
Abstract
Cnidarians rely on the innate immune defenses based on self/non-self recognition, signaling and effector responses to kill pathogens and heal wounds. Like other invertebrates, the immune system of cnidarians can be classified into several functional components and many of these elements have now been described in various cnidarian model systems. These include recognition receptors, toll-like receptors and peptidoglycan binding proteins, prophenoloxidase and melanin synthesis for an impermeable melanin barrier, anti-microbial proteins and molecules, reactive oxygen-producing and scavenging systems and wound repair and cellular systems. This chapter will summarize the current state of knowledge of cnidarian immune pathways and mechanisms. We will summarize the available data, guiding the reader through the steps of initiating and executing an immune response: recognition, signaling, effector and repair mechanisms. We will also explore the current research of constitutive immunity observed in healthy organisms and elicited immune responses seen in naturally infected organisms or organisms exposed to live pathogens or pathogen associated molecular patterns. We will connect these known immune pathways with how they are expressed and regulated and how this may influence the wide variation in disease resistance observed in the field, both within and between species. The overarching goal of this chapter is to take the reader from a genomic to phenotypic perspective, while keeping pathways and mechanisms in a whole organism and ecological context, whenever possible.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
- Innate immunity
- Pathogen associated molecular pattern
- Antimicrobial peptide
- Coral disease
- Pathogen recognition receptor
1 Introduction
Coral diseases are important processes that affect coral reef structure and function (Ruiz-Moreno et al. 2012). Not only can coral mortality be directly due to disease, but the effects on coral physiology , such as reproduction, could have long term effects on coral populations. Therefore understanding how corals fight disease is more critical than ever. It is likely that immunity, and consequentially disease resistance, play large roles in the varying temporal, spatial and species-specific patterns of disease distribution (Pinzon et al. 2014a).
The study of cnidarian immunity has benefited from advances in cell biology and genomics, enabling researchers to discover pathways and processes and compare them among the Cnidaria and other phyla (Miller et al. 2007, 2011). Full genome and transcriptome sequencing and analyses, along with complementary DNA (cDNA) microarrays and quantitative real-time polymerase chain reaction (qPCR), have opened the doors for discovery of immune-related genes, especially in non-model organisms. At this time, there are certainly gaps in our knowledge regarding how and when these pathways are controlled and the downstream and phenotypic effects of many of these pathways. However, the aforementioned resources have primed the field of cnidarian immunology to better piece together immune pathways and begin placing them into the context of coral disease ecology.
As invertebrates , cnidarians have to rely on the innate arm of immunity (Mydlarz et al. 2006; Palmer and Traylor-Knowles 2012) and do not appear to have the ability to develop resistance to a particular pathogen. In essence, they must use the same arsenal to repeatedly fight pathogens and prevent infections. However, currently the classic dichotomy between innate/no memory and adaptive immunity is being challenged as evidence of immune memory is found in cnidarians (Netea et al. 2011). To date, two lines of evidence support the existence of memory in the cnidarian immune system . For one, tissue transplantation of conspecifics and rejection among heterospecifics has long been attributed to immunological memory (Rinkevich 2004). A caveat here is that the mechanisms behind allorecognition may differ from those of ‘training’ an immune system. The second line of evidence is the discovery of many classes of receptors that recognize molecular patterns of different pathogens (Netea et al. 2011). Recognizing the difference between pathogen types and responding with the appropriate binding receptor seems like the first step in acquiring memory to that pathogen. In fact, heightened immune responses to a second exposure of the same or different pathogen is seen in other invertebrate models (Netea et al. 2011).
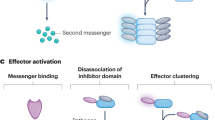
Cnidarian immunity is comprised of four categories of processes that enable the organism to recognize that it is under attack, kill the foreign perpetrator, and repair the wound. There are four major functions of the cnidarian immune system: (1) immune recognition, (2) intracellular signaling, (3) effector response and (4) tissue repair. The first step, recognition of a pathogen, leads to the activation of the immune system. This is accomplished by membrane bound receptors that bind directly to molecular patterns in pathogens (Rast and Messier-Solek 2008). For example, pattern recognition receptors, or PRRs, recognize pathogen /microbe-associated molecular patterns (PAMPs), such as lipopolysaccharides (LPS) from bacteria or β-glucans from fungi, and transmit the signal internally. The signal transduction mediators, signaling molecules and transcription factors that communicate the signal within the cell, comprise the intracellular signaling component of immunity (Fig. 28.1). The end-points of the signaling cascades are the proteins and molecules that kill the pathogen . These can comprise antibacterial proteins and small molecules, reactive oxygen and nitrogen molecules, and cellular activity and barriers. These effectors can be variable and novel among taxa (Rast and Messier-Solek 2008). Finally, the organism must seal off the infection and detoxify the tissue in order to heal the wound.

General scheme of signaling pathways (blue font) and related effector responses (green and red boxes) in cnidarians. Dashed line at Cell Activation indicates that not much is known about signaling controlling cell migration and phagocytosis in this group
There are several unique aspects of cnidarian and anthozoan physiology that influence immunity and health of the organism. Many cnidarians live with intracellular dinoflagellate symbionts from the genus Symbiodinium and extracellular microbial communities that reside in a host-derived mucus layer. This entire group of organisms (cnidarian host , algal symbionts and microbial associates ) is typically referred to as a holobiont (Rosenberg et al. 2007). This term has been used primarily in anthozoans , especially for reef-building scleractinian corals, but can be applied to many species of cnidarians and other taxa. Although the host is thought to be the primary contributor to cnidarian immune responses, there is mounting evidence for the important roles that the symbiont algal and microbial communities play. For example, the mechanisms used by the cnidarian host to establish and maintain these symbioses include many immune processes used to protect the host from pathogens . Thus, these components are of great interest when considering the immune system of cnidarians.
The health of the cnidarian holobiont relies on balance within these delicate symbioses and disruption of the symbiosis can lead to coral death . Breakdown of the symbiosis between the dinoflagellate symbiont and the host is referred to as bleaching. Bleaching can occur when the holobiont is stressed due to elevated temperature, excess ultraviolet radiation or pathogens (Baird et al. 2009). Additionally, shifts in the microbial communities of corals as a result of abiotic stressors (temperature, dissolved organic carbon, pH and nutrients) can lead to an increase in disease causing pathogens . Many of the cnidarian host responses to bleaching and shifting microbial communities can elicit immune responses, and we acknowledge the overlapping mechanisms throughout this chapter.
This chapter summarizes the current state of knowledge of cnidarian immune pathways and mechanisms by guiding the reader through the steps of initiating and executing an immune response as explained above. We draw examples from the presence of constitutive immunity observed in healthy organisms and elicited immune responses seen in natural infections or experimental exposures to live pathogens or PAMPs. We explore the functionality and expression/regulation of known immune pathways and their relation to wide variation, within and between species, in disease resistance observed in nature . The overarching goal of this chapter is to take the reader from a genomic to phenotypic perspective, while discussing pathways and mechanisms within a whole organism and ecological context, whenever possible.
2 Immune Recognition in Cnidarians
The activation of innate immunity in both vertebrates and invertebrates depends on pathogen recognition by various families of proteins known collectively as pattern recognition receptors, or PRRs. The term PRR is used to describe a diverse family of proteins that function in the recognition of potential pathogens by binding extracellular domains to proteins or other compounds characteristic of bacterial, fungal, protist, or parasitic pathogens (Akira et al. 2006). The cnidarian immune system is believed to include three main families of PRRs: Toll-like receptors (TLRs) , lectins , and nucleotide oligomerization domain (NOD )-like receptors (NLRs) (Miller et al. 2007; Schwarz et al. 2007, 2008b; Shinzato et al. 2011). Here we will discuss what is known of the presence and function of these three PRR groups across the Cnidaria as well as their expression patterns and importance to cnidarian immunity.
2.1 Toll-Like Receptors (TLRs)
Toll-like receptors (TLRs) are known to play an important role in the activation of innate immunity in many species (Aderem and Ulevitch 2000; Akira and Takeda 2004). These receptors are glycoproteins embedded within the host membrane which possess domains specific for binding to unique molecular patterns of microbes /pathogens , commonly referred to as pathogen-associated molecular patterns , or PAMPs, as the ligand.
There is an abundance of genomic data suggesting the presence of a wide repertoire of TLR type receptors in the Cnidaria. Binding of TLRs to PAMPs induces activation of both inflammatory and antimicrobial immune responses (Medzhitov 2001). Many TLR type proteins have been predicted from Nematostella vectensis genomic data (Miller et al. 2007). These include NvTLR-1, which is closely related to known Toll/TLR proteins , and three Toll/interleukin-1 (TIR) receptor proteins with immunoglobulin (Ig) domains, NvIL-1R1, NvIL-1R2, and NvIL-1R3. Finally, Nematostella vectensis also contains a MyD88 protein homolog, which functions in downstream transmission of TLR signaling (Miller et al. 2007).
In the gorgonian soft coral , Gorgonia ventalina , transcriptomic analysis contains annotations for three TLRs (TLR-2, -4, and -6; Burge et al. 2014). In scleractinian corals, several TLR type proteins were initially described from the genus Acropora. These acroporid TLRs are similar in sequence to NvIL-1R1, but they currently lack domain description (Miller et al. 2007). Subsequent genomic studies of Acropora digitifera have resulted in the discovery of many more putative TLR proteins (Shinzato et al. 2011) and the most recent studies have determined that up to 27 TIR proteins may be present in certain lineages of scleractinian corals (Poole and Weis 2014). Despite this knowledge, the exact functions of TLR proteins in corals and anemones are still not well understood. It can be assumed, based on sequence similarities, that cnidarian TLRs participate in recognition and activation of the immune system .
While components of TLRs have been detected in cnidarians, experimental evidence documenting their role in cnidarian immunity is still emerging. Two studies have documented conflicting patterns of TLR expression in cnidarians exposed to immune challenge (Burge et al. 2013; Libro et al. 2013). While the transcriptome of the sea fan Gorgonia ventalina includes annotations for three Toll-like receptors (TLR-2, -4, and -6), none of these were differentially expressed in response to immune challenge with the pathogen Aplanochytrium (Burge et al. 2013). In contrast, Acropora palmata displaying symptoms of White Band Disease show higher expression levels of two TLR-2 transcripts than do corals with signs of disease (Libro et al. 2013).
In contrast to Nematostella, Gorgonia ventalina , and acroporids, relatively few TLR proteins have been found in hydractinians. The Hydra magnipapillata genome includes two TIR domain-containing proteins, Toll-receptor-related (HyTRR)-1 and HyTRR-2 (Miller et al. 2007; Bosch et al. 2009) Both proteins contain an extracellular domain lacking characteristic leucine-rich repeats (LRRs). However, additional transcripts containing TLR-associated LRRs, HyLRR-1 and HyLRR- 2, have been discovered in the Hydra magnipapillata genome. HyLRR-2 is reactive to flagellin in the presence of human TIR domains, and is capable of modifying expression of HyTRR-1. Therefore it has been concluded that HyLRR-2 likely acts in tandem with HyTRR-1 to bind bacterial flagellin and induce innate immunity in Hydra magnipapillata (Bosch et al. 2009). TLRs in Hydra, particularly the combination of HyTRR-1 and HyLRR-2, are likely important in recognition of bacterial pathogens and activation of immunity.
Additional TIRs have been identified in a variety of cnidarians (Poole and Weis 2014). TIR domains have been identified in three anemone species , Anthopleura elegantissima, Aiptasia pallida (Lehnert et al. 2012), and Nematostella vectensis (Putnam et al. 2007), and six coral species, Acropora digitifera (Shinzato et al. 2011), Acropora millepora (Moya et al. 2012), Fungia scutaria (Poole and Weis 2014), Montastraea cavernosa (Poole and Weis 2014), Pocillopora damicornis (Traylor-Knowles et al. 2011), and Seriatopora hystrix (Poole and Weis 2014).
Even though TLR pathways are evolutionarily basal and present in most cnidarians, some lineages ( Hydra magnipapillata ) have since lost many major components of the pathways (Miller et al. 2007; Bosch et al. 2009). Other genomic studies have found the number of TLR and TIR proteins present in cnidarians to be lineage specific (Poole and Weis 2014); however, it is difficult to determine whether observed patterns are the result of gene expansion or gene loss (Miller et al. 2007; Bosch et al. 2009). Regardless of the mechanism of origin, it is believed that the wide variety of TLR proteins present in many coral genomes is important for the maintenance of the diverse microbial communities associated with corals (Poole and Weis 2014). Further experimental data will likely clarify the mechanisms driving TLR diversification between cnidarian lineages as well as the exact roles of Toll/TLR recognition in cnidarian immunity.
2.2 Lectins
Lectins are carbohydrate-binding proteins that act as important pattern recognition receptors (PRRs). Different classes of lectins have been described in cnidarians (Schwarz et al. 2007; Wood-Charlson and Weis 2009; Hayes et al. 2010; Kvennefors et al. 2010). Binding of lectins to potential pathogens is believed to trigger activation of the complement cascade and opsonization (Dunn 2009). Here we will focus on four main types of cnidarian lectins (tachylectin-2, millectin , rhamnospondin, and C-type lectins) and their roles in the activation of immune responses.
Tachylectin-2 was originally isolated from the Japanese horseshoe crab, Tachypleus tridentatus, and is capable of binding to N-acetylglucosamine and N-acetylgalactosamine (Beisel et al. 1999) and agglutinating bacteria during infections (Kawabata and Iwanaga 1999). Tachylectin-2 homologues have been well documented as PRRs in multiple lineages of octocorals, scleractinians, and hydractinians (DeSalvo et al. 2008; Hayes et al. 2010; Burge et al. 2013). Tachylectin-2 expression increased in Gorgonia ventalina following inoculation with the parasite Aplanochytrium (Burge et al. 2013). Furthermore, tachylectin-2 homologues have been identified in the scleractinian coral Orbicella (formerly Montastraea) faveolata (DeSalvo et al. 2008) and from the genus Oculina (Hayes et al. 2010). Interestingly, the Oculina homologue shows high amino acid diversity and a strong positive selection signal, indicating that it is not only part of the immune system, but is also undergoing rapid diversifying selection. This rapid selection indicates adaptive evolution, potentially as a result of rapidly changing microbial challenges (Hayes et al. 2010). Finally, Hydractinia echinata also contains a tachylectin-2 homologue gene named CTRN. However, no change in expression of CTRN was detected following exposure to LPS, suggesting that, although similar in sequence to immune functioning tachylectins, CTRN may not play a role in pathogen recognition (Mali et al. 2006).
Millectin , a mannose-binding lectin, has been well described in Acropora millepora as an important PRR and elicitor of immune response. Millectin is capable of binding and agglutinating a wide variety of Gram-positive and Gram-negative bacteria (Kvennefors et al. 2008). Millectin also shares a high degree of sequence similarly with other animal C-type lectins known to have immune functions. Due to these attributes, it is believed that millectin both recognizes and initiates opsonization and phagocytosis of potential pathogens (Kvennefors et al. 2010). Interestingly, the sensitivity of millectin depends upon the type of PAMP encountered. Millectin can exist in many different isoforms, enabling it to bind to a wide range of pathogens , thus increasing immune system efficiency (Kvennefors et al. 2008). Millectin is expressed in both nematocysts and the gastroderm, therefore coral pathogens are likely exposed to this PRR at some point (Kvennefors et al. 2010). Its diverse sensitivity, localization and expression, makes millectin a primary PRR and elicitor of immune response in Acropora millepora.
Several cnidarians express at least one rhamnose-binding lectin (RBL) gene, rhamnospondin, which contains multiple PRR-type domains (Lopez et al. 2011). The RBL domain of the rhamnose-binding lectin recognizes rhamnose, a component of the cell walls of some Gram-negative bacteria, and LPS in some Gram-positive bacteria. Expressed mainly in the mouth of Hydractinia symbiolongicarpus, rhamnospondin could prevent potential pathogenic bacteria from entering the organism through opsonization or agglutination, although there is no experimental evidence to support this (Schwarz et al. 2007). Since rhamnospondin and millectin are both highly expressed in areas that come into contact with the environment, they may serve homologous functions as the primary PRRs preventing infection in Hydractinia symbiolongicarpus and Acropora millepora , respectively.
In addition to the three main lectins described above, experimental and genomic studies have resulted in the discovery of a number of other potential C-type lectins (CTLs) in cnidarians. The Nematostella vectensis genome encodes a diversity of CTLs, many of which have extracellular domains suggesting their role in recognition and immune function (Reitzel et al. 2008; Wood-Charlson and Weis 2009). Furthermore, in Acropora palmata colonies infected with White Band Disease, three CTLs (C- type mannose receptor 2, macrophage lectin 2, and collectin-12) exhibited greater expression than in coral colonies with no signs of disease. These three CTLs can bind to a variety of bacterial, viral, and fungal compounds and likely trigger activation of innate immunity (Libro et al. 2013). While not well understood, it is likely that cnidarian CTLs function in recognition and activation of immune responses in a manner similar to the other lectins, tachylectin-2, millectin, and rhamnospondin.
Cnidarians possess a diverse and lineage-specific repertoire of lectins capable of recognizing pathogens and activating innate immunity. Localization of expression of some of these genes further suggests that lectins may serve as the first line of defense against pathogens in many cnidarians (Schwarz et al. 2007; Kvennefors et al. 2010). While lectins are essential to innate immunity in cnidarians, further research is necessary to understand the mechanistic basis of the involvement of these genes in activation of immunity.
