
Abstract
As the green tactics for enhancing plant growth and production using naturally occurring materials are highly needed, it is important to use the nanoformulation of these materials as an attractive novel technique. Therefore, this research has been performed to evaluate the plants’ morphological traits, the qualitative parameters, and molecular genetic characteristics using random amplified polymorphic DNA (RAPD) of French basil independence on growth biostimulators and their nanocomposite. The treatments included normal formulations and nanocomposite formulation of humic acid (5 mM HA), salicylic acid (1.4 mM SA), and glycyrrhizic acid ammonium salt (0.4 mM GA) and control treatment (water application). The results show that foliar spray with HA, SA, GA, and their nanocomposites significantly increased (p ≤ 0.05) on all vegetative growth characters, photosynthetic pigments, oil yield/plant, mineral content, and antibacterial activity as compared with control plants. Also, 1,1-diphenyl-2-picrylhydrazyl (DPPH) values of different samples used varied from 70.63 to 74.93%, with a significant increase compared to untreated plants. The most marked increases have been observed in treated plants with biostimulants in the nanocomposites form than in the natural form. On the other hand, GA and its nanocomposite showed variable effects on basil plants and gave the lowest increase values in all parameters than the other biostimulant but have high antimicrobial activity. For the molecular study, ten selected primers displayed a total of 288 amplified fragments scored per primer ranging from 7 to 46 fragments; 157 bands were polymorphic with 69% polymorphism. It could be concluded that humic acid and its nanocomposite are the most effective biostimulants that increased plant productivity and oil content.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The rising trend in the use of medicinal plants worldwide is increasingly concerned about their cultivation and development. Since medicinal plants are more natural compatible, herbal treatments have recently received particular attention, and the use of medicinal plants has become widespread and popular. Basil (Ocimum basilicum L.) is cultivated in temperate tropical, subtropical, and warm climates. Egypt, France, Spain, and Hungary are the world’s main producers of basil (Jadczak et al. 2006). It belongs to the Lamiaceae family. Basil products are fresh and dried herbs used for seasoning, but their extracts and oil are utilized in the food and perfume industries (Purushothaman et al. 2018). Basil herbs and oils are typically used to treat a wide range of illnesses, such as headache, cough, diarrhea, constipation, warts, worms, breakdown of the kidneys, antispasmodic, carminative, expectorating, and antimalarial (Pandey et al. 2014). The herb contains 0.04–0.70% essential oil, and its specific aromas are dominated by linalool, methyl chavicol, 1,8-cineol, and eugenol (Purushothaman et al. 2018).
There is currently a great deal of interest in agriculture (horticulture) in natural products which will increase the yields and at the same time the biological value of the crop without having any harmful effects on the natural environment (El-Beltagi et al. 2020a, b). These products are plant growth biostimulants that can be used in sustainable agriculture successfully by improving plant productivity and using environmental limited resources (e.g., water) (Jardin 2015). Plant biostimulants are various compounds from a natural origin and sources that have positive effects on the regulation and improvement of plant metabolic processes, therefore, boosting their vigor, yield, and quality (Sofy et al. 2020a; Akladious and Mohamed 2018). In addition, biostimulants do not provide plants with sufficient quantities of essential nutrients but increase their absorption by the root system and boost the growth of a stressed plant because of increased antioxidants (Sofy et al. 2020b). Plant-based biostimulants were widely regarded as environmentally friendly farming and are now among the instruments used in sustainable agriculture.
Humic acid (HA) formed from humic substances, known as humus, is a common plant biostimulant that triggers plant growth and development through nutrition, hormones, and elicitor paths (Conselvan et al. 2017). In addition, HA is shown to increase phenylpropanoid biosynthesis in maize in addition to its primary role in the absorption of nutrients (Schiavon et al. 2010). Many have assumed that the addition of HA would boost the biosynthesis of secondary metabolites in medicinal plants.
Salicylic acid (SA) is among the most readily accessible plant growth regulators which is effective in other forms such as acetylsalicylic acid and methyl salicylate in plants as well (Raskin 1992). SA is known as a plant growth regulator and a phenolic compound and serves as the central regulator of the signaling network in environmental conditions (Naeem et al. 2020; El-Beltagi et al. 2017). SA is directly involved in plant growth, ion absorption, yield, improvement in the quality of the chlorophyll pigments, and activities of certain essential enzymes (Sofy et al. 2020b; Moustafa-Farag et al. 2020). In addition, SA serves as a stress messenger that stimulates the plant to resist the susceptibility of plants to infection by microorganisms (Klessig and Malamy 1994). The percentage of essential oil contents and oil yield/plant (ml) as a result of salicylic acid or humic acid treatments was reported by Khazaie et al. (2011) who mentioned that the use of concentrations from 0- to 300-ml/l humic acid gave the highest production of total essential oil of hyssop plants; also, Abdol Rahman et al. (2013) on cumin and Farhood et al. (2017) noticed that coriander plants treated with salicylic acid had higher seed oil content and yield compared with untreated plants.
Glycyrrhizic acid (GA) is a triterpene glycoside that is naturally produced in the roots of licorice plants (Glycyrrhiza glabra) and is a plant grown in Egypt and many other world countries. Licorice’s root extract contains many compounds that have a similar effect to those stimulating production such as minerals, phenolics, flavonoids, amino acids, and vitamins (Abd El-Azim et al. 2017). GA is the most effective active ingredient in licorice and has a variety of pharmacological and biological activities (Yong 2012). The use of bio-fertilizer or licorice root extracts for agriculture in sandy soils gave enhancement effects on the plant growth and essential oil of Foeniculum valgare Mill (Abd El-Azim et al. 2017).
Furthermore, the use of nanoparticles in plant science draws the researchers’ interest because of its greatest benefits (Sofy et al. 20020c). Currently, nano-agro-technology is targeted by nanoparticles (NPs) with specific properties for improving crop productivity (Batsmanova et al. 2013). Natural nanoparticles may boost the nonactivity of natural products against plant pathogens, raise fruit shelf life at room temperature, reduce pesticide impact and toxicity, and reduce the negative impact of the use of insecure nanoparticles (Shoala 2018). The mobility of nanoparticles is extremely high, leading to fast nutrient transport to all parts of the plants (Sofy et al. 2020c). The nanosized mineral nutrient formulation may enhance solubility and dispersal of nutrient insoluble in the soil, minimize soil absorption and fixation, and boost the nanostructured particle bioavailability. Thus, nano nutrients (nano fertilizers) improve the nutrient quality and the plant’s ability to use nutrients Naderi and Danesh-Shahraki (2013).
Molecular markers help to complement a morphological and physiological plant characterization; for example, because they have many benefits, they are abundant and independent of the environmental effects and early in cultivar identity during plant production (Mohamed and Abdel-Hamid 2013; Mohamed, 2014; Ashry et al. 2018; Mohamed et al. 2018). Random amplified polymorphic DNA (RAPD) markers amplify anonymous DNA products with single, short, and arbitrary primers. Due to their simplicity, RAPD has found a wide range of uses in many biological fields because no previous understanding of a DNA sequence is needed. A significant proportion of genetic polymorphism is associated with RAPD markers (Williams et al. 1990). In comparison to morphological and physiological markers, the number of DNA markers is not restricted and does not suffer from plant growth stages (Collard et al. 2005).
Based on novel results from the application of some nano biostimulator sprays on some quantitative and qualitative parameters of marjoram and basil (Amer and Shoala 2020; Hammam and Shoala 2020), this comparative study was designed to examine the effect of some biostimulators and their nanoparticles on growth and phytochemical parameters as well as molecular genetic characteristics using the RAPD marker.
Materials and methods
Preparation and characterization of the growth stimulators nanoparticles
Nano synthesis of humic acid
Humic acid was purchased from Sigma-Aldrich company; 22.7 mg of humic acid was dissolved in 100-ml water and sonicated (XUBA3Analogue Ultrasonic Bath, Grant Company) with an ultrasonic power and frequency of 50 kHz for 2 h at room temperature (25°C). The concentration was prepared and applied to 1 and 5 mM in the normal and nanoform.
Nano synthesis of salicylic acid
Salicylic acid was purchased from Sigma-Aldrich company; 10 mg of SA was dissolved in 10-ml absolute ethanol and sonicated (XUBA3Analogue Ultrasonic Bath, Grant Company) with an ultrasonic power and frequency of 50 kHz for an hour at room temperature (25°C). The concentration was prepared and applied to 0.7 and 1.4 mM in the normal and nanoform.
