
Abstract
The search for citrus rootstocks that confer resistance and tolerance to biotic and abiotic factors is increasing. Among abiotic stresses, salinity is one of the main factors that significantly affect growth and development in this crop. Faced this situation, genetic improvement seeks, among other aspects, to generate rootstocks that are tolerant to salinity. In this sense, tissue culture constitutes an important tool for citrus improvement, as it helps in the pre-selection of genotypes that best express salinity tolerance. Thus, this work aimed to study the tolerance of 11 citrus rootstocks to saline stress in vitro, using different concentrations of sodium chloride (NaCl). For analysis of in vitro regeneration, 1 cm long stem apexes, originating from the in vitro cultivation of rootstocks, were introduced in basal WPM medium and supplemented with 0.64, 1.28, 1.92, 2.56 and 3.20 g L−1 of NaCl. After 120 days in a growth room, the plants were evaluated and the data were analyzed using the R statistical program. The genotypes showed different behaviors depending on the interaction between the studied factors. There was a reduction in most of the in vitro development variables of rootstocks grown in culture medium supplemented with NaCl. The ‘Riverside’ citrandarin genotypes, HTR 051, LRF (LCR × TR) - 005, BRS Bravo, TSK × TRBK - Colômbia and BRS H Montenegro, were more tolerant to salinity, while TSKC × CTSW - 028, the mandarin ‘Sunki Tropical’ and BRS Santana were the most sensitive.
Key Message
In this study, we identified genotypes of citrus rootstocks with different levels of salinity tolerance through tissue culture, contributing to genetic improvement and enhancement of citrus production.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Rootstocks play a significant role in citrus growing, helping to mitigate problems involving aspects related to climate, soils with low fertility, disease infestation, high plant size and planting density. However, despite the availability of citrus rootstocks adapted to adverse conditions, few are considered superior and suitable for situations of abiotic and biotic stress. As a result, the demand for high-quality rootstocks that meet resistance and tolerance criteria against various factors has been progressively increasing (Çimen 2020). In the case of saline stress, one of the methods to overcome its effects is to graft sensitive commercial canopy onto salinity-resistant rootstocks (Madani et al. 2022).
Salinity is one of the main abiotic stresses that considerably affect the development of agriculture and especially the citrus crop, which is highly sensitive to high levels of salt (Bilal et al. 2020). Water salinity affects crops in two ways: through specific ion toxicity and through osmotic effects. The first occurs when the irrigation used ends up salinizing the soil in places where there is little water availability. The second refers to the salinity of the soil itself, which comes in a small part directly from the water, but for the most part inert to the soil, which increases the level of salt ions in the conductive tissues of the plant, and, therefore, both cases interfere with plant growth and may affect citrus plantations. In the case of damage caused by salinity, induced by means of high osmotic potential, Alam et al. (2020) state that this occurs when the amount of salt in the plant’s growing environment is high enough to reduce growth.
Due to its negative influence on the yield of many crops, salinity is a major problem for citrus crops, as it has an adverse effect on tree growth and causes many physiological disturbances, as salt stress reduces liquid CO2 assimilation and stomatal conductance, as well as the water potential of tree leaves, in addition to promoting excessive accumulation of chloride or sodium in leaves (Al-Yassin 2004; Sharma et al. 2013). When subjected to saline irrigation water, citrus plants immediately stop their growth, negatively affecting fruit quality in much greater proportions than in other trees (Prior et al. 2007; Ziogas et al. 2021).
As mentioned by Kaushal et al. (2013) in the southwest region of Punjad, India, the main abiotic stress that citrus orchards face is salinity, where salt stress impedes plant growth and reduces productivity because of ionic toxicity, lack of water and imbalance nutrition, which results in a significant loss of yield. Age, position and genotype affect chlorine accumulation in citrus leaves; this suggests that factors other than the root system play a role in salt tolerance. As a result, when the salinity level increases during irrigation, the aerial part, the number of leaves and the fresh mass of the seedlings decrease, as well as the relative levels of chlorophyll, net photosynthesis and the rate of respiration in the leaves. Therefore, the use of tolerant genotypes and soil and water control can avoid these problems.
