
Abstract
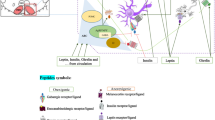
Several mechanisms both inside and outside the central nervous system (CNS) collaborate to control feeding. The hypothalamus received food-related messages from different regions of the brain, and exports appropriate output through specific hypothalamic nuclei. One of the most important of these hypothalamic nuclei is the lateral hypothalamus area (LHA). In this review, multiple valid papers from electronic sources (including Web of Science, Scopus, PubMed, SID, Google Scholar, and ISI databases) were used; which in them the role of LHA in the central regulation of feeding investigated. The hypothalamus is responsible for the central control of food intake in the CNS. The arcuate nucleus, paraventricular nucleus, and LHA are specific regions in the hypothalamus that regulate food intake. The LHA is considered to be the center of hunger or feeding among these nuclei. With the interaction of orexigenic and anorexigenic neurons, as well as various neurotransmitters in several neuronal pathways, this nucleus produces increased food intake. The LHA eventually improved food intake via extensive brain connections. The LHA via various neurons and synaptic connections with other special hypothalamic nuclei involved in the control of nutritional behavior (graphical abstract) stimulated food intake.
Graphical Abstract

Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Concerning the importance of nutrition as an essential ingredient of our lifestyle, we are going to acquire more knowledge about the information as a balance of energy that is was transmitted to the brain by several types of receptors such as smell, taste, vision, and gastrointestinal hormones (Zendehdel and Hassanpour 2014a, b; Zendehdel and Hassanpour 2014a, b; Hajinezhad et al. 2018). These data are under surveillance of a special part of the brain namely the forebrain and midbrain. In addition, it is transmitted by the function of neurotransmitters to the hypothalamus with inhibition and exhibition characters (Zendehdel et al. 2013a, b; Zendehdel et al. 2015; Marino et al. 2020).
Our behavior is the outcome of diverse circuit control of the brain alongside monitoring by considering full coordination at the level of the hypothalamus between foods related input messages and appropriate responses (Zendehdel et al. 2017; Qualls-Creekmore and Münzberg 2018).
By the last research in the field of neuroscience which had been implemented, among new technologies, investigator’s scientists have been proved to observe the neural intrinsic function of brain cells in tandem with control animal behavior (Zendehdel et al. 2020; Rahmani et al. 2021). This progression accommodates an atmosphere to innovative an array of massive populations of neurons and multiple neural circuits in the brain associated with nutritional behaviours (Zendehdel et al. 2019). This, in turn, scientists have found numerous regions deal with nutritional behavior. Many types of researches are still being done to identify different regions of the brain center controlling feeding (Clarke et al. 2018; Rossi and Stuber 2018). A large number of studies on special hypothalamus nuclei by the power of electrical stimulation consequently reveal that the lateral hypothalamus area (LHA) is the master responsible for the hunger center in the hypothalamus (Qualls-Creekmore and Münzberg 2018).
The LHA has several functions in the brain. Central control of food and water intake, blood pressure, heart rate, sleep, peripheral nervous system (PNS), and reward system are the most important functions of this nucleus (Farrokhi et al. 2020). One of the most important functions of the LHA is control of feeding. Multiple neuronal populations in this nucleus, along with orexigenic and anorexigenic neurotransmitters in LHA via multiple neural circuits send the message of increasing food intake to other upstream regions (Stuber and Wise 2016).
The LHA plays an important role in controlling food intake, and increasing feeding through interactions between multiple neurons and neurotransmitters. Review articles that have previously examined the role of LHA in appetite have merely described a specific neurotransmitter, or its neurotransmitter functions have been studied by various mechanisms. The current review article comprehensively surveyed the role of the most important neurotransmitters involved in the central regulation of food intake through the LHA, their receptors, and their associated second-order neurons. As well this review article studied the main role of LHA in the central regulation of food intake.
Study Design
In this study, the role of LHA in the central regulation of food intake was investigated. Multiple valid papers from electronic sources were used. Valuable articles indexed in the Web of Science, Scopus, PubMed, SID, Google Scholar, and ISI databases by using of Keywords: Central regulation of food intake, Hypothalamus, Lateral hypothalamus area, and Brain neurotransmitters.