Nano synthesis of glycyrrhizic acid ammonium salt
Glycyrrhizic acid ammonium salt will be purchased from Sigma-Aldrich company; 0.1 mg of glycyrrhizic acid ammonium salt was dissolved in 1-ml absolute ethanol and sonicated (XUBA3Analogue Ultrasonic Bath, Grant Company) with an ultrasonic power and frequency of 50 kHz for an hour at room temperature (25°C). The concentration was prepared and applied to 0.2 and 0.4 mM in the normal and nanoform.
Characterization of nanomaterials by using dynamic light scattering (DLS)
Measurements of nano-humic acid nanoparticles (HA NPs), nano-salicylic acid nanoparticles (SA NPs), and nano-glycyrrhizic acid ammonium salt nanoparticles (GA NPs) distribution and size were performed by a dynamic light scattering method using Zetasizer Nano ZS (Malvern Instruments, UK) at room temperature. Prior to measurement, 30 μl of the nanoparticles were diluted with 3 ml of water at 25°C. Particle size data were expressed as the mean of the Z average of three independent batches of the nanoparticles.
Experimental material
The field experiment was carried out in the period 2019 and 2020 at the Farm of Medicinal and Aromatic Plants Research Department, El-Kanater El-Khairiya, Kalubeia Governorate, Egypt (30°19′ N, 31°13′ E, 16.9 m above sea level rise). French basil seeds (cv. Grand verde) have been obtained from Medicinal and Aromatic Plants Research Department, Dokki, Giza, Egypt.
Experimental design
Basil seeds were sown at the beginning of April in both seasons in a seedbed mixture of vermiculite and peat moss (2:1). When the basil seedling reached 10 cm in plant height (45 days old), they were transplanted at a spacing of 60 × 25 cm on one side of the row. The experimental plots were 3.0 × 2.50 m, with 3 rows at a distance of 60 cm between them. Phosphorus fertilizer was mixed with soil before transplanting (200 kg/fed as calcium superphosphate); potassium sulfate (as a source of K) was applied at 75 kg/fed. Nitrogen (N) was applied at a rate of 300 kg/fed using ammonium nitrate. Half doses of N and K were applied 21 days after planting, and the second one was applied 1 month later. The physical and chemical analyses of the experimental soil were texture class: clay, organic matter 2.17%, electrical conductivity (dS/m) 0.26, and pH 7.5. Irrigation and other field practices had been done as needed. The experimental protocol was based on a randomized complete block design, with three replicates. Each replicate was composed of seven plots, each plot representing one group. The first group was foliar sprayed with distilled water (control); the second group was foliar sprayed with HA NPs; the third group was foliar sprayed with normal HA; the fourth group was foliar sprayed with SA NPs; the fifth group was foliar sprayed with normal SA; the sixth group was foliar sprayed with GA NPs; and the seventh group was foliar sprayed with normal GA. All groups were foliar sprayed twice at 60 and 90 days after sowing and during growth stages and cultivation season of basil. Sprays were carried out by means of an atomizer sprayer in a way that all above-ground parts of rosemary plants were covered. To increase the absorption of solutions by plants, foliar application of the treatments was done in conditions without wind and rain and before sunrise when plant stomata are open. Two cuts were achieved: in the middle of both July and September at the full bloom stage by cutting the herb at a height of about 5 cm. The data were calculated on the basis of two cutting means. The plants of the second year were recorded on the gas chromatography with flame ionization detection (GC-FID) analysis of oil, mineral content, antioxidant activity, and molecular traits.
Vegetative growth characters
Plant height, number of branches per plant, and fresh and dry weight of herb (g/plant).
Biochemical analysis
Determination of photosynthetic pigments
The photosynthetic pigment contents were determined in the fresh leaves in accordance with the extraction procedure described by Frooq et al. (2016). The aqueous acetone, 85%, was used for photosynthetic pigment extraction from 10 mg of fresh leaf disks. The extinction was evaluated against a blank of a pure 85% acetone at wavelengths of 663, 645, and 452.5 nm for Chl a, Chl b, and total carotenoids, respectively. Calculations were made as mg/g tissue according to the equations proposed by Porra et al. (1989):
Determination of essential oil % and GC-FID analysis
The essential oil obtained was analyzed in the Laboratory of Medicinal and Aromatic Plants Research Department, Horticulture Research Institute, using a DsCrom 6200 gas chromatograph equipped with a flame ionization detector (GC-FID) for separation of volatile oil constituents. The analysis conditions were as follows: the chromatograph fitted with capillary column DP-WAX-122-7032 polysillphenylene–siloxane 30 × 0.25 mm ID × 0.25 μ film and the temperature program ramp increase rate of 13°C/min from 60°C to 220°C. Flow rates of gases were nitrogen at 1 ml/min, hydrogen at 30 ml/min, and 330 ml/min for air detector; and injector temperatures were 280°C and 250°C, respectively. Standard compounds α-pinene, β-pinene, limonene, linalool, camphor, α-terpinol, geranyl acetate, eugenol, and β-caryophyllene obtained from Sigma-Aldrich were used to the chemical compounds of the essential oils from the basil oil. The obtained chromatogram and report of GC-FID analysis for each sample were analyzed to calculate the percentage of main components of essential oils.
Determination of mineral content
The herb of basil was carefully washed with double-distilled water to remove all debris and apoplastic contents. The dried plant samples were further digested using the method proposed by Linder (1944). For digestion, 200 mg of powdered sample was taken which was then digested in aqua regia (H2SO4:HNO3:HClO4, v/v) in ratio 1:3:1 using glass beakers on a hot induction plate. After that, digested samples were cooled and filtered using 0.22-μm nylon syringe filters. The samples were further diluted using double-distilled water to make up the final volume to 50 ml. These digested samples were stored at room temperature and used for further analysis. Nitrogen was determined using the micro-Kjeldahl apparatus of Parnos–Wagner as described by Van Schouwenburg and Walinga (1978). Phosphorus was estimated colorimetrically by using the chlorostannous-reduced molybdophosphoric blue color method according to Chapman and Parker (1961). Potassium was determined using the flame photometer according to the method described by Brown and Lilliland (1964).
Determination of antioxidant activity (DPPH)
One gram of each sample was weighed in vials, each with a 20-ml capacity, using a Mettler Toledo MS304 sensitive balance. All vials containing samples and solvent (ethanol) were allowed to macerate for 3 days. Finally, the supernatants were collected for DPPH free-radical scavenging activity analysis as described by Alam et al. (2013). The solution for the 70% (v/v) methanol DPPH (1,1-diphenyl-2-picrylhydrazyl, Sigma-Aldrich) was prepared, resulting in a DPPH concentration of 0.25 mM. In 335 μl of DPPH solution, various volumes have been added (from 2 to 10 μl) to the methanol leaf extract, reaching a final volume of 1 ml. The mixture was vortexed and stayed 30 min in the dark at room temperature. By mixing 70% methanol with radical DPPH solution, a blank solution (control) was formed. The absorbance was read at 517 nm using a spectrophotometer and using ascorbic acid as standard. The radical scavenger activity was expressed in terms of the amount of antioxidants necessary to decrease the initial DPPH absorbance by 50% (IC50). The IC50 value for each sample was determined graphically by plotting the percentage disappearance of DPPH as a function of the sample concentration.
The DPPH radical scavenging (%) was calculated using the formula:
\( =\frac{\mathrm{A}0-\mathrm{A}1}{\mathrm{A}0}\times 100 \)
where A0 is the absorbance of the control and A1 is the absorbance of the sample.
IC50 refers to its concentrations for 50% inhibition.
Antibacterial activity for essential oil
The paper disc diffusion method was used to determine antibacterial activities of essential oils (Berghe and Vlietinck 1991; Cappuccino and Sherman 1998). The gram-positive (Bacillus sp.) and gram-negative (Pseudomonas sp.) bacteria were used in these experiments. Nutrient agar medium has held the cultures of microorganisms. Briefly, microorganism suspensions were distributed over solid medium plates of Muller–Hinton (107–108 colony-forming units [CFU]/ml). Filter paper discs, 4-mm diameter (Whatman No. 1), were individually impregnated with 10 μl of each essential oil extracted from each treatment and then placed onto the surfaces of the inoculated plates. Positive antibacterial was formed by the presence of the measurable inhibition zone at the end of the incubation period (24 h at 37°C for bacteria) and registered in its width (mm) that comprises a disc diameter. Three replicates were conducted for each test.