In this sense, in addition to soil and water management, biotechnological approaches have been adopted as one of the alternatives to overcome salinity problems under laboratory conditions (Sharma et al. 2013). Among these tools, tissue culture is one of the most reliable and useful to determine the sensitivity of the genotype to salinity, as well as to evaluate basic aspects of plant growth and development, thus allowing to test genotypes early, in a short time and physical space (Pérez-Clemente et al. 2008; Pérez-Jiménez and Pérez-Tornero 2020a).
Thus, this technique contributes to conventional genetic improvement, considering that it identifies possible salinity-tolerant cultivars that can be previously tested for subsequent cultivation in different types of soil and under different environmental conditions (Mahmoud et al. 2020). Therefore, the objective of this work was to study the tolerance of 11 citrus rootstocks to saline stress in vitro, using different concentrations of sodium chloride (NaCl) in the culture medium.
Methodology
The work was developed at the Tissue Culture Laboratory of Embrapa Mandioca e Fruticultura, located in the city of Cruz das Almas, Bahia, Brazil. Stem apexes, 1 cm long, from plants previously grown in vitro on Common Sunki mandarin (TSKC) × ‘Swingle’ citrumelo (CTSW) - 028, citrandarin ‘Riverside’, citrandarin ‘Índio’, citrandarin ‘San Diego’, ‘Sunki Tropical’ mandarin, trifoliate hybrid (BRS Santana), BRS Victória, ‘Rugoso da Florida’ lemon tree (LRF) ((‘Rangpur lemon tree (LCR) × Poncirus trifoliata (TR) - 005, BRS Bravo, mandarin tree Sunki (TSK) × Poncirus trifoliata ‘Benecke’ (TRBK) - Colômbia and BRS H Montenegro were introduced into test tubes (2.5 × 15 cm) containing 10 mL of WPM culture medium (Lloyd; McCown 1980), in its basal composition and supplemented with NaCl at concentrations of 0.64 g L−1; 1.28 g L−1; 1.92 g L−1; 2.56 g L−1 and 3.20 g L−1 All media were added with 30 g L−1 of sucrose, gelled with 2.4 g L−1 of Phytagel® and had the pH adjusted to 5.7 before autoclaving, performed for 20 min at 120 °C.
After introduction into the culture medium, the explants were kept in a growth room, under conditions of temperature of 27 ± 1 °C, photon flux density of 30 µmol m−2 s−1 and photoperiod of 16 h.
The experimental design was completely randomized in a factorial 11 × 6 scheme (genotype and NaCl concentration), with 15 replications per treatment, with each experimental plot consisting of a mini-cutting grown in a test tube.
After 120 days of in vitro cultivation, the following variables were evaluated: aerial part height (APH), in cm; number of green leaves (NGL); number of minicuttings (NM) 1 cm long and containing at least one bud; fresh mass of aerial part (FMAP) and roots (FMR), in mg; aerial dry matter mass (ADMM) and root dry matter (RDM), in mg; number of roots (NR) and length of the largest root (LLR), in cm. To determine the dry matter mass, the plant material was placed in an oven with forced air circulation at a temperature of 60 ºC, until it reached a constant weight.
The data obtained were submitted to the F test of analysis of variance, with the aid of the statistical program R, version 3.4 (Core Team 2016). Means of genotypes were grouped by the Scott–Knott test at 5% probability and means of NaCl concentrations were adjusted for polynomial regression models. The count values for the number of green leaves, number of senescent leaves, number of minicuttings and number of roots were transformed into √(x + 0.5), aiming to meet the assumptions of the analysis of variance.
Results and discussion
There was a significant effect at 1% probability (P < 0.01) by the ANOVA F test for most of the isolated factors, with the exceptions being fresh and dry mass of roots for the NaCl factor. In the interaction of the factors, there was a highly significant effect at 1% probability for the variables shoot height, number of green leaves, number of minicuttings, number of roots, length of the largest root and root dry matter mass. In the other variables, factor interactions were obtained at 5% probability (P < 0.05) (Table 1).
The in vitro cultivation of the different genotypes submitted to different concentrations of NaCl resulted in a coefficient of variation (CV) that varied between 14.81 and 76.18%, in the variables number of minicuttings and fresh mass of roots, respectively.
In the adjustments of the models of linear and polynomial equations, coefficients of determination (R²) were obtained varying between 50.95%, for the number of senescent leaves, in the interaction of NaCl with the LRF genotype (LCR × TR) - 005, and 100%, for the length of the largest root, on the interaction of NaCl with ‘Sunki Tropical’ mandarin.
