulation was marked on the skin so that it could be repeated after the sustained contraction and between the two sessions. The anode was a 50 cm² (10 × 5 cm) rectangular electrode (Compex SA, Ecublens, Switzerland) located in the gluteal fold opposite the cathode. The optimal intensity of stimulation (i.e. that which recruited all knee extensor motor units) was considered to be reached when an increase in the stimulation intensity did not induce a further increase in the amplitude of the twitch force and the peak-to-peak amplitude of knee extensors compound muscle action potentials (M-wave, see Electrical recordings). The stimulus duration was 1 ms and the interval of the stimuli in the doublet was 10 ms. Once the optimal intensity was found, it was kept constant throughout the session for each subject.
Experimental procedures
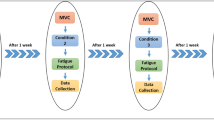
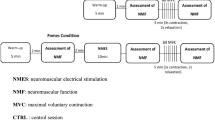
Subjects were required to participate in three testing sessions, each separated by at least 3 days. The measurements were similar but the fatiguing task differed between the three sessions. Subjects were asked to sustain a submaximal contraction until exhaustion with an EMG RMS feedback on (1) RF muscle (referred to as the RF task); (2) VL and VM muscles (referred to as the Vasti task) or (3) RF, VL and VM muscles (referred to as the Quadriceps task). The order of the fatiguing contractions was randomized for each subject. Before performing MVC, subjects performed a warm-up which included ∼10 brief and non-fatiguing submaximal contractions of the knee extensor and flexor muscles between ∼20 and 80% of the estimated MVC. The protocol was the same for the three tasks and involved (see Fig. 1 for a graphical overview):
-
1.
Three electrically evoked twitches separated by 3 s;
-
2.
Three paired stimuli (doublets) separated by 5 s;
-
3.
Two 5-s knee extensors MVC torque assessment (with a rest period of 60 s between trials) with doublet delivered over the isometric plateau (superimposed doublet) and 1.5 s after each MVC (potentiated doublet), to assess maximal voluntary activation level according to the interpolated twitch technique and potentiated peak doublet amplitude. A subsequent trial was performed if the difference in the peak force of the two MVCs were > 5%. The greatest level of torque achieved by the subjects before the fatiguing exercise was taken as the MVC torque and used for calculation of the submaximal target torque;
-
4.
One 5-s knee flexors MVC
-
5.
Randomized submaximal isometric contractions sustained for 5 s at target torques of 20, 40, and 60% MVC with the knee extensor muscles so as to evaluate the reliability of the EMG-force relation across sessions. These data were used to ensure that EMG behaviour during the fatiguing contraction represented physiological adjustments and were not due to differences in recording conditions. The EMG RMS level for the muscle(s) under investigation during the trial performed at 40% MVC was used as the target level during the fatiguing task;
-
6.
Performance of the RF, Vasti or Quadriceps task until failure (endurance time), i.e. the time at which the initial task could not be sustained any more for a 3 s period despite strong verbal encouragement by the investigators;
-
7.
Three electrically evoked twitches separated by 3 s;
-
8.
One knee extension MVC with doublet delivered 1.5 s before the MVC, over the isometric plateau and 1.5 s after the MVC. Only one unpotentiated doublet was delivered after fatigue to limit the recovery time between task failure and knee extensors MVC performance.

Graphical overview of the protocol for one session. The order and timing was the same for each subject and each of the three sessions
Data analysis
The three trials for mechanical parameters (single or paired stimuli) were averaged and further analyzed. The following variables were analyzed from the twitch response: peak twitch (Pt), time to peak twitch and half-relaxation time. Peak torque was also analyzed for the doublet. Postactivation potentiation was determined by computing the formula: [(amplitude of the potentiated doublet - amplitude of the nonpotentiated doublet) / amplitude of the nonpotentiated doublet × 100]. MVC was considered as the peak torque attained during the contraction. Maximal voluntary activation level was estimated according to the following formula, i.e. voluntary activation level = [1 − (superimposed doublet amplitude/potentiated doublet amplitude)] × 100 (Behm et al. 1996). In a few cases, in which the doublet was applied when the torque level was slightly below the real maximal voluntary torque, a correction was applied in the original equation, as suggested by Strojnik and Komi (1998). During the fatiguing contraction, the torque produced at the knee joint was analyzed at the time corresponding to 0, 1, 3, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90 and 100% of the endurance time for each of the three tasks.