2.3 NOD-Like Receptors
Nucleotide oligomerization domain (NOD) -like receptor (NLR) proteins are a type of PRR in vertebrates characterized by the presence of three domains: a NAIP, CIIA, HET-E and TP1 (NACHT) domain, one of three types of N-terminal effector domains, and a C-terminal leucine rich repeat (LRR) domain. While the genomes of several cnidarians have been found to contain NLR-like genes , the diversity of these genes is highly variable across the different lineages (Lange et al. 2011; Hamada et al. 2013). For example, very few identifiable NLRs were found in the Nematostella vectensis genome (Hamada et al. 2013). In contrast, the Acropora digitifera genome includes nearly 500 putative NLR loci that include NODs, one of the highest numbers among sequenced animal genomes. Furthermore, these putative NLRs have a wider diversity of protein domains compared to those seen in vertebrates (Hamada et al. 2013). In Acropora digitifera, the NACHT domain contains proteins with novel domain structures not seen in other organisms (Shinzato et al. 2011). The Acropora millepora genome also contains an unexpected number of NOD proteins, many of which have unique domain combinations (Lange et al. 2011).
Similar to Nematostella, Hydra appears to lack a wide diversity of NLR genes. Those NLRs that have been identified lack functional LRRs. Instead, NLRs in Hydra magnipapillata may function as a combination of two distinct proteins, the second of which contains a necessary LRR. A Hydra NLR protein has been shown to be capable of interacting with DD-Caspase, suggesting that it may be involved in the induction of apoptosis or downstream immune pathways (Lange et al. 2011). The lack of NLR diversity in Nematostella and Hydra, particularly in comparison to scleractinian corals, is still poorly understood. One hypothesis suggests that the diversity of NLRs in corals may serve as an adaptation to account for a lack of adaptive immunity ; however, that does not explain observed differences in diversity among cnidarian lineages, all of which lack adaptive immunity (Hamada et al. 2013). The need to maintain a diverse microbial community may have driven the expansion and subsequent diversity of NLRs (Hamada et al. 2013). Further genomic and experimental studies are necessary to understand the distribution of NLR diversity in cnidarians and to elucidate their function in immune activation .
2.4 Other PRR’s
A variety of immunoregulatory receptors that do not fall into one of the three major categories previously discussed have also been documented in cnidarians through a variety of molecular methods. These include LRR candidate PRRs (Schwarz et al. 2008b) and a transforming growth factor β (TGF-β) receptor (Detournay et al. 2012). Four LRR proteins, which may be important PRRs, were identified in expressed sequence tags (ESTs) from Orbicella faveolata and Acropora palmata . These may serve roles in recognition of and response to microbial infection (Schwarz et al. 2008b). Additionally, the presence of these receptors suggests that cnidarians possess a variety of other miscellaneous receptors, which contribute to recognition as well as immune activation and regulation.
3 Immune Signaling in the Cnidarians
Critical to the receptor binding detailed above is transmission of the signal and activation of the responses that will result in a successful immune response. These pathways are associated with the receptors and include many of the intracellular domains that propagate the signal though the cell. Some pathways are matched with specific receptors, whereas some are activated downstream from many potential sources. The TIRs introduced above, the mitogen-activated protein kinases (MAPK), the prophenoloxidase cascade and complement will be discussed in this section.
3.1 TLR Pathways and TIR Domains
Intracellular toll/interleukin-1 receptor (TIR) domains are found in several proteins associated with innate immunity signaling pathways. TIR domains are part of three classes of Toll receptor families: IL-1Rs, TLRs Myd88-dependent, and TLRs Myd88-independent (Poole and Weis 2014). These signaling pathways are most commonly initiated by cytokines or PAMPs. Upon activation, TIR proteins will employ downstream signaling and activate transcription factors. Transcription factors can then induce a variety of other pathways, including the melanin synthesis pathway and transforming growth factor beta (TGF-β) pathway (Medzhitov 2001).
As described above, several components of the Toll pathway have been identified in cnidarians. Four TIR domain-containing proteins were identified by Poole and Weis (2014), including TLR-like, IL-1R-like, Myd88 and TIR-only proteins. IL-1R-like and Myd88 proteins were identified in three anemone species , and in both Indo-Pacific and Caribbean scleractinian corals. However, no TIR domain-containing proteins have yet to be identified in gorgonians , although they are likely to exist (Miller et al. 2007). Myd88 proteins have also been identified in the freshwater hydrozoan, Hydra magnipapillata , in which they have been shown to be highly involved in bacterial recognition (Franzenburg et al. 2012).
One of the most commonly described transcription factors associated with Toll pathways in cnidarians has been the nuclear factor kappa light chain (NF-κB) transcription factor signaling pathway. NF-κB activation alters expression of target genes, including innate immune factors such as cytokines or antimicrobials (Hayden and Ghosh 2004). NF-κB gene homologs and proteins have been characterized in the anemone Nematostella vectensis (Wolenski et al. 2011) and identified in the scleractinian Acropora millepora and the hydrozoan Hydra magnipapillata (Miller et al. 2007). The role of NF-κB as an important immunity transcription factor has been demonstrated in Hydra, in which exposure to flagellin caused up-regulation of NF-κB (Bosch et al. 2009). However, functionality of the NF-κB transcription factor in relation to immunity has yet to be demonstrated in other cnidarian systems.
3.2 MAPK/ERK Pathway and GTPase Immune-Related Proteins
The multi-tiered kinase pathway consisting of mitogen-activated protein kinases (MAPK) and extracellular signal-regulated kinases (ERK) have also been identified in cnidarians as a component of TLR pathways (Miller et al. 2007). MAPK cascades direct many cellular responses and are likely involved in many of the immune pathways in cnidarians. To date, identified involvement and function of MAPK signaling have been limited to wound healing. In Nematostella vectensis, MAPK signaling-related genes were shown to be critical components of wound healing (Dubuc et al. 2014).
GTPases make up a large family of hydrolase enzymes that regulate a variety of cellular functions, including signaling and gene expression . GIMAPS, also known as immune-associated nucleotide-binding proteins, are a smaller family of GTPases. GIMAPs were once thought to only exist in vertebrates and play a role in T cell maturation (Filen and Lahesmaa 2010). Recently, however, several gene families of GIMAPs have been annotated in the transcriptome of the scleractinian coral Acropora millepora, where they are up-regulated when challenged with bacterial and viral PAMPs (Weiss et al. 2013). Unlike many of the other immune signaling pathways examined here, no GIMAP genes have been detected in either Nematostella vectensis or Hydra magnipapillata .
3.3 Prophenoloxidase Signaling Pathways
Melanin synthesis is a critical component of invertebrate immunity and is largely involved with pathogen encapsulation and wound healing. This cascade is initiated by a variety of PRRs, including TLRs and lectins , which subsequently trigger several proteolytic reactions that result in the synthesis of melanin. Upon activation, pro-phenoloxidase (PPO) enzymes are cleaved into phenoloxidases (PO) by trypsin-like serine proteases. POs then oxidize phenols in two steps: monophenols ➔ o-phenols ➔ o-quinones. Quinones are then polymerized to synthesize the final melanin product (Nappi and Ottaviani 2000).
Unlike many of the other signaling pathways, several components of the melanin synthesis cascade have been detected in cnidarians through both molecular and biochemical methods. Tyrosinase genes have been identified in the genomes of Nematostella vectensis and Hydra magnipapillata (Esposito et al. 2012) and a laccase-3 like gene was found in Pocillopora damicornis (Vidal-Dupiol et al. 2014). In addition, expression of a trypsin-like serine protease was highly up-regulated in Acropora millepora when challenged with bacterial and viral PAMPs (Weiss et al. 2013) and a gene homologous to a shrimp prophenoloxidase activating enzyme was also up-regulated in Pocillopora damicornis exposed to bacteria (Vidal-Dupiol et al. 2014). PPO/PO activity and deposits of melanin have been detected biochemically and/or histologically in many cnidarians, including sea anemones, gorgonian , alcyonacean and scleractinian corals, and hydrozoans (Palmer et al. 2008; Mydlarz et al. 2008; Mydlarz and Palmer 2011; Gimenez et al. 2014). Furthermore, Mydlarz and Palmer (2011) demonstrated that several classes of PO enzymes exist in scleractinian corals, including laccase, cresolase and catecholase, indicating the potential presence of different signaling cascades. In both gorgonian and scleractinian corals, higher PPO activity is found in infected versus healthy colonies (Mydlarz et al. 2008).
3.4 Complement Pathway
The complement pathway is a highly conserved component of innate immunity throughout metazoans . Activation of the pathway can occur through three different proteolytic pathways: classical, alternative, and lectin pathway. All pathways ultimately activate the complement protein (C3) complex to increase inflammatory responses such as phagocytosis, cell lysis, and coagulation. The lectin pathway has been most widely detected among cnidarians (Hemmrich et al. 2007). In this pathway, a lectin binds to a sugar on the pathogen, activating mannose-binding lectin associated serine proteases (MASPs), which then activate C2 and C4-like proteins to begin formation of the C3 complex (Endo et al. 2006). After the complex is formed, membrane-attack complex–perforin (MACPF) effector proteins are secreted to create a hole in the microbial membrane resulting in lysis of the pathogen.
In cnidarians, several components of the lectin complement pathway have been identified, including MASP, C3, and MACPF proteins. Mannose binding proteins have been found in the transcriptomes of Nematostella vectensis and two scleractinian corals, Acropora millepora and Orbicella faveolata (Brown et al. 2013; Pinzon et al. 2014a). Homologous genes for the C3 complex are found in the transcriptomes of numerous cnidarians, including the anemones Nematostella vectensis (Kimura et al. 2009) and Aptasia pallida (Sunagawa et al. 2009b), the octocorals Swiftia exserta and Gorgonia ventalina (Dishaw et al. 2005; Burge et al. 2013), and the scleractinian corals Acropora millepora (Miller et al. 2007) and Orbicella faveolata (Jorge et al. 2015).
Several additional complement effectors have been proposed and described among cnidarians. A gene matching human and abalone macrophage-expressed gene protein (MPEG-1) was identified in the Nematostella genome, but not in the Hydra genome (Miller et al. 2007). Although the function of MPEG in cnidarians has not been described, it is believed to represent an MACPF effector related to the complement system (Hemmrich et al. 2007). Toxic nematocyst venoms (TX-60A) from the anemones Actineria villosa and Phyllodiscus semoni belong to the MACPF protein family, form pores in target cell membranes and play roles in feeding and antipredator defense (Nagai et al. 2002; Oshiro et al. 2004), but could also kill potential pathogens . Two Nematostella vectensis MACPF proteins, one Acropora millepora transcript related to this protein type, and Hy-Mac from Hydra magnipapillata have been identified (Hemmrich et al. 2007; Miller et al. 2007). In Acropora millepora, expression of two complement pathway components, C3 and mannose-binding C-type lectin , were up-regulated when colonies were challenged with bacteria (Brown et al. 2013). Additional pore-forming toxins have been isolated from hydrozoans, scyphozoans and other anemones (Narat et al. 1994; Otero-Gonzalez et al. 2010), although antimicrobial functions have not been described. One other Hydra magnipapillata MACPF-domain protein, Hy-Apextrin, has an ortholog in Acropora millepora but not in Nematostella and is related to protein families in echinoderms and protozoans, although its function in Hydra remains unknown (Hemmrich et al. 2007; Miller et al. 2011).
4 Effector Responses in Cnidarians
A diversity of effector responses has been identified in cnidarians. These include a surface mucus layer that inhibits access of pathogens to cnidarian hosts , and antimicrobial activity due to antimicrobial peptides and secondary metabolites , reactive oxygen species (ROS) released as byproducts of various metabolic processes in both the host and symbiont, antioxidants that protect the host from those processes, and compounds produced as intermediates in various signaling pathways (e.g., during melanization activation). Melanin , the final product of this activation pathway, has antimicrobial activity, and provides a mechanism to encapsulate foreign cells and enhance structural support for impaired tissue. Foreign cells are also encapsulated, phagocytosed and expelled by cells that migrate to areas of tissue damage or infection.
4.1 Surface Mucus Layer
The first line of defense against invading pathogens in sedentary cnidarians, including anthozoans and hydrozoans, is the surface mucus layer (SML), a dynamic polysaccharide protein lipid complex covering the animal’s exterior (reviewed by Mullen et al. 2004; Brown and Bythell 2005). Mucous secretory cells are abundant in the epidermis and release mucus, which varies in composition among species. The mucus acts as a physical barrier, and the presence of cilia on the ectodermal cells aids in mucuciliary transport of particles, including foreign cells, towards the polyp mouth where they can be digested, or removed by sloughing of the mucus layer. In general, gorgonians have fewer mucous secretory cells than scleractinian corals, however, cilia on epidermal cells, mesenterial filaments, and the pharynx may also produce currents to remove wastes and potential foreign cells. Mucus production can vary in response to various stressors , including disease. Reneger et al. (2008) observed an increase in the size of mucocytes in Siderastrea siderea tissue affected by Dark Spot Syndrome.
The SML also serves as a nutrient source for diverse microorganisms, which can reach densities orders of magnitude higher in the mucus than in the surrounding seawater (Ritchie and Smith 1995; Krediet et al. 2009, 2013). The SML represents the natural coral surface microbiota , but may also include pathogenic species. Putative pathogens can take advantage of coral mucus as a nutritional resource and outcompete other commensals (Krediet et al. 2009). The surface microbial communities of healthy and diseased conspecifics vary considerably, representing not only changes in abundance of pathogens , but also in potential competitors and opportunistic commensals (Santavy 1995; Pantos et al. 2003; Gil-Agudelo et al. 2006; Bourne et al. 2008; Sunagawa et al. 2009a; Garcia et al. 2013; Meyer et al. 2014).
The ability of the cnidarian to control production and composition of the SML and its associated bacteria may serve as an important part of immunity. Antibacterial compounds produced by the cnidarian host , as well as its associated microbial consortium, are released into the mucus and play important roles in controlling the mucus-associated microbial community (Brown and Bythell 2005; Reshef et al. 2006; Ritchie 2006; Rosenberg et al. 2007; Shnit-Orland and Kushmaro 2009; Rypien et al. 2010; Krediet et al. 2013). In addition, the presence of quorum sensing molecules within the mucus also point to mechanisms by which the bacteria themselves can control the microbial community structure (Tait et al. 2010; Golberg et al. 2011). Quorum sensing molecules and activities have been detected in the polymicrobial consortium associated with Black Band Disease (Zimmer et al. 2014) and in gorgonian corals (Hunt et al. 2012).
4.2 Antimicrobial Activity
4.2.1 Antimicrobial Peptides
Antimicrobial peptides (AMPs) are a vital component of the innate immune system in both vertebrates and invertebrates , and several representatives have been identified in cnidarians (Zasloff 2002). AMPs are typically cationic membrane-bound peptides and have bactericidal activity against a diversity of microbes , largely due to their ability to disrupt microbial cell membranes. AMPs are often inducible and can be translocated where needed (Vidal-Dupiol et al. 2011).
The first AMP reported from a scleractinian coral is damicornin (Vidal-Dupiol et al. 2011), a compound of similar structure to the jellyfish AMP aurelin that exhibits antibacterial activity against Gram-positive and Gram-negative bacteria (Ovchinnikova et al. 2006).
Damicornin is constitutively transcribed in ectodermal granular cells in Pocillopora damicornis , where it is stored, probably as a precursor molecule subject to post-translational processing following immune challenge. In response to nonpathogenic Vibrio coralliilyticus (which is non-virulent at 25 °C), damicornin is released into the coral’s mucus . When exposed to virulent Vibrio coralliilyticus (at 28–32 °C), damicornin gene expression increased transiently at day 6 and was later repressed. Damicornin exhibited antimicrobial activity against Gram-positive bacteria and a fungus, although not against Gram-negative bacteria, including the coral pathogens Vibrio shiloi and Vibrio coralliilyticus (Vidal-Dupiol et al. 2011). More recently, mytimacin-like and LBP-BPI (lipopolysaccharide-binding protein and bactericidal/permeability-increasing protein) genes, which code for AMPs in other invertebrates (Krasity et al. 2011; Gerdol et al. 2012) were reported in Pocillopora damicornis (Vidal-Dupiol et al. 2014), and these were down-regulated in response to thermal stress and exposure to pathogens .
Three families of AMPs have been identified in Hydra by a combination of biochemical methods and by mining existing genomes (reviewed in Augustin et al. 2010; Bosch 2012, 2014). In Hydra spp., AMPs are sequestered into the epithelium and pathogen invasion causes increased expression of genes encoding for AMPs, which induces their production (Bosch et al. 2009; Augustin et al. 2010). Exposure to a culture filtrate of Pseudomonas aeruginosa, a potent immune inducer containing LPS and flagellin, induced expression of Hydramacin-1 (Bosch et al. 2009), which exhibits strong antibacterial activity against a diversity of human pathogens (Bosch et al. 2009; Jung et al. 2009). Hydramacin-1 promotes bacterial aggregation at low concentrations, representing the initial step of its bactericidal mechanism, which includes permeabilizing membranes of viable bacteria, although the bacteria die with intact cell membranes, suggesting that programmed cell death may be induced (Jung et al. 2009). However, Hydramacin-1 did not inhibit Pseudomonas aeruginosa, even though Hydra spp. tissue extract is active against this bacterium, suggesting that additional components of the Hydra tissue extract also possess antibacterial activity (Bosch et al. 2009).
The genomes of Hydra spp. contain genes for numerous arminins (Franzenburg et al. 2013). The first of these described was arminin 1a, an AMP with no sequence homology in other taxa (Augustin et al. 2009). Arminin 1a has strong broad-spectrum activity against human drug resistant pathogens , and kills bacteria by disrupting the bacterial cell wall without harming eukaryotic cells (Augustin et al. 2009). The expression of other arminims is species-specific (Franzenburg et al. 2013). Using arminin knockouts, Franzenburg et al. (2013) demonstrated that arminins function as species-specific AMPs, enabling Hydra spp. to control their associated bacterial communities.
