
Abstract
Cassava (Manihot esculenta Crantz), a ”miracle of the tropics,” is a critical component of the approaches to alleviate poverty, hunger, and malnutrition and increase livelihood security. Its high inherent photosynthetic efficiency and ability to sustain growth in challenging environments make it a potential food and nutrition security crop. However, water remains the most limiting factor for future cassava production, particularly under anticipated climatic variability. Though cassava is popularized as a drought-tolerant crop, seasonal or intermittent water stress episodes affect cassava productivity by influencing plant growth, storage root yield, and quality. Successful cassava production in drought-prone areas relies on the development of drought-tolerant cultivars along with tailored agronomic practices. We reviewed multi-faceted responses from morphological level to tissue/cell level biochemical changes, root development responses, and storage root quality alterations occurring under drought and potential targets for the future breeding program. This knowledge will pave the way for developing breeding strategies and implementable agronomic methods.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Environmental changes associated with global warming are potential threats to global food demand (FAO 2017) as the world population is projected to increase by 9–12.3 billion from 2050 to 2100 (Gerland et al. 2014; Frona et al. 2019). Furthermore, the increased drought and heatwave frequency (Naumann et al. 2018) will likely aggravate the anticipated limited land and water resource availability (Arora 2019). Moreover, the projected decline in productivity of major food crops will have severe implications for rural and semi-urban global populations in developing countries (Leng and Hall 2019; FAO 2020, 2021). Cassava, a staple for one billion people worldwide (Lebot 2020), thrives in challenging environments where other major food crops struggled, making it an ideal climate-resilient crop (Pushpalatha and Byju 2020). Cassava is generally grown by marginal and smallholder farmers, but its perception as a “poor man’s food” (FAO 2014) and “food-feed-fuel” crop (Howeler et al. 2013) has recently changed to a “future smart/climate-resilient crop” (Rosenthal and Ort 2012; Mukherjee et al. 2019; Pushpalatha and Byju 2020), “crop of industrial significance,” and “twenty-first century crop” (Howeler et al. 2013). It has immediate benefits against hunger, poverty, and malnutrition (Anikwe and Ikenganyia 2018; FAO 2018; Amelework et al. 2021) due to its higher energy production efficiency than maize and wheat (Howeler et al. 2013; Nayar 2014). The recent production statistics by FAOSTAT (2022) indicates that its production has increased significantly since 1961, and the global cassava market will reach up to US$66.84 billion by 2026, exhibiting a compound annual growth rate (CAGR) of 6.50% (GlobeNewswire 2021), showcasing its economic robustness, and importance. Apart from the cassava’s storage root, leaves are also consumed as a valuable protein source (Latif and Muller 2015), and extractable starch from storage roots forms the basis for numerous essential industrial products (Ravi et al. 2021; More et al. 2021).
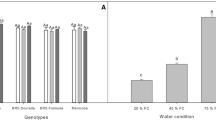
The average productivity of cassava is still far lower than its potential yield suggesting broad scope for genetic improvement and the development of agronomic practices to narrow yield gaps. Drought alone can lead to a notable reduction in storage root yield, though the quantum of reduction depends on genotype and the severity and timing of drought during the growing season (Fig. 1). Water deficit during early growth (1–3 MAP, months after planting) causes more yield losses as this period coincides with the critical yield attributes viz. root system development, canopy establishment, storage root initiation, multiplication, bulking, and starch accumulation (El-Sharkawy 2007; Duque and Setter 2019). The crop simulation model predicted the region-specific drought-induced yield reduction of cassava storage root ranging from − 3.7 to 17.5% across Africa (Jarvis et al. 2012), − 17.24 to − 21.26% in Thailand (Pipitpukdee et al. 2020) and − 10 to 12% in India (Pushpalatha et al. 2021) under various future climatic scenarios.

Quantum of yield loss in cassava subjected to early, mid, or terminal season drought stress
Additionally, drought alters storage roots’ physical and chemical properties hampering the remuneration potential for marginal cassava growers, end-product utilization, and consumption pattern (Santisopasri et al. 2001; Nhassico et al. 2008; Gleadow et al. 2016; Jyothi and Sajeev 2021). Understanding stage-specific crop responses and the underlying physiological process is a prerequisite to reducing the negative impacts of drought on cassava productivity (Zhu et al. 2020). There is a plethora of studies on drought stress in cereal (rice, wheat and maize), vegetables, fruits and other crops (Havrlentova et al. 2021; Giordano et al. 2021; Dietz et al. 2021). However, cassava remained neglected, and information on drought responses is limited. In the present review, we have revisited the detrimental effect of drought stress on the quality aspect of cassava and its impact on growth and yield. Moreover, this will help develop knowledge and understanding of cassava drought tolerance, which would help in the development of suitable traits tailored to region-specific cultivars and appropriate management practices to ameliorate the adverse effects of drought.
2 Cassava Water Relations Under Drought
Leaf water potential and relative water content are water stress indicators reflecting plant water availability (Alves and Setter 2004) as CO2 enters the leaf via stomata, the same channel as transpiration. Water availability controls the trade-off between CO2 absorption and water loss (Bhattacharya 2019).
The total water, free water, and the ratio of free water and bound water levels reduced in cassava leaves, while the bound water content increased with the intensification of drought (Shan et al. 2018). Reduced osmotic potential by osmotic adjustment under water stress facilitates turgor maintenance, stomatal regulation, CO2assimilation, and increased tolerance to dehydration (Alves and Setter 2004; Helal et al. 2013). Osmotic adjustment regulates turgidity so that plants can sustain low soil water potentials and create sufficiently low tissue water potential for continued soil water extraction. Under water stress, mature and expanding cassava leaves decreased osmotic potential to a relatively limited extent or had relatively modest osmotic potential, whereas immature leaves osmotically adjust to a greater extent than mature leaves (Ike and Thurtell 1981; Alves and Setter 2004). The majority (nearly 60%) of osmotic adjustment (OA) occurs as a result of the accumulation of K+salts in mature and expanding leaves, which is positively correlated with the extent of OA, while organic solutes such as sugars accounted for 25% of OA and their concentrations decreased as water stress progressed (Alves and Setter 2004; Okogbenin et al. 2013). Osmotic adjustment helps to increase water use efficiency and the water resupply system from stem to leaves during drought (Itani et al. 1999). However, OA alone is insufficient to provide drought tolerance and higher yields (Helal et al. 2013).
Water use efficiency [WUE; agronomic (yield produced per unit of water applied) or physiological, i.e., instantaneous WUE (A/E) and intrinsic WUE (A/gs)] is important for analyzing plant growth under water-deficit conditions (Hatfield and Dold 2019). In cassava, the WUE is highly dependent on water supply, stomatal conductance, photosynthetic efficiency, transpirational losses, growth stage, and atmospheric condition (El-Sharkawy 2006). Interestingly, cassava crop growth rate and short-term WUE are similar to C4 crops, which are considerably higher than other C3crops (El-Sharkawy 2004, 2006). However, optimizing WUE under water-limited environments would also enhance cassava’s productivity in challenging environments (Wongnoi et al. 2020).
3 Growth and Morphological Responses
3.1 Leaf and Shoot Growth Dynamics
Cassava leaves can take 10–12 days for emergence to full expansion (Cock 1984), with a leaf life of up to 30–180 days, depending on the cultivar, water availability, shade level, temperature, and growing conditions (Phosaengsri et al. 2019). Maximum leaf formation is completed within the first 90 DAP, followed by steady canopy development throughout the growing period (Alves and Setter 2004). Cassava canopy area increases significantly from 3 to 5 MAP, followed by a progressive decline in total canopy area during 8–9 MAP due to leaf loss (El‐Sharkawy and De Tafur 2007). The canopy development phase (3–5 MAP) is critical as storage root initiation, and bulking co-occurs, significantly affecting the storage root yield (Ceballos et al. 2012). Accumulation of optimum canopy with desirable leaf growth during the storage root bulking phase is critical to achieving higher productivity irrespective of the water status of the location (Phosaengsri et al. 2019). Prolonged water deprivation harmed leaf development, leaf cell division, leaf angle, leaf expansion, leaf retention capacity, and leaf area index (LAI), and leaf abscission was accelerated (de Oliviera et al. 2017; More et al. 2020) (Fig.2). According to Connor and Cock (1981), leaf expansion/leaf size was more sensitive to water deficit than the leaf production rate. Thus, leaf area is the key determinant factor for canopy development and productivity under water stress, not the leaf loss. Water stress within 2–3 MAP produced half the leaf area of non-stressed plants (Turyagyenda et al. 2013a, b) and caused 84.27% mortality (Turyagyenda et al. 2013a). Furthermore, diminished leaf area was associated with ethylene accumulation (Ogaddee and Girdthai 2019), which might be related to accelerated leaf senescence and ROS generation (Liao et al. 2016). Reduced leaf area is the first line of defense, which reduces the transpirational loss but substantially affects the photosynthetic efficiency (Pacheco et al. 2019), with improved water use efficiency (Vandegeer et al. 2013).

Cassava plants under drought show reduced canopy development. The most notable change in the leaf trait is the reduction in leaf area in stressed plants as compared to the leaf area of well-irrigated plants (A). In order to regulate the use of limited available water, cassava leaves are known to employ heliotropic effect to reduce the harmful effect of radiation caused by drought-induced high-temperature stress (B). While leaves continue to develop, they are smaller and grow more slowly, and the older leaves are lost (C). Cassava demonstrated remarkable post drought recovery. The newly developed leaves from the previously stressed plants had similar leaf area and photosynthetic efficiency as that of well-irrigated plants (D)
Plant height, stem diameter, and shoot biomass were decreased significantly, varying with the different genotypes, in response to water stress (Aina et al. 2007; Okogbenin et al. 2013; Shan et al. 2018), owing to the downregulation of proteins and photosynthetic activity resulting into impaired cell development and division (Shan et al. 2018) and overall plant vigor. Tissue turgidity is maintained to a minimum to prevent cell death by limiting cell growth (Calatayud et al. 2000). Inhibited apical meristem cell development and elongation, loss of turgor, significant leaf fall and senescence, and altered biomass partitioning contributed to decreased shoot growth (Helal et al. 2013; Shan et al. 2018; More et al. 2020). This information on leaf and stem is essential to understand response mechanism that cassava showcases under drought.
3.2 Root Growth and Development
The formation of roots is critical for the growth and productivity of cassava shoots (Duque and Villordon 2019). Investigating root development kinetics in resource-constrained production environments may provide additional insight into cassava crops’ drought-tolerance mechanisms (Adu et al. 2018). However, relatively few studies have identified the role of root growth and development in root and tuber crops, including cassava, compared to aboveground morphophysiological traits (Adu et al. 2018; Duque and Villordon 2019), particularly under conditions of water stress (Adu et al. 2020). Root emergence occurs 1 to 2 weeks after planting, followed by significant fibrous and lateral root development (Izumi et al. 1999). Early water stress coincides with the period of highest relative growth rate of root system during the first 30 DAP, which subsequently decreases to a nearly constant rate after 30–60 DAP (Adu et al. 2018) and becomes more detrimental to crop yield. The fibrous root system grows up to 1.4–2 m in depth to increase access to deeper water (de Tafur et al. 1997), or even up to 2.4–2.6 m Ekanayake et al. 1994, 1996) depending upon the timing of the water stress, and soil texture also significantly affected due to water stress (Adu et al. 2018). Aina et al. (2007) reported that drought reduced root growth more than shoot growth in cassava, depending on the timing and severity of the drought. Strikingly, reduced lateral and total root numbers; root length and diameter; adventitious root number and length; basal root branching density; lower and upper nodal root numbers; first, second, and third lateral root numbers; and root dry weight as a result of water stress cause sizeable reduction in storage root yield (Pardales and Yamauchi 2003; Helal et al. 2013; de Oliveira et al. 2015; Kengkanna et al. 2019; Adu 2020). Developing a cassava root phenotyping protocol and platform and identifying phenotypic variation in root systems is essential for developing highly drought-tolerant cassava genotypes. Recently developed root shovelomics-based phenotyping techniques combining manual and semi-automatic DIRT measurements (Kengkanna et al. 2019) or easy and low-cost three-dimensional cassava root crowns phenotyping platform (Sunvittayakul et al. 2022) could be proven extremely useful for root architectural trait phenotyping in cassava under various growing conditions. Moreover, a substantial amount of germplasm maintained at the International Center for Tropical Agriculture (CIAT), Colombia, Central Tuber Crops Research Institute (CTCRI), India, and other premier research institutes could be assessed in collaborative breeding program for identification of root-related traits for drought-tolerance.