Peak-to-peak amplitude, duration and RMS of the M-wave, calculated during the time interval corresponding to the area above baseline, i.e. 0 mV, were analyzed for VL, VM and RF muscles with the average of the three trials used for analysis. EMG of VL, VM, RF and BF muscles during the knee extensors MVC was quantified as the RMS for a 0.5 s interval at peak force (250 ms-interval either side of the peak force). Maximal RMS values for VL, VM and RF muscles were then normalized to the RMS of the M-wave for the respective muscles, in order to obtain RMS.M-1 ratio. This normalization procedure accounted for peripheral influences including neuromuscular propagation failure and changes in impedance from the EMG recordings. Maximal RMS of the BF muscle was quantified for the knee flexors MVC. BF RMS calculated during the knee extensors MVC was then normalized to that calculated when this muscle acted as an agonist, i.e. during the maximal knee flexion.
RMS activity was calculated for consecutive sampling intervals that were 10% of the endurance time for the VL, VM, RF, and BF muscles. The RMS of the EMG signal was normalized to that determined during the MVC performed before the fatiguing contraction. For the submaximal contractions at 20, 40 and 60% MVC, EMG signals were quantified as the RMS over a 0.5 s period after the torque had reached a plateau and were then normalized to that determined during the MVC performed before the fatiguing contraction.
Statistics
A one-factor (time) ANOVA with repeated measures was used to compare the endurance time and the initial target torque between the three tasks. Separate two-factor ANOVAs (time × task) with repeated measures on time and task were used to compare MVC torques, voluntary activation level, M-wave parameters for each muscle, twitch properties, peak doublet torque and postactivation potentiation. A three-factor ANOVA (task × intensity × muscle) with repeated measures on task was used to compare the RMS-force relation for the isometric contractions of the knee extensor muscles (VL, VM and RF) at four intensities of contraction (20, 40, 60 and 100% of MVC). A three-factor ANOVA (task × time × muscle) with repeated measures on time, task and muscle was used to compare the RMS for VL, VM and RF muscles at every 10% of the endurance time for the two tasks and the RMS.M−1 ratio for VL, VM and RF muscles before and after fatigue for each session. A two-factor ANOVA (task × time) with repeated measures on time and task was used to compare the antagonist BF muscle activity at every 10% of the endurance time. A two-factor ANOVA (task × time) with repeated measures was used to compare the torque at 0, 1, 3, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90 and 100% of the endurance time during the three fatiguing tasks. Post-hoc analyses (Tukey) were used to test for differences among pairs of means when appropriate. The statistical analyses were performed by using Statistica software for Windows (Statsoft, version 6.1, Statistica, Tulsa, OK). A significance level of P < 0.05 was used to identify statistical significance. Data are reported as means ± standard deviation (SD) within the text and table and displayed as means ± standard error (SE) in the figures.
Results
RMS-torque relation
The RMS activity for the knee extensor muscles was determined at the beginning of the three sessions with isometric contractions held at 20, 40, 60 and 100% MVC torque. RMS increased linearly with contraction intensity for all the knee extensor muscles (P < 0.05) and was similar across sessions (P > 0.05). There was no interaction for intensity and task (P > 0.05), indicating that the increase in RMS was consistent across the three tasks. There was no difference in the RMS activity of VL, VM and RF muscles at a given intensity, as indicated by the absence of interaction between muscle and intensity (P > 0.05). There was no interaction of muscle and task (P > 0.05), indicating that across sessions the increase in RMS was consistent for each of the knee extensor muscles.
Endurance time
Endurance times were not significantly different (P > 0.05) for the three conditions (166 ± 105 s, 173 ± 77 s, 208 ± 156 s for the RF, Vasti and Quadriceps tasks, respectively).