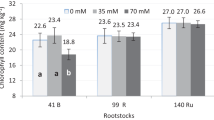
From the values estimated by the equations, it is observed that for all genotypes, the absence of NaCl favored the best response in shoot height, with a reduction in the mean according to the addition of salt (Fig. 1). Pérez-Jiménez and Pérez-Tornero (2020a) also observed a reduction in growth with the addition of 80 mM NaCl in Citrus macrophylla Wester. and in the mutant derivatives MM2A, MM3A and MM1B. As reported by Sotiropoulos et al. (2006), salinity is a limiting factor for plant growth, negatively affecting survival, biomass and height.

Aerial part height (cm) of citrus rootstocks, submitted to different concentrations of this salt in the WPM culture medium for 120 days
For the number of green leaves, 5 of the eleven genotypes showed the highest means in the absence of NaCl. However, the TSKC × CTSW - 28 genotypes, ‘Riverside’ and ‘Índio’ citrandarins, BRS Bravo and LCR × TR 001 showed a reduction in the number of green leaves up to concentrations of 1.30 g L−1, 2.00 g L−1, 1.79 g L−1, 1.99 g L−1 and 1.07 g L−1, respectively, where the lowest means were estimated (From the values estimated by the equations, it is observed that for all genotypes, the absence of NaCl favored the best response in shoot height, with a reduction in the mean according to the addition of salt (Fig. 2a). Regarding the number of senescent leaves, the lowest averages were observed in the absence of NaCl for T. ‘Sunki Tropical’, LRF (LCR × TR) - 005, BRS Bravo and LCR × TR - 001, as shown in Fig. 2b. According to Munns and Tester (2008), increased osmotic tolerance is evidenced by increased ability to continue producing new leaves, while tissue tolerance is related to increased survival of older leaves. Thus, it is observed that for most of the genotypes in this study, the increase in NaCl concentrations, considering the variables number of green leaves and the number of senescent leaves, promoted a reduction in osmotic and tissue tolerance, consequences of salt toxicity.

Number of green leaves (a) and number of senescent leaves (b) of citrus rootstocks, submitted to different concentrations of this salt in the WPM culture medium for 120 days
Ghaleb et al. (2010), when working with the rootstocks sour orange tree (Citrus aurantium L.) and ‘Volkamerian’ lemon tree (Citrus volkameriana Ten. et Pasq.) in vitro, observed a decrease in the length of the plant and in the number of green leaves with the increasing the dose of NaCl. This behavior was also observed by Shiyab et al. (2003) in sour orange, and this reduction is explained by the increase in salinity levels, and, consequently, as a function of the effect of osmotic potential and salt toxicity. Furthermore, the authors reported that symptoms of salt toxicity became apparent at the highest levels of NaCl, 200 and 300 mM, in the culture medium, causing complete death of the explant.
In irrigation studies, such as the one developed by Alam et al. (2020) with pomelo (Citrus maxima Merr.), ‘Cleópatra’ mandarin (Citrus reshni Hort. ex Tan.) and calamansi (Fortunella japonica Thunb.) seedlings, it was found that all growth parameters decreased with increasing salinity water level irrigation. However, these authors observed that pomelo and ‘Cleópatra’ mandarin trees were more tolerant to salinity conditions. Anjum (2008) observed this same decrease in plant development parameters from experiments with ‘Cleópatra’ mandarin and ‘Troyer’ citrang [Citrus sinensis (L.) Osb. × Poncirus trifoliata (L.) Raf.].
An in vitro study testing seed germination in different salinity gradients carried out by Garcia et al. (2002) with several genotypes, found that the ‘Volkamerian’ lemon tree was the most sensitive genotype to salt. In contrast, Yaman et al. (2020) obtained the highest germination rates in all salt concentrations on the rootstocks of this same lemon and sour orange and the lowest values on the rootstocks of rough lemon (Citrus jambhiri Lush.) and trifoliate orange (Poncirus trifoliata).
