
Abstract
The aims of this work were to evaluate the phosphate-solubilization and hydrogen peroxide (H2O2) production by the soil-borne micromycetes, Aspergillus japonicus, Penicillium italicum and Penicillium dipodomyicola, isolated from Phragmites australis rhizosphere and to study the effect of several concentrations of Cadmium (Cd2+) on both variables. Our results showed that P. italicum achieved a higher P-solubilization and H2O2 production than A. japonicus and P. dipodomyicola, as only P. italicum showed a positive correlation (R2 = 0.71) between P-solubilization and H2O2 production. In dose–response assays, P. italicum was also more tolerant to Cd2+ (0.31 mM) in comparison to A. japonicus (0.26 mM). Analysis of the 24 factorial experimental design showed that P-solubilization by P. italicum was negatively affected by increases in Cd2+ (p = 0.04) and yeast extract (p = 0.02) in the culture medium. The production of H2O2 was positively affected only by glucose (p = 0.002). Fungal biomass production was reduced significantly (p = 0.0009) by Cd2+ and increased (p = 0.0003) by high glucose concentration in the culture medium. The tolerance and correlation between P-solubilization and H2O2 production in the presence of Cd2+ was strain and species dependent. The effects of Cd2+, glucose, ammonium sulfate and yeast extract on those variables were evaluated through a two-level factorial design. P. italicum is promising for P-solubilization in soils contaminated with Cd2+ and may be an alternative for manufacture of biofertilizers to replace chemical fertilizers.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Given the negative environmental impact of chemical fertilizers and their increasing costs (Ahmed and Shahab 2009), the use of phosphate (P)-solubilizing microorganisms (PSM) as biofertilizers opens up a new horizon for better crop productivity, as well as for protecting agro-ecosystems from hazardous agrochemicals (Sharma et al. 2013), and has attracted great attention during the last decade (Morales et al. 2011). The rhizosphere is considered to be a major niche for microbial activity, where a substantial number of microorganisms may exert a beneficial effect on plant growth. In addition, P-solubilization ability by rhizosphere microorganisms is one of the most important features associated with plant nutrition (Morales et al. 2011). Extensive research (Nenwani et al. 2010) has shown that microorganisms with the ability to transform inorganic phosphate are mainly located in the rhizosphere of plants, where they play an integral ecological role and are involved in a wide range of processes that affect P-solubilization. It has been reported (Chakraborty et al. 2010) that the presence of several types of microorganisms associated with the rhizosphere could have the ability to solubilize phosphate complexes, guaranteeing a phosphorus supply for plants (Tripura et al. 2005).
The rhizosphere microorganisms, such as soil-borne micromycetes (SBM), arbuscular mycorrhizal fungi, soil yeasts, bacteria and actinomycetes (Clark and Zeto 2000; Al-Falih 2005; Rudresh et al. 2005; Khan et al. 2007; Hamdali et al. 2008), are documented phosphate solubilizers, although SBM have certain advantages in P-solubilization over other microorganisms. For example, SBM are more acid tolerant and therefore may have a much better potential as agents to convert insoluble inorganic molecules through environmental acidification (Chuang et al. 2007); usually, P-solubilization by SBM produces more organic acid than bacteria and consequently exhibits greater P-solubilizing activity (Scervino et al. 2011). Furthermore, SBM does not lose P-solubilizing ability upon repeated sub-culturing under laboratory conditions, as occurs with bacteria (Chuang et al. 2007). In addition, SBM can traverse long distances easier and faster than bacteria and hence could be more important for P-solubilization in soils (Khan et al. 2010). Among SBM that solubilize phosphorus, the genera Aspergillus and Penicillium are the most representative, although some species of Paecilomyces, Trichoderma, Rhizoctonia, Chaetomium, Cylindrocarpon, Fusarium, Gliocladium and Humicolahave have also been reported as phosphate solubilizers (Vazquez et al. 2000; Chen et al. 2002; Pandey et al. 2008; Posada et al. 2013).
Phosphorus (P) is one of the most dynamic elements in the environment and is present in both terrestrial and aquatic environments. However, limited availability of P in agricultural soils has led to the application of fertilizers, which are accumulated in soils due to the formation of insoluble complexes with iron (FePO4) and aluminum (AlPO4) in acidic environments and calcium (Ca3(PO4)2) in alkaline environments (Deepa et al. 2010; Naik et al. 2013). These phosphates can reach water bodies through surface runoff, contributing to eutrophication processes. According to Vymazal (2007), in aquatic environments free orthophosphate is the most common form of P that can be directly utilized by plants. Therefore, it represents a link between organic and inorganic phosphorus in wetlands. Additionally, as it occurs with negative charge and tends to bind to several cations, which are poorly soluble, the assimilation of these compounds is reduced, contributing to its accumulation in sediments. According with Maitra et al. (2015), these sediments may act as an important source of P by releasing sediment-bound P through microbial metabolism, which may be an ecologically sustainable manner of supplying this element to enhance the productivity of oligotrophic water bodies.
The use of aquatic plants is one of the most effective methods for removing phosphorus and pollutants from water (Rezaie and Sahlezadeh 2014). The role of aquatic plants in phytoremediation processes has been widely documented (Arthur et al. 2005), especially for invasive plants (Coats and Rumpho 2014), due to its rapid growth, adaptability and phytoremediation capability in heavy metal removal (Rai 2008). Based on this background, in this work we focus our study on SBM isolated from plant rhizosphere of Phragmites australis. This macrophyte has a cosmopolitan distribution and can be found in contaminated environments, mainly those with heavy metals, hydrocarbons and pesticides (Hechmi et al. 2014); in this sense, the P. australis-rhizosphere is an excellent reservoir of fungal communities and could be a source of new fungal species involved in ecological processes (Jung and Nechwatal 2008). Most studies regarding phosphate solubilization by fungi (Pradhan and Sukla 2005; Sharma et al. 2013) have been focused on the isolation and characterization of microorganisms from agricultural soils, but the presence of symbiotic microorganisms associated with the rhizosphere of aquatic plants and their ecological functions have not yet been fully defined (Stottmeister et al. 2003).
