
Abstract
The research was carried out to determine the salt stress tolerance status of some almond genotypes considered as rootstock candidates under in vitro conditions. The genotypes were cultured in the MS nutrient medium containing 0, 50, 100 and 150 mM NaCl. Apart from NaCl, 1.0 mg/l BAP, 0.01 mg/l IBA, 30 g/l sucrose and 7 g/l agar were added to the nutrients and the pH was adjusted to 5.7. In vitro shoots were incubated for 4 weeks in a climate room with 16‑h light and 24 °C temperature, and then the number of shoots per explant, the proline, chlorophyll, total phenolics, total flavonoids and total protein contents, superoxide dismutase, CAT and APX enzyme activities were evaluated. In parallel with the increase in salt stress level, it was determined that the number of shoots and chlorophyll contents decreased significantly in all genotypes as compared to the control treatment. The proline, total phenolic, total flavonoid and total protein contents and enzyme activities increased significantly with the increase in the salt level. In the study, no significant difference was observed regarding the tolerance status of the genotypes in the MS medium containing 50 mM NaCl. Considering the 100 and 150 mM NaCl applications, it was determined that the genotypes numbered 9, 29, 54, 120, 134, 183, 185, 196 and 241 showed better development and therefore they stood out in terms of salt tolerance as compared to the other genotypes.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
In recent years, serious productivity losses have been observed in agricultural production due to the problems experienced in the water resources and regional distribution as a result of global warming. Therefore, the breeding rootstocks resistant to the abiotic and biotic stress factors increase its importance day by day, and thus very large areas become suitable for production (Oluk and Sami 2007). In order to obtain high yields from fruit species, the species should be grown under suitable ecological conditions. However, the optimum conditions cannot always be fully provided for every fruit type (Köşkeroğlu 2006).
Salinity appears to be a stress factor that significantly limits the growth and development, as the various physiological and biochemical processes occurring in plants are negatively affected due to its excessive amount in the growth environment (Fu et al. 2013). Salts that cause stress in plants are in different forms and concentrations such as chlorides, sulfates, nitrates, borates, carbonates and bicarbonates, and NaCl is considered to be the salt with the highest toxic effect, especially due to its very high solubility (Dal et al. 2001; Ahmed et al. 2008). The excessive amounts of Na and Cl in the root zone increase the osmotic potential, thus causing physiological drought by preventing the plants from sufficiently using the water in the soil (Aziz and Khan 2001; Doğru and Canavar 2020).
Moreover, the excess Na and Cl in the root zone negatively affect the K and NO3 uptakes respectively and thus negatively affect the nitrogen and carbon metabolisms of plants by destructing the ion balance (Daşgan et al. 2006; Gupta and Huang 2014; Tilkat et al. 2017). As a result of this, the impairment of hormonal balance in plants, the decrease in photosynthesis capacity, the regression in root growth, the decline in protein synthesis as a result of decreased nitrate intake, the shortening of plant height, the small leaves and fruits, the fertilization disorders, and ultimately the low quality and yield may occur. The resistance of plants to salt varies according to their developmental stages, and the genetic characteristics of the rootstock and the variety used (Dölarslan and Gül 2012; Akça and Samsunlu 2012; Çelik and Eraslan 2015; Aras and Eşitken 2018). Salt stress also causes oxidative damage by inducing the formation of reactive oxygen species such as superoxide (O2−), hydrogen peroxide (H2O2) and hydroxyl radicals (HO) by creating a water deficiency in cells. In order to protect themselves from the toxic effects of salt stress, plants activate the enzymatic defense system such as superoxide dismutase (SOD), ascorbate peroxidase (APOD), monodehydro ascorbate reductase (MDHAR), dehydroascorbate reductase (DHAR), glutathione reductase (GR) and catalase (CAT) or they activate an effective defense system consisting of non-enzymatic components such as acid, glutathione, α‑tocopherol and carotenoids (Dat et al. 2000; Edrewa 2005; Yılmaz et al. 2020; Li et al. 2016; Doğru and Canavar 2020). The resistance to the stress conditions in plants is a feature directly related to their genetic structure. In this respect, the most effective method in obtaining the genotypes resistant to the stress conditions in fruit species is the method of selecting resistant genotypes from existing populations (Shalaby et al. 1993). Most of the other stone fruit types, including almonds, are generally sensitive to salt stress, and the salinity values above 1.50 dS/m cause a decrease in their productivity, and if it is above 4 dS/m, the yield decreases by approximately 50%. Therefore, the salt-sensitivity is among the most important breeding criteria in the salt-sensitive species such as almonds. For this reason, determining more salt-tolerant almond genotypes will offer important advantages to growers in terms of fruit quality and productivity. Therefore, the objective of this study was to determine the salt stress tolerance levels of some almond rootstocks under in vitro conditions.
Materials and Methods
In the study, promising rootstock candidates selected from Isparta region of Turkey by Yıldırım et al. (2007) were used as material. For this purpose, a few drops of tween 20 were added to the shoot tips taken at the beginning of vegetation, and they were disinfected by shaking in the 18% commercial sodium hypochlorite solution for 20 min. Then, the shoot tips were washed three times for 5 min with sterile distilled water. In the study, the shoot tips were cultured in the MS medium containing 1.00 mg/l BAP and 0.01 mg/l IBA (Channuntapipat et al. 2003). A total of 30 g/l sucrose and 7.00 g/l agar were added to the nutrient medium and the pH was adjusted to 5.70. To be used in the salt stress tolerance studies, it was subcultured four times in the MS medium containing the same growth regulator combination until a sufficient number of shoots were obtained.