4 Physiological-Biochemical Responses
4.1 Photosynthesis
Cassava is a biologically efficient crop with a higher leaf photosynthetic rate, as high as 40–50 μmol CO2 m−2 s−1under favorable environments (El-Sharkawy 2007). In a series of experiments, aboveground dry weight and storage root yield were positively correlated with the single-leaf photosynthesis or mean seasonal upper canopy photosynthesis (El-Sharkawy et al. 1990; El-Sharkawy 2006). However, this correlation depends on optimum PAR, plant age, leaf position, LAI, leaf duration, leaf anatomy, and the environment under which the leaves were formed (El-Sharkawy 2014; De Souza and Long 2018).
Inhibited CO2assimilation rate under water stress was primarily attributed to reduced leaf area, reduced stomatal conductance (Adjebeng-Danquah et al. 2016; Morgante et al. 2020), and reduced photosynthesis-related proteins (Chang et al. 2019; Wang et al. 2021a, b). The sensitivity of cassava’s stomata to varying water regimes and atmospheric humidity has been reviewed in detail (El-Sharkawy 2007). The presence of higher stomatal density on the abaxial side than on the adaxial side of the leaf also reduced gaseous exchange under water stress conditions due to leaf bending/rolling (El-Sharkawy and Cock 1987). Asymmetric stomatal density reduced gas exchange which optimized water use but impaired carbon gain. This is further affected due to restricted flow of water from leaf veins to guard cells (Ooba and Takahashi 2003). Hydro active stomatal closure (referred to as stomatal closure due to localized dehydration of guard cells and accessory cells as per the relative change in air humidity) was induced by the upregulation of ABA-dependent (ABA-D) and ABA-independent genes associated with decreased stomatal activity (El-Sharkawy 2006; Orek et al. 2020; Fu et al. 2016; Li et al. 2017a, b). Carbon isotope ratios (Δ13C), a key parameter to assess water use efficiency in relation to stomatal activity and gaseous exchange capacity remained more negative under water stress (Adjebeng-Danquah et al. 2016; More et al. 2019a, b2019, 2022). Drought-sensitive cultivars had lower photosynthetic quantum efficiency (Fv/Fm) than drought-tolerant cultivars (Zhao et al. 2015). The quantum yield of the light-adapted leaf (F′v/F′m), effective quantum yield (ΦPSII), photochemical quenching coefficient (qP), and ETR were decreased significantly under water stress. Efficiency of photosystem II and electron transport rate was reduced significantly as a result of water stress (Pereira et al. 2018), however, contrary to these findings, Shan et al. (2018) reported increased PS II and PS I subunits to modulate the photosystem repair and electron transfer to maintain photosynthetic efficiency to reduce the negative effect of water deficiency (Shan et al. 2018). These changes are ascribed to damage to the chlorophyll membrane, chlorophyll degradation, and associated photoinhibition (Morgante et al. 2020); downregulation of PSII synthesis; decreased oxygen-evolving complex protein production; and structural changes in oxygen-evolving complex proteins (Dalal and Tripathy 2018). Additionally, photosynthetic activity was harmed by ROS generation, decreased RuBisCO activity, and damage to multiple cellular components (Zhu et al. 2020), as well as downregulation of sedoheptulose-1,7-bisphosphatase (SBPase) and fructose-bisphosphate aldolase (both of which are required for the supply of RuBP to Rubisco) (Shan et al. 2018).
Reduced stomatal activity and carbon assimilation and the reduction in leaf area were the strategic responses to optimize the water use efficiency (Duque and Setter 2013; Zhao et al. 2015; Adjebeng-Danquah et al. 2016). Consequently, high water use efficiency will not always result in high yield, as it may be the result of reduced water consumption due to leaf abscission and consequently less CO2fixation (Blum 2009). A delicate balance between transpiration and CO2diffusion is crucial for optimizing water use efficiency and productivity under drought conditions in cassava. Evapotranspiration increases with a simultaneously higher stomatal conductance (Oguntunde 2005; El-Sharkawy 2012; Shan et al. 2018; Morgante et al. 2020). Transpiration also declined abruptly in conjunction with leaf ABA accumulation and severe leaf abscission (Duque and Setter 2013). Genotypic variation for gaseous exchange under varied ecosystems exists (Shan et al. 2018; More et al. 2020), which could be useful in fine-tuning the dual function of the stomatal aperture. More research is warranted to decipher biochemical and molecular characteristics of cassava photosynthesis in relation to storage root yield.
4.2 Assimilate Partitioning
Storage root yield of cassava is a function of source-sink interactions and the photosynthetic efficiency (Orek et al. 2020). It is severely impeded by water stress episodes (El-Sharkawy 2014) (Fig.3). Partitioning index (PI), a ratio of storage root yield to total biomass (aboveground biomass + storage root weight + fibrous root weight) determined at 4-6 MAP (Duque and Setter 2019), is an indicator of biomass accumulation efficiency of storage roots. Duque (2012) had established profuse relationship especially under water stress among PI and HI, for 45 diverse cassava genotypes. PI at 7 MAP and HI at 12 MAP (at harvesting) were significantly and positively correlated for drought-tolerant genotypes (Olasanmi 2010). Duque and Setter (2019) also suggested that root yield is the function of PI, based on observed correlation patterns. Water stress occurred during the 3–4 MAP period, significantly decreased PI, biomass, and yield. Assessment of PI at an early stage could have additional benefits as an indicator of higher storage root yield. Significantly less attention has been given to this particular trait in cassava, and further studies are sought to determine the pattern and quantum to which cassava alters carbohydrate accumulation and pattern of its diversion among different organs to sustain its partitioning capacity to sustain yield.

Tuberization at 5 and 10 months (harvesting) after planting in a variety H 165 subjected to drought from 61 to150 days after planting
4.3 Biochemical Changes
Drought episodes triggered the biosynthesis of protective biochemicals and secondary metabolites in cassava to counteract the water-deficit stress (Nuwamanya et al. 2014). Most changes were associated with the increases in the synthesis of defensive proteins (e.g., lectins and protease inhibitors) and other biomolecules like phenolics, proline, and tannins (Isah 2019). Cassava leaves accumulated significantly higher quantities of trehalose and proline during early drought stress (Ren et al. 2017). Sugar content in stems and immature/mature leaves decreased as water stress progressed (Duque and Setter 2013; Koundinya et al. 2018). Duque and Setter (2013) reported that while evaluating the composition of nonstructural carbohydrates in cassava organs under water stress, sugar was the primary form of nonstructural carbohydrate in the leaf blade and leaf petiole, while starch comprised approximately one-third. Contrastingly, stem and storage roots contained relatively high starch content. The stem’s starch reserves were remobilized in numerous organs to maintain development via storable carbohydrates depleted during periods of water stress due to reduced canopy photosynthesis and the generation of secondary metabolites and defense proteins (Duque and Setter 2013). However, they remained unaffected in tolerant and susceptible genotypes under water stress (Duque and Setter 2019). The accumulation of total reducing sugars (glucose) was negatively correlated with starch yield and positively correlated with protein and phenolic contents (Nuwamanya et al. 2014). Reverse translocation of sugars from storage roots occurred to cater for leaf and stem growth. Bound reducing sugars increased more than free reducing sugars with increasing water stress, suggesting possible losses in storage starch. Understanding biochemical responses is essential for a holistic perception of drought resistance mechanism that cassava adopts under drought
Reactive oxygen species (ROS) are generated during water-deficit stress, causing oxidative damage. ROSs are responsible for the negative impact on physiological processes leading to programmed cell death (PCD). Under oxidative stress, relative leaf electrical conductivity and malondialdehyde and hydrogen peroxide contents significantly increased with increasing drought stress (Petrov et al. 2015). Enzymatic (superoxide dismutase, ascorbate peroxidase, peroxidase, glutathione reductase, catalase) and non-enzymatic (ascorbic acid, glutathione, total phenolics, and total flavonoids) antioxidant mechanisms form to counteract the oxidative damage occurring under water stress (Zhu et al. 2020). Almost all antioxidants enhanced manifold under water stress, depending on the genotype. Some cultivars accumulated more ascorbic acid, glutathione, and superoxide dismutase triggered by the upregulation of Mn-SOD and CAT genes, while others synthesized more glutathione reductase and total phenols due to the upregulation of the GR gene (Zhu et al. 2020). Furthermore, drought abruptly affected the thylakoid membrane and grana stacking, and grana lamellae structure significantly affecting biochemical and physiological processes, whereas no change was observed in the thylakoid granum structure (Chang et al. 2019). Hence, it is needed to explore the role of chloroplast in drought resistance and adaptation in cassava.
Cassava’s complex multi-level drought responses involve employment of the antioxidant system (Yan et al. 2021). Proteomic analysis is a useful technique to reveal the role of genes associated with a specific protein involved with drought responses and adaptation mechanisms (Aslam et al. 2017). However, information regarding this aspect in cassava is very limited (Yan et al. 2021). In a Kyoto Encyclopedia of Genes and Genomes (KEGG)-based proteomics analysis in cassava, Wang et al. (2021a, b) reported distinct drought response and adaptation mechanisms in leaf vasculature and leaf mesophyll, respectively. Genes associated with amino acid biosynthesis (e.g., alanine, aspartic acid, and glutamic acid), pyruvate metabolism, and starch and sucrose metabolism were upregulated in leaf mesophyll, while genes associated with carbon metabolism, plant hormone signal transduction, pyrophosphatase activity, and oxidative phosphorylation were upregulated in leaf vasculature. In addition, the increased arginine biosynthesis in leaf mesophyll tissue was associated with enhanced adaptation to water deficiency. Shan et al. (2018), under long-term drought conditions, reported upregulation of 262 proteins potentially involved in carbohydrate energy metabolism, heat shock proteins, protein homeostasis, transcription, cell structure, cell membrane transport, signal transduction, and stress responses. Numerous reports depict the role of 90 kDa heat shock protein (HSP90, highly conserved molecular chaperone) as an aid in coping with multiple biotic and abiotic stresses in plants and animals. However, its role in cassava regarding drought resistance is not yet studied in detail (Wei et al. 2020). MeHSP90 proteins were identified as a critical factor for enhanced stress resistance. Out of 10 identified MeHSP90s, the MeHSP90.9 was abundantly induced as a result of drought stress. Apart from these, two potential HSP90 client proteins, viz., MeWRKY20, and MeCatalase1 interacted significantly with MeHSP90.9 to foster stress resistance. MeHSP90.9 proteins regulated the accumulation of ABA hormone through targeting the ABA biosynthesis gene MeNCED5 whereas along with MeCatalase1, it regulated the catalase activity and H2O2accumulation to facilitate cassava plants to cope up with drought. Li et al. (2021) characterized the functions of drought-responsive genes such asSQUAMOSA promoter binding protein-like 9 (SPL9) and MeSPL9. MeSPL9 and rMeSPL9-SRDXconferred drought tolerance with enhanced accumulation of proline, anthocyanin, jasmonic acid (JA), and soluble sugars. Chang et al. (2019) elucidated photosynthesis-related proteins’ downregulation, significantly affecting photosynthesis efficiency. However, the cassava plant could cope with the drought situation with the upregulation of carbon and nitrogen metabolism–associated proteins. In the very first report of integration of transcriptomic and proteomic analysis in cassava, Ding et al. (2019) reported 237 and 307 differentially expressed proteins (DEPs), in cassava leaf and root, enabling heat shock protein, secondary metabolism biosynthesis, and hormone biosynthesis to sustain under water-deficient environments.MeMYB1, MeMYB2, MeMYB4, and MeMYB9transcription factors belonging to the myeloblastosis (MYB) superfamily were involved in stress response mechanisms in cassava (Ruan et al. 2017). RNAi-driven repression ofMeMYB2resulted in drought and cold tolerance in transgenic cassava (Ruan et al. 2017). Wang et al. (2021a, b) reported that the MYB family proteinMeMYB26, a typical transcription factor with two MYB DNA-binding and transcriptional activation domains, was exclusively involved in water stress signal transduction in the root portion of cassava and crucial for plant stress resistance, growth, and biomass development. MeMYB transcription factors were differentially expressed in leaves upon exposure to cold and drought stress and thus could be important for signal transduction of abiotic stress responses (Ruan et al. 2017; Wang et al. 2021a, b). However, more experiments are needed to re-confirm these findings.
