
Abstract
It has been stated that central injection of ghrelin is acting as an anorexigenic peptide in chicken. Ghrelin activity was studied through some neuronal pathways. The present study was designed in 4 experiments to examine the hypophagic response of ghrelin through the central serotonergic system in chicken. The guide cannula was surgically implanted in the right lateral ventricle of the chickens. In experiment 1, intacerebroventricular injection with PCPA (1.5 mg) performed followed by ghrelin (0.6 nmol). In experiments 2, 3 and 4 prior to ghrelin injection, chickens received fluoxetine (10 μg), 8-OH-DPAT (15.25 nmol), SB242084 (1.5 μg) respectively via guide cannula intacerebroventricularly. Cumulative food intake was determined at 3 h post injection. The results of this study showed that flouxetine pretreatment significantly amplified ghrelin hypophagia in chicken (p < 0.05). The hypophagic effect of ghrelin was attenuated by pretreatment with PCPA and SB242084 (p < 0.05) but 8-OH-DPAT had no effect. These results suggest that hypophagic effect of ghrelin probably is mediated by serotonergic mechanisms via 5-HT2C receptor.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
A number of peptides comprise a complex network that regulates feeding behavior in vertebrates (Volkoff et al. 2005; Furuse et al. 2007; Valassi et al. 2008) including ghrelin, which was originally identified as a growth hormone (GH)-releasing peptide and isolated from the rat and human stomach (Kojima et al. 1999). Avian ghrelin was first isolated from chicken proventriculus and found to be a 26-amino acid peptide sharing 54 % amino acid sequence identity with rat and human ghrelin (Kaiya et al. 2002). Intracerebroventricular (ICV) or intraperitoneal (IP) injection of ghrelin stimulates food intake in rodents (Wren et al. 2001). It is known that, in rats, central and peripheral ghrelin increases food intake mainly through activation of neuropeptide Y and agouti-related protein (Wren et al. 2000; Kamegai et al. 2001). Interestingly, ghrelin has an opposite effect on food intake in the chicken compared to rat and human. In chickens, systemic and ICV injection of ghrelin or other growth hormone secretagogues like GHRP-2 strongly inhibits food intake in a dose- dependent manner in chickens (Furuse et al. 2001; Khan et al. 2006; Saito et al. 2005). The mechanism underlying the ghrelin-induced hypophagia in birds is still unclear, although the anorexic effect of ghrelin was reported to be mediated by corticotropin-releasing factor (CRF) but not neuropeptide Y (Saito et al. 2005).
Serotonergic (5-HTergic) mechanisms are involved in the regulation of numerous physiological functions. Central 5-HTergic mechanisms appear to have important roles in the control of ingestive behavior also in avian species. In birds, a dense 5-HTergic fiber distribution has been identified in diencephalic regions known to be involved in controlling feeding behavior (Parent 1981; Duchala et al. 1984; Sako et al. 1986; Challet et al. 1996). Anorexic and satiety responses involve 5-HT2C receptors and possibly 5-HT1A sites found in hypothalamic loci responsible for primary integration of feeding behavior (Blundell 1984, 1991; Curzon 1990).
On the basis of these findings and considering the same effects of ghrelin and 5-HT on feeding behavior in birds, we hypothesized that 5-HTergic system possibly mediates ghrelin signaling in the hypothalamus of birds. Thus, the present study was designed to investigate whether blocking or stimulating 5-HT receptors can influence ghrelin-induced feeding response in 3 h food-deprived chickens.
Materials and methods
Animals
Chickens (Eshragh Co. Iran) were reared in heated batteries with continuous lighting until 3 weeks of age. Birds were provided with a mash diet (21 % protein and 2,869 kcal/kg of metabolizable energy) and water ad libitum. At approximately 2 weeks of age, the birds were transferred to individual cages. The temperature and relative humidity of the animal room were maintained at 22 ± 1 °C and 50 %, respectively, in addition to the continuous lighting condition.
Drugs
Ghrelin, Fluoxetine (selective 5-HT reuptake inhibitor–SSRI), Parachlorophenylalanine (PCPA) (a brain’s 5-HT depletion), 8-OH-DPAT (5-HT1A receptor agonist) and SB-242,084 (selective antagonist for the 5-HT2C receptor) were purchased from Sigma (USA). All solutions were prepared in pyrogen-free 0.9 % NaCl solution (saline) that served as control.