The periculins represent a third group of AMPs in Hydra spp. with no known orthologs to other AMPs. Periculin-1 responds rapidly to bacterial flagellin and LPS, both of which cause a dose-dependent increase (Bosch et al. 2009). Periculin-1 may also respond to viral inducers (Bosch et al. 2009). Its strong bactericidal activity suggests that periculin is involved in Hydra spp. host defense (Bosch et al. 2009). Like the arminins, Perculin-1a also reduces overall bacterial load in embryonic Hydra vulgaris, and its strong selectivity against certain bacterial species highlights its importance in controlling bacterial community structure (Fraune et al. 2010). At least five members of the periculin family have been identified in Hydra spp. (Fraune et al. 2010).
4.2.2 Antimicrobial Secondary Metabolites
Antimicrobial secondary metabolites, also called “small molecules” or “natural products” are vital to the survival of sessile marine invertebrates . The need to protect surfaces from colonization by fouling organisms and pathogens is a critical role of these compounds (reviewed in Slattery and Gochfeld 2012). Although largely studied from a drug discovery perspective, recent studies have taken a more ecological approach to evaluating the roles of chemical extracts and, in some cases, purified compounds, within their host organisms or against ecologically relevant microbes . Unfortunately, little is known about the receptor mediated activation and signaling pathways associated with the biosynthesis of these compounds in cnidarians.
Whereas anemones produce highly toxic venoms that are released by their nematocysts and are important in feeding and providing protection from predators and competitors, these compounds probably play minimal roles in defense from microbial pathogens (Bosch 2008). However, toxic nematocyst venoms from the anemones Actineria villosa, Phyllodiscus semoni , and Stichodactyla helianthus include a class of compounds called the actinoporins, which form pores in target cell membranes and are suspected to be responsible for killing prey and deterring predators (Narat et al. 1994; Nagai et al. 2002; Oshiro et al. 2004; Otero-Gonzalez et al. 2010), but could also kill potential pathogens . Mucus and tissue extracts from anemones have shown only limited antimicrobial activity against microbes (Williams et al. 2007; Gunasundari et al. 2013), whereas mucus from the surface of Metridium senile stimulated the growth of 11 marine bacterial strains, suggesting that antibacterial compounds are not an effective defense against pathogenesis in this species (Astley and Ratcliffe 1989).
In addition to broad surveys of antimicrobial activity in gorgonians (reviewed in Slattery and Gochfeld 2012), some studies have targeted particular gorgonian pathogens . Nonpolar extracts of the sea fans Gorgonia ventalina , Gorgonia flabellum and 13 other Caribbean gorgonians inhibited spore germination of Aspergillus sydowii , the fungal pathogen of sea fans, at natural concentrations (Kim et al. 2000a, b; Dube et al. 2002). However, since Gorgonia spp. are the known hosts of Aspergillus sydowii, this suggests that other sources of immune function are necessary to inhibit pathogenesis. Extracts from Gorgonia marinae, which is less susceptible to aspergillosis than either of its conspecifics, had the strongest antifungal activity against Aspergillus sydowii (Mullen et al. 2006).
Because of their greater interest in the pharmaceutical arena, more effort has been directed towards identifying specific compounds associated with antimicrobial activity in the soft corals than in other cnidarians. Sinularia flexibilis produces the antibacterial diterpenes sinulariolide and flexibilide, which inhibit the growth of Gram-positive bacteria associated with human health (Aceret et al. 1998). The Antarctic soft coral Gersemia antarctica releases potent antimicrobial compounds into the surrounding seawater (Slattery et al. 1997). The most active of these compounds was identified as homarine, which is widespread in marine invertebrates , including cnidarians (e.g., gorgonians Leptogorgia virgulata, Leptogorgia setacea and Eunicella sinuglaris, hydrozoan Tubularia crocea, and scleractinian corals Montipora capitata and Porites spp.; Affeld et al. 2006; Targett et al. 1983; Maldonado et al. in review). In addition to Gersemia antarctica, homarine also exhibits antibacterial activity in Leptogorgia virgulata, Montipora capitata and Porites spp. (Shapo et al. 2007, Gochfeld unpublished data).
There is evidence that some of these antimicrobial metabolites are inducible during infections. Kim et al. (2000b) found antifungal activity to be higher in tissues closest to aspergillosis lesions and in diseased vs. healthy Gorgonia ventalina colonies. Antifungal activity also increased in Gorgonia ventalina following experimental inoculation with the fungal pathogen Aspergillus sydowii, providing strong evidence for an inducible defense (Dube et al. 2002). However, in broader geographic surveys, antifungal activity did not correlate with sea fan disease status or history, suggesting selection for resistance over time (Dube et al. 2002; Kim and Harvell 2004; Couch et al. 2008).
In addition, stimulation of immune effectors by disease can affect the production of other types of chemical defenses. For example, in the sea fan Gorgonia ventalina, the feeding deterrent compound julieannafuran decreased following fungal infection, resulting in increased levels of predation on affected fans (Slattery 1999). Likewise, colonies of Sinularia spp. affected by Sinularia Tissue Loss Disease were more palatable to butterflyfishes than healthy conspecifics (Slattery et al. 2013), suggesting a reduction in defenses in these corals.
Antimicrobial activity in scleractinians has received far less attention than in soft-bodied sessile taxa, largely due to the supposition that their hard calcareous skeletons precluded the need to synthesize chemical defenses. Clearly, that supposition undervalues the importance of immunity in scleractinian corals, and studies have demonstrated antimicrobial activity in scleractinian coral tissues, their mucus and in compounds released into the surrounding water. As in octocorals, broad surveys of extracts from scleractinian corals documented widespread but selective antibacterial activity (reviewed in Slattery and Gochfeld 2012). For example, aqueous extracts from 18 species of Caribbean corals (Pappas 2010; Gochfeld unpublished data) and 3 species of Hawaiian corals (Gochfeld and Aeby 2008) showed highly selective antibacterial activity against a panel of known coral pathogens , potential marine pathogens from human waste, and coral-associated bacteria . In contrast, Kelman (2009) found minimal antibacterial activity from nonpolar extracts of Red Sea corals, compared with widespread activity among the soft coral species tested. Some of these differences may reflect the overall greater antibacterial activity of aqueous extracts in the scleractinian corals, as compared to the more active nonpolar fractions in soft corals (Gochfeld unpublished data). Likewise, due to differences in extraction methods, antibacterial activity in protein extracts of corals may include activity due to AMPs (such as damicornin mentioned above), which would not necessarily be present in aqueous extracts. Protein extracts from tissues of the Caribbean corals Orbicella faveolata , Stephanocoenia intersepta and Porites astreoides also exhibit antibacterial activity (Mydlarz et al. 2009; Palmer et al. 2011b).
The isolation techniques and manipulation of the corals may have underestimated the antimicrobial compounds in scleractinian corals . For example, Geffen and Rosenberg (2004) described the rapid release of coral antibacterial compounds into the surrounding water following mechanical disturbance of several species of Red Sea corals. The release was fast and potent, killing Gram-negative and Gram-positive bacteria, including coral pathogens , within 1 min of release. In a follow-up study, antibacterial activity was released upon repeated disturbance , suggesting that it is constitutively present rather than inducible upon infection or damage (Geffen et al. 2009).
While antimicrobial activity now appears to be widespread in corals, there is significant intraspecific variability in activity due to a diversity of stressors , including disease, suggesting either that healthy corals remain healthy due to different constitutive levels of chemical defenses or that pathogenesis induces up- or down-regulation in the production of these defenses. Differences in antibacterial activity in extracts from colonies of Siderastrea siderea affected by Dark Spot Syndrome compared to healthy colonies may be related to differences in the concentrations of two uncharacterized compounds, suggesting the potential for induced antimicrobial defenses (Gochfeld et al. 2006; Pappas 2010). Extracts from healthy colonies of Montipora capitata exhibited greater antimicrobial activity against certain bacteria than did healthy tissues on colonies affected by Montipora White Syndrome, suggesting that greater constitutive levels of antibacterial defense in healthy corals may protect them from pathogens (Gochfeld and Aeby 2008). In addition, extracts from diseased tissues on Montipora White Syndrome-affected colonies exhibited higher levels of antibacterial activity against the coral pathogen Aurantimonas coralicida than did healthy tissues on those diseased colonies, indicative of a locally induced defense (Gochfeld and Aeby 2008). Protein extracts from Orbicella faveolata colonies affected by Yellow Band Disease had higher levels of antibacterial activity in both healthy and infected tissues of diseased colonies than in healthy colonies, suggesting a systemic response to infection (Mydlarz et al. 2009).
Surprisingly, in spite of their toxicity towards humans, little is known about antimicrobial activity in hydrozoans beyond their reported AMPs . Two species of fire coral, Millepora spp., from the Red Sea exhibited minor antibacterial and antifungal activity against human, animal and plant pathogens (Al-Lihaibi 2005).
4.2.3 Other Antimicrobial Mechanisms
Protease inhibitors counteract the impacts of proteases produced by many pathogenic microbes , which aid in colonization of host tissue and evasion of host immune defenses. By inhibiting these processes, serine protease inhibitors reduce pathogen growth or kill them outright (Augustin et al. 2010). Hydra magnipapillata produces a kazal-type serine protease inhibitor, Kazal-2, which has potent anti-staphyloccocal activity (Augustin et al. 2009). Gorgonia ventalina produces protease inhibitors that are active against proteases secreted by the fungal pathogen Aspergillus sydowii (Mann et al. 2014).
Lysozyme-like activity in sea fans is facilitated by a small lysozyme-like protein commonly grouped with AMPs due to its small size and ability to hydrolyze the peptidoglycans in bacterial cell walls (Couch et al. 2008). Lysozyme-like activity was greatest in colonies of Orbicella faveolata affected by Yellow Band Disease (Mydlarz et al. 2009). Mucus from the anemone Anthopleura elegantissima also contains a lysozyme-like enzyme that lyses bacteria (Phillips 1963, cited in Mullen et al. 2004).
Chitinases are enzymes that hydrolyze chitin, a major component of fungal cell walls. Exochitinase is found in healthy Gorgonia ventalina , but only a transient increase in exochitinase activity was observed in response to fungal infection, suggesting that it serves primarily as a constitutive defense (Douglas et al. 2006). Exochitinase was released into the water surrounding the sea fan and did exhibit limited antifungal activity against Aspergillus sydowii (Douglas et al. 2006). Chitinases have also been reported from three species of hydrozoans, Hydra attenuata, Hydra circumcincta and Podocoryne carnea (Klug et al. 1984). The first chitinase cDNA from a cnidarian was cloned from Hydractinia echinata by Mali et al. (2004), and in-situ hybridization indicated localized expression of this gene in the ectoderm and hyperplastic stolons, which are induced during allorecognition of histoincompatible colonies, suggesting a potential function in immunity .
4.3 Cellular Responses
Maintenance of an intact epithelium layer is essential to protection from pathogens and other stressors . In contrast to the hydrozoans, anthozoans have circulating populations of amoebocytes that can migrate to sites of injury and phagocytose foreign cells resulting in exposure to proteolytic enzymes and free radicals (Mullen et al. 2004). Many cnidarians can mount an inflammation response, which consists of an attempt to destroy or isolate the source of damage and any affected host cells. This typically involves rapid infiltration of injured tissue by immune cells and initiation of phagocytosis. Phagocytic cells can engulf and destroy pathogens or other foreign cells, often with the aid of ROS or antimicrobial compounds produced by the phagocytes, or encapsulate foreign cells and thereby provide a barrier between affected and healthy tissue (Palmer et al. 2008). In cnidarians, phagocytosis is also a primary means of acquiring nutrition and of maintaining host-symbiont relationships (Mullen et al. 2004).
4.3.1 Mobile Cytotoxic Cells and Reactive Oxygen and Nitrogen
In the anemone Actinia equina , amoebocytes are stored in the mesenterial filaments. Whereas only hyaline amoebocytes were observed to phagocytose and kill Gram-negative bacteria, mixed cultures of hyaline and granular amoebocytes also produced antibacterial activity upon stimulation by LPS, likely via the production of superoxide anions and other ROS through an oxidative burst (Hutton and Smith 1996).
In gorgonians , immediately following injury, a significant increase in amoebocytes was observed in the immediate vicinity (<1 mm) of the damaged tissue, with a decline in amoebocytes in adjacent tissues, suggesting that amoebocytes migrate from adjacent tissues to the site of repair (Meszaros and Bigger 1999). Following tissue damage in Swiftia exserta, granular cells staining positive for peroxidase were associated with phagocytosis of foreign particles, but the population of phagocytic cells was not uniform, indicating that multiple cell types may be involved in this process (Olano and Bigger 2000).
Amoebocytes also play critical roles in the cellular response to disease in gorgonians . In the sea fan Gorgonia ventalina , an inflammatory response is characterized by massive increases in granular acidophilic amoebocytes. Amoebocyte aggregation was associated with both naturally occurring infections and in the immediate area of experimental inoculations with the fungal pathogen Aspergillus sydowii (Mydlarz et al. 2008; Couch et al. 2013). However, there was no evidence that amoebocyte migration from immediately adjacent tissue was responsible for this increase (Mydlarz et al. 2008). Fungal hyphae infiltrate the gorgonin axial skeleton, where host-produced melanin is deposited, and melanosomes were detected in amoebocytes adjacent to these melanin bands in infected sea fans, pointing to granular amoebocytes as the sites of melanin synthesis. Since amoebocytes are responsible for phagocytosis and melanin production, they likely play a role in the antifungal activity observed in or adjacent to fungal lesions (Mydlarz et al. 2008).
In colonies of the hybrid soft coral Sinularia maxima x polydactyla affected by Sinularia Tissue Loss Disease, diseased tissue was also characterized by a proliferation of amoebocytes , which were frequently observed to envelop unidentified foreign eukaryotic cells (Slattery et al. 2013). Although not confirmed, these foreign cells may represent the disease agent, as all of these cells were engulfed by amoebocytes. Mesogleal amoebocyte area was also significantly increased in diseased tissue, although this appeared to be a localized response, as healthy tissue on diseased colonies did not differ in amoebocyte area from healthy colonies (Slattery et al. 2013).
In scleractinian corals, amoebocytes containing acidophilic granules have been observed in Montastraea cavernosa (Vargas-Angel et al. 2007), in which a rapid migration of granular amoebocytes was observed during tissue repair (Renegar et al. 2008). Chromophore cells infiltrating the calicoblastic epithelium in skeletal tissue anomalies of Porites compressa (Domart-Coulon et al. 2006) and melanin-containing granular cells in the epidermis or gastrodermis of several species of Indo-Pacific corals (Palmer et al. 2008, 2010) likely also represent amoebocytes .
An important role of mobile amoebocyte or phagocytic cells is the release of reactive oxygen and nitrogen species to aid in pathogen killing. Hydrogen peroxide as part of an oxidative burst has been identified in several gorgonian corals (Mydlarz et al. 2006), or as a constitutive release into the surrounding water (Shaked and Armoza-Zvuloni 2013; Armoza-Zvuloni and Shaked 2014). In a process called ETosis, chromatin is expelled from cell nuclei in response to an oxidative burst, LPS, or bacteria (Robb et al. 2014). To date, this has been studied in only a few invertebrates , but phagocytes in the mesoglea of the anemone Actinia equina produce ROS via a respiratory burst, which releases chromatin that may combine with the mucus to trap microbes , where they may come into contact with antimicrobial compounds or ROS that destroy them (Robb et al. 2014).
Nitric oxide synthase (NOS) has been measured in the sea anemone Aiptasia pallida, the scleractinian coral Acropora millepora and the soft coral Lobophytum pauciflorum, indicating that it is widespread among cnidarians (Safavi-Hemami et al. 2010). Nitric oxide (NO) was induced in response to LPS in Aiptasia pallida (Perez and Weis 2006), suggesting a potential role in response to pathogens .
Other cellular responses to infection have been observed in scleractinian corals. For example, zymogen granules were concentrated in the calicodermis in response to endolithic fungi associated with Dark Spot Syndrome in Siderastrea siderea , and mucocyte size increased in Dark Spot Syndrome-affected tissues (Renegar et al. 2008), suggesting the need for greater mucus production, a common sign of stress in corals. Hydra spp. do not have mobile phagocytes, and thus the epithelium serves as the sole center of the innate immune system , employing complex signaling pathways and defensive molecules, including AMPs (Bosch et al. 2009). However, upon exposure to culture filtrates of Pseudomonas aeruginosa, ectodermal epithelial cells of Hydra magnipapillata changed morphology and acquired a large number of granules, and endodermal epithelial cells were observed to engulf bacteria (Bosch et al. 2009).
4.3.2 Encapsulation
Scleractinians and gorgonians can also encapsulate invading microbes by increasing deposition of skeleton and proteinaceous organic material, such as collagen or gorgonin, to form a nodule surrounding the foreign organism (Mullen et al. 2004). Gorgonians may separate the host from the parasite by formation of a proteinaceous capsule (Goldberg et al. 1984). Gorgonia ventalina secretes gorgonin to surround invading algal filaments (Morse et al. 1977) and an unidentified parasite (Mullen et al. 2004). Scleractinians may exhibit accelerated growth in response to the presence of foreign organisms, such as epibionts or parasites (Aeby 2003; D’Angelo et al. 2012), which may encapsulate and attempt to minimize their damage to host tissues.
An increase in purple-pigmented calcium carbonate sclerites in gorgonians occurs in response to biotic stressors , including fungal infection (Alker et al. 2004; Mydlarz and Harvell 2007), and in response to predation (Puglisi et al. 2002). The purple pigment appears to be due to a carotenoid, some of which have antioxidant or antifungal activity (Leverette et al. 2008), but otherwise the function is unknown.