ABA, a stress hormone, plays a vital role in controlling the leaf development rate to reduce water losses. ABA accumulation is a part of the regulatory system to cope with water deficit (Alves and Setter 2000). ABA restricted stomatal activity, cell growth cycle and division, shoot and root growth, and leaf area growth to manage limited water availability under water stress (Alves and Setter 2004; Duque and Setter 2013; Felemban et al. 2019). Under water stress, young and mature leaves accumulate ABA. Strikingly, young leaves doubled the ABA content on a leaf area basis and always had higher ABA concentrations than mature leaves (Alves and Setter 2004; Duque and Setter 2013) because of their lesser metabolic breakdown capacity and translocation of ABA from mature leaves (Alves and Setter 2004). In addition to ABA, ethylene accumulation also increased due to drought stress, leading to the senescence of older leaves (Liao et al. 2016; Ogaddee and Girdthai 2019). Over a period of time, ABA has been referred to as principal drought-responsive hormone, but there are several drought-responsive hormones, namely, ethylene, jasmonic acid, and salicylic acid whose role under drought in cassava remained neglected.
5 Molecular Responses
Deciphering the molecular drought responses is the essential component for designing a breeding program to develop drought-tolerant crop species. Drought response in cassava involves the complex multi-faceted mechanisms with upregulation or downregulation of multiple genes and proteins (Muiruri et al. 2021). Upregulation of ABA-D genes and the downregulation of a large number of genes were observed during post-stress withdrawal. ABA-D genes regulate stress-related responses by mediating stomatal activity and ABA accumulation in cassava leaves (Orek et al. 2020). In addition to this, water stress induced ABA biosynthesis genes (NCED and ABA1) and ABA signaling elements (PP2C, SnRK2, and ABF) in vascular cells of cassava leaves. Gene expression pattern analysis revealed that the leaf vascular bundle responded more actively than mesophyll cells (Wang et al. 2021a, b). Ruan et al. (2018) identified 18 cassava-specific CC-type GRXs across the cassava genome, of whichMeGRXC3, C4, C7, C14, C15, and C18were involved in ABA signaling. Recently, Ruan et al. (2022) revealed that overexpression ofMeGRXC3, a cassava-specific CC-type glutaredoxin essential for redox homeostasis and ROS signaling, induced stress‑related transcription factor genes in Arabidopsis. Ren et al. (2017) employed bioinformatics approaches to identify and characterize candidateManihot esculenta ethylene receptor genes and transcription factor genes. The highest and the lowest expression of these genes was in the leaf and tuberous roots, respectively, subjected to drought stress through the ethylene signaling pathway.
Shang et al. (2021) elucidated the tissue-specific SOD, CAT, and APX gene expression consequent to osmotic stress, postharvest physiological deterioration (PPD) of storage roots, ABA, and Xanthomonas axonopodis infection. Seven SODs, 6 CATs, and 6 APX genes were identified in this research, of which SODsshowed high expression levels. Long non-coding RNAs (lncRNAs), important drought response regulators, were reported in autotetraploid cassava which conferred drought tolerance by increasing the stomatal density (Li et al. 2017b; Xiao et al. 2019). Li et al. (2017b) identified 318 long non-coding RNAs associated with biosynthesis of secondary metabolite, transduction of hormone signals, and sucrose metabolism in cassava. Yu et al. (2016) isolated stress-inducible homologous cassava plasma membrane gene,MePMP3-2, for the first time from cassava. The expression of this gene was upregulated under NaCl and PEG-mediated water stress suggesting its role in triggering stress responses. According to Yan et al. (2021), transcription factorMeRAV5 interacts physically with MePOD (peroxidase) and MeCAD15(lignin-related cinnamyl alcohol dehydrogenase 15), to govern hydrogen peroxide and lignin accumulation, respectively, to enhance drought resistance. Astonishingly, this pathway was ABA independent as it had no significant effect on ABA content of the leaves. Recently, genetic mechanism and genomics research focusing on drought tolerance is reviewed in detail by Muiruri et al. (2021).
5.1 GWAS Approaches for Identifying Breeding Targets
Cassava hybridization is challenging due to its long-life cycle (Bull et al. 2017), asynchronized flowering and sterility or non-flowering (Pineda et al. 2020), and polygenic nature of abiotic stress tolerance (Koundinya and More 2021). Marker-assisted selection (MAS) and quantitative trait loci (QTL) have been used to analyze genetic linkage maps of cassava for drought tolerance (Setter and Fregene 2007; Turyagyenda et al. 2013a, b). For example, the KASPar SNP genetic map was used to explore productivity-associated traits in cassava exposed to moderate drought stress in Africa using 505 polymorphic SNP markers distributed across 21 linkage groups, with 27 QTL identified for 11 productivity traits from 267 F1progeny (Ewa et al. 2021). The QTLsc3loc84.0, c6loc0.0, and c7loc13.0 were associated with stable productivity under moderate stress. However, according to Jannink et al. (2010), MAS may not improve polygenic traits as it ignores genes with minor effects when selecting quantitative traits such as drought tolerance. Genome-wide association studies (GWAS) can better analyze genomic regions associated with water deficit using genome-wide marker data. In a GWAS, Feng et al. (2019) observed that the abscisic acid (ABA)-responsive element (ABRE)-binding factors (ABFs) gene regulated the expression of target genes under drought stress by binding to ABRE (abscisic-acid-response element). Manihot esculenta ABFs (MeABFs) regulated the expression of Manihot esculenta betaine aldehyde dehydrogenase (MeBADHs) genes by binding the MeBADH1 promoter to modulate the accumulation of glycine betaine (GB) in cassava leaves to cope against drought stress. dos Santos Silva et al. (2021) identified 62 single nucleotide polymorphisms (SNPs) across 18 cassava chromosomes associated with drought-responsive-tolerance protein synthesis, including APETALA 2 domain (AP2), photosystem II oxygen-evolving enhancer protein, PR5-like receptor kinase-related, beta-fructofuranosidase/saccharase, leucine zipper, and bZIP transcription factors. Furthermore, numerous transcription factors associated with abiotic stresses identified in cassava, such as filamentous temperature-sensitive protein Z (Tubulin/FtsZ) and Manihot esculenta Crantz, 23 tubulin genes (MeTubulins) having important roles in IAA and GA3stress-induced responses (Li et al. 2021). GWAS is one of the feasible approaches for rapid identification of new/target genes and demands development of high-throughput phenotyping tool for GWAS in cassava to identify novel genes conferring drought tolerance.
5.2 Key genes for Transgenic Approaches
Molecular characterization of drought-responsive and tolerant genes is needed to improve drought tolerance in cassava. Researchers are keen to develop transgenic cassava plants with greater drought tolerance. Possible targets include the genes underlying the major QTL discussed previously, but other genes can also be considered. Cassava clone TMS 60444 was transformed with isopentenyl transferase (ipt) gene conferring drought tolerance via enhanced cytokinin production under drought (Zhang et al. 2010). Turyagenda et al. (2013) identified potential drought-tolerant candidate genes to develop transgenic cassava regulating oxidative stress by moderating reactive oxygen species (MeMSD and MeALDH) and osmotic adjustment (MeZFP and MeRD28). Application of PEG and ABA treatments significantly induced the expression of MeDREB1A, responsible for enhanced abiotic stress tolerance in transgenic Arabidopsisand cassava plants (An et al. 2017). Expression of theaquaporin (AQP) gene conferring drought tolerance has been characterized in cassava (Putpeerawit et al. 2017; Wahyuni et al. 2020). Wahyuni et al. (2020) observed differential expression of theAQP gene in three cassava varieties subjected to drought stress. Genome-wide analysis of the aquaporin gene family in cassava revealed the availability of many aquaporin isoforms. Expression analysis of the MeAQPs gene family under water stress confirmed the upregulation of MePIP2-1 and MePIP2-10. AQPgene enhanced drought tolerance through AQ PIP2-mediated greater water permeabilities in cellular membranes, influencing water efflux in guard cells to regulate stomatal activities (Putpeerawit et al. 2017). A recent study identified a novel gene,lncRNA, DROUGHT-INDUCED INTERGENIC lncRNA (DIR), enhancing proline accumulation and drought tolerance in transgenic cassava (Dong et al. 2022). In another study,MeHSP90.9 promoted the transcriptional activation of MeWRKY20 associated with ABA biosynthesis and the activation of MeCatalase1, conferring drought tolerance in cassava (Wei et al. 2020). According to Xu et al. (2013), transgenic cassava cultivars with increased expression of cytosolicMeCu/ZnSoD and peroxisomal MeCat1had greater drought tolerance than non-transgenic plants due to increased water content, proline content, superoxide dismutase, and catalase. Ethylene response factor family genes are involved in abiotic stress tolerance, inducing ethylene-mediated responses, and have been identified and characterized in cassava (Fan et al. 2016). Drought-stressed cassava had upregulated expression of TCP transcription factors (Lei et al. 2017), mitogen-activated protein kinase kinase kinases (MAPKKKs) genes (Ye et al. 2017), and the late embryogenesis abundant protein family (Wu et al. 2018), instrumental in stress responses through the ABA signaling pathway. In addition, calcium (Ca2+) sensors such as calmodulins (CaMs), calmodulin-like proteins (CMLs), and calcineurin B-like proteins (CBLs) are crucial for calcium-based abiotic stress sensing (Wei et al. 2018), and KT/HAK/KUP family genes crucial for potassium ion (K+) transport during stress (Ou et al. 2018) have been identified in cassava and could be used in breeding programs. Furthermore, Li et al. (2022) recently reviewed drought- and cold-responsive and tolerance genes in cassava. A summary of selected genomic studies on drought tolerance in cassava is presented in Table1.
6 Storage Root Quality
6.1 Physicochemical Properties
Cassava storage roots are a huge reservoir of starchy carbohydrates. In its original or modified form, starch is used in various industries due to its functional and chemical properties (Jyothi and Sajeev 2021). Starch accumulation in storage root initiates 4–6 weeks after planting (Cock 1984; El-Sharkawy 2004) and continues even when the root growth rate has declined until the harvest (Sriroth et al. 2001). Though the accumulation capacity of roots depends on the inherent varietal behavior, the reduced starch yield and quality are the likely consequence of water stress (Connor et al. 1981; Santisopasri et al. 2001). After prolonged water stress, water availability restores plant growth and starch synthesis (regardless of the stage of water stress, i.e., early/mid or terminal stress), but the effect on starch quality is compromised and cannot compare to the starch quality of plants supplemented with sufficient irrigation (Santisopasri et al. 2001; Sriroth et al. 2001). Starch physiochemical properties viz., hydration, gelatinization, and peak viscosity, were altered significantly under different water regimes (Teerawanichpan et al. 2008). Starch granules extracted from roots during early growth under normal moisture conditions had greater swelling capacity than those extracted from plants under moisture stress, which might be due to the moisture deficit significantly decreasing the granule size (Teerawanichpan et al. 2008) and influenced the paste viscosity with increased water absorption. Water stress had minimal influence on amylose: amylopectin contents (Defloor et al. 1998; Janket et al. 2020), though starch content (%) remained significantly lower in the plants exposed to water stress (Santisopasri et al. 2001). This reduction may help support aboveground growth by remobilization of starch under limited photosynthesis during water stress (Setter and Fregene 2007). Cassava starch grown under initial moisture stress has a considerably higher pasting temperature than cassava starch cultivated under no first moisture stress. In comparison, the peak viscosity of cassava starch extracted under early water stress was substantially lower than that extracted under normal conditions (Santisopasri et al. 2001). Water availability significantly changes the chloroplast structures. Chloroplasts from droughted plants were round in shape and further swollen with the progressive stress accumulating higher quantities of starch granules. On the other hand, chloroplast determined from the control plants was typically fusiform. Formation of higher number of starch granules might help plant to generate extra energy to combat drought (Chang et al. 2019).
The effect of drought on the physiochemical properties of cassava starch is inevitable (Santisopasri et al. 2001). Industrial application of cassava starch is predominantly reliant upon quality (Odeku 2013). As the economy of many countries (West and Central Africa, China, Thailand, Brazil, and India) revolves around cassava starch, undesired changes in yield and quality will lead to reduction in farm income. Therefore, additional research is needed to determine the effect of water availability on the functional, physiochemical, and structural features of starch.