The Role of LHA in Feeding
According to the central mechanism of feeding regulation, the interaction among of arcuate nucleus (ARC), paraventricular (PVN), and LHA is the most important central controller of feeding behaviour (Sahu 2003). The ARC is responsible for maintaining the metabolic status of the body. This nuclear function was performed by two neuronal populations including agouti-related protein (AgRP), and pro-opiomelanocortin (POMC) (Hamidi and Yusefvand 2017). When the body’s metabolic status changes, the starvation or hormonal changes, these neuronal populations are activated in the ARC, and by their functions, they move the body towards energy homeostasis (Yousefvand et al. 2020; Yousefvand and Hamidi 2021). These two neural populations by body’s hormonal status are changed. Finally, they send their outputs to the LHA, PVN, and bed nucleus of the stria terminalis (BNTS). As it’s obvious, the LHA is the center of hunger (feeding). As soon as the body’s energy status changes (due to starvation, or hormonal changes), the AGRP neuronal group activates, and inhibits the POMC neuronal group through the gamma-aminobutyric acid (GABA) branch. This, in turn, sends their excitatory output to multiple neuronal groups in the LHA (Chen et al. 2015, 2016). As well, the LHA achieves input from another region, such as the PVN. This nucleus also receives hormonal changes in the body directly. LHA is considered as the nucleus that integrates signals which are coming from the hypothalamus and other regions. Then it sends the message of increased feeding by changes in the activity of its neuronal pathways (Stuber and Wise 2016). LHA affects the central control of food intake through the function of various orexigenic, and anorexic neurotransmitters (Table 1), their receptors (Table 2), and second-order neurons. Orexigenic neurotransmitters include ghrelin (Toshinai et al. 2003), galanin (Schick et al. 1993), orexin (Zhu et al. 2002), and neuropeptide Y (NPY) (Gumbs et al. 2020a, b). Anorexigenic neurotransmitters include α-melanocortin stimulating hormone (α-MSH) (McMinn et al. 2000), insulin (Yousefvand et al. 2020), leptin (Sahu 2003), dopamine (Vucetic and Reyes 2010), and neurotensin (Schroeder and Leinninger 2018).
Glucose
Glucose is the primary source of energy for neurons. Brain neurons receive 10% of blood glucose. Blood glucose levels dropped during fasting, and less glucose reached the brain’s neurons. Glucose-sensing neurons detect glucose neurons in the brain. The LHA and VMH were the first areas where glucose-sensing neurons were discovered. The glucostatic theory states that the management of food intake in a starving state is primary (Routh 2010). Glucose-sensing neurons are divided into two types: glucose-inhibition (GI) neurons, and glucose-excited (GE) neurons. These two neurons are found in two different ARC populations. GE neuron located on the POMC/cocaine-amphetamine regulated transcript (CART), and GI neuron located on the NPY/AgRP neuron. Because glucose is a large polar molecule, it is not able to cross the blood–brain barrier (BBB) and must be transported by a carrier. Glucose transporters from the BBB are glucose transporters (GLUTs), sodium–glucose co-transporters (SGLTs), and sugars will eventually be exported transporters (Takeda et al. 2021). Because of GLT2, and SLC5A2 express in the brain, and glucose sensor in the brain. GLT2, and SLC5A2 inhibitors induced food intake in rats, they are expected to play a key role in central control of food intake, and via c-fos transcription factor upregulated in the LHA, and increased food intake (Takeda et al. 2021). When glucose blood drops, GI neurons on the NPY/AgRP neuron activate, and stimulate this neuron’s population, after that inhibited POMC. Activating NPY/AgRP neuron with GI neuron resulted in NPY releasing into the synaptic cleft. Then NPY in the synaptic cleft activated neuropeptide Y receptor (NPYR) on the orexin neuron of LHA and increases food intake. At the time of high energy levels (increasing leptin, and insulin), GE neurons located on the POMC are stimulated via different neurotransmitters. So GE neurons stimulated POMC, and inhibited NPY/AgRP neuron, then secreted α-MSH. α-MSH is located on the melanocortin stimulating 4 receptors (MC4R) on the MC4R neuron located on the LHA. Therefore decreased food intake (Song et al. 2001). So, LHA in hunger state via NPY synthesis, and in satiety state via α-MSH synthesis, increased and decreased food intake respectively.
Melanocortin Stimulating Hormone
MSH is a hypothalamic neurohormone that plays an important role in controlling food intake (Zendehdel et al. 2012). Intracerebroventricular injection (ICV) injection of this neurohormone decreased food intake in rats, and mice (Kim et al. 2000). This neurohormone synthesis in the POMC neuron and its receptors (MC4R) is in the ARC, PVN, and LHA. The target of the second-order neuron for the MSH is the corticotropin-releasing hormone (CRH) neuron in the LHA (McMinn et al. 2000; Michael et al. 2020). The MC4R is located on the MC4R and GE neurons on the LHA. When the body’s energy level rises (after feeding), leptin was released from adipose tissue. Leptin by leptin receptor (Lp-R) binding to the POMC neuron located on the ARC, and induced α-MSH synthesis and released. So, α-MSH binding to the MC4R on the MC4R, and GE neurons on the LHA. After that, activation of these hypophagic neurons decreased feeding (Kim et al. 2000; Guan et al. 2017; Takeda et al. 2021). Therefore, α-MSH via binding to the MCR4 on the MC4R neuron located on the LHA decreased food intake.
