
Abstract
A recent study found that guppies (Poecilia reticulata) can be trained to discriminate 4 versus 5 objects, a numerical discrimination typically achieved only by some mammals and birds. In that study, guppies were required to discriminate between two patches of small objects on the bottom of the tank that they could remove to find a food reward. It is not clear whether this species possesses exceptional numerical accuracy compared with the other ectothermic vertebrates or whether its remarkable performance was due to a specific predisposition to discriminate between differences in the quality of patches while foraging. To disentangle these possibilities, we trained guppies to the same numerical discriminations with a more conventional two-choice discrimination task. Stimuli were sets of dots presented on a computer screen, and the subjects received a food reward upon approaching the set with the larger numerosity. Though the cognitive problem was identical in the two experiments, the change in the experimental setting led to a much poorer performance as most fish failed even the 2 versus 3 discrimination. In four additional experiments, we varied the duration of the decision time, the type of stimuli, the length of training, and whether correction was allowed in order to identify the factors responsible for the difference. None of these parameters succeeded in increasing the performance to the level of the previous study, although the group trained with three-dimensional stimuli learned the easiest numerical task. We suggest that the different results with the two experimental settings might be due to constraints on learning and that guppies might be prepared to accurately estimate patch quality during foraging but not to learn an abstract stimulus–reward association.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Many vertebrates possess the ability to discriminate between sets of objects that differ in numerosity (reviewed in Agrillo and Bisazza 2014). This ability may benefit several fitness-related activities, such as detecting the largest available food source (Normand et al. 2009), assessing the number of potential opponents in a contest (Benson-Amram et al. 2011), or choosing the largest social group to increase protection from predators (Hager and Helfman 1991). Although the general mechanisms for numerical processing appear similar among vertebrates (reviewed in Feigenson et al. 2004), different species show different discrimination performance even within the same class. For example, apes can discriminate up to 9 versus 10 items while dogs (Canis lupus familiaris) generally discriminate up to 2 versus 3 items and domestic cats (Felis silvestris catus) can only discriminate up to 2 versus 5 items (Hanus and Call 2007; Bánszegi et al. 2016; Miletto Petrazzini and Wynne 2016).
Inter-species differences in cognitive abilities are usually ascribed to differences in ecology or to the complexity of neural systems (Deaner et al. 2007; Lefebvre et al. 2004; Sherry 2006). However, in many cases this variation could also be attributable to the different procedures that have been used in the different studies (Agrillo and Bisazza 2014). The effect of the procedure is also revealed by the fact that large between-studies differences in cognitive performance are sometimes observed even within the same species. For example, when presented with two sets of food items, the African grey parrot (Psittacus erithacus) selects the larger set in up to 2 versus 3 items (Al Aïn et al. 2009), but with training this species is able to discriminate up to 5 versus 6 items (Pepperberg 2006).
A recent study on guppies (Poecilia reticulata) found that when the length of the training is similar to that typically used in mammals and birds, this fish can discriminate up to 4 versus 5 items (Bisazza et al. 2014). Such numerical accuracy is higher than the maximum accuracy reported for many mammals and birds (reviewed in Agrillo and Bisazza 2014). However, Bisazza et al. (2014) used an experimental setting that deviated considerably from the setting of the classic two-choice discrimination task used with other species. The classic task generally consists in presenting two two-dimensional (2D) stimuli (e.g. sets of dots on a computer monitor) in front of the subject and releasing a food reward when it chooses the correct stimulus (Beran 2008; Emmerton and Renner 2006). Conversely, guppies were presented with two patches of yellow discs lying on the bottom of the test arena, each of which covered a small hole. The holes beneath the discs of the larger patch hid a food reward that the guppies could reach by pecking and dislodging the discs. This procedure was chosen because, in both the laboratory and the field, guppies spend much time pecking and moving small objects on the bottom (e.g. dead leaves and small stones) looking for hidden food (Dussault and Kramer 1981; Rodd et al. 2002).