6.2 Anti-nutritional Factors
Cassava is one of over 2000 plant species, including fruits and vegetables, which contain cyanogenic glycosides (CG) that release cyanide upon hydrolysis as a defense mechanism against herbivores (Møller 2010). It interacts with stress hormones and initiates actions, allowing plants to become used to biotic and abiotic challenges (Siegien and Bogatek 2006). Moreover, CG served as a storage house for nitrogen, which could be used in nitrogen assimilation under moisture stress to support the nitrogen-demanding sites to sustain growth (Mtunguja et al. 2016). Growing conditions, location, varietal differences, edaphic stresses, nutrient supply, and water availability are the crucial determinants of hydrogen cyanide (HCN) production in cassava leaves and storage roots (Burns et al. 2012; Imakumbili et al. 2019). Under water stress, HCN accumulation in leaves increased by 1.8–2.7 times as compared to leaves under no stress (Imakumbili 2019; Hular-Bograd et al. 2011) and increased with drought intensity (Santisopasri et al. 2001). The HCN content in dried storage roots of droughted plants was twofold higher than in control plants (Vandegeer et al. 2013). Higher HCN contents in roots could be due to the mobilization of resources, including HCN, from growing young leaves and tuberous roots (Vandegeer et al. 2013). According to the hypothesis of Vedengeer et al. (2013), if HCN is constitutive in nature, then accumulation of HCN as a result of water-stress will persist even after availability of sufficient water. Contrary to this hypothesis, if accumulating HCN is labile in nature, then glucosides synthesized under drought will be of same quantity as that of non-droughted plants (Møller 2010; Vandegeer et al. 2013). Quantification of HCN accumulation in leaves and storage roots under drought is extremely important and cannot be neglected while developing drought-tolerant cassava variety. The morphological, physiological, and molecular responses of cassava subjected to drought stress and their effects are summarized in Figs. 4 and 5.

Morphological and physiological responses of cassava subjected to drought stress and their effect on storage root and processing quality

Physiological and biochemical responses of cassava subjected to drought
7 Water Stress Adaptation Strategies
Cassava exhibited several strategies ranging from morphophysiological-biochemical responses to changes at the molecular level to mitigate drought-related consequences (Table 2). These strategies enable the plant to escape the drought by modifying life cycle/phenology or by regulating physiological functions. Strategies like stomata closure, shedding leaves, and regulating root system architecture (Muiruri et al. 2021) facilitate the reduction in photosynthetic leaf area, transpiration losses, and optimize water use (Okogbenin et al. 2013; Wongnoi et al. 2020). These changes correspond with impaired mesophyll conductance significantly hampering photosynthetic efficiency to manage drought episodes (El-Sharkawy 2004). The leaves became smaller with the progression of drought coupled with leaf rolling and change in leaf angle to prevent light interception and photooxidative damage (Calatayud et al. 2000). Along with leaves, stem length and thickness are greatly reduced to balance canopy growth and storage root development under drought. Balanced above-belowground growth under drought is achieved by modulating utilization of starch reserves. Non-structural carbohydrate reserves from leaves and stem are likely to be diverted to support the storage root growth under drought. It can be a potential strategy against drought stress (Duque and Setter 2013).
The employment of antioxidant system induces drought tolerance in cassava by scavenging harmful ROS generated under drought condition (Xu et al. 2013; Xu et al. 2013; Liao et al. 2016). Drought-tolerant cultivars tend to have a lower oxidative response than drought-susceptible cultivars (Zhu et al. 2020). According to Nuwamanya et al. (2014), cultivars with enhanced nitrogen metabolism enabled higher photosynthetic efficiency under water tress. Substantial quantities of osmolytes and biochemical factors are accumulated to maintain the physiology of plants. Proline and glycine-betaine accumulation has been found in plants after stress episodes (Ren et al. 2017; Wang et al. 2021a, b). Glutathione and ascorbic acid accumulation act as a protectant against water-stress–induced oxidative stress. Cultivars with higher mineral, osmoprotectant, osmolyte, and ascorbic acid contents had a higher degree of drought tolerance (Okogbenin et al. 2013; Zhu et al. 2020). Higher quantities of superoxide dismutase, peroxidase, catalase, ascorbate peroxidase, glutathione reductase, and ascorbic acid are part of the plant-defensive system (Zhu et al. 2020). At the molecular-genomic level, the genes, transcriptome, and enzymes expressed in cassava under drought conditions have been extensively studied and are covered in the preceding section. According to Zhao et al. (2015), boosting sucrose synthase substrate and aquaporin proteins is another technique for minimizing damage and enhancing protein stability, antioxidant activity, and secondary metabolism during times of water stress condition.
Plants under drought stress develop long deep roots as an escape mechanism to evade water stress (Okogbenin et al. 2013). A steeper root angle is associated with drought tolerance in various agronomic crops, including wheat (Comas et al. 2013), maize (Lynch 2013), and rice (Saengwilai et al. 2018). However, in cassava, wide root angle may not help in drought tolerance possibly due to the very shallow root system and complex storage root arrangements (Kengkanna et al. 2019). Accumulation of ABA and ethylene further increased the sustainability of plants under drought (Alves and Setter 2000; Duque and Setter 2013; Liao et al. 2016; Ogaddee and Girdthai 2019). Non-photochemical dissipation of energy mediated by enhanced carotenoid synthesis and enhanced PS-I and PS-II subunits (CP43 and R subunits) can be used as a protective mechanism against photooxidation in cassava (Pereira et al. 2018; Shan et al. 2018). Understanding the precise molecular, biochemical, and physiological level mechanism of drought tolerance is pivotal for enhancing storage root yield as the predicted climate change is likely to aggravate the drought intensity.
8 Key Traits for Enduring Drought Tolerance in Cassava
Selection based on highly heritable morphological, molecular, and physiological traits contributes directly to storage root yield under drought stress. Rapid storage root development followed by early bulking has been identified as one of the most important traits to enhance drought tolerance. This trait can be an effective drought avoidance strategy during early water stress period or intermittent water stress episode. Early bulking variety could have the upper hand over mid or late bulking varieties as late or mid bulking variety may not accumulate desired quantities of biomass under water limiting condition. Moreover, late bulking varieties might increase the cropping period beyond harvesting (12 months after planting in general) with serious implications in terms of storage root quality. Breeding cassava cultivars with highly regulated root system architecture with higher lateral branching density could result into drought tolerant cultivar. Additional traits like higher root length and number, dry biomass, higher axle length, and density of upper nodal roots can further intensify drought tolerance. Screening cassava genotypes for photosynthetic efficiency is easy and widely followed. Maintenance of reasonable photosynthetic rate under drought is essential to sustain higher productivity. It is suggested to breed varieties with a lower stomatal density on the abaxial surface of hypostomatous leaves in wet/humid zones or in ecosystems with short intermittent water drought episodes. On the other hand, selecting cultivars with a higher stomatal density on the abaxial surface of hypostomatous leaves would optimize water use efficiency and sustain productivity under prolonged drought period in subhumid/semi-arid ecosystems.
Selecting/development of medium or short canopy genotypes (one with developing extra branches late) with optimum LAI, HI, and PI is one of the strategies for developing drought-tolerant variety as storage root weight is positively correlated with leaf number, and cultivars that retained many leaves were high yielders. Increased leaf life under water stress is an important drought-tolerant trait. An evaluation of modern cassava cultivars based on leaf life, leaf turnover rate, leaf retention or recovery index interception efficiency of photosynthetically active radiation (PAR), and efficiency of conversion of that intercepted PAR have major opportunities for genetic improvement in water-deficient environments as it will help in sustaining optimum LAI throughout the growing season under stress. Furthermore, integrating anatomical plastic traits viz. increased epidermis thickness and blade adaxial face, increased palisade and spongy parenchyma thickness, and decreased xylem vulnerability would give additional benefit to drought-tolerant cultivars.
In the view of reduced photosynthesis under drought, the storage root bulking process becomes increasingly reliant of carbohydrate reserve in stem and leaves. Therefore, improving drought tolerance may involve selection of varieties based on regulation of carbohydrate reserves and its judicious use during drought episodes for canopy and storage root development. Additionally, increased pith density and pith biomass could serve as potential drought tolerance traits to develop drought-tolerant cultivars. The number of storage roots and storage root biomass are potential indicators of drought tolerance in cassava and should be incorporated into breeding programs. Cassava’s capacity to cope against drought is associated with accumulation of osmolytes to regulate osmotic adjustment. These traits have not been explored in detail in cassava. Yu et al. (2016) developed transgenic rice plants with constitutively expressing cassava plasma membrane–specificMePMP3-2 gene, conferring drought tolerance by upregulating stress-related genes OsProT and OsP5CS, enhanced proline content and lowered malondialdehyde (MDA) content. There is great scope for developing drought-tolerant transgenic cassava with increased MePMP3-2 gene expression. Integrating multiple traits into a comprehensive breeding program using advanced genomic and phenotyping tools could accelerate the development of high-yielding drought-tolerant cassava varieties with increased genetic gain.
9 Conclusion
Drought limited cassava plant’s productivity, by reducing leaf area, photosynthetic efficiency, biochemical parameters and quality. Phenotypic plastic responses instrumental in enhancing drought tolerance of cassava include abscission of old leaves, continued but limited photosynthetic area, highly regulated stomatal activity and starch reserve utilization, increased proline content, and modified sink-source relations. Screening and phenotyping drought adaptation traits during early stage would expedite the development of drought-tolerant varieties. While new omics-driven high-throughput techniques can be used to identify new genes linked with drought resistance, the immediate goal is to confirm the QTL and genes that have already been identified and include them in the breeding program. Starch exhibited vast portfolio of changes in its yield, quality, hydration, and functional properties. However, this aspect demands urgent intervention as very little information is available on changes in the physical, mechanical, and processing qualities of cassava starch under drought stress. In addition, these parameters should be integrated into breeding program to ensure nutritional benefits and its suitability for industrial application in the future. Furthermore, developing high-yielding drought-tolerant cassava cultivars with high starch content and low cyanogenic glucoside potential will be challenging for breeders as consumption of high HCN is associated with several diseases and disorders in humans. There remains a paucity of information on rhizospheric changes, viz., nutrient imbalance, microbial populations, and diversity associated with drought and their implications for the aboveground growth of cassava. Integrating multi-level adaptive traits and eco-physiological approaches will be essential to sustain the productivity and quality of cassava under drought stress.
Data Availability
Data sharing does not apply to this article as no datasets were generated or analyzed during the current study.