Tricalcium phosphate has not been highlighted as an important source of P-release, although it constitutes about 1–52 % of total P pools in rivers, lakes and wetlands sediments, and it is considered non-exchangeable, unless there is a rapid change in sediment pH and dissolution of metal oxides (Tong et al. 2005). In addition, cadmium is a potentially toxic substance of concern to human health. Cd2+ occurs naturally and is associated with phosphate rock (Roberts 2014), a source of fertilizers, and due to its nature, can form complexes with phosphorus and can also precipitate in sediments (Baars et al. 2014; Jacob et al. 2013). However, heavy metals are toxic to SMB, even at low concentrations; Baldrian (2003) and Gadd (2004) documented the fungitoxicity of heavy metals in SBM and white-rot basidiomycetes, respectively, which resulted in growth inhibition. On the other hand, the fungal tolerance to Cd+2 in soil is associated with adsorption mechanisms (Zafar et al. 2007) that depend on the functional groups of fungal cell walls, such as carbonyls, hydroxyls and amides, which are responsible for metal binding during biosorption processes (Xu et al. 2012). In the case of toxicity, presence of metals can result in the generation of reactive oxygen species (ROS) that may cause wide-ranging damage to proteins, nucleic acids and lipids, eventually leading to cell death (Baldrian and Gabriel 2003; Li et al. 2009). Non-redox active metals like Cd2+ and zinc (Zn2+) can deplete free-radical scavengers, resulting in ROS production, such as superoxide anion (O ·−2 ), hydrogen peroxide (H2O2) and hydroxyl radical (·OH) (Azevedo et al. 2007).
The main mechanism reported for phosphate solubilization by fungi is related to the pH decrease due to organic acids and phosphatases, as these play an important role in the mineralization of phosphate salts (Gyaneshwar et al., 2002; Naik et al., 2013). For example, Aspergillus and Penicillium fungi species can produce organic acids with a high potential to produce H+ and to promote greater phosphate solubilizing activity (Gharieb 2001). Nonetheless, these mechanisms are not the only ones involved in phosphate solubilization (Sharma et al. 2013) and even environmental factors and nutritional conditions can be involved (Relwani et al. 2008). For example, fungi glucose metabolism contributes to organic acid production, whereby glucose oxidase enzymes can generate H2O2 (Wong et al. 2008) that might further contribute to a pH decrease (Magnuson and Lasure 2004), favoring phosphate solubilization. Therefore, the aims of this study were: (1) to evaluate the ability of three SBM isolated from P. australis-rhizosphere for P-solubilization and H2O2 production in liquid culture medium; (2) to study the impact of Cd2+ in development growth and P-solubilization in agar culture medium under a dose–response assay and (3) to study the effect of Cd2+ and nutritional factors (glucose, ammonium sulphate and yeast extract) on P-solubilization and H2O2 production using a 24 factorial experimental design.
Materials and methods
Isolation and molecular identification of fungal strains
Three SBM were isolated from soil samples taken in the surrounding rhizosphere area of common reed plants [Phragmites australis (Cav.) Trin. exSteud.] collected from a natural wetland area denominated “La Mancha” located in the central coast of Veracruz state in Mexico (96° 23′09″W, 19° 35′19″N). The rhizospheric soil was characterized as a sandy loam soil (clay 8.41 %, sand 89.6 and silt 2 %), organic matter content was low, pH ranged between 6.67 and 6.73, total phosphorus values ranged from 20 to 125 mg/kg, the organic phosphorus was between 15 and 85 mg/kg, total nitrogen in soil ranged from 830 to 1 645 mg/kg. The pH values of the water samples were between 6.5 and 8.0, the chemical oxygen demand for water was between 11.66 and 15 mg O2/L, total phosphorus values for water ranged between 0.88 and 2.34 mg/L, total nitrogen values for water were around 8.82 mg/L.
Fungal isolation was carried out using serial dilution of rhizosphere soil sample up to 10−3 on rose bengal streptomycin agar with tricalcium phosphate [Ca3(PO4)2] as an insoluble source of inorganic phosphorus. In this culture media after 3 days of incubation at 25 °C, a halo around the colony indicated P-solubilization. Colonies with clear zones were further purified by replating on potato dextrose agar for maintenance and conservation. For molecular identification, the mycelia of studied strains were grown in yeast extract medium (standard CYM: dextrose, 20 g; peptone, 2 g; yeast extract, 2 g; MgSO4·7 H2O, 0.5 g; KH2PO4, 0.46 g; K2HPO4, 1 g and distilled water; 1000 mL) and incubated at 25 °C during five days. The mycelia were harvested and placed in Eppendorf tubes (1.5 mL), and then frozen in liquid nitrogen (−130 °C). Genomic DNA extraction and purification of dried mycelia was carried out according to Challen et al. (1995). DNA purity was determined by comparative agarose gel electrophoresis (1.5 %, w/v) and ethidium bromide staining. The PCR amplification of the internal transcribed spacers (ITS regions) was carried out using the following primers ITS1 (5´-TCCGTAGGTGAACCTGCGG-3´) and ITS4 (5´-TCCTCCGCTTATTGATATGC-3´). The amplification condition was according to Huerta et al. (2010). Purified PCR products were sequenced by Genomic Biotechnology Center—IPN (Reynosa, Tamaulipas, Mexico). Each sample was sequenced in the genomic. DNA sequence analyses were performed using the basic sequence alignment BLAST program, run against the NCBI database (www.ncbi.nlm.nih.gov). The Index Fungorum (www.indexfungorum.org) was used as a species authority.
Culture media and inoculation
The ability of three SBM for P-solubilization and H2O2 production were evaluated in modified-Pikovskaya culture medium (Pikovskaya 1948), which consisted of (g/L): (NH4)2SO4 (0.5), KCl (0.2), MgSO4·7H2O (0.1), MnSO4·H2O (0.004), NaCl (0.2), d-glucose (10), FeSO4·7H2O (0.002, Sigma-Aldrich), yeast extract (0.5), Ca3(PO4)2 (0.5, Sigma-Aldrich) and distilled water (1000 mL). Culture medium (120 mL) was deposited in 250-mL Erlenmeyer flasks and autoclaved for 15 min at 1.5 atm and 120 °C. Fungi were inoculated into the flasks using 200 µL of a spore solution (1 × 106 spores/mL) of each fungus. The spore solution was made by taking one mycelial disk of agar from 7-days-old culture of SBM, which was deposited in 1 mL of sterile distilled water supplemented with Tween-20® (Sigma-Aldrich, St. Louis, MO, USA) at 0.05 %. The spores in the solution were counted and adjusted using a Neubauer chamber (Hausser Sci., Horsham, PA, USA). Fungal cultures were incubated at 120 rpm at 25 °C during 16 days. Four replicates were established for each fungus, and a control without inoculation was used. Every 2 days, aliquots of 8 mL from each flask were taken and centrifuged at 7500 rpm during 10 min; the supernatant was used for P and H2O2 determinations.
