
Abstract
Parthenocarpy is becoming an essential trait for off-season greenhouse production of Zucchini squash. Given that winter conditions promote a reduction in the number of male flowers and in the activity of pollinators, the application of synthetic auxins is currently the most widespread method to induce fruit set. We have evaluated the parthenocarpic tendency of 48 long-fruited accessions of Cucurbita pepo spp. pepo, from morphotypes Zucchini, Vegetable marrow and Cocozelle, including 45 traditional cultivars and 3 commercial hybrids, with the goal of identifying new sources of parthenocarpy for breeding programs. After the first screening, 20 selected accessions were evaluated for the growth rate of unpollinated fruit. Twelve of the selected accessions identified as either strongly parthenocarpic or non-parthenocarpic, were compared for fruit rate growth, ethylene production and ethylene sensitivity. Apart from the three control hybrids, the fastest parthenocarpic fruit growth was observed in ‘CpCAL112’, ‘CM-37’, ‘E-27’, ‘PI261610’, and ‘V-185’. The source of the parthenocarpy of some of these accessions differs from that of the hybrids as it was not associated with the conversion of female into bisexual flowers or with the so-called “fruits with attached flowers” syndrome, which is an undesirable trait in current parthenocarpic hybrids. The alternative sources of parthenocarpy may be of great importance in current Zucchini breeding programs. We also demonstrate that the parthenocarpy of these accessions is associated with downregulation of ethylene production in unpollinated fruits during the first days post anthesis (DPA). In non-parthenocarpic accessions, unpollinated fruits boosted ethylene production at 3 DPA, concomitantly with fruit abortion and senescence, while in parthenocarpic accessions, fruits produced little ethylene at 3 DPA. Therefore, ethylene production in ovaries/fruits at 3 DPA can be used as a marker to identify and select parthenocarpy in Zucchini squash. However, in the cultivars tested here, ethylene production and sensitivity in vegetative organs and in male flowers earlier than 3 PDA do not appear well associated with parthenocarpy.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Cucurbita pepo is one of the most important species of the Cucurbitaceae family as regards both its worldwide distribution and its economic value (Nee 1990). Based on different molecular studies, the species has been divided into two subspecies: C. pepo ssp. pepo and C. pepo ssp. ovifera (Decker 1988; Jobst et al. 1998; Katzir et al. 2000). Moreover, the variability in size, shape, color and texture of fruits has led to the classification of C. pepo into eight different botanical morphotypes: Scallop, Acorn, Crookneck and Straightneck are included in the spp. ovifera, and Pumpkin, Vegetable Marrow, Zucchini and Cocozelle in the ssp. pepo (Paris 1986, 2000).
Cucurbita pepo is a monoecious crop with two or three sexual phases of development. In the first phase only male flowers are produced. This is followed by a phase during which male and female flowers alternate, and the final phase consists of only female flowers (Peñaranda et al. 2007; Manzano et al. 2010, 2013). This latter phase restricts pollination and fruit set, but only occurs in certain cultivars when grown under winter conditions (Peñaranda et al. 2007). Since female and male sexual organs are separated in unisexual flowers, fruit set and development in squash requires the activity of pollinators, a process that is restricted to the 6 h that male and female flowers remain open during the morning (Nepi and Pacini 1993), and which is also highly dependent on environmental conditions that affect both the number of female and male flowers per plant and pollinator activity (Wien 2002; Loy 2012). In off-season greenhouse production, i.e. under conditions in which plants produce a reduced number of male flowers and the activity of pollinators is very limited, natural pollination is difficult, and fruit set is currently induced by synthetic auxins (Sanz 1995; Wien 2002). Nevertheless, the demand for healthier and “greener” fresh produce is making parthenocarpy a priority trait in current breeding programs of Zucchini.
Cucumber, Cucumis sativus, was the first cucurbit species in which the introduction of parthenocarpic cultivars improved off-season production in greenhouses (de Ponti and Garretsen 1976), but parthenocarpy has been detected in other species of the family, including melon and squash (Rylski and Aloni 1990). The earliest studies in the parthenocarpy of C. pepo were made by Durham (1925), who closed 301 female flowers to prevent pollination, and found no parthenocarpic production in squash. The first squash variety described as parthenocarpic was ‘Royal Acorn’ (Nitsch et al. 1952), although its parthenocarpic potential was later questioned (Robinson and Reiners 1999). Subsequent studies identified a number of cultivars with parthenocarpic potential, including ‘Dg-4’ and ‘Poseidon’ (den Nijs and Balder 1983); the cultivar ‘Chefini’ (Robinson 1993), and ‘Whitaker’, whose parthenocarpy appears to be controlled by one major dominant gene (de Menezes et al. 2005). den Nijs and van Zanten (1982), Om and Hong (1989) and Robinson and Reiners (1999) studied the parthenocarpic potential of different C. pepo cultivars, concluding that the highest level of parthenocarpy is found in accessions with dark green skin from the morphotypes Zucchini and Cocozelle. It appears, therefore, that cultivars with elongated fruits exhibit the highest parthenocarpic fruit production. Nevertheless, the production of parthenocarpic fruits in a given cultivar is quite dependent on environmental conditions. It is known that winter conditions, especially low temperatures, are able to promote parthenocarpy in C. pepo (Globerson 1971; Rylski 1974; den Nijs and Balder 1983; Rylski and Aloni 1990; Robinson and Reiners 1999; Gómez et al. 2004).
