
Abstract
The Argentine ant is one among the 100 worst invasive animal species of the world. It has invaded six continents, and poses significant threats to the native invertebrate fauna. Prior knowledge on the suitable ecological niches of Argentine ants may help to devise better management plans. This study delineates the niches of Argentine ants using the ecological niche modeling approach by maintaining the temporal match between the distributional information and environmental data. We used an exhaustive methodological framework to understand the niche characteristics of Argentine ants in their native and invaded ranges. The model developed using combined native and invaded range distributional information provided better habitat suitability predictions in the invaded ranges. Efforts were also made to identify the suitable niches of this invasive species globally. The current global model was projected onto two future periods (2041–2070 and 2071–2100) under two different emission scenarios (SSP126 and SSP585). Our study highlights four important features related to the ecological niches of Argentine ants: (1) large habitat preference towards permanent streams in the native ranges, (2) native ecological niche is only partly conserved across invaded ranges, (3) invaded range niches are largely differentiated, and (4) global warming induces an increase in habitat suitability in the northern hemisphere. The results of this study may provide potential insights for the effective implementation of management strategies.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The Argentine ant (Linepithema humile Mayr, 1868, Formicidae) poses significant global threats to both vertebrates and invertebrates (Vega and Rust 2000; Wetterer et al. 2001, 2009; Suarez et al. 2005; Krushelnycky and Gillespie 2008; Angulo et al. 2024) due to its high invasion potential (Lowe et al. 2000; Boser et al. 2017; Angulo et al. 2022). The drastic ecological and economic damage caused by Argentine ants, led to their listing as one of the world’s 100 worst invasive species (Lowe et al. 2000). The estimated global economic damage per annum is ~ 0.5 M USD (Angulo et al. 2024). Introduction of Argentine ants has been deemed to have caused either complete loss or significant reduction in abundance of native ant populations (Walters 2006; Rowles and O’Dowd 2007; Boser et al. 2014; Naughton et al. 2020). In some of the invaded regions, they directly replace native ant communities (Silverman and Brightwell 2008), and disrupt food chain dynamics and ecological processes (Rowles and O’Dowd 2007). Key ecological processes, altered by Argentine ants include (1) disrupting the plant-pollinator interactions (Liang et al. 2022), (2) decreasing the rate of seed dispersal (Frasconi Wendt et al. 2022) and seed germination (Gómez et al. 2003), (3) reduced decomposition of litter (Stanley and Ward 2012), (4) limiting food resources of vertebrates (Suarez et al. 2000), and (5) affecting plant fitness via mutualistic interactions with pests (Silverman and Brightwell 2008), to name a few. In addition, they can cause damage to the drip irrigation systems in agricultural fields, resulting in serious economic losses (Ota and Chang 1991).
Native distribution of the Argentine ants is restricted to lower Paraná River basin in subtropical Argentina, Brazil, Paraguay, and Uruguay (Wetterer et al. 2009), and the ecosystems in native ranges are characterized by warm-temperate climatic conditions (De Fina 1992). The introduction of Argentine ants was initially documented in 1882 on Madeira Island, and through unintentional human-mediated dispersal (Park et al. 2023), it has invaded six continents where Mediterranean-like climatic conditions mostly prevail (Suarez et al. 2001; Wild 2004). However, invasion of Argentine ants is not only limited to regions with Mediterranean-type climate but have also successfully established themselves in areas with mild-temperate and sub-tropical conditions (Suarez et al. 2001). It is interesting to note that the natural range expansion of Argentine ants is extremely limited due to queens’ preference for stationary intranidal mating. Male and female mate within their natal nests, and afterwards the inseminated queens leave established nests on foot along with a group of workers, and form new nests nearby; thereby eliminating the chances of long-distance nuptial flights (Newell and Barber 1913; Silverman and Brightwell 2008). As clandestine travelers, Argentine ants travel long-distances via all modes of human-mediated transport (i.e., land, air, and water) (Silverman and Brightwell 2008). Documented evidence indicates that average point-to-point dispersal distance is ~ 160 km for L. humile in the United States, revealing that human-mediated jump/long distance dispersal is the most important reason behind the global introduction of this species (Silverman and Brightwell 2008). New introduction records are still accumulating (Masoni et al. 2020; Slimani et al. 2020; Charrier et al. 2020; López-Collar and Cabrero-Sañudo 2021; Castro-Cobo et al. 2021; Song et al. 2022; Park et al. 2023).
Using ecological niche modeling (ENM) framework to predict climatic niche suitability for invasive species in their native, invaded, and non-invaded regions can generate valuable insights pertaining to the possibility of their spread to novel areas in both space and time (Peterson 2003; Jiménez-Valverde et al. 2011; Cordier et al. 2020); such forecasts can help both public administrations and management agencies to develop integrated pest management policies to mitigate the ecological and economic impacts of invasive species (Beaury et al. 2020). Ecologists’ interest in identifying niche characteristics of invasive species (Cordier et al. 2020) is evident from the ongoing accumulation of scientific work on estimating the invasive potential of both terrestrial (e.g., Wang et al. 2022; Nair and Peterson 2023; Polaina et al. 2020) and aquatic (e.g., Gimenez et al. 2022; Marcelino and Verbruggen 2015; Cobos and Peterson 2023; Lo Parrino et al. 2023) invasive species using ENM framework. Although the utility of ENM in deciphering the patterns of species invasions in space and time has been strongly emphasized (Peterson 2003; Liu et al. 2020), choice of methods for implementing ENM deserves careful scrutiny (Jiménez-Valverde et al. 2011; Qiao et al. 2017; Feng et al. 2019; Dubos et al. 2023; Lo Parrino et al. 2023) to avoid erroneous interpretations.
Several efforts have been made to characterize the climatic niches of ants at global and local scales for different purposes: (i) identifying potential distribution, (ii) conservation prioritization, and (iii) assessing invasion risks (reviewed by Silva et al. 2022). Due to Argentine ants’ notoriety as one of the most damaging invasive pests (Lowe et al. 2000), assessment of its invasive potential at various spatio-temporal scales using ENM has been a topic of interest for ecologists since the beginning of the twenty-first century (Silva et al. 2022). Global-scale mapping of the potential distribution of L. humile has been attempted using various modeling algorithms under different assumptions to study the invasion risks worldwide (Roura-Pascual et al. 2004, 2011; Hartley et al. 2006; Bertelsmeier et al. 2015; Jung et al. 2022). Fine-scale mapping of ecological niche differentiation based on native and invaded ranges using cross-prediction approach was attempted by Roura-Pascual et al. (2006). Local-scale characterization of ecological niches aimed at identifying regional trends associated with the spread of Argentine ants has been made in China (Li et al. 2022), Catalonia (Abril et al. 2009), Catalonia and Costa Brava (Roura‐Pascual et al. 2009), California (Menke et al. 2009), Iberian Peninsula (Roura-Pascual et al. 2009a, b), New Zealand (Harris & Barker 2007; Ward 2009), and northern California (Fitzgerald et al. 2012).
Argentine ants’ invasion is still progressing (e.g., Charrier et al. 2020; Park et al. 2023; Song et al. 2022); timely efforts to re-calibrate the ecological niche models of invasive species using (i) updated climate data, (ii) occurrences from newly invaded regions, and (iii) current practices in modeling, are recommended to obtain reliable models for realistic interpretations (Pili et al. 2020). In this study, we re-assessed the niche characterization of L. humile by re-defining four components: (1) simulation-based estimation of the areas that can be reached by the species of interest via dispersal within the relevant timeframe (area M; Barve et al. 2011; Machado-Stredel et al. 2021) in native and invaded ranges, (2) filtered occurrence records based on the year of observation to avoid temporal mismatch with climate data (Heap and Culham 2010; Roubicek et al. 2010), (3) more-recent high resolution climate data from CHELSA version 2.1 (Karger et al. 2017), and (4) characterization of native and invaded niches in environmental space using 3D ellipsoid-based niche models (Farber and Kadmon 2003).
Enhancing the predictive capabilities of our models involves refining and updating the modeling components; our re-assessment of potential distribution of Argentine ants aimed to (1) quantify the niche overlap and niche similarities between native and invaded ranges; also, among invaded ranges, (2) investigate the performance of climate matching- (i.e., native model projected onto invaded ranges) and pooled-models (i.e., models calibrated using native + invaded and invaded only data sets) on accurately predicting the invasive trends of Argentine ants, and (3) forecast shifts in the global potential distribution of Argentine ants in two future climates (2041–2070 and 2071–2100) using five general circulation models (GCMs) under two socio-economic pathways (SSP126 and SSP585).
The phenomenon of heat accumulation in urban settings due to rapid socioeconomic developments is termed as Urban Heat Island (UHI) Effect (Yang et al. 2016). Considering the large influence of UHI effect in maintaining the populations of Argentine ants in invaded ranges (McGlynn et al. 2019; López-Collar and Cabrero-Sañudo 2021), we expect that the socio-economic development-induced climate changes may increase the habitat suitability of this species globally. We hypothesize that environmental niches between native and invaded ranges of Argentine ants are divergent due to the extensive geographic coverage of the latter, allowing this species to utilize potential novel environmental conditions.
Materials and methods
We have presented our exhaustive experimental approach to characterize the ecological niches of Argentine ants in native, invaded, and global ranges using recent distributional and climatic information in Fig. 1.

Flowchart of methodological framework. The exhaustive methodology used in this study is depicted in the form of a flow chart for the ease of comprehension. It shows the major criteria that we adopted at different stages of the study to investigate the niche characteristics of Argentine ants in various spatiotemporal scales. The arrows in red, green, and yellow indicate the utilization of modeling components such as occurrence records, environmental variables, and accessible areas respectively. Blue arrows indicate the model projections. Filled circles in pink represent the joint utilization of occurrence data and accessible areas
Occurrence data
Occurrence records of the Argentine ant were downloaded from Global biodiversity Information Facility (GBIF; https://doi.org/10.15468/dl.zfzfqy) using the rgbif R package (Chamberlain et al. 2017). The initial dataset contained 12,309 records spanning from 1885 to 2023. We excluded old records from the time range 1885–1980, and retained a total of 11,456 records that fell within 1981–2023 to be used in the subsequent data cleaning process. A five-step procedure was employed to clean the dataset by removing (1) duplicate records, (2) incomplete records (i.e., coordinates missing either longitude or latitude), (3) imprecise records (i.e., coordinates with no decimal places and/or only zeros in decimal places), (4) impossible records (i.e., −180 < longitude > + 180 and/or −90 < latitude > + 90), and (5) unlikely records (i.e., coordinates with zero values for both longitude and latitude). Coordinate cleaning was performed using the scrubr R package (Chamberlain 2016). Oceanic points were removed manually. The cleaned dataset (1981–2023) had 7,996 records of Argentine ants (Fig. 2, Table S1). Two distance filters were used to rarify the records at two different spatial scales: regional (2 km) and global (11 km). The specific thinning distances were identified based on the spatial resolution of the bioclimatic data layers (see below) to minimize issues with spatial autocorrelation (Sillero and Barbosa 2021). Spatial thinning procedure was performed using the spThin R package (Aiello‐Lammens et al. 2015).

