
Abstract
Animals are often required to estimate object sizes during several fitness-related activities, such as choosing mates, foraging, and competing for resources. Some species are susceptible to size illusions, i.e. the misperception of the size of an object based on the surrounding context, but other species are not. This interspecific variation might be adaptive, reflecting species-specific selective pressures; according to this hypothesis, it is important to test species in which size discrimination has a notable ecological relevance. We tested susceptibility to a size illusion in the guppy, Poecilia reticulata, a fish species required to accurately estimate sizes during mate choice, foraging, and antipredator behaviours. We focussed on the Delboeuf illusion, in which an object is typically perceived to be larger when surrounded by a smaller object. In experiment 1, we trained guppies to select the larger of two circles to obtain a food reward and then tested them using stimuli arranged in a Delboeuf-like pattern. In experiment 2, we tested guppies in a spontaneous food choice task to determine whether the subjective size perception of food items is affected by the surrounding context. Jointly, our experiments indicated that guppies perceived the Delboeuf illusion, but in a reverse direction relative to humans: guppies estimated as larger the stimulus that human perceived as smaller. Our results indicated susceptibility to size illusions also in a species required to perform accurate size discrimination and support previous evidence of variability in illusion susceptibility across vertebrates.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Animals are often required to make adaptive decisions based on the size of a given stimulus. For example, hermit crabs decide whether or not to evict a rival based on the perception of the rival’s shell size (Dowds and Elwood 1983). The female swordfish decides whether to mate with a male based on the tail length of that male (Basolo 1990). The rules underlying these decisions are often studied based on optimality models with the assumption that the animal recognises the real size of the available options (McNamara et al. 2001). However, visual information is often ambiguous: indeed, 3D form or scene is translated into a 2D retinal representation and this representation is in turn interpreted by cognitive processes in the brain (Kelley and Kelley 2014). This sensory process might cause misreading of information (i.e., visual illusions; Kelley and Kelley 2014) that in some cases, involves size perception.
Two size illusions commonly studied in humans are the Delboeuf illusion and the Ebbinghaus–Titchener illusion, whereby two same-sized objects are perceived as different depending on the surrounding context. Humans tend to perceive an object surrounded by a larger object to be smaller. A body of literature reveals similar effects across vertebrates. For example, rhesus monkeys, Macaca mulatta, and capuchin monkeys, Cebus apella, fall prey to the Delboeuf illusion as humans do (Parrish et al. 2015). A study on an individual bottlenose dolphin, Tursiops truncatus, has reported that the subject perceived the Ebbinghaus–Titchener illusion similarly to humans (Murayama et al. 2012). Likewise, two teleost fish species have been reported to perceive the Ebbinghaus–Titchener illusion (Sovrano et al. 2015) and the Delboeuf illusion (Fuss and Schluessel 2017). However, there are some exceptions: two independent studies on dogs, Canis lupus familiaris, one study on baboons, Papio papio, and one study on grey bamboo sharks, Chiloscyllium griseum, determined that these species are not sensitive to the Delboeuf illusion (Parron and Fagot 2007; Byosiere et al. 2017; Fuss and Schluessel 2017; Miletto Petrazzini et al. 2017). The homing pigeon, Columba livia, instead, has been reported to perceive the Ebbinghaus–Titchener illusion in a reversed fashion compared to humans, estimating a target circle surrounded by large circles as larger (Nakamura et al. 2014). Interestingly, in the chicken, Gallus gallus, two distinct studies have reported evidence of susceptibility to visual illusions in the same direction and in the opposite direction relative to humans, respectively (Rosa Salva et al. 2013; Nakamura et al. 2014).
To date, the origin of the observed variability of susceptibility to size illusions remains unclear. It has been proposed that such variability might actually hinge on research methodology (Feng et al. 2017). With respect to this issue, Parrish et al. (2015) found that, in monkeys, the perception of the Delboeuf illusion is influenced by the procedure adopted in the task and by the type of stimuli. When monkeys had to discriminate the relative sizes of two target dots, they did not demonstrate susceptibility to the illusion, perhaps because they also based their choices on the sizes of the rings surrounding the two target dots. Conversely, monkeys tested in an absolute size discrimination task, in which they had to classify a target dot as “large” or “small”, showed sensitivity to the Delboeuf illusion as reported for humans.
Apart from methodological issues, the observed variability in susceptibility to size illusions might be indicative of species-specific perceptual mechanisms that have evolved according to differential ecological demands (Feng et al. 2017). For instance, the perception of many visual illusions requires overall perception of the target and its surrounding context. However, animals may differ in the precedence of global or local features when perceiving the world around them. Chimpanzees display a rather robust global-to-local precedence (e.g., Fujita and Matsuzawa 1990; Hopkins 1997) while pigeons have been shown to have a local-to-global precedence (Cavoto and Cook 2001). Moreover, significant individual differences in global-to-local precedence have been reported within species (Pitteri et al. 2014), thereby complicating comparative investigation of visual illusions. Insights into the role of ecological adaptation may show up in the context of testing species with an ecology that allows the formulation of hypotheses on the susceptibility to size illusions. For example, it is conceivable that species more often required to discriminate sizes for key fitness-related activities might have been selected for accurate size perception and be not susceptible to size illusions.