Analytical methods
Phosphorus-solubilization was determined by phosphomolybdenum blue method. This determination is based on the reaction of ammonium heptamolybdate and potassium antimony (III) oxide tartrate in an acid medium with a diluted solution of phosphate to form antimony–phosphomolybdate complex. This complex is reduced to an intensely blue-colored complex by L (+) ascorbic acid (Method 4500-P, Standard Methods, 2005). The complex was measured at 880 nm in a Shimadzu 8000 UV/IS spectrophotometer (Shimadzu Corp Japan). The standard curve was: y = 0.2173 c + 0.0433 (y = OD880, c = P-solubilization in mg/L, R2 = 0.983).
Hydrogen peroxide concentrations were determined using the iodide/iodate method. Solutions A and B for the I3 − method were prepared according to Klassen et al. (1994). Solution A consisted of 33 g of KI, 1 g of NaOH and 0.1 g of (NH4)6Mo7O24·4H2O (ammonium molybdate tetrahydrate) diluted to 500 mL with water. Solution A was kept in darkness to inhibit oxidation of I−. Solution B (an aqueous buffer) contained 10 g of C8H4KO4 (potassium hydrogen phthalate) dissolved in 500 mL of water. The I3 − method consisted of mixing equal weights of A (3 mL) and B (3 mL), followed by addition of fungal supernatant (3 mL). The absorbance of the resulting solution was measured at 351 nm in a 3-cm3 cuvette. The blank absorbance was determined by substituting the enzyme extract with sterile deionized water in the reaction mixture. Hydrogen peroxide concentrations were calculated by substituting the absorbance in a standard curve with pure H2O2 reagent (30 %, J.T. Baker™, Center Valley, PA, USA) at known concentrations (0.5, 1.0, 1.5, 2.0, 2.5, 3.0 and 3.5 mg/L). The standard curve was: y = 0.134c + 0.0164 (y = OD351, c = H2O2 in mg/L, R2 = 0.997).
Dose–response bioassay
The effect of Cd2+ on mycelial growth and reduction in the solubilizing halo by soil fungi was investigated in dose–response bioassays. Pikovskaya-agar culture medium was supplemented with Cd2+ introduced as Cd(NO3)2·4H2O and at four different Cd2+ concentrations (0, 0.075, 0.15, 0.31 and 0.62 mM), and a culture medium without Cd2+ was used as a control. Culture media was emptied into Petri dishes (90 mm ø); the fungi were inoculated in the center of plates using 2 µL of a spore solution (1 × 106 spores/mL). Plates were incubated in darkness at 25 °C and 75 % relative humidity (RH). Four replicates of each Cd2+ concentration and control were used. The colonies and solubilizing halo were measured daily from the rear of the plates using a millimetric square, the dose–response bioassay finished when the control mycelia filled the Petri dishes.
From the colony diameter, a specific growth characteristic was calculated: the percentage of mycelial growth inhibition, as the percentage of inhibition (% in cm/cm) = [(C–T/C] × 100, where C and T are the mycelial diameter inside the control and treatment plates, respectively. Daily growth rate (DGR), was defined as DGR = Σ (R 1–R 0 )/(T 1–T 0), where R 0 and R 1 are the colony diameter at times T 0 and T 1, respectively. Both equations were used to calculate the DGR and percentage growth inhibition of the P-solubilization halo. The effective concentration inhibiting mycelial growth rate by 50 % (IC50) was calculated by Probit analysis using the software SAS 8.1. On the basis of the Probit analysis, the most tolerant strain was chosen for studying the effect of Cd2+ and nutritional factors on P-solubilization using a factorial experimental design.
Factorial experiment design
To study the Cd2+ effect and nutrimental factors on P-solubilization by P. italicum, a 24 factorial experimental design was applied. The studied factors were: Cd2+ concentration and amounts of glucose, (NH4)2SO4 and yeast extract. Table 1 shows the factor levels of independent variables. The values of each variable were coded as minus (–1) for lower and plus (+1) for upper limits. The studied responses were: P-solubilization (Y 1), and H2O2 (Y 2) and biomass production (Y 3).
The data analysis was carried out through Design Expert 8.0 (Stat-Ease Inc., Minneapolis, MN, USA). The Eq. 1 was used to describe the regression model, which includes interaction terms:
where β0 is the constant term, βi and βij are the regression coefficient, ε is the error, x i are the variables and n their number.
Biomass quantification
For fungal dry cell weight analysis, the suspension was then filtered through pre-weighed Whatman filter paper No. 1, the fungal biomass was washed with sterilized distilled water and this was repeated until the filtrate was clear. Then, the fungal pellets were vacuum filtered through a pre-weighed filter. For dry cell weight measurement, the filters were dried in an oven until constant weight and re-weighed after cooling the filters in a desiccator.
Results
Molecular identification
Polymerase chain reaction (PCR) products from genomic DNA of fungal isolates were analyzed using the BLAST program (blast.ncbi.nlm.nih.gov/Blast.cgi). The identified fungi showed similarities between the partial sequences derived from the ITS regions. A first isolate with PCR length 569 bp was Penicillium italicum Wehmer and had 100 % similarity with accession DQ991463.1 submitted by Hernández-Montiel and Ochoa (2007). A second isolate with PCR length 534 bp was identified as Penicillium dipodomyicola and had 100 % similarity with accession KM458817 submitted by Frisvad et al. (2000). A third isolate with PCR length 594 bp was identified as Aspergillus japonicus var. aculeatus Iizuka and had 100 % similarity with accession HM140184.1 by Ma et al. (2011).