For the number of minicuttings, it was possible to adjust the quadratic equation for ‘Riverside’ citrandarin, which showed a reduction in mean values up to 2.27 g L−1 of NaCl, obtaining the lowest average at this concentration, while ‘San Diego’ citrandarin showed the highest mean at the optimum dose of 1.54 g L−1 of salt. As for the BRS Victória and BRS Bravo genotypes, the adjustment of the linear equation showed that in the absence of NaCl it was possible to reach the highest averages for this Variable (Fig. 3).

Number of minicuttings of citrus rootstocks, submitted to different concentrations of this salt in the WPM culture medium for 120 days
Different from what was observed in the variables above, for the aerial fresh matter mass, citrandarin ‘San Diego’, BRS Victória and LRF (LCR × TR) - 005 showed the highest averages with the addition of NaCl in the concentrations of 1.77 g L−1, 2.25 g L−1 and 1.49 g L−1, respectively. In addition to these genotypes, BRS Bravo and BRS H Montenegro also followed this behavior, showing the highest averages at the optimal dose of 3.20 g L−1 (Fig. 4a). Thus, it is observed that the addition of NaCl to these genotypes, despite presenting a reduction in the other growth variables, did not affect the aerial fresh matter mass, that is, even with a reduction in variables such as height and number of green leaves, biomass accumulation occurred in the plant.
For aerial dry matter mass, it was possible to adjust the equation with a significant effect for BRS Victória, LRF (LCR × TR) - 005 and BRS H Montenegro genotypes, where it is observed that the addition of NaCl in concentrations of 2.10 g L−1, 1.62 g L−1 and 3.20 g L−1, respectively, resulted in the highest means (Fig. 4b). These results differ from those observed in a study conducted by Ghaleb et al. (2010), using sodium chloride and calcium chloride in the culture medium with sour orange and ‘Volkamerian’ lemon rootstocks, in which the dry weight of the plant decreased as the salinity level increased. Shiyab et al. (2003) also observed a reduction in dry weight with an increase in NaCl, in a study conducted in vitro with sour orange. However, corroborating the present study, Shibli et al. (2000), analyzing some apple varieties, observed an increase in dry matter mass with the increase in salinity for most varieties, except for the concentration of 100 mM. These favorable results, observed for the fresh and dry mass of shoots, can be explained by the positive effect exerted by low concentrations of NaCl on the proliferation of shoots in vitro, in response to the induction of increased osmolarity, as reported by Flowers and Läuchli (1983).

Aerial fresh matter mass (a) and aerial dry matter mass (b) of citrus rootstocks, submitted to different concentrations of this salt in the WPM culture medium for 120 days
The addition of NaCl reduced the number of roots for all studied genotypes (Fig. 5a). Similar results were obtained in the study conducted by Shiyab et al. (2003), in which no rooting occurred when microcuttings were grown in medium containing 150 mM or more of NaCl. For the length of the largest root, it was possible to adjust the equation with a significant effect only for the mandarin genotypes ‘Sunki Tropical’ and LRF (LCR × TR) - 005 which, according to the estimated values of the quadratic equation, presented the highest averages in the optimal doses of 0.44 g L−1 and 1.45 g L−1 of NaCl, respectively (Fig. 5b). Significant damage results in the rhizogenesis process with increasing NaCl concentration were also observed by Pérez-Tornero et al. (2009), who obtained 30% rooting and a 45% reduction in the number and length of the root of Citrus macrophylla with 100 mM NaCl, which is the most affected growth parameter in the in vitro growth of genotypes cultivated under different saline concentrations.

Number of roots (a) and length of the largest root (b) of citrus rootstocks, submitted to different concentrations of this salt in the WPM culture medium for 120 days
There was a significant effect only for the hybrid LRF (LCR × TR) - 005 when the fresh mass of roots was analyzed, adjusting a quadratic equation model where the estimated optimal dose was 1.45 g L−1, obtaining at this concentration the highest average (Fig. 6a). As for the dry mass of roots, the citrandarin ‘Riverside’ and LRF genotypes (LCR × TR) - 005 showed a significant effect, with the highest averages obtained in the absence and concentration of 1.46 g L−1 of NaCl, respectively (Fig. 6b). Sharma et al. (2013) evaluating seed germination and some growth variables of rough lemon trees, observed that the roots were less influenced by the addition of NaCl, when compared to the variables related to the aerial part. In agreement with the assertion of these authors, the results observed for the length of the largest root and the fresh and dry mass of roots demonstrate that the addition of salt, for most of the studied citrus genotypes, did not affect the growth nor the mass of the roots, when it is observed that there were no significant effects (Table 2).