Neuropeptide Y
NPY is a peptide with 36 amino acids. NPY is the most abundant neuropeptide in the central nervous system (CNS). This neuropeptide is stimulatory and plays an important role in the central control of food intake. NPY exerts its effect through NPY receptors such as NPYR1, NPYR2, NPYR4, NPYR5, NPYR6, and NPYR7. These receptors belong to the G protein-coupled receptor (GPCR) family. NPYR1, NPYR2, and NPYR5 mediated the hyperphagic effect of NPY (Yousefvand et al. 2018a, b; Yousefvand et al. 2019). NPY in the CNS synthesis by NPY/AgRP neurons in the ARC. Anorexigenic factors such as insulin, and leptin reduced NPY/AgRP neuron function; but orexigenic factors such as ghrelin, orexin, and galanin increased NPY/AgRP neuron function (Zhang et al. 2019). NPYR1, and NPYR5 are orexigenic receptors, but NPYR2 is anorexigenic receptor. NPYR2 is an auto-receptor (Hamidi and Yusefvand 2017). The most important mediator of the orexigenic action of NPY is NPYR1 (Zhang et al. 2019).NPYR1 is located on the LHA. When orexigenic hormones, such as ghrelin, are released into the bloodstream, this hormone crosses the BBB, and lodges at its receptors in the ARC. It then stimulates NPY/AgRP neurons in the ARC and releases NPY. As well GABA in the NPY/AgRP neuron inhibited the POMC neuron. After that NPY is located on NPYR1 in the orexin neurons of the LHA stimulates this neuron to increase food intake (Mercer et al. 2011; Gumbs et al. 2020a, b). NPY via binding to the NPYR1 in the orexin neuron located on the LHA increased food intake.
Orexin
Orexin is a hypothalamic neuropeptide that performs many important functions in the CNS. Orexin contains Orexin-A and B (Furuse et al. 1999). These neuropeptides are orexigenic and exert their function through orexin receptors (OXR) including OX1R and OX2R. These are receptors that belong to the GPCR family. OX2R has a widespread distribution in the LHA, ARC, PVN, and VMH (Yousefvand and Hamidi 2021). As well, orexin neurons are located on the LHA. In the hunger state or hypoglycemia, NPY synthesis in the NPY/AgRP neuron located on the ARC. So, NPY released to the synaptic cleft, and binding to the NPYR1 on the LHA orexin neuron, and via activated orexin neuron in this nucleus increased food intake (Hurley and Johnson 2014; Gumbs et al. 2020a, b). Indicated that, ICV injection of orexin-A, induced food intake in satiated rats (Sutcliffe and de Lecea 2002). During hypoglycemia, in the LHA glucose-sensing neurons, orexin neurons are activated and increased food intake (Liu et al. 2001). In the hunger status, orexin neuron stimulated and secreted orexin neuropeptide. So this neuropeptide binds to the OX1R in the NPY/AgRP located on the ARC. Orexin activated this neuron and released NPY to the synaptic cleft. After that, NPY binding to the NPYR1 on the orexigenic neurons located on the LHA, and with stimulation of these neurons increased food intake (Sahu 2003; Luan et al. 2017).
Insulin
Insulin is a special neurohormone in the CNS; which regulated food intake. The central and peripheral effects of insulin have differed. Central insulin reduces food intake, but peripheral insulin increases food intake. Insulin function in the CNS is mediated by insulin receptors (IR). IRs are members of the tyrosine kinase receptor family (Yousefvand et al. 2018a, b; Yousefvand and Hamidi 2020; Yousefvand et al. 2020). These receptors are located on the glucose-sensing neuron on the LHA. In the LHA, there is a population of MC4R neurons; which express MC4R, and Lp-R (Cui et al. 2012). MCRs are GPCR, and their actions are mediated by the activity of adenylyl cyclases and cyclic adenosine monophosphate (CAMP) (Yousefvand and Hamidi 2021). When central insulin or insulin passed through the BBB, is attached to IR located on the POMC in the ARC, it causes the synthesis of α-MSH in this neuron (Yousefvand et al. 2020). After that α-MSH is released to the synaptic cleft, and attached to the MC4R on the MC4R neuron (Cui et al. 2012), and thereby reducing food intake through the LHA. Also, when blood glucose and insulin are high, insulin is deposited on the IR receptor located on the glucose-sensing neurons in the LHA (Ono 2019). So insulin by inhibiting these neurons reduces food intake through the LHA. As well, the ICV injection of insulin reduced galanin expression (Chaillou and Tillet 2005). So, insulin via synthesis of α-MSH, and decreased function of glucose-sensing neurons, reduced food intake.
Leptin
Leptin informs the body’s energy status to the nutrition control centers in the hypothalamus and is an important signal of energy regulation in the body. Indicated that, central and peripheral leptin administration reduces food intake, and body weight in rats, monkeys, and mice (Sahu 2003). Leptin exerts its function by Lp-R. Lp-R is a member of the class 1 cytokine receptor family. There are two isoforms of LpR: short and long. Long isoforms are found in hypothalamic nuclei related to the central control of food intake, such as the ARC, VMH, PVN, DMH, and LHA (Yousefvand and Hamidi 2021). Lp-Rs are located on the galanin, and orexin neurons in the LHA (Berthoud and Münzberg 2011). These neurons are orexigenic. When the body’s energy levels are high, leptin binding to its receptors on these neurons, and inhibits increased food intake (Sahu 2003). As well, indicated that the ICV injection of leptin reduced galanin expression (Chaillou and Tillet 2005). Also, leptin from circulating via binding to its receptors on POMC neuron activates GE neurons located on the POMC. Activated GE neurons stimulated POMC, and this neuron released αMSH into the synaptic cleft. After that, α-MSH binding to the MC4R receptor located on the MC4R neuron on the LHA. So, α-MSH via activation of MC4R neuron inhibited feeding. In addition, MC4R neurons contain Lp-R. Therefore leptin via Lp-R is located directly on MC4R neurons reducing food intake (Cui et al. 2012; Ono 2019; Takeda et al. 2021). Neurotensin neurons are the gabaergic neurons and are presented in the LHA. The activity of this neuron is hypophagic. Lp-R is located on these neurons and stimulated neurotensin neuron, so reduces food intake (Brown et al. 2019). Leptin via increased synthesis of α-MSH decreased the function of orexin, and galanin neurons, but increased the function of neurotensin neurons, so decreased food intake.