Surgical preparation
At 3 weeks of age, chickens were anesthetized with sodium pentobarbital (Sagatal, Rhone Merieux) (25 mg/kg body weight, iv) and a 23-gauge thin-walled stainless steel guide cannula was stereotaxically implanted into the right lateral ventricle, according to the technique described previously (Denbow et al. 1981). The stereotaxic coordinates were AP = 6.7, L = 0.7, H = 3.5–4 mm below the duramater with the head oriented as described (Van Tienhoven and Juhasz 1962). The cannula was secured with three stainless steel screws placed in the calvaria surrounding each guide cannula, then acrylic dental cement (Pars acryl) was applied to the screws and guide cannula. An orthodontic # 014 wire (American Orthodontics) trimmed to the exact length of the guide cannula was inserted into the guide cannula while the chicks were not being used for experiments. Lincospectin (Razak) was applied to the incision to prevent infection. The birds were allowed a minimum of 5 days recovery prior to injection.
Experimental procedures
To determine the possible effects of ghrelin on food intake, four experiments were conducted. Twenty-four birds were used in each experiment (n = 6 for each group). Injections were made with a 29-gauge, thin-walled stainless steel injecting cannula which extends 1.0 mm beyond the guide cannula. This injecting cannula was connected through a 60-cm-long PE-20 tubing to 10-μl Hamilton syringe. Solutions were injected over a period of 60 s. A further 60-s period was allowed to permit the solution to diffuse from the tip of the cannula into the ventricle. Placement of the guide cannula into the ventricle was verified by the presence of cerebrospinal fluid and ICV injection of methylene blue followed by slicing the frozen brain tissue at the end of the experiments. All experimental procedures were performed during 10:00 a.m. to 02:00 p.m. period. Before injection, the birds were removed from their individual cages, restrained by hand, and then put back into their cages after injections. Birds were handled and mock injected daily during the 5-day recovery period, in order to be used to the injection procedure. Tubing and syringes were kept in 70 % ethanol, and the glassware was autoclaved to render materials pyrogen-free. Three hours before the start of the experiments, animals were deprived of food (FD3). Each individual bird received two injections with 15 min interval. Immediately after the second injection fresh food was supplied and cumulative food (gr) intake was recorded at 15, 30, 60, 120 and 180 min. In this study, all control groups like treatment groups received two injections, 5 μl saline, with 15 min interval.
In experiments 1, the first injection consisted of either 0 or 1.5 mg PCPA in 5-μl saline and the second injection consisted of either 0 or 0.6 nmol ghrelin in 5-μl saline.
Experiments 2, 3 and 4 were conducted parallel to the experiment 1, except that the chicks received 0 or 10 μg fluoxetine, 0 or 15.25 nmol 8-OH-DPAT, 0 or 1.5 μg SB242084, respectively instead of PCPA. Each birds received two injections with 15 min interval and cumulative food intake (gr) was recorded at 15, 30, 60, 120 and 180 min after the second injection.
In this study, the selection of PCPA, fluoxetine, 8-OH-DPAT and SB242084 doses were determined based on the preliminary and previous studies (Medeiros et al. 2005; von Meyenburg et al. 2003; Steffens et al. 1997; Lazartigues et al. 2000; Taati et al. 2011). These doses alone had no effect on food intake in FD3 chickens.
Statistical analysis
Cumulative food intake was analyzed by one-way analysis of variance (ANOVA), and is presented as mean ± SEM. For treatment showing a main effect by ANOVA, means have compared by post hoc bonferroni test. P < 0.05 was considered as significant differences between treatments.
Results
The food intake response to ICV injection of PCPA, fluoxetine, 8-OH-DPAT and SB-242084 in chickens is presented in Figs. 1, 2, 3 and 4.

Effect of ICV injection of PCPA (1.5 mg) followed by ghrelin (0.6 nmol) on food intake in 3-h food deprived broiler cockerels. Data are presented as mean ± SEM. Different letters (a and b) indicate significant differences between treatments (P<0.05)

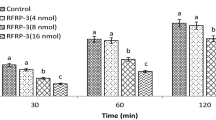
Effect of ICV injection of fluoxetine (10 µg) followed by ghrelin (0.6 nmol) on food intake in 3-h food deprived broiler cockerels. Data are presented as mean ± SEM. Different letters (a, b and c) indicate significant differences between treatments (P<0.05)

Effect of ICV injection of 8-OH-DPAT (15.25 nmol) followed by ghrelin (0.6 nmol) on food intake in 3-h food deprived broiler cockerels. Data are presented as mean ± SEM. Different letters (a and b) indicate significant differences between treatments (P<0.05)

Effect of ICV injection of SB242084 (1.5 µg) followed by ghrelin (0.6 nmol) on food intake in 3-h food deprived broiler cockerels. Data are presented as mean ± SEM. Different letters (a and b) indicate significant differences between treatments (P<0.05)
In experiment 1, the inhibitory effect of ghrelin on food intake was decreased by 1.5 mg PCPA pretreatment (P < 0.05), but 1.5 mg PCPA alone could not alter food intake in FD3 chickens (Fig. 1).