EMG activity and torque during the contractions
Vasti EMG activity remained constant during the three fatiguing tasks (see Fig. 2). There was no difference in EMG activity between the two mono-articular VL and VM muscles during any of the three conditions (P > 0.05). RF EMG activity was lower (P < 0.05) than vasti EMG activity from 60% of endurance time for RF task, from 40% of endurance time for Vasti task and from 30% of endurance time for Quadriceps task, respectively (Fig. 2). More interestingly, RF EMG activity decreased significantly from 60% of endurance time for the Vasti task (P < 0.001, Fig. 2b) and from 90% of endurance time for the Quadriceps task (P < 0.01, Fig. 2c). Antagonistic BF EMG activity remained constant (P > 0.05) during the three fatiguing contractions (on average from 3.2 ± 2.3 to 3.3 ± 2.3% MVC EMG, see Fig. 2).

Relative EMG RMS activity (% MVC EMG RMS) of the knee extensor (VL, VM and RF) and knee flexor (BF) muscles quantified at every 10% of the endurance time for the RF task (a), the Vasti task (b) and the Quadriceps task (c). RF EMG activity significantly lower than the first 10% value of the corresponding task: ** P < 0.01; *** P < 0.001. RF EMG activity significantly lower than VL and VM EMG activities at the same relative time: †† P < 0.01; ††† P < 0.001
Initial target torques were similar (P > 0.05) across the three contractions (41.1 ± 2.2 % MVC torque on average, Fig. 3). The decrease in exerted torque during the fatiguing contractions was significant (P < 0.001) from 10% (Vasti and Quadriceps tasks) or 15% (RF task) of endurance time. At exhaustion, final torque was greater for the RF task compared to the two other tasks (P < 0.005,Fig. 3).

Relative torque (% MVC torque) recorded during each of the three fatiguing tasks. Torque in the last 10% of endurance time of the RF task was significantly greater from torque in the last 10% endurance time of the Vasti and Quadriceps tasks: † P < 0.05
MVC and voluntary activation
MVC torque was similarly reduced after the three tasks (from 303 ± 48 to 215 ± 61 N.m for the RF task, from 304 ± 50 to 227 ± 55 N.m for the Vasti task and from 308 ± 48 to 219 ± 57 N.m for the Quadriceps task, P < 0.001; see Fig. 4a). Maximal voluntary activation level was significantly depressed (P < 0.01) to a similar extent (P > 0.05) after the three tasks (from 93.7 ± 4.8 to 87.2 ± 8.9 % on average, see Fig. 4b). Average RMS.M−1 losses of the RF, VL and VM muscles (P < 0.001) were not different for the three tasks (P > 0.05). No difference occurred in RMS.M−1 alterations between the three knee extensors for the three tasks (P > 0.05, Fig. 4c).

MVC torque loss (a), maximal voluntary activation level (b) and RMS.M−1 ratio for each knee extensor muscles (c) losses were significant after the fatiguing contractions: ** P < 0.01; *** P < 0.001. There was no difference between the three tasks. VL = vastus lateralis muscle, VM = vastus medialis muscle, RF = rectus femoris muscle
Muscle mechanical properties
Twitch properties remained relatively preserved after the fatiguing tasks (Table 1). The comparable (P > 0.05) decrease in time to peak twitch (−13 ± 16% on average, P < 0.01) in the three conditions was the only significant change in twitch properties observed in the present study. As for peak twitch, peak doublet remained unchanged at exhaustion of the fatiguing tasks (Table 1). In contrast, potentiated peak doublet (i.e. measured after MVC before and after fatiguing contractions) was significantly altered (on average −10.6 ± 11.8%, P < 0.05) with no difference between the three tasks (P > 0.05). Postactivation potentiation was similarly (P > 0.05) depressed following the three exercise bouts from 15 ± 11 to 9 ± 7% on average (P < 0.001, Table 1).
M-wave
M-wave properties (i.e. amplitude, duration and RMS) were preserved after each of the three tasks, as no significant change (P > 0.05) occurred after any of the fatiguing contractions. There was also no difference in M-wave properties between the three tasks (P > 0.05). For clarity reasons averaged M-waves (corresponding to the three knee extensors and the three experimental conditions) are given. M-wave amplitude, duration and RMS values before versus after fatigue were: 5.8 ± 2.9 mV versus 5.9 ± 2.9 mV, 7.5 ± 2.8 ms versus 7.9 ± 3.8 ms and 3.7 ± 1.5 mV versus 3.8 ± 1.5 mV, respectively.