Dopamine
Dopamine is an organic compound in the family of catecholamine and phenethylamines; which plays a vital role in the body and brain (Meguid et al. 2000; Yousefvand and Hamidi 2021). Among brain monoamines, dopamine is involved in controlling food intake. Dopamine is involved in regulating the pattern of nutrition and is a factor in initiating nutrition. At the beginning of feeding, the secretion of dopamine into the LHA is at the highest (Szczypka et al. 2000). Dopamine receptors in the LHA are predominantly dopamine receptor (DR)2 and to a lower level DR1 (Vucetic and Reyes 2010). DR1 and DR2 receptors belong to two different families of dopamine receptors and therefore have different functions. DR1 family receptors (DR1, DR5) are connected to Gαs, which raises the intracellular concentration of cAMP, whereas DR2 family receptors (DR2, DR3, DR4) are connected to Gαi, which directly decreases cAMP production by inhibiting adenylate cyclase (Björklund and Dunnett 2007). The impact of dopamine on food in the hypothalamus depends on the nucleus being examined, the type of receptor involved, and the nutritional status of the body. In the LHA, the DR2 is the dominant receptor (Vucetic and Reyes 2010). In the VMH, the DR1 is the dominant receptor (Yousefvand and Hamidi 2021). Therefore, the function of dopamine in this nucleus is different. It has been shown that dopamine levels in the LHA increase during food intake decrease after food intake. Therefore, dopamine in the LHA is involved in reducing food intake. Increasing the secretion of dopamine in the LHA inhibits food intake (Nieh et al. 2016; Stuber and Wise 2016). Maybe dopamine increased in the LHA, inhibited the orexigenic neurons in this nucleus, and decreased food intake. Also, this increase in dopamine secretion during eating in the LHA may be a signal to control the size of the meal (Van Gestel et al. 2014).
Ghrelin
Ghrelin is a peptide hormone secreted from the stomach with various paracrine and endocrine effects. Immuno-radioactive studies on ghrelin have shown that, high levels of this neurohormone in the hypothalamus, and some parts of the brain. The effect of ghrelin on nutritional behavior is dual (Zendehdel et al. 2013a, b). Ghrelin increased food intake in rats and mice, (Toshinai et al. 2003; Hsu et al. 2015; López-Ferreras et al. 2017) but decreases in broilers (Zendehdel et al. 2013a, b; Farrokhi et al. 2020). Ghrelin reduces food intake in birds through DR1 and CB1 receptors (Zendehdel et al. 2013a, b; Farrokhi et al. 2020). In rats and mice, ghrelin exerts its effect on food intake through growth hormone secretagogue receptor 1a (GHSR1a). This receptor has highly expressed in LHA, and ARC, and exerts its effect on food intake through direct and indirect effects (Prinz and Stengel 2017). Ghrelin exerts its effect on food intake due to the presence of GHSR1a on the LHA and activated the orexin neuron in the LHA, and finally increased food intake. As well, GHSR1 is present on NPY/AgRP neurons in the ARC. By stimulating these receptors by ghrelin, the NPY/AgRP neuron is motivated and release NPY into the synaptic space. Then NPY stimulated NPYR1 on the LHA, after that NPY fiber stimulates orexin neurons, and activation of this neuron increased food intake (Toshinai et al. 2003; Chen et al. 2004; Hsu et al. 2015). Orexin neurons are the target of ghrelin to exert its hyperphagic effect in the LHA (López-Ferreras et al. 2017). Ghrelin via NPY synthesis increased, and activation of orexin neurons increased food intake in rats and mice. In broiler chicken ghrelin via DR1 and CB1 receptors reduced food intake.
Neurotensin
Neurotensin is a neuropeptide that is released from CNS and digestive tract. This neurotransmitter via central effect induced hypophagia, and weight loss. The central neurotensin synthesis in the brain, and the peripheral neurotensin synthesis in the adrenal gland. Neurotensin expresses its effect on food intake through neurotensin receptor (NTs-R) 1, and 2. These receptors belong to the GPCR family. NTs-R1 is widely distributed in the CNS, and maybe mediated the hypophagic effect of neurotensin. The Neurotensin neurons express neurotensin and GABA. So this neuron is gabaergic neurons. Neurotensin neurons are distributed in the brain especially the hypothalamus. In the hypothalamus, this neuron is located on the LHA. This neuron is a hypophagic neuron, and induced reduction in food intake, and body weight (Opland et al. 2013; Brown et al. 2019). Physiologically, these neurons are stimulated by food-inhibiting hormones, including leptin (Patterson et al. 2015; Woodworth et al. 2017). Increased feeding due to NTsR1 blockage in the LHA. ICV injection of neurotensin lowered food intake in mice via an action on the LHA (Myers et al. 2016; Schroeder and Leinninger 2018). Lp-R is also found on the neurotensin neuron, and so leptin induces hypophagia via this neuron. (Leinninger et al. 2011; Brown et al. 2018). Food intake was reduced by neurotensin binding to the NTs-R1 receptor and neurotensin neuron stimulation.