Distributional information. The map shows the records that are used for global model calibration and testing. The blue circles indicate the points documented during the period 1981–2010, used for training the models. The red circles are the records observed during the period 2011–2023, used for testing the model performances
Climate data
We downloaded the full suite of 19 fine-scale bioclimatic variables (30 arcsec resolution; ~ 1 km at the equator) from CHELSA database version 2.1 (Karger et al. 2017; available at: https://chelsa-climate.org) for current conditions (1981–2010). We excluded four interactive variables that combine quarterly measurements of temperature and precipitation (Booth 2022): mean temperature of wettest quarter (bio8), mean temperature of driest quarter (bio9), precipitation of warmest quarter (bio18), and precipitation of coldest quarter (bio19) (Escobar et al. 2014). These variables showed discontinuities (abrupt changes) in their spatial interpolation (Booth 2022). From the set of 15 climatic layers, we created distinct matrices of environmental values using distinct sets of occurrence records using the ‘extract’ function of raster R package (Hijmans et al. 2015), specific to global and regional scales. Multifactorial statistical modeling approaches may be affected by overfitting due to multicollinearity within the independent variables (Saupe et al. 2015; Ntotsis and Karagrigoriou 2021). To reduce multicollinearity within the suite of 15 climatic variables, a stepwise algorithm using variance inflation factor (VIF) method was performed on each of these distinct matrices to form subsets of non-collinear environmental variables (Naimi et al. 2014); variables with VIF values > 10 were excluded from the final datasets (Chatterjee and Hadi 2015). Variance inflation factor analysis was implemented using the ‘vifstep’ function of the usdm R package version 2.1 (Naimi et al. 2014).
Climatic data layers for two future periods (2041–2070 and 2071–2100) were sourced from five general circulation models (GCMs) under two shared socio-economic pathways (SSP126 and SSP585) derived from Coupled Model Intercomparison Project Phase 6 (CMIP6) (Eyring et al. 2016). Future climatic layers were downloaded from CHELSA database version 2.1. GCMs used were (1) Geophysical Fluid Dynamics Laboratory's Earth System Model (GFDL-ESM4) (Dunne et al. 2020), (2) Institut Pierre-Simon Laplace Climate Model (IPSL-CM6A-LR) (Boucher et al. 2020), (3) Max Planck Institute Earth System Model (MPI-ESM1.2) (Gutjahr et al. 2019), (4) Meteorological Research Institute Earth System Model (MRI-ESM2) (Yukimoto et al. 2019), and (5) U.K. Earth System Model (UKESM1) (Sellar et al. 2019). The selection of GCMs was based on the full availability of climatic data for both periods in future under two emission scenarios.
Accessible area estimation
Estimation of the geographic areas that are accessible to species, taking into account the dispersal limitations over relevant time periods of the species’ history (area M in the BAM diagram; Soberon and Peterson 2005) is essential for delimiting the regions for model calibration (Barve et al. 2011). The BAM diagram explains how the interactions between abiotic (A), biotic (B), and dispersal/mobility (M) components shape the geographic distribution of a species (Fig. S2; Soberon and Peterson 2005). Delimiting the regions for model calibration based on accessibility has considerable effects on model predictions and model transfers (Machado-Stredel et al. 2021). We used a dispersal-based simulation approach under current climatic conditions (Machado-Stredel et al. 2021) to define individual Ms for native and invaded ranges of Argentine ants. We used region-wise occurrence records thinned at 2 km spatial resolution in conjunction with raw climatic layers of 30 arcsec resolution to construct simple ellipsoid models for the identification of suitability in these simulations. All simulation processes were performed using the grinnell R package (Machado-Stredel et al. 2021). The parameter settings used for the M simulation were provided in the supplementary information S3.
Ecological niche modeling
Maximum entropy modeling approach (Phillips et al. 2006; Maxent) implemented in the kuenm R package (Cobos et al. 2019) was used for modeling ecological niches in geographic space. Minimum volume ellipsoid models (3D ellipsoids or MVE models) to visualize ecological niches in environmental space were developed using the ellipsenm R package (Cobos et al. 2020).
For model calibrations, region-specific occurrence points (native and individual invaded regions) were extracted using selected accessible area polygons using the ‘over’ function of rgeos R package (Bivand et al. 2017). We have provided the details of region-specific occurrence records in table S4. We split the rarified occurrence records into two sets based on two temporal scales: training data (record within 1981–2010; maintaining temporal match between distributional information and climate data) and testing data (records within 2011–2023). However, as an exemption, for testing the performance of native-range model, we depended on traditional random data-partitioning approach for calibration and evaluation, as the observed clustering of records from 2011 to 2023 near calibration localities might cause inflated model performance estimates (Veloz 2009; Hijmans 2012; Boria et al. 2014).
Native-range modeling
For modeling the ecological niches within the native range, we used a random-partitioning approach to split the occurrence records of the native range, fell within the period 1981–2010, into two sets (training and testing data) in a proportion of 50:50. Data partitioning was performed using the caTools R package (Tuszynski and Khachatryan 2013). The parameter settings and the environmental data used for the modeling experiments were provided in table S5. We evaluated: (1) statistical significance of models, (2) performance of models in terms of correctly predicting occurrences in testing data set, and (3) model complexity (Warren and Seifert 2011; Radosavljevic and Anderson 2014). Statistical significance of models was assessed using partial ROC tests (pROC, Peterson et al. 2008). Statistically significant models were filtered using a user-defined omission rate threshold (E = 10%, i.e., models with omission rate > 10% were omitted; Anderson et al. 2003). Models meeting omission rate criteria were sorted by AICc values in ascending order; models that are within the first 2 AICs units of the minimum were selected as final models (Warren and Seifert 2011). From this set, we selected the model with lowest AICc value as our best model. The best model was replicated 10 times using the bootstrap method, and the median output was used for interpreting model predictions (Cobos et al. 2019). This performance evaluation strategy (Cobos et al. 2019) was maintained in all the subsequent maxent modeling experiments.
Invaded range modeling (Maxent) I & II: climate matching
We chose six major invaded ranges of Argentine ants: western United States, southern United States, Europe, southern Africa, Australia, and New Zealand. Our aim was to better understand whether native range climate alone could predict Argentine ants’ invasive potential. We performed two distinct experiments using native-range model, and its corresponding parameter settings. Firstly, we projected the validated native-range model onto each of the selected invaded ranges using three different modes of extrapolation (see the final subsection of methods for details of extrapolation). Secondly, we trained separate models for invaded-ranges using the whole set of occurrence records from the time range 1981–2010 from the corresponding regions, maintaining the same set of variables and parameter settings, as used by the final native-range model. In these experiments, no testing was performed as our focus was solely on visually comparing the pattern of predictions.
3D ellipsoid niche modeling: niche characterization in environmental space
Ecological niches of Argentine ants in both native and invaded ranges were characterized in environmental space using three approaches (Alkishe et al. 2022): (1) a niche overlap test based on 3D ellipsoid models (Nuñez-Penichet et al. 2021), (2) visualization of climatic suitability in three-dimensional environmental space (Nuñez-Penichet et al. 2021), and (3) a niche similarity test using Schoener's D index (Broennimann et al. 2012). The environmental variables deemed as relevant in maxent modeling experiments were used in these experiments (Cuervo et al. 2021; Mathur et al. 2023), as correlative niche models can identify the variables that define the niches and potential distribution of species (Broennimann et al. 2012).
Ellipsoid models were used to perform the pairwise comparison of climatic niches to assess the overlap between the niches of Argentine ants in native and invaded ranges. Two methods were used to measure the niche overlap employing ellipsoids: full overlap and background overlap (Nuñez-Penichet et al. 2021). In the full overlap experiment, the entire set of environmental conditions that overlap between two ellipsoids was considered. To determine how these ellipsoids overlap, full overlap analysis was conducted against the background of 106 points that are uniformly distributed, filling the multidimensional space containing two ellipsoids (Qiao et al. 2016). In background overlap, environmental conditions that exist within the Ms of native and invaded ranges were only considered (Nuñez-Penichet et al. 2021). The Jaccard index (Mammola 2019), calculated as the proportion of the number of points within intersection of two ellipsoids (E1 ∩ E2) to the entire set of points within the union of two ellipsoids (E1 ⋃ E2), was used to measure the overlap. Statistical significance of niche overlap was assessed only for the background overlap method that focuses on environmental conditions within accessible areas (Nuñez-Penichet et al. 2021; Alkishe et al. 2022). Random ellipsoid pairs were created by sampling points from the accessible areas of Argentine ants in native and invaded ranges. This process was repeated 100 times to create a null distribution of overlap values. The null hypothesis in this test is that observed overlap reflects environmental similarity within the M; alternate hypothesis is that environmental conditions within the Ms are divergent (Nuñez-Penichet et al. 2021; Alkishe et al. 2022). The observed value of niche overlap is compared with the null distribution of overlap values; if the observed value of overlap falls within the upper 95% of the null distribution, null hypothesis of niche similarity cannot be rejected (Nuñez-Penichet et al. 2021; Alkishe et al. 2022).
To visualize the climatic suitability in a three-dimensional space, we used three highly relevant variables (i.e., first three environmental variables with a high percent contribution to the final native-range maxent model; bio2, bio4, and bio15) to source environmental information of Argentine ants’ occurrences in both native and invaded ranges. This allowed us to fit three-dimensional minimum volume ellipsoids (Farber and Kadmon 2003; Van Aelst and Rousseeuw 2009; Osorio-Olvera et al. 2020).
The Broennimann’s method of niche similarity assessment (Broennimann et al. 2012) was performed only on those pairs of ranges where M-area environmental similarity was statistically confirmed via elliptical envelope overlaps. Niche similarity test was performed using the ecospat R package (Cola et al. 2017). If the observed D value falls within the upper 95% of the null distribution, the null hypothesis of niche similarity cannot be rejected (Tocchio et al. 2015; Nair and Peterson 2023).
Invaded range modeling (Maxent) III & IV: pooled invaded range models
We tested the effect of combining the distributional information of both native and invaded ranges on predicting the habitat suitability in invaded ranges by performing two distinct modeling experiments: native + invaded ranges (NI) and invaded ranges only (IO). Pooled models enhance the model outcomes in invaded ranges, as species may utilize novel environmental conditions in these regions compared to its native ranges (Broennimann and Guisan 2008). Separate accessible area polygons for NI and IO models were constructed using the ‘merge’ option in QGIS desktop version 3.34.0. The details regarding the parameter settings and environmental variables were provided in table S5.
