
Abstract
In this paper we compare the sensitivity of different squash genotypes to ethylene and brassinosteroids by studying the effects of different ethylene and brassinosteroid treatments on the sexual expression and flower development of different C. pepo genotypes: Bolognese (Bog) and Vegetable Spaghetti (Veg), two contrasting lines for ethylene production and sensitivity, as well as Cora, a standard commercial hybrid. Results have demonstrated that ethylene has a much greater effect on sexual expression and flower development in C. pepo than brassinosteroids. Ethephon increases the number of female flowers per plant and reduces the first male phase of development, while treatments with the ethylene inhibitors AVG and STS reduce the number of female flowers per plant and expand the first male phase of development. The differential response observed between genotypes appears to be related to ethylene production and sensitivity. Bog, which produces more ethylene and is more sensitive to this hormone, responded much better to AVG and STS, reducing the number of female flowers per plant, while Veg, which is characterised by lower production of and sensitivity to ethylene, responded better to ethephon by reducing the first male phase of development and increasing the number of female flowers per plant. The differential abortion of female and male flowers in ethephon, AVG and STS treatments, as well as the occurrence of bisexual flowers in the AVG and STS treated plants of the more ethylene sensitive genotypes, demonstrate that ethylene is also involved in the development of female flowers. Female flower buds require a minimal level of ethylene not only to complete their development and maturation without a premature abortion, but also to arrest the development of stamens in the third whorl and to promote the appropriate growth of the carpels. On the contrary, the role of brassinosteroids in the sexual expression of C. pepo was not so evident. The application of brassinazole, an inhibitor of brassinosteroid biosynthesis slightly changes the production of ethylene in the three analysed genotypes, but those changes have little effect on their sexual phenotypes, and they do not alter the development of the unisexual flowers.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Sex determination and flower development in monoecious and dioecious plants are under the control of different phytohormones, whose functional roles depend on the species (Grant et al. 1994). Different approaches, including external treatments as well as measurements of the internal level of each hormone in male and female flowers, have been used to elucidate the role of each hormone in both sex determination and flower development. In Mercurialis annua, which can be dioecious, monoecious or androdioecious, depending on local environmental conditions (Pannell 1997a, b), sexual expression is sensitive to the external application of auxins (Durand 1969; Ainsworth 2000), and the level of indol-3-acetic acid (IAA) is three to six times higher in male than in female flowers (Louis et al. 1990). Cytokinins, on the other hand, play a role in sex determination of dioecious species such as Vitis vinifera, Cannabis sativa and Spinacea oleracea (Chailakhyan 1979; Durand 1969; Dellaporta and Calderon-Urrea 1993; Grant et al. 1994). Giberellins (GAs) and abscisic acid (ABA) also regulate the sexual expression of certain species. In maize, GAs are required for the abortion of the stamen primordia in female flowers (Lebel-Hardenack and Grant 1997), but they promote the production of male flowers in Cucumis sativus, C. sativa, S. oleracea and A. officinalis (Ainsworth 2000). By contrast, ABA induces the development of female flowers in many monoecious and dioecious species, maybe by reducing the activity of GAs (Khryanin 2002). Finally, sexual expression of certain species depends on ethylene and brassinosteroids (Mcmurray and Miller 1968; Ram and Jaiswal 1970; Arteca et al. 1983; Vardhini and Rao 2002; Woeste et al. 1999).