Root fresh matter mass (a) and root dry matter mass (b) of citrus rootstocks, submitted to different concentrations of this salt in the WPM culture medium for 120 days
The genotypes showed different behavior depending on the variables studied with the different concentrations of NaCl. The highest means of aerial part height were obtained for citrandarins ‘Riverside’ and ‘Índio’, in the absence of NaCl, and for the hybrid LRF (LCR × TR) - 005 at a concentration of 0.64 g L−1. At the concentration of 1.28 g L−1, most genotypes showed higher means, not statistically different from each other, except for genotypes TSKC × CTSW - 028, mandarin ‘Sunki Tropical’, BRS Santana and BRS Victória, which presented inferior to the others. At concentrations of 1.92 g L−1 and 2.56 g L−1 of NaCl, the highest averages were observed in the ‘Riverside’ and ‘Índio’ citrandarins genotypes, LRF (LCR × TR) - 005, TSKC × (LCR × TR) - 059 and TSK × TRBK - Colômbia. In addition to these genotypes, for the concentration of 2.56 g L−1, the BRS Victória genotype also stood out. At a dose of 3.20 g L−1 of salt, citrandarin ‘Riverside’, and again the hybrids LRF (LCR × TR) - 005, BRS Bravo and TSK × TRBK - Colômbia presented the highest averages (Table 2).
The number of green leaves was higher for citrandarin ‘Riverside’ and ‘Índio’, mandarin ‘Sunki Tropical’ and BRS Bravo in the absence of NaCl. At the concentration of 0.64 g L−1, the BRS Bravo genotype presented the highest average, while the lowest average was for HTR - 069. The citrandarin genotypes ‘Riverside’ and ‘Índio’, LRF (LCR × TR) - 005 and BRS Bravo were responsible for the highest averages at the concentration of 1.28 g L−1. These same genotypes together with the hybrids BRS Victória and TSK × TRBK - Colômbia were superior to the others at concentrations of 1.92 g L−1. For the concentration of 2.56 g L−1 of salt, all genotypes, except for ‘Riverside’ and ‘San Diego’ citrandarins, resulted in higher mean values. At the concentration of 3.20 g L−1 of NaCl, the highest averages were observed in citrandarins ‘Riverside’ and ‘Índio’, BRS Bravo, TSK × TRBK - Colômbia and BRS H Montenegro (Table 2).
Regarding the number of senescent leaves, in the absence of NaCl, the highest averages occurred in TSKC × CTSW - 28 and mandarin ‘Sunki Tropical’. At the concentrations of NaCl 0.64 g L−1 and 1.28 g L−1, the rootstock ‘Sunki Tropical’ was also responsible for the highest averages. At a dose of 1.92 g L−1 of salt, the citrandarin ‘San Diego’, BRS Bravo and BRS H Montenegro genotypes had the highest averages. At the concentrations of 2.56 g L−1 and 3.20 g L−1, there were no statistical differences between the genotypes. In general, for most genotypes, low leaf senescence was recorded even at the highest NaCl doses, as can be seen in Table 2.
For the number of minicuttings, in the absence of salt, citrandarin ‘Riverside’, LRF (LCR × TR) - 005, and BRS Bravo presented the highest values, while in the concentration of 0.64 of NaCl the highest averages were registered in these last two genotypes and in TSK × TRBK - Colômbia. In the ‘Riverside’ and ‘Índio’ citrandarin genotypes, and again in the LRF (LCR × TR) - 005 and in BRS Bravo, the highest number of minicuttings was observed at the concentration of 1.28 g L−1. In addition to these genotypes, TSK × TRBK - Colômbia also showed high averages in the highest NaCl concentration (3.20 g L−1). At concentrations of 1.92 g L−1 and 2.56 g L−1, the citrandarin genotypes ‘Índio’, BRS Victória, LRF (LCR × TR) - 005, TSKC × (LCR × TR) - 059 and TSK × TRBK - Colômbia were responsible for the highest averages, in addition to CSF × TR - 001 at a dose of 2.56 g L−1 of NaCl (Table 2).