Discussion
Present findings show that task failure occurs when maintaining a submaximal EMG biofeedback for as long as possible; indeed, we found comparable endurance time and MVC loss for the three tasks investigated. Besides, neuromuscular fatigue after the three different fatiguing tasks was similar among the superficial heads of the quadriceps. To our knowledge, this study is the first that has attempted to determine the relative activation of mono- versus bi-articular muscles during fatiguing isometric tasks by using EMG biofeedback on separate heads of the quadriceps. The results clearly showed that activation pattern was similar for VL, VM and RF at the beginning of each task, but we reported a different behaviour for bi-articular RF muscle compared to mono-articular vasti muscles during fatigue.
Failure of sustaining a submaximal level of EMG
As hypothesized, sustaining a submaximal level of EMG leads ultimately to task failure. It appears that failure involved both central (processes located above the neuromuscular junction) and peripheral components. In the present study, central fatigue was assessed by both an overall index (maximal voluntary activation level) and a more sensitive one (RMS.M−1 ratio) which allowed the investigation of the neural activation of individual knee extensor muscles. We found that both indexes (measured during maximal effort) indicate a similar alteration of neural drive whatever the condition.
By evoking maximal M-waves of the involved muscles, we aimed to assess neuromuscular propagation - transmission and/or sarcolemma excitability changes after the sustained contraction. We found no significant alteration in M-wave properties, suggesting that neuromuscular excitability within the periphery was not affected for any muscle by any of the fatiguing tasks. This finding confirms previous observations (Place et al. 2006) that fatigue of a prolonged task involving an EMG feedback did not originate from failure to drive action potentials to the muscle cell. The mechanical Pt response associated with the electrical M-wave and peak doublet amplitude did not change after fatigue, contrasting with the reduction in the more fatigue-sensitive potentiated peak doublet amplitude (Kufel et al. 2002; Place et al. 2006). Here potentiated amplitudes were used to prevent from potentiation that could occur during prolonged isometric contraction (Place et al. 2005). Indeed, the reduced time to peak twitch reported in the present study after each of the three tasks, which is generally observed for a potentiated twitch (Hamada et al. 2000), indicated that fatigue and potentiation effects likely coexisted in the twitch evoked immediately after the task (Rassier and Macintosh 2000). In addition, post-activation potentiation, which is an indicator of contractile protein sensitivity to Ca2+ (Sweeney et al. 1993), was depressed after the three tasks. Taken together, these observations suggest that peripheral impairment could be attributed to failure occurring beyond the membrane ionic process. Nevertheless, changes within the periphery (∼10%) were smaller than central impairment (a reduction from ∼15 to ∼35% for RMS.M−1 ratio depending on muscle), leading us to mainly attribute task failure to neural factors. The similar neural and contractile acute changes after each of the three tasks investigated here are consistent with the observed comparable endurance times and maximal torque generating capacity reductions.
Physiologically, the inability to maintain the target EMG level means that the subject is unable to (1) continue activating their fatigued motor units and/or (2) de-recruit the fatigued motor units, and replace their contribution to the electrical activity by other, unfatigued ones. Recently, Noakes (1997) and Noakes et al. (2001) propose a “central governor theory” during endurance events, which suggests that afferents from the heart inform the brain of any threat of hypoxia or ischemia that may develop in this organ. In response, the central governor acts via the motor cortex to reduce the efferent neural activation of the exercising muscles (Noakes et al. 2001). Our observation of similar MVC loss and central fatigue at termination of the three tasks enlarge this theory; here we assume that the inability to sustain the initial voluntary drive (leading directly to task failure) was necessary to preserve integrity of the fatigued motor units and more generally of the neuromuscular system. This hypothesis is consistent with the lower level of peripheral fatigue in comparison to central impairment at exhaustion and with the comparable endurance times across sessions. In addition, this novel suggestion can be generalized to prolonged submaximal contraction with torque feedback, where failure has been shown to occur before complete recruitment of available motor units in the active limb, an observation interpreted as central fatigue (Loscher et al. 1996). The comparable MVC reduction at the end of the three tasks investigated here and at exhaustion of many studies cited in the Introduction strengthens our idea that the reduction in voluntary drive prevents the neuromuscular system from complete exhaustion.