Exceptional cognitive abilities are usually associated with specific ecological adaptations such as homing in pigeons, food storing in parids and corvids, and vocal learning in parrots. For example, food-storing birds can memorize the location of thousands of food items, but in other contexts their memory performance is similar to that of other birds (Shettleworth and Hampton 1998); among food-storing birds, performance in spatial tasks tends to positively correlate with dependence on cached food (Olson et al. 1995). Apart from these striking examples, biological predispositions to learn or to solve specific problems are extremely diffuse and can affect the comparison of cognitive abilities of the different species with laboratory tasks (Shettleworth 1972; Hinde 1973). For example, adopting a more ecologically relevant procedure dramatically affected the performance of two primate species in a complex learning task (Prétôt et al. 2016). Similarly, pigeons acquired matching- and oddity-to-sample tasks in <30 trials when the procedure exploited their natural foraging habit, but they required many more trials (from 1000 to 2500) to acquire the same task in a Skinner box (Wright and Delius 1994). Inangas (Galaxias maculatus) were unable to associate predation risk with a specific place at a specific time of the day; however, they could promptly solve a similar time–place learning task to obtain a food reward (Reebs 1999).
Because the study by Bisazza et al. (2014) used a training method attuned to guppies’ foraging behaviour, it is not clear whether guppies possess exceptional numerical abilities or whether their remarkable performance was due to a specific predisposition to discriminate between differences in the quality of patches while foraging. In this study, we tried to disentangle these possibilities. In the first experiment, we trained guppies to select the larger quantity in the same numerical discriminations administered by Bisazza et al. (2014), but we used a training setting more similar to that used in experiments with other vertebrates and which was previously used to study discrimination learning in zebrafish (Parker et al. 2012a, b; Proulx et al. 2014; Wang et al. 2015). The stimuli were sets of dots presented on a computer screen in two different chambers. We rewarded the guppies with food when they entered the correct chamber.
If guppies do possess numerical abilities equal or superior to many warm-blooded vertebrates, we expected the same numerical accuracy as Bisazza et al. (2014), even with our training setting, similar to that used in experiments with other vertebrates. As we found a considerable difference between the performances in the two settings, we pursued four further experiments, varying the task parameters to identify the possible factors responsible for the difference.
Experiment 1
In experiment 1, we tested guppies on numerical discriminations of increasing difficulty starting from 2 versus 3, as in the experiment of Bisazza et al. (2014), but with a different training setting, a two-choice discrimination task devised for zebrafish (Parker et al. 2012a, b; Proulx et al. 2014; Wang et al. 2015). To keep the two experiments as similar as possible, we used subjects of same strain, sex, and age, which were maintained under the same conditions until the experiment started.
Materials and methods
Subjects
The subjects were eight adult guppies of an ornamental strain (‘snakeskin cobra green’) bred in our laboratory in the Department of General Psychology (University of Padova, Italy). Bisazza et al. (2014) used only females in their experiment. In some contexts, guppies show sex differences in numerical discrimination abilities (Lucon-Xiccato et al. 2016); to avoid that these sex differences affected the comparison between our experiments and the previous study, here we used only females. The maintenance aquaria (150 L) had a gravel bottom, abundant natural and artificial plants, water filters, and 15-W fluorescent lamps (12:12-h light/dark photoperiod). We kept water temperature at 26 ± 1 °C and fed the fish with commercial food flakes (Fioccomix, Super Hi Group, Ovada, Italy) and Artemia salina nauplii three times per day.
Apparatus
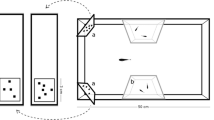
The apparatus resembled the one used in previous works on fish (Parker et al. 2012a, b; Proulx et al. 2014; Wang et al. 2015; Fig. 1). In a 60 × 40 × 38 cm glass tank with gravel bottom and filled with 30 cm of water, we built a central compartment (60 × 16 cm) using green plastic. The two lateral compartments housed abundant natural vegetation and water filters to ensure that the water characteristics and odour were similar to the maintenance tanks. The central compartment also had two trapezoidal sectors made of transparent plastic, containing one immature guppy each as a social companion. We presented the stimuli at one end of the central compartment with an LCD monitor connected to a laptop. Here, we built two ‘choice chambers’ (8 × 9 cm) separated from the central compartment by means of a transparent guillotine door. Two pumps connected to the filters constantly drained water (4 L/min) from the choice chambers to the lateral compartments to remove the olfactory cues of the food reward. Two 15-W fluorescent lamps placed above the tank on the opposite side of the monitor illuminated the apparatus. Outside of the training sessions, we maintained the subjects in a 50 × 20 × 38 cm ‘home tank’ with immature guppies as social companions, vegetation, and water filters.