Here, we investigated the susceptibility to the Delboeuf illusion in a teleost fish, the guppy, Poecilia reticulata. The guppy is an interesting species for the study of size illusions because it relies on size discrimination for many fitness-related decisions. First, under threat of predation, guppies exploit the size-association strategy, which requires the ability to discriminate between social companions, in terms of size (Croft et al. 2009). Individual guppies join social groups formed by conspecifics with a body size similar to their own. This increases individual survival because predators attacking groups with same-sized individuals show reduced rates of capture. Second, vision plays an important role in guppies’ foraging (Rodd et al. 2002; Birk and White 2014); this species has been reported to make very accurate size discriminations while choosing among food items (Lucon-Xiccato et al. 2015; Lucon-Xiccato and Dadda 2017a), demonstrating the ability to discriminate between food items with a 0.75 size ratio, a threshold unattained even by some mammals observed in similar tasks (cats: Bánszegi et al. 2016; dogs: Ward and Smuts 2007; horses: Uller and Lewis 2009; prosimians: Santacà et al. 2017). It is for the sake of reproduction, that guppies have shown their outstanding ability to discriminate among sizes. Males are characterised by a variable pattern of orange and black spots on body flanks (Houde 1997). Females precisely estimate the size of males’ spots to choose among available mates (Houde 1997; Magurran 2005), managing to distinguish even subtle variations in male spots due to different diets (Karino and Shinjo 2004). Producing orange and black spots is costly for males; thus, males that express more colours are in better condition (Rahman et al. 2013). As genes related to condition are hereditary in this species, females choosing more colourful males achieve an indirect genetic advantage that consists of producing offspring with better genes and thus in better condition (Evans et al. 2004). In this species, males contribute to reproduction only via genes and there are not direct benefits of mate choice because males do not provide females with, for example, a territory and do not engage in parental care. Therefore, indirect genetic advantages that result from choosing colourful males are used by females to increase their reproductive fitness (Houde 1997; Magurran 2005). Moreover, several males show a Delboeuf-like pattern in colour spots with a black spot surrounded by an iridescent spot (Fig. 1a). Iridescent spots are not costly for males, at least in some guppy populations (Devigili et al. 2012; Rahman et al. 2013). This pattern might cause females to overestimate the size of the central, black spot. Though the discrimination of colour spots is less studied in males, there is evidence to suggest that they perform such discrimination to evaluate rivals (Gasparini et al. 2013). Last, females also base their reproductive choice on male tail size, showing a preference for males with larger tails (Bischoff et al. 1985), which may also provide selective pressure for enhanced size discrimination abilities.

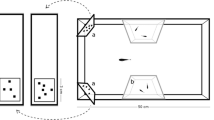
a Example of the colour pattern displayed by male guppies in which the central, black spot is surrounded by an iridescent spot, b view from above of the apparatus used in the two experiments, and c perspective of a subject approaching the two stimuli during the experiment
To investigate guppies’ susceptibility to the Delboeuf illusion, we adopted two methodological approaches commonly used in comparative perception studies: a discrimination learning task with two-dimensional objects as stimuli (Parrish et al. 2015; Byosiere et al. 2017) and a spontaneous choice task with food items as stimuli (Parrish and Beran 2014; Santacà et al. 2017). We chose this twofold approach because several studies have highlighted the importance of using multiple tasks to investigate a given species’ cognitive characteristics (Prétôt et al. 2016a, b; Gatto et al. 2017).
Experiment 1—discrimination learning
Experiment overview
We initially trained guppies to select the larger of two differently-sized target circles surrounded by concentric rings (training phase). The ratio between the areas of the two target circles was 1:0.5, calculated as ‘area of the larger circle/area of the smaller circle’. In the test phase, which took place after that the guppies attained the learning criterion, we presented guppies with both discrimination trials (three types of trials, with area of the stimuli differing by a ratio 1:0.5, 1:0.66 or 1:0.75) and illusory trials.
If fish perceive the Delboeuf illusion as humans do, we predicted that their size-judgment performance would be impacted by the outer ring size in two different ways. First, in some of the discrimination trials, we presented a condition in which a large ring circumscribed the larger target circle and a small ring circumscribed the smaller target circle (hereafter ‘congruent trials’; see Fig. 2). This pattern was expected to facilitate an assimilation effect of the smaller target circle with the surrounding small ring, and a contrast effect of the larger target circle in the larger surrounding circle. Both effects were expected to reduce the subjective size difference of the two target circles. In other discrimination trials (hereafter ‘incongruent trials’), the condition was reversed, with a large ring that circumscribed the smaller target circle (a condition that facilitates the contrast effect) and a small ring that circumscribed the large target circle (assimilation effect). This is supposed to increase the subjective size difference between the two target circles. Hence, if guppies are susceptible to the Delboeuf illusion, one might expect them to be facilitated in size judgments in incongruent trials compared to the congruent trials (Prediction 1; see Parrish et al. 2015). Second, for the illusory trials, the stimuli were two same-sized target circles surrounded by concentric rings which differed in size. The stimuli of the illusory trials produce a size illusion in humans, causing the target circle surrounded by the larger ring to be perceived as smaller. If guppies—trained to select the larger circle—were susceptible to the Delboeuf illusion as humans are, then one would expect them to choose the target circle surrounded by the smaller ring in illusory trials (Prediction 2).

Examples of the stimuli used in the experiments. a In experiment 1, we used white cards with printed orange target circles. b In experiment 2, we used cards with a black background, displaying a white circle and a piece of food pasted in the middle of it
Materials and methods
Experimental subjects
We used 12 female and 12 male guppies, which were adult (6–8-months old) at the time of the experiment. We collected these guppies from a large (> 5000), self-sustaining population in an artificial warm-water pond in Padova, Italy. This population descends from wild guppies collected from the lower Tacarigua River (Trinidad) in 2002. Two months before the start of the experiment, we collected the subjects and moved them to the laboratory, in 400-L tanks. We provided these maintenance tanks with natural vegetation (Hygrophila corymbosa and Taxiphyllum barbieri) and a gravel bottom, and we kept a 12:12-h light:dark photoperiod. Biomechanical filtres and a heating system set at 26 ± 1 °C maintained a constant water condition. We fed the guppies three times per day, alternating commercial food flakes and live brine shrimp, Artemia salina, nauplii.
Apparatus
Like previous studies, we tested each subject in a 20 × 50 cm glass tank (Dadda et al. 2015; Lucon-Xiccato et al. 2015; Lucon-Xiccato and Dadda 2017a; Fig. 1b, c). The tank contained 28 cm of water and had a bottom made of gravel. An air stone aerated the water; we removed the air stone during the trials to avoid affecting the choice of the subjects. Green plastic panels covered the long walls of the tank and white panels covered the short walls, where we presented the stimuli during the trials. Two transparent plastic panels formed a pair of trapezoidal lateral compartments (10 × 5 × 25 cm each) in the middle of the tank. Each lateral compartment housed plants and two social companions. The social companions were immature fish because we have previously observed that immature fish interact less with the subjects than adult fish, and thereby do not interfere with the experiments. To further minimise social deprivation and provide adult social stimuli, we also placed one 28 × 5 cm mirror in each lateral compartment, as guppies seem to perceive their mirror images as other fish (Cattelan et al. 2017; Lucon-Xiccato and Dadda 2017b). Holes on the transparency allowed for water exchange between the lateral compartments and the main compartment of the tank. Because of the lateral compartments, the central portion of the tank formed a narrow 10-cm corridor; this corridor forced the subject to swim in the middle of the tank while approaching the stimuli (see Procedure). We placed a 15-W fluorescent lamp oriented downward above the apparatus to provide illumination (12:12-h light:dark cycle). We performed the experiment in a dark room. Because of the plastic walls of the tank and the strong light above the water surface, we expected the guppies to be unable to see the experimenter and not be affected during their choice.