In the salt stress experiments, NaCl was added at 0, 50, 100 and 150 mM doses in addition to the propagation medium. After the cultures were grown in the climate chamber for 4 weeks, the genotypes were examined in terms of their responses to the stress conditions. The condition of climate chamber was 25 ± 1 °C with a 16‑h photoperiod under cool white fluorescent light and the light intensity of inside was set to 140 ± 10 mmol/m2s. After the incubation, the genotypes were evaluated in terms of the total number of shoots per explant. In addition, the biochemical analyses such as the total proline, chlorophyll, phenolic, flavonoid and protein contents, and superoxide dismutase, CAT and APX enzyme activities of the shoots were determined.
Analysis Performed in Stress Applications
The Determination of Proline Content
The proline amounts of the samples obtained from in vitro conditions, were determined as described by Liu et al. (2012). A total of 0.1 g of sample was taken and homogenized in 2 ml of 3% sulfosalicylic acid. In all, 200 µl of the prepared plant extraction was taken and the same amounts of ninhydrin and glacial acetic acid were added on it, and then it was incubated in a water bath for 1 h at 100 °C. Then, its activation was stopped on ice. After this mixture was extracted with 1 ml of toluene, the absorbance of the toluene fraction aspirated from the liquid phase was read in the spectrophotometer at 520 nm. Proline concentration was calculated as μmol proline g−1 fresh weight with the help of calibration curve.
Determination of the Chlorophyll Content
In order to determine the chlorophyll content, 0.1 g of sample was weighed and homogeneously disintegrated in 100% DMF. The homogenate obtained was centrifuged at 10,000 g for 10 min. The absorbances of the upper phases (liquid phase) taken after centrifugation were measured at 664 and 647 nm wavelengths and the amounts of chlorophyll a, chlorophyll b and chlorophyll a + b were calculated with the help of the following formulas (Sibley et al. 1996; Inskeep and Bloom 1985; Aono et al. 1993).
Determination of the Total Phenolic Content
A 0.2‑g leaf sample was added to 10 ml of 80% methanol and crushed with a homogenizer and then mixed in a shaker incubator for 15 min at room temperature. Then, after being centrifuged at 4000 × rpm for 10 min, the liquid part was separated, and after adding 80% methanol on the solid part again, the same processes were repeated. The final volume is completed to 25 ml with 80% methanol. The total amounts of phenolic compounds in the leaf samples extracted with methanol were determined using Folin-Ciocalteu colorimetric method as described by Singleton and Rossi (1965). The total amount of phenolic compounds was determined as mg g−1 wet weight in terms of gallic acid by making measurements at a wavelength of 765 nm in the spectrophotometer and using the curves prepared with standard gallic acid solution.
Total Amount of Flavonoid Substances
Total amount of flovanoid substances was determined using the method described by Zhishen et al. (1999). For this purpose, 0.2 g of sample was homogenized in 10 ml of 80% methanol with a homogenizer and mixed for 15 min at room temperature in a shaking incubator. Afterwards, it was centrifuged at 4000 × rpm for 10 min, the supernatant part was separated and 80% methanol was added on the pellet again and the same process was repeated. The final volume was completed to 25 ml with 80% methanol. A total of 1.5 ml of distilled water and 75 µL of 5% sodium nitrite solution were added onto 0.25 ml of methanol extract and incubated at room temperature for 6 min. After incubation, 0.15 ml of 10% AlCl3 was added to the mixture and re-incubated for 5 min. Then, 0.5 ml of 1M NaOH was added and the absorbance values were read at 510 nm wavelength. The results were calculated according to the catechin standard and expressed as mg/g.
Total Soluble Protein Content
The total amount of protein was determined using the method described by Hartree-Lowry (1972). In all, 5 ml of cold EtOH was added to 1 g of sample and it was disintegrated with the help of a homogenizer. Subsequently, the samples were centrifuged at 10,000 g at 4 °C for 20 min and the liquid part was removed. Then, 8.333 ml of 80% cold EtOH was added on the remaining solid part and the solid part was thoroughly dissolved. The liquid part was then removed by centrifugation under the same conditions. The pellet was dissolved by adding 5 ml of protein extraction buffer (50 mM Tris + 1.2 M NaCl Ph: 7.0) on the solid part and incubated on ice for 30 min. Then it was centrifuged under the same conditions and the supernatant part was filtered with miracloth. Following this, 1 ml of protein extract was added to 0.9 ml of reagent A and vortexed. It was incubated at 50 °C for 10 min and then cooled at the room temperature. Then 0.1 ml reagent B was added to the tubes and mixed. It was again incubated at the room temperature for 10 min. After incubation, 3 ml reagent C was quickly added to the tubes and mixed. After incubation at 50 °C for 10 min, it was cooled to room temperature. Finally, the absorbance values of the samples were read at 650 nm wavelength. The results were calculated according to the BSA standardization and expressed as mg/ml.
Determination of the Superoxide Dismutase (SOD) Enzyme Activity
Superoxide dismutase (SOD) enzyme activity was performed according to the method reported by Constantine and Stanley (1977). Accordingly, 3 ml reaction mixture containing 50 mM potassium phosphate solution (pH: 7.3), 13 mM L-methionine, 75 µM Nitroblue Tetrazolium (NBT), 0.1 mM EDTA, 4 µM riboflavin and 0.25 ml enzyme extract, was kept under 48 µmol photons m−2 s−1 light intensity for 10 min and the absorbance values were measured at 560 nm in the spectrophotometer. Since one unit of SOD activity is defined as the amount of enzyme required to inhibit 50% of the photoreduction of NBT in the presence of riboflavin and light, SOD activity was determined accordingly and the unit was evaluated as mg protein−1.
Determination of the Catalase (CAT) Enzyme Activity
CAT enzyme activity was determined as described by Beers and Sizer (1952). The enzyme activity was determined by the method of determining the decrease in the absorbance of H2O2 at 240 nm by spectrophotometer. For this purpose, 3 ml of the prepared reaction mixture contained 50 mM potassium phosphate solution (pH: 7.0), 15 mM H2O2 and 50 µl enzyme extract. The reaction was started by the addition of the enzyme. The CAT activity was calculated using the extinction coefficient (ε:39.4 mM−1 cm−1) and expressed as µm min−1 mg protein−1.