Activation pattern differs between mono- and bi-articular muscles but fatigue is the same between synergists
It was surprising that EMG activities of VL, VM and RF muscles were similar when starting the fatiguing tasks, whatever the biofeedback involved. This consistency of the descending drive led to comparable initial torque values across the three tasks and constitutes strong evidence of an undifferentiated descending drive from the CNS to the quadriceps synergists. We attributed this observation to the anatomical specificity of the knee extensors that have a common insertion onto the patella tendon, possibly limiting independent activation by the CNS. As the descending command seems to be directed to the whole knee extensors group and not specifically to each muscle, it is not surprising to observe a non-selective central fatigue among knee extensors at exhaustion, as assessed with the sensitive RMS.M−1 ratio.
Despite the undifferentiated command to knee extensor muscles at the beginning of the fatiguing contractions, we observed a reduction in RF EMG activity towards the latter stages of the Vasti task and Quadriceps task; it seems that fatigue induced a change in RF activation pattern during these two tasks. We attributed the comparable activation pattern for RF muscle across Vasti task and Quadriceps task to the anatomical function of the bi-articular RF muscle, which regulates the distribution of net moments about the hip joint (Van Ingen Schenau et al., 1992; Doorenbosch and van Ingen Schenau 1995) whatever the task. We can suppose that maintaining RF EMG activity led to earlier fatigue, as even during the Quadriceps task (where RF electrical activity is displayed in the feedback), activation of RF muscle is decreased and that of vasti muscles is slightly increased to keep the signal constant. Furthermore, it has been reported that RF is more fatigable (Shinohara et al. 1998; Akima et al. 2000) and much smaller than vasti muscles (Trappe et al. 2001), which can account for its depressed activation during fatigue. However, endurance time for the RF task, when RF EMG activity remained constant, was not shorter than for the two other tasks, suggesting that other mechanisms occurred.
We believe that subjects failed to perform the task because of a reduced muscle activation via inhibition from muscle afferents. Both Ia spindle afferents (Macefield et al. 1991) and small diameter afferents among group III and IV fibre populations (Garland 1991) have been reported to reflexively inhibit the α-motoneurons. Moreover, Gandevia (1998) supposed that a reduction in drive may be mediated by supraspinal effects of group III and IV afferents, probably via Renshaw cells (for a review see Windhorst 1996). As CNS is not able to regulate the descending drive specifically to the knee extensors (see above), we propose that the local environment of the active units deteriorated during fatigue, which then produced afferent activity that directly inhibited the motoneurons and/or affected the voluntary drive, progressively leading to task failure. We can speculate that the decrease in EMG activity observed for the bi-articular RF muscle during fatigue was due to its different anatomical function compared to vasti muscles, which led to a faster decline in RF motoneurons’ activity at the spinal level.
The absence of any change in vasti EMG activity during the RF task was unpredictable, as it serves apparently no useful purpose to maintain their initial electrical activity, vasti muscles crossing only the knee joint. This result could be interpreted as a waste of energy within the quadriceps muscle but is consistent with the unique descending drive to the knee extensors; it is also interesting to note that this apparent waste of energy did not induce a larger amount of neuromuscular fatigue compared to the other tasks. Besides, the finding of the greater torque production at the end of the RF task—the only task where RF EMG activity remains constant—in comparison to the two other tasks indicates that RF muscle significantly contributes to force production during a submaximal prolonged contraction in contrast to a recent suggestion (Kooistra et al. 2005), at least near the end of the contraction.
Finally, despite all the precautions taken with the placement of the recording electrodes, cross-talk might have occurred, especially with the RF muscle as recently shown by Byrne et al. (2005). This could constitute a potential limitation of the present results. However, as RF behaviour differs between the three tasks, we assume that cross-talk was limited. In addition, despite contradictory results (Giannesini et al. 2003; Todd et al. 2005), EMG signal could be affected by the accumulation of metabolites (Kent-Braun 1999) and/or a change in muscle temperature within the muscle (Petrofsky and Laymon 2005) that might occur during sustained isometric contraction. As endurance time was comparable across the three tasks, we suppose that the influence of temperature on EMG signal was the same whatever the task. Moreover, the effect of metabolites might be limited when sustaining a submaximal EMG level compared to a torque level (Place et al. 2006).