Stimuli
We used orange target circles surrounded by black concentric rings as stimuli. We chose orange target circles because guppies are attracted by orange objects (Rodd et al. 2002) and this was expected to improve subjects’ performance (Lucon-Xiccato and Bisazza 2014; Gatto et al. 2017). We drew the stimuli using Microsoft Office Software to precisely determine the size of each circle and ring (measured as the diameter by the software). Using a spreadsheet, we calculated the area of the circles and the rings from the diameter, and paired stimuli according to ratios useful for the experiment (see below). We printed the stimuli on 3 × 3 cm white cards. During the trials, we presented two stimuli to the subject by affixing each card to the terminal part of a transparent panel (3.5 × 15 cm). An L-shaped support allowed the panel to be placed on the short wall of the tank.
We used stimuli with different combinations of target circles and surrounding rings across the different phases of the experiment. In each trial of the training phase, the stimuli were two differently sized target circles (ratio between areas of the two circles 1:0.5; Fig. 2a). In half of the trials, we used identical surrounding rings for both stimuli (either large or small surrounding rings). In the remaining half of the trials, we used rings with different size: in particular, in one quarter of the trials, we used a large ring that surrounded the larger target circle and a small ring that surrounded the smaller circle; in the remaining quarter of the trials, we used a large ring that surrounded the smaller circle and a small ring that surrounded the larger circle. The diameter of the circles and the rings varied from 0.57 to 1.17 cm and from 1.58 to 2.35 cm, respectively.
During the test phase, we presented subjects with a discrimination task with four types of trials (Fig. 2a), alternating trials of each type according to a predetermined pseudo-random schedule. In one type of trials (60 trials overall), the stimuli differed in size with a 1:0.5 ratio, as in the training phase. We also presented two more challenging types of trials: 20 trials with 1:0.66 ratio between the sizes of the stimuli and 20 trials with 1:0.75 ratio between the sizes of the stimuli. We arranged stimuli for these latter two types of trials in four conditions: a condition with a large ring encompassing the larger target circle and a small ring encompassing the smaller target circle (congruent trials), and another condition with a large ring surrounding the smaller target circle and a small ring surrounding the larger target circle (incongruent trials). The congruent and incongruent trials allowed us to analyse the impact of the outer rings on the subjective size estimation of target stimuli with true physical differences (Prediction 1; Parrish et al. 2015). The diameter of the target circles and the rings used in these discriminations varied from 0.70 to 1.19 cm and from 1.52 to 2.35 cm, respectively. The fourth type of trials presented in the test phase consisted of 20 illusory trials, in which the size of the two target circles was identical, but the rings varied. The diameter of the circles and the rings used in the illusory trials varied from 1.22 to 1.47 cm and from 1.5 to 2.82 cm, respectively.
Procedure
We adapted a procedure previously used to study trained numerical abilities in guppies (Dadda et al. 2015) that consisted of three phases: a habituation phase, a training phase and a test phase. For the habituation phase, we placed each individual subject in an experimental tank with four immature social companions for a period of 2 days (we removed the companions before the beginning of the test phase). During the first day of habituation, we fed the fish eight times with a procedure simulating the one of the following phases: we placed a single transparent panel with a card randomly chosen from the set of the stimuli in front of one short side of the tank (alternating the two sides between trials). Using a Pasteur pipette, we delivered a small amount of food (brine shrimp nauplii) to the subject when it approached the card. During the second day of habituation, we performed 12 habituation trials using two cards with differently sized target circles. Here, we pasted the food reward (a small piece of commercial flake) on the panels holding the card, above both stimuli, above water level. We also affixed the reward on the panel with the wrong stimulus to prevent the fish from using the visual food cue to select the correct stimulus. We did not expect the olfactory cues afforded by the food to affect fish behaviour because the food was outside the water when guppies were making the choice and also because the food was present above both stimuli. After placing the panels in the water, we refrained from touching them to avoid affecting the subject’s choice. When the subject approached (swam at less than 1 body length) the card with the larger target circle, we delivered the food by immersing the panel with the larger target circle in the water. We simultaneously and gently removed the panel with the smaller target circle. In the event that the subject selected the smaller stimulus, we continued the trials until it corrected its choice and obtained the food reward. We counterbalanced both the relative left/right position of the two stimuli and the use of the two sides of the tank to present the stimuli across trials.
In the training phase, we administered 12 daily trials (6 in the morning and 6 in the afternoon) for a period of up to 10 days with the size of the stimuli differing with a 1:0.5 ratio. We conducted the trials as described for the second day of the habituation phase, with a single exception: if the subject chose the smaller stimulus, we removed both cards and did not provide a reward. We set the learning criterion at 70% correct choices over two consecutive days (17 out of 24 trials). This learning criterion was statistically significant (chi-squared test: χ2 = 4.167, P = 0.042) and previously used for experiments involving this species (e.g., Miletto Petrazzini et al. 2018).
In the test phase, we collected data aimed at the analysis of the sensitivity to the illusion. We administered 12 trials per day for 10 days, following the procedure laid out in the training phase. We randomly alternated the four types of stimuli (discrimination trials with 1:0.5, 1:0.66 or 1:0.75 ratios between the sizes of the two target circles, and illusory trials) and the conditions of the discrimination trials (congruent and incongruent trials). We administered the food reward in response to correct choices in the trials with 1:0.5 ratio between size of the target circle but not in the other trials. Undergraduate students unaware of the study hypothesis collected the data. A female subject ceased to participate after 5 days of the test phase; we analysed its performance only up to that point.
Statistical analysis
We analysed the data in R version 3.4.0 (The R Foundation for Statistical Computing, Vienna, Austria, http://www.r-project.org). Initially, we performed a repeated measures analysis using a generalised mixed-effects model for binomial data on the choice of the larger stimulus in the discrimination trials and on the choice of the stimulus perceived as larger by humans in the illusory trials. In this model, we tested for the effect of the day of training to ensure that the subjects’ performance was constant, for the effect of the sex, and for the effect of the type of stimuli (discrimination trials with ratios between sizes of 1:0.5, 1:0.66 and 1:0.75, and illusory trials). Then, we used one-sample t tests to compare the choice of the larger target circles and, for the illusory trials, the choice of the target circle that humans perceive as larger with chance (50%) in the entire test phase. A one-sample t test on the illusory trials was also performed on the first day of the test phase only; this latter analysis was based on two choices per fish and could, therefore, have reduced power due to binomial error but was useful in case of large effect sizes to confirm our findings. We also tested for correlation between individual performance in the illusory trials and in the discrimination trials with differently sized target circles using Pearson correlation test. We checked the data for normality before conducting these analyses.