Despite the above studies, to date few Zucchini squash cultivars are marketed as parthenocarpic. The previously identified parthenocarpic cultivars, such as ‘Whitaker’ and ‘Chefini’, showed no parthenocarpic potential in greenhouse production in the southeast of Spain (unpublished data). Under these specific conditions, we found only three hybrids that are commercialized as parthenocarpic: ‘Cavili’, ‘Partenon’ and ‘Argo’. These hybrids have been subjected to different evaluations by our research group since 2004, and they all produce parthenocarpic fruits with attached flowers, an undesirable associated trait that results from the delay in maturation and abscission of floral organs (Peñaranda et al. 2007; Martínez et al. 2013). As the fruit is not commercialized with the flower, its manual abscission causes a wound which leads to rapid loss of water and decay during postharvest storage. In ‘Cavili’, parthenocarpic fruits have been found to be derived not from female flowers but from bisexual ones, which are induced in these cultivars when greenhouse temperature rises above 30 °C (Martínez et al. 2013). This type of parthenocarpy has no value for Zucchini greenhouse production, as the temperature easily exceeds 30 °C not only during spring and summer, but also on occasions in the autumn–winter season. Given these considerations, it would be necessary to identify new sources of parthenocarpy for Zucchini greenhouse breeding programs.
Fruit set and subsequent fruit growth is a vital phase of plant development that is largely dependent on the biosynthesis and crosstalk of phytohormones, such as auxins, GAs and cytokinins, produced in the pollinated ovaries (Ozga and Reinecke 2003; Srivastava and Handa 2005; de Jong et al. 2009). To date, however, the involvement of other hormones such as ethylene in this developmental process has not been studied in depth. Our research group has recently found that fruit set and early fruit development of Zucchini after pollination and fertilization requires a low level of ethylene in the pollinated ovaries for a few days after anthesis, and that the lack of pollination induces a boost of ethylene in the fruit 3 days after anthesis, which is concomitant with fruit abortion and senescence (Martínez et al. 2013). Therefore, although the parthenocarpic development of the fruit is normally achieved by the coordinated action of GAs, auxins and cytokinins (Srivastava and Handa 2005; Serrani et al. 2010), we have recently demonstrated that the parthenocarpy of the cultivar ‘Cavili’ is also associated with a low level of ethylene in the unpollinated fruit for a few days after anthesis (Martínez et al. 2013). This reduction in ethylene production could be responsible for the conversion of female into bisexual flowers and the delay in the maturation and abscission of floral organs that is normally associated with the parthenocarpy of this cultivar (Payán et al. 2006; Peñaranda et al. 2007; Martínez et al. 2013).
Our objective was to screen C. pepo accessions for parthenocarpic tendency and to determine whether ethylene production is associated with parthenocarpy. Results indicate the existence of a negative correlation between the production of ethylene in the unpollinated ovaries of squash at 3 DPA and the parthenocarpic development of fruit.
Materials and methods
Plant material and culture conditions
The present study used forty-five accessions of C. pepo from the core collection of the germplasm bank at the Polytechnic University of Valencia (COMAV), as well as certain accessions from the seed bank of the University of Almería (BSUAL) (Table 1). All produce fruits of elongated phenotype, belonging to types Zucchini, Vegetable Marrow and Cocozelle of C. pepo spp. pepo. The evaluated germplasm represents Spanish accessions from different regions of the Iberian peninsula, mainly Valencia and Andalucía, but also from the Canary Islands. However, four of the accessions originated in Greece, Yugoslavia, Morocco and Turkey (Table 1). The hybrids ‘Argo’, ‘Cavili’ and ‘Partenon’ were used as positive controls of parthenocarpy.
Evaluations were made in autumn–winter 2009–2010 and spring 2011. On both occasions, seeds from each accession were germinated on nursery trays under high humidity conditions for a period of 15 days, and then transplanted to soil. Plants were grown in a greenhouse in Almeria (Spain) following standard local practices for both plant nutrition and pest and disease control.
Phenotypic evaluation of accessions
In 2009–2010 ten plants of each of the 48 accessions were classified according to the fruit morphotype, and evaluated for their habit of growth (bushy or vine), monoecy stability (monoecious or partially andromonoecious), and for the level of parthenocarpy (Table 1). From this first evaluation, we selected a number of accessions that were again evaluated in 2011 for parthenocarpy, monoecy stability and ethylene production and sensitivity.
We have previously observed that under high temperature conditions, certain accessions of C. pepo show unstable monoecy (or partial andromonoecy), characterized by the partial conversion of female into bisexual flowers with differing degrees of stamen development (Martínez et al. 2014). To evaluate monoecy instability, pistillate flowers were scored from 0 to 3 according to their degree of stamen development. Female flowers with no stamen development were scored as 0, flowers with primordial stamens as 1, those with medium-sized stamens and anthers as 2, and bisexual flowers with complete stamens and anthers able to produce pollen were scored as 3. On the basis of these bisexuality scores, we defined an andromonoecy index (AI) for each accession, calculated as the average bisexuality score from at least 10 plants with a minimum of 5 pistillate flowers evaluated per plant. Plants and accessions with an AI of 0–0.9 were considered to be monoecious, while those with scores of 1–3 were considered unstable for monoecy or partially andromonoecious (Martínez et al. 2014).