Global range modeling
Considering the computational challenges associated with using variables at 30 arcsec resolution for global-scale modeling, we aggregated global climatic layers to 5 arcmin resolution (~ 10 km at the equator) using pixel averages (Pillet et al. 2022) by employing the ‘aggregate’ function in the raster R package. In this experiment, we aimed to assess the sufficiency of distributional information associated with the native and the chosen invaded ranges to predict the global invasive potential of Argentine ants. The distributional information used for calibrating native + invaded ranges models (NI) was employed for calibrating the global model, keeping the entire world except Antarctica as the background (Fig. 2). Occurrence points were thinned at 11 km to maintain one record per pixel. The parameter settings and environmental variables used for the global range modeling were provided in table S5. Binary presence-absence maps were created using two minimum training presence thresholds: most desirable (5%) and maximum permitted (10%) (Nair and Peterson 2023) to distinguish suitable regions from unsuitable ones.
Future models
Future climatic layers were aggregated to a resolution of 5 arcmin to match with the resolution of the current global model (Sillero et al. 2021). Final global model was projected individually onto five GCM conditions per period per SSP. We estimated the changes in habitat suitability between current, and two future periods (2041–2070 and 2071–2100) under two SSPs (i.e., 126 and 585). Predictions based on each GCM were compared with the current model, and changes in suitability were represented in three ways: (1) stable, (2) gain, and (3) loss. Final models were presented in three distinct modes of extrapolation (Cobos et al. 2019): (1) free extrapolation (E), (2) extrapolation + clamping (EC), and (3) no extrapolation (NE). Free extrapolation allows the response surface of novel environmental conditions in future scenarios to follow the environmental patterns in the area of calibration (Owens et al. 2013). Under extrapolation with clamping, values of response surface in areas with novel environmental conditions are clamped to the boundary values of environmental space in the calibration region (Phillips et al. 2006). No extrapolation sets the values of response surface to zero in areas where environmental conditions are novel compared to those in calibrated regions (Owens et al. 2013; Cobos et al. 2019).
Results
Preference towards water bodies in native ranges
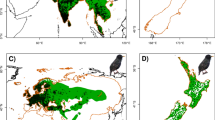
Accessible areas for the native and invaded populations of Argentine ants in their respective ranges were estimated (Fig. S6). The simulated M for native ranges included regions of Parana River basin in Argentina, Uruguay, Brazil, Paraguay, and Bolivia. The stepwise VIF method identified five non-collinear environmental variables: three temperature variables (mean diurnal range = bio2, temperature seasonality = bio4, mean temperature of warmest quarter = bio10) and two precipitation variables (precipitation of wettest month = bio13 and precipitation seasonality = bio15). Native-range modeling experiment developed 490 models, and only one model met our three-step performance evaluation criteria (see above; Cobos et al. 2019). Our best native-range model utilized a relatively higher RM value of 3, only one feature class (q = quadratic), and maximum amount of climatic information (set 7: bio2, bio4, bio10, bio13, and bio15) (Table S7). High habitat suitability for Argentine ants was predicted near water areas in native-range model (Fig. 3). All distinct patches of high suitability were in close proximity to water bodies (rivers, wetlands, and coastal areas) within Argentina, Uruguay, Brazil, and Paraguay. Overall, a pattern of West to East increase in suitability was noticed across the native range. Although high predicted suitability was concentrated in Central East, South, and South-East limits of the region, the Mar Chiquita Lake in Argentina, lying close to the western limit was predicted to be the only potentially suitable area in the West of native range.

Native range model. The map shows the potential distribution of Argentine ants in their native ranges. It also reveals the large preference of Argentine ants towards permanent streams in their native habitats
Native range climatic conditions are insufficient to predict the invasion potential
We projected our final native-range model onto six invaded regions (Fig. 4). However, the native-range model failed to predict accurately the known distributional areas in most of the invaded ranges, regardless of the extrapolation modes employed. The West Coast of the United States, hosting one of the largest super colonies, was not predicted as suitable under any of the three modes of extrapolation (Fig. 4). When the response was set to zero for non-analogous climatic conditions (i.e., NE mode) in invaded ranges, extremely low or no suitability was predicted in all invaded regions, although with some exceptions (e.g., Florida in the United States). When free extrapolation was allowed, extreme suitability was noticed throughout the southern ranges for Argentine ants, with the exception of Florida in the United States. A pattern of West to East increase in suitability was noticed in Europe under the mode E; however, overall in EC mode, a South to North increase in suitability was observed. North Island of New Zealand was predicted as suitable for Argentine ants in both E and EC modes. No potentially suitable habitats were predicted in Africa. In Australia, the predicted distribution based on native-range model showed a shift towards North in both modes (E and EC). The models developed using distributional information in invaded ranges, coupled with constraints sourced from native-range model in the form of parameter settings (RM = 3, feature class = q, and climatic variables = bio2, bio4, bio10, bio13, and bio15) were not able to predict the currently known-distributional pattern of the long-established populations of Argentine ants in Europe.

Climate matching models. The insufficiency of native range conditions alone to accurately predict the distribution of Argentine ants in their invaded ranges is shown. The top to bottom row-wise positioning of invaded ranges is in the following order: western USA, southern USA, Europe, southern Africa, Australia, and New Zealand. The columns indicate (i) the native model, (ii) Iall—individually trained invaded range models using the best native model parameters (RF = 3, feature class = q, and climatic variables = bio2, bio4, bio10, bio13, and bio15) and the whole set of corresponding occurrence records, (iii) PE—projections with free extrapolation, (iv) PEC—projections with extrapolation + clamping, and (v) PNE—projections with no extrapolation
Invaded range niches are largely differentiated among themselves with an exception to those in southern hemisphere
The first three variables with high contribution-percentage to the native-range maxent model were mean diurnal range (bio2; 38.7%), precipitation seasonality (bio15; 30.1%), and temperature seasonality (bio4; 16.3%). We fitted minimum volume ellipsoid models to the occurrence data using these highly relevant variables (Mathur et al. 2023) to compare the niches between native and invaded ranges, as well as between invaded ranges, in the environmental space (Fig. 5). The hyper-volume of environmental space in native range was 6.6 × 104 oC2.mm. Among the invaded ranges, Africa (2.05 × 105 °C2.mm) had the largest hyper-volume of environmental space, while New Zealand had the least (9.604 × 103 °C2.mm).

Ellipsoid models. The ecological niches of Argentine ants in their native and invaded ranges are visualized in three dimensional environmental spaces to assess possible niche overlaps. Blue ellipsoid models in rows represent the regions mentioned along the diagonal. Red ellipsoid models in columns indicate the regions mentioned at the top of each column
Full overlap analysis
Full overlap values between native and invaded ranges, as well as between invaded ranges were ranging from 0 to 0.286, indicating low climatic overlap. Overall, the overlap values in the background overlap approach (i.e., comparing conditions within the M areas) varied from 0 to 0.533, revealing a spectrum of low to relatively high climatic overlap within the M areas (Table S8).
Background analysis
In the background overlap approach, native range conditions considerably overlapped with those of Australia (J = 0.533). Among the pairs of invaded ranges, Southern USA and Europe (J = 0.374), and Africa and Australia (J = 0.334) had relatively higher climatic overlapping. When comparing native range conditions with those of invaded ranges, no niche overlap was observed between the niches of the native range and western USA. We failed to reject the null hypothesis of niche overlap, reflecting M-area environmental similarity (Alkishe et al. 2022), between native range with remaining invaded ranges, with the exception of New Zealand (Fig. 5). The observed overlap value between the native range and New Zealand fell below the upper 95% of the null distribution of randomly generated overlap values (Fig. S9). We could not reject the null hypothesis of niche overlap in five cases among the pairs of invaded ranges: (1) southern USA and Africa, (2) Europe and Africa, (3) Africa and Australia, (4) Africa and New Zealand, and (5) Australia and New Zealand, as the observed overlap values fell within the upper 95% of the null distribution (P > 0.05; Fig. S9). Although largely overlapping climatic conditions exist between the M areas of southern USA and Europe, and southern USA and Australia (Fig. 5), niche differences were observed between those regions (Fig. S9). In addition, the rejection of the null hypothesis of niche overlap in the remaining 10 pairs of invaded range comparisons (Fig. 5; Fig. S9) revealed that the niches of Argentine ants in invaded ranges are largely differentiated. Further, niche similarity using Schoener's D index was performed on range pairs for which niche overlap was found to be better than random expectations in ellipsoid niche overlapping experiments. With the exception of Southern USA versus Africa, all other pairs of ranges indicated niche similarity (Fig. S10). We found that among the six invaded regions analyzed (i.e., three in each hemisphere), a southern hemisphere-specific pattern of climatic niche similarity exists between the M areas of the invaded ranges (Africa, Australia, and New Zealand).
Pooled distributional information provides better understanding on invasive potential
Native + invaded range (NI) and invaded ranges only (IO) modeling experiments developed 280 and 210 models respectively. Only one model from each of these experiments met the three-step selection criteria (Table S7). Both NI and IO final models used only two feature classes (l = linear and q = quadratic), low RM values (≤ 0.3), and all available environmental information. Both the NI and IO models predicted a large part of the southern USA as unsuitable for Argentine ants (Fig. 6). A great extent of the long-established ranges of Argentine ants in Europe was predicted as unsuitable in the IO model. The IO model predicted suitability mostly in Iberian Peninsula, whereas the NI model predicted suitability in Iberian Peninsula, North-West Europe, and Mediterranean Europe, revealing a better picture of the invasive potential of Argentine ants in Europe. Another noteworthy difference was the prediction of the South Island of New Zealand as largely unsuitable by the IO model, while the NI model predicted both South and North Islands of New Zealand as largely suitable for Argentine ants. Potential habitats in central Australia, and broader suitability in Tasmania were only predicted by the NI model. Both models predicted the East and North-East ranges of South Africa as potentially suitable for Argentine ants; however, the NI model alone predicted the central ranges of Namibia as suitable for invasion. These results indicate that the IO model was inferior to the NI model in predicting the invasive potential of Argentine ants.