Aerial view of the experimental apparatus. In experiment 2, an additional guillotine door (not drawn) was used to increase decision time; in experiment 4, an additional fluorescent lamp (not drawn) was placed above the choice chambers to light the 3D stimuli
The stimuli were two sets of black dots on a white background made with Microsoft PowerPoint (Fig. 2a). During trials, we presented each set of dots in correspondence of one choice chamber. Two sets with a different number of objects may also differ by non-numerical attributes that covary with numerosity, and fish might solve the discrimination exploiting these attributes. To prevent this confound, Bisazza et al. (2014) controlled the stimuli for the three most important non-numerical attributes that covary with numerosity (cumulative surface area of the objects, overall space encompassed by the object arrays, and density of the objects) by using discs of different diameter and by displacing the discs on different arrays. We controlled our stimuli following the same scheme. We used dots with different diameter (range 0.3–0.5 cm) to control for the cumulative surface area: in one-third of trials, the ratio between the cumulative surface area of the smaller over the larger set was between 76 and 85 %; in one-third, between 86 and 95 %; and in one-third between 96 and 105 %. Further, we varied the position of the dots in order to equate the overall space encompassed by the two arrays in half of the trials and to equate the density of the dots in the other half of the trials.

Frontal view of the stimuli (above) and aerial view of their position during the trials (below). a In experiments 1, 2, 3, and 5, the stimuli were black dots on a white background projected on a LCD monitor; b in experiment 4, the stimuli were yellow discs fixed on green plastic panels
Familiarization with the apparatus and the procedure
The day before the experiment started, we randomly selected two female guppies from the maintenance tanks and moved them to the apparatus for habituation. In the strain of guppies that we tested, individual females show different fin colouration; we used this characteristic to recognize each individual subject. Only in this phase, we provided three immature guppies in the central compartment to facilitate familiarization with the new environment (Lucon-Xiccato et al. 2015; Miletto Petrazzini et al. 2015). The computer monitor presented a white slide. When one subject swam inside a choice chamber, we supplied crumbled food flakes suspended in water with a Pasteur pipette. Usually, the remaining subject and the social companions rapidly joined to feed on the flakes. After both subjects finished the food and left the choice chamber, we closed the guillotine door and presented a black slide for 20 min. Then, the monitor presented again a white slide to attract the attention of the fish, and we repeated the procedure of above three times. As a consequence of this procedure, the subjects learned to seek food in the choice chambers when the white slide appeared. This resembled the situation of the training trials (see below) in which food could be obtained when the monitor presented the white slides with the stimuli. The subjects stayed all night in the apparatus, and the following morning we moved them to the home tank. The following 2 days, we moved one of the subjects to the apparatus and we administered 20 trials similar to those during the training phase. We then repeated the procedure for the second subject. In these trials, we presented 1 versus 2 dots as stimuli on the monitor. If the subject entered the chamber corresponding to the set with the larger number of dots, we immediately rewarded it with food. In the initial 16 trials, if the subject chose the set with the smaller numerosity, we rewarded it when it switched to the other chamber. In the last four trials of this series, if the subject entered the chamber corresponding to the smaller set, we closed the guillotine door immediately after the fish exited from the first chosen chamber. Thus, the subject could not correct initial wrong choices, a situation similar to the one of the training phase. The following day, the training began.
Training
During the training, we performed 12 daily trials, which were subdivided into two sessions of six trials each. The break between the two sessions was at least 2 h. In each session, we moved one subject from the home tank to the apparatus. The guillotine door was closed, and the monitor showed a black slide. Then, we presented the slide with the two sets of stimuli to be discriminated in the trial and raised the guillotine door to allow the subject to enter one of the choice chambers. If the subject chose the chamber associated with the larger numerosity, we administered a small quantity of food flakes as reward; if the subject chose the chamber with the smaller numerosity, we removed the stimuli and presented a black slide on the monitor. After the subject exited the choice chamber, we closed the guillotine door and started the following trial after 5 min. We varied the position (left or right) of the larger set of stimuli in each trial according to the pseudo-random scheme used by Bisazza et al. (2014).