Galanin
Galanin is a peptide with 29 amino acids (Schick et al. 1993). Galanin has a stimulating effect on food intake in rodents (Chaillou and Tillet 2005). Galanin’s influence on food intake is mediated by galanin-receptor-1 (GalR1). GalR1 is a member of the GPCR family (Laque et al. 2015). On the LHA, there is a Galanin orexigenic neuron. This neuron is a gabaergic neuron. (Berthoud and Münzberg 2011). When galanin-expressing neurons are stimulated, these neurons secrete the neurohormone galanin. Therefore galanin is located on the ARC via GalR1 and stimulates NPY/AgRP neurons. After that, NPY is released to the synaptic cleft. Then NPY is located on NPYR1 in the orexin neuron in the LHA. Therefore, stimulate this gabaergic neuron to be increasing food intake. The galanin receptor is also located on the neurons of orexin, and by directly stimulating these neurons, it increases food intake (Sahu 2003; Qualls-Creekmore et al. 2017). Galanin via increased NPY synthesis, and stimulation of orexin neuron increased food intake.
Conclusion
Feeding central regulation is a complicated process that requires the cooperation of the CNS, PNS, and body. Various hormones reflect the hormonal changes that occur in the body during hunger and satiety to the CNS. The hypothalamus receives these many messages from the CNS. The hypothalamus is the brain’s fundamental regulating center for nutrition. The hypothalamus regulates food intake by interactions between the ARC and, in particular, the LHA. Hypoglycemia, which is perceived by glucose-sensing neurons and influences the function of other neurons in this nucleus, is the most important stimuli in the LHA in controlling food intake. NPY and -MSH are the most critical brain neurotransmitters in the LHA’s regulation of food intake. To control food intake, these two neurotransmitters affect separate neuron papulations in the LHA. An increase in food intake is the outcome of the interaction of several neuronal circuits in the LHA.
Abbreviations
- LHA:
-
Lateral hypothalamus area
- PNS:
-
Peripheral nervous system
- ARC:
-
Arcuate nucleus
- PVN:
-
Paraventricular nucleus
- AgRP:
-
Agouti-related protein
- POMC:
-
Pro-opiomelanocortin
- BNTS:
-
Bed nucleus of the stria terminalis
- GABA:
-
Gamma-aminobutyric acid
- GI:
-
Glucose-inhibition
- GE:
-
Glucose-excited
- CART:
-
Cocaine-amphetamine regulated transcript
- BBB:
-
Blood–brain barrier
- GLUTs:
-
Glucose transporters
- SGLTs:
-
Sodium–glucose co-transporters
- NPYR:
-
Neuropeptide Y receptor
- α-MSH:
-
α-Melanocortin stimulating hormone
- MC4R:
-
Melanocortin stimulating 4 receptor
- ICV:
-
Intracerebroventricular injection
- CRH:
-
Corticotropin-releasing hormone
- Lp-R:
-
Leptin receptor
- CNS:
-
Central nervous system
- GPCR:
-
G protein-coupled receptor
- OXR:
-
Orexin receptors
- IR:
-
Insulin receptors
- CAMP:
-
Cyclic adenosine monophosphate
- GHSR1a:
-
Growth hormone secretagogue receptor 1a
- NTs-R:
-
Neurotensin receptor
- GalR1:
-
Galanin-receptor1
References
Berthoud HR, Münzberg H (2011) The lateral hypothalamus as integrator of metabolic and environmental needs: from electrical self-stimulation to opto-genetics. Physiol Behav 104(1):29–39
Björklund A, Dunnett SB (2007) Dopamine neuron systems in the brain: an update. Trends Neurosci 30(5):194–202
Brown J, Sagante A, Mayer T, Wright A, Bugescu R, Fuller PM, Leinninger G (2018) Lateral hypothalamic area neurotensin neurons are required for control of orexin neurons and energy balance. Endocrinology 159(9):3158–3176
Brown JA, Wright A, Bugescu R, Christensen L, Olson DP, Leinninger GM (2019) Distinct subsets of lateral hypothalamic neurotensin neurons are activated by leptin or dehydration. Sci Rep 9(1):1–16
Chaillou E, Tillet Y (2005) Nutrition and hypothalamic neuropeptides in sheep: histochemical studies. Histol Histopathol
Chen HY, Trumbauer ME, Chen AS, Weingarth DT, Adams JR, Frazier EG, Shen Z, Marsh DJ, Feighner SD, Guan XM, Ye Z (2004) Orexigenic action of peripheral ghrelin is mediated by neuropeptide Y and agouti-related protein. Endocrinology 145(6):2607–2612
Chen Y, Lin YC, Kuo TW, Knight ZA (2015) Sensory detection of food rapidly modulates arcuate feeding circuits. Cell 160(5):829–841
Chen Y, Lin YC, Zimmerman CA, Essner RA, Knight ZA (2016) Hunger neurons drive feeding through a sustained, positive reinforcement signal. Elife 5:e18640