In relation to the fresh mass of aerial part, there were, in the absence and concentration of 0.64 g L−1, statistically higher averages for the genotypes citrandarin ‘Riverside’, LRF (CRL × TR) - 005 and BRS Bravo, plus citrandarin ‘Indio’ in the absence of NaCl. The LRF (LCR × TR) - 005 and BRS Bravo genotypes were also responsible for the highest averages at the concentration of 1.28 g L−1. Again, TSKC × (CSF × TR) - 059 stood out in NaCl doses of 1.92, 2.56 and 3.20 g L−1, showing means that differed statistically from the others (Table 2).
The same response of the genotypes obtained in the above variable is observed in the absence and in the concentrations of 0.64 g L−1 and 1.28 g L−1 of NaCl for the dry matter mass of aerial part. At the concentration of 1.92 g L−1, in the genotypes citrandarin ‘Riverside’ and ‘Índio’, LRF (LCR × TR) - 005 and BRS Bravo, the highest averages were observed. These same genotypes, except for citrandarin ‘Riverside’, also showed the highest values at the dose of 2.56 g L−1 of NaCl. At the highest concentration of NaCl, 3.20 g L−1, the highest average was observed in the hybrid BRS Bravo and the lowest in BRS Victória (Table 2).
For the number of roots, all genotypes except TSKC × CTSW - 028, HTR - 069, BRS Victória and BRS Bravo showed high means in the absence of NaCl. At a concentration of 0.64 g L−1, the genotypes citrandarin ‘Riverside’ and ‘Índio’, LRF (LCR × TR) - 005, TSK × TRBK - Colômbia and BRS H Montenegro were responsible for the highest averages. At the concentration of 1.28 g L−1, in the genotypes citrandarin ‘Índio’ and ‘San Diego’, LRF (LCR × TR) - 005 and LCR × TR - 001, the highest number of roots was obtained. At a concentration of 1.96 g L−1, citrandarin ‘Índio’ and the hybrids HTR-051, LRF (LCR × TR) - 005 and BRS H Montenegro were responsible for the highest means, while at a concentration of 2.56 g L−1 1 the BRS Victória and BRS H Montenegro genotypes showed the highest averages. At the highest dose of NaCl (3.20 g L−1) it was observed that there were no statistical differences between the genotypes, however it was verified that there was a smaller number of roots in relation to the other concentrations, indicating that the addition of NaCl affected root development, mainly in citrandarins ‘Índio’ and ‘San Diego’ that did not show any root (Table 3).
The length of the largest root showed the highest averages for the LRF genotype (LCR × TR) - 005 and the mandarin ‘Sunki Tropical’ tree in the absence of NaCl. At concentrations of 0.64, 1.28, 1.96 and 2.56 g L−1 of NaCl the LRF (CRL × TR) - 005 remained higher, differing statistically from the other genotypes, while at the dose of 3.20 g L−1 there were no statistical differences between the genotypes. Root fresh matter mass showed the same behavior for the genotypes at all doses studied (Table 3).
For root dry matter mass, citrandarins ‘Riverside’ and ‘San Diego’, mandarin ‘Sunki Tropical’, LRF (LCR × TR) - 005 and BRS Bravo genotypes presented the highest averages in the absence of NaCl. At the concentration of 0.64 g L−1, the highest average occurred in mandarin ‘Sunki Tropical’ and LRF (LCR × TR) - 005. At NaCl concentrations of 1.28, 1.96 and 2.56 g L−1 the highest average was obtained in the LRF (CRL × TR) - 005, while when the 3.20 g L−1 was used, the salt did not provide statistical differences between the genotypes (Table 3).
According to Maas (1993), the ability of citrus plants to tolerate salinity varies among different species, although it is well known that salt tolerance is particularly dependent on the rootstock, as observed in the results obtained in this study.
According to Syvertsen et al. (2010), rootstocks such as ‘Rangpur’ lime and ‘Cleópatra’ mandarin have relatively high resistance to salt, due to their ability to limit the accumulation of Cl- in leaves of the canopy variety. Rootstocks such as Poncirus trifoliata and its hybrids, such as ‘Carrizo’ citrange, are considered sensitive to salt, as they accumulate high levels of Cl- in the leaves of the scion variety, although they have also demonstrated the ability to restrict the transport of Na + to the stem (García-Sánchez and Syvertsen 2006).