Abbreviations
- A:
-
Carbon assimilation
- ABA:
-
Abscisic acid
- Cal:
-
Calorie
- CAT:
-
Catalase
- CG:
-
Cyanogenic glycosides
- Ci/Ca :
-
Ratio of internal (Ci) to atmospheric CO2 concentration
- Ca :
-
Leaf internal carbon dioxide concentration
- DAP:
-
Days after planting
- DEGs:
-
Differentially expressed genes
- DIRT:
-
Digital imaging of root traits
- E:
-
Transpiration
- ETR:
-
Electron transfer rate
- FC:
-
Field capacity
- FW:
-
Fresh weight
- g:
-
Gram
- GR:
-
Glutathione reductase
- gs :
-
Stomatal conductance
- h:
-
Hour
- ha:
-
Hectare
- HCN:
-
Hydrogen cyanide
- HI:
-
Harvest index
- K+ :
-
Potassium ion
- kg:
-
Kilogram
- LAI:
-
Leaf area index
- m:
-
Meter
- MAP:
-
Months after planting
- NSC:
-
Nonstructural carbohydrates
- OA:
-
Osmotic adjustment
- PAR:
-
Photosynthetically active radiation
- PCD:
-
Program cell death
- PEPC:
-
Phosphoenolpyruvate carboxylase
- PSII:
-
Photosystem II
- qP:
-
Photochemical quenching coefficient
- ROS:
-
Reactive oxygen species
- RSA:
-
Root system architecture
- RuBisCO:
-
Ribulose-1,5-bisphosphate carboxylase-oxygenase
- s:
-
Second
- SOD:
-
Superoxide dismutase
- t:
-
Ton
- UN:
-
United Nations
- US$:
-
United States dollars
- WDS:
-
Water-deficit stress
- WP:
-
Water potential
- WUE:
-
Water use efficiency
- ΦPSII:
-
Effective quantum yield
- ICAR:
-
Indian Council of Agricultural Research
References
Adjebeng-Danquah J, Manu-Aduening J, Gracen VE, Offei SK, Asante IK (2016) Genotypic variation in abscisic acid content, carbon isotope ratio and their relationship with cassava growth and yield under moisture stress and irrigation. J Crop Sci Biotechnol 19:263–273. https://doi.org/10.1007/s12892-016-0004-9
Adu MO (2020) Causal shoot and root system traits to variability and plasticity in juvenile cassava (Manihot esculenta Crantz) plants in response to reduced soil moisture. Physiol Mol Biol Plants 26:1799–1814. https://doi.org/10.1007/s12298-020-00865-4
Adu MO, Asare PA, Asare-Bediako E, Amenorpe G, Ackah F, Afutu E, Amoah MN, Yawson D (2018) Characterizing shoot and root system trait variability and contribution to genotypic variability in juvenile cassava (Manihot esculenta Crantz) plants. Heliyon 4:e00665. https://doi.org/10.1016/j.heliyon.2018.e00665
Adu MO, Asare PA, Yawson DO, Nyarko MA, Abdul Razak A, Kusi AK, Tachie-Menson JW, Afutu E, Andoh DA, Ackah FK, Vanderpuije GC (2020) The search for yield predictors for mature field-grown plants from juvenile pot-grown cassava (Manihot esculenta Crantz). PLoS ONE 15:e0232595. https://doi.org/10.1371/journal.pone.0232595
Aina OO, Dixon AGO, Akinrinde EA (2007) Effect of soil moisture stress on growth and yield of cassava in Nigeria. Pakistan J Biological Sci 10:3085–3090. https://doi.org/10.3923/pjbs.2007.3085.3090
Alves AAC, Setter TL (2000) Response of cassava to water deficit: leaf area growth and abscisic acid. Crop Sci 40:131–137. https://doi.org/10.2135/cropsci2000.401131x
Alves AAC, Setter TL (2004) Response of cassava leaf area expansion to water deficit: cell proliferation, cell expansion and delayed development. Annals Bot 94:605–613. https://doi.org/10.1093/aob/mch179
Alves AAC, Setter TL (2004) Abscisic acid accumulation and osmotic adjustment in cassava under water deficit. Environ Exp Bot 51:259–271. https://doi.org/10.1016/j.envexpbot.2003.11.005
Amelework AB, Bairu MW, Maema O, Venter S, Laing M (2021) Adoption and promotion of resilient crops for climate risk mitigation and import substitution: a case analysis of cassava for South African agriculture. Front Sustain Food Syst 5:617783. https://doi.org/10.3389/fsufs.2021.617783
An D, Ma Q, Wang H, Yang J, Zhou W, Zhang P (2017) Cassava C-repeat binding factor 1 gene responds to low temperature and enhances cold tolerance when overexpressed in Arabidopsis and cassava. Plant Mol Biol 94:109–124
Anikwe MAN, Ikenganyia EE (2018) Ecophysiology and production principles of cassava (Manihot species) in southeastern Nigeria. In: Waisundara V (ed) Cassava. InTech. https://doi.org/10.5772/intechopen.70828
Arora NK (2019) Impact of climate change on agriculture production and its sustainable solutions. Environ Sustain 2:95–96. https://doi.org/10.1007/s42398-019-00078-w
Aslam B, Basit M, Nisar MA, Khurshid M, Rasool MH (2017) Proteomics: technologies and their applications. J Chromatogr Sci 55(2):182–196. https://doi.org/10.1093/chromsci/bmw167
Ballen-Taborda C, Plata G, Ayling S, Rodriguez-Zapata F, Lopez-Lavalle LAB, Duitama J (2013) Identification of cassava microRNAs under abiotic stress. Int J Genomics. https://doi.org/10.1155/2013/857986
Bhattacharya A (2019) Nitrogen-use efficiency under changing climatic conditions. In: Bhattacharya A (Ed) Changing Climate and Resource Use Efficiency in Plants. Academic Press, pp 181-240. https://doi.org/10.1016/C2017-0-04681-5
Blum A (2009) Effective use of water (EUW) and not water-use efficiency (WUE) is the target of crop yield improvement under drought stress. Field Crops Res 112(2–3):119–123. https://doi.org/10.1016/j.fcr.2009.03.009
Bull SE, Alder A, Barsan C, Kohler M, Hennig L, Gruissem W, Vanderschuren H (2017) FLOWERING LOCUS T triggers early and fertile flowering in glasshouse cassava (Manihot esculenta Crantz). Plants (Basel) 6(2):22. 10.3390%2Fplants6020022
Burns A, Gleadow R, Cliff J, Zacarias A, Cavagnaro T (2010) Cassava: the drought, war and famine crop in a changing world. Sustainability 2:3572–3607. https://doi.org/10.3390/su2113572
Burns AE, Gleadow RM, Zacarias AM, Cuambe CE, Miller RE, Cavagnaro TR (2012) Variations in the chemical composition of cassava (Manihot esculenta Crantz) leaves and roots as affected by genotypic and environmental variation. J Agric Food Chem 60:4946–4956. https://doi.org/10.1021/jf2047288
Calatayud PA, Llovera E, Bois JF, Lamaze T (2000) Photosynthesis in drought-adapted cassava. Photosynt 38:97–104. https://doi.org/10.1023/A:1026704226276
Cardoso AP, Mirione E, Ernesto M, Massaza F, Cliff J, Haque MR, Bradbury JH (2005) Processing of cassava roots to remove cyanogens. J Food Compos Anal 18(5):451–460. https://doi.org/10.1016/j.jfca.2004.04.002
Ceballos H, De la Cruz A, Gabriel A (2012) Cassava taxonomy and morphology. In: Cassava in the third millennium: modern production, processing, use, and marketing systems/Compiled and directed by: Bernardo Ospina and Hernán Ceballos. Centro Internacional de Agricultura Tropical (CIAT), Cali, Colombia, pp 15–28; Latin American and Caribbean Consortium to Support Cassava Research and Development (CLAYUCA); TechnicalCentre for Agricultural and Rural Cooperation (CTA), p 574. – (CIAT publication no. 377). http://ciat-library.ciat.cgiar.org/Articulos_Ciat/cassava_in_third_millennium_1.pdf
Chang L, Wang L, Peng C, Tong Z, Wang D, Ding G, Xiao J, Guo A, Wang X (2019) The chloroplast proteome response to drought stress in cassava leaves. Plant Physiol Biochem 142:351–362. https://doi.org/10.1016/j.plaphy.2019.07.025
Cock JH (1984) Cassava. In: Goldsworthy PR, Fisher NM (eds) The physiology of tropical field crops. Wiley, New York, pp 529–549
Cock JH, Franklin D, Sandoval G, Juri P (1979) The ideal cassava plant for maximum yield. Crop Sci 19:271–279. https://doi.org/10.2135/cropsci1979.0011183X001900020025x
Comas L, Becker S, Cruz VM, Byrne PF, Dierig DA (2013) Root traits contributing to plant productivity under drought. Front Plant Sci 4:442. https://doi.org/10.3389/fpls.2013.00442
Connor DJ, Cock JH (1981) Response of cassava to water shortage II. Canopy dynamics. Field Crops Res 4:285–296. https://doi.org/10.1016/0378-4290(81)90079-4
Connor DJ, Cock JH, Parra GE (1981) Response of cassava to water shortage I. Growth and yield. Field Crops Res 4:181–200. https://doi.org/10.1016/0378-4290(81)90071-X
Dalal VK, Tripathy BC (2018) Water-stress induced downsizing of light-harvesting antenna complex protects developing rice seedlings from photo-oxidative damage. Sci Rep 8:5955. https://doi.org/10.1038/s41598-017-14419-4
Daryanto S, Wang L, Jacinthe PA (2016) Drought effects on root and tuber production: a meta-analysis. Agric Water Manage 176:122–131. https://doi.org/10.1016/j.agwat.2016.05.019
de Olanda Souza GH, de Oliveira Aparecido LE, de Moraes JRdSC, Botega Guilherme Torsoni (2022) Climate change and its influence on planting of cassava in the Midwest region of Brazil. Environ Dev Sustain. https://doi.org/10.1007/s10668-021-02088-3
de Oliveira EJ, Aidar SD, Morgante CV, Chaves AR, Cruz JL, Coelho Filho MA (2015) Genetic parameters for drought-tolerance in cassava. Pesq Agropec Bras 50:233–241. https://doi.org/10.1590/S0100-204X2015000300007
de Oliveira EJ, Morgante Carolina Vianna, de Tarso Saulo, Aidar Agnaldo Rodrigues, de Melo Chaves, Rafaela Priscila Antonio, Jailson Lopez Cruz, Maurício Antônio Coelho Filho, (2017) Evaluation of cassava germplasm for drought tolerance under field conditions. Euphytica 213:188. https://doi.org/10.1007/s10681-017-1972-7
De Souza AP, Long SP (2018) Towards improving photosynthesis in cassava: characterizing photosynthetic limitations in four current African cultivars. Food Energy Secur 7:e00130
De Souza AP, Massenburg LN, Jaiswal D, Cheng S, Shekar R, Long SP (2017) Rooting for cassava: insights into photosynthesis and associated physiology as a route to improve yield potential. New Phytol 213:50–65. https://doi.org/10.1111/nph.14250
de Tafur MS, El-Sharkawy MA, Calle F (1997) Photosynthesis and yield performance of cassava in seasonally dry and semiarid environments. Photosynthetica 33:249–257
Defloor I, Swennen R, Bokanga M, Delcour JA (1998) Moisture stress during growth affects the breadmaking and gelatinization properties of cassava (Manihot esculenta Crantz) flour. J Sci Food Agr 76(2):233–238
Dietz KJ, Zorb C, Geilfus CM (2021) Drought and crop yield. Plant Biol J 23:881–893. https://doi.org/10.1111/plb.13304
Ding Z, Fu L, Tie W, Yan Y, Wu C, Hu W, Zhang J (2019) Extensive post-transcriptional regulation revealed by transcriptomic and proteomic integrative analysis in cassava under drought. J Agric Food Chem 67(12): 3521-3534. https://doi.org/10.1021/acs.jafc.9b00014 dos Santos Silva Carlos Andre, Lívia Maria Batista Vilela, Roberta Lane de Oliveira-Silva, Jéssica Barboza da Silva, Alexandre Reis Machado, João Pacífico Bezerra-Neto, Sergio Crovella, Ana Maria Benko-Iseppon (2021) Cassava (Manihot esculenta) defensins: Prospection, structural analysis and tissue-specific expression under biotic/abiotic stresses. Biochimie 186: 1-12. https://doi.org/10.1016/j.biochi.2021.03.012
Dong S, Xiao L, Li Z, Shen J, Yan H et al (2022) A novel long noncoding RNA, DIR, increases drought tolerance in cassava by modifying stress-related gene expression. J Integrative Agriculture 21:2588–2602
dos Santos Silva PP, Bandeira e Sousa M, de Oliveira EJ, Morgante CV, de Oliveira CRS, Vieira SL, Borel JC (2021) Genome-wide association study of drought tolerance in cassava. Euphytica 217(4):1–26
Duque LO (2012) Cassava drought tolerance mechanisms revisited: evaluation of drought tolerance in contrasting cassava genotypes under water stressed environments. PhD dissertation submitted to Cornell University. https://hdl.handle.net/1813/29398
Duque LO, Setter TL (2013) Cassava response to water deficit in deep pots: root and shoot growth, ABA, and carbohydrate reserves in stems, leaves and storage roots. Tropical Plant Biol 6:199–209. https://doi.org/10.1007/s12042-013-9131-3
Duque LO, Setter TL (2019) Partitioning index and nonstructural carbohydrate dynamics among contrasting cassava genotypes under early terminal water stress. Environ Exp Bot 163:24–35. https://doi.org/10.1016/j.envexpbot.2019.03.023
Duque LO, Villordon A (2019) Root branching and nutrient efficiency: status and way forward in root and tuber crops. Front Plant Sci 10:237. https://doi.org/10.3389/fpls.2019.00237
Ekanayake IJ, Porto MC, Dixon AG (1994) Response of cassava to dry weather: potential and genetic variability. African Crop Science Conference Proceedings 1:115–119
Ekanayake IJ, Dixon AGO, Porto MCM (1996) Performance of various cassava clones in the dry savannah region of Nigeria. In: Kurup C, Palaniswamy M, Potty V, Padmaja G, Kabeerathumar AS, Pillai S (eds) Tropical tuber crops: problems, prospects and future strategies. Science Publishers, Lebanon, PA, USA, pp 207–215
El-Sharkawy MA (2004) Cassava biology and physiology. Plant Mol Biol 56:481–501. https://doi.org/10.1007/s11103-005-2270-7
El-Sharkawy MA (2006) International research on cassava photosynthesis, productivity, eco-physiology, and responses to environmental stresses in the tropics. Photosynt 44:481–512. https://doi.org/10.1007/s11099-006-0063-0
El-Sharkawy MA (2007) Physiological characteristics of cassava tolerance to prolonged drought in the tropics: implications for breeding cultivars adapted to seasonally dry and semiarid environments. Braz J Plant Physiol 19:257–286. https://doi.org/10.1590/S1677-04202007000400003
El-Sharkawy MA (2012) Stress-tolerant cassava: the role of integrative ecophysiology-breeding research in crop improvement. Open J Soil Sci 2:162–186
El-Sharkawy MA (2014) Global warming: causes and impacts on agroecosystems productivity and food security with emphasis on cassava comparative advantage in the tropics/subtropics. Photosynt 52:161–178. https://doi.org/10.1007/s11099-014-0028-7
El-Sharkawy MA (2016) Prospects of photosynthetic research for increasing agricultural productivity, with emphasis on the tropical C4 amaranthus and the cassava C3–C4 crops. Photosynt 54:161–184. https://doi.org/10.1007/s11099-016-0204-z
El-Sharkawy MA, Cock JH (1987) C3–C4 intermediate photosynthetic characteristics of cassava (Manihot esculenta Crantz): I. Gas exchange. Photosynth Res 12:219–235. https://doi.org/10.1007/BF00055122
El-Sharkawy MA, De Tafur SM (2007) Genotypic and within canopy variation in leaf carbon isotope discrimination and its relation to short-term leaf gas exchange characteristics in cassava grown under rain-fed conditions in the tropics. Photosynt 45:515–526. https://doi.org/10.1007/s11099-007-0089-y
El-Sharkawy MA, Cock JH, Lynam JK, Hernandez ADP, Cadavid LF (1990) Relationships between biomass, root yield and single-leaf photosynthesis in field-grown cassava. Field Crops Res 25:183–201
Ernesto M, Cardoso AP, Nicala D, Mirione E, Massaza F, Cliff J, Haque MR, Bradbury JH (2002) Persistent konzo and cyanogen toxicity from cassava in northern Mozambique. Acta Tropica 82(3):357–362
Ewa F, Asiwe JNA, Okogbenin E, Ogbonna AC, Egesi C (2021) KASPar SNP genetic map of cassava for QTL discovery of productivity traits in moderate drought stress environment in Africa. Sci Rep 11(1):11268. https://doi.org/10.1038/s41598-021-90131-8
Fan W, Hai M, Guo Y, Ding Z, Tie W, Ding X, Yan Y, Wei Y, Liu Y, Chunlai W, Shi H, Li K, Wei H (2016) The ERF transcription factor family in cassava: genome-wide characterization and expression analyses against drought stress. Sci Rep 6:37379. https://doi.org/10.1038/srep37379
FAO (2011) Climate change, water and food security. Water Reports 36, FAO, Rome.