In experiment 2, hypophagic effect of ghrelin was significantly increased by administration of 10 μg fluoxetine in FD3 chickens (Fig. 2) (P < 0.05), while 10 μg fluoxetine alone had no effect on food intake.
In experiment 3, pretreatment with 15.25 nmol 8-OH-DPAT had no effect on the anorexia induced by ghrelin in FD3 chickens (Fig. 3) (P > 0.05). Furthermore, this dose of 8-OH-DPAT had no effect on food intake.
In experiment 4, the hypophagic effect of ghrelin was significantly attenuated by SB-242084 pretreatment in FD3 chickens (Fig. 4) (P < 0.05). Also, 1.5 μg SB242084 alone had no effect on food intake in chickens.
Discussion
The present study designed for the first time to investigate the possible involvement of 5-HTergic circuits in ghrelin control mechanisms of feeding behavior in chicken. The results obtained from experiment 1 (Fig. 1) show that ghrelin decreases food intake and pretreatment with PCPA attenuate this effect of ghrelin. It is known that opposite to mammals ICV or IV administration of ghrelin strongly inhibits food intake in a dose dependent manner in chickens (Furuse et al. 2001). Saito et al. (2005) demonstrated that the inhibitory effect of central ghrelin on food intake in chickens is caused by activating the CRF system, so that co-injection of a CRF receptor antagonist, astressin, attenuated ghrelin-induced anorexia in neonatal chicks (Saito et al. 2005). Also, as demonstrated in neonatal chickens, the CRF system may mediate the inhibitory action of ghrelin on food intake in birds whereas in mammals it appears that the stimulatory effects of ghrelin on food intake are mediated by neuropeptides such as NPY and orexin (Toshinai et al. 2003). In this regard, the effect of ICV injection of CRF on various behaviors in chicks was determined at 15-min intervals over a 30-min period. Ohgushi et al. (2001) showed that food intake of chicks was significantly decreased, and pecking rhythm was significantly delayed by CRF during the first 15-min post-injection. Also, the effects of central injection of 5-HT in avian species have been sparsely studied. 5-HT exerts an inhibitory effect on feeding behavior in animals, including birds (Steffens et al. 1997), and studies with agonists have demonstrated that 5-HT acts on different receptor subtypes. In 24-h food-deprived or satiated leghorn chicken, ICV injection of 5-HT induces potent decrease in food intake (Denbow 1989; Saadoun and Cabrera 2002; Zendehdel et al. 2012). In this study we showed that inhibitory effect of ghrelin on food intake was significantly attenuated with PCPA pretreatment (Fig. 1), and this effect of ghrelin was amplified with pretreatment of fluoxetine (Fig. 2). In addition, the decreased food consumption induced by the ICV injection of ghrelin was not significantly attenuated by pretreatment with 8-OH-DPAT (5-HT1A receptor agonist) (Fig. 3). While, pretreatment with SB242084 (selective antagonist for the 5HT2C receptor) significantly attenuated hypophagic effect of ghrelin in chickens (Fig. 4). Based on this study maybe there is an interaction between ghrelin and 5-HT neurons on feeding behavior in chicken. In birds it appears that ghrelin activates CRF neurons and that CRF released from these neurons is involved in inhibiting food intake. However, it is also possible that other factors participate in decreasing food intake (Saito et al. 2005). Furthermore, the effects of 5-HTergic agonists and antagonists on CRF secretion have been investigated by explanted rat hypothalamus. In this regard, 5-HT stimulates CRF secretion by explanted rat hypothalami and that this effect appears to be mediated mainly through a 5HT2c receptor mechanism (Calogero et al. 1989). In chickens, both CRF and 5-HT stimulate the hypothalamo-pituitary-adrenal (HPA) axis and the stimulatory role of 5-HT on CRF secretion was mediated mainly through 5HT2c receptor (Zhang et al. 2004). It has been also reported that pharmacological blockade of 5-HT reuptake by fluoxetine increases CRF in the portal plasma (Gibbs and Vale 1983). Taken together it is concluded that ghrelin probably stimulates 5-HT release and/or synthesis, thereby CRF secretion is stimulated resulting in food intake decreases in chicken.