To evaluate parthenocarpy, plants were grown in a greenhouse free of pollinators and other insects, which ensured that all the fruits were parthenocarpic. In the first trial, conducted in autumn–winter 2009–2010 with 10 plants of each accession, cultivars were scored from 0 to 5, according to the number of marketable parthenocarpic fruits produced in 10 days. The score of each accession was the mean of three evaluations, and scoring was as follows: at least 10 marketable fruits scored 5, 8 to 10 fruits scored 4, 5 to 7 fruits 3, 2 to 4 fruits scored 2, 1 fruit scored 1 and no fruits scored 0. On the basis of these results, some accessions having diverse scores were selected for re-evaluation. In this second trial, parthenocarpy was evaluated by measuring the length and width of at least 10 parthenocarpic fruits of each accession over a total of 7 days post-anthesis (DPA). Measurements were taken at anthesis and 3, 5 and 7 DPA. For fruits whose growth aborted before 7 days, the last measurement before abortion was considered in the subsequent days. The findings allowed us to determine the mean fruit growth rate in each accession, and to compare the increase in the fruit length and width between anthesis and each of the control points (3, 5 and 7 DPA) for each accession.
To assess the ethylene sensitivity of each accession, 10 male flower buds (1–2 days before anthesis) were removed from the plant and placed in glass jars with water in a hermetic 50 L container. The floral buds were incubated in 20 ppm of ethylene for 2 days, and the number of hours until abscission was scored in each flower up to a total of 3 days post-treatment. As a control, the same number of male flowers were treated with air and maintained in the same conditions. The assay was performed twice, and ethylene sensitivity was measured as the percentage of reduction in abscission time caused by the ethylene treatment.
Ethylene production in vegetative organs was determined in 3 replicates per accession, each containing at least 10 young leaves of 2–3 cm length. Leaves were excised from the plant and incubated at room temperature for 24 h in sealed containers in the dark for the accumulation of ethylene. On the other hand, ethylene production in fruits at 3 DPA was also measured in 3 replicates per accession, each containing 3 fruits. In this case 3 fruits at the same developmental stage were harvested and enclosed in sealed containers for 6 h. Ethylene production was determined in each sample 3 times in a Varian 3900 gas chromatograph fitted with a flame ionization detector (FID), previously calibrated to determine ethylene production.
Statistical analysis
In order to determine differences in fruit development and ethylene production and sensitivity among accessions, mean and standard error and standard deviations were calculated. Dependence between variables was studied using regression analysis. The results were evaluated fitting a linear model. The ANOVA table tells us whether there is a relationship between variables: the F statistic tests the null hypothesis, i.e. that the slope of the regression line is equal to 0 and there is no relation between variables; the R value indicates the strength of the relationship among variables; while the model p value establishes the significance level of the analysis. All analyses were performed with Statgraphics Plus v 5.1 software (Statpoint Technologies Inc, Warrenton, VA).
Results
Screening for parthenocarpy in C. pepo
The first assessment of parthenocarpy was carried out in autumn–winter 2009–2010, under conditions that favor the growth of parthenocarpic fruits. The three commercial hybrids (‘Argo’, ‘Cavili’ and ‘Partenon’) and 2 accessions (‘CpCAL112’ and ‘CpCAL110’) belonging to the morphotypes Zucchini and Cocozelle received scores of over 4 (Table 1). A total of 11 accessions that obtained a mean parthenocarpic score of over 2.75 were selected for further analysis. Another 6 accessions with a mean score of less than 1.50 were also selected as contrasting materials.
The C. pepo accessions were also classified according to their growth habit (vine or bushy type), sex expression (monoecious or partially andromonoecious), and fruit morphology (Zucchini, Vegetable Marrow or Cocozelle) (Table 1). In certain accessions some of these traits were still segregating, producing plants whose fruits were classified in different morphotypes. In accordance with visual observations and previous evaluations (Ferriol et al. 2003), ‘AFR-12’, ‘A-17’, ‘C-1’, ‘C-9’, ‘V-142’, ‘V-171’ and ‘V-32’ were classified as both Zucchini and Vegetable Marrow; ‘V-116’ and ‘V-185’ produced fruits of the morphotypes Zucchini and Cocozelle; while accessions ‘CA-154’ and ‘CpCAL097’ produced fruits of morphotypes Cocozelle and Vegetable Marrow (Table 1). In these cases, plants from different morphotypes were reproduced separately, and the offspring from plants belonging to the morphotype with the higher production of parthenocarpic fruits, normally Zucchini, were used for the second evaluation.
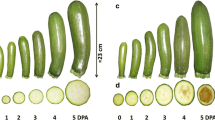
The second screening was conducted with 20 accessions selected from the first assay in spring 2011, under environmental conditions that did not favor the parthenocarpic growth of the fruit and where it could be easier, therefore, to discriminate between parthenocarpic and non-parthenocarpic accessions. Since the number of varieties tested was lower, we studied the longitudinal and diametrical growth rate of at least 10 unpollinated fruits of each accession from anthesis up to 7 DPA, when a pollinated fruit has already reached marketable size. In many of the accessions that were classified as non-parthenocarpic, the unpollinated fruit aborted after 3 DPA, while in others the fruit growth rate was so low that they did not reach marketable size at 7 DPA (Fig. 1). In some accessions, however, the growth of the fruit increased throughout the 7-day observation period and attained marketable size. Thus, these accessions were classified as parthenocarpic (Fig. 1). In 6 of the 11 accessions that were initially selected as parthenocarpic, ‘CpCAL110’, ‘AFR-12’, ‘PI169462’, ‘Pascual 40’, ‘Greece 6’ and ‘CpCAL044’, the potential parthenocarpic growth of their fruits proved insufficient to reach marketable size at 7 DPA (Fig. 1). These accessions were finally classified as non-parthenocarpic (Fig. 1).