In the Cucurbitaceae family sex expression is known to be controlled by different genetic, environmental and hormonal factors. In C. sativus and C. melo, sex is controlled by three major independent genes, the combination of which explains the main sex phenotypes found in these two species (Kubicki 1969; Kenigsbuch and Cohen 1990; Rudich 1990; Pierce and Wehner 1990; Perl-Treves 1999). Sex expression in this family of plants can be modified by environmental factors, including light intensity, photoperiod and temperature. Winter conditions, with short days, low light intensity and low night temperatures promote the production of female flowers, while summer conditions increase the production of male flowers (Wien 1997; Peñaranda et al. 2007). In C. pepo, low temperature inhibits the development of male flowers and increases the number of female flower per plant (Wien et al. 2004), while high temperature induces both a partial transformation of female flowers into hermaphrodites and a complete transformation of female flowers into male ones (Peñaranda et al. 2007). Phytohormones are the main modulators of sex expression in the Cucurbitaceae family. GAs, auxins, brassinosteroids and especially ethylene regulate sex expression of cucurbit species. GAs and auxins promote the production of male and female flowers, respectively (Rudich et al. 1972; Wien 1997; Trebitsh et al. 1987), although their functions appear to be mediated by ethylene. Undoubtedly, ethylene is the principal hormone regulating sex expression in the Cucurbitaceae family. The level of ethylene in flower buds seems to be essential for both sex determination and female flower development. Thus, ethylene treatments promote the production of female flowers, while treatments with inhibitors of ethylene biosynthesis or perception, such as aminoethoxyvinylglycine (AVG) or silver thiosulphate (STS), increase the number of male flowers per plant (Rudich et al. 1969; Byers et al. 1972; DenNijs and Visser 1980; Owens et al. 1980; Rudich 1990; Payán et al. 2006). Recently it has been demonstrated that the monoecious phenotype of melon and cucumber is controlled by a 1-AMINOCYCLOPROPANE-1-CARBOXYLATE SYNTHASE (ACS) gene that is specifically expressed in carpels and that is involved in the arrest of stamen development in female flowers. Andromonoecious varieties are indeed mutants for this gene, which results in an inhibition of the arrest of stamen development in the ovary-bearing flowers, leading to the formation of hermaphrodite flowers instead of female ones (Boualem et al. 2008; Boualem et al. 2009; Feng et al. 2009). Moreover, cucumber and melon gynoecious lines produce more ethylene than monoecious or andromonoecious ones (Owens et al. 1980; Yamasaki et al. 2001; Manzano et al. 2008). In fact, gynoecious cucumber varieties have an additional ACS gene (Trebitsh et al. 1997; Mibus and Tatlioglu 2004; Knopf and Trebitsh 2006). Nevertheless, the gynoecious phenotype of melon results from a transposon in the promoter of the transcription factor CmWIP1, a gene that controls carpel abortion and indirectly promotes the development of stamens by repressing the expression of the andromonoecious gene CmACS7 (Martin et al. 2009).
The role of brassinosteroids in the control of sex determination of cucurbit species is not clear. Papadopoulou and Grumet (2005) performed a number of external treatments with brassinolide in cucumber, melon and zucchini and concluded that cucumber was more sensitive than zucchini, by reducing the number of male flowers in the initial phase of development and promoting the initiation of the first female flower in the main shoot. Given that brassinolide treated plants of cucumber produced more ethylene than control plants, the authors concluded that the effect of brassinosteroids on sexual expression of cucumber appears to be mediated by ethylene (Papadopoulou and Grumet 2005).
We have previously characterised two inbred lines of C. pepo, Bolognese (Bog) and Vegetable spaghetti (Veg) that differ in their sexual expression. While Bog develops the first female flowers between nodes 2–4, and produces a mean of 69–75% female flowers, the sex phenotype of Veg was strongly male, delaying the development of the first female flower to nodes 10–20, and producing less than 25% female flowers, with some of the plants producing less than 5% (Manzano et al. 2010). This extreme male phenotype of Veg inbred line is determined by a single gene that confers reduced ethylene sensitivity (Manzano et al. 2010). Therefore the inbred line Veg appears to have a mutation that reduces ethylene sensitivity in the plants and consequently delays and reduces the production of female flowers. In order to study the differential sensitivity of different squash genotypes to ethylene and brassinosteroids for the control of sex expression and flower development in C. pepo, in the present paper we assess the effects of external application of different ethylene and brassinazole treatments on sexual expression and flower development in Bog and Veg inbred lines. The results demonstrate that the effects of ethylene and brassinazole treatments depend on the production and sensitivity of each genotype to ethylene.
Materials and methods
Plant material, growing conditions and evaluation of sexual expression
Two inbred lines of Cucurbita pepo subspecies pepo of the Vegetable Marrow Group, Vegetable Spaghetti (Veg) and Bolognese (Bog) have been analyzed. The Veg and Bog lines are the results of more than eight generations of selfing, and they were selected for their contrasting sensitivity to ethylene in C. pepo (Manzano et al. 2010). The hybrid variety Cora was used for comparison in all the experiments. Seeds were germinated on wet filter paper in Petri dishes for 3 days, and then transplanted individually to 20 cm wide pots containing peat, perlite and vermiculite in the proportion 2:1:1. Plants were grown in a climate controlled chamber at 25°C, 75% relative humidity and with a long photoperiod regime of 16 h light/8 h night. Watering and nutrition of the plants were carried out automatically following standard practices.
Sex expression in each plant was determined as both the number of initial nodes with male flowers before the production of the first female flower (earliness), and as the percentage of female flowers per plant in the first 20 nodes of the main stem (femaleness).