Results
The repeated measures analysis showed that the performance of guppies was stable across the 10 days of training (χ29 = 8.838, P = 0.452) and did not vary as a function of sex (χ21 = 0.071, P = 0.790). The performance of guppies varied according to the type of trial (discrimination trials with 1:0.5, 1:0.66 and 1:0.75 ratios between sizes, and illusory trials; χ23 = 18.008, P < 0.001). Tukey post hoc test revealed no significant difference between the guppies’ performance in the trials of the three size discriminations (1:0.5 versus 1:0.66 ratio: P = 0.894; 1:0.5 versus 1:0.75 ratio: P = 0.860; 1:0.66 versus 1:0.75 ratio: P > 0.999), but in the illusory trials, the guppies’ choice of the stimulus that humans perceive as larger was significantly lower than the choice of the larger stimulus in the size discrimination trials (versus 1:0.5 ratio: P < 0.001; versus 1:0.66 ratio: P = 0.021; versus 1:0.75 ratio: P = 0.025). The repeated measures analysis also showed a significant sex × day interaction (χ29 = 21.931, P = 0.009). There was no impact of sex on the perception of the illusion because the sex × type of trial and the day × sex × type of trial interactions were not significant (χ23 = 1.532, P = 0.675; and χ227 = 34.568, P = 0.150, respectively). The day × type of trial interaction was not significant (χ227 = 21.604, P = 0.757).
Data of individual subjects from experiment 1 are shown in Table 1. In the test trials in which the stimuli were circles with 1:0.5 ratio between the size, guppies chose the larger stimulus more often than expected by chance (one-sample t test: t23 = 6.832, P < 0.001; Fig. 3). Guppies also chose the larger target circle in the two novel and unrewarded types of trial with a difference between the size of the target circle (0.66 size ratio: t23 = 2.967, P = 0.007; 0.75 size ratio: t23 = 2.905, P = 0.009; Fig. 3), indicating that they were able to generalise the rule learned in the training phase, even when confronting with novel and harder to identify ratios.

Results of experiment 1. Percentage of choice (mean ± standard error) for the larger target circle in the discrimination between target circles with different sizes and percentage of choice for the target circle that humans perceive as larger
When we considered the discrimination trials with 1:0.66 and 1:0.75 ratios with different surrounding circles, we found that guppies did not show a significant preference for the larger stimulus in the incongruent trials (mean ± standard deviation: 55.83 ± 17.42% choices for the larger stimulus; one-sample t test: t23 = 1.640, P = 0.115); conversely, guppies showed a significant preference for the larger stimulus in the congruent trials (59.00 ± 12.85%; t23 = 3.415, P = 0.002). This result indicated that the presence and size of the surrounding circles influenced the guppies’ choice of the larger stimulus, thereby suggesting susceptibility to the Delboeuf illusion (Prediction 1), though in a reverse direction relative to the illusion as it has been described for humans.
In the illusory trials (Prediction 2), guppies did not show a significant choice for the stimulus that humans perceive as larger (t23 = 1.877, P = 0.073; Fig. 3). The analysis restricted to the first day of the testing phase did not show a significant choice for the stimulus that humans perceive as larger (t23 = 1.000, P = 0.328). The preference for the stimulus that humans perceive as larger did not correlate significantly with the preference for the larger circles in the discrimination trials with 1:0.5, 1:0.66 and 1:0.75 ratio between sizes (1:0.5: r22 = − 0.171, P = 0.426; 1:0.66: r22 = 0.034, P = 0.875; and 1:0.75: r22 = 0.280, P = 0.185).
Experiment 2—spontaneous food choice
Experiment overview
In experiment 2, we exploited the guppies’ spontaneous preference for larger food items to assess their susceptibility to the Delboeuf illusion. In the test phase of experiment 2, we presented guppies with two target food items circumscribed by circles drawn on the background. In some of the trials (size discrimination trials), the food items were differently sized (1:0.66 ratio between surface area) and the surrounding circles were all the same size; here, we expected guppies to choose the larger food item (Lucon-Xiccato et al. 2015; Lucon-Xiccato and Dadda 2017a). In the remaining trials (illusory trials), the food items were identical (in terms of size) but the surrounding circles were differently sized; here, we expected guppies to exhibit a preference for one of the two food items if they perceived any sort of traditional (or reversed) Delboeuf illusion.
Materials and methods
Experimental subjects
We tested 16 adult female guppies of the same strain used in experiment 1; of these fish, 4 did not complete the experiment and we, therefore, did not analyse their performance (final sample size: 12 guppies; see below). Because we used different individual subjects, the data of the two experiments are independent. We did not test males because experiment 1 furnished us with no evidence of sex differences in the perception of the illusion and because a previous study had already reported an absence of sex differences associated with choosing differently sized food items in guppies (Lucon-Xiccato et al. 2015).
Apparatus and stimuli
We performed this experiment in the same apparatus described in experiment 1 (Fig. 1b, c). The stimuli were 3 × 3 cm plastic cards with a black background (as opposed to the rings characterising the stimuli of experiment 1) and a central white circle. The diameter of the central white circle was either Ø 2.2 cm (‘large’) or 0.7 cm (‘small’). The size of the white circle varied across trials to induce illusory stimuli. In the middle of the white circle, we pasted the target, a small piece of food (GVG mix, Sera GmbH, Heinsberg, Germany), by adding a drop of water and letting it dry. We cut the food items with a surgical scalpel guided by circular moulds printed on paper. We used food items of two distinct diameters: 0.43 cm (surface area: 0.145 cm2) and 0.35 cm (surface area: 0.096 cm2).
In the size discrimination trials of the test phase, we used two differently-sized food items, pasted on two cards with same-sized inducer circles. The ratio between the areas of the two food items was 1:0.66, the same ratio used in previous studies on Delboeuf illusion among mammals (Miletto Petrazzini et al. 2017; Santacà et al. 2017). In the illusory trials, we used two same-sized food items (diameter 0.43 cm) pasted on different sized surrounding circles (Fig. 2b).