Although less studied than in corals, the melanin-synthesizing pathway has been found to occur in several species of anemones. The Argentinian anemones Aulactinia marplatensis and Bunodosoma zamponii exhibited PO activity in all tissues, and the high constitutive levels in B. zamponii were considered likely to provide resistance against disease (Gimenez et al. 2014). Upon exposure to the polymicrobial consortium associated with Caribbean Yellow Band Disease in corals, PPO activity in Aiptasia pallida polyps was up-regulated 8.5-fold over healthy polyps (Zaragoza et al. 2014). To corroborate the end product of this pathway in anemones, melanin granules were observed in Aiptasia pallida following infection with Serratia marcescens (Zaragoza et al. 2014).
In Gorgonia ventalina , melanin is deposited as a localized band along the axial skeleton in areas adjacent to fungal hyphae, and acts both to strengthen the skeleton and to encapsulate the fungal hyphae so they are unable to penetrate surrounding tissue (Petes et al. 2003; Mullen et al. 2004; Mydlarz et al. 2008). An increase in PPO activity was observed in conjunction with this increase in melanin deposition and granular amoebocytes in fungal-infected tissues (Mydlarz et al. 2008; Couch et al. 2013). Melanin deposition in Gorgonia ventalina was also observed in response to an unidentified parasite (Mullen et al. 2004) and to aggregations of Labyrinthulomycetes (Burge et al. 2012). PO, PPO and melanin were found constitutively in colonies of the soft corals Sarcophyton sp. and Lobophyton sp., with Sinularia sp. having the highest level of enzyme activity (Palmer et al. 2010, 2012).
The melanin synthesis pathway is also active in scleractinian corals. Interspecific constitutive differences in immune responses may play a role in relative susceptibility to disease. For example, Acropora millepora had constitutive levels of PO that were half those of Porites sp., with no melanin deposition, whereas Porites sp. had higher constitutive levels of PO and an inducible melanin response. These factors may contribute to the greater susceptibility of Acropora spp. than Porites spp. to diseases (Palmer et al. 2008). A more extensive survey of scleractinian coral species from the Great Barrier Reef found that all contained PO, PPO and melanin-containing granular cells, and that these constitutive indicators of immunity, along with fluorescent protein concentration, were good predictors of relative susceptibility to bleaching and disease (Palmer et al. 2010, 2012). Likewise, Porites astreoides had significantly higher PPO activity and melanin concentration than Orbicella faveolata (Palmer et al. 2011b). Melanin concentration was significantly correlated with phylogeny at the species level in Caribbean corals and was higher in corals that experienced less disease pressure (Pinzon et al. 2014a).
Palmer et al. (2008) found up-regulation of PO activity in abnormally pigmented tissues of Acropora millepora , although no melanin deposition was observed, confirming the inducibility of PO activity in corals, but also indicating that PO activity does not necessarily lead to melanin deposition. In contrast, a fourfold increase in the density of granular cells containing melanin was observed in pigmented regions of Porites sp. colonies and assumed to be involved in encapsulation (Palmer et al. 2008). Melanin-containing granular cells are also present in the epidermis of pigmented tissues in Porites trematodiasis (Palmer et al. 2009b). These studies confirmed that melanin represents the biochemical content of the granular cells observed in anthozoan cellular responses (i.e., amoebocytes) to invasion. Since abnormally pigmented tissues are commonly observed in corals and are due to a diversity of causes, it appears that melanization is a generalized defensive response to localized stress.
Exposure to commercial LPS, in combination with elevated seawater temperature, induced a significant increase in PPO activity, but not melanin concentration, in Stephanoecoenia intersepta and Porites astreoides, although not in Orbicella faveolata (Palmer et al. 2011b). PO activity was significantly reduced in colonies of Acropora millepora affected by White Syndrome relative to healthy colonies, although PO activity in diseased colonies was induced at the border of the lesion (Palmer et al. 2011a). Porites compressa affected by Porites Bleaching and Tissue Loss Disease also showed a decline in melanin containing granular cell densities in affected tissues (Sudek et al. 2012).
5 Repair Mechanisms in Cnidarians
Pathogenic infections and physical injuries trigger cellular and tissue repair mechanisms in all organisms. These responses are critical for individual survival and species fitness (Reitzel et al. 2008). In cnidarians, responses to stressors and repair mechanisms are complex and dynamic (Olano and Bigger 2000), integrating several cellular components and events (Reitzel et al. 2008). Among the best characterized repair mechanisms are antioxidants , apoptosis , and wound healing (Fig. 28.2). These mechanisms are an essential part of an efficient immune response in Cnidaria (Wenger et al. 2014).

Schematic representation of the steps involved in tissue repair and wound healing after a pathogen attack and lesion
5.1 Antioxidants
Continuous production of exposure to oxygen radicals can result in oxidative stress (Tarrant et al. 2014), an imbalance between ROS production and antioxidant defenses (Sies 2000; Betteridge 2000; Lesser 2006). Common sources of ROS in the immune response include respiratory burst and other disruptions of cellular and physiological processes (Betteridge 2000; Landis and Tower 2005; Tarrant et al. 2014). High levels of ROS affect cellular integrity, lead to tissue damage, diminish whole organism defensive responses and/or cause death (Betteridge 2000; Palmer et al. 2009a; Tarrant et al. 2014). To limit ROS levels and prevent oxidative stress, the cell uses various enzymatic and non-enzymatic antioxidants (Halliwell 1994; Hawkridge et al. 2000; Kim et al. 2011; Tarrant et al. 2014). The enzymes superoxide dismutase (SOD) , catalase and peroxidase play important roles in controlling free radical levels and preventing oxidative stress. SOD catalyzes superoxide oxidation to molecular oxygen and reduction to hydrogen peroxide, while catalases and peroxidases convert hydrogen peroxide to oxygen and water (Halliwell 1994; Betteridge 2000). Together, these three enzymes control ROS levels; however, during stress, up-regulation of heat shock proteins also contributes to regulating ROS and repairing cells (Feder and Hofmann 1999; Tarrant et al. 2014).
Through genome and transcriptome sequencing , many antioxidant genes have been identified in the Cnidaria. Antioxidant genes have been found in the sea anemones, Nematostella vectensis (Goldstone 2008; Reitzel et al. 2008) and Aiptasia pallida (Sunagawa et al. 2009b). An analysis of the Nematostella vectensis genome described the “defensome” (the battery of genes in charge of immune production), in which antioxidants play a key role in immune responses (Goldstone 2008). Two kinds of SOD (CuZnSOD and MnSOD ) and several catalase genes were identified in the genomes and transcriptomes of the corals Acropora digitifera (Shinzato et al. 2011), Acropora palmata (DeSalvo et al. 2010) and Orbicella faveolata (DeSalvo et al. 2008). Antioxidant genes were also part of the immune responses of the sea fan Gorgonia ventalina to the unicellular parasite Aplanochytrium (Burge et al. 2013). Genes for catalase have been identified in Hydra vulgaris (Wenger et al. 2014). These sequences have helped characterize the protein structure of cnidarian SOD and catalases (Dash et al. 2007; Dash and Phillips 2012).
Protein activity levels of antioxidants, such as SOD, catalase and peroxidase, have been measured in many different species. Anemonia viridis has two kinds of SOD (CuZnSOD and MnSOD ) enzymes, both of which are found in the ectoderm and the endoderm of this anemone. In the ectoderm, SOD activity is low and catalase activity is high, in an apparent discordance between the functions and relation between these enzymes (Richier et al. 2003). The reason could be that catalase acts against external stressors in addition to scavenging hydrogen peroxide produced by SOD (Merle et al. 2007). SOD and catalase activity were observed in 24 cnidarians from the Great Barrier Reef (Shick and Dykens 1985), and demonstrated a positive correlation across most of the collected species, implying that catalase activity is related to the production of hydrogen peroxide by SOD.
Hawkridge et al. (2000) used immunocytochemical techniques to establish the presence, location and distribution of SOD, catalase and glutathione peroxidase in the sea anemone Anemonia viridis and in the coral Goniopora stokesi . Antioxidants in these two cnidarians are associated with granular vesicles and accumulation bodies, indicating the conformation of peroxisomes (Hawkridge et al. 2000). Five peroxidase isoforms occur in the sea fan Gorgonia ventalina and were induced following experimental inoculation with the fungal pathogen Aspergillus sydowii (Mydlarz and Harvell 2007). Furthermore, peroxidase was localized in granular amoebocytes in the process of phagocytosis in the gorgonian Swiftia exserta (Olano and Bigger 2000). Peroxidase, along with other antioxidant enzymes, occurs constitutively in seven species of healthy Caribbean scleractinian corals (Mydlarz and Palmer 2011). Peroxidase activity varied at the genus and species level in Caribbean corals and was correlated with disease prevalence (Pinzon et al. 2014a).
There are several other antioxidant systems that may serve as supplemental systems when enzymatic antioxidant systems are overwhelmed. Fluorescent proteins (FPs) have multiple functions in anthozoan symbioses , primarily in photoprotection (Salih et al. 2000). With respect to immunity, their most important role is to scavenge hydrogen peroxide, and thereby contribute to the host’s antioxidant defense (Palmer et al. 2009a). Coral tissue often develops abnormal pigmentation in response to injury or infection, and red FPs, which exhibit significantly greater total fluorescence and hydrogen peroxide scavenging activity, are often associated with this pigmentation response (Palmer et al. 2009a, b). FPs are locally induced in Porites compressa tissue harboring parasitic trematodes (Palmer et al. 2008), with the resulting pink tissue attracting coral-feeding butterflyfishes that consume the parasite and enable the coral polyps to recover (Aeby 1992).
Mycosporine-like amino acids and dimethylsulfonioproprionate represent other antioxidant molecules and mechanisms identified in cnidarians (Lesser 2006; Van Alstyne et al. 2006). Their roles have mainly been characterized in response to environmental stress , but like FPs, they may be important in immunity as supplemental antioxidant systems .
5.2 Apoptosis
Apoptosis is a key biological process important in many aspects of the development and functionality of all organisms, including defense against diseases (Zmasek et al. 2007; Di Pietro et al. 2009). In fact, as a testament to the critical role of apoptosis in immunity, some pathogens develop regulating factors to control apoptosis as a way to increase virulence (Hedrick et al. 2010). During apoptosis, old, damaged and/or infected cells are destroyed. Cell death starts with signaling recognition and activation of the initiation caspases, and is followed by the activation of executing caspases and cell deletion (Dunn et al. 2006). This process is highly conserved in nature and is regulated by pro- (e.g., Bax) and anti- (e.g., Bcl2) apoptosis proteins. Apoptosis-related proteins include NB-ARC, caspases, caspase recruitment domain (CARD), death domain (DD) and death domain effectors (DDE) (Zmasek et al. 2007). These proteins form the same complexes, with similar domain organization (i.e. CARD-NB-ARC-repeats) among cnidarians and across vertebrates. Genomic data from Nematostella vectensis and Hydra magnipapillata indicate that cnidarians possess some of the most variable apoptosis-protein cores among animals, with 4–11 proteins (Zmasek et al. 2007).
Several genes associated with apoptosis, including both pro- and anti-apoptotic regulators were differentially affected in White Band Disease infected Acropora cervicornis . Tumor necrosis factor receptor superfamily member 1A (TNFRSF1A) and caspase 3 (CASP-3) genes were up-regulated, while caspase 8 (CASP-8) was down-regulated in White Band Disease infected corals (Libro et al. 2013). Similarly, in Indo-Pacific acroporid corals, individuals infected with White Band Disease had cellular markers indicative of apoptosis. The proportion of apoptotic cells was dramatically increased in diseased corals; that, combined with the absence of a clear pathogen, suggested that apoptosis was part of the disease etiology (Ainsworth et al. 2007). Characteristic blebbing of apoptotic cells was also detected in the wound-healing clot of Porites cylindrica (Palmer et al. 2011c).
5.3 Wound Healing
Wound healing is important as it re-establishes the function(s) of injured areas and prevents infections (Poss 2010; Galliot 2012). In sessile organisms, including most cnidarians, which are continuously exposed to physical damage by both abiotic and biotic stressors , including disease, wound healing is extremely important (Reitzel et al. 2008). Immediately after injury, the host response is activated to prevent infection and start the removal of cellular debris around the wound area (Peiris et al. 2014). Wound healing involves several phases: plug formation, inflammation , tissue proliferation and maturation (DesVoigne and Sparks 1968; Fontaine and Lightner 1973; Sherratt and Murray 1990; Theopold et al. 2004; Palmer et al. 2011c). The coordination and synchronization of the different components of this process is complex and involves several pathways (Godwin and Brockes 2006; Gurtner et al. 2008; Godwin and Rosenthal 2014). Differences in the healing and regeneration capabilities across taxa have been linked to the complexity of the immune system (Godwin and Brockes 2006; Godwin and Rosenthal 2014; Peiris et al. 2014). In general, organisms with “simple” immune systems (e.g., cnidarians) have a higher regeneration capacity, while higher animals with “complex” immune systems (e.g., mammals ) usually show low regeneration capabilities (Peiris et al. 2014).
Genomic analyses are now revealing details of the wound healing process in cnidarians (Pang et al. 2004; Reitzel et al. 2008). In Nematostella vectensis, the cells around an injury tend to cluster together, triggering the expression of genes related to wound healing and regeneration. The MAPK/ERK and Wnt pathways are actively involved in this process (Pang et al. 2004). MAPK/ERK regulates cell movement and recognition while Wnt regulates axis formation (Pang et al. 2004). In Hydra magnipapillata , Wnt3 signaling appears to be the primary tissue regeneration mechanism (Lengfeld et al. 2009; Galliot and Chera 2010). In these two cnidarians, regeneration and wound healing act in conjunction with apoptosis to control cell proliferation (Pang et al. 2004; Galliot and Chera 2010).
Several cell types with wound healing potential are known in the anemone Actinia equina (Hutton and Smith 1996) and the gorgonians Swiftia exserta and Plexaurella fusifera (Meszaros and Bigger 1999; Olano and Bigger 2000). Amoebocytes play a key role by aggregating near wound sites and initiating the production of connective mesogleal fibers to seal the wound (Meszaros and Bigger 1999; Mydlarz et al. 2008). This process was detailed in higher resolution studies of wound healing in Porites cylindrica and Acropora millepora . A sequence of four main events was characterized (Palmer et al. 2008, 2011c). First, plug formation via degranulation of melanin-containing granular cells occurs, indicated by a lack of melanin-containing granular cells and an increase in extracelluar melanin. This degranulation likely releases antimicrobial and cytotoxic material, including melanin, which can kill foreign organisms, and may also explain an immediate decline in zooxanthellae in the wound vicinity. In stage 2, immune cells, including eosinophilic (acidophilic) granular amoebocytes infiltrate the area as part of an inflammatory response, and spread out along the lesion edge. Agranular hyaline cells and fibroblasts with extensive pseudopodia help to connect the tissue components, and are assumed to secrete collagen. Stage 3 is characterized by granular tissue formation (proliferation) with an increase in melanin, possibly to provide structural support. Stage 4, maturation, is characterized by tissue differentiation, re-melanization of eosinophilic granular amoebocytes and apoptosis or programmed cell death of some immune cells.
Finally, another wound-healing related pathway in cnidarians is the Notch signaling pathway. This pathway regulates cell fate and differentiation, controls proliferation, and has been associated with wound healing (Chigurupati et al. 2007; Marlow et al. 2012). In Nematostella, Notch is fundamental for cellular differentiation during development and for cnidocyte cell fate (Marlow et al. 2012). In Hydra vulgaris, Notch signaling is essential to control the differentiation of interstitial cells (Kasbauer et al. 2007). All of these functions play significant roles during the final stages of wound healing.
6 Overlap Between Immunity and Symbiosis
Many Cnidarians, predominantly anthozoans (such as anemones and corals), form mutualistic relationships with dinoflagellate symbionts that provide much of the nutritional resource for the host in exchange for protection (Baird et al. 2009). Since the symbiont is intracellular, both the establishment and breakdown of the symbiosis involves host immunity (Vidal-Dupiol et al. 2009). Even though this review has focused on the immune responses of cnidarians to pathogens , we will briefly highlight the overlap and integration of immune functions in both facilitating the formation of the symbiosis and protecting the host from pathogens . A primary function of innate immunity is discrimination of self vs. non-self. The cnidarian immune system must thus be able to differentiate between non-self cells that can be tolerated, including the symbionts, and pathogens that need to be eliminated.
Cellular and physiological processes need to be modified to facilitate the formation of the symbiosis. A number of changes in gene expression are observed during the establishment of symbiosis in corals. These changes involve genes regulating cellular processes, such as those involved in cell cycle progression and apoptosis, and genes coding for lectin and other PRR domains (Schwarz et al. 2008a). Many of these patterns of gene expression are similar to those that occur during host-pathogen interactions (Rodriguez-Lanetty et al. 2006). Therefore, there is strong evidence for the involvement of multiple recognition genes that have shared function in both host defense and symbiont recognition (Rodriguez-Lanetty et al. 2006; Schwarz et al. 2008a).