Molecular analysis
DNA isolation
Leaves of seven French basil were collected for DNA isolation by Doyle and Doyle (1987) method.
RAPD analysis polymerase chain reaction (PCR)
RAPD-PCR analyses were performed as described by Williams et al. (1990) with some modifications. Amplifications were carried out in an Eppendorf Mastercycler Gradient Thermocycler. In total, 10 oligonucleotides arbitrary primers from Operon Technologies Inc. (Alameda, CA, USA) were used for RAPD amplification. Ten primers were selected based on the repeatable and scorable banding patterns to detect the polymorphisms of French basil. Polymerase chain reactions were performed in a final volume of 20 μl containing 50-ng DNA template, 4-μl Taq buffer, 10 mM each dNTP, Operon Primer Technologies, and 5 units of Taq DNA polymerase (Sigma). RAPD-PCR conditions were as follows: initial DNA denaturation for 5 min at 94°C and DNA amplification for 35 cycles (30 s at 94°C, 45 s at 37°C, and 1 min 30 s at 72°C). In the end, the reaction products were stored at 4°C after a concluding elongation for 5 min at 72°C. After PCR amplification, amplified DNA fragments were separated by gel electrophoresis on a 2% (w/v) agarose gel, in a 1× TAE buffer solution for 2 h at 4 V/cm. The gels were stained for 15 min with ethidium bromide (5 mg/ml) and visualized under UV transillumination (UV Star 365 nm, Biometra). The reproducibility of the amplification products was checked for each primer. Table 1 lists the RAPD primer sequences producing informative polymorphic bands.
Statistical analysis
The experimental design was completely randomized, and statistical analysis was performed using SPSS (Social Science version 26.00) statistical software at a significance level of 0.05 (Gomez and Gomez 1984). Quantitative analyses were obtained using one-way ANOVA with Fisher’s post hoc test variance analysis with the parametric distribution of Levene’s sample. The confidence interval was set at 95% and the negotiated margin of error at 5%. The genetic similarity between individual genotyping pairs was analyzed using the computer NTSYSpc 2.1 (Rohlf 2008) and was determined by comparing all basil treatments with MEGA 5.2.2 in a similar manner.
Results
Dynamic light scattering (DLS)
Dynamic light scattering technique was performed to understand the size distribution and the stability of prepared HA, SA, and GA nanoparticles, showing that the size distribution ranges mainly within 100–110, 20–26.6, and 5–10 nm, respectively, as shown in Figs. 1, 2, and 3. The zeta potential is a key indicator of the stability of colloidal dispersions. So colloids with high zeta potential (positive) are electrically stabilized (Figs. 4, 5, and 6).

Z average size of HA NPs, 100–110nm

Z average size of SA NPs, 26.6 nm

Z average size of GA NPs, 5–10nm

Zeta potential of HA NPs

Zeta potential of SA NPs nanoparticles

Zeta potential of GA NPs
Vegetative growth characters
The studied plant vegetative growth parameters detected significant effects concerning the foliar spray with the different treatments (Table 2). In comparison with control plants, all treatments cause a substantial boost in all plant growth characteristics. The maximum values for both seasons of plant height were about 23.2% and 28.05%; the number of branches/plant (87.4% and 52.4%), fresh weight/plant (172.2% and 164.9%), and dry weight/plant (151.7% and 146.6%) were obtained from a plant treated with HA NPs. In addition, in both seasons, foliar spray with GA illustrated the lowest increases in plant height, about 10.9% and 7.4%; the number of branches/plant (20.4% and 10.5%); fresh weight/plant (5.7% and 3.8%); and dry weight/plant (11.0% and 10.6%) as compared with control plants. Nanocomposite biostimulants were observed to cause the most marked increase in vegetative growth characteristics than natural biostimulants.
Biochemical analysis
Photosynthetic pigments
The treatment with HA, HA NPs, SA, SA NPs, GA, and GA NPs caused substantial increases in Chl a, Chl b, carotenoid, and total chlorophyll as compared to control plants (Table 3). Moreover, treatment with GA in the two seasons showed the lowest increases in Chl a (16.3% and 13.3%), Chl b (36.5% and 28.4%), carotenoid (25.8% and 25.6%), and total chlorophyll content (22.6% and 19.9%) as compared to untreated plants. The higher values of Chl a are about (54.4% and 48.9%); Chl b (84.1% and 82.1%), carotenoid (74.2% and 75.0%), and total chlorophyll content (65.8% and 63.0%) were recorded in plants treated with HA NPs in both seasons. Moreover, the foliar plants sprayed with nanocomposites have seen the most marked increases in chlorophyll content.
Essential oil content and GC-FID analysis
The percentage of essential oil content ranged from 0.12 to 0.32% in the first season and 0.14 to 0.34% in the second season (Table 4). All biostimulants in the natural form or nanocomposite form caused a considerable increase in % of oil and oil yield/plant. The highest values (0.32% and 0.34%) were observed in the treatment with HA NPs, while the lowest values (0.16 and 0.18%) were verified in the plants treated with natural biostimulant SA and GA in both growing seasons. Moreover, Table 4 indicated that the uppermost values from oil yield per plant were 89.5 and 84.8 and 72.5 and 79.8 ml/plant and were shown in the application of HA NPs and HA, respectively, followed by SA NPs (49.2 and 54.9 ml/plant).
With regard to GC-FID analysis, it is clear that there were varying chemical compositions of essential oil in the treated basil plants (Fig. 7). The results of GC-FID analysis of the French basil volatile oils in the second season showed that the major compounds identified were alpha-pinene, beta-pinene, limonene, linalool, camphor, alpha-terpinol, geranyl acetate, eugenol, and beta-caryophyllene (Fig. 7). The oxygenated monoterpenes (linalool) are considered the main components, and their content ranged from 51.02% in the control to 57.30% in the treatment of HA natural. Monoterpene hydrocarbons (limonene) showed content ranging from 13.38 for HA to 15.82 for SA NPs %, and the range of monoterpene ketone (camphor) being 5.12% for SA NPs to 9.41% for HA. The ß-caryophyllene compound (sesquiterpene hydrocarbons) was found and ranged from 5.63% in plants treated with HA NPs % to 8.79% in the control.

Effect of HA, SA, GA, and their nanocomposite on GC-FID analysis of the essential oil of French basil during the second growing season
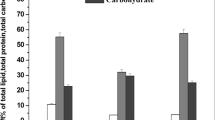
Mineral content
Data demonstrated in Fig. 8 clearly indicated significant differences between the treatments in N, P, and K % contents of basil dried leaves relative to control. In plants treated with foliar application of HA NPs, they showed the highest amounts of N, P, and K (5.06%, 1.46%, and 6.53%, respectively); while the SA NPs and GA NPs were the favored treatment for increasing nitrogen (4.87% and 4.57%), phosphorus (1.05% and 0.98%), and potassium (4.09% and 3.65%) as compared to control plants.

Effect of HA, SA, GA, and their nanocomposite on mineral content (%) in dried basil leaves during the second growing season. According to Fisher’s test, the different letters are significantly different between treatments at the 0.05 level. Vertical bars represent the means of three independent determinations ± standard error (SE)
Antioxidant DPPH scavenging activity
The free-radical scavenging activity of basil leaves was measured by the DPPH assay. The results of antioxidant activity of the extracted basil leaves of plants sprayed by the treatments are reported in Fig. 9. DPPH values of different samples used varied from 70.63 to 74.93%, with a significant increase compared to untreated plants. The highest DPPH scavenging activity corresponds to the plants foliar sprayed with HA NPs, being of 74.93% with IC50 (42.2 μg/ml), followed by the treatment of SA NPs and GA NPs which showed 73.89% and 72.74% with IC50 (45.5 and 49.6 μg/ml), respectively, as compared to control plants (70.63%) with IC50 (55.3 μg/ml), while the other treatments showed slight increases in DPPH.