P-solubilizing and H2O2 production by SBM
All the evaluated strains were able to solubilize tricalcium phosphate at different rates (Fig. 1a). P. italicum and A. japonicus achieved the maximum P-solubilization values with 107.0 and 104.0 mg/L, respectively, and after 16 days of incubation both fungi solubilized 100 % of the initial P content introduced as Ca3(PO4)2. Figure 1b shows the H2O2 production by the strains tested through 16 d of culture. P. italicum had significantly (p < 0.05) higher H2O2 production than P. dipodomyicola in all evaluations. P. italicum achieved the highest H2O2 production (2.3–4.4 mg/L) compared to the other tested strains during 12–16 days. Pearson’s correlation analysis was used to determine the relationship between P-solubilization and H2O2 production by SBM (Table 2), in order to verify involvement of H2O2 in phosphorus release. P. italicum showed a significantly (p < 0.05) higher correlation coefficient (0.71) compared to A. japonicus (0.63) and P. dipodomyicola (0.65). On the basis of CE50 and correlation analysis, P. italicum was chosen for the factorial experiment to assess the effect of Cd2+ and nutritional factors on P-solubilization, H2O2 and biomass production.

P-solubilization (a) and H2O2 production (b) by Aspergillus japonicus (filled triangle), Penicillium italicum (filled diamond), Penicillium dipodomyicola (filled circle) and control (filled square) in modified-Pikovskaya culture medium
Dose–response assay
Penicillium italicum and A. japonicus, due to their greater capability of P-solubilization, were selected for the performance of a dose response assay under in vitro conditions (at darkness, 25 °C and 75 % RH) using a Pikovskaya-agar medium added with Cd+2. Toxicological effects on fungal growth and colony morphology were observed at the several Cd2+ levels in the culture medium. A wide range of fungal morphological changes such as, discoloration, decrease in mycelial density and low sporulation were observed during mycelial growth in culture medium when Cd2+ was added (Fig. 2). P. italicum showed a reduction in DGR per 0.15, 0.31 and 0.62 mM of Cd2+ added by 18.3, 39.8 and 61.15 %, respectively. Halo formation due to P-solubilization by P. italicum was stimulated by low Cd2+ concentration, as the halo DGR was significantly (p = 0.001) higher at 0.078 (0.78 cm/d) and 0.155 mM Cd2+ (0.73 cm/d) in comparison to controls without Cd2+ (0.68 cm/d). Reduction in the halo DGR was only achieved at 0.31 and 0.62 mM Cd2+ with 26.3 and 40.5 % reductions, respectively (Table 3). A. japonicus was less susceptible to Cd2+, because its mycelial DGR was stimulated by low Cd2+ concentrations. Its mycelial DGR was significantly (p = 0.001) higher for 0.07 and 0.155 mM Cd2+ (0.86 and 0.77 cm/d, respectively) than for controls (0.75 cm/d). Only high Cd2+ concentrations reduced the colony DGR by 29.6 and 88.5 % (0.31 and 0.62 mM Cd2+, respectively). However, halo DGR was more affected than colony DGR, with reductions of 9.0, 19.6, 44.6 and 92.4 % for 0.078, 0.155, 0.31 and 0.62 mM Cd2+, respectively (Table 4). A wide range of fungal morphological changes, such as discoloration, decrease in mycelial density and low sporulation were observed during mycelial growth in culture medium with added Cd2+. The same influence was also detected in liquid media for concentrations of 0.25 and 50 mM Cd2+. When compared with other fungal species, such as Fusarium sp. and Alternaria tenuis, the observed effect on growth and morphology was the same, regardless of Cd2+ source and culture media (Gharieb 2001).

Dose-response assay in Pikovskaya-agar medium contaminated with Cd2+ levels: (Up) Penicillium italicum and (Down) Aspergillus japonicus. Control (a), 0.07 mM (b), 0.015 mM (c), 0.031 mM (d) and 0.62 mM (e)
Factorial experimental design and mathematical models
On the basis of CE50 and correlation analysis, P. italicum was chosen for the factorial experiment design to assess the effect of Cd2+ and nutritional factors on dependent variables. The results of 20 experiments were used to obtain three mathematical models (Table 5), each obtained from Eq. 1 through the study of the partial coefficients of each factor and interactions between factors. The fitted models were designed to explain the relationship between the independent factors and dependent responses.
P-solubilization
The mathematical model for the P-solubilization allowed us to define the optimal media composition and to know the combined effects of studied factors. In addition, the predicted responses (Figs. 3, 4) as well as the residual values between experimental and theoretical data were calculated (Fig. 5a). The ANOVA for phosphate solubilization of P. italicum through the 20 assays of the factorial experiment is shown in Table 6. The theoretical data showed a functional relationship with the experimental data, indicating that the predictability of the model was significant at 95 % of confidence level. The coefficient of determination (R2) value of 80.26 % showed that Eq. 2 was highly reliable. Figure 3 shows observed responses versus those from the statistical model of Eq. 2.

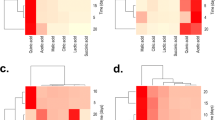
Graphical analyses of the interaction Cd+2 and yeast for P solubilization obtained through the analysis with Design-Expert software

Interaction analyses a Cd+2 and yeast and b Cd2+ and glucose for biomass production obtained through the analysis with Design-Expert Sofware

Graphical analyses of the model fitted for Y1 = P solubilization (a), Y2 = hydrogen production (c), and Y3 = biomass (e). The regression analyses were set between predicted responses and experimental values. Statistically analysis was by normality plot of the residual Y1 (b), Y2 (d), and Y3 (f)
The coefficient of determination (R2 = 0.8) indicated a moderately strong relationship between predicted data of P-solubilization from empirical observations and the experimentally obtained data. A normal probability plot of the residuals to assess normality resulted in a straight line (Fig. 5b). In addition, the negative coefficients corresponding to Cd2+ (−11.49) and yeast extract (−13.90) indicated that P-solubilization decreased with increasing concentration of Cd2+ and yeast extract.