6.1 Establishment of Symbiosis
The recognition of symbionts by cnidarian hosts is believed to be largely dependent on lectin /glycan interactions. In order for symbionts to be recognized by the host, they must possess an intact cell surface. Furthermore, α-mannose/α-glucose and α-galactose residues have been identified as potential Symbiodinium cell-surface ligands essential in recognition (Wood-Charlson et al. 2006). One of the first lectins described as having a potential role in the recognition of symbionts was SLL-2, a D-galactose binding lectin isolated from the soft coral Sinularia lochmodes. SLL-2 is highly expressed near the surface of symbionts, suggesting its role in the maintenance of symbiosis (Jimbo et al. 2000). A second lectin important in symbiosis is CecL from the scleractinian coral Fungia echinata, which binds to melibiose, lactose, and D-galactose. Exposure of symbionts to CecL triggers phenotypic changes, such as reduced motility and growth, suggesting that CecL, may play an important role in regulating symbiotic relationships in cnidarians (Jimbo et al. 2010). Finally, Pocillopora damicornis C type lectin (PdC-Lectin), which is specific for mannose, is believed to mediate molecular interactions between the host and symbiont. PdC-Lectin is highly expressed in endodermal tissue that is in contact with transient free zooxanthellae. Furthermore, expression of this protein has been shown to decrease just prior to bleaching, suggesting a role in the maintenance of symbiosis (Vidal-Dupiol et al. 2009).
Other lectins that have been identified as crucial to symbiotic processes in cnidarians also have immune functions. The best documented of these molecules is millectin (Kvennefors et al. 2008, 2010), which binds both bacterial pathogens and certain members of the genus Symbiodinium. Millectin binds Symbiodinium clades C1, C2, and A1, leading to agglutination and opsonisation of the potential symbionts (Kvennefors et al. 2008). Furthermore, millectin has been observed in coral epithelial cells and surrounding Symbiodinium (Kvennefors et al. 2010). Based on these findings, it is hypothesized that millectin plays a role in triggering the phagocytosis of Symbiodinium, while other proteins determine whether the phagocytosed cells will become resident in host cells (Kvennefors et al. 2008). In addition to millectin, the complement protein C3 is also believed to play a role in symbiosis. C3 is expressed in areas of the gastroderm in close association with symbionts. C3 may serve a similar role to millectin, mediating phagocytosis of Symbiodinium to initiate symbiosis ; however, further evidence is needed to fully understand its role in mediating this symbiosis (Kvennefors et al. 2010).
In addition to the dual use of cnidarian receptors for immune function and establishment of symbiosis, a number of effector responses may also affect both immunity and symbiosis. The sphingosine rheostat, a homeostatic cell regulatory pathway consisting of the sphingolipids sphingosine (Sph) and sphingosine 1-phosphate (S1P), impacts a number of cellular functions, including immunity, cell fate, and cell differentiation (Spiegel and Milstien 2000). Transcriptomic analysis has identified the sphigosine rheostat as a potential pathway involved in the regulation of symbiosis in cnidarians (Rodriguez-Lanetty et al. 2006). Furthermore, excess S1P may reduce the prevalence of heat-induced bleaching in cnidarians, while excess Sph may increase bleaching frequency in response to heat stress (Detournay and Weis 2011). Whereas evidence indicates that the sphigosine rheostat plays a role in regulating symbiosis, the potential impacts of this system on immunity are unknown.
In contrast, recent studies have suggested that TGF-β may have effects on both establishment of symbiosis and immunity in cnidarians (Detournay et al. 2012). Binding of TGF-β to its cellular receptor results in the production of SMAD intermediate compounds which can regulate expression of immune genes, resulting in immunosuppression (Massague 1998). Genes encoding for both TGF-β and its receptor have been found in Aiptasia pallida, in which Symbiodinium may up-regulate host expression of TGF-β, thereby resulting in immunosuppression and tolerance of symbiosis (Detournay et al. 2012). While most known cnidarian immune receptors are designed to elicit an up-regulation of immune response, immunosuppression by a TGF-β receptor suggests that other mechanisms may exist in cnidarians in which immunity may be suppressed via receptor action.
In conclusion, the significant systemic overlap in processes involved in both the establishment of symbiosis and the immune system suggests that many of the genes and molecules essential to the establishment and maintenance of symbioses have been evolutionarily co-opted from existing immune pathways. Therefore, when considering questions of cnidarian immunity, it is essential to also consider the potential effects of symbiotic processes on immune functions of the host organism .
6.2 Breakdown of Symbiosis
The breakdown of symbiosis (known as bleaching) also involves many overlapping processes with immunity. ROS are produced as part of metabolism in photosynthetic symbionts (Lesser 2006, 2007; McGinty et al. 2012). During stress , the levels of ROS can exceed constitutive antioxidant systems and become harmful to the host coral. This is believed to be one of the drivers of coral bleaching (Lesser 2006, 2007). Since ROS is also a powerful signaling molecule and part of the oxidative burst in phagocytes, cross-talk between algal ROS and host immune pathways may occur (Mydlarz et al. 2009).
Antioxidants, such as SOD , are expressed by both the cnidarian host and the algal symbiont , and can aid in regulating oxidative stress during pathogen-induced host responses (Mydlarz et al. 2006) and during thermal and UV stress in corals (Lesser 1997; Lesser and Farrell 2004). In the anemone Anemonia viridis and the coral Goniopora stokesi , SOD and catalase were found throughout the tissue in both species, but were localized in the cnidae , intracellular granules throughout the host, and the symbiont itself (Hawkridge et al. 2000). Downs et al. (2002) and McGinty et al. (2012) also showed that various Symbiodinium clades produce antioxidants and can increase production at elevated temperatures.
Apoptosis is a key player during the breakdown of symbiosis in cnidarians. The activation of apoptotic pathways (along with autophagy ) during bleaching may remove symbiont cells identified as dysfunctional (Yu et al. 2006). This is essentially an activation of immune pathways to reject and remove foreign invaders (“symbiosis gone wrong”). During thermal stress , the corals Acropora aspera and Stylophora pistillata regulate pro- (Bax-like) proteins and anti- (Bcl2-like) apoptosis genes by differentially expressing them in different areas of the polyps . Apoptosis is initially active in the gastrodermis. Later, as stress continues, and Symbiodinium cell density is reduced, the activity is centered in the epithelium (Ainsworth et al. 2008; Kvitt et al. 2011). In Acropora millepora , up-regulation of Bcl2-like genes may be a protective response in the unaffected or surviving cells during high-temperature stress (Pernice et al. 2011). Bcl2 up-regulation might also help surviving corals to recover faster from thermal related bleaching events (Kvitt et al. 2011).
In addition to the intrinsic Bax-Bcl2 controlled activation, apoptosis can be activated by extrinsic mechanisms such as the tumor necrosis factor (TNF) pathway (Hedrick et al. 2010). Several TNF receptors are critical for the onset of apoptosis in corals (Quistad et al. 2014). Activation of the TNF pathway is important against biological stressors (Ainsworth et al. 2007; Libro et al. 2013) and may also play a significant role in the survival and adaptation of corals to climate change (Barshis et al. 2013; Palumbi et al. 2014). Additionally, MAPK signaling is likely involved in exocytosis of Symbiodinium in two scleractinians, Acropora palmata and Orbicella faveolata , and in Nematostella vectensis (Granados-Cifuentes et al. 2013).
Immune responses in some coral diseases seem to originate from Symbiodinium. Caribbean Yellow Band Disease appears to be caused by Vibrio spp. that infect the symbionts and cause chlorosis (Mydlarz et al. 2009). The coral Orbicella faveolata up-regulates immune effector responses when infected with Caribbean Yellow Band Disease, illustrating the crossover of immune and symbiosis signaling. In several other disease systems, such as Oculina bacterial bleaching (Rosenberg et al. 2009) and Acropora White Syndrome (Sussman et al. 2008), bacterial virulence factors affect the Symbiodinium directly, eliciting disease etiology in the coral. Since the health of the symbiont is inextricably linked to health of the coral host , adopting a ‘your fight is my fight’ approach against symbiont diseases would be advantageous to both members of the symbiosis .
7 General Conclusions and Discussion
Despite their basal phylogenetic placement, cnidarians possess a highly complex innate immune system . To recognize invaders , three main groups of receptors (TLRs , lectins , and NLRs) likely initiate downstream immune responses. The diversification and variation in the PRRs across different cnidarian lineages may be attributed to each lineage’s unique ecological needs, primarily the demand for maintenance of diverse holobionts (Miller et al. 2007; Schwarz et al. 2007, 2008b; Shinzato et al. 2011). The need to recognize and distinguish between commensal and pathogenic microorganisms, including algal symbionts , may be a selective force driving this diversity of PRRs in cnidarians (Hayes et al. 2010; Hamada et al. 2013; Poole and Weis 2014). Signaling cascades in the Cnidaria are among the most complex components of the immune response. Many of the cascades are highly conserved within invertebrate innate immunity (i.e., PPO and TLR ), while others are homologous with mammalian cascades (e.g., GIMAP GTPases). Since many of these cascades work behind the scenes, identifying effector responses that have clear phenotypes has been more fruitful. Antimicrobial activity (e.g., AMPs and secondary metabolites) and encapsulation (cellular and skeletal) have been identified in cnidarians, and many of these cellular mechanisms play roles in would healing and tissue repair.
One of the goals of this chapter was to connect what is known about the presence of immune genes to phenotypes of either immunity itself or disease resistance. It was clear after compiling the data that for many of the known immune pathways, information providing this direct connection either exists in fragmented form or does not exist at all. There is also a disparity in the type of information available on each of the four stages of immunity we described. For example, receptor identity and signaling cascades are heavily reflected in genomic data , with the presence of gene homologs in genomes and transcriptomes , while information on effector responses and, to some extent wound repair , rely predominantly on observations of cellular, biochemical and antimicrobial activity.
In addition to understanding the genome to phenotype links within one arm of immunity, we still need more information on the identities of and mechanisms by which PRRs lead to specific effector responses. For example, in other invertebrate systems, activation of TLRs and subsequent signaling pathways lead to up-regulation of AMPs (Aderem and Ulevitch 2000; Akira et al. 2006). While these elements have been identified in the Cnidaria, especially within the Anthozoa, the connections between the entire cascade are not fully understood (Fig. 28.3). The discovery of the AMP damicornin in the coral Pocillopora damicornis , and identification of its genetic sequence, cellular localization, antibacterial activity and response to pathogen challenge (Vidal-Dupiol et al. 2011) bring us a few steps further to this goal. Likewise, while it is clear that amoebocytes and other mobile cytotoxic cells respond to infections and aggregate near lesions or wounds (Mydlarz et al. 2008; Palmer et al. 2008, 2011c), it is less clear what genes or upstream processes control differentiation and movement of these cells. If this process is analogous to other invertebrates, it is likely the integrin or lectin /complement pathway are involved, but again, this is still unknown. In many cases, as the tools and experimental techniques advance, so will our understanding of the connection between genotypes and phenotypes.

Details of three coral immune pathways (lectins , antimicrobial peptides and prophenoloxidase) and the known and (yet) unknown linkages between the genome (i.e. gene identity) and phenotype (e.g., effectors and role in disease or pathogen exposure). Select examples and references for Response to Disease are as follows: (1) Acropora cervicornis with White Band Disease (Libro et al. 2013), (2) Pocillopora damicornis exposed to bacteria (Vidal-Dupiol et al. 2014), (3) Acropora millepora exposed to bacteria (Brown et al. 2013), (4) Gorgonia ventalina following inoculation with the parasite Aplanochytrium (Burge et al. 2013), (5) Acropora millepora following exposure to many pathogens (Kvennefors et al. 2008), (6) Pocillopora damicornis exposed to bacteria (Vidal-Dupiol et al. 2011), (7) Gorgonia ventalina infected with Aspergillus sydowii (Mydlarz et al. 2008), (8) Acropora millepora exposed to PAMPs (Weiss et al. 2013), (9) Wound healing in Pocillopora damicornis (Palmer et al. 2012). Two-headed arrow signifies the interaction between genome and phenome that can go in either direction, images are generic representations of genome to phenome in coral immunity (left to right): generic heat map of gene expression, generic cell signaling pathway , ameobocytes aggregating in Gorgonia ventalina , melanin deposition in Gorgonia ventalina , clear zone of inhibition showing antibacterial activity , Gorgonia ventalina showing whole organismal immune response to Aspergillus sydowii
Another main goal of understanding cnidarian immunity is elucidating which elements lead to organismal disease resistance. Diseases affecting scleractinian corals, one of the cnidarian groups hardest hit by disease, are caused by a diversity of etiological agents (e.g., bacteria, fungi, viruses, etc.), increasing the challenge of this endeavor. It is unlikely that one cascade or PRR is responsible for resistance against all types of pathogens , but identifying the genetic basis of variation in susceptibility to a particular disease will help aid in predicting population-wide consequences of a disease outbreak. To date, several phenotypes have been linked to species-specific disease resistance, including PO/PPO (Palmer et al. 2010; Pinzon et al. 2014a) and antimicrobial activity (Pinzon et al. 2014a), but the basis for this variation is not well understood (Pinzon et al. 2014b).
Despite significant progress in the last 10–15 years, much is still unknown about cnidarian immunity, mainly with respect to how it is coordinated and executed, and how key processes are translated from receptor to effector. Confirming these specific functional linkages will require empirical controlled experiments in which cnidarian host responses (both genotypic and phenotypic) to specific stressors , such as pathogen challenge, are evaluated. Advances in the field of cnidarian immunity are expected to increase exponentially in the coming years as more genomic and molecular resources become available. With tools in place, discerning and linking gene networks to their function in fighting pathogens and their roles in disease resistance will become possible.