Hormonal treatments and evaluation of ethylene production
To study the implication of ethylene in sexual determination and development of male and female flowers of C. pepo, plants were treated with the ethylene releasing agent ethephon, with the ethylene biosynthesis inhibitor aminoethoxyvinylglycine (AVG) or with the ethylene perception inhibitor silver thiosulphate (STS). For ethephon treatments, the apical shoots of plants with 4 true leaves were sprayed with 0.1% tween 20 and 500 ppm of ethephon. Ethylene inhibiting treatments were with solutions containing 0.1% tween 20 and 1 mM of AVG or 0.1% tween 20 and 0.5 M of STS. Control plants were sprayed with only 0.1% tween 20. 10 plants were used for each treatment.
To evaluate the role of brassinosteroids, apical shoots of plants with 4-true leaves were sprayed with 10 μM of brassinazole, an inhibitor of brassinosteroid biosynthesis. The treatment was repeated three times a week for 4 weeks. Control plants were treated with water. The production of ethylene in the leaves of control and brassinosteroid treated plants was measured as described in Manzano et al. (2010).
Statistical differences between lines in each hormonal treatments (control plants and treated plants) were determined by analysis of variance (ANOVA), followed by Tuckey’s multiple comparison test using the STATISTIX 8.0 software package.
Results
Effects of ethephon, AVG and STS on sexual expression of C. pepo
Three genotypes of C. pepo were selected on the basis of their differential sexual expression. Two of them, Veg and Bog, correspond to two inbred lines with a very different sex phenotype. The third one, Cora, is a standard hybrid variety for zucchini production in the South of Spain. Under controlled environmental conditions in culture chambers, the three genotypes have a very different sexual expression (Fig. 1). While Bog developed the first female flowers between nodes 3–4 and produced a mean of 69–75% female flowers, in the Veg genotype the development of the first female flower was delayed to node 20 and fewer than 5% of flowers are female. Cora developed the first female flowers between nodes 6–7 and its sexual expression was intermediate between Bog and Veg.

Effects of ethephon, AVG and STS on the sexual expression of three genotypes of C. pepo. At each node, black, white and red bars represent the percentages of male, female and bisexual flowers of the plants analysed (n ≥ 10 for each genotype). At each node, grey bars represent the percentage of aborted flowers
To determine the effects of ethylene on the sexual expression of Veg, Bog and Cora we have compared two parameters in control and treated plants: the number of female flowers per plant, and the number of initial male flowers until the appearence of the first female flowers. Ethephon treatments had a feminizing effect, but altered differentially the sexual expression of the different genotypes (Fig. 1). Ethephon significantly increased the average number of female flowers per plant in Cora and Veg, but no significant difference was detected between treated and control plants in Bog (Table 1). In the Veg line ethephon also significantly promoted the initiation of female flower production from node 20 to node 7, i.e. there were fewer male flowers on the main stem until the development of the first female flower (Table 1).
External application of the ethylene biosynthesis inhibitor aminoethoxyvinylglycine (AVG) and the ethylene perception inhibitor silver thiosulphate (STS) produced a masculinizing effect on all treated plants of the three genotypes (Table 1). In relation to the number of female flowers per plant, both AVG and STS induced a reduction of this parameter, although the decrease was more pronounced in AVG-treated plants than in STS-treated ones. AVG significantly reduced the number of female flowers per plant in Cora and Bog, but no significant difference was found for Veg (Table 1). STS significantly decreased the number of female flowers per plant in Bog, but significant differences were not detected in the other two genotypes (Table 1). Regarding the extension of the first male phase of development, AVG significantly increased the average number of nodes to the first female flower in Cora, delaying female flowering. However, no significant differences for this trait were observed among treated and control plants in Bog and Veg (Table 1). STS treatment did not alter earliness of female flowering in any of the studied genotypes.
Effects of ethephon, AVG and STS on flower development of C. pepo
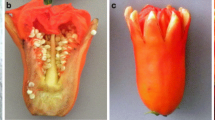
In addition to complete conversions between flower sex phenotypes, AVG and STS also promoted partial transformations of female flowers into male ones, resulting in the formation of bisexual flowers (Fig. 1). Besides developing ovary, style and stigma, these flowers were characterized by their long peduncles and a full or partial development of stamens, two distinctive characteristics of male flowers (Fig. 2). The formation of bisexual flowers did not affect the different genotypes equally. In the Veg line bisexual flowers were not detected, maybe because the number of female flowers produced by this genotype was very low. In Cora, bisexual flowers were detected in AVG-treated plants but not in STS- treated ones, and they were characterised by partial development of stamens (Fig. 1). The Bog line was the most sensitive one, developing bisexual flowers (in plants) under both treatments (Fig. 1). Some of these Bog bisexual flowers, like those of Cora, showed partial development of stamens and a normal development of style and stigma, but some of them developed full stamens but no style or stigma (Fig. 2). Ovary development was not affected, although its thickness decreased considerably compared to a normal female flower (Fig. 2).