Effect of HA, SA, GA, and their nanocomposite on antioxidant activity (%) in dried basil leaves during the second growing season. According to Fisher’s test, the different letters are significantly different between treatments at the 0.05 level. Vertical bars represent the means of three independent determinations ± standard error (SE)
Antibacterial activity of essential oil
Using the filter paper disc method for the examination of antibacterial activity of the essential oils extracted from different treatments against the reference bacteria of gram-positive Bacillus sp. and gram-negative Pseudomonas sp., Fig. 10 shows images of zones of inhibition around filters loaded with oils extracted from the different plant treatments and were found to inhibit bacterial strains with different inhibition degrees based on inhibition zone size, the essential oil extracted from basil treated with SA NPs followed by GA NPs, showing the highest antibacterial activity of the tested bacteria of gram-negative Pseudomonas sp., while the oil extracted from basil treated with HA NPs, GA, SA NPs, and SA showed the highest antibacterial activity of the tested bacteria of gram-positive Bacillus sp. (Table 5). Moreover, the inhibition zone observed on the extracted oil from basil treated with HA natural indicated that it had the minimum antibacterial properties of gram-negative Pseudomonas sp. and positive bacteria Bacillus sp. On the other hand, Bacillus sp. and Pseudomonas sp. were sensitive strains to the essential oils extracted from untreated basil plants.

Image showing inhibition zones of the microbial growth of tested microorganisms: a gram-positive (Bacillus sp.) and b gram-negative (Pseudomonas sp.) as affected by the different treatments
Molecular analysis
RAPD markers analysis
Genetic analyses were derived directly from differences in the plant genome. The genetic relationships of French basil treated with HA, SA, GA, and their nanocomposites using RAPD markers were identified during the present research. Ten RAPD primers were used, 228 bands were obtained, and 157 bands were polymorphic with 69% polymorphism. Total bands, polymorphic bands, and unique bands were presented in Tables 6, 7, 8, and 9 and Fig. 11. Primer BA-10 had the largest number of bands (46 bands), and primer AN-01 have the lowest number of bands (7 bands). The data in Table 6 show that the highest number of bands was detected in plants foliar sprayed with HA NPs about 36 bands, followed by GA NPs which recorded 34 bands than SA NPs which showed 33 bands.

Amplification patterns of basil plants treated with HA, SA, GA, and their nanocomposites generated using arbitrary different primer. M, marker; 1, HANP; 2, HA; 3, SANP; 4, SA; 5, GANP; 6, GA; 7, control
The highest polymorphism was obtained by primer O-16 with polymorphism 76.7%, and the lowest polymorphism was 6.7% with primer D-12. Control characterized with 4 unique bands: three bands were obtained with primer C-20 at (130-720-950 bp) and one band was detected with primer Z-10 at 800 bp. GA was characterized by four unique bands: two bands were detected with primer BA-10 at 500 and 900 bp, one with primer O-01 at 700 bp, and one with primer Z-04 at 650 bp. While HA NPs were characterized with two unique bands at 950 and 400 bp with primer BA-10 and D-10, respectively. Unique bands 400, 480, and 700bp characterized plants treated with HA, SA NPs, and SA, respectively, with primer C-20, BA-10, and D-12.
A significant genetic variation was identified within available basil treatments with genetic similarities between 40% and 80%. The highest genetic similarity is 80% between GA NPs and SA, and the lowest similarity is 40% between GA and HA NPs. The dendrogram resulted by RAPD marker divided French basil treated with nanomaterial into two groups each having two subclusters: the first cluster has two groups, the first has HA and GA and the second group has SA NPs and GA NPs. While the second cluster has two subclusters, separating control in one subcluster and the other subcluster has HA NPs and SA (Fig. 12).

The UPGMA dendrogram based on RAPD marker for basil treated with HA, SA, GA, and their nanocomposite
Discussion
The use of fertilizers has exponentially increased in recent years worldwide, causing severe environmental problems. The increment use of fertilizer in the soil and plant systems may accumulate heavy metal and may reach the food supply chain. So it is very important to use nano fertilizer to reduce the aggregation in soil. Natural or synthetic nanoparticles are materials of 1 to 100 nm in size (Kelkar et al. 2014). Due to their high surface area, high reactivity, pores, and morphology, nanoparticles have special physical–chemical properties. A major scope is opened for new applications in the biotechnological, agricultural, and fertilizer industries (Siddiqui et al. 2015).
In comparison with control plants, foliar spray with HA, SA, GA, and their nanoparticles caused enhancement in growth characteristics, photosynthetic pigments, oil %, and mineral content (N, P, K %) as compared to control plants. The biostimulants in the form of nanocomposites were more effective than in the natural form. These findings are related to Roosta et al. (2015) who observed that the use of various levels of nano-fertile fertilizer containing 60% humic acid boosts the fresh and dry weight of shoot and root, chlorophyll, and soluble sugar contents of mint plants as related to untreated plants. Enhancement of plant production and improved physiological process may be associated with increased content in plants of K, P, Ca, Fe, Mn, and Cu. In addition, salicylic acid nanoparticle pretreatment has substantially boosted the shoots and roots fresh weight of Isatis cappadocica (Souri et al. 2017).
Earlier studies on humic acid influence relate to improving root nutrition via various mechanisms. An important contribution of humic acids to root nutrition is the increased absorption and translocation of nutrients, which also help to stimulate plasma membrane H+-ATPases, transferring the free energy released through ATP hydrolysis into the transmembrane electrochemical potential used to supply nitrate and other nutrients (Jardin 2015). In addition to nutrient absorption, ATPase membrane pump protons also lead to the loosening of cell walls, and expansion and elongation of cells which lead to an increase in plant growth and chlorophyll content (Afify et al. 2012; Abdelgawad et al. 2018;). Also, the use of humic acid induces changes in plant gene expression and the content of the primary and secondary metabolite compounds (Jindo et al. 2020) that participate in various plant physiological processes (e.g., Krebs cycle, metabolism, photosynthesis) (Conselvan et al. 2018; Sofi et al. 2018).
SA NPs may have a positive impact on the growth criteria as it may affect germination rate, cell proliferation, respiration, stomatal closure, and fruit yield (Yusuf et al. 2013; Khokon et al. 2011) and increase phenolics accumulation (Kovácik et al. 2009) and enhance the oxidative stress tolerance (Li et al. 2014). This is due to the fact that the synthesis or signaling of some hormones, such as ethylene, jasmonic acid, and auxin, is modulated indirectly by SA (Bagherifard et al. 2015). Moreover, enhanced vegetative growth, nutrient uptake, photosynthesis, and changes in the number of secretory glands of essential oil in the leaves and flowers may lead to a boost production of essential oils (Gharib 2007). The role of SA in boosting rubisco activity, photosynthesis, and chlorophyll content (Gao et al. 2012) and thus improving the output of dry matter can be a reason for increasing the secretory glands of essential oil (Singh and Usha 2003). In some studies, secondary metabolites of plants may be affected by SA application and therefore cause an increment in the essential oils in Ocimum basilicum L. (Mirzajani et al. 2015) and triterpenes in Nigella sativa (Elyasi et al. 2016).
GA is abundant in antioxidants; amino acids; prolines; soluble sugars; salicylic acid; ascorbic acid; glutathione; vitamins; selenium; phytohormones including substantial quantities of auxin, gibberellins, and cytokinins (zeatin); and nutrients (Thanaa et al. 2016). The positive effect of GA and GA NPs may be due to its effect in increasing endogenous hormones such as GA3 in treated plants which increased the metabolic processes role and its effect on the mineral content in tissues (Al-Obady 2015; El-Gohary et al. 2021). The spraying of plants with the licorice extract resulted in increased flowering (%) of the plant. This increase may be due to the similarity of licorice extract in its behavior with GA3 in stimulating flowering, increasing plant height, number of branches, and decreasing the fresh and dry weight of plants. It contains mephalonic acid which improves vegetative growth as a result of stimulating the enzymes necessary for the conversion of complex compounds into simple compounds and energy-efficient processing required for plant growth (Cuong et al. 2017). The application of licorice extract maintains a suitable level of endogenous GA in various phases for stimulated growth which activates cell division and lengthening and stay-green effect of cytokinins found in licorice extract, which promotes photosynthesis (Abd El-Hamied and El-Amary, 2015). In addition, the low concentrations of GA have numerous cellular benefits such as antioxidants and antimicrobial effects (Wang et al. 2015).