Clarke RE, Verdejo-Garcia A, Andrews ZB (2018) The role of corticostriatal–hypothalamic neural circuits in feeding behaviour: implications for obesity. J Neurochem 147(6):715–729
Cui H, Sohn JW, Gautron L, Funahashi H, Williams KW, Elmquist JK, Lutter M (2012) Neuroanatomy of melanocortin-4 receptor pathway in the lateral hypothalamic area. J Comp Neurol 520(18):4168–4183
Farrokhi R, Babapour V, Zendehdel M, Asghari A, Gilanpour H (2020) The role of dopaminergic and cannabinoidergic receptors on ghrelin-induced hypophagia in neonatal chicken. Arch Razi Inst 76(4):941–954
Furuse M, Ando R, Bungo T, Shimojo M, Masuda Y (1999) Intracerebroventricular injection of orexins does not stimulate food intake in neonatal chicks. Br Poult Sci 40(5):698–700
Guan HZ, Dong J, Jiang ZY, Chen X (2017) α-MSH influences the excitability of feeding-related neurons in the hypothalamus and dorsal vagal complex of rats. BioMed Res Int 2017:1–9
Gumbs MC, Eggels L, Kool T, Unmehopa UA, van den Heuvel JK, Lamuadni K, Mul JD, la Fleur SE (2020a) Neuropeptide Y signaling in the lateral hypothalamus modulates diet component selection and is dysregulated in a model of diet-induced obesity. Neuroscience 447:28–40
Gumbs MC, Eggels L, Vuuregge AH, Unmehopa UA, Mul JD, la Fleur SE (2020) Effects of Neuropeptide Y administration into the lateral hypothalamus on intake of free-choice high-fat high-sucrose diet components of the male Wistar rat. Nutr Neurosci 25(3):621–630
Hajinezhad MR, Hasanein P, Mokhtarpour A (2018) Nociceptin/orphanin FQ (N/OFQ) receptors are involved in adrenaline-induced feeding behavior in broiler cockerels. Int J Pept Res Ther 24(3):403–407
Hamidi F, Yusefvand S (2017) Role of the hypothalamic arcuate nucleus in regulation of food intake (review study). J Neyshabur Univ Med Sci 5(1):52–65
Hsu TM, Hahn JD, Konanur VR, Noble EE, Suarez AN, Thai J, Nakamoto EM, Kanoski SE (2015) Hippocampus ghrelin signaling mediates appetite through lateral hypothalamic orexin pathways. Elife 4:e11190
Hurley SW, Johnson AK (2014) The role of the lateral hypothalamus and orexin in ingestive behavior: a model for the translation of past experience and sensed deficits into motivated behaviors. Front Syst Neurosci 8:216
Kim MS, Rossi M, Abusnana S, Sunter D, Morgan DG, Small CJ, Edwards CM, Heath MM, Stanley SA, Seal LJ, Bhatti JR (2000) Hypothalamic localization of the feeding effect of agouti-related peptide and alpha-melanocyte-stimulating hormone. Diabetes 49(2):177–182
Laque A, Yu S, Qualls-Creekmore E, Gettys S, Schwartzenburg C, Bui K, Rhodes C, Berthoud HR, Morrison CD, Richards BK, Münzberg H (2015) Leptin modulates nutrient reward via inhibitory galanin action on orexin neurons. Mol Metab 4(10):706–717
Leinninger GM, Opland DM, Jo YH, Faouzi M, Christensen L, Cappellucci LA, Rhodes CJ, Gnegy ME, Becker JB, Pothos EN, Seasholtz AF (2011) Leptin action via neurotensin neurons controls orexin, the mesolimbic dopamine system and energy balance. Cell Metab 14(3):313–323
Liu XH, Morris R, Spiller D, White M, Williams G (2001) Orexin a preferentially excites glucose-sensitive neurons in the lateral hypothalamus of the rat in vitro. Diabetes 50(11):2431–2437
López-Ferreras L, Richard JE, Anderberg RH, Nilsson FH, Olandersson K, Kanoski SE, Skibicka KP (2017) Ghrelin’s control of food reward and body weight in the lateral hypothalamic area is sexually dimorphic. Physiol Behav 176:40–49
Luan X, Sun X, Guo F, Zhang D, Wang C, Ma L, Xu L (2017) Lateral hypothalamic Orexin-A-ergic projections to the arcuate nucleus modulate gastric function in vivo. J Neurochem 143(6):697–707
Marino RA, McDevitt RA, Gantz SC, Shen H, Pignatelli M, Xin W, Wise RA, Bonci A (2020) Control of food approach and eating by a GABAergic projection from lateral hypothalamus to dorsal pons. Proc Natl Acad Sci 117(15):8611–8615
McMinn JE, Wilkinson CW, Havel PJ, Woods SC, Schwartz MW (2000) Effect of intracerebroventricular α-MSH on food intake, adiposity, c-Fos induction, and neuropeptide expression. Am J Physiol Regul 279(2):695–703
Meguid MM, Fetissov SO, Varma M, Sato T, Zhang L, Laviano A, Rossi-Fanelli F (2000) Hypothalamic dopamine and serotonin in the regulation of food intake. Nutrition 16(10):843–857