Pooled invaded range models. The top panel indicates the invaded range models (IO) developed using the distributional information from invaded ranges only. The bottom panel represents the invaded range models (NI) developed using the distributional information from native and invaded ranges
Global warming increases suitability in the northern hemisphere
Global modeling experiment developed 280 models. No model met the user-defined omission threshold (< 10%). We accepted a model with an omission rate of 13% as our final global model (Table S7), as omission rate < 15% is considered to be acceptable (Saupe et al. 2014). The global model used only linear features with a relatively low value of RM (0.6). Mean diurnal range, mean temperature of warmest quarter, mean temperature of coldest quarter, and precipitation of wettest month were the variables used by the global model. Almost the entire southern hemisphere was predicted to be suitable for invasion by Argentine ants, with high suitability localized mainly in the West to East corridor of central Africa and the central to northern parts of South America (Fig. 7). Although the South-East Asia, New Zealand, and Australia were largely suitable for Argentine ants, high suitability was predicted mainly along the coastal regions. In the northern hemisphere, large parts of North Asia and North America were predicted as unsuitable or not-very-suitable for Argentine ants. The suitability prediction in Greenland might be an overestimation owing to the prevailing cold conditions with no history of occurrences. Presence-absence maps (Fig. 7), thresholded at E = 5%, revealed a suitability cover of 6.38 × 108 km2; increased threshold (E = 10%) reduced the suitability area by ~ 2% (6.25 × 108 km2), excluding the area of Greenland. In the future scenarios, as time progresses and emission increases, there is a corresponding increase in the gain of suitability in the northern hemisphere, particularly in North America, central Asia, and North Asia (Fig. 8). In contrast, in the southern hemisphere, a trend of losing suitability with no signs of further gain was observed in the future scenarios. The pattern of suitability gain in the northern hemisphere was consistent in all three modes of extrapolation in all scenarios. This clearly indicated that climate change might help the Argentine ants to invade rapidly in the currently non-invaded regions in the northern hemisphere.

Global-scale model. The top panel represents the global potential distribution of Argentine ants. The bottom panel shows the thresholded presence-absence map (E = 5% and E = 10%). The estimated suitability for Argentine ants in Greenland could be an overestimation

Future projections. The shifts in the potential distribution of Argentine ants in future periods under different emission scenarios are represented. In all modes of extrapolation, the suitability of Argentine ants is predicted to be increasing in the northern hemisphere with advancement in time. The highest loss of suitability in the southern hemisphere is observed to be in 2071–2100 under the SSP585 scenario, particularly when no extrapolation was allowed (bottom-right corner)
Discussion
This study primarily focused on the native and invaded ranges of Argentine ants to understand the ecological niche characteristics in both geographic and environmental spaces. Previous studies (see introduction for details), attempted to characterize the ecological niches of Argentine ants, overlooked the potential need to avoid the temporal mismatch between the time range of species occurrence records and climate data (Roubicek et al. 2010; Zurell and Engler 2019). The reliability of ecological niche models, trained with data points whose time range does not necessarily match with climatic baselines, is questionable (Roubicek et al. 2010). Therefore, our modeling efforts maintained the temporal match (1981–2010) between the training points and climatic layers.
The previous attempt (Roura-Pascual et al. 2006) to characterize the ecological niches of Argentine ants in their native range alone, did not include the climatic variables. In the case of Argentine ants, inclusion of climatic variables is essential, as temperature affects the brood development and the rate of survival at different developmental stages (Abril et al. 2010). High temperatures cause detrimental effects to the eggs, while low temperatures hamper the larval development (Abril et al. 2010). Precipitation is another important abiotic factor, as the abundance of Argentine ants is directly proportional to the levels of soil moisture content (Walters and Mackay 2003; Menke and Holway 2006; Heller et al. 2008). Despite the non-inclusion of climatic data, no explicit hypothesis for the accessible area (area M; Soberon and Peterson 2005) was stated by Roura-Pascual et al. (2006) for calibrating the native range model. In comparison to the most conservative delimitation of native range (Wild 2004), our simulated M-area for the native range covered slightly wider areas that are accessible to the natural populations through dispersal (Barve et al. 2011; Machado-Stredel et al. 2021). High habitat suitability, predicted largely along the river basins in the native ranges, indicates that the presence of permanent stream flow would be one of the major determinants of Argentine ant’s survival and colonizing ability not only in the introduced ranges (Holway 1998; Walters and Mackay 2003) but in the native ranges also (Wild 2004). However, in the North-West region of Argentina of the native range, no clear habitat preferences along the permanent streams (Bermejo and Pilcomayo rivers) were observed. A possible reason could be the substantial decrease of rainfall in the western margin along the longitudinal band of 60-65o W (Depetris 2007). The eastern region of the native habitat experiences greater precipitation compared to the western counterpart (Depetris 2007). The predicted high suitability in the eastern area aligns with the Argentine ants' documented preference for regions with abundant rainfall (Heller et al. 2008).
The Argentine ants form large supercolonies owing to their extraordinary social organization (Giraud et al. 2002). The long established major supercolonies of Argentine ants include Australian (Walters 2006; Rowles and O’Dowd 2007; Lach and Thomas 2008; Suhr et al. 2011), Californian (Van Wilgenburg et al. 2010; Gordon and Heller 2014; Menke and Holway 2020), European (Giraud et al. 2002), South African (Luruli 2007; Mothapo and Wossler 2014, 2016), and New Zealand colonies (Corin et al. 2007; Ward 2009). The colonies of Argentine ants in the southern USA are distributed in small patches within the urban areas (Buczkowski et al. 2004). The inadequacy of native-range climates to correctly predict the potential distribution of Argentine ants in their invaded ranges is best revealed by the omission of the West coast of USA, along the Californian coast, where the long-established distribution of Argentine ants (Californian large supercolony) exists. Although the native model projections largely covered the Mediterranean coastal regions in the E and EC modes, the currently occupied habitats of the supercolonies of Argentine ants along the Atlantic coasts of southern Europe (Giraud et al. 2002; Charrier et al. 2020; Castro-Cobo et al. 2021) were not correctly predicted in any of the projections. Majority of the supercolonies of Argentine ants in New Zealand are distributed in the North of the North Island (Corin et al. 2007; Ward et al. 2010; Ward 2009); in our study, all models confirmed the habitat suitability of this region, with the exception of the model developed under NE mode. However, none of these models predicted the current known-distribution of Argentine ants along the South-East coastal regions of the South Island (Fig. 2). In Australia, the Melbourne-Adelaide-Perth near-coast distribution of Argentine ants, spanning ~ 2700 km (Suhr et al. 2011), was largely predicted by the Iall model. In South Africa, contrary to our expectation, no suitability was predicted in the Western Cape, where this species is abundant (Mothapo and Wossler 2011). However, all the native model projections almost entirely omitted these regions. These results support the previous findings that the native model projections are not always dependable in correctly predicting the species’ potential distribution in the invaded ranges (Broennimann and Guisan 2008; Sales et al. 2021; Nguyen and Leung 2022).
Ecological niches are not only linked to the geographic space but to the environmental space also (Qiao et al. 2016), and this dual nature (i.e., Hutchinsonian duality; Colwell and Rangel 2009) has not been properly investigated in most of the previous studies on Argentine ants. The characterization of ecological niches in geographic space alone can introduce biases (Qiao et al. 2016), as a point in geographic space can be represented by only one point in environmental space, but the reverse does not always hold true, because a point in environmental space may be expressed by more than one point in geographic space (Castaneda-Guzman 2022; Nuñez-Penichet et al. 2022; Nair and Peterson 2023). Considering the large inadequacy of native range conditions to accurately predict the long-established geographic distribution of Argentine ants in their invaded ranges, the characterization of niches in the respective environmental spaces has become essential to assess how similar/dissimilar are the niches in native and invaded ranges (Nuñez-Penichet et al. 2021; Issaly et al. 2023). The comparison of niches in environmental space resulted in three notable findings: (1) the ecological niches of native ranges and the western USA (Californian distribution) were not even overlapping in environmental space, (2) the overlapped niches of native range and New Zealand were dissimilar, and (3) the environmental space in the invaded ranges is broader than that in the native ranges, with an exception of Western USA and New Zealand. The first two findings are critical, as the previous study (Roura-Pascual et al. 2006), in which climatic data were excluded, reached the conclusion that Argentine ants’ niches were conserved between the native and invaded ranges. Furthermore, they did not consider the long-established populations in the Australian, South African, and New Zealand ranges for assessing the niche similarities. In our study, we could not find a consistent trend of niche conservatism (Wiens and Graham 2005; Wiens et al. 2010; Peterson 2011), existing between the native and invaded ranges, at least with respect to the climatic data. The absence of niche conservatism, and niche differentiation among the invaded ranges, do not allow us to reject our hypothesis that the climatic niches of Argentine ants are evolving. The third finding, the larger volume of environmental space utilization by invasive species in their invaded ranges, has been observed in a few studies (Fitzpatrick et al. 2007; Loo et al. 2007; Kearney et al. 2008; Issaly et al. 2023). The variation in soil temperature is limited in the lower bounds of the southern hemisphere, relative to that in the northern hemisphere (Nigam and DeWeaver 2015). The established invaded distributions in the lower bounds of the southern hemisphere, along with the existing niche similarity between those ranges, indicate that the behavioral stress of Argentine ants, associated with defending against the fluctuations in soil temperatures (Parr and Bishop 2022) by tracking for thermally suitable regions (Pecl et al. 2017), may be lower in the currently invaded ranges of southern hemisphere than that in the northern hemisphere.
The assumption that the invasive species conserve the native climatic niche in its invaded ranges (Peterson 2003, 2011; Broennimann et al. 2007) has still been utilized to predict the invasive potential (Liu et al. 2020). However, there is no consensus among ecologists regarding the applicability of niche conservatism in assessing the potential distribution in invaded ranges, as several studies on invasive species have revealed niche shifts in the invaded ranges (e.g., Broennimann et al. 2007; Christina et al. 2020; Mata et al. 2010; Escobar et al. 2016; Hill et al. 2013; Issaly et al. 2023; Kelly et al. 2023; Tang et al. 2021; Tingley et al. 2014). Hence, we adopted a better consensus approach, which was employed in several previous studies; i.e., calibrating models with combined data from native and invasive ranges (e.g., Beaumont et al. 2009; Broennimann and Guisan 2008; Mainali et al. 2015; Sales et al. 2017, 2021), to assess the invasive potential of Argentine ants. Our efforts to compare the performance of the NI models (i.e., models fitted with pooled data from native and invaded ranges) with that of IO models (i.e., models fitted with pooled data from invaded ranges only) support the relative superiority of NI models (Broennimann and Guisan 2008; Sales et al. 2021) in predicting extent of invasion of Argentine ants. For instance, although both the models omitted the southern USA, the current extent of invasion in Europe and New Zealand is better predicted by the NI models (Figs. 2 and 6). Furthermore, the reported occurrence in Namibia, and the associated forecast of the northward expansion of its range (Espadaler et al. 2004) is indicated only by the NI model.