According to this experimental design, the stimuli in the illusory trials differed for the surrounding circle sizes, whereas the other stimuli had always an identical surrounding circle. In case of a preference for one of the two stimuli in the illusory trials, guppies might have made a choice based only on the size of the surrounding circle, regardless of the size of the food item. To rule out this possibility, we introduced a control phase at the end of experiment 2: we presented guppies with control trials with the external circles used in the illusory trials, but in the absence of food items. We alternated these control trials with the size discrimination trials as in the previous phase.
Procedure
Following previous studies that relied on this paradigm (Lucon-Xiccato et al. 2015; Lucon-Xiccato and Dadda 2017a), we used a three-step procedure (habituation, test and control phase). Seven days before the experiment began, we moved each subject into an experimental tank together with the social companions (see “Experiment 1—discrimination learning”). For 3 days, we fed the fish three times per day from alternating short sides of the tank using a Pasteur pipette. On the 4th day, we fed the subjects four times; in these trials, we presented an empty stimulus card right before administering the food. On days 5 and 6 of the habituation phase, we fed the subjects six times per day using one card with small pieces of food pasted onto it. We employed a longer period of habituation relative to experiment 1 to habituate the subjects to touching the card and eating the food pasted onto it. Four fish did not become habituated to feeding from the card; we discarded them from the experiment and substituted with new subjects. We did not feed the subjects on the last day of the habituation phase to motivate them to perform the trials in the test phase that followed.
In the test phase, we tested guppies for 48 trials subdivided across 6 days. Overall, each guppy participated in 32 size discrimination trials and 16 illusory trials. We determined the order of these trials according to a pseudo-random schedule. During each trial, we simultaneously placed two cards in one short side of the tank and left subjects free to select one of them. We removed the card that the subject did not choose and left the chosen card in the water until the fish consumed the food. We scored the subject’s choice as the first card that it touched.
The control phase started 1 day after the test phase. Guppies underwent 64 trials subdivided over 8 days. We administered 32 discrimination trials and 32 control trials following the procedure of the previous phase. In the control trials, we scored the subject’s choice as the first card that it touched. One subject stopped participating during the control phase; we, therefore, evaluated its performance only up to the test phase. Other procedural details were identical to those of experiment 1.
Statistical analysis
We performed the analysis similarly to our analysis of experiment 1: we used a repeated measures analysis to study the trend in performance across the days of the test phase, followed by one-sample t tests to compare the choice of a stimulus with chance level (50%), a paired-sample t test to compare the choice among the different types of stimuli, and a Pearson correlation to compare individual performance between discrimination trials and illusory trials.
Results
Test phase
The repeated measures analysis showed that the performance of guppies was stable across the days of the test phase (χ25 = 0.536, P = 0.991). There was a significant effect of trial type (χ21 = 82.123, P < 0.001): guppies chose the stimulus with the larger target circle in the discrimination trials more frequently than they chose the stimulus that humans perceive as larger in the illusory trials (Fig. 4a). The interaction between day and type of trial was not significant (χ25 = 3.180, P = 0.672).

a Results of experiment 2. Percentage of choice (mean ± standard error) for the larger food item in the discrimination task with 1:0.66 ratio between sizes and percentage preference for the food item that human perceive as larger in the illusory trials. b Results of control phase of experiment 2: percentage of choice (mean ± standard error) for the larger food item in the discrimination task with 1:0.66 ratio between sizes and percentage of choice for the larger surrounding circle in control trials
Data of individual subjects from experiment 2 are shown in Table 2. In the size discrimination trials (ratio between the size of the stimuli 1:0.66), guppies showed a significant preference for the larger food item (one-sample t test: t11 = 12.202, P < 0.001; Fig. 4a). We found that guppies chose the larger food item more frequently when the surrounding circle was small (choice for the larger item with small surrounding circle: 71.35 ± 9.02%; with large surrounding circle: 60.94 ± 7.60%; paired-sample t test: t11 = 2.600, P = 0.025).
In the illusory trials, guppies showed a marked preference for the food item surrounded by the larger circle, i.e. for the stimulus that humans perceive as smaller (t11 = 6.611, P < 0.001; Fig. 4a). This result was additionally confirmed in the analysis of the first test day (t11 = 5.745, P < 0.001), which indicated a particularly strong effect that could be detected even with a small number of trials. The preference for the stimulus that humans perceive as smaller was negatively correlated with the preference for the larger food item in the discrimination trials (r10 = − 0.620, P = 0.031), which indicated that individual guppies with greater size discrimination performance were more sensitive to the illusory effect.
Control phase
The repeated measures analysis showed that the performance of guppies was stable across the days of the control phase (χ27 = 0.916, P = 0.996). There was a significant effect of trial type (χ21 = 5.550, P = 0.019): guppies chose the larger food item in the discrimination trials more often than they chose the stimulus with the smaller circle (Fig. 4b). The interaction between day and type of trial was not significant (χ27 = 3.422, P = 0.843).
In the size discrimination trials (ratio between the size of the stimuli 1:0.66), guppies showed a significant preference for the larger food item (one-sample t test: t10 = 8.181, P < 0.001; Fig. 4b). In the control trials (without food items), guppies did not show any significant preference, though they tended to choose the stimulus with the smaller circle (t10 = 2.014, P = 0.072; Fig. 4b). This excluded the possibility that the results of the test phase were due to any sort of spontaneous bias for the surrounding circle itself.
Discussion
There is an increasing evidence of substantial interspecific variability in susceptibility to size illusions, but the causes of this variability remain unknown. We investigated susceptibility to the Delboeuf illusion in the guppy, a species that performs very accurate size discrimination for fitness-related decisions and thus might not be susceptible to size illusions. Our study indicates that guppies are susceptible to the Delboeuf illusion, but that the direction of such size misperception is reversed relative to humans; guppies seem to perceive as larger the stimulus that humans and non-human primates perceive as smaller (e.g., Parrish and Beran 2014; Parrish et al. 2015). This finding suggests that interspecific variability in size illusion perception may be unrelated to the need to accurately perform size discrimination tasks of a given species.