Determination of Ascorbate Peroxidase (APX) Enzyme Activity
APX enzyme activity was determined using the method described by Nakano and Asada (1981). The samples (4 g) were homogenized in 12 ml of 50 mM potassium phosphate buffer (Ph: 7.3) containing 1 mM EDTA, 2 mM DTT and 1 mM ascorbic acid, and centrifuged at 10,000 g for 15 min at 4 °C. After centrifugation, the supernatant portion was used for the analysis. In all, 0.1 ml of the enzyme extract was added to the 0.9 ml 0.05 M sodium phosphate buffer (0.5 mM ascorbate, 0.1 mM Na2EDTA and 1.2 mM H2O2) (pH:7.0), and the absorbance values were immediately read at 470 nm wavelength. Then, the samples were kept for 3 min and their absorbances were read again at the same wavelength. The results were expressed as mol/min/g protein.
Statistical Analyses
A completely randomized design was used in an in vitro drought stress experiment with six replications per treatment and five plants per replication. The data were subjected to the analyses of variance (ANOVA) using MINITAB statistical software (MINITAB LTD., Coventry, UK). Means were separated by Tukey’s multiple range test at p ≤ 0.05.
Results and Discussion
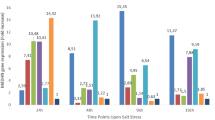
The effects of salt stress applications performed on the 13 different genotypes on the number of shoots and chlorophyll contents in the study are presented in Table 1. In the study, it was determined that the number of shoots in all genotypes decreased significantly as compared to the control in parallel with the increase in the salt stress level. In addition, it was determined that the 50 mM NaCl application had no significant effect on the total shoot number in some genotypes (29, 120, 163, 183, 228 and 241) as compared to the control. In the study, it was determined that the genotypes 9, 54, 120, 134 and 183 were more tolerant to the salt treatment (100 mM NaCl application) as compared to the other genotypes in terms of the shoot numbers. However, it was determined that the other genotypes, except for the genotype 196, did not give shoots and the plants were damaged in 150 mM NaCl application. It was determined that the total chlorophyll contents of the genotypes decreased by 4.35 to 60.07% in 50 mM NaCl application and 17.90 to 68.13% in 100 mM NaCl application. Moreover, the application of 150 mM NaCl caused 60.37% decrease in the chlorophyll content of the genotype 196. The effects of salt stress applications on the total flovonoid, proline, total phenolic and total protein contents performed in 13 different genotypes in the study are presented in Table 2. In all genotypes, it was found that the total flavonoid content increased with increasing doses of NaCl applications. These increases were prominent in the genotypes of 129 (31.03%), 185 (47.50%) and 241 (42.55%) in the 50 mM NaCl application as compared to the control, while in the 100 mM NaCl application the increases were higher in the 40 (101.00%), 134 (50.00%), 163 (54.35%), 185 (77.5%) and 241 (57.45%) genotypes respectively. Moreover, the increase in the content of flavonoids in the application of 150 mM NaCl in the genotype 196 was 56.71%. In parallel to the increase in the amount of salt application in the study, very high increases in the amount of proline in all genotypes were found. Proline contents in the genotypes varied between 9.77% (genotype 54) and 279.48% (genotype 9) in the 50 mM NaCl application, and between 49.22% (genotype 54) and 368.60% (genotype 185) in the 100 mM NaCl application. On the other hand, this increase was 272.58% in the genotype 196 in response to 150 mM NaCl application. In the study, the increases in the total phenolic substance contents were detected in all genotypes in response to the increasing salt concentrations, and these increases were observed at higher rates in the genotypes 134 (190.91%) and 185 (75.00%) in 50 mM NaCl application as compared to the other genotypes. The increases in the total phenolic substance amounts continued in 100 mM NaCl application, and these increases reached to their highest levels in the genotypes 9 (119.15%), 134 (245.45%), 163 (107.02%) and 185 (103.95%). In addition, the increase in the amount of total phenolic substances continued in the application of 150 mM NaCl in the genotype 196, and it was found to be 90.65%. In the study, the protein contents of the genotypes varied between 0.29 mg/ml (genotype 9) and 0.68 mg/ml (genotype 183) in the control application. In parallel with the increase in the salt amount, the increases in protein contents were detected in all genotypes except for the genotype 134, and the highest increases were recorded in the 29 (297.37%) and 129 genotypes (208.77%). However, it was determined that 100 mM NaCl application decreased the protein content in the genotype 134 as compared to the control. In addition, the increase in the protein amount continued in the treatment of 150 mM NaCl in the genotype 196, and the increase was determined as 108.70%. The effects of salt stress applications on the APX, Catalase and SOD enzyme activities performed in 13 different genotypes are presented in Table 3. In the research, in parallel with the increase in salt applications, the increases in the enzyme activities occurred in all genotypes. Especially, the APX enzyme activity was 40 times, 14 times and 50 times higher in the genotypes 9, 134 and 185, respectively, in 100 mM NaCl application. The catalase enzyme activity was more than two-fold higher in the genotypes 9 and 120 as compared to the control in the 100 mM NaCl application, and was more than five times higher in the genotype 163. On the other hand, it was determined that the SOD enzyme activity increased four-fold, five-fold, and more than six-fold, in the genotypes 29, 120 and 241, respectively, as compared to the control in the 100 mM NaCl application.
In some abiotic stress conditions such as salinity, the physiological and metabolic changes that occur in the plants prevent them from being damaged by stress conditions or help them to maintain their vitality by being less damaged (Öztürk 2015). However, plants that are tolerant to the stress conditions can maintain their osmotic balance by operating their protective mechanisms (such as color pigments, increase in enzyme activity, phenolic substance synthesis, proline synthesis and some morphological changes) under stress conditions and maintain their vitality during stress conditions (Meloni et al. 2003; Candan and Tarhan 2003; Demiral and Türkan 2006; De Britto et al. 2011; Caverzan et al. 2012; Turfan 2016).