We developed global-scale ecological niche models to identify areas that are highly suitable for invasion by Argentine ants, considering three factors: (1) almost two-thirds of the efforts for controlling the spread of Argentine ants have failed (Hoffmann et al. 2016), (2) the invasion is still continuing (Charrier et al. 2020; Park et al. 2021; Anastasio et al. 2021), and (3) climatic factors determine the current global distribution of Argentine ants (Roura-Pascual et al. 2011; Bertelsmeier 2021). The overall habitat suitability predicted by the previously developed global-scale models (Roura-Pascual et al. 2004, 2011; Hartley et al. 2006; Bertelsmeier et al. 2015; Jung et al. 2022) were lower compared to our model. The prediction of climatic suitability in the Mediterranean region was consistent across our model and all previous models, with an exception of the most recent model developed by Jung et al. (2022). In their model, high suitability was restricted to eastern parts of the USA, South America, and Australia, and central Africa. South-East Asia was earlier mentioned as vulnerable to invasion (Roura-Pascual et al. 2004); however, those regions were not largely predicted as suitable in any of the other studies. Our model also has indicated the potential suitability of climatic conditions in South-East Asia for Argentine ants. Large suitability in central Africa, predicted by our model, was only in agreement with a few of the previous studies (Roura-Pascual et al. 2004; Jung et al. 2022). In Australia, predictions of all previous models were restricted to the southern and eastern coastal regions where known-distribution occurs (Suarez et al. 2001), while our model indicates a broader suitability. Considering the large suitability indicated in the African continent, South-East Asia, and Australia, we feel that extensive field explorations in these regions may result in the inclusion of more areas under the invaded distributional regime of Argentine ants. In contrast with the previous models, our model indicates the broadest climatic suitability in the southern hemisphere for Argentine ants.
The urban development and environmental space in urban settings share a reciprocal relationship (Kleerekoper et al. 2012). The climate has a significant role in determining the design of cities; in turn, cities influence the regional environmental conditions (Kleerekoper et al. 2012). The urban heat island effect (UHI) explains the phenomenon of increased temperature prevalence in urban settings, compared to other underdeveloped adjacent areas (Heisler and Brazel 2010; Heaviside et al. 2017). Among the different scenarios of shared socioeconomic pathways (SSPs), SSP126 emphasizes less-intensive utilization of non-renewable energy resources and implementation of measures for the prevention of environmental degradation, while SSP585 features a world characterized by competition for rapid industrial growth coupled with intensive exploitation of fossil fuels, with little concern for environmental degradation (O’Neill et al. 2017). By the year 2100, SSP126 predicts an expected increase in temperature by 0.9 °C, while SSP585 forecasts an increase of 3.5 °C from the reference period 1995–2014 (Lee et al. 2021). Considering the above facts, we have taken four potential interconnections between the UHI effect, SSPs, and Argentine ants into the consideration to explain the habitat suitability increase, as predicted in the future models: (1) If invasive ants are capable of adaptation to urban settings, they may expand their range as a response to the climate change (Menke et al. 2011), (2) Argentine ants are highly capable of adaptation to urban settings (Knight and Rust 1990; Rust et al. 2003; Clarke et al. 2008; Sang-Hyun et al. 2014; López-Collar and Cabrero-Sañudo 2021), (3) Urbanization facilitates the range expansion of Argentine ants (Newell and Barber 1913; Suarez et al. 2001; Clarke et al. 2008), and (4) UHI effect may help to maintain the permanence of Argentine ants’ population in invaded urban settings (Borden and Flory 2021; López-Collar and Cabrero-Sañudo 2021). All the above points, in line with the latitudinal gradient of socioeconomic development, i.e., only less than 13% of the total human population lives South of equator, and socioeconomic developments would be higher in higher latitudes (Kummu and Varis 2011), support the large-scale habitat suitability increase in the higher latitudes of northern hemisphere, as observed in the future models. Our future models indicate higher risk for countries located above 45oN, where considerable socioeconomic growth has been happening since 1960 (Kummu and Varis 2011). We could not find a consistent trend of predicted increase in habitat suitability worldwide, as we expected; however, this finding does not allow us to reject our hypothesis of climate change associated habitat suitability increase for Argentine ants in future scenarios. Our finding of northern hemisphere specific patterns of suitability increase in climate change scenarios is not consistent across previous studies. However, a similar trend of predicted increase of suitability for Argentine ants in the northern hemisphere by 2050, with a considerably lesser geographic cover, was noticed in only one study (Roura-Pascual et al. 2004). A recent study predicted no such hemisphere-specific habitat suitability patterns by 2080 (Bertelsmeier et al. 2015).
Conclusion
Major findings of our study provide valuable insights, pertaining to the ecological niches of Argentine ants in their native and invaded ranges: (1) large habitat preference towards permanent streams in the native ranges, (2) native ecological niche is only partly conserved across invaded ranges, (3) invaded range niches are largely differentiated, and (4) global warming induces increase in habitat suitability in the northern hemisphere. In addition, the present study has identified highly vulnerable areas to Argentine ants’ invasion at a global scale, for both present and future conditions. The failure rate of effective implementation of management strategies to control Argentine ants is relatively high (Hoffmann et al. 2016). Hence, the results of this study may provide potential insights for the effective implementation of management strategies.
Data availability
The occurrence data used in the modeling experiments are provided in the electronic supplementary materials. The R scripts used are available upon request.
References
Abril S, Roura-Pascual N, Oliveras J, Gómez C (2009) Assessing the distribution of the Argentine ant using physiological data. Acta Oecologica 35:739–745. https://doi.org/10.1016/j.actao.2009.07.005
Abril S, Oliveras J, Gómez C (2010) Effect of temperature on the development and survival of the Argentine ant Linepithema Humile. J Insect Sci 10:97. https://doi.org/10.1673/031.010.9701
Aiello-Lammens ME, Boria RA, Radosavljevic A et al (2015) spThin: an R package for spatial thinning of species occurrence records for use in ecological niche models. Ecography 38:541–545
Alkishe A, Cobos ME, Osorio-Olvera L, Peterson AT (2022) Ecological niche and potential geographic distributions of Dermacentor marginatus and Dermacentor reticulatus (Acari: Ixodidae) under current and future climate conditions. Web Ecol 22:33–45. https://doi.org/10.5194/we-22-33-2022
Anastasio OE, Mathis KA, Rivera MJ (2021) Impacts of invasive ant-hemipteran interaction, edge effects and habitat complexities on the spatial distribution of ants in citrus orchards. Agric Ecosyst Environ 310:107299. https://doi.org/10.1016/j.agee.2021.107299
Anderson RP, Lew D, Peterson AT (2003) Evaluating predictive models of species’ distributions: criteria for selecting optimal models. Ecol Model 162:211–232. https://doi.org/10.1016/S0304-3800(02)00349-6
Angulo E, Hoffmann BD, Ballesteros-Mejia L et al (2022) Economic costs of invasive alien ants worldwide. Biol Invasions 24:2041–2060. https://doi.org/10.1007/s10530-022-02791-w
Angulo E, Guénard B, Balzani P et al (2024) The Argentine ant, Linepithema humile: natural history, ecology and impact of a successful invader. Entomol Gen 44:41–61
Barve N, Barve V, Jiménez-Valverde A et al (2011) The crucial role of the accessible area in ecological niche modeling and species distribution modeling. Ecol Model 222:1810–1819. https://doi.org/10.1016/j.ecolmodel.2011.02.011
Beaumont LJ, Gallagher RV, Thuiller W et al (2009) Different climatic envelopes among invasive populations may lead to underestimations of current and future biological invasions. Divers Distrib 15:409–420
Beaury EM, Fusco EJ, Jackson MR et al (2020) Incorporating climate change into invasive species management: insights from managers. Biol Invasions 22:233–252. https://doi.org/10.1007/s10530-019-02087-6
Bertelsmeier C (2021) Globalization and the anthropogenic spread of invasive social insects. Curr Opin Insect Sci 46:16–23. https://doi.org/10.1016/j.cois.2021.01.006
Bertelsmeier C, Luque GM, Hoffmann BD, Courchamp F (2015) Worldwide ant invasions under climate change. Biodivers Conserv 24:117–128. https://doi.org/10.1007/s10531-014-0794-3
Bivand R, Rundel C, Pebesma E et al (2017) Package ‘rgeos.’ Compr R Arch Netw CRAN
Booth TH (2022) Checking bioclimatic variables that combine temperature and precipitation data before their use in species distribution models. Austral Ecol 47:1506–1514. https://doi.org/10.1111/aec.13234
Borden JB, Flory SL (2021) Urban evolution of invasive species. Front Ecol Environ 19:184–191. https://doi.org/10.1002/fee.2295
Boria RA, Olson LE, Goodman SM, Anderson RP (2014) Spatial filtering to reduce sampling bias can improve the performance of ecological niche models. Ecol Model 275:73–77. https://doi.org/10.1016/j.ecolmodel.2013.12.012
Boser CL, Hanna C, Faulkner KR et al (2014) Argentine ant management in conservation areas: results of a pilot study. Monogr West North Am Nat 7:518–530. https://doi.org/10.3398/042.007.0140
Boser CL, Hanna C, Holway DA et al (2017) Protocols for Argentine ant eradication in conservation areas. J Appl Entomol 141:540–550. https://doi.org/10.1111/jen.12372
Boucher O, Servonnat J, Albright AL et al (2020) Presentation and evaluation of the IPSL-CM6A-LR climate model. J Adv Model Earth Syst 12:e2019MS002010. https://doi.org/10.1029/2019MS002010