FAO (2014) Food Outlook Biannual Report On Global Food Markets. FAO, Rome
FAO (2017) The future of food and agriculture - trends and challenges. FAO, Rome
FAO (2018) The state of agricultural commodity markets 2018. Agricultural trade, climate change and food security. Rome.
FAO (2020) The State of Food and Agriculture 2020. Overcoming water challenges in agriculture, Rome. https://doi.org/10.4060/cb1447en
FAO (2021) Crop prospects and food situation - quarterly global report no. 1, March 2021. Rome. https://doi.org/10.4060/cb3672en
FAOSTAT (2022) https://www.fao.org/faostat/en/#data/QCL/visualize accessed on 10 May 2022
Felemban A, Braguy J, Zurbriggen MD, Al-Babili S (2019) Apocarotenoids involved in plant development and stress response. Front Plant Sci 27(10):1168. https://doi.org/10.3389/fpls.2019.01168
Feng RJ, Ren MY, Lu LF, Peng M, Guan X, Zhou DB, Zhang MY, Qi DF, Li K, Tang W, Yun TY, Chen YF, Wang F, Zhang D, Shen Q, Liang P, Zhang YD, Xie JH (2019) Involvement of abscisic acid-responsive element-binding factors in cassava (Manihot esculenta) dehydration stress response. Sci Rep 9:12661. https://doi.org/10.1038/s41598-019-49083-3
Fernandez MD, Tezara W, Rengifo E, Herrera A (2002) Lack of downregulation of photosynthesis in a tropical root crop, cassava, grown under an elevated CO2 concentration. Funct Plant Biol 29(7):805–814. https://doi.org/10.1071/PP01165
Frona D, Szenderak J, Harangi-Rakos M (2019) The challenge of feeding the world. Sustainability 11(20):5816. https://doi.org/10.3390/su11205816
Fu L, Ding Z, Han B, Hu W, Li Y, Zhang J (2016) Physiological investigation and transcriptome analysis of polyethylene glycol (PEG)-induced dehydration stress in cassava. Int J Mol Sci 17:283. https://doi.org/10.3390/ijms17030283
Gerland P, Raftery AE, Sevčíková H, Li N, Gu D, Spoorenberg T, Alkema L, Fosdick BK, Chunn J, Lalic N, Bay G, Buettner T, Heilig GK, Wilmoth J (2014) World population stabilization unlikely this century. Science 346(6206):234–237. https://doi.org/10.1126/science.1257469
Giordano M, Petropoulos SA, Rouphael Y (2021) Response and defence mechanisms of vegetable crops against drought, heat and salinity stress. Agriculture 11(5):463. https://doi.org/10.3390/agriculture11050463
Gleadow R, Pegg A, Blomstedt CK (2016) Resilience of cassava (Manihot esculenta Crantz) to salinity: implications for food security in low-lying regions. J Exp Bot 67:5403–5413. https://doi.org/10.1093/jxb/erw302
GlobeNewswire (2021) https://www.globenewswire.com/en/news-release/2020/03/10/1997843/0/en/Cassava-Starch-Market-Size-Worth-USD-66-84-Billion-by-2026-Rising-Demand-for-Functional-Food-to-Augment-Growth-Globally-states-Fortune-Business-Insights.html accessed on 30 October 2021
Hatfield JL, Dold C (2019) Water-use efficiency: advances and challenges in a changing climate. Front Plant Sci 10:103. https://doi.org/10.3389/fpls.2019.00103
Havrlentova M, Kraic J, Gregusova V, Kovacsova B (2021) Drought stress in cereals - a review. Agriculture 67:47–60. https://doi.org/10.2478/agri-2021-0005
Helal NAS, Eisa SS, Attia A (2013) Morphological and chemical studies on influence of water deficit on cassava. World J Agril Sci 9(5):369–376
Howeler R, Lutaladio N, Thomas G (2013) Save and grow cassava: a guide to sustainable production intensification. FAO Plant Production and Protection Division, Rome
Hular-Bograd J, Sarobol E, Rojanaridpiched C, Sriroth K (2011) Effect of supplemental irrigation on reducing cyanide content of cassava variety Kasetsart 50. Agric Nat Resour 45(6):985–994
Ike IF, Thurtell GW (1981) Osmotic adjustment in indoor grown cassava in response to water stress. Physiol Plant 52:257–262. https://doi.org/10.1111/j.1399-3054.1981.tb08502.x
Imakumbili ML, Semu E, Semoka JM, Abass A, Mkamilo G (2019) Cyanogenic glucoside production in cassava: the comparable influences of varieties, soil moisture content and nutrient supply. bioRxiv. 1: 649236. https://doi.org/10.1101/649236
Isah T (2019) Stress and defense responses in plant secondary metabolites production. Biol Res 52:39. https://doi.org/10.1186/s40659-019-0246-3
Itani J, Oda T, Numao T (1999) Studies on mechanisms of dehydration postponement in cassava leaves under short-term soil water deficits. Plant Prod Sci 2(3):184–189
Izumi Y, Yuliadi E, Sunyoto S, Iijima M (1999) Root system development including root branching in cuttings of cassava with reference to shoot growth and tuber bulking. Plant Prod Sci 2:267–272. https://doi.org/10.1626/pps.2.267
Janket A, Vorasoot N, Toomsan B, Kaewpradit W, Jogloy S, Theerakulpisut P, Holbrook CC, Kvien CK, Banterng P (2020) Starch accumulation and granule size distribution of cassava cv. Rayong 9 grown under irrigated and rainfed conditions using different growing seasons. Agronomy 10(3): 412. https://doi.org/10.3390/agronomy10030412
Jannink JL, Lorenz AJ, Iwata Hiroyoshi (2010) Genomic selection in plant breeding: from theory to practice. Brief Funct Genom 9(2):166–177. https://doi.org/10.1093/bfgp/elq001
Jarvis A, Ramirez-Villegas J, Herrera Campo BV, Navarro-Racines C (2012) Is cassava the answer to African climate change adaptation? Tropical Plant Biol 5(1):9–29. https://doi.org/10.1007/s12042-012-9096-7
Joseph Adjebeng-Danquah, Gracen Vernon Edward, Offei Samuel Kwame, Asante Isaac Kwadwo, Manu-Aduening Joseph (2016) Genetic variability in storage root bulking of cassava genotypes under irrigation and no irrigation. Agric & Food Secur 5:9. https://doi.org/10.1186/s40066-016-0055-7
Jyothi AN, Sajeev MS (2021) Industrial products from tropical tuber crops. In: More SJ, Namrata AG, Suresh Kumar J, Visalakshi CC, Sirisha T (eds) Recent Advances in Root and Tuber Crops. Brillion Publishing, New Delhi, India, pp 391–406
Kengkanna J, Jakaew P, Amawan S, Busener N, Bucksch A, Saengwilai P (2019) Phenotypic variation of cassava root traits and their responses to drought. Appl Plant Sci 7:e01238. https://doi.org/10.1002/aps3.1238
Koundinya AVV, More SJ (2021) Breeding for drought tolerance in cassava. In: More SJ, Namrata AG, Suresh Kumar J, Visalakshi CC, Sirisha T (eds) Recent Advances in Root and Tuber Crops. Brillion Publishing, New Delhi, India, pp 51–64
Koundinya AVV, Hegde Vivek, Sheela MN, Visalakshi Chandra C (2018) Evaluation of cassava varieties for tolerance to water deficit stress conditions. J Root Crops 44(1):70–75
Koundinya AV, Ajeesh BR, Hegde V, Sheela MN, Mohan C, Asha KI (2021) Genetic parameters, stability and selection of cassava genotypes between rainy and water stress conditions using AMMI, WAAS. BLUP and MTSI. Sci Hortic 281:109949. https://doi.org/10.1016/j.scienta.2021.109949
Latif S, Muller J (2015) Potential of cassava leaves in human nutrition: a review. Trends Food Sci Technol 44(2):147–158. https://doi.org/10.1016/j.tifs.2015.04.006
Lebot V (2020) Tropical root and tuber crops: cassava, sweet potato, yams and aroids. Crop Production Science in Horticulture Series 17. Wallingford, CT: CABI. https://doi.org/10.1079/9781789243369.0000
Lei N, Yu X, Li S, Zeng C, Zou L, Liao W, Peng M (2017) Phylogeny and expression pattern analysis of TCP transcription factors in cassava seedlings exposed to cold and/or drought stress. Sci Rep 7:10016. https://doi.org/10.1038/s41598-017-09398-5
Leng G, Hall J (2019) Crop yield sensitivity of global major agricultural countries to droughts and the projected changes in the future. Sci Total Environ 654:811–821. https://doi.org/10.1016/j.scitotenv.2018.10.434
Lenis JI, Calle F, Jaramillo G, Perez JC, Ceballos H, Cock JH (2006) Leaf retention and cassava productivity. Field Crops Res 95:126–134. https://doi.org/10.1016/j.fcr.2005.02.007
Li S, Yu X, Cheng Z, Xiaoling Y, Mengbin R, Wenbin L, Ming P (2017a) Global Gene expression analysis reveals crosstalk between response mechanisms to cold and drought stresses in cassava seedlings. Front Plant Sci 8:1259. https://doi.org/10.3389/fpls.2017.01259
Li S, Yu X, Lei N, Cheng Z, Zhao P, He Y, Wang W, Peng M (2017b) Genome-wide identification and functional prediction of cold and/or drought-responsive lncRNAs in cassava. Sci Rep 7:45981. https://doi.org/10.1038/srep45981
Li S, Cao P, Wang C, Guo J, Zang Y, Wu K, Ran F, Liu L, Wang D, Min Y (2021) Genome-wide analysis of tubulin gene family in cassava and expression of family member FtsZ2-1 during various stress. Plants 10(4):668. https://doi.org/10.3390/plants10040668
Li S, Zhao P, Yu X, Liao W, Peng M, Ruan M (2022) Cell signaling during drought and/or cold stress in cassava. Tropical Plants 1(1):1–7
Liao W, Wang G, Li Y, Wang B, Zhang P, Peng M (2016) Reactive oxygen species regulate leaf pulvinus abscission zone cell separation in response to water-deficit stress in cassava. Sci Rep 6:21542. https://doi.org/10.1038/srep21542
Lokko YV, Anderson JV, Rudd ST, Raji AD, Horvath D, Mikel MA, Kim RY, Liu L, Hernandez AL, Dixon AG, Ingelbrecht IL (2007) Characterization of an 18,166 EST dataset for cassava (Manihot esculenta Crantz) enriched for drought-responsive genes. Plant Cell Rep 26(9):1605–1618. https://doi.org/10.1007/s00299-007-0378-8
Lynch JP (2013) Steep, cheap and deep: an ideotype to optimize water and N acquisition by maize root systems. Annals Bot 112:347–357. https://doi.org/10.1093/aob/mcs293
Møller BL (2010) Functional diversifications of cyanogenic glucosides. Curr Opin Plant Biol 13(3):337–346. https://doi.org/10.1016/j.pbi.2010.01.009
More SJ, Ravi V, Saravanan R (2019a) Tropical tuber crops. In: de Freitas ST, Pareek S (eds) Postharvest physiological disorders in fruits and vegetables. CRC Press, Boca Raton, pp 719–757. https://doi.org/10.1201/b22001
More SJ, Ravi V, Suresh Kumar J (2019b) Cassava under water deficit stress: differential carbon isotope discrimination, 19–21 December 2019, National Conference of Plant Physiology NCPP-2019 Plant Productivity and Stress Management. Kerala Agricultural University, Thrissur, Kerala, Department of Plant Physiology