In fruit growing, it is extremely important to know the tolerance status and limits of rootstocks in order to eliminate the negative effects of stress factors such as salt on the development. Stress resistance mechanism emerges as a feature directly related to the genetic structure of the plant. In this respect, it is the most economical and most effective method to obtain stress resistant types from existing populations (Shalaby et al. 1993; Paschke et al. 2005).
In the study, it was determined that the number of shoots decreased as the salt concentration added to the nutrient medium increased. It was determined that only the genotype 196 formed new shoots in a nutrient medium containing 150 mM NaCl. In addition, it was determined that the genotypes 9, 29, 54, 120, 134, 183, 185 and 241 could continue their development by forming new shoots in the nutrient medium containing 100 mM NaCl. Therefore, it is possible to say that these genotypes are more tolerant to salt than the others. Shıyab et al. (2003) examined the effects of 0, 50, 100, 150, 200 and 300 mM NaCl applications on the development of citrus rootstocks under in vitro conditions. The researchers found that the shoot growth decreased with the increase in salt concentration in 0 and 150 mM NaCl applications, and the plants died completely in 200 and 300 mM NaCl applications. Similarly, Shibli et al. (2003) examined the shoot formation in the nutrient media containing 0, 50, 75 and 100 mM NaCl under in vitro conditions, and they stated that the shoot length decreased with the increasing application doses and that some almond genotypes studied were salt tolerant. Najafian et al. (2008) applied 0, 15, 30, 45, 60 and 75 mM NaCl doses in order to determine the responses of two almond genotypes to salt stress, and they determined that the stem length and number of nodes decreased in both rootstocks with the increase in salt concentration, but the chlorophyll content did not change. Different doses of NaCl were applied to the GiselA 5 rootstock under in vitro conditions, and it was stated that the growth retardation caused by salinity and the decreases in chlorophyll amount occurred in shoots (Ertürk et al. 2007). Similarly, the GF 677 and Nemared rootstocks were cultured in the MS medium containing 0, 5, 10, 15, 20, 40 and 80 mM KCl doses under in vitro conditions, and it was determined that the number of shoots was not affected in the 0–40 mM KCl applications, there were more shoots of the Nemared rootstock than the GF 677 rootstock, and there was a decrease in the development of both rootstocks in response to the 80 mM KCl treatment (Sotiropoulos et al. 2006). It has also been stated in the other studies that with the increase of NaCl concentrations in salt stress applications, there were significant decreases in the number of shoots and shoot development, and plant deaths were observed in very high doses (Okubo and Sakuratani 2000; Bolat et al. 2006; Zrig et al. 2011; Feng et al. 2017). With the increase in the salt concentrations in our study, there was a decrease in the chlorophyll content, as reported by Ranjbarfordoei et al. (2006), who have determined the similar results in the research conducted on P. dulcis almond type. In addition, the decrease in the chlorophyll content in the leaves with the increase of NaCl and KCl doses in the salt stress applications performed on different fruit species has also been reported by the other researchers (Bolat et al. 2006; Ertürk et al. 2007; Zrig et al. 2011; Akçay and Eşitken 2017).
Phenolic compounds (such as flavonoid and phenolics), which show strong antioxidant properties by binding reactive oxygen radicals to themselves, constitute the most important secondary metabolite products of plant metabolism (Çetin et al. 2012; Berli et al. 2010; Halliwell 2008). It has been stated that these substances play a role in the elimination of free oxygen radicals. In addition, similar to our study, it has been reported by many researchers that there are increases in the amount of substances such as total flavonoids and total phenolics synthesized in plant tissues under stress conditions (Rezazadeh et al. 2019; Valifard et al. 2014; Rebey et al. 2017).
In the study, it was found that the CAT enzyme activities increased in all genotypes with the increase in salt concentration, but these increases were higher in the genotypes 9, 120, 134, 163 and 185. However, it has been observed that the CAT enzyme activity has not increased much with the salt concentration in the genotype 196, which has given good results in terms of the shoot development. Therefore, this result leads to the conclusion that the increase in CAT enzyme activity in the almond genotypes under the salt stress conditions may not be an indicator of the resistance. In the salt stress applications, there are the research results indicating both the increases (Ertürk et al. 2007; Sivritepe et al. 2008; Bolat et al. 2014) and the decreases (Sorkheh et al. 2012) in the CAT enzyme activity in response to the increases in the salt concentrations.
Zrig et al. (2011) have stated that the responses of plants to the stress conditions can vary according to the species and they develop different resistance mechanisms. Similarly, it was determined that the amount of SOD increased significantly in all genotypes with the increase of salt concentration, and the increase was higher in the genotypes 29, 120 and 241 as compared to the other genotypes. However, in some genotypes (129 and 176), there was no significant increase in the SOD enzyme activity in parallel with the increase in the salt concentration. Meloni et al. (2003) obtained similar results in their study and stated that the change in the SOD enzyme activity varied according to the cultivars with the increase in salt concentration. It has been emphasized in some studies that the SOD enzyme activity in plants increases under stress conditions and eliminates superoxide and hydrogen peroxide radicals in cells (Scandalios 1996; Zhu et al. 2004; Yıldız et al. 2010). In the study, it was found that the proline content increased significantly in all genotypes with the increase in salt concentration, and this increase was five-fold higher than the control in the genotypes including 9, 40, 120, 134, 185 and 196.