Broennimann O, Guisan A (2008) Predicting current and future biological invasions: both native and invaded ranges matter. Biol Lett 4:585–589. https://doi.org/10.1098/rsbl.2008.0254
Broennimann O, Treier UA, Müller-Schärer H et al (2007) Evidence of climatic niche shift during biological invasion. Ecol Lett 10:701–709. https://doi.org/10.1111/j.1461-0248.2007.01060.x
Broennimann O, Fitzpatrick MC, Pearman PB et al (2012) Measuring ecological niche overlap from occurrence and spatial environmental data. Glob Ecol Biogeogr 21:481–497. https://doi.org/10.1111/j.1466-8238.2011.00698.x
Buczkowski G, Vargo EL, Silverman J (2004) The diminutive supercolony: the Argentine ants of the southeastern United States. Mol Ecol 13:2235–2242. https://doi.org/10.1111/j.1365-294X.2004.02261.x
Castaneda-Guzman M (2022) Modeling species geographic distributions in aquatic ecosystems using a density-based clustering algorithm. Virginia Tech
Castro-Cobo S, Blight O, Espadaler X, Angulo E (2021) Long-term spread of Argentine ant (Hymenoptera: Formicidae) European supercolonies on three Mediterranean islands. Myrmecol News 31:185. https://doi.org/10.25849/myrmecol.news_031:185
Chamberlain S (2016) scrubr: clean biological occurrence records. R Package Version 01(1):162
Chamberlain S, Ram K, Barve V et al (2017) Package ‘rgbif.’ Interface Glob Biodivers Inf Facil ‘API
Charrier NP, Hervet C, Bonsergent C et al (2020) Invasive in the North: new latitudinal record for Argentine ants in Europe. Insectes Soc 67:331–335. https://doi.org/10.1007/s00040-020-00762-9
Chatterjee S, Hadi AS (2015) Regression analysis by example. John Wiley & Sons
Christina M, Limbada F, Atlan A (2020) Climatic niche shift of an invasive shrub (Ulex europaeus): a global scale comparison in native and introduced regions. J Plant Ecol 13:42–50. https://doi.org/10.1093/jpe/rtz041
Clarke KM, Fisher BL, LeBuhn G (2008) The influence of urban park characteristics on ant (Hymenoptera, Formicidae) communities. Urban Ecosyst 11:317–334. https://doi.org/10.1007/s11252-008-0065-8
Cobos ME, Peterson AT (2023) Broad-scale factors shaping the ecological niche and geographic distribution of Spirodela polyrhiza. PLoS ONE 18:e0276951. https://doi.org/10.1371/journal.pone.0276951
Cobos ME, Peterson AT, Barve N, Osorio-Olvera L (2019) kuenm: an R package for detailed development of ecological niche models using Maxent. PeerJ 7:e6281. https://doi.org/10.7717/peerj.6281
Cobos ME, Osorio-Olvera L, Soberón J et al (2020) Ellipsenm: ecological niche’s characterizations using ellipsoids. R package. Available at https://github.com/marlonecobos/ellipsenm
Colwell RK, Rangel TF (2009) Hutchinson’s duality: the once and future niche. In: Proceedings national academy science, 106:19651–19658. https://doi.org/10.1073/pnas.090165010
Cordier JM, Loyola R, Rojas-Soto O, Nori J (2020) Modeling invasive species risk from established populations: insights for management and conservation. Perspect Ecol Conserv 18:132–138. https://doi.org/10.1016/j.pecon.2020.06.001
Corin SE, Abbott KL, Ritchie PA, Lester PJ (2007) Large scale unicoloniality: the population and colony structure of the invasive Argentine ant (Linepithema humile) in New Zealand. Insectes Soc 54:275–282. https://doi.org/10.1007/s00040-007-0942-9
Cuervo PF, Flores FS, Venzal JM, Nava S (2021) Niche divergence among closely related taxa provides insight on evolutionary patterns of ticks. J Biogeogr 48:2865–2876. https://doi.org/10.1111/jbi.14245
da Mata RA, Tidon R, Côrtes LG et al (2010) Invasive and flexible: niche shift in the drosophilid Zaprionus indianus (Insecta, Diptera). Biol Invasions 12:1231–1241. https://doi.org/10.1007/s10530-009-9542-0
De Fina AL (1992) Aptitud agroclimática de la república Argentina. Academia nacional de agronomía y veterinaria, Buenos Aires
Depetris PJ (2007) The parana river under extreme flooding: a hydrological and hydro-geochemical insight. Interciencia 32:656–662
Di Cola V, Broennimann O, Petitpierre B et al (2017) ecospat: an R package to support spatial analyses and modeling of species niches and distributions. Ecography 40:774–787. https://doi.org/10.1111/ecog.02671
Dubos N, Fieldsend TW, Roesch MA et al (2023) Choice of climate data influences predictions for current and future global invasion risks for two Phelsuma geckos. Biol Invasions 25:1–20
Dunne JP, Horowitz LW, Adcroft AJ et al (2020) The GFDL earth system model version 4.1 (GFDL-ESM 4.1): overall coupled model description and simulation characteristics. J Adv Model Earth Syst 12:e20195. https://doi.org/10.1029/2019MS002015
Escobar LE, Lira-Noriega A, Medina-Vogel G, Peterson AT (2014) Potential for spread of the white-nose fungus (Pseudogymnoascus destructans) in the Americas: use of Maxent and NicheA to assure strict model transference. Geospat Health 9:221–229
Escobar LE, Qiao H, Phelps NBD et al (2016) Realized niche shift associated with the Eurasian charophyte Nitellopsis obtusa becoming invasive in North America. Sci Rep 6:29037. https://doi.org/10.1038/srep29037
Espadaler X, Blancafort X, Gómez C (2004) The Argentine ant, Linepithema humile (Mayr, 1868) in Namibia (Hymenoptera, Formicidae). Nouv Rev Entomol 21:42
Eyring V, Bony S, Meehl GA et al (2016) Overview of the coupled model intercomparison project phase 6 (CMIP6) experimental design and organization. Geosci Model Dev 9:1937–1958. https://doi.org/10.5194/gmd-9-1937-2016
Farber O, Kadmon R (2003) Assessment of alternative approaches for bioclimatic modeling with special emphasis on the Mahalanobis distance. Ecol Model 160:115–130. https://doi.org/10.1016/S0304-3800(02)00327-7
Feng X, Park DS, Walker C et al (2019) A checklist for maximizing reproducibility of ecological niche models. Nat Ecol Evol 3:1382–1395. https://doi.org/10.1038/s41559-019-0972-5
Fitzgerald K, Heller N, Gordon DM (2012) Modeling the spread of the Argentine ant into natural areas: habitat suitability and spread from neighboring sites. Ecol Model 247:262–272. https://doi.org/10.1016/j.ecolmodel.2012.07.036
Fitzpatrick MC, Weltzin JF, Sanders NJ, Dunn RR (2007) The biogeography of prediction error: why does the introduced range of the fire ant over-predict its native range? Glob Ecol Biogeogr 16:24–33
Frasconi Wendt C, Nunes A, Lobo Dias S et al (2022) Seed removal decrease by invasive Argentine ants in a high nature value farmland. J Nat Conserv 67:126183. https://doi.org/10.1016/j.jnc.2022.126183
Gimenez LH, Rivera RJ, Brante A (2022) One step ahead of sea anemone invasions with ecological niche modeling: potential distributions and niche dynamics of three successful invasive species. Mar Ecol Prog Ser 690:83–95
Giraud T, Pedersen JS, Keller L (2002) Evolution of supercolonies: the Argentine ants of southern Europe. In: Proceedings national academy science, 99:6075–6079. https://doi.org/10.1073/pnas.092694199
Gómez C, Pons P, Bas JM (2003) Effects of the Argentine ant Linepithema humile on seed dispersal and seedling emergence of Rhamnus alaternus. Ecography 26:532–538. https://doi.org/10.1034/j.1600-0587.2003.03484.x
Gordon DM, Heller NE (2014) The invasive Argentine ant Linepithema humile (Hymenoptera: Formicidae) in northern California reserves: from foraging behavior to local spread. Myrmecol News 19:103–110
Gutjahr O, Putrasahan D, Lohmann K et al (2019) Max Planck institute earth system model (MPI-ESM1.2) for the high-resolution model intercomparison project (HighResMIP). Geosci Model Dev 12:3241–3281. https://doi.org/10.5194/gmd-12-3241-2019
Harris RJ, Barker G (2007) Relative risk of invasive ants (Hymenoptera: Formicidae) establishing in New Zealand. N Z J Zool 34:161–178. https://doi.org/10.1080/03014220709510075
Hartley S, Harris R, Lester PJ (2006) Quantifying uncertainty in the potential distribution of an invasive species: climate and the Argentine ant. Ecol Lett 9:1068–1079
Heap MJ, Culham A (2010) Automated pre-processing strategies for species occurrence data used in biodiversity modelling. In: International conference on knowledge-based and intelligent information and engineering systems. Springer, pp 517–526
Heaviside C, Macintyre H, Vardoulakis S (2017) The urban heat island: implications for health in a changing environment. Curr Environ Health Rep 4:296–305. https://doi.org/10.1007/s40572-017-0150-3
Heisler GM, Brazel AJ (2010) The urban physical environment: temperature and urban heat islands. Urban Ecosystem Ecology. John Wiley & Sons, Ltd, Hoboken, pp 29–56
Heller NE, Sanders NJ, Shors JW, Gordon DM (2008) Rainfall facilitates the spread, and time alters the impact, of the invasive Argentine ant. Oecologia 155:385–395. https://doi.org/10.1007/s00442-007-0911-z
Hijmans RJ (2012) Cross-validation of species distribution models: removing spatial sorting bias and calibration with a null model. Ecology 93:679–688
Hijmans RJ, Van Etten J, Cheng J et al (2015) Package ‘raster.’ R Package 734:473
Hill MP, Chown SL, Hoffmann AA (2013) A predicted niche shift corresponds with increased thermal resistance in an invasive mite, Halotydeus destructor. Glob Ecol Biogeogr 22:942–951. https://doi.org/10.1111/geb.12059
Hoffmann BD, Luque GM, Bellard C et al (2016) Improving invasive ant eradication as a conservation tool: a review. Biol Conserv 198:37–49
Holway DA (1998) Factors governing rate of invasion: a natural experiment using Argentine ants. Oecologia 115:206–212. https://doi.org/10.1007/s004420050509
Issaly EA, Baranzelli MC, Rocamundi N et al (2023) Too much water under the bridge: unraveling the worldwide invasion of the tree tobacco through genetic and ecological approaches. Biol Invasions 26:1–19
Jiménez-Valverde A, Peterson AT, Soberón J et al (2011) Use of niche models in invasive species risk assessments. Biol Invasions 13:2785–2797. https://doi.org/10.1007/s10530-011-9963-4
Jung J-M, Kim S-H, Jung S, Lee W-H (2022) Spatial and climatic analyses for predicting potential distribution of an invasive ant, Linepithema humile (Hymenoptera: Formicidae). Entomol Sci 25:e12527. https://doi.org/10.1111/ens.12527
Karger DN, Conrad O, Böhner J et al (2017) Climatologies at high resolution for the earth’s land surface areas. Sci Data 4:1–20
Kearney M, Phillips BL, Tracy CR et al (2008) Modelling species distributions without using species distributions: the cane toad in Australia under current and future climates. Ecography 31:423–434
Kelly CL, Gordon IJ, Schwarzkopf L et al (2023) Invasive wild deer exhibit environmental niche shifts in Australia: where to from here? Ecol Evol 13:e10251. https://doi.org/10.1002/ece3.10251