Hydrogen peroxide production
According to the ANOVA, the glucose amount has a significant effect on H2O2 production (p = 0.002); the other studied factors and their interactions did not show significant effects. The highest H2O2 production (3.23 mg/L) was found in the experiment with a high concentration of Cd2+ (0.45 mM) and a high amount of glucose (15 g/L), (NH4)2 SO4(0.75 g/L) and yeast extract (0.75 g/L). Assay 17 with low concentration of Cd2+ and glucose but high concentration of ammonium nitrate and yeast extract resulted in the lowest H2O2 production (2.16 mg/L). The positive coefficient for glucose indicates that the maximum production of H2O2 occurred for the highest amount of glucose. Figure 5c contrasts the observed response and the predicted response. The normality test was used to construct a normal probability plot, resulting in a straight line (Fig. 5d). The Pearson’s correlation coefficient was high (0.89), indicating a moderately strong relationship between observed and predicted responses.
Biomass production
The model for biomass production (Eq. 3) was significant and reliable (p = 0.004 and R2 = 0.88). The factors showing positive effects with a confidence level of 95 % were Cd2+ (p = 0.0009) and glucose (p = 0.0003). Similarly, the interactions of Cd2+*glucose (p = 0.02) and Cd2+*yeast (p = 0.005) were significant. The ANOVA for biomass production through the 20 assays of the factorial experiment is shown in Table 7.
The highest value of biomass was achieved in assay 9 (5.43 mg/L), which contained a low amount of Cd2+ and high levels of glucose, ammonium nitrate and yeast extract. In contrast, the lowest biomass was for assay 8, which contained high values of Cd2+ and yeast extract and low amounts of glucose and ammonium nitrate. The negative coefficient corresponding to Cd2+ (−0.82) in Eq. 3 suggested that fungal biomass decreased with increasing Cd2+ concentration in the culture medium. The yeast extract (−0.28) showed a similar but weaker effect to Cd2+ (Fig. 4a). Glucose played an important role in biomass production by P. italicum (Fig. 4b) as shown by the positive coefficient of glucose (0.96) in Eq. 3. Ammonium nitrate was important for biomass production, although less important than glucose. Figure 5e shows the contrast of experimental and predicted responses, and Fig. 5f is a normal probability plot.
Discussion
Fungus genera, such as Penicillium and Aspergillus, have been shown to have phosphate-solubilizing capacities (Mittal et al. 2008; Naik et al. 2013; Saxena et al. 2013). In this study, three strains isolated from rhizosphere soil of P. australis have not been reported for phosphate solubilization under in vitro conditions. Fungal strains P. italicum and A. japonicus strains presented higher P-solubilization ability and were selected for the implementation of the dose response assay. The IC50 calculated by Probit analysis suggested that P. italicum was more tolerant to Cd+2 than A. japonicus, although the latter showed a high DGR under low Cd2+ concentrations in a Pikovskaya-agar medium. Some studies have reported the minimum inhibitory concentration (MIC) for some fungal species under semi-solid cultivation; the MICs for Cd2+ of A. japonicus and Penicillium sp. isolated from metal-contaminated soils were 35.5 and 44.4 mM (Zafar et al. 2007). Ahmad et al. (2005) investigated the biosorption of Cd2+ by Rhizopus and found a MIC of 17.8 mM. These values are high compared to the CI50 of P. italicum (0.31 mM) and A. japonicus (0.26 mM), although the differences could be due to differences in methodology and statistical methods used to determine MICs and CI50. Similarly, the solubilization halo in both P. italicum and A. japonicus showed differences in percentage inhibition, with the most evident reduction at high Cd2+ concentrations (0.31 and 62 mM); however, P. italicum showed less reduction in halo solubilization at 0.62 mM Cd2+ in comparison to A. japonicus. The decrease in mycelial growth was due to the toxic effect of Cd2+. Nonetheless, the solubilization halo present in solid media could suggest that fungi under Cd2+ stress can divert energy from growth to cell maintenance functions (Muhammad et al. 2005).
The factorial experiment confirmed that Cd2+ decreased the ability of soil fungi to solubilize P and produce biomass. We suggest that the decrease in P-solubilization was correlated with the fungal biomass reduction due to the toxic effect of Cd2+ (Vig et al. 2003). Tuason and Arocena (2009) demonstrated that P-solubilization was positively correlated with fungal biomass, glucose oxidation and other intermediates of energy metabolism pathways that are important in the production of organic acids (Chaiharn and Lumyong 2009), corresponding to decreases in pH (Walpola and Yoon 2013). This phenomenon has been directly related to the release of phosphorus of tricalcium phosphate (Johnson and Loeppert 2006; Onthong et al. 2007; Scervino et al. 2010). However, this does not seem to apply to all cases, as there have been reports of positive but non-significant correlations between the amount of soluble P and low molecular weight acids produced by fungi (Pradhan and Sukla 2005). Thus, although organic acid generation is important to fungal metabolism, it may not be the only factor involved in P-solubilization (Rashid et al. 2004). The production of oxalic acid by fungi provides a means of immobilizing soluble metal ions or complexes as insoluble oxalates, thus decreasing bioavailability and increasing tolerance of these metals (Gadd 2007). In our study, decreasing metal bioavailability could be associated to metal chelation by organic acids under acidic conditions (pH below 5), as it was reported by Gadd (2004), rather than precipitation that occurs at alkaline pH (Hong et al. 2010).
In the presence of Cd2+, organic acids can act as chelating agents, which decrease the metal toxicity (Khan 2005); however, the presence of this metal may in turn influence the generation of organic acids. Nonetheless, this may not be generalized for all species of fungi, as this activity is influenced by metal concentration, nitrogen form and other culture conditions (Sazanova et al. 2015). The mechanisms used to solubilize microbial phosphate include acidification, chelation and exchange reactions, and are affected by nutritional conditions. Most of the previous reports (Pradhan and Sukla 2005) state that calcium phosphates are dissolved by acidification when glucose and ammonium sulphate are used, favoring a decrease in pH. In addition to P-solubilization, we studied H2O2 production as a response mechanism to physiological stress due to Cd2+ concentration in the culture medium. Although it was possible to determine a correlation between P-solubilization and H2O2 production, we did not find a significant effect of Cd2+on H2O2 production. It was likely that other physiological responses were involved in the stress induced by Cd2+, such as enzyme activities, organic acid production or another ROS types. There are some reports (Hammel et al. 2000; Tanaka et al. 2006) concerning the production of ROS in several fungal species; however, their role in fungal metabolism is unclear (Bai et al. 2003). Therefore, more detailed study of the particular case of H2O2 is necessary to determine its role in Cd2+-induced stress and therefore P-solubilization.