All biostimulants in the natural form or nanocomposite form caused a considerable increase in % of oil and oil yield/plant. These results are in accordance with Talaat et al. (2014), reporting that in plants treated with phenolic compounds such as salicylic and amino acids, the fruit production and oil yield (ml plant−1) increased. The increase in the percentage of oil and protein may be caused by increasing vegetative growth and absorption of nutrients. In addition, Noreen and Ashraf (2010) referred to the high concentrations of salicylic acid led to marked rises in the content of the sunflower achene oil and certain important fatty acids. The effect of different treatments on essential oil and its constituents may be due to its effect on enzyme activity and metabolism of essential oil production (Talaat et al. 2014). Also, SA may change secondary metabolites and their pathway by effects on plastid, chlorophyll level, and tolerate condition stress. The SA-like stress manipulated the quality and quantity of essential oil of Salvia macrosiphon. The yield of essential oil was increased (Talaat et al. 2014). Furthermore, the rise of the humic acid concentration in comparison with the control increased the essential oil content of Thymus vulgaris. The oil content of essential oil ranged from 0.8 (control) to 2.0% (75 g m−2) (Noroozisharaf and Kaviani 2018).
Basil leaves contain a number of secondary metabolites such as flavonoids, alkaloids, phenol, and essential oil. The main group of phenolic compounds consists of flavonoids. Due to its antioxidant ability, the interest in phenolic and flavonoid compounds increased over the last 10 years. Their radical-scavenging features help avoid stress-related chronic and oxidative stresses (Mohamed et al. 2016, 2018a, b). The antioxidants activity of basil oil may be due to that, containing high concentrations of flavonoids, phenolic, and methyl eugenol in basil (3.68%) (Warsi and Sholichah 2017). The highest DPPH scavenging activity corresponds to the plants foliar sprayed with HA NPs, followed by the treatment of SA NPs and GA NPs as compared to control plants, while the other treatments showed slight increases in DPPH. Similar findings showed that SA promotes the antioxidant activity of Alternanthera tenella leaves and can be linked to an increase in betacyanin contents which are compounds with an antioxidant action recognized (Rodrigues-Brandão et al. 2014). Moreover, yellow pepper treated with biostimulants had been significantly higher in DPPH and ABTS (p < 0.05) (Paradikovi et al. 2011).
In the antibacterial activity test, our findings showed different sensitivity to the bacteria used. Essential oils are related specifically to their chemical composition by their antibacterial activity (Akhtar et al. 2014; Swamy et al. 2016). Essential oils are a complex combination of compounds from plants that are commonly known for their scents, flavors, primarily monoterpenes, sesquiterpenes, and their corresponding oxygenated derivatives (alcohols, aldehydes, esters, ethers, ketones, phenolics, and oxides). Previous studies indicate that the key component that is responsible for antibacterial activities is oxygenated monoterpenes; linalool, oxygenated monoterpene (Chaturvedi et al. 2018; Hussain et al. 2008) which affect cell membrane integrity and function, alter membrane potential, causing loss of cytoplasmic material and obstructing the respiratory chain (Greay and Hammer 2015). Studies have shown that camphor has substantial antifungal activity along with 1,8-cineole, as shown against Candida albicans and Candida krusei (Viljoen et al. 2003). The antimicrobial activity exhibited may have a major or minor component present in the essential oil (Lopes-Lutz et al. 2008). Most studies promote direct cell membrane disruption and concentration-dependent cytotoxic effects, though some studies indicate that the essential oil components are able to penetrate the microorganism and respond to enzyme sites and/or interfere with cell metabolism (Aly et al. 2013).
In addition, our findings suggest that gram-positive bacteria are more susceptible to basil oil than gram-negative bacteria. Besides, some oils seemed more specific to inhibit gram-positive bacteria, but gram-negative bacteria are often reported to be more resistant to essentially plant-based oils (Smith-Palmer et al. 1998; El-Beltagi et al. 2018, 2019a, b). Lipopolysaccharides significantly constitute the hydrophilic cell wall structure of gram-negative bacteria which is able to spread out the hydrophobic oil and prevent essential oils from accumulating in the target cell membrane (Bezic et al. 2003). This is the reason that gram-positive bacteria were found more susceptible to essential oils, methanol extracts, and various subfractions of basil than gram-negative bacteria (Hossain et al. 2010)
In the RAPD markers analysis in the different treatments of basil plants, a total of 10 RAPD primers were included in the evaluation and a total of 228 bands were obtained. DNA molecular analysis is commonly used in herbal medicinal analysis for the authentication of medicinal plant species (Giachino et al. 2014). The base sequences, total band numbers, and polymorphic band numbers of polymorphic band ratios are presented in Tables 6, 7, and 8. Giachino et al. (2014) used 24 RAPD primers to characterize essential oil in Turkish basil, and 273 out of a total of 375 bands provided by RAPD primers were polymorphically observed and genetic similarity ranging from 46 to 87%. The polymorphic band ratios of primers varied from 6.7 (D-12) to 76.7% (O-01). These values indicate that there is a large variance in the polymorphic band ratio of primers. The genetic similarity index values among the basil genotypes were stated in the RAPD study by De Masi et al. (2006), ranging from 0.64 to 1.00. Genetic similarity values for 12 adhesions of Ocimum gratissimum present in previous analysis have shifted between 0.20 and 067 Vieira and Simon (2001). Vieira et al. (2003) have also identified a narrow genetic variation of RAPD markers in O. basilicum genotypes.
The results have shown that foliar spray with the biostimulants affected the genome response of basil plants. The highest number of bands was detected in plants foliar sprayed with HA NPs of about 36 bands followed by GA NPs which recorded 34 bands than SA NPs which showed 33 bands. Also, the total number of bands in natural biostimulants reported the highest number in plants foliar spraying with GA (35 bands) and then HA (33 bands) and SA (30 bands) as compared with untreated plants (30 bands). Similar results are reported by Ražná et al. (2018) who found that seed priming with the plant-derived elicitor affected the genome response of wheat. DNA fingerprinting generated by RAPD polymorphism has sensitively reflected different concentrations of cobalt diglycyrrhizinate, while this response was tissue specific, as confirmed by several experiments. Priming treatments were found to induce DNA replication in the embryo root meristems. A significant difference was observed in DNA content in priming seeds when compared with the unprimed seeds (Ražná et al. 2018). The significant effect of cobalt diglycyrrhizinate seed priming was observed and represented genomic changes of the wheat genomes detectable by RAPD molecular markers. These changes located in primer binding sites create polymorphism. The changes caused by the elicitor’s treatment may be due to the differential expressions of plant genes, which do not have to directly relate to plant protection mechanisms. In this way, the ontogenesis or metabolism of the plants is regulated (Chandra and Gunasekaran 2017). In addition, data palm offshoot (Phoenix dactylifera L.) cultivars showed that RAPD technology shows polymorphism between some of the treatments such as salicylic acid and humic acid. Differences in the primers’ connection with DNA or for the deletion or addition of a base or number of nitrogen base composites composed of a bar of DNA or of a replacement, reversing and rearrangement processes will result from those polymorphisms in the DNA fragments (Shareef et al. 2017). This would serve to activate specific genes (gene on) or silence other genes (gene off) (Kunert et al. 2003).
For instance, in dendrograms, all genotypes were located in two clusters (Fig. 12, cluster I: control, HA NPs, and SA; cluster II: HA, GA, SA NPs, and GA), Furthermore, the RAPD analysis clustered the genotypes in cluster I: cluster control C separated from HA NP and SA; and cluster II have two groups: one of them has HA and GA and the other group has SA NP and GA NP. De Masi et al. (2006) observed the two separated dendrograms by using the Euclidian and unweighted pair group method with arithmetic mean approaches in essential oil and RAPD basil genotype analysis. In addition, Trindade et al. (2008) observed that two distinct clusters of Thymus caesptitius essential oils were obtained by using RAPD markers and genetic analysis.