In experiment 1 (discrimination learning), guppies learned to discriminate between the stimuli used during the training phase (circles with 1:0.5 ratio between surface areas). In the test phase, guppies also succeeded at the two more challenging types of trial (1:0.66 and 1:0.75 ratio between areas), which squares with the finding of a previous study (Lucon-Xiccato et al. 2015). In the congruent and incongruent size discrimination trials of the test phase, we found evidence that guppies showed a facilitation effect due to the Delboeuf illusion (Prediction 1) in the reversed direction relative to humans. Guppies chose the larger target circle at a rate above chance when it was surrounded by a large ring and the small target circle was surrounded by a small ring; conversely, guppies failed to discriminate between stimuli when the larger target circle was surrounded by a small ring and the smaller target circle was surrounded by a large ring. In the illusory trials of the test phase (Prediction 2), in which the two target circles were the same size, we found no significant preference both in the entire sample of subjects and in a tentative analysis at the individual level. It should be noted that, on this point, our analysis might not be conclusive. It is possible that guppies have shown a preference for the target circle surrounded by the larger circle, as we would have expected in case of a reversed Delboeuf illusion, but we lacked sufficient statistical power to reject the null hypothesis. Indeed, the group statistic P value approached the threshold for statistical significance (P = 0.073). Moreover, in the individual-level analysis, a fish should have chosen a stimulus in 15 out of 20 trials (75%) to achieve data indicating a significant preference. This choice accuracy is uncommon in fish experiments, and guppies showed it only when required to discriminate between stimuli with areas that differed according to a 0.5 ratio (Lucon-Xiccato et al. 2015). Because the illusory effect was not expected to be strong enough to make two identical stimuli appear as if one were twice the size of the other, the individual-level analysis was perhaps unlikely to detect a significant result using this experimental design.
One could argue that, in experiment 1, fish made their choices on the basis of the size of the concentric rings, as monkeys evidently did when tested on a similar discrimination task (Parrish et al. 2015). This might have yielded an increase in the chance to select the larger circle in the ‘congruent’ condition, as actually did occur. However, if fish relied meaningfully on this cue, we should also have found a preference for the larger ring in the incongruent condition, but this was not observed in our study. Hence, although we cannot exclude the possibility that this cue played a role in our training phase, we do not believe that it can account for our data on its own. As advanced in a study with baboons (Parron and Fagot 2007), one may alternatively suppose that subjects only focused on the target stimuli, virtually ignoring the outer rings, and thereby explaining the non-significant preference in the presence of the illusory pattern with identical targets. Although it is possible that the attention of some subjects was largely focused on the target stimuli, the low performance demonstrated in the incongruent condition does not support this hypothesis.
Regarding sex differences, one might expect that females would demonstrate smaller susceptibility to size illusions relative to males because females of this species must be able to perform accurate size discriminations more frequently than males do (i.e., during mate choice). However, the results of experiment 1 seem to exclude the presence of sex differences in the perception of the illusion.
In experiment 2 (spontaneous food choice), guppies demonstrated a preference for the larger food item (1:0.66 ratio). The accuracy in choosing the larger food item (66%) was greater compared to the accuracy in choosing the larger target circle in experiment 1 (57%). Because we used identical apparatuses in the two experiments, this difference may be due to individual differences between the subject samples used in the different experiments (Lucon-Xiccato and Bisazza 2017) or to the type of stimuli. In particular, the food stimuli used in experiment 2 might be more salient for guppies than the two-dimensional objects printed on the cards used in experiment 1. In the illusory trials of experiment 2, guppies showed a marked preference for the food item surrounded by the larger circle. The control phase of experiment 2 excluded that guppies focussed on the external ring during their choice. Further, individual-level analysis found four subjects that significantly choose the food item surrounded by the larger circle, despite the high threshold for significance in this experiment. Moreover, seven of the eight remaining subjects showed a preference that was similar in direction but not as strong. The illusory effect seems substantially greater than that observed in experiment 1. This might be due to the stimuli’ greater salience in experiment 2; alternatively, guppies might use different strategies to discriminate between objects (experiment 1) and food items (experiment 2; Lucon-Xiccato et al. 2015), and distinct strategies might account for the differential illusory effect.
Considering both experiments, we found two significant pieces of evidence that guppies are susceptible to a reverse Delboeuf illusion (comparison of congruent and incongruent trials in experiment 1 and analysis of illusory trials in experiment 2) and a third analysis that, while not conclusive, was not significant but apparently consistent with the other two (illusory trials of experiment 1). Therefore, we believe that the most reasonable interpretation of the whole study indicates that guppies perceive (and are, therefore, susceptible to) a reversed Delboeuf illusion. It is worth remembering that we used two methodological approaches, one that employed a training procedure (experiment 1) and another that relied on a free choice test (experiment 2); furthermore, the type of stimuli was different (two-dimensional objects in experiment 1 and food items in experiment 2). Using different methods to test the same hypothesis is important to ensure that the results are unaffected by the methodology adopted (Agrillo and Miletto Petrazzini 2012). In this sense, the relatively analogous results between the two experiments here reportedly strengthen the claim of susceptibility to a reversed Delboeuf illusion in this species.
One would hardly anticipate the susceptibility to the Delboeuf illusion in guppies because of the importance of accurate size discrimination in the guppies’ ecology: one would instead expect to find that guppies were not susceptible to the illusion because this would increase size estimation accuracy. Hence, our results do not support the hypothesis that the ecological demand for high accuracy in size discrimination causes selection against sensitivity to illusions. The reversed illusion was even more unexpected considering the literature on other teleost fish species. In the other teleosts for which the Delboeuf illusion has been investigated, the damselfish, Chromis chromis, three out of the five subjects were found to perceive the illusion in the same direction that humans did, whereas the remaining two subjects showed a similar, though not significant, trend (Fuss and Schluessel 2017). Furthermore, damselfish and another teleost fish, the redtail splitfins, Xenotoca eiseni, showed a human-like perception of a closely-related size illusion, the Ebbinghaus–Titchener illusion (Sovrano et al. 2015; Fuss and Schluessel 2017). The comparison between these prior studies and the present study seems to indicate the existence of species-specific differences in the susceptibility to size illusions within teleost fish, which squares with the broad variation previously observed in warm-blooded vertebrates (Feng et al. 2017).