P-solubilization by soil-borne fungi is complex and not completely understood (Narula et al. 2000). In the present study, we found that yeast extract negatively affected the P-solubilization, as yeast extract is a nitrogen source but more complex that other sources, like sulfate ammonium form (Nahas 2007). The increase of Cd2+ in Pikovskaya-liquid medium negatively affected P-solubilization; however, high concentrations of glucose reduced the effect of Cd2+ on P-solubilization. In addition to nutritional factors (Fomina et al. 2003), the influence of abiotic factors such as pH, temperature, available oxygen and light can affect P-solubilization by soil fungi. The effects of these factors and their interactions with heavy metals should also be considered for P-solubilization by SBM.
Conclusion
Penicillium italicum achieved faster P-solubilization and higher H2O2 production than A. japonicus and P. dipodomyicola, although there was a correlation between P-solubilization and H2O2 production only for P. italicum; however, no correlation was observed with addition of Cd2+. The Cd2+ amount negatively affected both the DGR of mycelium and halo solubilization. P. italicum was more tolerant to Cd2+ compared to A. japonicus and P. dipodomyicola in liquid culture medium, where the Cd2+ presence and yeast extract negatively affected the P-solubilization by P. italicum. Cd2+ levels did not influence H2O2 production, although a positive effect was observed with high glucose concentration. Fungal biomass decreased in the presence of Cd2+ and increased with addition of high glucose concentration in the culture media. Our results confirmed that the Cd2+ effect on P-solubilizing fungi was species- and strain-dependent, and we suspect that different mechanisms are involved in Cd2+-induced stress, which should be studied in more detail.
References
Ahmad I, Zafar S, Ahmad F (2005) Heavy metal biosorption potential of Aspergillus and Rhizopussp. isolated from waste water treated soil. J Appl Sci Environ Manag 9(1):123–126
Ahmed N, Shahab S (2009) Phosphate solubilization: their mechanism genetics and application. Int J Microbiol 9(1):4408–4412
Al-Falih MA (2005) Phosphate solubilization in vitro by some soil yeasts. Qatar Univ Sci J 25:119–125
Arthur E, Rice P, Anderson T, Baladi S, Henderson K, Coats J (2005) Phytoremediation: an overview. Crit Rev Plant Sci 24:109–122
Azevedo M, Carvalho A, Pascoal C, Rodrigues F, Cassio F (2007) Responses of antioxidant defenses to Cu and Zn stress in two aquatic fungi. Sci Total Environ 377(2–3):233–243
Baars O, Abouchami W, Galer SJ, Boye M, Croot PL (2014) Dissolved cadmium in the Southern Ocean: distribution, speciation, and relation to phosphate. Limnol Oceanogr 59(2):385–399
Bai Z, Harvey LM, McNeil B (2003) Oxidative stress in submerged cultures of fungi. Crit Rev Biotechnol 23(4):267–302
Baldrian P (2003) Interactions of heavy metals with white-rot fungi. Enzym Microb Technol 32(1):78–91
Baldrian P, Gabriel J (2003) Lignocellulose degradation by Pleurotus ostreatus in the presence of cadmium. FEMS Microbiol Lett 220(2):235–240
Chaiharn M, Lumyong S (2009) Phosphate solubilization potential and stress tolerance of rhizobacteria from rice soil in Nothern Thailand. World J Microbiol Biotechnol 25(2):305–314
Chakraborty BN, Chakraborty U, Saha A, Sunar K, Dey PL (2010) Evaluation of phosphate solubilizers from soils of North Bengal and their diversity analysis. World J Agric Sci 6(2):195–200
Challen MP, Moore AJ, Martinez-Carrera D (1995) Facile extraction and purification of filamentous fungal DNA. Biotechniques 18(6):975–978
Chen CR, Condron LM, Davis, Sherlock RR (2002) Phosphorus dynamics in the rhizosphere of perennial ryegrass (Lolium perenne L.) and radiata pine (Pinus radiata D. Don.). Soil Biol Biochem 34(4):487–499
Chuang CC, Kuo YL, Chao CC, Chao WL (2007) Solubilization of inorganic phosphates and plant growth promotion by Aspergillus niger. Biol Fertil Soils 435:575–584
Clark RB, Zeto SK (2000) Mineral acquisition by arbuscular mycorrhizal plants. J Plant Nutr 23(7):867–902
Coats V, Rumpho ME (2014) The rhizosphere microbiota of plant invaders: an overview of recent advances in the microbiomics of invasive plants. Front Microbiol 5(1):1–10
Deepa V, Aadarsh P, Balakrishna MP, Sridhar R (2010) Efficient phosphate solubilization by fungal strains isolated from rice-rhizosphere soils for the phosphorus release. Res J Agric Biol Sci 6(4):487–492
Fomina M, Ritz K, Gadd GM (2003) Nutritional influence on the ability of fungal mycelia to penetrate toxic metal-containing domains. Mycol Res 107(7):861–871
Frisvad JC, Filtenborg O, Lund F, Samson RA (2000) The homogeneous species and series in subgenus Penicillium are related to mammal nutrition and excretion. In: Samson RA, Pitt JI (eds) Integration of modern taxonomic methods for Penicillium and Aspergillus classification. Harwood Academic, Amsterdam, pp 265–283
Gadd GM (2004) Microbial influence on metal mobility and application to bioremediation. Geoderma 122(2–4):109–119
Gadd GM (2007) Geomycology: biogeochemical transformations of rocks, minerals, metals and radionuclides by fungi, bioweathering and bioremediation. Mycol Res 111(1):3–49
Gharieb MM (2001) Pattern of cadmium accumulation and essential cations during growth of cadmium-tolerant fungi. BioMetals 14(2):143–151