Mercer RE, Chee MJ, Colmers WF (2011) The role of NPY in hypothalamic mediated food intake. Front Neuroendocrinol 32(4):398–415
Michael NJ, Caron A, Lee CE, Castorena CM, Lee S, Zigman JM, Williams KW, Elmquist JK (2020) Melanocortin regulation of histaminergic neurons via perifornical lateral hypothalamic melanocortin 4 receptors. Mol Metab 35:100956
Myers MG, Olson DP, Low MJ, Elias CF, Ahima RS (2016) Brain regulation of feeding and energy homeostasis. Metabolic syndrome. Springer, Cham, pp 347–368
Nieh EH, Vander Weele CM, Matthews GA, Presbrey KN, Wichmann R, Leppla CA, Izadmehr EM, Tye KM (2016) Inhibitory input from the lateral hypothalamus to the ventral tegmental area disinhibits dopamine neurons and promotes behavioral activation. Neuron 90(6):1286–1298
Ono H (2019) Molecular mechanisms of hypothalamic insulin resistance. Int J Mol Sci 20(6):1317
Opland D, Sutton A, Woodworth H, Brown J, Bugescu R, Garcia A, Christensen L, Rhodes C, Myers M Jr., Leinninger G (2013) Loss of neurotensin receptor-1 disrupts the control of the mesolimbic dopamine system by leptin and promotes hedonic feeding and obesity. Mol Metab 2(4):423–434
Patterson CM, Wong JM, Leinninger GM, Allison MB, Mabrouk OS, Kasper CL, Gonzalez IE, Mackenzie A, Jones JC, Kennedy RT, Myers MG Jr (2015) Ventral tegmental area neurotensin signaling links the lateral hypothalamus to locomotor activity and striatal dopamine efflux in male mice. Endocrinology 156(5):1692–1700
Prinz P, Stengel A (2017) Control of food intake by gastrointestinal peptides: mechanisms of action and possible modulation in the treatment of obesity. J Neurogastroenterol Motil 23(2):180
Qualls-Creekmore E, Münzberg H (2018) Modulation of feeding and associated behaviors by lateral hypothalamic circuits. Endocrinology 159(11):3631–3642
Qualls-Creekmore E, Yu S, Francois M, Hoang J, Huesing C, Bruce-Keller A, Burk D, Berthoud HR, Morrison CD, Münzberg H (2017) Galanin-expressing GABA neurons in the lateral hypothalamus modulate food reward and noncompulsive locomotion. J Neurosci 37(25):6053–6065
Rahmani B, Ghashghayi E, Zendehdel M, Khodadadi M, Hamidi B (2021) The crosstalk between brain mediators regulating food intake behavior in birds: a review. Int J Pept Res Ther 27:2349–2370
Rossi MA, Stuber GD (2018) Overlapping brain circuits for homeostatic and hedonic feeding. Cell Metab 27(1):42–56
Routh VH (2010) Glucose sensing neurons in the ventromedial hypothalamus. Sensors 10(10):9002–9025
Sahu A (2003) Leptin signaling in the hypothalamus: emphasis on energy homeostasis and leptin resistance. Front Neuroendocrinol 24(4):225–253
Schick RR, Samsami SO, Zimmermann JP, Eberl TH, Endres CH, Schusdziarra VO, Classen ME (1993) Effect of galanin on food intake in rats: involvement of lateral and ventromedial hypothalamic sites. Am J Physiol Regul Integr Comp Physiol 264(2):355–361
Schroeder LE, Leinninger GM (2018) Role of central neurotensin in regulating feeding: implications for the development and treatment of body weight disorders. Biochim Biophys Acta BBA Mol Basis Dis 1864(3):900–916
Song Z, Levin BE, McArdle JJ, Bakhos N, Routh VH (2001) Convergence of pre-and postsynaptic influences on glucosensing neurons in the ventromedial hypothalamic nucleus. Diabetes 50(12):2673–2681
Stuber GD, Wise RA (2016) Lateral hypothalamic circuits for feeding and reward. Nat Neurosci 19(2):198–205
Sutcliffe JG, de Lecea L (2002) The hypocretins: setting the arousal threshold. Nat Rev Neurosci 3(5):339–348
Szczypka MS, Rainey MA, Palmiter RD (2000) Dopamine is required for hyperphagia in Lep ob/ob mice. Nat Genet 25(1):102–104
Takeda K, Ono H, Ishikawa K, Ohno T, Kumagai J, Ochiai H, Matumoto A, Yokoh H, Maezawa Y, Yokote K (2021) Central administration of sodium–glucose cotransporter-2 inhibitors increases food intake involving adenosine monophosphate-activated protein kinase phosphorylation in the lateral hypothalamus in healthy rats. BMJ Open Diabetes Res Care 9(1):e002104
Toshinai K, Date Y, Murakami N, Shimada M, Mondal MS, Shimbara T, Guan JL, Wang QP, Funahashi H, Sakurai T, Shioda S (2003) Ghrelin-induced food intake is mediated via the orexin pathway. Endocrinology 144(4):1506–1512
Van Gestel MA, Kostrzewa E, Adan RA, Janhunen SK (2014) Pharmacological manipulations in animal models of anorexia and binge eating in relation to humans. Br J Pharmacol 171(20):4767–4784