Reversed perception of size illusions has also been identified in birds (Nakamura et al. 2008, 2014; Watanabe et al. 2011, 2013). These results have often been interpreted in the light of different sensitivities to assimilation-contrast effects. Indeed, in humans the illusion is believed to be due to a combination of assimilation and contrast effects (King 1988). When the ring is close to the target circle ‘X’, then the ring is meant to assimilate, leading to a perception of ‘X’ as larger; whereas when the ring is far from the target circle ‘Y’, then the ring is meant to contrast, leading to an underestimation of ‘Y’ in size. It has been proposed that some bird species are more sensitive to assimilation effects (Watanabe et al. 2011), and thereby perceive a reversed illusion (as the stimulus assimilated to the larger ring is meant to appear larger than the one assimilated to the smaller ring). It is possible that these perceptive mechanisms are under selective pressures other than those promoting accurate size discrimination, and this caused the results observed in guppies. Alternatively, it is worth noting that the assimilation mechanism adopted to solve the task might also depend on the stimuli and the experimental procedure, as evidenced by the presence of conflicting results between different studies of the same species (Nakamura et al. 2014; Rosa Salva et al. 2013).
Another novel finding of experiment 2 was that the sensitivity to the illusion of each subject was related to its ability to solve the size discrimination task. In humans, it has been often reported that individuals exhibit different cognitive performance and that these individual differences correlate across similar tasks (Hunt et al. 1973). Conversely, in fish, the study of individual differences is less developed (for a recent review, see Lucon-Xiccato and Bisazza 2017) and there are no clear results regarding the perception of visual illusions. Interestingly, three independent studies (two on guppies and one on another poeciliid fish) have reported that the performance of individual fish covaries across different quantity discrimination tasks, including a food size discrimination task (guppies: Miletto Petrazzini and Agrillo 2016; Lucon-Xiccato and Dadda 2017a; Gambusia affinis: Etheredge et al. 2018). These results seem analogous to those of experiment 2 and can be interpreted as an indication of the relevance of individual differences in the ability of fish to process quantity information that also affects the perception of size illusions. This effect clearly requires confirmation by studies specifically aimed at investigating individual differences. These studies should test the same set of individuals in multiple tasks, such as with diverse size illusions and/or with different versions of the same illusory pattern, to assess whether the within-individual variance in performance explains a substantial proportion of the total variance in performance relative to the variance between individuals.
In conclusion, our study has provided the first evidence of reversed perception of a visual illusion in a fish species. Future studies should try to confirm this susceptibility to size illusion in guppies using different illusory patterns and different paradigms, such as those based on a single choice that allow testing a large number of subjects. Critically, future studies should also try to understand the causes of interspecific and intraspecific variability in size illusions. For example, it would be interesting to compare individuals of the same species that have experienced different developmental conditions to test whether intraspecific variability is due to ontogenetic factors. Alternatively, it would be compelling to study species with different ecological demands to test hypotheses based on the selective pressures that might act on the perceptual mechanisms for processing size information.
References
Agrillo C, Miletto Petrazzini ME (2012) The importance of replication in comparative psychology: the lesson of elephant quantity judgments. Front Psychol 3:181
Bánszegi O, Urrutia A, Szenczi P, Hudson R (2016) More or less: spontaneous quantity discrimination in the domestic cat. Anim Cogn 19:879–888
Basolo AL (1990) Female preference for male sword length in the green swordtail, Xiphophorus helleri (Pisces: Poeciliidae). Anim Behav 40:332–338
Birk MA, White JW (2014) Experimental determination of the spatial scale of a prey patch from the predator’s perspective. Oecologia 174:723–729
Bischoff RJ, Gould JL, Rubenstein DI (1985) Tail size and female choice in the guppy (Poecilia reticulata). Behav Ecol Sociobiol 17:253–255
Byosiere SE, Feng LC, Woodhead JK, Rutter NJ, Chouinard PA, Howell TJ, Bennett PC (2017) Visual perception in domestic dogs: susceptibility to the Ebbinghaus–Titchener and Delboeuf illusions. Anim Cogn 20:435–448
Cattelan S, Lucon-Xiccato T, Pilastro A, Griggio M (2017) Is the mirror test a valid measure of fish sociability? Anim Behav 127:109–116
Cavoto KK, Cook RG (2001) Cognitive precedence for local information in hierarchical stimulus processing by pigeons. J Exp Psychol-Anim Behav Proces 27:3–16
Croft DP, Darden SK, Ruxton GD (2009) Predation risk as a driving force for phenotypic assortment: a cross-population comparison. Proc R Soc B Biol Sci 276:1899–1904
Dadda M, Agrillo C, Bisazza A, Brown C (2015) Laterality enhances numerical skills in the guppy, Poecilia reticulata. Front Behav Neurosci 9:285
Devigili A, Kelley JL, Pilastro A, Evans JP (2012) Expression of pre-and postcopulatory traits under different dietary conditions in guppies. Behav Ecol 24:740–749
Dowds BM, Elwood RW (1983) Shell wars: assessment strategies and the timing of decisions in hermit crab shell fights. Behaviour 85:1–24
Etheredge RI, Avenas C, Armstrong MJ, Cummings ME (2018) Sex-specific cognitive–behavioural profiles emerging from individual variation in numerosity discrimination in Gambusia affinis. Anim Cogn 21:37–53
Evans JP, Kelley JL, Bisazza A, Finazzo E, Pilastro A (2004) Sire attractiveness influences offspring performance in guppies. Proc R Soc B Biol Sci 271:2035–2042
Feng LC, Chouinard PA, Howell TJ, Bennett PC (2017) Why do animals differ in their susceptibility to geometrical illusions? Psychon Bull Rev 24:262–276
Fujita K, Matsuzawa T (1990) Delayed figure reconstruction by a chimpanzee (Pan troglodytes) and humans (Homo sapiens). J Comp Psychol 104:345–351
Fuss T, Schluessel V (2017) The Ebbinghaus illusion in the gray bamboo shark (Chiloscyllium griseum) in comparison to the teleost damselfish (Chromis chromis). Zoology 123:16–29