In the study, although there was a regular increase in the total amount of phenolic substances together with the salt concentration, it was found that this increase was not as high as the increases in the CAT, proline and SOD enzyme activities. It is known that proline is one of the compounds whose synthesis increases the most when plants are exposed to the stress conditions and this increase in the amount of proline plays a key role in the protection of cellular structures, osmotic adaptation and regulation of available nitrogen accumulation (Demiral and Türkan 2006; Yıldız et al. 2010). It has been reported that the proline synthesis varies according to the genotypes under stress conditions and can be synthesized in 100 folds higher amounts as compared to the amount synthesized in the control plants (Yıldız et al. 2010). While some studies have reported that the proline synthesis can be used as a selection criteria in terms of tolerance to salt stress, some studies have reported that the proline level does not change or even decrease under stress conditions, and it has been stated that it cannot be used as a selection criteria (Ramanjulu and Sudhakar 2001; Naik and Joshi 1983). In addition, it has been stated that the proline synthesis increased up to 5.2-fold in the salt-resistant species, but this increase has reached up to 27-fold in the salt-sensitive species (Kafi et al. 2003). It has been reported that the total phenolic substance synthesis is promoted in plants under salt stress conditions and these phenolic substances play the role as antioxidants in the removal of free oxygen radicals (Rezazadeh et al. 2019; Valifard et al. 2014; Rebey et al. 2017). Similar to our research results, Çetin et al. (2012) reported that phenolic substance synthesis increased in some vine rootstocks under salt stress conditions. In our study, it was determined that the total protein contents increased with the increase in salt concentrations, in all genotypes except the genotype 134. It has been reported in the previous studies that protein synthesis decreased with the increase in salt concentration (Gadallah 1999; Sorkheh et al. 2012), but it increased in some others (Mashayekhi et al. 2015; Zrig et al. 2016; Hatami et al. 2018). Increasing the amount of salt in plants increases the amount of active oxygen species, which causes enzyme inhibition, protein oxidation, and ultimately damage to DNA and RNA until the death of the plant. Proteins participate in many reactions in plants in order to minimize the damages that may occur at the cellular level of the excess salt formed in the environment. In particular, the proteins that act as sensors enable the functioning of regulatory proteins that regulate the expression of genes responsible for stress (phosphorylation/dephosphorylation). As a result of the work of this mechanism, the response of plants to stress occurs as either cell death, growth restriction or stress tolerance (Çulha and Çakırlar 2011). In addition, the excess amount of salt in the plant, especially the Na+ ion, negatively affects the uptake of Ca+2 ions which regulate the protein phosphorylation stages and the physiological and functional structure of the cell membranes, and the uptake of K+ ion which is one of the essential elements for growth and development, effective in the functionality of many enzymes in plant cells and in the maintenance of the osmotic balance, regulation of enzyme activity, protein synthesis, neutralization of negatively charged proteins and the movement of stomata, and thus negatively affects the ion balance of Na+/K+ and Na+/Ca+2. In addition, the synthesis of protective molecules such as LEA proteins, which are effective in regulating the ion concentration, protecting the cell membrane structure with proteins, and taking a role in regaining the structural form of the structurally disrupted proteins in plants exposed to salt stress, is also increasing (Çulha and Çakırlar 2011; Büyük et al. 2012). These proteins, which have a protective effect on the stress defense in plants, have been identified in seed embryos. In addition, Pareek et al. (1997) have suggested that the stress proteins are important molecular indicators in increasing salt tolerance, and they reported that tolerance may vary depending on the plant species, variety and genotype.
Conclusion
The importance of breeding rootstocks tolerant to abiotic and biotic stress factors increases day by day, so that very large areas can be made suitable for production. At this point, seedling rootstocks constitute an important advantage against climate changes (drought, sudden floods, etc.) and increasing environmental stress conditions (salinization, chemical pollution, etc.), which are considered certain to occur in the world in the future. The rootstocks used in almond cultivation are generally known to be sensitive to salt. Likewise, Zrig et al. (2011) reported that the almond seedlings were more sensitive to salinity than GN15 and GF 677 clone rootstocks in their study. In addition, Moore and Janick (1996) have reported that almond rootstocks are widely used in unfavorable soil conditions, they do not show incompatibility with almond varieties, and their ability to adapt to calcareous soils is better. As a result, it has been determined that almond genotypes are generally sensitive to salt. In general, it was determined that only the genotype 196 formed new shoots in the nutrient medium containing 150 mM NaCl. In addition, it was determined that the genotypes 9, 29, 54, 120, 134, 183, 185 and 241 could continue their development by forming new shoots in the nutrient medium containing 100 mM NaCl. In this respect, these genotypes stand out in terms of salt tolerance as compared to the other genotypes.