It is observed that at the concentrations of 1.92, 2.56 and 3.20 g L−1 of NaCl, for the mandarin genotypes ‘Sunki Tropical’ and HTR - 069, all of them died, and at doses of 2.56 g L−1 and 3.20 g L−1 the same occurred with TSKC × CTSW - 028. For citrandarins ‘Índio’ and ‘San Diego’ there was a significant reduction in root development at the highest dose of the product (Table 3). This is one of the consequences of exposure to salinity, where the excessive absorption of these ions has a notable effect on the performance and yield of the plants, which can, in addition to reducing growth, cause their death. However, some plants have different mechanisms for exclusion and/or extrusion of ions to survive in saline environments (Ghaleb et al. 2010; Pérez-Jiménez and Pérez-Tornero 2020b), mechanisms that may be present in rootstocks which, even with a reduction in development for some growth variables, managed to form aerial parts and roots at higher NaCl doses, with less damage than those observed in more sensitive genotypes, such as the rootstock citrandarin ‘Riverside’, BRS Victória, LRF (LCR × TR) - 005, BRS Bravo, TSK x TRBK - Colômbia and BRS H Montenegro, presenting, therefore, by this aspect, some degree of resistance to salinity.
Conclusion
The addition of NaCl to the culture medium reduced the growth of citrus rootstocks in most of the growth variables analyzed.
The genotypes citrandarin ‘Riverside’, BRS Victória, LRF (LCR × TR) - 005, BRS Bravo, TSK × TRBK - Colômbia and BRS H Montenegro were more tolerant to salinity, while TSKC × CTSW - 028, the mandarin ' Sunki Tropical’ and BRS Santana were more sensitive.
Data availability
All data generated were analyzed during this study and are included in this published article.
References
Al-Yassin ADNAN (2004) Influence of salinity on citrus: a review paper. J Cent Eur Agric 5(4):263–272
Alam A, Ullah H, Attia A, Datta A (2020) Effects of salinity stress on growth, mineral nutrient accumulation and biochemical parameters of seedlings of three citrus rootstocks. Int J Fruit Sci 20(4):786–804
Anjum MA (2008) Effect of NaCl concentrations in irrigation water on growth and polyamine metabolism in two citrus rootstocks with different levels of salinity tolerance. Acta Physiol Plant 30(1):43
Bilal HH, Zulfiqar R, Adnan M, Umer MS, Islam H, Zaheer H, Ahmad RI (2020) Impact of salinity on citrus production; a review. Int J Appl Res 6(8):173–176
Çimen B (2020) Efficient protoplast isolation from ovule-derived embryogenic callus in Citrus Volkameriana. Turkish J Agric Forestry 44(6):567–576
Flowers TJ, Läuchli A (1983) Sodium versus potassium: substitution and compartmentation. In: Pearson A, Zimmerman MH (eds) Encyclopedia of plant physiology. Springer, Berlin
Garcia MR, Bernet GP, Puchades J, Gomez I, Carbonell EA, Asins MJ (2002) Reliable and easy screening technique for salt tolerance of citrus rootstocks under controlled environments. Aust J Agric Res 53(6):653–662
García-Sánchez F, Syvertsen JP (2006) Salinity tolerance of Cleopatra mandarin and Carrizo citrange citrus rootstock seedlings is affected by CO2enrichment during growth. J Am Soc Hortic Sci 131(1):24–31
Ghaleb WS, Sawwan JS, Akash MW, Al-Abdallat AM (2010) Vitro response of two citrus rootstocks to salt stress. Int J Fruit Sci 10(1):40–53
Kaushal M, Kumar L, Gill MIS, Choudhary OP, Bali SK (2013) Effect of salinity on survival and growth performance of in vitro grown rough lemon (Citrus jambhiri Lush.) seeds. Indian J Biotechnol 12:284–286
Lloyd G, Mccown B (1980) Use of microculture for production and improvement of Rhododendron spp. HortScience 15(3):416–417
Maas EV (1993) Salinity and citriculture. Tree Physiol 12(2):195–216
Madani SM, Piri S, Sedaghathoor S (2022) The response of three mandarin cultivars grafted on sour orange rootstock to salinity stress. Int J Fruit Sci 22(1):264–274
Mahmoud LM, Dutt M, Vincent CI, Grosser JW (2020) Salinity-induced physiological responses of three putative salt tolerant citrus rootstocks. Horticulturae 6(4):90