Fruit length in 20 accessions of C. pepo during the first 7 days post-anthesis (DPA). The fruit length at each control point represents the mean of at least 10 unpollinated fruits from the same accession. The accession-specific fruit growth rates allowed the separation of parthenocarpic and non-parthenocarpic accessions in squash, defining as parthenocarpic those accessions whose fruits are able to reach 16 cm in length
Although the most parthenocarpic accessions had larger ovaries at anthesis, marketable fruit size was most consistently attained in accessions with the highest fruit growth rate (Fig. 1). We have compared the increase in fruit length between anthesis and 3, 5 and 7 DPA (Fig. 2). Results indicate that the most parthenocarpic accessions grew faster and that the greatest differences between accessions occur at 5 and 7 DPA (Fig. 2). Between anthesis and 3 DPA many accessions showed an increase in fruit length, but in some growth was arrested after 3 DPA, indicating that the increase in length at 3 DPA is not a suitable parameter to distinguish between parthenocarpic and non-parthenocarpic accessions (Fig. 2). However, if parthenocarpic accessions are those whose fruits reach marketable size at 7 DPA, the increase in fruit length between anthesis and 5 DPA or between anthesis and 7 DPA proves to be a simple and useful parameter for the identification of parthenocarpy in a given accession (Fig. 2). Moreover, if there is little variation in the ovary size at anthesis, the average size of 10 fruits at 5 or at 7 DPA could be a simpler parameter for evaluating the parthenocarpic potential of a cultivar in summer squash (Fig. 2).

Increase in fruit length 3, 5, and 7 DPA in 20 accessions of C. pepo ssp. pepo. Data indicate the mean of at least 10 fruits per accession
Association between parthenocarpy and ethylene production and sensitivity
Six parthenocarpic and six non-parthenocarpic genotypes were compared for tendency to andromonoecism and ethylene production and sensitivity in different plant organs. The Index of Andromonoecy (AI) (see “Materials and methods” section) was not associated with parthenocarpy in all the analyzed accessions (Fig. 3). In the hybrids ‘Cavili’, ‘Partenon’ and ‘Argo’, but also in accession ‘V-185’, the parthenocarpic development of the fruit was associated with the development of stamen in pistillate flowers and with AI higher than 1, but this association was not found in the parthenocarpic accessions ‘PI261610’ and ‘CpCAL112’ (Fig. 3). The latter accessions therefore represent one or two new sources of parthenocarpy to add to current commercial hybrids.

Regression of Andromonoecy Index (AI) onto parthenocarpic fruit growth rate during the first 7 DPA in six parthenocarpic and six non-parthenocarpic accessions of C. pepo. Although the parthenocarpy of the hybrids ‘Argo’, ‘Cavili’ and ‘Partenon’ is correlated with an AI higher than 1, i.e. a partial conversion of female into bisexual flowers, the parthenocarpic potential of accessions ‘CpCAL112’ and ‘PI261610’ is not associated with andromonoecy (AI = 0). Horizontal and vertical bars represent standard errors for AI or parthenocarpic fruit growth rate, respectively
The profile of ethylene production in pollinated and unpollinated fruits of each accession differed between parthenocarpic and non-parthenocarpic materials. In the non-parthenocarpic cultivars such as ‘CpCAL044’, pollination maintained or even reduced the production of ethylene during the days immediately after anthesis, while the lack of pollination induced a boost in ethylene production at 3 DPA (Fig. 4). However, in the parthenocarpic accessions this did not occur, and ethylene production remained at a low level in both pollinated and unpollinated fruits (Fig. 4). These data indicate that fruit set and early fruit development are correlated with a low level of ethylene at 3 DPA, while the induction of ethylene production at 3 DPA appears to be a signal that precedes abortion and senescence of unpollinated fruits in non-parthenocarpic accessions.

Ethylene production in pollinated and unpollinated ovaries/fruits of two accessions of C. pepo that differ in their parthenocarpic potential. a ‘CpCAL044’, which has been classified as non-parthenocarpic, and b ‘V-185’, which has been classified as parthenocarpic. Bars represent standard errors (n = 3). Ethylene production profiles in other parthenocarpic and non-parthenocarpic accessions were similar
We have observed a high negative correlation between parthenocarpy, expressed as the fruit length growth rate of unpollinated fruits during the first 7 DPA, and ethylene production at 3 DPA (Fig. 5). The fruit of parthenocarpic accessions produced less than 60 nL ethylene/g FW, whereas the fruit of the non-parthenocarpic accessions produced up to 180–190 nL/g FW (Fig. 5). Given that this peak of ethylene production at 3 DPA is practically absent in the unpollinated fruits of the parthenocarpic accessions, the lower production of ethylene in the parthenocarpic fruits during the days immediately after anthesis could be a good marker for the identification and selection of parthenocarpic accessions in C. pepo.