Abbreviations
- AMP:
-
Antimicrobial peptide
- C3:
-
Complement protein
- LPS:
-
Lipopolysaccharide
- LRR:
-
Leucine rich repeat
- MACPF:
-
Membrane-attack complex–perforin protein
- MAPK/ERK:
-
Mitogen activation protein kinases/extracellular signal-regulated kinases
- MASP:
-
Mannose-binding lectin associated serine protease
- Myd88:
-
Myeloid differentiation primary response protein 88
- NF-κB:
-
Nuclear factor kappa light chain of activated B cells
- PAMP:
-
Pathogen/microbe-associated molecular pattern
- PO/PPO:
-
Phenoloxidase/prophenoloxidase
- PRR:
-
Pathogen recognition receptor
- ROS:
-
Reactive oxygen species
- SML:
-
Surface mucus layer
- SOD:
-
Superoxide dismutase
- TGF-β:
-
Transforming growth factor beta
- TIR:
-
Toll/interleukin-1 receptor
- TLR:
-
Toll-like receptor
References
Aceret TL, Coll JC, Uchio Y, Sammarco PW (1998) Antimicrobial activity of the diterpenes flexibilide and sinulariolide derived from Sinularia flexibilis Quoy and Gaimard 1833 (Coelenterata: Alcyonacea, Octocorallia). Comp Biochem Physiol C Pharmacol Toxicol Endocrinol 120(1):121–126
Aderem A, Ulevitch RJ (2000) Toll-like receptors in the induction of the innate immune response. Nature 406(6797):782–787. doi:10.1038/35021228
Aeby GS (1992) The potential effect of the ability of a coral intermediate host to regenerate has had on the evolution of its association with a marine parasite. In: Proceedings of the 7th international coral reef symposium, University of Guam Marine Laboratory, Guam, 22–27 June 1992
Aeby GS (2003) Corals in the genus Porites are susceptible to infection by a larval trematode. Coral Reefs 22(3):216–216. doi:10.1007/s00338-003-0310-9
Affeld S, Wagele H, Avila C, Kehraus S, Konig GM (2006) Distribution of homarine in some Opisthobranchia (Gastropoda: Mollusca). In: 2nd international workshop on Opisthobranchia, Zoologisches Forschungsmuseum Koenig, 20–22 September 2006
Ainsworth TD, Kvennefors EC, Blackall LL, Fine M, Hoegh-Guldberg O (2007) Disease and cell death in white syndrome of Acroporid corals on the Great Barrier Reef. Mar Biol 151(1):19–29. doi:10.1007/S00227-006-0449-3
Ainsworth TD, Hoegh-Guldberg O, Heron SF, Skirving WJ, Leggat W (2008) Early cellular changes are indicators of pre-bleaching thermal stress in the coral host. J Exp Mar Biol Ecol 364(2):63–71. doi:10.1016/J.Jembe.2008.06.032
Akira S, Takeda K (2004) Toll-like receptor signalling. Nat Rev Immunol 4(7):499–511. doi:10.1038/nri1391
Akira S, Uematsu S, Takeuchi O (2006) Pathogen recognition and innate immunity. Cell 124(4):783–801. doi:10.1016/j.cell.2006.02.015
Alker AP, Kim K, Dube DH, Harvell CD (2004) Localized induction of a generalized response against multiple biotic agents in Caribbean sea fans. Coral Reefs 23(3):397–405. doi:10.1007/s00338-004-0405-y
Al-Lihaibi SS (2005) Chemical and pharmacological characterization of Fire coral from Eastern Red Sea. Res J Chem Environ Sci 68:28–32
Armoza-Zvuloni R, Shaked Y (2014) Release of hydrogen peroxide and antioxidants by the coral Stylophora pistillata to its external milieu. Biogeosciences 11(17):4587–4598. doi:10.5194/Bg-11-4587-2014
Astley MR, Ratcliffe NA (1989) Marine invertebrate mucus – agglutinating and antibacterial activity, with emphasis on Metridium senile. Symp Soc Exp Biol 43:367–377
Augustin R, Anton-Erxleben F, Jungnickel S, Hemmrich G, Spudy B, Podschun R, Bosch TC (2009) Activity of the novel peptide arminin against multiresistant human pathogens shows the considerable potential of phylogenetically ancient organisms as drug sources. Antimicrob Agents Chemother 53(12):5245–5250. doi:10.1128/AAC.00826-09
Augustin R, Fraune S, Bosch TC (2010) How Hydra senses and destroys microbes. Semin Immunol 22(1):54–58. doi:10.1016/j.smim.2009.11.002
Baird AH, Bhagooli R, Ralph PJ, Takahashi S (2009) Coral bleaching: the role of the host. Trends Ecol Evol 24(1):16–20. doi:10.1016/j.tree.2008.09.005
Barshis DJ, Ladner JT, Oliver TA, Seneca FO, Traylor-Knowles N, Palumbi SR (2013) Genomic basis for coral resilience to climate change. Proc Natl Acad Sci U S A 110(4):1387–1392. doi:10.1073/pnas.1210224110
Beisel HG, Kawabata S, Iwanaga S, Huber R, Bode W (1999) Tachylectin-2: crystal structure of a specific GlcNAc/GalNAc-binding lectin involved in the innate immunity host defense of the Japanese horseshoe crab Tachypleus tridentatus. EMBO J 18(9):2313–2322. doi:10.1093/emboj/18.9.2313
Betteridge DJ (2000) What is oxidative stress? Metabolism 49(2):3–8
Bosch TC (2008) The path less explored: innate immune reactions in Cnidarians. In: Heine H (ed) Innate immunity of plants, animals, and humans. Springer, Berlin, pp 27–42. doi:10.1007/978-3-540-73930-2_2
Bosch TC (2012) Understanding complex host-microbe interactions in Hydra. Gut Microbes 3(4):345–351. doi:10.4161/gmic.20660
Bosch TC (2014) Rethinking the role of immunity: lessons from Hydra. Trends Immunol 35(10):495–502. doi:10.1016/j.it.2014.07.008
Bosch TC, Augustin R, Anton-Erxleben F, Fraune S, Hemmrich G, Zill H, Rosenstiel P, Jacobs G, Schreiber S, Leippe M, Stanisak M, Grotzinger J, Jung S, Podschun R, Bartels J, Harder J, Schroder JM (2009) Uncovering the evolutionary history of innate immunity: the simple metazoan Hydra uses epithelial cells for host defence. Dev Comp Immunol 33(4):559–569. doi:10.1016/j.dci.2008.10.004
Bourne D, Iida Y, Uthicke S, Smith-Keune C (2008) Changes in coral-associated microbial communities during a bleaching event. ISME J 2(4):350–363. doi:10.1038/ismej.2007.112
Brown BE, Bythell JC (2005) Perspectives on mucus secretion in reef corals. Mar Ecol Prog Ser 296:291–309
Brown T, Bourne D, Rodriguez-Lanetty M (2013) Transcriptional activation of c3 and hsp70 as part of the immune response of Acropora millepora to bacterial challenges. PLoS One 8(7):e67246. doi:10.1371/journal.pone.0067246
Burge CA, Douglas NL, Conti-Jerpe I, Weil E, Roberts S, Friedman CS, Harvell CD (2012) Friend or foe: the association of Labyrinthulomycetes with the Caribbean sea fan Gorgonia ventalina. Dis Aquat Org 101(1):1–12. doi:10.3354/dao02487
Burge CA, Mouchka ME, Harvell CD, Roberts S (2013) Immune response of the Caribbean sea fan, Gorgonia ventalina, exposed to an Aplanochytrium parasite as revealed by transcriptome sequencing. Front Physiol 4:180. doi:10.3389/fphys.2013.00180
Burge CA, Mark Eakin C, Friedman CS, Froelich B, Hershberger PK, Hofmann EE, Petes LE, Prager KC, Weil E, Willis BL, Ford SE, Harvell CD (2014) Climate change influences on marine infectious diseases: implications for management and society. Annu Rev Mar Sci 6:249–277. doi:10.1146/annurev-marine-010213-135029
Chigurupati S, Arumugam TV, Son TG, Lathia JD, Jameel S, Mughal MR, Tang SC, Jo DG, Camandola S, Giunta M, Rakova I, McDonnell N, Miele L, Mattson MP, Poosala S (2007) Involvement of notch signaling in wound healing. PLoS One 2(11):e1167. doi:10.1371/journal.pone.0001167
Couch CS, Mydlarz LD, Harvell CD, Douglas NL (2008) Variation in measures of immunocompetence of sea fan coral, Gorgonia ventalina, in the Florida Keys. Mar Biol 155(3):281–292. doi:10.1007/s00227-008-1024-x
Couch CS, Weil E, Harvell CD (2013) Temporal dynamics and plasticity in the cellular immune response of the sea fan coral, Gorgonia ventalina. Mar Biol 160(9):2449–2460. doi:10.1007/s00227-013-2240-6
D’Angelo C, Smith EG, Oswald F, Burt J, Tchernov D, Wiedenmann J (2012) Locally accelerated growth is part of the innate immune response and repair mechanisms in reef-building corals as detected by green fluorescent protein (GFP)-like pigments. Coral Reefs 31(4):1045–1056. doi:10.1007/s00338-012-0926-8
Dash B, Phillips TD (2012) Molecular characterization of a catalase from Hydra vulgaris. Gene 501(2):144–152. doi:10.1016/j.gene.2012.04.015
Dash B, Metz R, Huebner HJ, Porter W, Phillips TD (2007) Molecular characterization of two superoxide dismutases from Hydra vulgaris. Gene 387(1–2):93–108. doi:10.1016/j.gene.2006.08.020
DeSalvo MK, Voolstra CR, Sunagawa S, Schwarz JA, Stillman JH, Coffroth MA, Szmant AM, Medina M (2008) Differential gene expression during thermal stress and bleaching in the Caribbean coral Montastraea faveolata. Mol Ecol 17(17):3952–3971. doi:10.1111/j.1365-294X.2008.03879.x
DeSalvo MK, Sunagawa S, Fisher PL, Voolstra CR, Iglesias-Prieto R, Medina M (2010) Coral host transcriptomic states are correlated with Symbiodinium genotypes. Mol Ecol 19(6):1174–1186. doi:10.1111/j.1365-294X.2010.04534.x
DesVoigne DM, Sparks AK (1968) The process of wound healing in the Pacific oyster, Crassostrea gigas. J Invertebr Pathol 12(1):53–65. doi:10.1016/0022-2011(68)90243-7
Detournay O, Weis VM (2011) Role of the sphingosine rheostat in the regulation of cnidarian-dinoflagellate symbioses. Biol Bull 221(3):261–269
Detournay O, Schnitzler CE, Poole A, Weis VM (2012) Regulation of cnidarian-dinoflagellate mutualisms: evidence that activation of a host TGFbeta innate immune pathway promotes tolerance of the symbiont. Dev Comp Immunol 38(4):525–537. doi:10.1016/j.dci.2012.08.008
Di Pietro C, Ragusa M, Barbagallo D, Duro LR, Guglielmino MR, Majorana A, Angelica R, Scalia M, Statello L, Salito L (2009) The apoptotic machinery as a biological complex system: analysis of its omics and evolution, identification of candidate genes for fourteen major types of cancer, and experimental validation in CML and neuroblastoma. BMC Med Genom 2(1):20. doi:10.1186/1755-8794-2-20
Dishaw LJ, Smith SL, Bigger CH (2005) Characterization of a C3-like cDNA in a coral: phylogenetic implications. Immunogenetics 57(7):535–548. doi:10.1007/s00251-005-0005-1
Domart-Coulon IJ, Traylor-Knowles N, Peters EC, Elbert D, Downs CA, Price K, Stubbs J, McLaughlin S, Cox E, Aeby G, Brown PR, Ostrander GK (2006) Comprehensive characterization of skeletal tissue growth anomalies of the finger coral Porites compressa. Coral Reefs 25(4):531–543. doi:10.1007/s00338-006-0133-6
Douglas NL, Mullen KM, Talmage SC, Harvell CD (2006) Exploring the role of chitinolytic enzymes in the sea fan coral, Gorgonia ventalina. Mar Biol 150(6):1137–1144. doi:10.1007/s00227-006-0444-8
Downs CA, Fauth JE, Halas JC, Dustan P, Bemiss J, Woodley CM (2002) Oxidative stress and seasonal coral bleaching. Free Radic Biol Med 33(4):533–543
Dube D, Kim K, Alker AP, Harvell CD (2002) Size structure and geographic variation in chemical resistance of sea fan corals Gorgonia ventalina to a fungal pathogen. Mar Ecol Prog Ser 231:139–150
Dubuc TQ, Traylor-Knowles N, Martindale MQ (2014) Initiating a regenerative response; cellular and molecular features of wound healing in the cnidarian Nematostella vectensis. BMC Biol 12:24. doi:10.1186/1741-7007-12-24
Dunn SR (2009) Immunorecognition and immunoreceptors in the Cnidaria. Invertebr Surviv J 6(1):7–14
Dunn SR, Phillips WS, Spatafora JW, Green DR, Weis VM (2006) Highly conserved caspase and Bcl-2 homologues from the sea anemone Aiptasia pallida: lower metazoans as models for the study of apoptosis evolution. J Mol Evol 63(1):95–107. doi:10.1007/s00239-005-0236-7
Endo Y, Takahashi M, Fujita T (2006) Lectin complement system and pattern recognition. Immunobiology 211(4):283–293. doi:10.1016/j.imbio.2006.01.003
Esposito R, D’Aniello S, Squarzoni P, Pezzotti MR, Ristoratore F, Spagnuolo A (2012) New insights into the evolution of metazoan tyrosinase gene family. PLoS One 7(4):e35731. doi:10.1371/journal.pone.0035731
Feder ME, Hofmann GE (1999) Heat-shock proteins, molecular chaperones, and the stress response: evolutionary and ecological physiology. Annu Rev Physiol 61(1):243–282. doi:10.1146/annurev.physiol.61.1.243
Filen S, Lahesmaa R (2010) GIMAP proteins in T-lymphocytes. J Signal Transduct 2010:268589. doi:10.1155/2010/268589
Fontaine CT, Lightner DV (1973) Observations on the process of wound repair in penaeid shrimp. J Invertebr Pathol 22(1):23–33. doi:10.1016/0022-2011(73)90005-0
Franzenburg S, Fraune S, Kunzel S, Baines JF, Domazet-Loso T, Bosch TC (2012) MyD88-deficient Hydra reveal an ancient function of TLR signaling in sensing bacterial colonizers. Proc Natl Acad Sci U S A 109(47):19374–19379. doi:10.1073/pnas.1213110109
Franzenburg S, Walter J, Kunzel S, Wang J, Baines JF, Bosch TC, Fraune S (2013) Distinct antimicrobial peptide expression determines host species-specific bacterial associations. Proc Natl Acad Sci U S A 110(39):E3730–E3738. doi:10.1073/pnas.1304960110
Fraune S, Augustin R, Anton-Erxleben F, Wittlieb J, Gelhaus C, Klimovich VB, Samoilovich MP, Bosch TC (2010) In an early branching metazoan, bacterial colonization of the embryo is controlled by maternal antimicrobial peptides. Proc Natl Acad Sci U S A 107(42):18067–18072. doi:10.1073/pnas.1008573107
Galliot B (2012) Hydra, a fruitful model system for 270 years. Int J Dev Biol 56(6–8):411–423. doi:10.1387/ijdb.120086bg
Galliot B, Chera S (2010) The Hydra model: disclosing an apoptosis-driven generator of Wnt-based regeneration. Trends Cell Biol 20(9):514–523. doi:10.1016/j.tcb.2010.05.006
Garcia GD, Gregoracci GB, Santos Ede O, Meirelles PM, Silva GG, Edwards R, Sawabe T, Gotoh K, Nakamura S, Iida T, de Moura RL, Thompson FL (2013) Metagenomic analysis of healthy and white plague-affected Mussismilia braziliensis corals. Microb Ecol 65(4):1076–1086. doi:10.1007/s00248-012-0161-4
Geffen Y, Rosenberg E (2004) Stress-induced rapid release of antibacterials by scleractinian corals. Mar Biol 146(5):931–935. doi:10.1007/s00227-004-1505-5
Geffen Y, Ron EZ, Rosenberg E (2009) Regulation of release of antibacterials from stressed scleractinian corals. FEMS Microbiol Lett 295(1):103–109. doi:10.1111/j.1574-6968.2009.01590.x
Gerdol M, De Moro G, Manfrin C, Venier P, Pallavicini A (2012) Big defensins and mytimacins, new AMP families of the Mediterranean mussel Mytilus galloprovincialis. Dev Comp Immunol 36(2):390–399. doi:10.1016/j.dci.2011.08.003
Gil-Agudelo DL, Myers C, Smith GW, Kim K (2006) Changes in the microbial communities associated with Gorgonia ventalina during aspergillosis infection. Dis Aquat Org 69(1):89–94. doi:10.3354/dao069089
Gimenez AV, Haran NS, Pereira NA, Acuña FH (2014) First report of phenoloxidase and peroxidase activities in two intertidal sea anemone species of Argentina. Invertebr Surviv J 11:192–196
Gochfeld DJ, Aeby GS (2008) Antibacterial chemical defenses in Hawaiian corals provide possible protection from disease. Mar Ecol Prog Ser 362:119–128. doi:10.3354/meps07418
Gochfeld DJ, Olson JB, Slattery M (2006) Colony versus population variation in susceptibility and resistance to dark spot syndrome in the Caribbean coral Siderastrea siderea. Dis Aquat Org 69(1):53–65. doi:10.3354/dao069053
Godwin JW, Brockes JP (2006) Regeneration, tissue injury and the immune response. J Anat 209(4):423–432. doi:10.1111/j.1469-7580.2006.00626.x
Godwin JW, Rosenthal N (2014) Scar-free wound healing and regeneration in amphibians: immunological influences on regenerative success. Differentiation 87(1–2):66–75. doi:10.1016/j.diff.2014.02.002
Golberg K, Eltzov E, Shnit-Orland M, Marks RS, Kushmaro A (2011) Characterization of quorum sensing signals in coral-associated bacteria. Microb Ecol 61(4):783–792. doi:10.1007/s00248-011-9848-1
Goldberg WM, Makemson JC, Colley SB (1984) Entocladia endozoica sp. nov., a pathogenic chlorophyte: structure, life history, physiology, and effect on its coral host. Biol Bull 166:368–383
Goldstone JV (2008) Environmental sensing and response genes in cnidaria: the chemical defensome in the sea anemone Nematostella vectensis. Cell Biol Toxicol 24(6):483–502. doi:10.1007/s10565-008-9107-5
Granados-Cifuentes C, Bellantuono AJ, Ridgway T, Hoegh-Guldberg O, Rodriguez-Lanetty M (2013) High natural gene expression variation in the reef-building coral Acropora millepora: potential for acclimative and adaptive plasticity. BMC Genomics 14:228. doi:10.1186/1471-2164-14-228
Gunasundari V, Ajith Kumar TT, Kumaresan S, Balagurunathan R, Balasubramanian T (2013) Isolation of aliphatic-antibiotic compounds from marine invertebrate, Heteractis magnifica ‘Quoy & Gaimard, 1833’ against captive marine ornamental fish pathogens. Indian J Geomarine Sci 42:807–811