In conclusion, task failure for a submaximal isometric contraction with EMG feedback on the different heads of the quadriceps is attributed to the inability to sustain the initial descending drive and/or to spinal inhibitions via muscle afferents in order to prevent any impairment of the neuromuscular system, leading to comparable endurance time, similar amount of MVC reduction and central fatigue whatever the biofeedback involved. We also showed that muscle activation was similar among knee extensors at the beginning of the three tasks, suggesting that CNS is not able to differentiate descending drive to knee extensor muscles that have a common insertion to the patella tendon. However, fatigue-induced EMG adjustments for RF muscle during the Quadriceps and Vasti tasks occurred, but the mechanisms involved in the different activation patterns between mono- and bi-articular muscles requires further investigation.
References
Akima H, Kuno S, Takahashi H, Fukunaga T, Katsuta S (2000) The use of magnetic resonance images to investigate the influence of recruitment on the relationship between torque and cross-sectional area in human muscle. Eur J Appl Physiol 83:475–480
Behm DG, St-Pierre DM, Perez D (1996) Muscle inactivation: assessment of interpolated twitch technique. J Appl Physiol 81:2267–2273
Bigland-Ritchie B, Cafarelli E, Vollestad NK (1986) Fatigue of submaximal static contractions. Acta Physiol Scand Suppl 556:137–148
Bilodeau M, Henderson TK, Nolta BE, Pursley PJ, Sandfort GL (2001) Effect of aging on fatigue characteristics of elbow flexor muscles during sustained submaximal contraction. J Appl Physiol 91:2654–2664
Byrne CA, Lyons GM, Donnelly AE, O’Keeffe D T, Hermens H, Nene A (2005) Rectus femoris surface myoelectric signal cross-talk during static contractions. J Electromyogr Kinesiol 15:564–575
Clark BC, Collier SR, Manini TM, Ploutz-Snyder LL (2005) Sex differences in muscle fatigability and activation patterns of the human quadriceps femoris. Eur J Appl Physiol 94:196–206
Doorenbosch CAM, van Ingen Schenau G (1995) The role of mono- and bi-articular muscles during contact control leg tasks in man. Hum Mov Sci 14:279–300
Fallentin N, Jorgensen K, Simonsen EB (1993) Motor unit recruitment during prolonged isometric contractions. Eur J Appl Physiol Occup Physiol 67:335–341
Gandevia SC (1998) Neural control in human muscle fatigue: changes in muscle afferents, motoneurones and motor cortical drive [corrected]. Acta Physiol Scand 162:275–283
Garland SJ (1991) Role of small diameter afferents in reflex inhibition during human muscle fatigue. J Physiol 435:547–558
Garland SJ, Enoka RM, Serrano LP, Robinson GA (1994) Behavior of motor units in human biceps brachii during a submaximal fatiguing contraction. J Appl Physiol 76:2411–2419
Giannesini B, Cozzone PJ, Bendahan D (2003) Non-invasive investigations of muscular fatigue: metabolic and electromyographic components. Biochimie 85:873–883
Hamada T, Sale DG, Macdougall JD (2000) Postactivation potentiation in endurance-trained male athletes. Med Sci Sports Exerc 32:403–411
Housh TJ, deVries HA, Johnson GO, Evans SA, Housh DJ, Stout JR, Bradway RM, Evetovich TK (1996) Neuromuscular fatigue thresholds of the vastus lateralis, vastus medialis and rectus femoris muscles. Electromyogr Clin Neurophysiol 36:247–255
Hunter SK, Enoka RM (2001) Sex differences in the fatigability of arm muscles depends on absolute force during isometric contractions. J Appl Physiol 91:2686–2694
Hunter SK, Enoka RM (2003a) Changes in muscle activation can prolong the endurance time of a submaximal isometric contraction in humans. J Appl Physiol 94:108–118
Hunter SK, Lepers R, MacGillis CJ, Enoka RM (2003b) Activation among the elbow flexor muscles differs when maintaining arm position during a fatiguing contraction. J Appl Physiol 94:2439–2447
Hunter SK, Rochette L, Critchlow A, Enoka RM (2005) Time to task failure differs with load type when old adults perform a submaximal fatiguing contraction. Muscle Nerve 31:730–740