Regression of ethylene production in unpollinated fruits at 3 DPA onto fruit length growth rate for the first 7 DPA in six parthenocarpic and six non-parthenocarpic accessions of C. pepo. The linear regression analysis (R = −0.799, p = 0.0018) indicates a strong negative correlation between variables with a significance level of 99 %. Horizontal and vertical bars represent standard error for ethylene production and fruit length rate of growth, respectively, in each accession
As ethylene production in 3 DPA fruits is measurable only in mature plants, we have attempted to determine whether ethylene production in young plant organs could serve as an indicator of parthenocarpic tendency. Figure 6 shows the data of ethylene production in young leaves from plants which had already developed 10 true leaves. Although some parthenocarpic cultivars such as ‘Cavili’, ‘CpCAL112’ and ‘V-185’ showed reduced ethylene production in leaves, others like ‘Argo’ and ‘Partenon’ produced a high level of ethylene (Fig. 6), indicating that the reduced level of ethylene in the unpollinated fruits of parthenocarpic accessions is not expressed in other plant organs, but rather is specific to this developmental process. The production of ethylene in the leaves of the non-parthenocarpic accessions was very diverse, ranging from 27 to 151 nL/g FW. Therefore, the production of ethylene in leaves cannot be used as a marker for the selection of parthenocarpy in squash.

Regression of ethylene production in young leaves onto fruit length growth rate during the first 7 DPA in six parthenocarpic and six non-parthenocarpic accessions of C. pepo. The lineal regression analysis (R = −0.420; p = 0.1700) denotes no correlation between variables with a level of significance of 90 %. Horizontal and vertical bars represent standard error for ethylene production and fruit length rate of growth, respectively, in each accession
The sensitivity to ethylene in each accession was assessed by determining the effect of external ethylene treatments on the abscission time of male flowers. Male flowers at the same developmental stage of each accession were treated with air (control) or ethylene for 48 h, and the number of flowers that reached abscission was recorded every 12 h for 3 days following the treatment. The sensitivity to ethylene was expressed by comparing the times at which abscission occurred in ethylene treated flowers and in controls of the same accession. The regression analysis (R = −0.690; p = 0.0113) demonstrated a significant negative correlation between the level of parthenocarpy in each accession and sensitivity to ethylene (Fig. 7), but correlation was not as high as for the production of ethylene in fruits at 3 DPA. In fact, the parthenocarpic accessions ‘CpCAL112’ and ‘V-185’, as well as the parthenocarpic hybrids ‘Cavili’, ‘Partenon’ and ‘Argo’, showed a reduced response to ethylene, with values of below 20 %, while the parthenocarpic accession ‘PI261610’ was found to be highly sensitive to ethylene (Fig. 7). Moreover, all non-parthenocarpic accessions, except ‘AFR-12’, showed ethylene sensitivity values of over 25 %. Consequently, although parthenocarpy in squash appears to be correlated with reduced ethylene sensitivity, this rule does not hold true for certain accessions.

Regression of ethylene sensitivity (expressed as the percentage of time at which the abscission of ethylene treated male flowers is reduced with respect to non-treated control flowers) onto fruit length growth rate during the first 7 DPA in six parthenocarpic and six non-parthenocarpic accessions of C. pepo. The lineal regression analysis (R = −0.699, p = 0.0113) indicates a significant correlation between the two variables, with parthenocarpic accessions showing reduced ethylene sensitivity. Horizontal and vertical bars represent standard error for ethylene sensitivity and fruit growth rate, respectively, in each accession
Discussion
Sources of parthenocarpy for Zucchini breeding programs
With a view to searching for new sources of parthenocarpy that are useful for current Zucchini breeding programs in off-season greenhouse production, we have analyzed 45 accessions with elongated fruits from the core collection of the COMAV germplasm bank at the Polytechnic University of Valencia (Spain) and from the BSUAL at the University of Almería (Spain). In concordance with other previous studies (Om and Hong 1989; Robinson and Reiners 1999) under greenhouse environmental conditions, the highest parthenocarpic potential was detected in accessions having the most elongated fruits, i.e. morphotypes Zucchini and Cocozelle. In fact, four accessions that showed parthenocarpic potential in the trials of both 2009–2010 and 2011, ‘CpCAL112’, ‘PI261610’, ‘V-185’ and ‘E-27’, all belonged to the morphotype Zucchini or Cocozelle, and only one of the detected parthenocarpic accessions, ‘CM-37’, belonged to the morphotype Vegetable Marrow (Table 1; Figs. 1, 2). Moreover, all the parthenocarpic accessions detected in this paper produced dark green fruits, with the exception of ‘Cavili’, which is a commercial white-fruited hybrid. den Nijs and van Zanten (1982) and Robinson and Reiners (1999) also noted that Zucchini types with dark green fruit had the highest level of parthenocarpy, although not all dark green Zucchini accessions had good parthenocarpic fruit set either in this work or in previous studies.
The parthenocarpic accessions detected in this work are not all of the same origin. Four of the identified parthenocarpic accessions, ‘CM-37’, ‘PI261610’, ‘E-27’ and ‘V-185’, as well as six accessions that have been classified as non-parthenocarpic, had been previously included in genetic diversity studies of Spanish accessions of squash (Formisano et al. 2012). The parthenocarpic accessions ‘E-27’, ‘CM-37’, ‘PI261610’, ‘V-185’ of our study all belong to subspecies pepo, and were grouped close to each other when analyzed by SRAP molecular markers. These Spanish accessions are open pollinated and have been traditionally developed by farmers, which would also explain the diversity of morphotypes detected in some of the accessions (Ferriol et al. 2003).