Bisexual flowers from Bog plants treated with AVG (b–c) y STS (e–g). a Detail of a normal female flower. b Detail of a bisexual flower with rudimentary stamens in the third whorl and normal development of carpels. c Detail of a bisexual flower with fully developed stamens and complete absence of style and stigma. d Development of ovary style and stigma in a normal female flower. (e, f and g) Different levels of development of stamens in the third whorl of ovary-bearing flowers, and variations in the morphology of the ovary of female flowers from plants treated with STS
Effects of brassinosteroids on sexual expression and flower development of C. pepo
The influence of brassinosteroids on sexual expression and flower development of C. pepo was studied by determining the effects of external applications of brassinazole (Brz), an inhibitor of brassinosteroid biosynthesis, on the sexual expression of Veg, Bog and Cora. Table 2 shows the effects of Brz on both the earliness and production of female flowers. The application of Brz produced no alteration of the number of female flowers per plant in the three genotypes, but it significantly increased the number of male nodes until the occurrence of the first female flower in Cora, though not in either Bog or Veg.
The production of ethylene in leaves of control and Brz-treated plants agreed with the sexual expression effects in the three genotypes (Fig. 3). Under controlled environmental conditions in culture chambers, the leaves of Cora and Bog genotypes produced more ethylene than the Veg line, and the Bog line showed the highest production (of ethylene). The application of Brz significantly reduced the production of ethylene in Cora and Bog, but the same treatment was able to increase significantly the production of ethylene in the Veg line, reaching a level of ethylene similar to that observed in control plants of Cora and Bog. No alteration was detected in the development of Brz-treated female or male flowers of the three genotypes studied.

Effects of the brassinosteroid inhibitor Brassinazole on ethylene production in leaves of three genotypes of C. pepo. Values are the mean (± standard error) of 8 repetitions expressed in nl/g FW after 24 h incubation
Discussion
Implication of ethylene production and sensitivity on the sexual expression of C. pepo
Ethylene has been described as the principal regulator of sexual expression in different cultivated species of the Cucurbitaceae family, including cucumber, melon, watermelon and zucchini (Rudich 1990; Tongjia and Quinn 1995; Yamasaki et al. 2005). This small gaseous hormone is able to control the duration of the sexual phases of development in these species, as well as the proportion of male and female flowers per plant. In zucchini, previous studies had demonstrated that external treatments with agents that stimulate or inhibit ethylene production or perception are able to modify the production of male and female flowers per plant in different varieties (Wien 2006; Payán et al. 2006). Our results of ethylene treatments on the genotypes Cora, Bog and Veg indicate that ethylene promotes earliness and the production of female flowers, while ethylene inhibitors such as AVG and STS delay and reduce the production of female flowers per plant. The effects are similar to those observed in other cucurbit species such as cucumber and melon (Takahashi and Jaffe 1984; Takahashi and Suge 1982; Atsmon and Tabbak 1979; Byers et al. 1972; Owens et al. 1980; Kamachi et al. 1997; Trebitsh et al. 1997).
The results also indicate variation in sensitivity to ethylene among the analysed genotypes. The hybrid variety Cora responded to ethephon, AVG and STS treatments in a very similar way to other commercial varieties of Zucchini (Payán et al. 2006). Nevertheless, the contrasting lines Bog and Veg responded in a differential way to the same treatments. Ethephon induced earlier and higher production of female flowers in the Veg line, but did not significantly alter the sexual expression of Bog. On the contrary, the ethylene inhibitors AVG and STS significantly delayed and diminished the production of female flowers in Bog, but they did not affect the sexual expression of Veg. These results indicate that Veg and Bog genotypes may differ in their production of ethylene, not only in leaves, as we report in this paper, but also in reproductive organs (non-published data). The Bog line, with higher production of ethylene, would respond to a decrease in the level of the hormone (AVG), while the Veg line, with lower ethylene production, would be more affected by the increase of ethylene caused by ethephon. The abortion of female flower development induced by ethephon in the Veg line, but not in Bog and Cora, may also be related to low production of this hormone in the female flowers of Veg.