In conclusion, our data in this study and previous studies shows that ghrelin can affect the 5-HTergic system in central regulation of food intake in chicken. Nevertheless, we cannot rule out the interaction between 5-HTergic and CRF systems in ghrelin induced inhibition of food intake in broiler chickens. It is known that ghrelin and 5-HT stimulate the release of CRF. On the other hand the appetite-inhibiting activity of ghrelin and 5-HT could be mediated by release of CRF. These results indicate that central ghrelin and 5-HT inhibit food intake and presumably these effects may occure on the same pathway by interacting with the endogenous CRF and its receptors.
References
Blundell JE (1984) Serotonin and appetite. Neuropharmacology 23(12B):1537–1551
Blundell J (1991) Pharmacological approaches to appetite suppression. Trends Pharmacol Sci 12(4):147–157
Calogero AE, Bernardini R, Margioris AN, Bagdy G, Gallucci WT, Munson PJ, Tamarkin L, Tomai TP, Brady L, Gold PW et al (1989) Effects of serotonergic agonists and antagonists on corticotropin-releasing hormone secretion by explanted rat hypothalami. Peptides 10(1):189–200
Challet E, Miceli D, Pierre J, Reperant J, Masicotte G, Herbin M, Vesselkin NP (1996) Distribution of serotonin-immunoreactivity in the brain of the pigeon (Columba livia). Anat Embryol (Berl) 193(3):209–227
Curzon G (1990) Serotonin and appetite. Ann N Y Acad Sci 600:521–530, discussion 530–521
Denbow DM (1989) Peripheral and central control of food intake. Poult Sci 68(7):938–947
Denbow DM, Cherry JA, Siegel PB, Van Krey HP (1981) Eating, drinking and temperature response of chicks to brain catecholamine injections. Physiol Behav 27(2):265–269
Duchala CS, Ottinger MA, Russek E (1984) The developmental distribution of monoamines in the brain of male Japanese quail (Coturnix coturnix japonica). Poult Sci 63(5):1052–1060
Furuse M, Tachibana T, Ohgushi A, Ando R, Yoshimatsu T, Denbow DM (2001) Intracerebroventricular injection of ghrelin and growth hormone releasing factor inhibits food intake in neonatal chicks. Neurosci Lett 301(2):123–126
Furuse M, Yamane H, Tomonaga S, Tsuneyoshi Y, Denbow DM (2007) Neuropeptidergic regulation of food intake in the neonatal chick: a review. J Poult Sci 44(4):349–356
Gibbs DM, Vale W (1983) Effect of the serotonin reuptake inhibitor fluoxetine on corticotropin-releasing factor and vasopressin secretion into hypophysial portal blood. Brain Res 280(1):176–179
Kaiya H, van der Geyten S, Kojima M, Hosoda H, Kitajima Y, Matsumoto M, Geelissen S, Darras VM, Kangawa K (2002) Chicken ghrelin: purification, cDNA cloning, and biological activity. Endocrinology 143(9):3454–3463. doi:10.1210/en.2002-220255
Kamegai J, Tamura H, Shimizu T, Ishii S, Sugihara H, Wakabayashi I (2001) Chronic central infusion of ghrelin increases hypothalamic neuropeptide Y and Agouti-related protein mRNA levels and body weight in rats. Diabetes 50(11):2438–2443
Khan MSI, Dodo K-I, Yahata K, Nishimoto S, Ueda H, Taneike T, Kitazawa T, Hosaka Y, Bungo T (2006) Intracerebroventricular administration of growth hormone releasing peptide-6 (GHRP-6) inhibits food intake, but not food retention of crop and stomach in neonatal chicks. J Poult Sci 43(1):35–40
Kojima M, Hosoda H, Date Y, Nakazato M, Matsuo H, Kangawa K (1999) Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature 402(6762):656–660. doi:10.1038/45230
Lazartigues E, Brefel-Courbon C, Bagheri H, Costes S, Gharib C, Tran MA, Senard JM, Montastruc JL (2000) Fluoxetine-induced pressor response in freely moving rats: a role for vasopressin and sympathetic tone. Fundam Clin Pharmacol 14(5):443–451