Gurtner GC, Werner S, Barrandon Y, Longaker MT (2008) Wound repair and regeneration. Nature 453(7193):314–321. doi:10.1038/nature07039
Halliwell B (1994) Free radicals, antioxidants, and human disease: curiosity, cause, or consequence? Lancet 344(8924):721–724
Hamada M, Shoguchi E, Shinzato C, Kawashima T, Miller DJ, Satoh N (2013) The complex NOD-like receptor repertoire of the coral Acropora digitifera includes novel domain combinations. Mol Biol Evol 30(1):167–176. doi:10.1093/molbev/mss213
Hawkridge JM, Pipe RK, Brown BE (2000) Localisation of antioxidant enzymes in the cnidarians Anemonia viridis and Goniopora stokesi. Mar Biol 137(1):1–9. doi:10.1007/S002270000324
Hayden MS, Ghosh S (2004) Signaling to NF-kappaB. Genes Dev 18(18):2195–2224. doi:10.1101/gad.1228704
Hayes ML, Eytan RI, Hellberg ME (2010) High amino acid diversity and positive selection at a putative coral immunity gene (tachylectin-2). BMC Evol Biol 10:150. doi:10.1186/1471-2148-10-150
Hedrick SM, Ch’en IL, Alves BN (2010) Intertwined pathways of programmed cell death in immunity. Immunol Rev 236(1):41–53. doi:10.1111/j.1600-065X.2010.00918.x
Hemmrich G, Miller DJ, Bosch TC (2007) The evolution of immunity: a low-life perspective. Trends Immunol 28(10):449–454. doi:10.1016/j.it.2007.08.003
Hunt LR, Smith SM, Downum KR, Mydlarz LD (2012) Microbial regulation in gorgonian corals. Mar Drugs 10(6):1225–1243. doi:10.3390/md10061225
Hutton DMC, Smith VJ (1996) Antibacterial properties of isolated amoebocytes from the sea anemone Actinia equina. Biol Bull 191:441–451
Jimbo M, Yanohara T, Koike K, Koike K, Sakai R, Muramoto K, Kamiya H (2000) The D-galactose-binding lectin of the octocoral Sinularia lochmodes: characterization and possible relationship to the symbiotic dinoflagellates. Comp Biochem Physiol B Biochem Mol Biol 125(2):227–236
Jimbo M, Yamashita H, Koike K, Sakai R, Kamiya H (2010) Effects of lectin in the scleractinian coral Ctenactis echinata on symbiotic zooxanthellae. Fish Sci 76(2):355–363. doi:10.1007/s12562-009-0204-z
Jorge HP, Kamel B, Colleen AB, Harvell CD, Medina M, Weil E, Laura DM (2015) Whole transcriptome analysis reveals changes in expression of immune-related genes during and after bleaching in a reef-building coral. R Soc Open Sci 2015(2):140214. doi:10.1098/rsos.140214
Jung S, Dingley AJ, Augustin R, Anton-Erxleben F, Stanisak M, Gelhaus C, Gutsmann T, Hammer MU, Podschun R, Bonvin AM, Leippe M, Bosch TC, Grotzinger J (2009) Hydramacin-1, structure and antibacterial activity of a protein from the basal metazoan Hydra. J Biol Chem 284(3):1896–1905. doi:10.1074/jbc.M804713200
Kasbauer T, Towb P, Alexandrova O, David CN, Dall’armi E, Staudigl A, Stiening B, Bottger A (2007) The Notch signaling pathway in the cnidarian Hydra. Dev Biol 303(1):376–390. doi:10.1016/j.ydbio.2006.11.022
Kawabata S, Iwanaga S (1999) Role of lectins in the innate immunity of horseshoe crab. Dev Comp Immunol 23(4–5):391–400
Kelman D, Kashman Y, Hill RT, Rosenberg E, Loya Y (2009) Chemical warfare in the sea: the search for antibiotics from Red Sea corals and sponges. Pure Appl Chem 81(6):1113–1121. doi:10.1351/pac-con-08-10-07
Kim K, Harvell CD (2004) The rise and fall of a six-year coral-fungal epizootic. Am Nat 164(Suppl 5):S52–S63. doi:10.1086/424609
Kim K, Harvell CD, Kim PD, Smith GW, Merkel SM (2000a) Fungal disease resistance of Caribbean sea fan corals (Gorgonia spp.). Mar Biol 136:259–267
Kim K, Kim PD, Alker AP, Harvell CD (2000b) Chemical resistance of gorgonian corals against fungal infections. Mar Biol 137:393–401
Kim RO, Rhee JS, Won EJ, Lee KW, Kang CM, Lee YM, Lee JS (2011) Ultraviolet B retards growth, induces oxidative stress, and modulates DNA repair-related gene and heat shock protein gene expression in the monogonont rotifer, Brachionus sp. Aquat Toxicol 101(3–4):529–539. doi:10.1016/j.aquatox.2010.12.005
Kimura A, Sakaguchi E, Nonaka M (2009) Multi-component complement system of Cnidaria: C3, Bf, and MASP genes expressed in the endodermal tissues of a sea anemone, Nematostella vectensis. Immunobiology 214(3):165–178. doi:10.1016/j.imbio.2009.01.003
Klug M, Tardent P, Smid I, Holstein TW (1984) Presence and localization of chitinase in Hydra and Podocoryne (Cnidaria, Hydrozoa). J Exp Zool 229(1):69–72. doi:10.1002/jez.1402290109
Krasity BC, Troll JV, Weiss JP, McFall-Ngai MJ (2011) LBP/BPI proteins and their relatives: conservation over evolution and roles in mutualism. Biochem Soc Trans 39(4):1039–1044. doi:10.1042/BST0391039
Krediet CJ, Ritchie KB, Paul VJ, Teplitski M (2013) Coral-associated micro-organisms and their roles in promoting coral health and thwarting diseases. Proc Biol Sci 280(1755):20122328. doi:10.1098/rspb.2012.2328
Krediet CJ, Ritchie KB, Cohen M, Lipp EK, Sutherland KP, Teplitski M (2009) Utilization of mucus from the coral Acropora palmata by the pathogen Serratia marcescens and by environmental and coral commensal bacteria. Appl Environ Microbiol 75(12):3851–3858. doi:10.1128/AEM.00457-09
Kvennefors EC, Leggat W, Hoegh-Guldberg O, Degnan BM, Barnes AC (2008) An ancient and variable mannose-binding lectin from the coral Acropora millepora binds both pathogens and symbionts. Dev Comp Immunol 32(12):1582–1592. doi:10.1016/j.dci.2008.05.010
Kvennefors EC, Leggat W, Kerr CC, Ainsworth TD, Hoegh-Guldberg O, Barnes AC (2010) Analysis of evolutionarily conserved innate immune components in coral links immunity and symbiosis. Dev Comp Immunol 34(11):1219–1229. doi:10.1016/j.dci.2010.06.016
Kvitt H, Rosenfeld H, Zandbank K, Tchernov D (2011) Regulation of apoptotic pathways by Stylophora pistillata (Anthozoa, Pocilloporidae) to survive thermal stress and bleaching. PLoS One 6(12):e28665. doi:10.1371/journal.pone.0028665
Landis GN, Tower J (2005) Superoxide dismutase evolution and life span regulation. Mech Ageing Dev 126(3):365–379. doi:10.1016/j.mad.2004.08.012
Lange C, Hemmrich G, Klostermeier UC, Lopez-Quintero JA, Miller DJ, Rahn T, Weiss Y, Bosch TC, Rosenstiel P (2011) Defining the origins of the NOD-like receptor system at the base of animal evolution. Mol Biol Evol 28(5):1687–1702. doi:10.1093/molbev/msq349
Lehnert EM, Burriesci MS, Pringle JR (2012) Developing the anemone Aiptasia as a tractable model for cnidarian-dinoflagellate symbiosis: the transcriptome of aposymbiotic A. pallida. BMC Genomics 13:271. doi:10.1186/1471-2164-13-271
Lengfeld T, Watanabe H, Simakov O, Lindgens D, Gee L, Law L, Schmidt HA, Ozbek S, Bode H, Holstein TW (2009) Multiple Wnts are involved in Hydra organizer formation and regeneration. Dev Biol 330(1):186–199. doi:10.1016/j.ydbio.2009.02.004
Lesser MP (1997) Oxidative stress causes coral bleaching during exposure to elevated temperatures. Coral Reefs 16(3):187–192. doi:10.1007/s003380050073
Lesser MP (2006) Oxidative stress in marine environments: biochemistry and physiological ecology. Annu Rev Physiol 68:253–278. doi:10.1146/annurev.physiol.68.040104.110001
Lesser MP (2007) Coral reef bleaching and global climate change: can corals survive the next century? Proc Natl Acad Sci U S A 104(13):5259–5260. doi:10.1073/pnas.0700910104
Lesser MP, Farrell JH (2004) Exposure to solar radiation increases damage to both host tissues and algal symbionts of corals during thermal stress. Coral Reefs 23(3):367–377. doi:10.1007/s00338-004-0392-z
Leverette CL, Warren M, Smith MA, Smith GW (2008) Determination of carotenoid as the purple pigment in Gorgonia ventalina sclerites using Raman microscopy. Spectrochim Acta A Mol Biomol Spectrosc 69(3):1058–1061. doi:10.1016/j.saa.2007.07.018
Libro S, Kaluziak ST, Vollmer SV (2013) RNA-seq profiles of immune related genes in the staghorn coral Acropora cervicornis infected with white band disease. PLoS One 8(11):e81821. doi:10.1371/journal.pone.0081821
Lopez JA, Fain MG, Cadavid LF (2011) The evolution of the immune-type gene family Rhamnospondin in cnidarians. Gene 473(2):119–124. doi:10.1016/j.gene.2010.11.013
Maldonado A, Johnson A, Gochfeld DJ, Slattery M, Ostrander GK, Bingham JP, Schlenk D (in review) The role of cytochrome P450 in biotransformation of a hard coral (Porites lobata) extract and homarine in butterflyfishes. J Chem Ecol
Mali B, Mohrlen F, Frohme M, Frank U (2004) A putative double role of a chitinase in a cnidarian: pattern formation and immunity. Dev Comp Immunol 28(10):973–981. doi:10.1016/j.dci.2004.04.002
Mali B, Soza-Ried J, Frohme M, Frank U (2006) Structural but not functional conservation of an immune molecule: a tachylectin-like gene in Hydractinia. Dev Comp Immunol 30(3):275–281. doi:10.1016/j.dci.2005.04.004
Mann WT, Beach-Letendre J, Mydlarz LD (2014) Interplay between proteases and protease inhibitors in the sea fan – Aspergillus pathosystem. Mar Biol 161(10):2213–2220. doi:10.1007/s00227-014-2499-2
Marlow H, Roettinger E, Boekhout M, Martindale MQ (2012) Functional roles of Notch signaling in the cnidarian Nematostella vectensis. Dev Biol 362(2):295–308. doi:10.1016/j.ydbio.2011.11.012
Massague J (1998) TGF-beta signal transduction. Annu Rev Biochem 67:753–791. doi:10.1146/annurev.biochem.67.1.753
McGinty ES, Pieczonka J, Mydlarz LD (2012) Variations in reactive oxygen release and antioxidant activity in multiple Symbiodinium types in response to elevated temperature. Microb Ecol 64(4):1000–1007. doi:10.1007/s00248-012-0085-z
Medzhitov R (2001) Toll-like receptors and innate immunity. Nat Rev Immunol 1(2):135–145. doi:10.1038/35100529
Merle PL, Sabourault C, Richier S, Allemand D, Furla P (2007) Catalase characterization and implication in bleaching of a symbiotic sea anemone. Free Radic Biol Med 42(2):236–246. doi:10.1016/j.freeradbiomed.2006.10.038
Meszaros A, Bigger C (1999) Qualitative and quantitative study of wound healing processes in the coelenterate, Plexaurella fusifera: spatial, temporal, and environmental (light attenuation) influences. J Invertebr Pathol 73(3):321–331. doi:10.1006/jipa.1999.4851
Meyer JL, Paul VJ, Teplitski M (2014) Community shifts in the surface microbiomes of the coral Porites astreoides with unusual lesions. PLoS One 9(6):e100316. doi:10.1371/journal.pone.0100316
Miller DJ, Hemmrich G, Ball EE, Hayward DC, Khalturin K, Funayama N, Agata K, Bosch TC (2007) The innate immune repertoire in cnidaria – ancestral complexity and stochastic gene loss. Genome Biol 8(4):R59. doi:10.1186/gb-2007-8-4-r59
Miller DJ, Ball EE, Forêt S, Satoh N (2011) Coral genomics and transcriptomics – ushering in a new era in coral biology. J Exp Mar Biol Ecol 408(1–2):114–119. doi:10.1016/j.jembe.2011.07.031
Morse DE, Morse ANC, Duncan H (1977) Algal ‘tumors’ in the Caribbean sea-fan Gorgonia ventalina. In: Proceedings of third international coral reef symposium, University of Miami, May 1977
Moya A, Ganot P, Furla P, Sabourault C (2012) The transcriptomic response to thermal stress is immediate, transient and potentiated by ultraviolet radiation in the sea anemone Anemonia viridis. Mol Ecol 21(5):1158–1174. doi:10.1111/j.1365-294X.2012.05458.x
Mullen KM, Peters EC, Harvell CD (2004) Coral resistance to disease. In: Rosenberg E, Loya Y (eds) Coral health and disease. Springer, Berlin, pp 377–395
Mullen KM, Harvell CD, Alker AP, Dube D, Jordán-Dahlgren E, Ward JR, Petes LE (2006) Host range and resistance to aspergillosis in three sea fan species from the Yucatan. Mar Biol 149(6):1355–1364. doi:10.1007/s00227-006-0275-7
Mydlarz LD, Harvell CD (2007) Peroxidase activity and inducibility in the sea fan coral exposed to a fungal pathogen. Comp Biochem Physiol A Mol Integr Physiol 146(1):54–62. doi:10.1016/j.cbpa.2006.09.005
Mydlarz LD, Palmer CV (2011) The presence of multiple phenoloxidases in Caribbean reef-building corals. Comp Biochem Physiol A Mol Integr Physiol 159(4):372–378. doi:10.1016/j.cbpa.2011.03.029
Mydlarz LD, Jones LE, Harvell CD (2006) Innate immunity environmental drivers and disease ecology of marine and freshwater invertebrates. Annu Rev Ecol Evol Syst 37:251–288. doi:10.1146/Annurev.Ecolsys.37.091305.110103
Mydlarz LD, Holthouse SF, Peters EC, Harvell CD (2008) Cellular responses in sea fan corals: granular amoebocytes react to pathogen and climate stressors. PLoS One 3(3):e1811. doi:10.1371/journal.pone.0001811
Mydlarz LD, Couch CS, Weil E, Smith G, Harvell CD (2009) Immune defenses of healthy, bleached and diseased Montastraea faveolata during a natural bleaching event. Dis Aquat Org 87(1–2):67–78. doi:10.3354/dao02088
Nagai H, Oshiro N, Takuwa-Kuroda K, Iwanaga S, Nozaki M, Nakajima T (2002) A new polypeptide toxin from the nematocyst venom of an Okinawan sea anemone Phyllodiscus semoni (Japanese name “unbachi-isoginchaku”). Biosci Biotechnol Biochem 66(12):2621–2625. doi:10.1271/bbb.66.2621
Nappi AJ, Ottaviani E (2000) Cytotoxicity and cytotoxic molecules in invertebrates. Bioessays 22(5):469–480. doi:10.1002/(SICI)1521-1878(200005)22:5<469::AID-BIES9>3.0.CO;2-4
Narat M, Macek P, Kotnik V, Sedmak B (1994) The humoral and cellular immune response to a lipid attenuated pore-forming toxin from the sea anemone Actinia equina L. Toxicon 32(1):65–71
Netea MG, Quintin J, van der Meer JW (2011) Trained immunity: a memory for innate host defense. Cell Host Microbe 9(5):355–361. doi:10.1016/j.chom.2011.04.006
Olano CT, Bigger CH (2000) Phagocytic activities of the gorgonian coral Swiftia exserta. J Invertebr Pathol 76(3):176–184. doi:10.1006/jipa.2000.4974
Oshiro N, Kobayashi C, Iwanaga S, Nozaki M, Namikoshi M, Spring J, Nagai H (2004) A new membrane-attack complex/perforin (MACPF) domain lethal toxin from the nematocyst venom of the Okinawan sea anemone Actineria villosa. Toxicon 43(2):225–228. doi:10.1016/j.toxicon.2003.11.017
Otero-Gonzalez AJ, Magalhaes BS, Garcia-Villarino M, Lopez-Abarrategui C, Sousa DA, Dias SC, Franco OL (2010) Antimicrobial peptides from marine invertebrates as a new frontier for microbial infection control. FASEB J 24(5):1320–1334. doi:10.1096/fj.09-143388
Ovchinnikova TV, Balandin SV, Aleshina GM, Tagaev AA, Leonova YF, Krasnodembsky ED, Men’shenin AV, Kokryakov VN (2006) Aurelin, a novel antimicrobial peptide from jellyfish Aurelia aurita with structural features of defensins and channel-blocking toxins. Biochem Biophys Res Commun 348(2):514–523. doi:10.1016/j.bbrc.2006.07.078
Palmer CV, Traylor-Knowles N (2012) Towards an integrated network of coral immune mechanisms. Proc Biol Sci 279(1745):4106–4114. doi:10.1098/rspb.2012.1477
Palmer CV, Mydlarz LD, Willis BL (2008) Evidence of an inflammatory-like response in non-normally pigmented tissues of two scleractinian corals. Proc Biol Sci 275(1652):2687–2693. doi:10.1098/rspb.2008.0335
Palmer CV, Modi CK, Mydlarz LD (2009a) Coral fluorescent proteins as antioxidants. PLoS One 4(10):7298–7298. doi:10.1371/journal.pone.0007298
Palmer CV, Roth MS, Gates RD (2009b) Red fluorescent protein responsible for pigmentation in trematode-infected Porites compressa tissues. Biol Bull 216(1):68–74
Palmer CV, Bythell JC, Willis BL (2010) Levels of immunity parameters underpin bleaching and disease susceptibility of reef corals. FASEB J 24(6):1935–1946. doi:10.1096/fj.09-152447
Palmer CV, Bythell JC, Willis BL (2011a) A comparative study of phenoloxidase activity in diseased and bleached colonies of the coral Acropora millepora. Dev Comp Immunol 35(10):1098–1101. doi:10.1016/j.dci.2011.04.001
Palmer CV, McGinty ES, Cummings DJ, Smith SM, Bartels E, Mydlarz LD (2011b) Patterns of coral ecological immunology: variation in the responses of Caribbean corals to elevated temperature and a pathogen elicitor. J Exp Biol 214(Pt 24):4240–4249. doi:10.1242/jeb.061267
Palmer CV, Traylor-Knowles NG, Willis BL, Bythell JC (2011c) Corals use similar immune cells and wound-healing processes as those of higher organisms. PLoS One 6(8):e23992. doi:10.1371/journal.pone.0023992
Palmer CV, Bythell JC, Willis BL (2012) Enzyme activity demonstrates multiple pathways of innate immunity in Indo-Pacific anthozoans. Proc Biol Sci 279(1743):3879–3887. doi:10.1098/rspb.2011.2487