Munns R, Tester M (2008) Mechanisms of salinity tolerance. Annu Rev Plant Biol 59:651–681
Pérez-Clemente RM, Montoliu A, López P, López-Climent MF, Arbona V, Gómez-Cadenas A (2008) In vitro tissue culture approaches for the study of salt stress in citrus. In biosaline agriculture and high salinity tolerance, Birkhäuser Basel (pp. 37–42)
Pérez-Jiménez M, Pérez-Tornero O (2020) Vitro plant evaluation trial: reliability test of salinity assays in citrus plants. Plants 9(10):1352
Pérez-Jiménez M, Pérez-Tornero O (2020) Mutants of Citrus macrophylla rootstock obtained by gamma radiation improve salt resistance through toxic ion exclusion. Plant Physiol Biochem 155:494–501
Pérez-Tornero O, Tallón CI, Porras I, Navarro JM (2009) Physiological and growth changes in micropropagated Citrus macrophylla explants due to salinity. J Plant Physiol 166(17):1923–1933
Prior LD, Grieve AM, Bevington KB, Slavich PG (2007) Long-term effects of saline irrigation water on ‘Valencia’orange trees: relationships between growth and yield, and salt levels in soil and leaves. Aust J Agric Res 58(4):349–358
R Core Team, 2016. R: A Language and Environment For Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria http://www.R-project.org/. Acesso em 27 de outubro de 2019.
Sharma LK, Kaushal M, Bali SK, Choudhary OP (2013) Evaluation of rough lemon (Citrus jambhiri Lush.) as rootstock for salinity tolerance at seedling stage under invitro conditions. Afr J Biotechnol 12(44):6267–6275
Shibli R, Mohammad M, Abu-Ein A, Shatnawi M (2000) Growth and micronutrient acquisition of some apple varieties in response to gradual in vitro induced salinity. J Plant Nutr 23(9):1209–1215
Shiyab SM, Shibli RA, Mohammad MM (2003) Influence of sodium chloride salt stress on growth and nutrient acquisition of sour orange in vitro. J Plant Nutr 26(5):985–996
Sotiropoulos TE, Dimassi KN, Tsirakoglou V, Therios IN (2006) Responses of two Prunus rootstocks to KCl induced salinity in vitro. Biol Plant 50(3):477–480
Syvertsen JP, Melgar JC, García-Sánchez F (2010) Salinity tolerance and leaf water use efficiency in Citrus. J Am Soc Hortic Sci 135(1):33–39
Yaman M, Pınar H, Seday U, Altınöz D, Uzun A, Çabuk NS (2020) screening for Salt Tolerance of some Citrus rootstocks. Turkish J Agriculture-Food Sci Technol 8(5):1117–1121
Ziogas V, Tanou G, Morianou G, Kourgialas N (2021) Drought and salinity in citriculture: optimal practices to alleviate salinity and water stress. Agronomy 11(7):1283
Acknowledgments
The author expresses her sincere thanks to Dr. Antônio da Silva Souza for his invaluable support, valuable guidance and excellent work, as well as for the fundamental review of the manuscript. Your dedication and contribution were essential to the quality of this work.
Funding
To CAPES and FAPESB, for their financial support for the scholarships of the students involved in the study. To Embrapa Mandiocae Fruticultura, for providing the physical structure and financial.
support for the development of the study.
Author information
Authors and Affiliations
Contributions
DSVV, MISM and LVCN: collected the data and interpreted the results. DSVV and MISM: wrote the manuscript. DSVV, MISM, ASS, KCFS and WSSF: provided technical support to set up and conduct the study. ASS, KCFS, and WSSF: defined the main lines of the experiments. DSVV: supported the statistical analysis. ASS, KCFS and WSSF: were the supervisors of this research. ASS, WSSF and KCFS: contributed with the financial acquisition of the experiments.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no known competing fnancial interests or personal relationships that could have appeared to infuence the work reported in this paper.
Additional information
Communicated by Maria Antonietta Germanà.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Vila Verde, D.d., Mendes, M.I.d., Nobre, L.V.d. et al. In vitro tolerance of citrus rootstocks under saline stress. Plant Cell Tiss Organ Cult 156, 13 (2024). https://doi.org/10.1007/s11240-023-02627-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11240-023-02627-y