Although ethephon increases the production of female flowers per plant in Veg and Cora, these genotypes never reached the level of production of female flowers in Bog. Moreover, AVG treatments significantly diminished the production of female flowers in Bog, but never to the levels observed in Veg or Cora. These results indicate that the differential sexual expression of Bog and Veg lines cannot be based merely on differences in the production of ethylene, but must also be caused by differences in sensitivity to this hormone, as observed previously by Manzano et al. (2010). Higher sensitivity to ethylene in the Bog line would promote a limited physiological response in the production of female flowers which cannot be overcome by external ethylene applications. Similarly, limited sensitivity to ethylene in the Veg line makes the production of male flowers in this line very high, a feature that can not be changed by decreasing the level of ethylene. Therefore, though it seems that Veg responds much more to ethephon than Bog, the relative effects on the sexual expression in both lines indicate that sensitivity to ethylene in Veg is less than in Bog and Cora. Moreover, the fact that untreated Bog plants produce more female flowers than ethylene-treated Cora plants indicates that the Bog line is also more sensitive to ethylene than Cora. The response of Bog to STS treatments (this line was the only one that responded to STS) also support the conclusion that Bog is the most sensitive of the three analysed genotypes to ethylene.
The role of ethylene during floral development of C. pepo
Besides controlling the duration of the two sexual phases of development and the number of female and male flowers per plant, the treatments with ethephon, AVG and STS have demonstrated that ethylene also regulates the individual development of floral primordia in squash. Thus, in those flower buds that are already determined as male or female, treatments with AVG or STS do not induce a complete transformation of female flowers into male ones, as is observed in the youngest flower buds, but rather a partial transformation that results in bisexual flowers with different degrees of stamen development. These bisexual flowers may be produced as a consequence of a decrease in ethylene in already determined female flower buds, as was proposed by Yamasaki et al. (2003) in cucumber. If so, our results would indicate that ethylene is not only necessary to determine the sexual identity of a certain floral meristem, but it is also required to maintain the arrest of stamen development and therefore the sexual identity of a flower throughout female flower development. The maintenance of stamen arrest in the female flowers would need, therefore, a minimal level of ethylene as well as suitable activity of the ethylene receptors. Therefore, the inhibition of ethylene biosynthesis (treatment with AVG) or perception (treatment with STS) in the early stages of female flower development would result in the abnormal development of stamen in female flowers. In melon and cucumber, it has recently been demonstrated that the arrest of the stamen in female flowers of monoecious varieties is controlled by CmACS-7 and CsACS2, respectively, two orthologous genes that are expressed specifically in the pistils of female flowers of monoecious varieties, and which have a loss of function mutation in the andromonoecious varieties (Boualem et al. 2008, 2009). Therefore, the pistil seems to be the floral organ that produces ethylene in female flowers, whereas the perception of ethylene seems to reside in the stamens (Little et al. 2007). In zucchini, our treatments with AVG and STS demonstrate that a decrease in both production (probably in the pistils) and perception of ethylene in female flowers (probably in the stamen primordia) favour the development of stamen in the female flowers, indicating that the control of stamen development arrest in this species is also dependent on the production of and response to ethylene.
Previous results had indicated that the development of the stamen in C. pepo female flowers was not linked to the development of the carpels (Payán et al. 2006). The results obtained in the Bog line, where it has been possible to analyze a higher number of bisexual flowers, demonstrate that these flowers are not only altered in the development of the stamens, but also showed different degrees of style and stigma development. Some Bog bisexual flowers were found to show poor development of stamens but complete development of style and stigma, while others showed complete development of stamens accompanied by an absolute arrest of style and stigma development. These results support the results of Papadopoulou et al. (2005) and Little et al. (2007) indicating that ethylene production and perceptions are also necessary for the appropriate development of carpels in the female flowers. The flowers of transgenic melon plants with the mutation etr1-1 of Arabidopsis under the specific promoter of petals and stamen AP3 show very poor development of the pistils, which indicates that the ethylene perceived by stamens or petals promotes the development of the pistils in the fourth whorl of the flowers (Little et al. 2007). The fact that this effect on C. pepo female flower development has been detected only in Bog might be related to the higher production of and/or sensitivity to ethylene of the female flowers of this line.
Finally, it is also necessary to emphasize that our results demonstrate that ethylene regulates the growth of both female and male flowers of zucchini. The normal development of female flowers in this species needs a higher level of ethylene than the development of male flowers. In fact, the treatments with ethephon significantly increased the number of aborted male flowers in the three genotypes, whereas the treatments with AVG favoured the abortion of female flowers. In Veg, the treatments with ethephon also increased the number of female flower abortions, which would indicate that this line is less sensitive to ethylene (Manzano et al. 2010), and though the external ethylene is capable of inducing the formation of a female flower, it does not produce the necessary response for correct development and maturation.