van Ingen Schenau GJ, Boots PJ, de Groot G, Snackers RJ, van Woensel WW (1992) The constrained control of force and position in multi-joint movements. Neuroscience 46:197–207
Kent-Braun JA (1999) Central and peripheral contributions to muscle fatigue in humans during sustained maximal effort. Eur J Appl Physiol Occup Physiol 80:57–63
Kleine BU, Stegeman DF, Mund D, Anders C (2001) Influence of motoneuron firing synchronization on SEMG characteristics in dependence of electrode position. J Appl Physiol 91:1588–1599
Kooistra RD, de Ruiter CJ, de Haan A (2005) Muscle activation and blood flow do not explain the muscle length-dependent variation in quadriceps isometric endurance. J Appl Physiol 98:810–816
Kufel TJ, Pineda LA, Mador MJ (2002) Comparison of potentiated and unpotentiated twitches as an index of muscle fatigue. Muscle Nerve 25:438–444
Lloyd AJ (1971) Surface electromyography during sustained isometric contractions. J Appl Physiol 30:713–719
Loscher WN, Cresswell AG, Thorstensson A (1996) Excitatory drive to the alpha-motoneuron pool during a fatiguing submaximal contraction in man. J Physiol 491(Pt 1):271–280
Macefield G, Hagbarth KE, Gorman R, Gandevia SC, Burke D (1991) Decline in spindle support to alpha-motoneurones during sustained voluntary contractions. J Physiol 440:497–512
Maluf KS, Shinohara M, Stephenson JL, Enoka RM (2005) Muscle activation and time to task failure differ with load type and contraction intensity for a human hand muscle. Exp Brain Res 167:165–177
Noakes TD (1997) 1996 J.B. Wolffe Memorial Lecture. Challenging beliefs: ex Africa semper aliquid novi. Med Sci Sports Exerc 29:571–590
Noakes TD, Peltonen JE, Rusko HK (2001) Evidence that a central governor regulates exercise performance during acute hypoxia and hyperoxia. J Exp Biol 204:3225–3234
Petrofsky J, Laymon M (2005) Muscle temperature and EMG amplitude and frequency during isometric exercise. Aviat Space Environ Med 76:1024–1030
Place N, Maffiuletti NA, Ballay Y, Lepers R (2005) Twitch potentiation is greater after a fatiguing submaximal isometric contraction performed at short vs. long quadriceps muscle length. J Appl Physiol 98:429–436
Place N, Martin A, Lepers R (2006) Changes in neuromuscular function after tasks involving control of EMG versus torque feedback of the same duration. Neurosci Lett 394:37–41
Rassier DE, Macintosh BR (2000) Coexistence of potentiation and fatigue in skeletal muscle. Braz J Med Biol Res 33:499–508
de Ruiter CJ, Elzinga MJ, Verdijk PW, van Mechelen W, de Haan A (2004) Voluntary drive-dependent changes in vastus lateralis motor unit firing rates during a sustained isometric contraction at 50% of maximum knee extension force. Pflugers Arch 447:436–444
Shinohara M, Kouzaki M, Yoshihisa T, Fukunaga T (1998) Mechanomyogram from the different heads of the quadriceps muscle during incremental knee extension. Eur J Appl Physiol Occup Physiol 78:289–295
Strojnik V, Komi PV (1998) Neuromuscular fatigue after maximal stretch-shortening cycle exercise. J Appl Physiol 84:344–350
Sweeney HL, Bowman BF, Stull JT (1993) Myosin light chain phosphorylation in vertebrate striated muscle: regulation and function. Am J Physiol 264:C1085–C1095
Todd G, Butler JE, Taylor JL, Gandevia SC (2005) Hyperthermia: a failure of the motor cortex and the muscle. J Physiol 563:621–631
Trappe TA, Lindquist DM, Carrithers JA (2001) Muscle-specific atrophy of the quadriceps femoris with aging. J Appl Physiol 90:2070–2074
Windhorst U (1996) On the role of recurrent inhibitory feedback in motor control. Prog Neurobiol 49:517–587
Acknowledgments
The authors thank Dr Sandra Hunter and Dr Julie Erskine for the helpful comments concerning the manuscript, as well as all the volunteers who participated to the study.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Place, N., Matkowski, B., Martin, A. et al. Synergists activation pattern of the quadriceps muscle differs when performing sustained isometric contractions with different EMG biofeedback. Exp Brain Res 174, 595–603 (2006). https://doi.org/10.1007/s00221-006-0504-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-006-0504-9