Based on the study of Formisano et al. (2012) it was possible to separate two of the identified parthenocarpic accessions, ‘E-27’ and ‘V-185’. While ‘E-27’ was grouped among the traditional cultivars alongside others such as the non-parthenocarpic ‘AFR-12’, ‘V-185’ was clustered in the branch of the commercial cultivars (Formisano et al. 2012). These data indicate that at least the parthenocarpy of these two accessions has a different origin, providing evidence, therefore, of two different sources of parthenocarpy and genetic variability for current Zucchini breeding programs. The phenotypic analysis carried out in this work also showed the existence of different sources of parthenocarpy among selected squash cultivars. Thus, parthenocarpic cultivars can be easily separated according to their monoecy stability or andromonoecy index (Fig. 3). The parthenocarpic fruit growth potential of the hybrid ‘Cavili’ is known to be associated with a partial conversion of female into bisexual flowers and therefore with a conversion of monoecious into partially andromonoecius plants (Martínez et al. 2013). The results presented in Fig. 3 demonstrate that the parthenocarpy of the other two parthenocarpic hybrids, ‘Argo’ and ‘Partenon’, as well as that of accession ‘V-185’, is related to a high andromonoecious index, i.e. a high conversion of female into bisexual flowers. Therefore, the parthenocarpy of the commercial hybrids and ‘V-185’ appears to have the same origin and is associated with andromonoecy. In these cultivars, parthenocarpic fruit is developed from a bisexual flower, and their floral organs remain attached to the fruit even after harvesting. These fruits with attached flowers occur because of a delay in floral organ maturation and abscission, caused by a reduction in flower ethylene production (Peñaranda et al. 2007; Martínez et al. 2013). The attached flower is, however, an undesirable trait for Zucchini fruit exportation, since the senescence of floral organs, or the wound left after their manual removal, produce a rapid decay of the fruit during postharvest storage (Payán et al. 2006). Nevertheless, we have also identified accessions, such as ‘CpCAL112’ and ‘PI261610’, which are completely stable for monoecy and showed no conversion of female into bisexual flowers, and consequently produced no fruit with attached flowers (Fig. 3). These results indicate that not all sources of parthenocarpy in C. pepo are dependent on andromonoecy, and that some of the identified accessions constitute alternative sources of variability for this trait in Zucchini squash.
Relationship between parthenocarpy and ethylene production and sensitivity
The relationship between ethylene and fruit set has been studied in Arabidopsis (Carbonell-Bejerano et al. 2011) and Pisum sativum (Orzáez and Granell 1997) by analyzing the effects of ethylene inhibitors on fruit growth. Treatments extended the time of response to GAs and delayed ovarian senescence in both species, resulting in an increase in the final size of the fruit. The ethylene insensitive mutants of Arabidopsis also have a larger window of response to GA than wild type plants (Carbonell-Bejerano et al. 2011). Furthermore, massive expression analyses in tomato have shown a down-regulation of ethylene biosynthesis and signaling genes after fruit set via pollination/fertilization or GA treatments (Vriezen et al. 2008; Pascual et al. 2009). Pascual et al. (2009) also observed a differential expression pattern of ethylene genes in pat3/pat4 mutants of tomato, suggesting that ethylene regulates carpel development and parthenocarpic fruit set. In C. pepo, we have recently demonstrated that ethylene is directly involved in fruit set, and that the initiation of fruit growth in this species requires low levels of ethylene production and signaling in the fruit during the days immediately after anthesis (Martínez et al. 2013). This reduced ethylene production is associated with partial andromonoecy and parthenocarpy in the unpollinated fruits of the hybrid ‘Cavili’, but can also be maintained via pollination or auxin treatments in non-parthenocarpic cultivars (Martínez et al. 2013).
In the unpollinated fruits of parthenocarpic cultivars other than ‘Cavili’, including the commercial hybrids ‘Argo’ and ‘Partenon’, and the accessions ‘CpCAL112’, ‘V-185’ and ‘PI261610’, ethylene is not induced in the days after anthesis (Fig. 5). By contrast, in the unpollinated fruits of the non-parthenocarpic cultivars, including ‘CpCAL110’, ‘PI169462’, ‘CpCA097’, ‘AFR-12’, ‘CpCAL003’ and ‘CpCAL044’, the abortion of fruit growth is coupled with a peak of ethylene production at 3 DPA. Moreover, there is a negative correlation between ethylene production in fruits at 3 DPA and fruit growth rate (R = −0.799, p value = 0.0018) (Fig. 5). In this sense the parthenocarpic hybrid ‘Cavili’ is not exceptional, because it is likely that the parthenocarpic development of the fruit in this species is always accompanied by a reduction of ethylene during the days immediately after anthesis, while the increase of ethylene would be a signal that triggers fruit abortion and senescence in the absence of natural parthenocarpy or pollination. Therefore, the measurement of ethylene production in the unpollinated fruits of summer squash at 3 DPA could be a suitable marker for the identification and selection of parthenocarpic materials in this species. This may be of special interest in segregating populations, where the number of fruits per plant may not be enough to determine parthenocarpic potential on the basis of fruit size.