The role of brassinosteroids on the sexual expression and floral development of C. pepo
Our results indicate that, in comparison with ethylene, brassinosteroids play a minor role in the control of sexual expression in C. pepo. The inhibition of brassinosteroids biosynthesis by Brz significantly delayed the production of female flowers only in Cora, but it did not alter this parameter in the other two genotypes. Moreover, the brassinosteroid inhibitor did not affect the number of female flowers per plant or the development of male and female flowers. These results suggest that brassinosteroids may regulate the induction of the female flower phase of development in C. pepo, although this regulation is genotype dependent. By using different species of the Cucurbitaceae family, Papadopoulou and Grumet (2005) observed that the application of epi-brassinolide promotes earlier and higher production of female flowers, although the effect was much higher in cucumber than in zucchini. This response could be mediated by ethylene since it is known that brassinosteroids induce the production of ethylene in different plant species (Arteca et al. 1983; Yi et al. 1999; Woeste et al. 1999; Vardhini and Rao 2002; Arteca and Arteca 2008), including cucurbits (Papadopoulou and Grumet 2005). In this paper we have observed that the ethylene induced by brassinosteroids appears to be dependent on ethylene response, since we have observed that Brz applications reduced the production of ethylene in leaves of Bog and Cora (as is expected if brassinosteroids induce the production of ethylene), but increase the production of ethylene in the ethylene insensitive line Veg. The differential effect of brassinosteroids on the sexual expression of the different genotypes of C. pepo could therefore depend on the sensitivity to ethylene of the different genotypes, as proposed by Papadopoulou and Grumet (2005). Thus, the partial insensitivity of Veg to ethylene (Manzano et al. 2010) makes this line unresponsive to Brz treatment, even though ethylene production was increased by the Brz treatment to the normal levels in control plants of the other two genotypes. In the other two genotypes, Bog and Cora, the reduction in ethylene production induced by the Brz treatment was accompanied by a reduction in the number of female flowers per plant as well as an increase in the number of male flowers during the first phase of development, although these differences were only significant in Bog. The development of male or female flowers of C. pepo was not altered by Brz treatment, suggesting that although brassinosteroids may regulate the production of ethylene and have minor effects on the sexual expression of C. pepo, they do not appear to be involved in the development of female or male flowers.
References
Ainsworth C (2000) Boys and girls come out to play: the molecular biology of dioecious plants. Ann Bot 86:211–221
Arteca RN, Arteca JM (2008) Effects of brassinosteroid, auxin, and cytokinin on ethylene production in Arabidopsis thaliana plants. J Exp Bot 59:3019–3026
Arteca RN, Tsai DS, Schlagnhaufer C, Mandava NB (1983) The effect of brassinosteroid on auxin-induced ethylene production by etiolated mung bean segments. Physiol Plantarum 59:539–544
Atsmon D, Tabbak C (1979) Comparative effects of gibberellin silver nitrate and aminoethoxyvinylglycine on sexual tendency and ethylene evolution in the cucumber plant (Cucumis sativus L.). Plant Cell Physiol 20:1547–1555
Boualem A, Fergany M, Fernandez R, Troadec C, Martin A, Morin H, Sari M, Collin F, Flowers JM, Pitrat M, Purugganan MD, Dogimont C, Bendahmane A (2008) A conserved mutation in an ethylene biosynthesis enzyme leads to andromonoecy in melons. Science 321:836–838
Boualem A, Troadec C, Kovalski I, Sari M, Perl-Treves R, Bendahmane A (2009) A conserved ethylene biosynthesis enzyme leads to andromonoecy in two Cucumis species. PLoS ONE 4:1–10
Byers RE, Baker LR, Sell HM, Herner RC, Dilley DR (1972) Ethylene: a natural regulator of sex expression in Cucumis melo L. Proc Natl Acad Sci USA 69:712–720
Chailakhyan MK (1979) Genetic and hormonal regulation of growth, flowering, and sex expression in plants. Am J Bot 66:717–736
Dellaporta SL, Calderon-Urrea A (1993) Sex determination in flowering plants. Plant Cell 5:1241–1251