Gyaneshwar P, Kumar GN, Parekh LJ, Poole PS (2002) Role of soil microorganisms in improving P nutrition of plants. Plant Soil 245(1):83–93
Hamdali H, Bouizgarne B, Hafidi M, Lebrihi A, Virolle MJ, Ouhdouch Y (2008) Screening for rock phosphate solubilizing Actinomycetes from Moroccan phosphate mines. Appl Soil Ecol 38(1):12–19
Hammel KE, Kapich AN, Jensen KA, Ryan ZC (2000) Reactive oxygen species as agents of wood decay by fungi. Enzym Microb Technol 30(4):445–453
Hechmi N, Aissa NB, Jedidi N (2014) Evaluating the phytoremediation potential of Phragmites australis growth in pentachlorophenol and cadmium co-contaminated soils. Environ Sci Pollut Res Int 21(2):1304–1313
Hernández-Montiel LG, Ochoa JL (2007) Fruit rot caused by Penicillium italicum on Lemon (Citrus aurantifolia) in Colima, Mexico. Plant Dis 91(6):767–777
Hong CO, Chung DY, Lee DK, Kim PJ (2010) Comparison of phosphate materials for immobilizing cadmium in soil. Arch Environ Contam Toxicol 58(2):268–274
Huerta G, Martínez-Carrera D, Sánchez JE, Leal-Lara E, Vilgalys R (2010) Genetic relationships between Mexican species of Pleurotus analyzing the ITS-region from rDNA. Micol Apl Int 22(1):15–25
Jacob DL, Yellick AH, Kissoon LTT, Asgary A, Wijeyaratne DN, Saini-Eidukat B, Otte ML (2013) Cadmium and associated metals in soils and sediments of wetlands across the Northern Plains, USA. Environ Pollut 178(1):211–219
Johnson SE, Loeppert RH (2006) Role of organic acids in phosphate mobilization from iron oxide. Soil Sci Soc Am J 70(1):222–234
Jung T, Nechwatal J (2008) Phytophthora gallica sp. nov.a new species from rhizosphere soil of declining oak and reed stands in France and Germany. Mycol Res 112(10):1195–1205
Khan AG (2005) Role of soil microbes in the rhizospheres of plants growing on trace metal contaminated soils in phytoremediation. J Trace Elem Med Biol 18(4):355–364
Khan MS, Zaidi A, Wani PA (2007) Role of phosphate-solubilizing microorganisms in sustainable agriculture—a review. Agron Sustain Dev 27(1):29–43
Khan MS, Zaidi A, Ahemad M, Oves M, Wani PA (2010) Plant growth promotion by phosphate solubilizing fungi-current perspective. Arch Agron Soil Sci 56(1):73–98
Klassen NV, Marchington D, Mcgowan HCE (1994) H2O2 Determination by the I3-method and by KMnO4 titration. Anal Chem 66(18):2921–2925
Li Q, Harvey LM, McNeil B (2009) Oxidative stress in industrial fungi. Crit Rev Biotechnol 29(3):199–213
Ma ZY, Pu SC, Jiang JJ, Huang B, Fan MZ, Li ZZ (2011) A novel thermostable phytase from the fungus Aspergillus aculeatus RCEF 4894: gene cloning and expression in Pichia pastoris. World J Microbiol Biotechnol 27(3):679–686
Magnuson JK, Lasure LL (2004) Organic acid production by filamentous fungi. In: Tkacz J, Lange L (eds) Advances in fungal bio/technology for industry, agriculture and medicine. Springer, Berlin, pp 307–340
Maitra N, Manna SK, Samanta S, Sarkar K, Debnath D, Bandopadhyay C, Sahu SK, Sharma AP (2015) Ecological significance and phosphorus release potential of phosphate solubilizing bacteria in freshwater ecosystems. Hydrobiologia 745(1):69–83
Mittal V, Singh O, Nayyar H, Kaur J, Tewari R (2008) Stimulatory effect of phosphate solubilizing fungal strains (Aspergillus awamori and Penicillium citrinum) on the yield of chickpea (Cicerarietinum L. cv. GPF2). Soil Biol Biochem 40(3):718–727
Morales A, Alvear M, Valenzuela E, Castillo CG, Borie F (2011) Screening, evaluation and selection of phosphate-solubilizing fungi as potential biofertilizer. J Soil Sci Plant Nutr 11(4):89–103
Muhammad A, Xu J, Li Z, Wang H, Yao H (2005) Effects of lead and cadmium nitrate on biomass and substrate utilization pattern of soil microbial communities. Chemosphere 60(4):508–514
Nahas E (2007) Phosphate solubilizing microorganisms In: Velázquez E, Rodriguez-Barrueco C (eds.) Effect of carbon, nitrogen, and phosphorus sources. First International Meeting on Microbial Phosphate Solubilization, Salamanca, Spain. pp. 111–115
Naik SK, Maurya S, Kumar R, Sadhna K, Gagrai S, Das B, Kumar S, Bhatt BP (2013) Inorganic phosphate solubilization by phosphate solubilizing fungi isolated from acidic soils. Afr J Microbiol Res 7(34):4310–4316
Narula N, Kumar V, Behl RK, Deubel A, Gransee A, Merbach W (2000) Effect of P solubilizing Azotobacter chroococcum on N, P, K uptake in P-responsive wheat genotypes grown under greenhouse conditions. J Plant Nutr Soil Sci 163(4):393–398
Nenwani V, Doshi P, Saha T, Rajkumar S (2010) Isolation and characterization of a fungal isolate for phosphate solubilization and plant growth promoting activity. J Yeast Fungal Res 1:9–14
Onthong J, Gimsanguan S, Pengnoo A, Nilnond C, Osaki M (2007) Effecf of pH and some cations on activity of acid phosphatase secreted from Ustilago sp. Isolated from acid sulphate soil. Songklanakarin. J Sci Technol 29:275–286
Pandey A, Das N, Kumar B, Rinu K, Trivedi P (2008) Phosphate solubilization by Penicillium spp. isolated from soil samples of Indian Himalayan region. World J Microbiol Biotechnol 24(1):97–102
Pikovskaya RI (1948) Mobilization of phosphorus in soil in connection with vital activity of some microbial species. Mikrobiologiya 17:362–370
Posada RH, Heredia-Abarca G, Sieverding E, Sánchez de Prager M (2013) Solubilization of iron and calcium phosphates by soil fungi isolated from coffee plantations. Arch Agron Soil Sci 59(2):185–196