Medeiros MA, Costa-e-Sousa RH, Olivares EL, Cortes WS, Reis LC (2005) A reassessment of the role of serotonergic system in the control of feeding behavior. An Acad Bras Cienc 77(1):103–111
Ohgushi A, Bungo T, Shimojo M, Masuda Y, Denbow DM, Furuse M (2001) Relationships between feeding and locomotion behaviors after central administration of CRF in chicks. Physiol Behav 72(1–2):287–289
Parent A (1981) Comparative anatomy of the serotoninergic systems. J Physiol (Paris) 77(2–3):147–156
Saadoun A, Cabrera MC (2002) Effect of the 5-HT1A receptor agonist 8-OH-DPAT on food and water intake in chickens. Physiol Behav 75(3):271–275. doi:10.1016/s0031-9384(01)00665-5
Saito ES, Kaiya H, Tachibana T, Tomonaga S, Denbow DM, Kangawa K, Furuse M (2005) Inhibitory effect of ghrelin on food intake is mediated by the corticotropin-releasing factor system in neonatal chicks. Regul Pept 125(1–3):201–208. doi:10.1016/j.regpep. 2004.09.003
Sako H, Kojima T, Okado N (1986) Immunohistochemical study on the development of serotoninergic neurons in the chick: I. Distribution of cell bodies and fibers in the brain. J Comp Neurol 253(1):61–78. doi:10.1002/cne.902530106
Steffens SM, Casas DC, Milanez BC, Freitas CG, Paschoalini MA, Marino-Neto J (1997) Hypophagic and dipsogenic effects of central 5-HT injections in pigeons. Brain Res Bull 44(6):681–688
Taati M, Nayebzadeh H, Zendehdel M (2011) The effects of DL-AP5 and glutamate on ghrelin-induced feeding behavior in 3-h food-deprived broiler cockerels. J Physiol Biochem 67(2):217–223. doi:10.1007/s13105-010-0066-y
Toshinai K, Date Y, Murakami N, Shimada M, Mondal MS, Shimbara T, Guan JL, Wang QP, Funahashi H, Sakurai T, Shioda S, Matsukura S, Kangawa K, Nakazato M (2003) Ghrelin-induced food intake is mediated via the orexin pathway. Endocrinology 144(4):1506–1512
Valassi E, Scacchi M, Cavagnini F (2008) Neuroendocrine control of food intake. Nutr Metab Cardiovasc Dis 18(2):158–168. doi:10.1016/j.numecd.2007.06.004
Van Tienhoven A, Juhasz LP (1962) The chicken telencephalon, diencephalon and mesencephalon in sterotaxic coordinates. J Comp Neurol 118:185–197
Volkoff H, Canosa LF, Unniappan S, Cerda-Reverter JM, Bernier NJ, Kelly SP, Peter RE (2005) Neuropeptides and the control of food intake in fish. Gen Comp Endocrinol 142(1–2):3–19. doi:10.1016/j.ygcen.2004.11.001
von Meyenburg C, Langhans W, Hrupka BJ (2003) Evidence that the anorexia induced by lipopolysaccharide is mediated by the 5-HT2C receptor. Pharmacol Biochem Behav 74(2):505–512
Wren AM, Small CJ, Ward HL, Murphy KG, Dakin CL, Taheri S, Kennedy AR, Roberts GH, Morgan DG, Ghatei MA, Bloom SR (2000) The novel hypothalamic peptide ghrelin stimulates food intake and growth hormone secretion. Endocrinology 141(11):4325–4328
Wren AM, Small CJ, Abbott CR, Dhillo WS, Seal LJ, Cohen MA, Batterham RL, Taheri S, Stanley SA, Ghatei MA, Bloom SR (2001) Ghrelin causes hyperphagia and obesity in rats. Diabetes 50(11):2540–2547. doi:10.2337/diabetes.50.11.2540
Zendehdel M, Hamidi F, Babapour V, Mokhtarpouriani K, Mazaheri Nezhadfard R (2012) The effect of melanocortin (Mc3 and Mc4) antagonists on serotonin- induced food and water intake of broiler cockerels. J Vet Sci 13(3):229–234. doi:10.4142/jvs.2012.13.3.229
Zhang R, Tachibana T, Takagi T, Koutoku T, Denbow DM, Furuse M (2004) Serotonin modifies corticotropin-releasing factor-induced behaviors of chicks. Behav Brain Res 151(1–2):47–52. doi:10.1016/j.bbr.2003.08.005
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Zendehdel, M., Mokhtarpouriani, K., Hamidi, F. et al. Intracerebroventricular injection of ghrelin produces hypophagia through central serotonergic mechanisms in chicken. Vet Res Commun 37, 37–41 (2013). https://doi.org/10.1007/s11259-012-9544-8
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11259-012-9544-8