Conclusion
HA, SA, GA, and their nanocomposites are realistic techniques to increase the productivity characteristics of basil crops and essential oil. All the biostimulants used increased vegetative growth, chlorophyll content, mineral content, antioxidant activity, and antimicrobial activity (Fig. 13). The most pronounced increases were observed in plants treated with humic acid. In addition, the biostimulants in nanocomposites form are more effective than in natural form. The results indicate that RAPD markers are very useful to study the effect of different biostimulants on basil. These data will make breeding studies of the basil species and treatments substantially easier. On the basis of the results of the current study, it appears that using nanocomposites from biostimulants that play an important role in sustainable agriculture will reduce the use of chemical fertilizers that increased environmental pollution.

Model showing the effects of HA, SA, GA, and their nanocomposite on the plant
Abbreviations
- RAPD:
-
random amplified polymorphic DNA
- HA:
-
humic acid
- HA NPs:
-
nanocomposite of humic acid
- SA:
-
salicylic acid
- SA NPs:
-
nanocomposite of salicylic acid
- GA:
-
glycyrrhizic acid
- GA NPs:
-
nanocomposite of glycyrrhizic acid
- DNA:
-
deoxyribonucleic acid
- DPPH:
-
1,1-diphenyl-2-picrylhydrazyl
- PCR:
-
polymerase chain reaction
- DLS:
-
dynamic light scattering
References
Abd El-Azim WM, Khater RMR, Badawy MYM (2017) Effect of bio-fertilization and different licorice extracts on growth and productivity of Foeniculum vulgare. Mill Plant Middle East J Agri Res 06(01):01–12
Abd El-Hamied SA, El-Amary EI (2015) Improving growth and productivity of “pear” trees using some natural plants extracts under North Sinai conditions. J Agri Veter Sci 8(1):1–9
Abdelgawad KF, Mhmoud AA, Mohamed HFY (2018) Foliar spraying with some biostimulants improves growth, chemical constituents, and yield of head lettuce plant Middle East. J Agric Res 7(4):1268–1277
Abdol Rahman R, Asad R, Sheno A (2013) Effect of salicylic acid and methyl jasmonate on growth and secondary metabolites in Cuminum cyminum L. J Biod Environ Sci 3(12):140–149
Afify AMR, El-Beltagi HS, Aly AA, AE E-A (2012) Antioxidant enzyme activities and lipid peroxidation as biomarker for potato tuber stored by two essential oils caraway and clove and its main component carvone and eugenol. Asian Pac J Trop Biomed 2:S772–S780
Akhtar MS, Degaga B, Azam T (2014) Antimicrobial activity of essential oils extracted from medicinal plants against the pathogenic microorganisms: a review. Bio Sci Pharma Res 2(1):1–7
Akladious SA, Mohamed HI (2018) Ameliorative effects of calcium nitrate and humic acid on the growth, yield component and biochemical attribute of pepper (Capsicum annuum) plants grown under salt stress. Sci Hortic 236:244–250
Alam MN, Bristi NJ, Rafiquzzaman M (2013) Review on in vivo and in vitro methods evaluation of antioxidant activity. Saudi Pharm J 21:143–152
Al-Obady RM (2015) Effect of foliar application with garlic extract and liquorice root extract and salicylic acid on vegetative growth and flowering and flower set of tomato and under unheated houses. J Appl Sci Res 3(1):11–22
Aly AA, Mohamed HI, Mansour MTM, Omar MR (2013) Suppression of powdery mildew on flax by foliar application of essential oils. J Phytopathol 161:376–381
Amer A, Shoala T (2020) Physiological and phenotypic characters of sweet marjoram in response to hydrogen peroxide and chitosan nanoparticles treatments. Sci Hortic 268:109374
Ashry NA, Ghonaim MM, Mohamed HI, Mogazy AM (2018) Physiological and molecular genetic studies on two elicitors for improving the tolerance of six Egyptian soybean cultivars to cotton leaf worm. Plant Physiol Biochem 130:224–234
Bagherifard A, Bagheri A, Sabourifard H, Bagherifard G, Najar M (2015) The effect of salicylic acid on some morphological and biochemistry parameters under salt stress in herb artichoke (Cynara Scolymus L.). Res J Fish Hydrobiol 10:745–750
Batsmanova LM, Gonchar LM, Taran NY, Okanenko AA (2013) Using a colloidal solution of metal nanoparticles as micronutrient fertiliser for cereals. In: Proceedings of the international conference on nanomaterials: applications and properties, Crimea, Ukraine, Proc NAP2, 04NABM14
Berghe DAV, Vlietinck AJ (1991) Screening methods for antibacterial and antiviral agents from higher plants. Plant Biochem 6:47–69
Bezic N, Skocibusic M, Dinkic V, Radonic A (2003) Composition and antimicrobial activity of Achillea clavennae L. essential oil. Phytother Res 17:1037–1040
Brown JD, Lilliland O (1964) Rapid determination of potassium and sodium in plant material and soil extracts by flame photometer. Proc Amer Soc Hort Sci 48:341–346
Cappuccino JG, Sherman N (1998) Microbiology: a laboratory manual, 5th ed.; Benjamin/Cumming Science. Publishing: San Francisco, CA, USA, 254.
Chandra JH, Gunasekaran H (2017) Screening of phytochemical, antimicrobial and antioxidant activity of glycyrrhiz glabra root extract. J Environ Biol 138:161–165
Chapman HD, Parker F (1961) Methods of analysis for soil, plant and water. J Plant Nutr 22(1):121–128
Chaturvedi T, Kumar A, Kumar A, Verma RS, Padalia RC, Sundaresan V, Chauhan A, Saikia D, Singh VR, Venkatesha KT (2018) Chemical composition, genetic diversity, antibacterial, antifungal and antioxidant activities of camphor-basil (Ocimum kilimandscharicum Guerke). Industrial Crops Products 118:246–258
Collard BCY, Jahufer MZZ, Brouwer JB, Pang ECK (2005) An introduction to markers, quantitative trait loci (QTL) mapping and marker-assisted selection for crop improvement: the basic concepts. Euphytica 142:169–196
Conselvan GB, Pizzeghello D, Francioso O, Di Foggia M, Nardi S, Carletti P (2017) Biostimulant activity of humic substances extracted from leonardites. Plant Soil 420:119–134
Conselvan GB, Fuentes D, Merchant A, Peggion C, Francioso O, Carletti P (2018) Effects of humic substances and indole-3-acetic acid on arabidopsis sugar and amino acid metabolic profile. Plant Soil 426:17–32
Cuong TX, Ullah H, Datta A, Hanh TC (2017) Effects of silicon-based fertilizer on growth, yield and nutrient uptake of rice in tropical zone of Vietnam. Rice Sci 24(5):283–290
De Masi L, Siviero P, Esposito C, Castaldo D, Siano F, Laratta B (2006) Assessment of agronomic, chemical and genetic variability in common basil (Ocimum basilicum L.). Eur Food Res Technol 223:273–281
Doyle JJ, Doyle JL (1987) A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochemical Bull 19:11–15
El-Beltagi HS, Ahmed SH, Namich AAM, Abdel-Sattar RR (2017) Effect of salicylic acid and potassium citrate on cotton plant under salt stress. Fresenius Environ Bull 26(1a):1091–1100
El-Beltagi HS, Mohamed HI, Safwat G, Megahed BMH, Gamal M (2018) Evaluation of some chemical constituents, antioxidant, antibacterial and anticancer activities of Beta vulgaris L. root. Fresenius Environ Bull 27(9):6369–6378
El-Beltagi HS, Mohamed HI, Elmelegy AA, Eldesoky SE, Safwat G (2019a) Phytochemical screening, antimicrobial, antioxidant, anticancer activities and nutritional values of cactus (Opuntia Ficus Indicia) pulp and peel. Fresenius Environ Bull 28(2A):1534–1551
El-Beltagi HS, Mohamed HI, Safwat G, Gamal M, Megahed BMH (2019b) Chemical composition and biological activity of Physalis peruviana L. Gesunde Pflanzen 71:113–122
El-Beltagi HS, Sofy MR, Aldaej MI, Mohamed HI (2020a) Silicon alleviates copper toxicity in flax plants by up-regulating antioxidant defense and secondary metabolites and decreasing oxidative damage. Sustainability 12(11)