The level of endogenous ethylene has been also found to be associated with sex phenotypes in cucurbit species (Byers et al. 1972; Rudich 1990; Owens et al. 1980; Yamasaki et al. 2001; Saito et al. 2007; Martínez et al. 2008). Gynoecious lines of melon and cucumber produce more ethylene than monoecious ones, and the level of ethylene production in monoecious cucumber lines increases upon transition from the male to the female phase of development in both apical shoots and in leaves. Moreover, ethylene production and sensitivity in seedlings was found to be correlated with sex expression in C. pepo (Manzano et al. 2011). These data suggest that the production of ethylene in vegetative organs could be used as a marker for sex expression in cucurbits. We have analyzed the possible correlation between parthenocarpy and early measurements of ethylene production and sensitivity in vegetative organs and in male flowers. We found that ethylene production in young leaves at early stages of plant development was not correlated with parthenocarpic fruit growth rate in the different accessions (Fig. 6). Ethylene sensitivity, measured as the ethylene response to male flower abscission, was significantly correlated with parthenocarpy, with most parthenocarpic accessions being less sensitive to ethylene. Nevertheless, one parthenocarpic accession (‘PI261610’) showed the same sensitivity as non-parthenocarpic ones (Fig. 7), and one non-parthenocarpic accession (‘AFR-12’) showed the same ethylene sensitivity as parthenocarpic ones. Neither ethylene production in leaves nor ethylene sensitivity in male flowers are unambiguous parameters to determine the parthenocarpic potential of a Zucchini cultivar.
References
Byers R, Baker L, Dilley D, Sell H (1972) Chemical induction of perfect flowers on a gynoecious line of muskmelon, Cucumis melo L. HortSci 9:321–331
Carbonell-Bejerano P, Urbez C, Granell A, Carbonell J, Perez-Amador MA (2011) Ethylene is involved in pistil fate by modulating the onset of ovule senescence and the GA-mediated fruit set in Arabidopsis. BMC Plant Biol 11:84
de Jong M, Mariani C, Vriezen WH (2009) The role of auxin and gibberellin in tomato fruit set. J Exp Bot 60:1523–1532
de Menezes CB, Maluf WR, De Azevedo SM, Faria MV, Nascimento IR, Nogueira DW, Gomes LAA, Bearzoti E (2005) Inheritance of parthenocarpy in summer squash (Cucurbita pepo L.). Genet Mol Res 4:39–46
de Ponti OMB, Garretsen F (1976) Inheritance of parthenocarpy in pickling cucumbers (Cucumis sativus L.) and linkage with other characters. Euphytica 25:633–642
Decker DS (1988) Origin(s), evolution, and systematics of Cucurbita pepo (Cucurbitaceae). Econ Bot 42:4–15
den Nijs APM, Balder J (1983) Growth of parthenocarpic and seed-bearing fruits of zucchini squash. Cucurbit Genet Coop Rep 6:84–85
den Nijs APM, van Zanten N (1982) Parthenocarpic fruit set in glasshouse grown zucchini squash. Cucurbit Genet Coop Rep 5:44–45
Durham G (1925) Has parthenogenesis been confused with hermaphroditism in Cucurbita? Am Nat 59:283–294
Ferriol M, Picó B, Nuez F (2003) Genetic diversity of a germplasm collection of Cucurbita pepo using SRAP and AFLP markers. Theor Appl Genet 107:271–282
Formisano G, Roig C, Esteras C, Ercolano MR, Nuez F, Monforte AJ, Picó MB (2012) Genetic diversity of Spanish Cucurbita pepo landraces: an unexploited resource for summer squash breeding. Genet Resour Crop Evol 59:1169–1184
Globerson D (1971) Effects of pollination on set and growth of summer squash (Cucumis pepo) in Israel. Expt Agr 7:183–188
Gómez P, Peñaranda A, Garrido D, Jamilena M (2004) Evaluation of flower abscission and sex expression in different cultivars of zucchini squash (Cucurbita pepo). In: Lebeda A, Paris H (eds) Progress in Cucurbit genetics and breeding research. Eucarpia-Cucurbitaceae 2004. Palacký University, Olomouc, pp 347–352
Jobst J, King K, Hemleben V (1998) Molecular evolution of the internal transcribed spacers (ITS1 and ITS2) and phylogenetic relationships among species of the family Cucurbitaceae. Mol Phylogenet Evol 9:204–219
Katzir N, Tadmor Y, Tzuri G, Leshzeshen E, Mozes-Daube N, Danin-Poleg Y, Paris HS (2000) Further ISSR and preliminary SSR analysis of relationships among accessions of Cucurbita pepo. Acta Hortic 510:433–439
Loy JB (2012) Breeding squash and pumpkins. In: Wang Y, Behera T, Kole C (eds) Genetics, genomics and breeding of cucurbits. Hew Hampshire, Science Publisher, Enfield, pp 93–139
Manzano S, Martínez C, Domínguez V, Avalos E, Garrido D, Gómez P, Jamilena M (2010) A major gene conferring reduced ethylene sensitivity and maleness in Cucurbita pepo. J Plant Growth Regul 29:73–80
Manzano S, Martínez C, Megías Z, Gómez P, Garrido D, Jamilena M (2011) The role of ethylene and brassinosteroids in the control of sex expression and flower development in Cucurbita pepo. Plant Growth Regul 65:213–221