Gasparini C, Serena G, Pilastro A (2013) Do unattractive friends make you look better? Context-dependent male mating preferences in the guppy. Proc R Soc B Biol Sci 280:20123072
Gatto E, Lucon-Xiccato T, Savaşçı BB, Dadda M, Bisazza A (2017) Experimental setting affects the performance of guppies in a numerical discrimination task. Anim Cogn 20:187–198
Hopkins WD (1997) Hemispheric specialization for local and global processing of hierarchical visual stimuli in chimpanzees (Pan troglodytes). Neuropsychol 35:343–348
Houde AE (1997) Sex, color, and mate choice in guppies. Princeton University Press, Princeton
Hunt E, Frost N, Lunneborg C (1973) Individual differences in cognition: a new approach to intelligence. Psychol Learn Motiv 7:87–122
Karino K, Shinjo S (2004) Female mate preference based on male orange spot patterns in the feral guppy Poecilia reticulata in Japan. Ichthyol Res 51:316–320
Kelley LA, Kelley JL (2014) Animal visual illusion and confusion: the importance of a perceptual perspective. Behav Ecol 25:450–463
King DL (1988) Assimilation is due to one perceived whole and contrast is due to two perceived wholes. New Ideas Psychol 6:277–288
Lucon-Xiccato T, Bisazza A (2014) Discrimination reversal learning reveals greater female behavioural flexibility in guppies. Biol Lett 10:20140206
Lucon-Xiccato T, Bisazza A (2017) Individual differences in cognition among teleost fishes. Behav Proc 141:184–195
Lucon-Xiccato T, Dadda M (2017a) Individual guppies differ in quantity discrimination performance across antipredator and foraging contexts. Behav Ecol Sociobiol 71:13
Lucon-Xiccato T, Dadda M (2017b) Personality and cognition: sociability negatively predicts shoal size discrimination performance in guppies. Front Psychol 8:1118
Lucon-Xiccato T, Miletto Petrazzini ME, Agrillo C, Bisazza A (2015) Guppies discriminate between two quantities of food items but prioritize item size over total amount. Anim Behav 107:183–191
Magurran AE (2005) Evolutionary ecology: the Trinidadian guppy. Oxford University Press, Oxford
McNamara JM, Houston AI, Collins EJ (2001) Optimality models in behavioral biology. SIAM Rev 43:413–466
Miletto Petrazzini ME, Agrillo C (2016) Turning to the larger shoal: are there individual differences in small-and large-quantity discrimination of guppies? Ethol Ecol Evol 28:211–220
Miletto Petrazzini ME, Bisazza A, Agrillo C (2017) Do domestic dogs (Canis lupus familiaris) perceive the Delboeuf illusion? Anim Cogn 20:427–434
Miletto Petrazzini ME, Parrish AE, Beran MJ, Agrillo C (2018) Exploring the solitaire illusion in guppies (Poecilia reticulata). J Comp Psychol 132:48–57
Murayama T, Usui A, Takeda E, Kato K, Maejima K (2012) Relative size discrimination and perception of the Ebbinghaus illusion in a bottlenose dolphin (Tursiops truncatus). Aquat Mamm 38:333–342
Nakamura N, Watanabe S, Fujita K (2008) Pigeons perceive the Ebbinghaus–Titchener circles as an assimilation illusion. J Exp Psychol Anim Behav Process 34:375–387
Nakamura N, Watanabe S, Fujita K (2014) A reversed Ebbinghaus–Titchener illusion in bantams (Gallus gallus domesticus). Anim Cogn 17:471–481
Parrish AE, Beran MJ (2014) When less is more: like humans, chimpanzees (Pan troglodytes) misperceive food amounts based on plate size. Anim Cogn 17:427–434
Parrish AE, Brosnan SF, Beran MJ (2015) Do you see what I see? A comparative investigation of the Delboeuf illusion in humans (Homo sapiens), rhesus monkeys (Macaca mulatta), and capuchin monkeys (Cebus apella). J Exp Psychol Anim Learn Cogn 41:395–405
Parron C, Fagot J (2007) Comparison of grouping abilities in humans (Homo sapiens) and baboons (Papio papio) with the Ebbinghaus illusion. J Comp Psychol 121:405–411
Pitteri E, Mongillo P, Carnier P, Marinelli L (2014) Hierarchical stimulus processing by dogs (Canis familiaris). Anim Cogn 17:869–877
Prétôt L, Bshary R, Brosnan SF (2016a) Factors influencing the different performance of fish and primates on a dichotomous choice task. Anim Behav 119:189–199
Prétôt L, Bshary R, Brosnan SF (2016b) Comparing species decisions in a dichotomous choice task: adjusting task parameters improves performance in monkeys. Anim Cogn 19:819–834
Rahman MM, Kelley JL, Evans JP (2013) Condition-dependent expression of pre-and postcopulatory sexual traits in guppies. Ecol Evol 3:2197–2213
Rodd FH, Hughes KA, Grether GF, Baril CT (2002) A possible non-sexual origin of mate preference: are male guppies mimicking fruit? Proc R Soc B Biol Sci 269:475–481
Rosa Salva O, Rugani R, Cavazzana A, Regolin L, Vallortigara G (2013) Perception of the Ebbinghaus illusion in 4-day-old domestic chicks (Gallus gallus). Anim Cogn 16:895–906
Santacà M, Regaiolli B, Miletto Petrazzini ME, Spiezio C, Agrillo C (2017) Preliminary study to investigate the Delboeuf illusion in ring-tailed lemurs (Lemur catta): methodological challenges. Anim Behav Cogn 4:365–377
Sovrano VA, Albertazzi L, Rosa Salva O (2015) The Ebbinghaus illusion in a fish (Xenotoca eiseni). Anim Cogn 18:533–542
Uller C, Lewis J (2009) Horses (Equus caballus) select the greater of two quantities in small numerical contrasts. Anim Cogn 12:733–738
Ward C, Smuts BB (2007) Quantity-based judgments in the domestic dog (Canis lupus familiaris). Anim Cogn 10:71–80
Watanabe S, Nakamura N, Fujita K (2011) Pigeons perceive a reversed Zöllner illusion. Cognition 119:137–141
Watanabe S, Nakamura N, Fujita K (2013) Bantams (Gallus gallus domesticus) also perceive a reversed Zöllner illusion. Anim Cogn 16:109–115
Acknowledgements
We thank the undergraduate students who helped in the project. We have no competing interests. Funding was provided by STARS 2017 Grant (Prot. 19215) and PRIN 2015 Grant (Prot. 2015FFATB7) from University of Padova.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
We have no conflicts of interest.
Ethical approval
We followed all applicable international, national, and/or institutional guidelines for the care and use of animals (Italy, D.L. 4 Marzo 2014, n. 26). All procedures performed in studies involving animals were in accordance with the ethical standards of the institution or practice at which the studies were conducted (Protocol n. 13/2018).
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Lucon-Xiccato, T., Santacà, M., Miletto Petrazzini, M.E. et al. Guppies, Poecilia reticulata, perceive a reversed Delboeuf illusion. Anim Cogn 22, 291–303 (2019). https://doi.org/10.1007/s10071-019-01237-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10071-019-01237-6