El-Beltagi HS, Mohamed HI, Sofy MR (2020b) Role of ascorbic acid, glutathione and proline applied as singly or in sequence combination in improving chickpea plant through physiological change and antioxidant defense under different levels of irrigation intervals. Molecules 25:1702. https://doi.org/10.3390/molecules25071702
El-Gohary AE, Wahba HE, Hendawy SF, Hussein MS (2021) Effect of licorice root and cabbage leaf extracts as a natural fertilizer on growth and productivity of Cynara cardunculus L. Egyptian Pharmace J 20:17–22
Elyasi R, Majdi M, Bahramnejad B, Mirzaghaderi G (2016) Spatial modulation and abiotic elicitors responses of the biosynthesis related genes of mono/triterpenes in black cumin (Nigella sativa). Ind Crop Prod 79:240–247
Farhood Y, Saeid ZS, Jalil SK (2017) Evaluation of some morphological traits and oil content of coriander seeds in response to bio-fertilizer and salicylic acid under water stress. J Biod Environ Sci 10(1):140–149
Frooq MA, Ali S, Hameed A, Bharwana SA, Rizwan M, Ishaque W, Farid M, Mahmood K, Iqbal Z (2016) Cadmium stress in cotton seedlings: physiological, photosynthesis and oxidative damages alleviated by glycinebetaine. S Afr J Bot 104:61–68
Gao Z, Meng C, Zhang X, Xu D, Miao X, Wang Y, Yang L, Lv H, Chen L, Ye N (2012) Induction of salicylic acid (SA) on transcriptional expression of eight carotenoid genes and astaxanthin accumulation in Haematococcus pluvialis. Enzym Microb Technol 51:225–230
Gharib FA (2007) Effect of salicylic acid on the growth, metabolic activities and oil content of basil and marjoram. Inter J Agri Biol 4:485–492
Giachino RRA, Sönmez Ç, Tonk FA, Bayram E, Yüce S, Telci I (2014) RAPD and essential oil characterization of Turkish basil (Ocimum basilicum L.). Plant Syst Evol 300(8):1779–1791
Gomez KA, Gomez AA (1984) Statistical procedures for agricultural research. John Wiley & Sons Inc., Singapore
Greay SJ, Hammer KA (2015) Recent developments in the bioactivity of mono- and diterpenes: anticancer and antimicrobial activity. Phytochem Rev 14:1–6
Hammam KA, Shoala T (2020) Influence of spraying nano-curcumin and nano-rosemarinic acid on growth, fresh herb yield, chemicals composition and postharvest criteria of french basil (Ocimum basilicum l. Var. Grand vert) plants. J Agri Rural Res 5(1):1–22
Hossain MA, Kabir MJ, Salehuddin SM, Mizanur Rahman SM, Das AK, Singha SK, Alam MK, Rahman A (2010) Antibacterial properties of essential oils and methanol extracts of sweet basil Ocimum basilicum occurring in Bangladesh. Pharm Biol 48(5):504–551
Hussain AI, Anwar F, Hussain Sherazi ST, Przybylski R (2008) Chemical composition, antioxidant and antimicrobial activities of basil (Ocimum basilicum) essential oils depends on seasonal variations. Food Chem 108(3):986–995
Jadczak D, Błaszczuk A, Rekowska E (2006) Effect of covering on the content of macroelements in yield of Basil (Ocimum basilicum L.) cultivated for bunch harvest. J Elem 11(2):135–141
Jardin DUP (2015) Plant biostimulants: definition, concept, main categories and regulation. Sci Hortic 196:3–14
Jindo K, Canellas LP, Albacete A, Figueiredo dos Santos L, Rocha RLF, Baia DC, Canellas NOA, Goron TL, Olivares FL (2020) Review on interaction between humic substances and plant hormones for phosphorous acquisition. Agronomy 10(640). https://doi.org/10.3390/agronomy10050640
Kelkar AJ, Herr DJC, Ryan JG (2014) Nanoscience and nanoengineering: advances and applications. Boca Raton: CRC Press, Taylor and Francis Group 331 978-1-4822-3119-9
Khazaie HR, Eyshi Rezaie E, Bannayan M (2011) Application times and concentration of humic acid impact on aboveground biomass and oil production of hyssop (Hyssopus officinalis). J Med Plant Res 5(20):5148–5154
Khokon MAR, Okuma E, Hossain MA, Munemasa S, Uraji M, Nakamura Y et al (2011) Involvement of extracellular oxidative burst in salicylic acid-induced stomatal closure in Arabidopsis. Plant Cell Environ 34:434–443
Klessig DF, Malamy J (1994) The salicylic acid signal in plants. Plant Mol Biol 26(5):1439–1458
Kovácik J, Grúz J, Backor M, Strnad M, Repcák M (2009) Salicylic acid-induced changes to growth and phenolic metabolism in Matricaria chamomilla plants. Plant Cell Rep 28:135–143
Kunert KJ, Baaziz M, Cullis CA (2003) Techniques for determination of true-to-type date palm (Phoenix dactylifera L.) plants: a literature review. Emirates J Agric Sci 15:1–16
Li T, Hu Y, Du X, Tang H, Shen C, Wu J (2014) Salicylic acid alleviates the adverse effects of salt stress in Torreya grandis cv. merrillii seedlings by activating photosynthesis and enhancing antioxidant systems. PLoS One 9:e109492
Linder RC (1944) Rapid analytical methods for some of the more common inorganic constituents of plant tissues. Plant Physiol 19(1):76–79
Lopes-Lutz D, Alviano DS, Alviano CS, Kolodziejczyk PP (2008) Screening of chemical composition, antimicrobial and antioxidant activities of artemisia essential oils. Phytochemistry 69(8):1732–1738
Mirzajani Z, Hadavi E, Kashi A (2015) Changes in the essential oil content and selected traits of sweet basil (Ocimum basilicum L.) as induced by foliar sprays of citric acid and salicylic acid. Ind Crop Prod 76:269–274
Mohamed HI, AG AE-H (2014) Molecular and biochemical markers of some Vicia faba L. genotype in response to storage insect pests infestation. J Plant Interact 9(1):618–626
Mohamed HI, Abdel-Hamid AME (2013) Molecular and biochemical studies for heat tolerance on four cotton genotypes (Gossypium hirsutum L.). Romanian Biotechnol Letters 18(6):7223–7231
Mohamed HI, Elsherbiny EA, Abdelhamid MT (2016) Physiological and biochemical responses of Vicia faba plants to foliar application with zinc and iron. Gesunde Pflanzen 68:201–212
Mohamed HI, Akladious SA, Ashry NA (2018) Evaluation of six soybean genotypes using retroelements and some physiological parameters under water stress. Gesunde Pflanzen 70:205–215
Mohamed HI, Akladious SA, El-Beltagi HS (2018a) Mitigation the harmful effect of salt stress on physiological, biochemical and anatomical traits by foliar spray with trehalose on wheat cultivars. Fresenius Environ Bull 27(10):7054–7065
Mohamed HI, El-Beltagi HS, Aly AA, Latif HH (2018b) The role of systemic and non systemic fungicides on the physiological and biochemical parameters in Gossypium hirsutum plant, implications for defense responses. Fresenius Environ Bull 27(12):8585–8593
Moustafa-Farag M, Mohamed HI, Mahmoud A, Elkelish A, Misra AN, Guy KM, Kamran M, Ai S, Zhang M (2020) Salicylic acid stimulates antioxidant defense and osmolyte metabolism to alleviate oxidative stress in watermelons under excess boron. Plants 9(724). https://doi.org/10.3390/plants9060724
Naderi MR, Danesh-Shahraki A (2013) Nanofertilizers and their roles in sustainable agriculture. Int J Agric Crop Sci 5(19):2229–2232
Naeem M, Basit A, Ahmad I, Mohamed HI, Wasila H (2020) Effect of salicylic acid and salinity stress on the performance of tomato plants. Gesunde Pflanzen 72:393–402
Noreen S, Ashraf M (2010) Modulation of salt (NaCl) induced effects on oil composition and fatty acid profile of sunflower (Helianthus annuus L.) by exogenous application of salicylic acid. J Sci Food Agric 90(15):2608–2616
Noroozisharaf A, Kaviani M (2018) Effect of soil application of humic acid on nutrients uptake, essential oil and chemical compositions of garden thyme (Thymus vulgaris L.) under greenhouse conditions. Physiol Mol Biol Plants 24(3):423–431
Pandey AK, Singh P, Tripathi NN (2014) Chemistry and bioactivities of essential oils of some Ocimum species: an overview. Asian Pac J Trop Biomed 4:682–694