References
Ahmed CB, Rouina BB, Boukhris M (2008) Changes in water relations, photosynthetic activity and proline accumulation in one-year-old olive trees (Olea europaea L. cv. Chemlali) in response to NaCl salinity. Acta Physiol Plant 30:553–560. https://doi.org/10.1007/s11738-008-0154-6
Akça Y, Samsunlu E (2012) The effect of salt stress on growth, chlorophyll content, proline and nutrient accumulation, and K/Na ratio in walnut. Pak J Bot 44(5):1513–1520
Akçay D, Eşitken A (2017) MM106 anacı ve üzerine aşılı Golden Delicious elma çeşidine tuz stresinin etkileri. Selçuk Tarım Bilimleri Dergisi 3(2):228–232
Aono M, Kubo A, Saji H, Tanaka K, Kondo N (1993) Enhanced tolerance to photooxidative stress of transgenic Nicotiana tabacum with high chloroplastic glutathione reductase activity. Plant Cell Physiol 34(1):129–135. https://doi.org/10.1093/oxfordjournals.pcp.a078386
Aras S, Eşitken A (2018) Physiological responses of cherry rootstocks to short term salinity. Erwerbs-Obstbau 60(2):161–164. https://doi.org/10.1007/s10341-017-0350-x
Aziz I, Khan MA (2001) Experimental assessment of salinity tolerance of Ceriops tagal seedlings and saplings from the Indus delta, Pakistan. Aquat Bot 70(3):259–268. https://doi.org/10.1016/S0304-3770(01)00160-7
Beers RF, Sizer IW (1952) A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase. J Biol Chem 195(1):133–140
Berli FJ, Moreno D, Piccoli P, Hespanhol-Vıana L, Silva MF, Bressan-Smith R, Cavagnaro JB, Bottini R (2010) Abscisic acid is involved in the response of grape (Vitis vinifera L.) cv. Malbec leaf tissues to ultraviolet‑B radiation by enhancing ultraviolet-absorbing compounds, antioxidant enzymes and membrane sterols. Plant Cell Environ 33(1):1–10. https://doi.org/10.1111/j.1365-3040.2009.02044.x
Bolat I, Kaya C, Almaca A, Timucin S (2006) Calcium sulfate improves salinity tolerance in rootstocks of plum. J Plant Nutr 29(3):553–564. https://doi.org/10.1080/01904160500526717
Bolat I, Dikilitas M, Ercisli S, Ikinci A, Tonkaz T (2014) The effect of water stress on some morphological, physiological, and biochemical characteristics and bud success on apple and quince rootstocks. Sci World J. https://doi.org/10.1155/2014/769732
Büyük İ, Soydam-Aydın S, Aras S (2012) Bitkilerin stres koşullarına verdiği moleküler cevaplar. Turk Bull Hyg Exp Biol. https://doi.org/10.5505/TurkHijyen.2012.40316
Candan N, Tarhan L (2003) The correlation between antioxidant enzyme activities and lipid peroxidation levels in Mentha pulegium organs grown in Ca2+, Mg2+, Cu2+, Zn2+ and Mn2+ stress conditions. Plant Sci 165(4):769–776. https://doi.org/10.1016/S0168-9452(03)00269-3
Caverzan A, Passaia G, Rosa SB, Ribeiro CW, Lazzarotto F, Margis-Pinheiro M (2012) Plant responses to stresses: role of ascorbate peroxidase in the antioxidant protection. Genet Mol Biol 35(4):1011–1019. https://doi.org/10.1590/S1415-47572012000600016
Çelik A, Eraslan F (2015) Nitrik oksit uygulamasının tuz stresi altında yetiştirilen mısır bitkisinin mineral beslenmesi ve bazı fizyolojik özellikleri üzerine etkisi. Ziraat Fakültesi Dergisi 10(1):55–64
Çetin ES, Babalık Z, Göktürk Baydar N (2012) Bazı sofralık üzüm çeşitlerinde tanelerdeki toplam karbonhidrat, fenolik madde, antosiyanin, β‑karoten ve C vitamini içeriklerinin belirlenmesi. In: IV. Ulusal Üzümsü Meyveler Sempozyumu, pp 3–5
Channuntapipat C, Sedgley M, Collins G (2003) Micropropagation of almond cultivars Nonpareil and Ne Plus Ultra and the hybrid rootstock Titan x Nemaguard. Sci Hortic 98:473–484. https://doi.org/10.1016/S0304-4238(03)00067-0
Constantine NG, Stanley KR (1977) Superoxide dismutases. Plant Physiol 59(309):e314
Çulha Ş, Çakırlar H (2011) Tuzluluğun bitkiler üzerine etkileri ve tuz tolerans mekanizmaları. Afyon Kocatepe Üniversitesi Fen Ve Mühendislik Bilimleri Dergisi 11(2):11–34
Dal Bİ, Yazıcı K, Baktır İ (2001) Tuz stresi ve bahçe bitkileri üzerindeki etkileri. Derim 18(3):122–131
Daşgan HY, Koç S, Ekici B, Aktaş H, Abak K (2006) Bazı fasulye ve börülce genotiplerinin tuz stresine tepkileri. Alatarım 5(1):23–31
Dat JS, Vandenabeele E, Vranova M, Van Montagu D, Inze F, Breusegem FV (2000) Dual action of the active oxygen species during plant stress responses. Cell Mol Life Sci 57:779–795
De Britto AJ, Gracelin DHS, Sebastian SR (2011) Antibacterial activity of a few medicinal plants against Xanthomonas campestris and Aeromonas hydrophila. J Biopestic 4(1):57
Demiral T, Türkan I (2006) Exogenous glycinebetaine affects growth and proline accumulation and retards senescence in two rice cultivars under NaCl stress. Environ Exp Bot 56(1):72–79. https://doi.org/10.1016/j.envexpbot.2005.01.005
Doğru A, Canavar S (2020) Bitkilerde tuz toleransının fizyolojik ve biyokimyasal bileşenleri. Akademik Platf Mühendislik Ve Fen Bilimleri Dergisi 8(1):155–174. https://doi.org/10.21541/apjes.541620
Dölarslan M, Gül E (2012) Toprak bitki ilişkileri açısından tuzluluk. Türk Bilimsel Derlemeler Dergisi 5(2):56–59
Edrewa A (2005) Generation and scavenging of reactive oxygen species in chloroplasts: a submolecular approaches. Agric Ecosyst Environ 106:119–133. https://doi.org/10.1016/j.agee.2004.10.022
Ertürk U, Sivritepe N, Yerlikaya C, Bor M, Ozdemir F, Turkan I (2007) Responses of the cherry rootstock to salinity invitro. Biol Plant 51(3):597–600
Feng X, Chen L, Lei N, Wang S, Xu X, Zhou G, Li Z (2017) Emulsifying properties of oxidatively stressed myofibrillar protein emulsion gels prepared with (−)-epigallocatechin-3-gallate and NaCl. J Agric Food Chem 65(13):2816–2826. https://doi.org/10.1021/acs.jafc.6b05517
Fu M, Li C, Ma F (2013) Physiological responses and tolerance to nacl stress in different biotypes of malus prunifolia. Euphytica 189:101–109
Gadallah MAA (1999) Effects of proline and glycinebetaine on Vicia faba responses to salt stress. Biol Plant 42(2):249–257
Gupta B, Huang B (2014) Mechanism of salinity tolerance in plants: physiological, biochemical, and molecular characterization. Int J Genomics. https://doi.org/10.1155/2014/701596
Halliwell B (2008) Are polyphenols antioxidants or pro-oxidants? What do we learn from cell culture and in vivo studies? Arch Biochem Biophys 476(2):107–112. https://doi.org/10.1016/j.abb.2008.01.028t
Hartree EF (1972) Determination of protein: a modification of the lowry method that gives a linear photometric response. Anal Biochem 48(2):422–427. https://doi.org/10.1016/0003-2697(72)90094-2
Hatami E, Shokouhian AA, Ghanbari AR, Naseri LA (2018) Alleviating salt stress in almond rootstocks using of humic acid. Sci Hortic 237:296–302. https://doi.org/10.1016/j.scienta.2018.03.034
Inskeep WP, Bloom PR (1985) Extinction coefficients of chlorophyll a and B in n,n-dimethylformamide and 80 % acetone. Plant Physiol 77(2):483–485. https://doi.org/10.1104/pp.77.2.483
Kafi M, Stewart WS, Borland AM (2003) Carbohydrate and proline contents in leaves, roots, and apices of salt-tolerant and salt-sensitive wheat cultivars1. Russ J Plant Physiol 50(2):155–162
Köşkeroğlu S (2006). Tuz ve stresi altındaki mısır (Zea Mays L.) bitkisinde prolin birikim düzeyleri ve stres parametrelerinin araştırılması. Yüksek Lisans Tezi, Muğla Üniversitesi, Fen Bilimleri Enstitüsü.