More SJ, Ravi V, Raju S, Suresh Kumar J (2020) The quest for high yielding drought-tolerant cassava variety. J Pharmacogn Phytochem SP6:433–439
More SJ, Namrata AG, Suresh Kumar J, Visalakshi CC, Sirisha T (2021) Recent advances in root and tuber crops. Brillion Publishing, New Delhi, India
More SJ, Ravi V, Raju S (2022) Carbon isotope discrimination studies in plants for abiotic stress. In: Shanker AK, Shanker Chitra, Anand Anjali, Maheswari M (eds) Climate Change and Crop. Academic Press, Cambridge, MA, pp 493–537. https://doi.org/10.1016/B978-0-12-816091-6.00004-3
Morgante CV, Nunes SL, de Melo Chaves AR, Ferreira CF, de Tarso Aidar S, Vitor AB, de Oliveira EJ (2020) Genetic and physiological analysis of early drought response in Manihot esculenta and its wild relative. Acta Physiol Plant 42:22. https://doi.org/10.1007/s11738-019-3005-8
Mtunguja MK, Laswai HS, Kanju E, Ndunguru J, Muzanila YC (2016) Effect of genotype and genotype by environment interaction on total cyanide content, fresh root, and starch yield in farmer-preferred cassava landraces in Tanzania. Food Sci Nutr 4:791–801. https://doi.org/10.1002/fsn3.345
Muiruri SK, Ntui VO, Tripathi L, Tripathi JN (2021) Mechanisms and approaches towards enhanced drought tolerance in cassava (Manihot esculenta). Current Plant Biol 28:100227. https://doi.org/10.1016/j.cpb.2021.100227
Mukherjee A, Immanuel S, Sreekumar J, Sivakumar PS (2019) Tropical tubers: journey from ‘life saving food’ to ‘future smart crops’. In: Chadha KL, Singh SK, Jai Prakash, Patel VB (eds) Shaping the future of horticulture. Kruger Brentt, UK, pp 511–522
Naumann G, Alfieri L, Wyser K, Mentaschi L, Betts RA, Carrao H, Spinoni J, Vogt J, Feyen L (2018) Global changes in drought conditions under different levels of warming. Geophys Res Lett 45(7):3285–3296. https://doi.org/10.1002/2017GL076521
Nayar NM (2014) The contribution of tropical tuber crops towards food security. J Root Crops 40(1):3–14
Nhassico D, Muquingue H, Cliff J, Cumbana A, Bradbury JH (2008) Rising African cassava production, diseases due to high cyanide intake and control measures. J Sci Food Agric 88(12):2043–2049
Nuwamanya E, Rubaihayo PR, Mukasa S, Kyamanywa S, Hawumba JF, Baguma Y (2014) Biochemical and secondary metabolites changes under moisture and temperature stress in cassava (Manihot esculenta Crantz). Afr J Biotechnol 13:3173–3186. https://doi.org/10.5897/AJB2014.13663
Odeku OA (2013) Potentials of tropical starches as pharmaceutical excipients: a review. Starch/Stärke 65:89–106. https://doi.org/10.1002/star.201200076
Ogaddee P, Girdthai T (2019) Physiological and ethylene accumulation responses of cassava under drought stress. IOP Conf Ser: Earth Environ Sci 346:012092. https://doi.org/10.1088/1755-1315/346/1/012092
Oguntunde PG (2005) Whole-plant water use and canopy conductance of cassava under limited available soil water and varying evaporative demand. Plant Soil 278:371–383. https://doi.org/10.1007/s11104-005-0375-z
Okogbenin E, Setter TL, Ferguson M, Mutegi R, Ceballos H, Olasanmi B, Fregene M (2013) Phenotypic approaches to drought in cassava: review. Front Physiol 4:93. https://doi.org/10.3389/fphys.2013.00093
Olasanmi O (2010) Cassava drought tolerance mechanisms re-visited: evaluation of drought tolerance in contrasting cassava genotypes under water stressed environments. Ph.D. thesis, Department of Agronomy (Plant breeding), University of Ibadan
Ooba M, Takahashi H (2003) Effect of asymmetric stomatal response on gas-exchange dynamics. Ecol Model 164(1):65–82. https://doi.org/10.1016/S0304-3800(03)00012-7
Orek C, Gruissem W, Ferguson M, Vanderschuren H (2020) Morpho-physiological and molecular evaluation of drought tolerance in cassava (Manihot esculenta Crantz). Field Crops Res 255:107861. https://doi.org/10.1016/j.fcr.2020.107861
Ou W, Mao X, Huang C, Tie W, Yan Y, Ding Z, Wu C, Xia Z, Wang W, Zhou S, Li K, Hu W (2018) Genome-wide identification and expression analysis of the KUP family under abiotic stress in cassava (Manihot esculenta Crantz). Front Physiol 9:17. https://doi.org/10.3389/fphys.2018.00017
Pacheco RI, Macias MP, Campos FC, Izquierdo AJ, Izquierdo GA (2019) Agronomic and physiological evaluation of eight cassava clones under water deficit conditions. Rev Fac Nac Agron Medellin 73(1):9109–9119. https://doi.org/10.15446/rfnam.v73n1.75402
Pardales JR Jr, Yamauchi A (2003) Regulation of root development in sweetpotato and cassava by soil moisture during their establishment period. Plant Soil 255:201–208. https://doi.org/10.1023/A:1026160309816
Pearce F (2007) Cassava comeback. New Scientist 194(2600):38–39. https://doi.org/10.1016/S0262-4079(07)61001-X
Pereira LFM, Zanetti S, de Silva M, A, (2018) Water relations of cassava cultivated under water-deficit levels. Acta Physiol Plant 40:13. https://doi.org/10.1007/s11738-017-2590-7
Pereira LF, Santos HL, Zanetti S, de Oliveira Brito IA, dos Santos Tozin LR, Rodrigues TM, de Almeida Silva M (2022) Morphology, biochemistry, and yield of cassava as functions of growth stage and water regime. S Afr J Bot 149:222–239. https://doi.org/10.1016/j.sajb.2022.06.003
Petrov V, Hille J, Mueller-Roeber B, Gechev TS (2015) ROS-mediated abiotic stress-induced programmed cell death in plants. Front Plant Sci 18(6):69. https://doi.org/10.3389/fpls.2015.00069
Petsakos A, Prager SD, Gonzalez Carlos Eduardo, Gama Arthur Chibwana, Sulser TB (2019) Understanding the consequences of changes in the production frontiers for roots, tubers and bananas. Global Food Sec 20:180–188. https://doi.org/10.1016/j.gfs.2018.12.005
Phosaengsri W, Banterng P, Vorasoot N, Jogloy S, Theerakulpisut P (2019) Leaf performances of cassava genotypes in different seasons and its relationship with biomass. Turk J Field Crop 24:54–64. https://doi.org/10.17557/tjfc.564116
Pineda M, Morante N, Salazar S, Cuásquer J, Hyde PT, Setter TL, Ceballos H (2020) Induction of earlier flowering in cassava through extended photoperiod. Agronomy 10(9):1273. https://doi.org/10.3390/agronomy10091273
Pipitpukdee S, Attavanich W, Bejranonda S (2020) Impact of climate change on land use, yield and production of cassava in Thailand. Agriculture 10(9):402. https://doi.org/10.3390/agriculture10090402
Pujol B, Salager JL, Beltran M, Bousquet S, McKey D (2008) Photosynthesis and leaf structure in domesticated cassava (Euphorbiaceae) and a close wild relative: have leaf photosynthetic parameters evolved under domestication? Biotropica 40:305–312. https://doi.org/10.1111/j.1744-7429.2007.00373.x
Pushpalatha R, Byju G (2020) Is cassava (Manihot esculenta Crantz) a climate “smart” crop? A review in the context of bridging future food demand gap. Tropical Plant Biol 13:201–211. https://doi.org/10.1007/s12042-020-09255-2
Pushpalatha R, Shiny R, Kutty G, Dua VK, Sunitha S, Santhosh Mithra VS, Byju G (2021) Testing of cassava (Manihot esculenta) varieties for climate resilience under Kerala (India) conditions. Agric Res. 22:1–8. https://doi.org/10.1007/s40003-021-00547-x
Putpeerawit P, Sojikul P, Thitamadee S, Narangajavana J (2017) Genome-wide analysis of aquaporin gene family and their responses to water-deficit stress conditions in cassava. Plant Physiol Biochem 121:118–127. https://doi.org/10.1016/j.plaphy.2017.10.025
Ravi V, Suja G, Saravanan R, More SJ (2021) Advances in cassava‐based multiple‐cropping systems. In: Warrington I (ed), 1st edn. Wiley, pp 153-232. https://doi.org/10.1002/9781119750802.ch3
Ren MY, Feng RJ, Shi HR, Lu LF, Yun TY, Peng M, Guan X, Zhang H, Wang JY, Zhang XY, Li CL (2017) Expression patterns of members of the ethylene signaling-related gene families in response to dehydration stresses in cassava. PLoS ONE 12:e0177621. https://doi.org/10.1371/journal.pone.0177621
Ribeiro MNO, Carvalho SP, Pereira FJ, Castro EM (2012) Anatomia foliar de mandioca em fun¸cao do potencial para tolerancia a diferentes condi¸c~oes ambientais. Rev Cienc Agron 43(2):354–361. https://doi.org/10.1590/S1806-66902012000200019
Rogers ED, Benfey PN (2015) Regulation of plant root system architecture: implications for crop advancement. Curr Opin Biotech 32:93–98. https://doi.org/10.1016/j.copbio.2014.11.015
Rosenthal DM, Ort DR (2012) Examining cassava’s potential to enhance food security under climate change. Tropical Plant Biol 5:30–38. https://doi.org/10.1007/s12042-011-9086-1
Ruan MB, Guo X, Wang B, Yang YL, Li WQ, Yu XL, Zhang P, Peng M (2017) Genome-wide characterization and expression analysis enables identification of abiotic stress-responsive MYB transcription factors in cassava (Manihot esculenta). J Ex Bot 68:3657–3672
Ruan MB, Yang YL, Li KM, Guo X, Wang B, Yu XL, Peng M (2018) Identification and characterization of drought-responsive CC-type glutaredoxins from cassava cultivars reveals their involvement in ABA signalling. BMC Plant Biol 18:329. https://doi.org/10.1186/s12870-018-1528-6
Ruan MB, Yu XL, Guo X, Zhao PJ, Peng M (2022) Role of cassava CC-type glutaredoxin MeGRXC3 in regulating sensitivity to mannitol-induced osmotic stress dependent on its nuclear activity. BMC Plant Biol 22:41. https://doi.org/10.1186/s12870-022-03433-y
Saengwilai P, Klinsawang S, Sangachart M, Bucksch A (2018) Comparing phenotypic variation of root traits in Thai rice (Oryza sativa L.) across growing systems. Appl Ecol Environ Res 16:1069–1083
Santisopasri V, Kurotjanawong K, Chotineeranat S, Piyachomkwan K, Sriroth K, Oates CG (2001) Impact of water stress on yield and quality of cassava starch. Ind Crops Prod 13:115–129. https://doi.org/10.1016/S0926-6690(00)00058-3
Setter TL, Fregene MA (2007) Recent advances in molecular breeding of cassava for improved drought stress tolerance. In: Jenks MA, Hasegawa PM, Jain SM (eds) Advances in Molecular Breeding Toward Drought and Salt Tolerant Crops. Springer, Netherlands, Dordrecht, pp 701–711