Manzano S, Martínez C, Megías Z, Garrido D, Jamilena M (2013) Involvement of ethylene biosynthesis and signalling in the transition from male to female flowering in the monoecious Cucurbita pepo. J Plant Growth Regul 1–10
Martínez C, Manzano S, Kraaman P, Jamilena M (2008) Producción de etileno: un marcador temprano para seleccionar ginoecia en melón. Actas Hortic 51:197–198
Martínez C, Manzano S, Megías Z, Garrido D, Picó B, Jamilena M (2013) Involvement of ethylene biosynthesis and signalling in fruit set and early fruit development in zucchini squash (Cucurbita pepo L.). BMC Plant Biol 13:139
Martínez C, Manzano S, Megías Z, Barrera A, Boualem A, Garrido D, Bendahmane A, Jamilena M (2014) Molecular and functional characterization of CpACS27A gene reveals its involvement in monoecy instability and other associated traits in squash (Cucurbita pepo L.). Planta 1–15
Nee M (1990) The domestication of Cucurbita (Cucurbitaceae). Econ Bot 44:56–68
Nepi M, Pacini E (1993) Pollination, pollen viability and pistil receptivity in Cucurbita pepo. Ann Bot 72:527–536
Nitsch J, Kurtz E, Liverman J, Went F (1952) The development of sex expression in cucurbit flowers. Am J Bot 39:32–43
Om Y, Hong K (1989) Evaluation of parthenocarpic fruit set in zucchini squash. Res Rpt Rural Dev Adm (Suweon) 31:30–33
Orzáez D, Granell A (1997) DNA fragmentation is regulated by ethylene during carpel senescence in Pisum sativum. Plant J 11:137–144
Owens K, Peterson C, Tolla G (1980) Production of hermaphrodite flowers on gynoecious muskmelon by silver nitrate and aminoethoxyvinylglycine. HortSci 15:654–655
Ozga JA, Reinecke DM (2003) Hormonal interactions in fruit development. J Plant Growth Regul 22:73–81
Paris HS (1986) A proposed subspecific classification for Cucurbita pepo. Phytologia 61:133–138
Paris HS (2001) History of the cultivar-groups of Cucurbita pepo. In: Janick J (ed) Horticultural reviews, vol 25. Wiley, New York, pp 71–170
Pascual L, Blanca JM, Cãizares J, Nuez F (2009) Transcriptomic analysis of tomato carpel development reveals alterations in ethylene and gibberellin synthesis during pat3/pat4 parthenocarpic fruit set. BMC Plant Biol 9:67
Payán M, Peñaranda A, Rosales R, Garrido D, Gómez P, Jamilena M (2006) Ethylene mediates the induction of fruits with attached flower in Zucchini squash. In: Holmes GJ (ed) Proceedings of Cucurbitaceae 2006. Universal Press, Raleigh, pp 171–179
Peñaranda A, Payan MC, Garrido D, Gómez P, Jamilena M (2007) Production of fruits with attached flowers in zucchini squash is correlated with the arrest of maturation of female flowers. J Hortic Sci Biotechnol 82:579–584
Robinson RW (1993) Genetic parthenocarpy in Cucurbita pepo L. Cucurbit Genet Coop Rep 16:55–57
Robinson RW, Reiners S (1999) Parthenocarpy in summer squash. HortSci 34:715–717
Rudich J (1990) Biochemical aspects of hormonal regulation of sex expression in cucurbits. In: Bates DM, Robinson RW, Jeffrey C (eds) Biology and utilization of the Cucurbitaceae. Cornell University Press, Ithaca, pp 269–280
Rylski I (1974) Effects of season on parthenocarpic and fertilized summer squash (Cucumis pepo L.). Expt Agr 10:39–44
Rylski I, Aloni B (1990) Parthenocarpic fruit set and development in Cucurbitaceae and Solanaceae under protected cultivation in a mild winter climate. Acta Hortic 287:117–126
Saito S, Fujii N, Miyazawa Y, Yamasaki S, Matsuura S, Mizusawa H, Fujita Y, Takahashi H (2007) Correlation between development of female flower buds and expression of the CS-ACS2 gene in cucumber plants. J Exp Bot 58:2897–2907
Sanz M (1995) Fitorreguladores para el calabacín. Hortofruticultura 33:46–48
Serrani JC, Carrera E, Ruiz-Rivero O, Gallego-Giraldo L, Peres LEP, García-Martínez JL (2010) Inhibition of auxin transport from the ovary or from the apical shoot induces parthenocarpic fruit-set in tomato mediated by gibberellins. Plant Physiol 153:851–862
Srivastava A, Handa A (2005) Hormonal regulation of fruit development: a molecular perspective. J Plant Growth Regul 24:67–82
Vriezen WH, Feron R, Maretto F, Keijman J, Mariani C (2008) Changes in tomato ovary transcriptome demonstrate complex hormonal regulation of fruit set. New Phytol 177:60–76
Wien HC (2002) The cucurbits: cucumber, melon, squash and pumpkin. In: Wien HC (ed) The physiology of vegetable crops. CABI, New York, pp 345–386
Yamasaki S, Fujii N, Matsuura S, Mizusawa H, Takahashi H (2001) The M locus and ethylene-controlled sex determination in andromonoecious cucumber plants. Plant Cell Physiol 42:608–619
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Martínez, C., Manzano, S., Megías, Z. et al. Sources of parthenocarpy for Zucchini breeding: relationship with ethylene production and sensitivity. Euphytica 200, 349–362 (2014). https://doi.org/10.1007/s10681-014-1155-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10681-014-1155-8