Vucetic Z, Reyes TM (2010) Central dopaminergic circuitry controlling food intake and reward: implications for the regulation of obesity. Wiley Interdiscip Rev Syst Biol Med 2(5):577–593
Woodworth HL, Beekly BG, Batchelor HM, Bugescu R, Perez-Bonilla P, Schroeder LE, Leinninger GM (2017) Lateral hypothalamic neurotensin neurons orchestrate dual weight loss behaviors via distinct mechanisms. Cell Rep 21(11):3116–3128
Yousefvand S, Hamidi F (2020) Role of paraventricular nucleus in regulation of feeding behaviour and the design of intranuclear neuronal pathway communications. Int J Pept Res Ther 26(3):1231–1242
Yousefvand S, Hamidi F (2021) The role of ventromedial hypothalamus receptors in the central regulation of food intake. Int J Pept Res Ther 27(1):689–702
Yousefvand S, Hamidi F, Zendehdel M, Parham A (2018a) Effects of insulin and somatostatin on water intake in neonatal chickens. Iran J Physiol Pharmacol 2(3):165–158
Yousefvand S, Hamidi F, Zendehdel M, Parham A (2018b) Hypophagic effects of insulin are mediated via NPY1/NPY2 receptors in broiler cockerels. Can J Physiol Pharmacol 96(12):1301–1307
Yousefvand S, Hamidi F, Zendehdel M, Parham A (2019) Interaction of neuropeptide Y receptors (NPY1, NPY2 and NPY5) with somatostatin on somatostatin-induced feeding behaviour in neonatal chicken. Br Poult Sci 60(1):71–78
Yousefvand S, Hamidi F, Zendehdel M, Parham A (2020) Survey the effect of insulin on modulating feed intake via NPY receptors in 5-day-old chickens. Int J Pept Res Ther 26(1):467–476
Zendehdel M, Hassanpour S (2014) Central regulation of food intake in mammals and birds: a review. Neurotransmitter 1:1–7
Zendehdel M, Hassanpour S (2014b) Ghrelin-induced hypophagia is mediated by the β 2 adrenergic receptor in chicken. J Physiol Sci 64(5):383–391
Zendehdel M, Hamidi F, Babapour V, Mokhtarpouriani K, Fard RM (2012) The effect of melanocortin (Mc3 and Mc4) antagonists on serotonin-induced food and water intake of broiler cockerels. J Vet Sci 13(3):229
Zendehdel M, Mokhtarpouriani K, Babapour V, Pourrahimi M, Hamidi F (2013a) The role of 5-HT2A and 5-HT2C receptors on harmalineinduced eating behavior in 24-h food-deprived broiler cockerels. Iran J Vet Res 14(2):94–99
Zendehdel M, Mokhtarpouriani K, Hamidi F, Montazeri R (2013b) Intracerebroventricular injection of ghrelin produces hypophagia through central serotonergic mechanisms in chicken. Vet Res Commun 37(1):37–41
Zendehdel M, Hamidi F, Hassanpour S (2015) The effect of histaminergic system on nociceptin/orphanin FQ induced food intake in chicken. Int J Pept Res Ther 21(2):179–186
Zendehdel M, Parvizi Z, Hassanpour S, Baghbanzadeh A, Hamidi F (2017) Interaction between nociceptin/orphanin FQ and adrenergic system on food intake in neonatal chicken. Int J Pept Res Ther 23(1):155–161
Zendehdel M, Ebrahimi-Yeganeh A, Hassanpour S, Koohi MK (2019) Interaction of the dopaminergic and nociceptin/orphanin FQ on central feed intake regulation in chicken. Br Poult Sci 60(3):317–322
Zendehdel M, Hassanpour S, Movahedi N (2020) Central and peripheral methylamine-induced hypophagia is mediated via nitric oxide and TAAR1 in neonatal layer-type chicken. Neurosci Lett 739:135408
Zhang L, Hernandez-Sanchez D, Herzog H (2019) Regulation of feeding-related behaviors by arcuate neuropeptide Y neurons. Endocrinology 160(6):1411–1420
Zhu Y, Yamanaka A, Kunii K, Tsujino N, Goto K, Sakurai T (2002) Orexin-mediated feeding behavior involves both leptin-sensitive and-insensitive pathways. Physiol Behav 77(2–3):251–257
Acknowledgements
The authors thank the Ferdowsi University of Mashhad for their cooperation and support of this article.
Funding
This review article did not receive any specific grant from funding agencies in the public, commercial, and not-for-profit sectors.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical Approval
In this review article, no laboratory work has been done on animals.
Research Involving Human Participants and/or Animals
No humans/or animals participated in this review article.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Yousefvand, S., Hamidi, F. Role of Lateral Hypothalamus Area in the Central Regulation of Feeding. Int J Pept Res Ther 28, 83 (2022). https://doi.org/10.1007/s10989-022-10391-4
Accepted:
Published:
DOI: https://doi.org/10.1007/s10989-022-10391-4