Den Nijs APM, Visser DL (1980) Induction of male flowering in gynoecious cucumbers (Cucumis sativus L.) by silver ions. Euphytica 29:273–280
Durand B (1969) Selection de genotypes males de Mercurialis annua L. (2n = 16) en fonction de leur sensibilite aux cytokinins. CR Acad Sci Paris Ser D 268:2049–2051
Feng H, Li X, Liu Z, Wei P, Ji R (2009) A co-dominant molecular marker linked to the monoecious gene CmACS-7 derived from gene sequence in Cucumis melo L. African J Biotech 8:3168–3174
Grant S, Houben A, Vyskot B, Siroky J, Pan W, Macas J, Saedler H (1994) Genetics of sex determination in flowering plants. Dev Genet 15:214–230
Kamachi S, Sekimoto H, Kondo N, Sakai S (1997) Cloning of a cDNA for a 1-Aminocyclopropane-1-Carboxylate synthase that is expressed during development of female flowers at the apices of Cucumis sativus L. Plant Cell Physiol 38:1197–1206
Kenigsbuch D, Cohen Y (1990) The inheritance of gynoecy in muskmelon. Genome 33:317–320
Khryanin VN (2002) Role of phytohormones in sex differentiation in plants. Russian J Plant Physiol 49:545–551
Knopf RR, Trebitsh T (2006) The female-specific Cs-ACS1G gene of cucumber. A case of gene duplication and recombination between the non-sex-specific 1-aminocyclopropane-1-carboxylate synthase gene and a branched-chain amino acid transaminase gene. Plant Cell Physiol 47:1217–1228
Kubicki B (1969) Investigation of sex determination in cucumber (Cucumis sativus L.). Genet Pol 10:101–121
Lebel-Hardenack S, Grant SR (1997) Genetics of sex determination in flowering plants. Trends Plant Sci 2:130–137
Little HA, Papadopoulou E, Hammar SA, Grumet R (2007) The influence of ethylene perception on sex expression in melon (Cucumis melo L.) as assessed by expression of the mutant ethylene receptor, At-etr1–1, under the control of constitutive and floral targeted promoters. Sex Plant Reprod 20:123–136
Louis J, Augur C, Teller G (1990) Cytokinins and differentiation processes in Mercurialis annua: Genetic regulation, relations with auxins, indoleacetic acid oxidases, and sexual expression patterns. Plant Physiol 94:1535–1541
Manzano S, Martínez C, Kraakman P, Jamilena M (2008) Ethylene Use of ethylene production as a marker for the selection of gynoecy in melon (Cucumis melo). In: Pitrat M (ed) IXth EUCARPIA Meeting on Genetics and Breeding of Cucurbitaceae. INRA, Avignon, pp 557–561
Manzano S, Martínez C, Domínguez V, Avalos E, Garrido D, Gómez P, Jamilena M (2010) A major gene conferring reduced ethylene sensitivity and maleness in Cucurbita pepo. J Plant Growth Regul 29:73–80
Martin A, Troadec C, Boualem A, Rajab M, Fernandez R, Morin H, Pitrat M, Dogimont C, Bendahmane A (2009) A transposon-induced epigenetic change leads to sex determination in melon. Nature 461:1135–1138
Mcmurray AL, Miller CH (1968) Cucumber sex expression modified by 2-chloroethanephosphonic acid. Science 162:1397–1398
Mibus H, Tatlioglu T (2004) Molecular characterization and isolation of the F/f gene for femaleness in cucumber (Cucumis sativus L.). Theor Appl Genet 109:1669–1676
Owens KW, Peterson CE, Tolla GE (1980) Prodution of hermaphrodite flowers on gynoecious muskmelon by silver nitrate and aminoethyoxyvinylglycine. HortScience 15:654–655
Pannell J (1997a) Mixed genetic and environmental sex determination in an androdioecious population of Mercurialis annua. Heredity 78:50–56
Pannell J (1997b) Widespread functional androdioecy in Mercurialis annua L. (Euphorbiaceae). Biol J Linn Soc 61:95–116
Papadopoulou E, Grumet R (2005) Brassinosteriod-induced femaleness in cucumber and relationship to ethylene production. HortScience 40:1763–1767
Papadopoulou E, Little HA, Hammar SA, Grumet R (2005) Effect of modified endogenous ethylene production on sex expression bisexual flower development and fruit production in melon (Cucumis melo L.). Sex Plant Reprod 18:131–142
Payán MC, Peñaranda A, Rosales R, Garrido D, Gomez P, Jamilena M, Holmes GJ (2006) Ethylene mediates the induction of fruits with attached flower in zucchini squash. In: Holmes GJ (ed) Proceedings Cucurbitaceae 2006. Universal Press, Raleigh, pp 171–179
Peñaranda A, Payán MC, Garrido D, Gómez P, Jamilena M (2007) Production of fruits with attached flowers in zucchini squash is correlated with the arrest of maturation of female flowers. J Hort Sci Biotechnol 82:579–584