Paradikovi N, Vinkovi´ CT, Vinkovi´ CI, Zuntar I, Boji´ CCM, Sari´ CCM (2011) Effect of natural biostimulants on yield and nutritional quality: an example of sweet yellow pepper (Capsicum annuum L.) plants. J Sci Food Agric 91:2146–2152
Porra RJ, Thompson WA, Kriedemann PE (1989) Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: verification of the concentration of chlorophyll standards by atomic absorption spectroscopy
Purushothaman B, Prasanna SR, Suganthi P, Ranganathan B, Gimbun J, Shanmugam KA (2018) Comprehensive review on Ocimum basilicum. J Nat Remedies 18:71–85
Raskin I (1992) Role of salicylic acid in plants. Annu Rev Plant Biol 43(1):439–463
Ražná K, Ablakulova N, Žiarovská J, Kyseľ M, Cagáň Ľ, Khojiboboevich KK, Bakievich GM (2018) The effect of seed-priming by cobalt-diglycyrrhizinate on wheat (Triticum aestivum L.) genome. J Agri. Res 3(7):000184
Rodrigues-Brandão I, Kleinowski AM, Einhardt AM, Lima MC, DO Amarante L, Peters JA, EJB B (2014) Salicylic acid on antioxidant activity and betacyan in production from leaves of Alternanthera tenella. Ciência Rural Santa Maria 44(10):1893–1898
Rohlf FJ (2008) NTSYS. Pc., Version 2.20U. Exeter Software, Setauket, New York
Roosta HR, Hosseinkhani M, Vakili SMA (2015) Effects of foliar application of nano-fertile fertilizer containing humic acid on growth, yield and nutrient concentration of mint (Mentha sativa) in aquaponic system. J Sci Technol Greenhouse Culture 6(24):1–10
Schiavon M, Pizzeghello D, Muscolo A, Vaccaro S, Francioso O, Nardi S (2010) High molecular size humic substances enhance phenylpropanoid metabolism in maize (Zea mays L.). J Chem Ecol 36:662–669
Shareef HJ, Jasim AM, Abbas MF (2017) Molecular analysis of anti-salinity compounds on date palm offshoots (Phoenix dactylifera L.) cultivars using RAPD techniques. J Environ Sci Comp Sci Eng Technol 6(1):061–071
Shoala T (2018) Positive impacts of nanoparticles in plant resistance against different stimuli 267–279. In: “Nanobio-technology applications in plant protection” (K.A. Abd- -Elsalam, R. Prasad, eds). 1st ed. Springer International Publishing AG, part of Springer Nature
Siddiqui MH, Al-Whaibi MH, Mohammad F (2015) Nanotechnology and plant sciences. Springer International Publishing 303 https://doi.org/10.1007/978-3-319-14502-0
Singh B, Usha K (2003) Salicylic acid induced physiological and biochemical changes in wheat seedlings under water stress. Plant Growth Regul 39:137–141
Smith-Palmer A, Stewart J, Fyfe L (1998) Antimicrobial properties of plant essential oils and essences against five important food-borne pathogens. Lett Appl Microbiol 26:118–122
Sofi A, Ebrahimi M, Shirmohammadi E (2018) Effect of humic acid on germination, growth, and photosynthetic pigments of Medicago sativa L. under salt stress. Ecopersia 6:21–30
Sofy AR, Dawoud RA, Sofy MR, Mohamed HI, Hmed AA, El-Dougdoug NK (2020a) Improving regulation of enzymatic and non-enzymatic antioxidants and stress-related gene stimulation in cucumber mosaic cucumovirus-infected cucumber plants treated with glycine betaine, chitosan and combination. Molecules 25(10):2341. https://doi.org/10.3390/molecules25102341
Sofy MR, Seleiman MF, Alhammad BA, Alharbi BM, Mohamed HI (2020b) Minimizing adverse effects of pb on maize plants by combined treatment with jasmonic, salicylic acids and proline. Agronomy 10:699. https://doi.org/10.3390/agronomy10050699
Sofy AR, Hmed AA, Alnaggar A, Dawoud RA, Elshaarawy R, Sofy MR (2020c) Mitigating effects of bean yellow mosaic virus infection in faba bean using new carboxymethyl chitosan-titania nanobiocomposites. Int J Biol Macromol 163:1261–1275
Souri Z, Karimi N, Sarmadi M, Rostami E (2017) Salicylic acid nanoparticles (SA NPs) improve growth and phytoremediation efficiency of Isatis cappadocica Desv., under As stress. IET Nanobiotechnology:1–10
Swamy MK, Akhtar MS, Sinniah UR (2016) Antimicrobial properties of plant essential oils against human pathogens and their mode of action: an updated review. Evid Based Complement Alternat Med 21:3012462. https://doi.org/10.1155/2016/3012462
Talaat IM, Khattab HI, Ahmed AM (2014) Changes in growth, hormones levels and essential oil content of Ammi visnaga L. plants treated with some bioregulators. Saudi J Biol Sci 21:355–365
Thanaa SM, Nabila EK, AbouRayya MS, Eisa RA (2016) Response of nonpareil seedlings almond to foliar application of licorice root extract and bread yeast suspend under south Sinai conditions. J Innovations Pharma Biol Sci 3:123–132
Trindade H, Costa MM, Sofia BLA, Pedro LG, Figueiredo AC, Barroso JG (2008) Genetic diversity and chemical polymorphism of Thymus caespititius from Pico, Sao Jorge and Terceira islands (Azore). Biochem Syst Ecol 36:790–797
Van-Schouwenburg JCH, Walinga I (1978) Methods of analysis of plant material. Agric Univ, Wagejnen, Netherlands
Vieira RF, Simon JE (2001) Chemical characterization of basil (Ocimum spp.) founding the markets and used in traditional medicine in Brazil. Econ Bot 54(2):207–216
Vieira RF, Goldsbrough P, Simon JE (2003) Genetic diversity of basil (Ocimum spp.) based on RAPD markers. J Am Soc Hortic Sci 128:94–99
Viljoen A, Van Vuuren S, Ernst E, Klepser M, Demirci B, Baser H, Van Wyk BE (2003) Osmitopsis asteriscoides (Asteraceae)-the antimicrobial activity and essential oil composition of a Cape-Dutch remedy. J Ethnopharmacol 88(2-3):137–143
Wang L, Yang R, Yuan B, Liu Y, Liu C (2015) The antiviral and antimicrobial activities of licorice, a widely-used Chinese herb. ActaPharmac Sin B 5(4):310–315
Warsi A, Sholichah R (2017) Phytochemical screening and antioxidant activity of ethanolic extract and ethyl acetate fraction from basil leaf (Ocimum basilicum L.) by DPPH radical scavenging method. IOP Conf Ser Mater Sci Eng 259:012008
Williams JGK, Kubelik AR, Livak KJ, Rafalski JA, Tingey SV (1990) DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res 18:6531–6535
Yong JP (2012) Advances in studies on the synthesis of glycyrrhizic acid, glycyrrhetinic acid derivatives and their biological activities. Lishizhen Medicine and Materia Medica Res 23(6):1174–1182
Yusuf M, Hayat S, Alyemeni MN, Fariduddin Q, Ahmad A (2013) Salicylic acid: physiological roles in plants. In: Hayat S, Ahmad A, Alyemeni M (eds) Salicylic acid. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-6428-6_2
Acknowledgements
We thank Professor Mahasen Sedky, Department of Medicinal and Aromatic Plants Research, as that the seeds have been obtained from her.
Availability of data and materials
Not applicable.
Author information
Authors and Affiliations
Contributions
A A: conceptualization, methodology, data curation, writing — original draft of physiological data, visualization, and investigation; M G: conceptualization, methodology, data curation, and writing — original draft of molecular data; T S and HI M: validation, resources, and writing — review and editing.
Corresponding author
Ethics declarations
Ethics approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Conflict of interest
The authors declare no competing interests.
Additional information
Responsible Editor: Philippe Garrigues
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Amer, A., Ghoneim, M., Shoala, T. et al. Comparative studies of eco-friendly compounds like humic acid, salicylic, and glycyrrhizic acids and their nanocomposites on French basil (Ocimum basilicum L. cv. Grand verde). Environ Sci Pollut Res 28, 47196–47212 (2021). https://doi.org/10.1007/s11356-021-14022-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-021-14022-1