Palumbi SR, Barshis DJ, Traylor-Knowles N, Bay RA (2014) Mechanisms of reef coral resistance to future climate change. Science 344(6186):895–898. doi:10.1126/science.1251336
Pang K, Matus DQ, Martindale MQ (2004) The ancestral role of COE genes may have been in chemoreception: evidence from the development of the sea anemone, Nematostella vectensis (Phylum Cnidaria; Class Anthozoa). Dev Genes Evol 214(3):134–138. doi:10.1007/s00427-004-0383-7
Pantos O, Cooney RP, Le Tissier MD, Barer MR, O’Donnell AG, Bythell JC (2003) The bacterial ecology of a plague-like disease affecting the Caribbean coral Montastrea annularis. Environ Microbiol 5(5):370–382
Pappas KE (2010) Effects of bleaching and disease stress on antimicrobial chemical defenses in corals. Honors thesis, University of Mississippi
Peiris TH, Hoyer KK, Oviedo NJ (2014) Innate immune system and tissue regeneration in planarians: an area ripe for exploration. Semin Immunol 26(4):295–302. doi:10.1016/j.smim.2014.06.005
Perez S, Weis V (2006) Nitric oxide and cnidarian bleaching: an eviction notice mediates breakdown of a symbiosis. J Exp Biol 209(Pt 14):2804–2810. doi:10.1242/jeb.02309
Pernice M, Dunn SR, Miard T, Dufour S, Dove S, Hoegh-Guldberg O (2011) Regulation of apoptotic mediators reveals dynamic responses to thermal stress in the reef building coral Acropora millepora. PLoS One 6(1):e16095. doi:10.1371/journal.pone.0016095
Petes LE, Harvell CD, Peters EC, Webb MAH, Mullen KM (2003) Pathogens compromise reproduction and induce melanization in Caribbean sea fans. Mar Ecol Prog Ser 264:167–171
Phillips JH (1963) Immune mechanisms in the phylum Coelenterata. In: Dougherty EC (ed) The lower metazoa. University of California Press, Berkley, pp 425–432
Pinzon CJ, Beach-Letendre J, Weil E, Mydlarz LD (2014a) Relationship between phylogeny and immunity suggests older Caribbean coral lineages are more resistant to disease. PLoS One 9(8):e104787. doi:10.1371/journal.pone.0104787
Pinzon CJ, Dornberger L, Beach-Letendre J, Weil E, Mydlarz LD (2014b) The link between immunity and life history traits in scleractinian corals. Peer J 2:e628. doi:10.7717/peerj.628
Poole AZ, Weis VM (2014) TIR-domain-containing protein repertoire of nine anthozoan species reveals coral-specific expansions and uncharacterized proteins. Dev Comp Immunol 46(2):480–488. doi:10.1016/j.dci.2014.06.002
Poss KD (2010) Advances in understanding tissue regenerative capacity and mechanisms in animals. Nat Rev Genet 11(10):710–722. doi:10.1038/nrg2879
Puglisi MP, Paul VJ, Biggs J, Slattery M (2002) Co-occurrence of chemical and structural defenses in the gorgonian corals of Guam. Mar Ecol Prog Ser 239:105–114
Putnam NH, Srivastava M, Hellsten U, Dirks B, Chapman J, Salamov A, Terry A, Shapiro H, Lindquist E, Kapitonov VV, Jurka J, Genikhovich G, Grigoriev IV, Lucas SM, Steele RE, Finnerty JR, Technau U, Martindale MQ, Rokhsar DS (2007) Sea anemone genome reveals ancestral eumetazoan gene repertoire and genomic organization. Science 317(5834):86–94. doi:10.1126/science.1139158
Quistad SD, Stotland A, Barott KL, Smurthwaite CA, Hilton BJ, Grasis JA, Wolkowicz R, Rohwer FL (2014) Evolution of TNF-induced apoptosis reveals 550 My of functional conservation. Proc Natl Acad Sci U S A 111(26):9567–9572. doi:10.1073/pnas.1405912111
Rast JP, Messier-Solek C (2008) Marine invertebrate genome sequences and our evolving understanding of animal immunity. Biol Bull 214(3):274–283
Reitzel AM, Sullivan JC, Traylor-Knowles N, Finnerty JR (2008) Genomic survey of candidate stress-response genes in the estuarine anemone Nematostella vectensis. Biol Bull 214(3):233–254
Renegar D-EA, Blackwelder P, Miller J, Gochfeld D, Moulding AL (2008) Ultrastructural and histological analysis of dark spot syndrome in Siderastrea siderea and Agaricia agaricites
Reshef L, Koren O, Loya Y, Zilber-Rosenberg I, Rosenberg E (2006) The coral probiotic hypothesis. Environ Microbiol 8(12):2068–2073. doi:10.1111/j.1462-2920.2006.01148.x
Richier S, Merle PL, Furla P, Pigozzi D, Sola F, Allemand D (2003) Characterization of superoxide dismutases in anoxia- and hyperoxia-tolerant symbiotic cnidarians. Biochim Biophys Acta 1621(1):84–91
Rinkevich B (2004) Allorecognition and xenorecognition in reef corals: a decade of interactions. Hydrobiologia 530:443–450. doi:10.1007/S10750-004-2686-0
Ritchie K (2006) Regulation of microbial populations by coral surface mucus and mucus-associated bacteria. Mar Ecol Prog Ser 322:1–14
Ritchie K, Smith GW (1995) Carbon-source utilization of coral-associated marine heterotrophs. J Mar Biotechnol 3:107–109
Robb CT, Dyrynda EA, Gray RD, Rossi AG, Smith VJ (2014) Invertebrate extracellular phagocyte traps show that chromatin is an ancient defence weapon. Nat Commun 5:4627. doi:10.1038/ncomms5627
Rodriguez-Lanetty M, Phillips WS, Weis VM (2006) Transcriptome analysis of a cnidarian-dinoflagellate mutualism reveals complex modulation of host gene expression. BMC Genomics 7:23. doi:10.1186/1471-2164-7-23
Rosenberg E, Koren O, Reshef L, Efrony R, Zilber-Rosenberg I (2007) The role of microorganisms in coral health, disease and evolution. Nat Rev Microbiol 5(5):355–362. doi:10.1038/nrmicro1635
Rosenberg E, Kushmaro A, Kramarsky-Winter E, Banin E, Yossi L (2009) The role of microorganisms in coral bleaching. ISME J 3(2):139–146. doi:10.1038/ismej.2008.104
Ruiz-Moreno D, Willis BL, Page AC, Weil E, Croquer A, Vargas-Angel B, Jordan-Garza AG, Jordan-Dahlgren E, Raymundo L, Harvell CD (2012) Global coral disease prevalence associated with sea temperature anomalies and local factors. Dis Aquat Org 100(3):249–261. doi:10.3354/dao02488
Rypien KL, Ward JR, Azam F (2010) Antagonistic interactions among coral-associated bacteria. Environ Microbiol 12(1):28–39. doi:10.1111/j.1462-2920.2009.02027.x
Safavi-Hemami H, Young ND, Doyle J, Llewellyn L, Klueter A (2010) Characterisation of nitric oxide synthase in three cnidarian-dinoflagellate symbioses. PLoS One 5(4):e10379. doi:10.1371/journal.pone.0010379
Salih A, Larkum A, Cox G, Kuhl M, Hoegh-Guldberg O (2000) Fluorescent pigments in corals are photoprotective. Nature 408(6814):850–853. doi:10.1038/35048564
Santavy DL (1995) The diversity of microorganisms associated with marine invertebrates and their roles in the maintenance of ecosystems. In: Allsop D, Cowell RR, Hawsworth DL (eds) Microbial diversity and ecosystem function. CAB International, Wallingford, pp 211–229
Schwarz RS, Hodes-Villamar L, Fitzpatrick KA, Fain MG, Hughes AL, Cadavid LF (2007) A gene family of putative immune recognition molecules in the hydroid Hydractinia. Immunogenetics 59(3):233–246. doi:10.1007/s00251-006-0179-1
Schwarz JA, Brokstein PB, Voolstra C, Terry AY, Manohar CF, Miller DJ, Szmant AM, Coffroth MA, Medina M (2008a) Coral life history and symbiosis: functional genomic resources for two reef building Caribbean corals, Acropora palmata and Montastraea faveolata. BMC Genomics 9:97. doi:10.1186/1471-2164-9-97
Schwarz RS, Bosch TC, Cadavid LF (2008b) Evolution of polydom-like molecules: identification and characterization of cnidarian polydom (Cnpolydom) in the basal metazoan Hydractinia. Dev Comp Immunol 32(10):1192–1210. doi:10.1016/j.dci.2008.03.007
Shaked Y, Armoza-Zvuloni R (2013) Dynamics of hydrogen peroxide in a coral reef: sources and sinks. J Geophys Res Biogeosci 118(4):1793–1801. doi:10.1002/2013jg002483
Shapo JL, Moeller PD, Galloway SB (2007) Antimicrobial activity in the common seawhip, Leptogorgia virgulata (Cnidaria: Gorgonaceae). Comp Biochem Physiol B Biochem Mol Biol 148(1):65–73. doi:10.1016/j.cbpb.2007.04.019
Sherratt JA, Murray JD (1990) Models of epidermal wound healing. Proc Biol Sci 241(1300):29–36. doi:10.1098/rspb.1990.0061
Shick JM, Dykens JA (1985) Oxygen detoxification in algal-invertebrate symbioses from the great barrier-reef. Oecologia 66(1):33–41. doi:10.1007/Bf00378549
Shinzato C, Shoguchi E, Kawashima T, Hamada M, Hisata K, Tanaka M, Fujie M, Fujiwara M, Koyanagi R, Ikuta T, Fujiyama A, Miller DJ, Satoh N (2011) Using the Acropora digitifera genome to understand coral responses to environmental change. Nature 476(7360):320–U382. doi:10.1038/Nature10249
Shnit-Orland M, Kushmaro A (2009) Coral mucus-associated bacteria: a possible first line of defense. FEMS Microbiol Ecol 67(3):371–380. doi:10.1111/j.1574-6941.2008.00644.x
Sies H (2000) What is oxidative stress? In: Keaney JF Jr (ed) Oxidative stress and vascular disease. Springer, Norwell, pp 1–8. doi:10.1007/978-1-4615-4649-8_1
Slattery M (1999) Fungal pathogenesis of the sea fan Gorgonia ventalina: direct and indirect consequences. Chemoecology 9:97–104
Slattery M, Gochfeld DJ (2012) Chemically mediated competition and host–pathogen interactions among marine organisms. In: Fattorusso E, Gerwick WH, Taglialatela-Scafati O (eds) Handbook of marine natural products. Springer, Berlin, pp 823–859. doi:10.1007/978-90-481-3834-0_15
Slattery M, Hamann MT, McClintock JB, Perry TL, Puglisi MP, Yoshida WY (1997) Ecological roles for water-borne metabolites from Antarctic soft corals. Mar Ecol Prog Ser 161:133–144
Slattery M, Renegar DA, Gochfeld DJ (2013) Direct and indirect effects of a new disease of alcyonacean soft corals. Coral Reefs 32(3):879–889. doi:10.1007/s00338-013-1035-z
Spiegel S, Milstien S (2000) Sphingosine-1-phosphate: signaling inside and out. FEBS Lett 476:55–57
Sudek M, Work TM, Aeby GS, Davy SK (2012) Histological observations in the Hawaiian reef coral, Porites compressa, affected by Porites bleaching with tissue loss. J Invertebr Pathol 111(2):121–125. doi:10.1016/j.jip.2012.07.004
Sunagawa S, DeSantis TZ, Piceno YM, Brodie EL, DeSalvo MK, Voolstra CR, Weil E, Andersen GL, Medina M (2009a) Bacterial diversity and White Plague Disease-associated community changes in the Caribbean coral Montastraea faveolata. ISME J 3(5):512–521. doi:10.1038/ismej.2008.131
Sunagawa S, Wilson EC, Thaler M, Smith ML, Caruso C, Pringle JR, Weis VM, Medina M, Schwarz JA (2009b) Generation and analysis of transcriptomic resources for a model system on the rise: the sea anemone Aiptasia pallida and its dinoflagellate endosymbiont. BMC Genomics 10(1):258. doi:10.1186/1471-2164-10-258
Sussman M, Willis BL, Victor S, Bourne DG (2008) Coral pathogens identified for white syndrome (WS) epizootics in the Indo-Pacific. PLoS Biol 3(6):e2393. doi:10.1371/journal.pone.0002393
Tait K, Hutchison Z, Thompson FL, Munn CB (2010) Quorum sensing signal production and inhibition by coral-associated vibrios. Environ Microbiol Rep 2(1):145–150. doi:10.1111/j.1758-2229.2009.00122.x
Targett NM, Bishop SS, McConnell OJ, Yoder JA (1983) Antifouling agents against the benthic marine diatom, Navicula salinicola: homarine from the Gorgonians Leptogorgia virgulata and L. setacea and analogs. J Chem Ecol 9:817–829
Tarrant AM, Reitzel AM, Kwok CK, Jenny MJ (2014) Activation of the cnidarian oxidative stress response by ultraviolet radiation, polycyclic aromatic hydrocarbons and crude oil. J Exp Biol 217(Pt 9):1444–1453. doi:10.1242/jeb.093690
Theopold U, Schmidt O, Soderhall K, Dushay MS (2004) Coagulation in arthropods: defence, wound closure and healing. Trends Immunol 25(6):289–294. doi:10.1016/j.it.2004.03.004
Traylor-Knowles N, Granger BR, Lubinski TJ, Parikh JR, Garamszegi S, Xia Y, Marto JA, Kaufman L, Finnerty JR (2011) Production of a reference transcriptome and transcriptomic database (PocilloporaBase) for the cauliflower coral, Pocillopora damicornis. BMC Genomics 12:585. doi:10.1186/1471-2164-12-585
Van Alstyne KL, Schupp P, Slattery M (2006) The distribution of dimethylsulfoniopropionate in tropical Pacific coral reef invertebrates. Coral Reefs 25(3):321–327. doi:10.1007/s00338-006-0114-9
Vargas-Angel B, Peters EC, Kramarsky-Winter E, Gilliam DS, Dodge RE (2007) Cellular reactions to sedimentation and temperature stress in the Caribbean coral Montastraea cavernosa. J Invertebr Pathol 95(2):140–145. doi:10.1016/j.jip.2007.01.003
Vidal-Dupiol J, Adjeroud M, Roger E, Foure L, Duval D, Mone Y, Ferrier-Pages C, Tambutte E, Tambutte S, Zoccola D, Allemand D, Mitta G (2009) Coral bleaching under thermal stress: putative involvement of host/symbiont recognition mechanisms. BMC Physiol 9:14. doi:10.1186/1472-6793-9-14
Vidal-Dupiol J, Ladriere O, Destoumieux-Garzon D, Sautiere PE, Meistertzheim AL, Tambutte E, Tambutte S, Duval D, Foure L, Adjeroud M, Mitta G (2011) Innate immune responses of a scleractinian coral to vibriosis. J Biol Chem 286(25):22688–22698. doi:10.1074/jbc.M110.216358
Vidal-Dupiol J, Dheilly NM, Rondon R, Grunau C, Cosseau C, Smith KM, Freitag M, Adjeroud M, Mitta G (2014) Thermal stress triggers broad Pocillopora damicornis transcriptomic remodeling, while Vibrio coralliilyticus infection induces a more targeted immuno-suppression response. PLoS One 9(9):e107672. doi:10.1371/journal.pone.0107672
Weiss Y, Foret S, Hayward DC, Ainsworth T, King R, Ball EE, Miller DJ (2013) The acute transcriptional response of the coral Acropora millepora to immune challenge: expression of GiMAP/IAN genes links the innate immune responses of corals with those of mammals and plants. BMC Genomics 14:400. doi:10.1186/1471-2164-14-400
Wenger Y, Buzgariu W, Reiter S, Galliot B (2014) Injury-induced immune responses in Hydra. Semin Immunol 26(4):277–294. doi:10.1016/j.smim.2014.06.004
Williams GP, Babu S, Ravikumar S, Kathiresan K, Prathap SA, Chinnapparaj S, Marian MP, Alikhan SL (2007) Antimicrobial activity of tissue and associated bacteria from benthic sea anemone Stichodactyla haddoni against microbial pathogens. J Environ Biol 28(4):789–793
Wolenski FS, Garbati MR, Lubinski TJ, Traylor-Knowles N, Dresselhaus E, Stefanik DJ, Goucher H, Finnerty JR, Gilmore TD (2011) Characterization of the core elements of the NF-κB signaling pathway of the sea anemone Nematostella vectensis. Mol Cell Biol 31(5):1076–1087. doi:10.1128/MCB.00927-10
Wood-Charlson EM, Weis VM (2009) The diversity of C-type lectins in the genome of a basal metazoan, Nematostella vectensis. Dev Comp Immunol 33(8):881–889. doi:10.1016/j.dci.2009.01.008
Wood-Charlson EM, Hollingsworth LL, Krupp DA, Weis VM (2006) Lectin/glycan interactions play a role in recognition in a coral/dinoflagellate symbiosis. Cell Microbiol 8(12):1985–1993. doi:10.1111/j.1462-5822.2006.00765.x
Yu L, Wan F, Dutta S, Welsh S, Liu Z, Freundt E, Baehrecke EH, Lenardo M (2006) Autophagic programmed cell death by selective catalase degradation. Proc Natl Acad Sci U S A 103(13):4952–4957. doi:10.1073/pnas.0511288103
Zaragoza WJ, Krediet CJ, Meyer JL, Canas G, Ritchie KB, Teplitski M (2014) Outcomes of infections of sea anemone Aiptasia pallida with Vibrio spp. pathogenic to corals. Microb Ecol 68(2):388–396. doi:10.1007/s00248-014-0397-2
Zasloff M (2002) Antimicrobial peptides of multicellular organisms. Nature 415(6870):389–395. doi:10.1038/415389a
Zimmer BL, May AL, Bhedi CD, Dearth SP, Prevatte CW, Pratte Z, Campagna SR, Richardson LL (2014) Quorum sensing signal production and microbial interactions in a polymicrobial disease of corals and the coral surface mucopolysaccharide layer. PLoS One 9(9):e108541. doi:10.1371/journal.pone.0108541
Zmasek CM, Zhang Q, Ye Y, Godzik A (2007) Surprising complexity of the ancestral apoptosis network. Genome Biol 8(10):R226. doi:10.1186/gb-2007-8-10-r226
Acknowledgments
The authors would like to acknowledge funding from the National Science Foundation (awards IOS 1017458 to LDM, OCE-PRF 1225163 to JHPC, and OCE 0961384 to DJG), and from the National Oceanic and Atmospheric Administration’s National Institute for Undersea Science and Technology (award NA16RU1496 to DJG).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Mydlarz, L.D., Fuess, L., Mann, W., Pinzón, J.H., Gochfeld, D.J. (2016). Cnidarian Immunity: From Genomes to Phenomes. In: Goffredo, S., Dubinsky, Z. (eds) The Cnidaria, Past, Present and Future. Springer, Cham. https://doi.org/10.1007/978-3-319-31305-4_28
Download citation
DOI: https://doi.org/10.1007/978-3-319-31305-4_28
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-31303-0
Online ISBN: 978-3-319-31305-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)