Kleerekoper L, van Esch M, Salcedo TB (2012) How to make a city climate-proof, addressing the urban heat island effect. Resour Conserv Recycl 64:30–38. https://doi.org/10.1016/j.resconrec.2011.06.004
Knight RL, Rust M (1990) The urban ants of California with distribution notes of imported species. Southwest Entomol
Krushelnycky PD, Gillespie RG (2008) Compositional and functional stability of arthropod communities in the face of ant invasions. Ecol Appl 18:1547–1562. https://doi.org/10.1890/07-1293.1
Kummu M, Varis O (2011) The world by latitudes: a global analysis of human population, development level and environment across the north–south axis over the past half century. Appl Geogr 31:495–507. https://doi.org/10.1016/j.apgeog.2010.10.009
Lach L, Thomas ML (2008) Invasive ants in Australia: documented and potential ecological consequences. Aust J Entomol 47:275–288. https://doi.org/10.1111/j.1440-6055.2008.00659.x
Lee J-Y, Marotzke J, Bala G et al (2021) Future global climate: scenario-based projections and near-term information. In: Climate change 2021: the physical science basis. contribution of working group i to the sixth assessment report of the intergovernmental panel on climate change. Cambridge University Press, pp 553–672
Li M, Xian X, Zhao H et al (2022) Predicting the potential suitable area of the invasive ant Linepithema humile in China under future climatic scenarios based on optimized MaxEnt. Diversity 14:921
Liang CT, Shiels AB, Haines WP et al (2022) Invasive predators affect community-wide pollinator visitation. Ecol Appl 32:e2522. https://doi.org/10.1002/eap.2522
Liu C, Wolter C, Xian W, Jeschke JM (2020) Most invasive species largely conserve their climatic niche. In: Proceedings national academy science, 117:23643–23651. https://doi.org/10.1073/pnas.2004289117
Lo Parrino E, Falaschi M, Manenti R, Ficetola GF (2023) All that changes is not shift: methodological choices influence niche shift detection in freshwater invasive species. Ecography 2023:e06432. https://doi.org/10.1111/ecog.06432
Loo SE, Nally RM, Lake PS (2007) Forecasting New Zealand mudsnail invasion range: model comparisons using native and invaded ranges. Ecol Appl 17:181–189
López-Collar D, Cabrero-Sañudo FJ (2021) Update on the invasion status of the Argentine ant, Linepithema humile (Mayr, 1868), in Madrid, a large city in the interior of the Iberian Peninsula. J Hymenopt Res 85:161–177
Lowe S, Browne M, Boudjelas S, De Poorter M (2000) 100 of the world’s worst invasive alien species: a selection from the global invasive species database. Invasive Species Specialist Group, Auckland
Luruli NM (2007) Distribution and impact of the Argentine ant, Linepithema humile (Mayr), in South Africa. Stellenbosch University, Stellenbosch
Machado-Stredel F, Cobos ME, Peterson AT (2021) A simulation-based method for selecting calibration areas for ecological niche models and species distribution models. Front Biogeogr. https://doi.org/10.21425/F5FBG48814
Mainali KP, Warren DL, Dhileepan K et al (2015) Projecting future expansion of invasive species: comparing and improving methodologies for species distribution modeling. Glob Change Biol 21:4464–4480. https://doi.org/10.1111/gcb.13038
Mammola S (2019) Assessing similarity of n-dimensional hypervolumes: which metric to use? J Biogeogr 46:2012–2023. https://doi.org/10.1111/jbi.13618
Marcelino VR, Verbruggen H (2015) Ecological niche models of invasive seaweeds. J Phycol 51:606–620. https://doi.org/10.1111/jpy.12322
Masoni A, Frizzi F, Giannini F, Santini G (2020) First record of the Argentine ant, Linepithema humile (Mayr, 1868), in the Tuscan Archipelago (Italy). BioInvasions Rec 9:37–43
Mathur M, Mathur P, Purohit H (2023) Ecological niche modelling of a critically endangered species Commiphora wightii (Arn.) Bhandari using bioclimatic and non-bioclimatic variables. Ecol Process 12:8. https://doi.org/10.1186/s13717-023-00423-2
McGlynn TP, Meineke EK, Bahlai CA et al (2019) Temperature accounts for the biodiversity of a hyperdiverse group of insects in urban Los Angeles. In: Proceedings of the royal society B biology science, 286:20191818. https://doi.org/10.1098/rspb.2019.1818
Menke SB, Holway DA (2006) Abiotic factors control invasion by Argentine ants at the community scale. J Anim Ecol 75:368–376
Menke SB, Holway DA (2020) Historical resurvey indicates no decline in Argentine ant site occupancy in coastal southern California. Biol Invasions 22:1669–1679
Menke SB, Holway DA, Fisher RN, Jetz W (2009) Characterizing and predicting species distributions across environments and scales: Argentine ant occurrences in the eye of the beholder. Glob Ecol Biogeogr 18:50–63. https://doi.org/10.1111/j.1466-8238.2008.00420.x
Menke SB, Guénard B, Sexton JO et al (2011) Urban areas may serve as habitat and corridors for dry-adapted, heat tolerant species; an example from ants. Urban Ecosyst 14:135–163. https://doi.org/10.1007/s11252-010-0150-7
Mothapo NP, Wossler TC (2011) Behavioural and chemical evidence for multiple colonisation of the Argentine ant, Linepithema humile, in the Western Cape. South Africa BMC Ecol 11:6. https://doi.org/10.1186/1472-6785-11-6
Mothapo NP, Wossler TC (2014) Resource competition assays between the African big-headed ant, Pheidole megacephala (Fabricius) and the invasive Argentine ant, Linepithema humile (Mayr): mechanisms of inter-specific displacement. Ecol Entomol 39:501–510. https://doi.org/10.1111/een.12126
Mothapo NP, Wossler TC (2016) “You are not always what you eat”: diet did not override intrinsic nestmate recognition cues in Argentine ants from two supercolonies in South Africa. Afr Zool 51:161–171. https://doi.org/10.1080/15627020.2016.1236670
Naimi B, Hamm NAS, Groen TA et al (2014) Where is positional uncertainty a problem for species distribution modelling? Ecography 37:191–203. https://doi.org/10.1111/j.1600-0587.2013.00205.x
Nair RR, Peterson AT (2023) Mapping the global distribution of invasive pest Drosophila suzukii and parasitoid Leptopilina japonica: implications for biological control. PeerJ 11:e15222
Naughton I, Boser C, Tsutsui ND, Holway DA (2020) Direct evidence of native ant displacement by the Argentine ant in island ecosystems. Biol Invasions 22:681–691. https://doi.org/10.1007/s10530-019-02121-7
Newell W, Barber TC (1913) The Argentine ant. U.S. Department of Agriculture Bureau of Entomology, Washington, DC
Nguyen D, Leung B (2022) How well do species distribution models predict occurrences in exotic ranges? Glob Ecol Biogeogr 31:1051–1065. https://doi.org/10.1111/geb.13482
Nigam S, DeWeaver E (2015) Dynamical meteorology. Stationary waves (orographic and thermally forced). In: North GR, Pyle J, Zhang F (eds) Encyclopedia of atmospheric sciences (Second Edition). Academic Press, Oxford, pp 431–445
Ntotsis K, Karagrigoriou A (2021) The impact of multicollinearity on big data multivariate analysis modeling. Applied modeling techniques and data analysis 1. John Wiley & Sons, Ltd, Hoboken, pp 187–202
Nuñez-Penichet C, Cobos ME, Soberon J (2021) Non-overlapping climatic niches and biogeographic barriers explain disjunct distributions of continental Urania moths. Front Biogeogr. https://doi.org/10.21425/F5FBG52142
Nuñez-Penichet C, Cobos ME, Soberón J et al (2022) Selection of sampling sites for biodiversity inventory: effects of environmental and geographical considerations. Methods Ecol Evol 13:1595–1607. https://doi.org/10.1111/2041-210X.13869
O’Neill BC, Kriegler E, Ebi KL et al (2017) The roads ahead: narratives for shared socioeconomic pathways describing world futures in the 21st century. Glob Environ Change 42:169–180. https://doi.org/10.1016/j.gloenvcha.2015.01.004
Osorio-Olvera L, Yañez-Arenas C, Martínez-Meyer E, Peterson AT (2020) Relationships between population densities and niche-centroid distances in North American birds. Ecol Lett 23:555–564. https://doi.org/10.1111/ele.13453
Ota V, Chang AK (1991) Ant control in Hawaiian drip irrigation systems. Applied myrmecology. CRC Press, Boca Raton
Owens HL, Campbell LP, Dornak LL et al (2013) Constraints on interpretation of ecological niche models by limited environmental ranges on calibration areas. Ecol Model 263:10–18. https://doi.org/10.1016/j.ecolmodel.2013.04.011
Park S-H, Ha Y-H, Kim DE et al (2021) Distribution and mitochondrial DNA tracing of the invasive Argentine ants (Linepithema humile) in South Korea. Entomol Res 51:118–126
Park S-H, Kim I-K, Kim C-J, Choi MB (2023) Settlement and population competition assessments of invasive Argentine ants (Linepithema humile) in South Korea. BioInvasions Rec 12:545–554. https://doi.org/10.3391/bir.2023.12.2.17
Parr CL, Bishop TR (2022) The response of ants to climate change. Glob Change Biol 28:3188–3205. https://doi.org/10.1111/gcb.16140
Pecl GT, Araújo MB, Bell JD et al (2017) Biodiversity redistribution under climate change: impacts on ecosystems and human well-being. Science 355:eaai9214. https://doi.org/10.1126/science.aai9214
Peterson AT (2003) Predicting the geography of species’ invasions via ecological niche modeling. Q Rev Biol 78:419–433. https://doi.org/10.1086/378926
Peterson AT (2011) Ecological niche conservatism: a time-structured review of evidence. J Biogeogr 38:817–827
Peterson AT, Papeş M, Soberón J (2008) Rethinking receiver operating characteristic analysis applications in ecological niche modeling. Ecol Model 213:63–72
Phillips SJ, Anderson RP, Schapire RE (2006) Maximum entropy modeling of species geographic distributions. Ecol Model 190:231–259
Pili AN, Tingley R, Sy EY et al (2020) Niche shifts and environmental non-equilibrium undermine the usefulness of ecological niche models for invasion risk assessments. Sci Rep 10:7972. https://doi.org/10.1038/s41598-020-64568-2
Pillet M, Goettsch B, Merow C et al (2022) Elevated extinction risk of cacti under climate change. Nat Plants 8:366–372
Polaina E, Pärt T, Recio MR (2020) Identifying hotspots of invasive alien terrestrial vertebrates in Europe to assist transboundary prevention and control. Sci Rep 10:11655. https://doi.org/10.1038/s41598-020-68387-3
Qiao H, Peterson AT, Campbell LP et al (2016) NicheA: creating virtual species and ecological niches in multivariate environmental scenarios. Ecography 39:805–813. https://doi.org/10.1111/ecog.01961