Li W, Chen M, Wang E, Hu L, Hawkesford MJ et al (2016) Genome-wide analysis of autophagyassociated genes in foxtail millet (Setaria italica L.) and characterization of the function of SiATG8a in conferring tolerance to nitrogen starvation in rice. BMC Genomics 17(1):797
Liu B, Li M, Cheng L, Liang D, Zou Y, Ma F (2012) Influence of rootstock on antioxidant system in leaves and roots of young apple trees in response to drought stress. Plant Growth Regul 67(3):247–256
Mashayekhi M, Amiri ME, Habibi F (2015) Study of the biochemical responses and enzymatic activity of GF677 (peach and almond hybrid) rootstock to in vitro salinity stress. J Hortic Sci 29(2):207–2015
Meloni DA, Oliva MA, Martinez CA, Cambraia J (2003) Photosynthesis and activity of superoxide dismutase, peroxidase and glutathione reductase in cotton under salt stress. Environ Exp Bot 49(1):69–76. https://doi.org/10.1016/S0098-8472(02)00058-8
Moore JN, Janick J (1996) Fruit breeding: tree and tropical fruits; 2. Vine and small fruits; 3. Nuts. John Wiley & Sons, Oxford
Naik GR, Joshi GV (1983) Ineffectual role of proline metabolism in salt-stressed sugarcane leaves. Proc Plant Sci 92(3):265–269
Najafian SH, Rahemi M, Tavallali V (2008) Effect of salinity on tolerance of two bitter almond rootstock. American Eurasian Journal of Agricultural and Environmental Sciences 3(2):264–268
Nakano Y, Asada K (1981) Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol 22(5):867–880. https://doi.org/10.1093/oxfordjournals.pcp.a076232
Okubo M, Sakuratani T (2000) Effects of sodium chloride on survival and stem elongation of two Asian pear rootstock seedlings. Sci Hortic 85(1):85–90. https://doi.org/10.1016/S0304-4238(99)00141-7
Oluk EA, Sami O (2007) Yüksek öğretim öğrencilerinin sera etkisi, küresel isınma ve iklim değişikliği algılarının analizi. Dokuz Eylül Üniversitesi Buca Eğitim Fakültesi Dergisi 22:45–53.
Öztürk NZ (2015) Bitkilerin kuraklık stresine tepkilerinde bilinenler ve yeni yaklaşımlar. Turk J Agric Sci Technol 3(5):307–315. https://doi.org/10.24925/turjaf.v3i5.307-315.307
Pareek A, Singla SL, Grover A (1997) Salt responsive proteins/genes in crop plants. Oxford, IBH, New Delhi
Paschke M, Bernasconi G, Schmid B (2005) Effects of inbreeding and pollen donor provenance and diversity on offspring performance under environmental stress in the rare plant Cochlearia bavarica. Basic Appl Ecol 6(4):325–338. https://doi.org/10.1016/j.baae.2005.02.005
Ramanjulu S, Sudhakar C (2001) Alleviation of NaCl salinity stress by calcium is partly related to the increased proline accumulation in mulberry (Morus alba L.) callus. J Plant Biol 28:203–206
Ranjbarfordoei A, Samson R, Van Damme P (2006) Chlorophyll fluorescence performance of sweet almond [Prunus dulcis (Miller) D. Webb] in response to salinity stress induced by NaCl. Photosynt 44(4):513–522
Rebey IB, Bourgou S, Rahali FZ, Msaada K, Ksouri R, Marzouk B (2017) Relation between salt tolerance and biochemical changes in cumin (Cuminum cyminum L.) seeds. J Food Drug Anal 25(2):391–402. https://doi.org/10.1016/j.jfda.2016.10.001
Rezazadeh S, Ilkaee M, Aghayari F, Paknejad F, Rezaee M (2019) The physiological and biochemical responses of directly seeded and transplanted maize (Zea mays L.) supplied with plant growth-promoting rhizobacteria (PGPR) under water stress. Iran J Plant Physiol 10(1):3009–3021. https://doi.org/10.22034/IJPP.2019.670787
Scandalios JG (1996) Oxidative stress and moleculer biologoy of antioxidant defenses. Cold Spring Harbor Laboratory, Cold Spring Harbor
Shalaby EE, Epstein E, Qualset CO (1993) Variation in salt tolerance among some wheat and triticale genotypes. J Agron Crop Sci 171(5):298–304. https://doi.org/10.1111/j.1439-037X.1993.tb00144.x
Shibli RA, Shatnawi MA, Swaidat IQ (2003) Growth, osmotic adjustment, and nutrient acquisition of bitter almond under induced sodium chloride salinity invitro. Commun Soil Sci Plant Anal 34(13–14):1969–1979. https://doi.org/10.1081/CSS-120023231
Shiyab SM, Shibli RA, Mohammad MM (2003) Influence of sodium chloride salt stress on growth and nutrient acquisition of sour orange invitro. J Plant Nutr 26(5):985–996. https://doi.org/10.1081/PLN-120020070