Perl-Treves R (1999) Male to female conversion along the cucumber shoot: Approaches to studying sex genes and floral development in Cucumis sativus. In: Ainsworth CC (ed) Sex determination in plants. BIOS Scientific, Oxford, pp 189–215
Pierce LK, Wehner TC (1990) Review of genes and linkage groups in cucumber. HortScience 25:605–615
Ram HYM, Jaiswal VS (1970) Induction of female flowers on male plants of Cannabis sativa L. by 2-chloroethanephos-phonic acid. Experientia 26:214–216
Rudich J (1990) Biochemical aspects of hormonal regulation of sex expression in Cucurbits. In: Bates DM, Robinson RW, Jeffry C (eds) Biology and Utilization of the Cucurbitaceae. Cornell University Press, New York, pp 288–304
Rudich J, Halevy AH, Kedar N (1969) Increase in femaleness of three cucurbits by treatment with ethrel, an ethylene releasing compound. Planta 86:69–76
Rudich J, Halevy AH, Kedar N (1972) Ethylene evolution from cucumber plants as related to sex expression. Plant Physiol 49:998–999
Takahashi H, Jaffe MJ (1984) Further studies of auxin and ACC induced feminization in the cucumber plant using ethylene inhibitors. Phyton 44:81–86
Takahashi H, Suge H (1982) Sex expression and ethylene production in cucumber plants as affected by 1-aminocyclopropane-1-carboxylic acid. J Japan Soc Hor Sci 51:51–55
Tongjia Y, Quinn JA (1995) Tests of a mechanistic model of one hormone regulating both sexes in Cucumis sativus (Cucurbitaceae). Am J Bot 82:1537–1546
Trebitsh T, Rudich J, Riov J (1987) Auxin, biosynthesis of ethylene and sex expression in cucumber (Cucumis sativus). Plant Growth Regul 5:105–113
Trebitsh T, Staub JE, O’Neill SD (1997) Identification of a 1-aminocyclopropane-1-carboxylic acid synthase gene linked to the Female (F) locus that enhances female sex expression in cucumber. Plant Physiol 113:987–995
Vardhini BV, Rao SSR (2002) Acceleration of ripening of tomato pericarp discs by brassinosteroids. Phytochemistry 61:843–847
Wien HC (1997) The cucurbits: Cucumber, melon, squash and pumpkin. In: Wien HC (ed) The Physiology of Vegetable Crops. CAB International, New York, pp 345–386
Wien HC (2006) Ethephon treatment may alleviate the suppression of female flowers of Cucurbita pepo under high temperatures. HortScience 41:1421–1422
Wien HC, Stapleton SC, Maynard DN, McClurg C, Riggs D (2004) Flowering, sex expression and fruiting of pumpkin (Cucurbita sp.) cultivars under various temperatures in greenhouse and distant field trials. HortScience 39:239–242
Woeste KE, Ye C, Kieber JJ (1999) Two Arabidopsis mutants that overproduce ethylene are affected in the posttranscriptional regulation of 1-aminocyclopropane-1-carboxylic acid synthase. Plant Physiol 119:521–529
Yamasaki S, Fujii N, Matsuura S, Mizusawa H, Takahashi H (2001) The M locus and ethylene-controlled sex determination in andromonoecious cucumber plants. Plant Cell Physiol 42:608–619
Yamasaki S, Fujii N, Takahashi H (2003) Photoperiodic regulation of CS-ACS2, CS-ACS4 and CS-ERS gene expression contributes to the femaleness of cucumber flowers through diurnal ethylene production under short-day conditions. Plant Cell Environ 26:537–546
Yamasaki S, Fujii N, Takahashi H (2005) Hormonal regulation of sex expression in plants. Vitamins Hormones 72:79–110
Yi HC, Joo S, Nam KH, Lee JS, Kang BG, Kim WT (1999) Auxin and brassinosteroid differentially regulate the expression of three members of the 1-aminocyclopropane-1-carboxylate synthase gene family in mung bean (Vigna radiata L.). Plant Mol Biol 41:443–454
Acknowledgments
This work was supported by the Ministerio de Ciencia e Innovación of Spain [AGL2005-06677-CO2 and AGL2008-05619-C02-02/ALI], and by the Consejería de Innovación, Ciencia y Empresa, Junta de Andalucía, Spain [CVI-02617]. We would like to thank Dr. Miriam Nuñez from the research institute INCA of Cuba for supplying the brassinazole used in this study.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Manzano, S., Martínez, C., Megías, Z. et al. The role of ethylene and brassinosteroids in the control of sex expression and flower development in Cucurbita pepo . Plant Growth Regul 65, 213–221 (2011). https://doi.org/10.1007/s10725-011-9589-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10725-011-9589-7