Qiao H, Escobar LE, Peterson AT (2017) Accessible areas in ecological niche comparisons of invasive species: recognized but still overlooked. Sci Rep 7:1213. https://doi.org/10.1038/s41598-017-01313-2
Radosavljevic A, Anderson RP (2014) Making better Maxent models of species distributions: complexity, overfitting and evaluation. J Biogeogr 41:629–643
Roubicek AJ, VanDerWal J, Beaumont LJ et al (2010) Does the choice of climate baseline matter in ecological niche modelling? Ecol Model 221:2280–2286. https://doi.org/10.1016/j.ecolmodel.2010.06.021
Roura-Pascual N, Suarez AV, McNyset K et al (2006) Niche differentiation and fine-scale projections for Argentine ants based on remotely sensed data. Ecol Appl 16:1832–1841
Roura-Pascual N, Bas JM, Thuiller W et al (2009a) From introduction to equilibrium: reconstructing the invasive pathways of the Argentine ant in a Mediterranean region. Glob Change Biol 15:2101–2115
Roura-Pascual N, Brotons L, Peterson AT, Thuiller W (2009b) Consensual predictions of potential distributional areas for invasive species: a case study of Argentine ants in the Iberian Peninsula. Biol Invasions 11:1017–1031
Roura-Pascual N, Suarez AV, Gómez C et al (2004) Geographical potential of Argentine ants (Linepithema humile Mayr) in the face of global climate change. In: Proceedings of the royal society of London. Series B: biological sciences, 271:2527–2535. https://doi.org/10.1098/rspb.2004.2898
Roura-Pascual N, Hui C, Ikeda T et al (2011) Relative roles of climatic suitability and anthropogenic influence in determining the pattern of spread in a global invader. In: Proceedings national academy science, 108:220–225. https://doi.org/10.1073/pnas.1011723108
Rowles AD, O’Dowd DJ (2007) Interference competition by Argentine ants displaces native ants: implications for biotic resistance to invasion. Biol Invasions 9:73–85. https://doi.org/10.1007/s10530-006-9009-5
Rust MK, Reierson DA, Klotz JH (2003) Pest Management of Argentine Ants (Hymenoptera: Formicidae)1. J Entomol Sci 38:159–169. https://doi.org/10.18474/0749-8004-38.2.159
Sales LP, Ribeiro BR, Hayward MW et al (2017) Niche conservatism and the invasive potential of the wild boar. J Anim Ecol 86:1214–1223. https://doi.org/10.1111/1365-2656.12721
Sales LP, Reboucas R, Toledo LF (2021) Native range climate is insufficient to predict anuran invasive potential. Biol Invasions 23:2635–2647
Sang-Hyun P, Shingo H, Kazuo O (2014) Long-term impacts of Argentine ant invasion of urban parks in Hiroshima, Japan. J Ecol Environ 37:123–129. https://doi.org/10.5141/ecoenv.2014.015
Saupe EE, Hendricks JR, Townsend Peterson A, Lieberman BS (2014) Climate change and marine molluscs of the western North Atlantic: future prospects and perils. J Biogeogr 41:1352–1366. https://doi.org/10.1111/jbi.12289
Saupe EE, Qiao H, Hendricks JR et al (2015) Niche breadth and geographic range size as determinants of species survival on geological time scales. Glob Ecol Biogeogr 24:1159–1169. https://doi.org/10.1111/geb.12333
Sellar AA, Jones CG, Mulcahy JP et al (2019) UKESM1: description and evaluation of the U.K. earth system model. J Adv Model Earth Syst 11:4513–4558. https://doi.org/10.1029/2019MS001739
Sillero N, Barbosa AM (2021) Common mistakes in ecological niche models. Int J Geogr Inf Sci 35:213–226. https://doi.org/10.1080/13658816.2020.1798968
Sillero N, Arenas-Castro S, Enriquez-Urzelai U et al (2021) Want to model a species niche? a step-by-step guideline on correlative ecological niche modelling. Ecol Model 456:109671. https://doi.org/10.1016/j.ecolmodel.2021.109671
Silva PS, de Koch EB, Arnhold A, Delabie JHC (2022) A review of distribution modeling in ant (Hymenoptera: Formicidae) biogeographic studies. Sociobiology 69:e7775–e7775. https://doi.org/10.13102/sociobiology.v69i4.7775
Silverman J, Brightwell RJ (2008) The Argentine Ant: challenges in managing an invasive unicolonial pest. Annu Rev Entomol 53:231–252. https://doi.org/10.1146/annurev.ento.53.103106.093450
Slimani S, Berrai H, Meridji R et al (2020) New reports of the Argentine ant Linepithema humile (Mayr, 1868) (Hymenoptera: Formicidae) in Algeria. Ukr J Ecol 10:248–252
Soberon J, Peterson AT (2005) Interpretation of models of fundamental ecological niches and species’ distributional areas. Biodivers Inf. https://doi.org/10.17161/bi.v2i0.4
Song J, Won LeeJeong SJ-C et al (2022) Ant occurrence dataset regarding Linepithema humile (Mayr, 1868)(Hymenoptera, Formicidae), first recorded in Busan. South Korea ARPHA Prepr 3:e97545
Stanley MC, Ward DF (2012) Impacts of Argentine ants on invertebrate communities with below-ground consequences. Biodivers Conserv 21:2653–2669. https://doi.org/10.1007/s10531-012-0324-0
Suarez AV, Richmond JQ, Case TJ (2000) Prey selection in horned lizards following the invasion of Argentine ants in southern California. Ecol Appl 10:711–725. https://doi.org/10.1890/1051-0761(2000)010[0711:PSIHLF]2.0.CO;2
Suarez AV, Yeh P, Case TJ (2005) Impacts of Argentine ants on avian nesting success. Insectes Soc 52:378–382. https://doi.org/10.1007/s00040-005-0824-y
Suarez AV, Holway DA, Case TJ (2001) Patterns of spread in biological invasions dominated by long-distance jump dispersal: insights from Argentine ants. In: Proceedings national academy science, 98:1095–1100
Suhr EL, O’Dowd DJ, McKechnie SW, Mackay DA (2011) Genetic structure, behaviour and invasion history of the Argentine ant supercolony in Australia. Evol Appl 4:471–484. https://doi.org/10.1111/j.1752-4571.2010.00161.x
Tang X, Yuan Y, Liu X, Zhang J (2021) Potential range expansion and niche shift of the invasive Hyphantria cunea between native and invasive countries. Ecol Entomol 46:910–925. https://doi.org/10.1111/een.13028
Tingley R, Vallinoto M, Sequeira F, Kearney MR (2014) Realized niche shift during a global biological invasion. In: Proceedings national academy science, 111:10233–10238. https://doi.org/10.1073/pnas.1405766111
Tocchio LJ, Gurgel-Gonçalves R, Escobar LE, Peterson AT (2015) Niche similarities among white-eared opossums (Mammalia, Didelphidae): is ecological niche modelling relevant to setting species limits? Zool Scr 44:1–10. https://doi.org/10.1111/zsc.12082
Tuszynski J, Khachatryan MH (2013) Package ‘caTools’. Available at https://cran.r-project.org/web/packages/caTools/index.html.
Van Aelst S, Rousseeuw P (2009) Minimum volume ellipsoid. Wires Comput Stat 1:71–82. https://doi.org/10.1002/wics.19
Van Wilgenburg E, Torres CW, Tsutsui ND (2010) The global expansion of a single ant supercolony. Evol Appl 3:136–143
Vega SJ, Rust M (2000) The Argentine ant - a significant invasive species in agricultural, urban and natural environments. Sociobiology 37:3–25
Veloz SD (2009) Spatially autocorrelated sampling falsely inflates measures of accuracy for presence-only niche models. J Biogeogr 36:2290–2299
Walters AC (2006) Invasion of Argentine ants (Hymenoptera: Formicidae) in South Australia: impacts on community composition and abundance of invertebrates in urban parklands. Austral Ecol 31:567–576. https://doi.org/10.1111/j.1442-9993.2006.01592.x
Walters AC, Mackay DA (2003) An experimental study of the relative humidity preference and survival of the Argentine ant, Linepithema humile (Hymenoptera, Formicidae): comparisons with a native Iridomyrmex species in South Australia. Insectes Soc 50:355–360. https://doi.org/10.1007/s00040-003-0685-1
Wang A, Melton AE, Soltis DE, Soltis PS (2022) Potential distributional shifts in North America of allelopathic invasive plant species under climate change models. Plant Divers 44:11–19. https://doi.org/10.1016/j.pld.2021.06.010
Ward D, Green C, Harris R et al (2010) Twenty years of Argentine ants in New Zealand: past research and future priorities for applied management. N Z Entomol 33:68–78. https://doi.org/10.1080/00779962.2010.9722193
Ward DF (2009) Potential social, economic and biodiversity impacts of the Argentine ant, Linepithema humile, in the Hawke’s Bay region. Landcare Res Contract Rep LC0809087 Hawke’s Bay Reg Counc Envirolink Proj 700HBRC93
Warren DL, Seifert SN (2011) Ecological niche modeling in Maxent: the importance of model complexity and the performance of model selection criteria. Ecol Appl 21:335–342
Wetterer J, Cirranello A, Hebard E (2001) Impact of the Argentine ant, Linepithema humile on the native ants of Santa Cruz Island, California. Sociobiology 38:709–721
Wetterer J, Wild A, Suarez A et al (2009) Worldwide spread of the Argentine ant, Linepithema humile (Hymenoptera: Formicidae). Myrmecol News 12:187–194
Wiens JJ, Graham CH (2005) Niche conservatism: integrating evolution, ecology, and conservation biology. Annu Rev Ecol Evol Syst 36:519–539
Wiens JJ, Ackerly DD, Allen AP et al (2010) Niche conservatism as an emerging principle in ecology and conservation biology. Ecol Lett 13:1310–1324
Wild AL (2004) Taxonomy and distribution of the Argentine Ant, Linepithema humile (Hymenoptera: Formicidae). Ann Entomol Soc Am 97:1204–1215. https://doi.org/10.1603/0013-8746(2004)097[1204:TADOTA]2.0.CO;2
Yang L, Qian F, Song D-X, Zheng K-J (2016) Research on urban heat-island effect. In: Procedia engineering, 169:11–18. https://doi.org/10.1016/j.proeng.2016.10.002
Yukimoto S, Kawai H, Koshiro T et al (2019) The meteorological research institute earth system model version 2.0, MRI-ESM2.0: description and basic evaluation of the physical component. J Meteorol Soc Jpn Ser II 97:931–965. https://doi.org/10.2151/jmsj.2019-051
Zurell D, Engler JO (2019) Ecological niche modelling. In: Dunn PO, Møller AP (eds) Effects of climate change on birds. Oxford University Press, Oxford, pp 60–73
Funding
This research has not received any funding.
Author information
Authors and Affiliations
Contributions
RRN conceived the research idea. RRN formulated the methodology. RRN and ANS analyzed the data. RRN, DEG, MP, and ANS interpreted the results. RRN and ANS wrote the first draft of manuscript. All authors commented on the previous versions of manuscript. All authors read and approved the final version of manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cite this article
Nair, R.R., Gurvich, D.E., Pereyra, M. et al. Clandestine travelers, a boon for south and a bane for north? Warming-induced shifts in global invasion potential of Argentine ants. Biol Invasions 26, 3369–3392 (2024). https://doi.org/10.1007/s10530-024-03389-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-024-03389-0