Shan Z, Luo X, Wei M, Huang T, Khan A, Zhu Y (2018) Physiological and proteomic analysis on long-term drought resistance of cassava (Manihot esculenta Crantz). Sci Rep 8:17982. https://doi.org/10.1038/s41598-018-35711-x
Shang S, Tang Y, Dai J, Wu C, Yan Y, Tie W, Li M, Yang J, Zeng J, Chen M, Wei H (2021) Genomic analysis of the principal members of antioxidant enzymes in simulated stresses response and postharvest physiological deterioration in cassava. Tropical Plant Biol 14:419–428. https://doi.org/10.1007/s12042-021-09304-4
Shuxia Li, Cheng Zhihao, Li Zhibo, Dong Shiman, Xiaoling Yu, Zhao Pingjuan, Liao Wenbin, Xiang Yu, Peng Ming (2021) MeSPL9 Attenuates drought resistance by regulating ja signaling and protectant metabolite contents in cassava. Theor Appl Genet 135:817–832. https://doi.org/10.1007/s00122-021-04000-z
Siegien I, Bogatek R (2006) Cyanide action in plants-from toxic to regulatory. Acta Physiol Plant 28:483–497. https://doi.org/10.1007/BF02706632
Sriroth K, Piyachomkwan K, Santisopasri V, Oates CG (2001) Environmental conditions during root development: drought constraint on cassava starch quality. Euphytica 120:95–102. https://doi.org/10.1023/A:1017511806128
Sunvittayakul P, Kittipadakul P, Wonnapinij P, Chanchay P, Wannitikul P, Sathitnaitham S, Phanthanong P, Changwitchukarn K, Suttangkakul A, Ceballos H, Vuttipongchaikij S (2022) Cassava root crown phenotyping using three-dimension (3D) multi-view stereo reconstruction. Sci Rep 12:10030. https://doi.org/10.1038/s41598-022-14325-4
Teerawanichpan P, Lertpanyasampatha M, Netrphan S, Varavinit S, Boonseng O, Narangajavana J (2008) Influence of cassava storage root development and environmental conditions on starch granule size distribution. Starch/Starke 60:696–705. https://doi.org/10.1002/star.200800226
Turyagyenda FL, Kizito EB, Baguma Y, Osiru D (2013a) Evaluation of Ugandan cassava germplasm for drought tolerance. Intl J Agri Crop Sci 5(3):212–226
Turyagyenda LF, Kizito EB, Ferguson M, Baguma Y, Agaba M, Harvey JJW, Osiru DSO (2013b) Physiological and molecular characterization of drought responses and identification of candidate tolerance genes in cassava. AoB PLANTS 5:plt007. https://doi.org/10.1093/aobpla/plt007
Utsumi Y, Tanaka MA, Morosawa T, Kurotani A, Yoshida T, Mochida K, Matsui A, Umemura Y, Ishitani M, Shinozaki K, Sakurai T, Seki M (2012) Transcriptome analysis using a high-density oligomicroarray under drought stress in various genotypes of cassava: an important tropical crop. DNA Research 19(4):335–345. https://doi.org/10.1093/dnares/dss016
Utsumi Y, Utsumi C, Tanaka M, Ha CV, Takahashi S, Matsui A, Matsunaga TM, Matsunaga S, Kanno Y, Seo M, Okamoto Y (2019) Acetic acid treatment enhances drought avoidance in Cassava (Manihot esculenta Crantz). Front Plant Sci 10:521. https://doi.org/10.3389/fpls.2019.00521
Vandegeer R, Miller RE, Bain M, Gleadow RM, Cavagnaro TR (2013) Drought adversely affects tuber development and nutritional quality of the staple crop cassava (Manihot esculenta Crantz). Funct Plant Biol 40:195–200. https://doi.org/10.1071/FP12179
Wahyuni Y, Supatmi Sri Hartati, N, Sudarmonowati E, (2020) Phenotypic and molecular changes in cassava (Manihot esculenta Crantz) genotypes as a response to water-deficit stress IOP Conf Ser. Earth Environ Sci 572:012013. https://doi.org/10.1088/1755-1315/572/1/012013
Wang B, Guo X, Zhao P, Ruan M, Yu X, Zou L, Yang Y, Li X, Deng D, Xiao J, Xiao Y, Hu C, Wang X, Wang X, Wang W, Peng M (2017) Molecular diversity analysis, drought related marker-traits association mapping and discovery of excellent alleles for 100-day old plants by EST-SSRs in cassava germplasms (Manihot esculenta Cranz). PLoS One 12(5):e0177456. https://doi.org/10.1371/journal.pone.0177456
Wang B, Guo X, Zhao P, Liao W, Zeng C, Li K, Zhou Y, Xiao J, Ruan M, Peng M, Bai Y (2021a) MeMYB26, a drought-responsive transcription factor in cassava (Manihot esculenta Crantz). Crop Breed Appl Biotechnol 21(1):e34432114. https://doi.org/10.1590/1984-70332021v21n1a4
Wang S, Lu C, Chen X, Wang H, Wang W (2021b) Comparative transcriptome profiling indicated that leaf mesophyll and leaf vasculature have different drought response mechanisms in cassava. Trop Plant Biol 14(4):396–407. https://doi.org/10.1007/s12042-021-09302-6
Wei Hu, Yan Yan, Tie Weiwei, Ding Zehong, Chunlai Wu, Ding Xupo, Wang Wenquan, Xia Zhiqiang, Guo Jianchun, Peng Ming (2018) Genome-wide analyses of calcium sensors reveal their involvement in drought stress response and storage roots deterioration after harvest in cassava. Genes (Basel) 9(4):221. https://doi.org/10.3390/genes9040221
Wei Y, Liu W, Hu W, Yan Y, Shi H (2020) The chaperone MeHSP90 recruits MeWRKY20 and MeCatalase1 to regulate drought stress resistance in cassava. New Phytol 226(2):476–491. https://doi.org/10.1111/nph.16346
Wongnoi S, Banterng P, Vorasoot N, Jogloy S, Theerakulpisut P (2020) Physiology, growth and yield of different cassava genotypes planted in upland with dry environment during high storage root accumulation stage. Agronomy 10:576. https://doi.org/10.3390/agronomy10040576
Wu C, Hu W, Yan Y, Tie W, Ding Z, Guo J, He G (2018) The late embryogenesis abundant protein family in Cassava (Manihot esculenta Crantz): genome-wide characterization and expression during abiotic stress. Molecules 23(5):1196. https://doi.org/10.3390/molecules23051196
Wu C, Ding X, Ding Z, Tie W, Yan Y, Wang Y, Yang H, Hu W (2019) The class III peroxidase (POD) gene family in cassava: identification, phylogeny, duplication, and expression. Int J Mol Sci 20(11):2730. https://doi.org/10.3390/ijms20112730
Xiao L, Shang XH, Cao S, Xie XY, Zeng WD, Lu LY, Chen SB, Yan HB (2019) Comparative physiology and transcriptome analysis allows for identification of lncRNAs imparting tolerance to drought stress in autotetraploid cassava. BMC Genom 20:514. https://doi.org/10.1186/s12864-019-5895-7
Xu J, Duan X, Yang J, Beeching JR, Zhang P (2013) Coupled expression of cu/Zn-superoxide dismutase and catalase in cassava improves tolerance against cold and drought stresses. Plant Signal Behav 8(6):e24525. https://doi.org/10.4161/psb.24525
Xu J, Duan X, Yang J, Beeching JR, Zhang P (2013) Enhanced reactive oxygen species scavenging by overproduction of superoxide dismutase and catalase delays postharvest physiological deterioration of cassava storage roots. Plant Physiol 161(3):1517–1528. https://doi.org/10.1104/pp.112.212803
Yan Y, Wang P, Lu Y, Bai Y, Wei Y, Liu G, Shi H (2021) MeRAV5 promotes drought stress resistance in cassava by modulating hydrogen peroxide and lignin accumulation. Plant J 107:847–860. https://doi.org/10.1111/tpj.15350
Yao NR, Goue B (1992) Water use efficiency of a cassava crop as affected by soil water balance. Agric For Meteorol 61:187–203. https://doi.org/10.1016/0168-1923(92)90049-A
Ye J, Yang H, Shi H, Wei Y, Tie W, Ding Z, Yan Y, Luo Y, Xia Z, Wang W, Peng M, Li K, Zhang H, Hu W (2017) The MAPKKK gene family in cassava: genome-wide identification and expression analysis against drought stress. Sci Rep 7(1):14939. https://doi.org/10.1038/s41598-017-13988-8
Yu Y, Cui YC, Ren C, Rocha PS, Peng M, Xu GY, Wang ML, Xia XJ (2016) Transgenic rice expressing a cassava (Manihot esculenta Crantz) plasma membrane gene MePMP3-2 exhibits enhanced tolerance to salt and drought stresses. Genet Mol Res 15(1):15017336. https://doi.org/10.4238/gmr.15017336
Zeng C, Ding Z, Zhou F, Zhou Y, Yang R, Yang Z, Wang W, Peng M (2017) The discrepant and similar responses of genome-wide transcriptional profiles between drought and cold stresses in cassava. Int J Mol Sci 18(12):2668. https://doi.org/10.3390/ijms18122668
Zhang P, Wang WQ, Zhang GL, Kaminek M, Dobrev P, Xu J, Gruissem W (2010) Senescence-inducible expression of isopentenyl transferase extends leaf life, increases drought stress resistance and alters cytokinin metabolism in cassava. J Integr Plant Biol 52(7):653–669. https://doi.org/10.1111/j.1744-7909.2010.00956.x
Zhao P, Liu P, Shao J, Li C, Wang B, Guo X, Yan B, Xia Y, Peng M (2015) Analysis of different strategies adapted by two cassava cultivars in response to drought stress: ensuring survival or continuing growth. J Exp Bot 66:1477–1488. https://doi.org/10.1093/jxb/eru507
Zhu Y, Luo X, Wei M, Khan A, Munsif F, Huang T, Pan X, Shan Z (2020) Antioxidant enzymatic activity and its related genes expression in cassava leaves at different growth stages play key roles in sustaining yield and drought tolerance under moisture stress. J Plant Growth Regul 39:594–607. https://doi.org/10.1007/s00344-019-10003-4
Zinsou V, Wydra K, Ahohuendo B, Schreiber L (2006) Leaf waxes of cassava (Manihot Esculenta Crantz) in relation to ecozone and resistance to Xanthomonas blight. Euphytica 149:189–198. https://doi.org/10.1007/s10681-005-9066-3
Zou Z, Yang J (2019) Genome-wide comparison reveals divergence of cassava and rubber aquaporin family genes after the recent whole-genome duplication. BMC Genom 20(1):1–16. https://doi.org/10.1186/s12864-019-5780-4
Author information
Authors and Affiliations
Contributions
SJM and KB: conceived the idea and design the outline of the first draft, designed the figures, and wrote the first draft. VR and RP: interpretation and critical revision. MKL: critical revision and designed the figures. AKC and KHMS: critical revision for important intellectual content and provided critical feedback. All the authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
More, S.J., Bardhan, K., Ravi, V. et al. Morphophysiological Responses and Tolerance Mechanisms in Cassava (Manihot esculenta Crantz) Under Drought Stress. J Soil Sci Plant Nutr 23, 71–91 (2023). https://doi.org/10.1007/s42729-023-01127-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42729-023-01127-4