Sibley JL, Eakes DJ, Gilliam CH, Keever GJ, Dozier WA, Himelrick D (1996) Foliar SPAD-502 meter values, nitrogen levels, and extractable chlorophyll for red maple selections. HortScience 31(3):468–470. https://doi.org/10.21273/HORTSCI.31.3.468
Singleton VL, Rossi JA (1965) Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am J Enol Vitic 16(3):144–158
Sivritepe N, Erturk U, Yerlikaya C, Turkan I, Bor M, Ozdemir F (2008) Response of the cherry rootstock to water stress induced invitro. Biol Plant 52(3):573
Sorkheh K, Shiran B, Rouhi V, Khodambashi M, Sofo A (2012) Salt stress induction of some key antioxidant enzymes and metabolites in eight Iranian wild almond species. Acta Physiol Plant 34(1):203–213
Sotiropoulos TE, Dimassi KN, Tsirakoglou V, Therios IN (2006) Responses of two Prunus rootstocks to KCl induced salinity invitro. Biol Plant 50(3):477–480
Tilkat EA, Kaplan A, Hoşer A, Tilkat E (2017) In vitro şartlarda yetiştirilen buttum (Pistacia khinjuk Stocks)’da çözünür karbonhidrat değerleri ile antioksidan peroksidaz aktivitesi üzerine tuz stresinin etkileri. Batman Üniversitesi Yaşam Bilimleri Dergisi 7(2/2):90–97
Turfan N (2016) Yerel ceviz çeşidinde (Juglans regia L.) abiyotik stres faktörlerine karşı dayanıklılık mekanizmasının belirlenmesi. Anadolu Tarım Bilimleri Dergisi 31(3):321–331. https://doi.org/10.7161/omuanajas.269984
Valifard M, Mohsenzadeh S, Kholdebarin B, Rowshan V (2014) Effects of salt stress on volatile compounds, total phenolic content and antioxidant activities of Salvia mirzayanii. S Afr J Bot 93:92–97. https://doi.org/10.1016/j.sajb.2014.04.002
Yıldırım AN, Tekintaş E, Koyuncu F (2007) Isparta bölgesinde geç çiçeklenen ve üstün nitelikli meyve veren badem (Prunus amygdalus Batsch.) genotiplerinin seleksiyonu. Adnan Menderes Üniversitesi Ziraat Fakültesi Dergisi 4(1–2):39–48
Yıldız M, Terzi H, Cenkci S, Terzi E, Uruşak B (2010) Bitkilerde tuzluluğa toleransın fizyolojik ve biyokimyasal markörleri. Anadolu Univ Sci Technol Life Sci Biotechnol 1(1):33
Yılmaz S, Temizgül R, Yürürdurmaz C, Kaplan M (2020) Oxidant and antioxidant enzyme response of redbine sweet sorghum under NaCl salinity stress. Bioagro 32(1):31–38
Zhishen J, Mengcheng T, Jianming W (1999) The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem 64(4):555–559. https://doi.org/10.1016/S0308-8146(98)00102-2
Zhu Z, Wei G, Li J, Qian Q, Yu J (2004) Silicon alleviates salt stress and increases antioxidant enzymes activity in leaves of salt-stressed cucumber (Cucumis sativus L.). Plant Sci 167(3):527–533. https://doi.org/10.1016/j.plantsci.2004.04.020
Zrig A, Tounekti T, Vadel AM, Mohamed HB, Valero D, Serrano M, Chaker C, Habib K (2011) Possible involvement of polyphenols and polyamines in salt tolerance of almond rootstocks. Plant Physiol Biochem 49(11):1313–1322. https://doi.org/10.1016/j.plaphy.2011.08.009
Zrig A, Mohamed HB, Tounekti T, Khemira H, Serrano M, Valeroc D, Vadelet AM (2016) Effect of rootstock on salinity tolerance of sweet almond (cv. Mazzetto). S Afr J Bot 102:50–59. https://doi.org/10.1016/j.sajb.2015.09.001
Acknowledgements
This research is an output of project number 215O779 supported by The Scientific and Technological Research Council of Turkey (TUBITAK).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
A.N. Yildirim, B. Şan, F. Yildirim, C. Çelik, B. Bayar and Y. Karakurt declare that they have no competing interests.
Rights and permissions
Springer Nature oder sein Lizenzgeber (z.B. eine Gesellschaft oder ein*e andere*r Vertragspartner*in) hält die ausschließlichen Nutzungsrechte an diesem Artikel kraft eines Verlagsvertrags mit dem/den Autor*in(nen) oder anderen Rechteinhaber*in(nen); die Selbstarchivierung der akzeptierten Manuskriptversion dieses Artikels durch Autor*in(nen) unterliegt ausschließlich den Bedingungen dieses Verlagsvertrags und dem geltenden Recht.
About this article

Cite this article
Yildirim, A.N., Şan, B., Yildirim, F. et al. Determining the Tolerance of Selected Almond Rootstock Genotypes to Salt Stress Under in vitro Conditions. Erwerbs-Obstbau 65, 299–310 (2023). https://doi.org/10.1007/s10341-022-00827-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10341-022-00827-y