Pradhan N, Sukla LB (2005) Solubilization of inorganic phosphates by fungi isolated from agriculture soil. Afr J Biotechnol 5(10):850–854
Rai PK (2008) Heavy-metal pollution in aquatic ecosystems and its phytoremediation using wetland plants: an eco sustainable approach. Int J Phytoremediation 10(2):133–160
Rashid M, Khalil S, Ayub N, Alam S, Latif F (2004) Organic acids production and phosphate solubilization by phosphate solubilizing microorganisms (PSM) under in vitro conditions. Pak J Biol Sci 7(2):187–196
Relwani L, Krishna P, Reddy M (2008) Effect of carbon and nitrogen sources on phosphate solubilization by a wild-type strain and UV-induced mutants of Aspergillus tubingensis. Curr Microbiol 57(1):401–406
Rezaie H, Sahlezadeh M (2014) Performance removal nitrate and phosphate from treated municipal wastewater using Phragmites australis and Typha latifolia aquatic plants. J Civil Eng Urban 4(3):315–321
Roberts TL (2014) Cadmium and phosphorus fertilizers: the issues and the science. Proc Eng 83:52–59
Rudresh DL, Shivaprakash MK, Prasad RD (2005) Tricalcium phosphate solubilizing abilities of Trichoderma sp. in relation of P uptake and growth and yield parameters of chickpea (Cicerarietinum L.). Can J Microbiol 51(3):217–222
Saxena J, Basu P, Jaligam V, Chandra S (2013) Phosphate solubilization by a few fungal strains belonging to the genera Aspergillus and Penicillium. Afr J Microbiol Res 7(41):4862–4869
Sazanova K, Osmolovskaya N, Schiparev S, Yakkonen K, Kuchaeva L, Vlasov D (2015) Organic acids induce tolerance to zinc- and copper-exposed fungi under various growth conditions. Curr Microbiol 70(4):520–527
Scervino JM, Mesa MP, Della Mónica I, Recchi M, Moreno NS, Godeas A (2010) Soil fungal isolates produce different organic acid patterns involved in phosphate salts solubilization. Biol Fertil Soils 46(1):755–763
Scervino JM, Papinutti VL, Godoy MS, Rodriguez MA, Della Mónica I, Recchi M, Pettinari MJ, Godeas AM (2011) Medium pH, carbon and nitrogen concentrations modulate the phosphate solubilization efficiency of Penicillium purpurogenum through organic acid production. J Appl Microbiol 110:1215–1223
Sharma SB, Sayyed R, Trivedi MH, Gobi TA (2013) Phosphate solubilizing microbes: sustainable approach for managing phosphorus deficiency in agricultural soils. Springer Plus 2:587–591
Standard Methods for the Examination of Water and Wastewater (2005) 21st ed. American Public Health Association APHA, Washington, DC
Stottmeister U, Wiessner A, Kuschk P, Kappelmeyer U, Kastner M, Bederski O, Muller RA, Moormann H (2003) Effects of plants and microorganisms in constructed wetlands for wastewater treatment. Biotechnol Adv 22(1):93–117
Tanaka A, Christensen MJ, Takemoto D, Park P, Scott B (2006) Reactive oxygen species play a role in regulating a fungus-perennial rye grass mutualistic association. Plant Cell 18(4):1052–1066
Tong Y, Lin G, Ke X, Liu F, Zhu G, Gao G, Shen J (2005) Comparison of microbial community between two shallow freshwater lakes in middle Yangtze basin, East China. Chemosphere 60(1):85–92
Tripura CB, Sashidhar B, Podile AR (2005) Transgenic mineral phosphate solubilizing bacteria for improved agricultural productivity. In: Satyanarayana T, Johri BN (eds) Microbial diversity current perspectives and potential applications. I. K. International Pvt. Ltd., New Delhi, pp 375–392
Tuason MM, Arocena JM (2009) Calcium oxalate biomineralization by Piloderma fallax in response to various levels of calcium and phosphorus. Appl Environ Microbiol 72(22):7079–7085
Vazquez P, Holguin G, Puente ME, Lopez-Cortes A, Bashan Y (2000) Phosphate-solubilizing microorganisms associated with the rhizosphere of mangroves in a semiarid coastal lagoon. Biol Fert Soils 30:460–468
Vig K, Megharaj M, Sethunathan N, Naidu R (2003) Bioavailability and toxicity of cadmium to microorganisms and their activities in soil: a review. Adv Environ Res 8(1):121–135
Vymazal J (2007) Removal of nutrients in various types of constructed wetlands. Sci Total Environ 380:48–65
Walpola BC, Yoon M (2013) In vitro solubilization of inorganic phosphates by phosphate solubilizing microorganisms. Afr J Microbiol Res 7(27):3534–3541
Wong CM, Wong KH, Chen CD (2008) Glucose oxidase: natural occurrence, function, properties and industrial applications. Appl Microbiol Biotechnol 78(1):927–938
Xu X, Huang Q, Gu JD, Chen W (2012) Biosorption of cadmium by metal-resistant filamentous fungus isolated from chicken manure compost. Environ Technol 33(13–15):1661–1670
Zafar SF, Aqil F, Ahmad I (2007) Metal tolerance and biosorption potential of filamentous fungi isolated from metal contaminated agricultural soil. Biores Technol 98(13):2557–2561
Acknowledgments
The authors are greatly indebted to Gisselly Mendoza for their technical assistance. This work was funded by own resources from the Laboratory of Xenobiotics of CINVESTAV-IPN, Mexico. Also the first author wish to thank the Mexican National Council of Science and Technology (CONACyT) by his doctoral scholarship.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
Zúñiga-Silva, J.R., Chan-Cupul, W., Kuschk, P. et al. Effect of Cd+2 on phosphate solubilizing abilities and hydrogen peroxide production of soil-borne micromycetes isolated from Phragmites australis-rhizosphere. Ecotoxicology 25, 367–379 (2016). https://doi.org/10.1007/s10646-015-1595-5
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10646-015-1595-5