We initially presented the subjects with up to 120 trials (for a total of 10 days of training) with a 2 versus 3 discrimination. Each day, we computed the number of correct choices in which the subject initially entered the chamber with the larger numerosity. Similarly to the previous study, we considered the subjects to achieve the discrimination if they reached the criterion of 75 % correct responses in two consecutive days. We presented successful subjects with a more difficult discrimination (3 versus 4). We also introduced a secondary, less stringent criterion: subjects that did not reach the primary learning criterion but reached a significant performance in the overall 10 days of training (Chi-square test) were admitted to the more difficult discrimination task as well. We used the proportion of correct responses in the analysis (arcsine square root transformed to meet parametric assumptions; Sokal and Rohlf 1995). Statistical tests were two-tailed; significance threshold was α = 0.05, unless stated otherwise. In the text, the M ± SD percentage of correct choices are given.
Results
Only one out of eight subjects achieved the primary criterion in the 2 versus 3 discrimination. This proportion of successful guppies was significantly smaller than the one observed in Bisazza et al. (2014) work, where seven out of eight subjects achieved the learning criterion in the 2 versus 3 discrimination (Fisher’s exact test: P = 0.010). Considering the secondary criterion, two additional subjects achieved the 2 versus 3 discrimination (Table 1, column 1). In the 3 versus 4 discrimination, none of these three subjects achieved the task through either the primary or the secondary criterion (62/120, 70/120, and 65/120 correct responses; all P > 0.06). An analysis with a Chi-square test revealed that one subject expressed a significant preference bias for the left choice chamber.
An overall analysis of the performance of the eight subjects failed to reveal a significant acquisition of the 2 versus 3 discrimination (53.29 ± 6.30 % correct responses; one-sample t test: t(7) = 1.478, P = 0.183). A linear mixed model (LMM) on the proportion of correct responses of each day of training (using the day as the fixed effect and the subject ID as a random factor) showed no significant improvement in the performance across the 10 days of the 2 versus 3 discrimination (F(9,57) = 1.295, P = 0.260).
Discussion
The guppies’ performances in the numerical discrimination task were strikingly lower in our experiment compared to the ones observed in the previous study (Bisazza et al. 2014). In our experiment, only one out of eight fish learned to discriminate up to 2 versus 3 objects; in the previous study, seven out of eight fish did so. None of the subjects in our experiment achieved the 3 versus 4 discrimination, while in the previous study, six out of eight fish were able to discriminate up to 3 versus 4 objects and five out of eight fish were even able to discriminate up to 4 versus 5 objects.
As in the two experiments we used subjects of the same sex, age, and strain, we maintained them in identical conditions before the test, we followed the same training schedule and the cognitive problem was identical, the different outcome is most likely due to the differences in the experimental setting. In the previous study, the setting reproduced an ecologically relevant context in which guppies had to remove small objects on the bottom of a tank to find food concealed below (Rodd et al. 2002), while we used a training setting more similar to that used in experiments with other vertebrates. It is therefore possible that the improved performance of guppies in the former study was due to a predisposition to learn within the specific setting. Before considering this hypothesis, we need to evaluate whether other differences in task parameters might account for the different outcome of the two studies. We attempted to keep the conditions (characteristics of the subjects, numerical contrasts, control of non-numerical variables, number of trials) as similar as possible in the two studies, but there are at least three important differences.
The first difference is the decision time allowed to the subject. In the previous work, subjects were kept behind a transparent guillotine door to allow them to look at both stimuli for 10 s before making their choice. With our procedure, as soon as the stimuli appeared on the monitor, the subject swam to one of the two chambers and could no longer see the alternative stimulus. Consequently, the decision time was normally very short, around 1–2 s. In many animals, there is a trade-off between decision time and accuracy (Chittka et al. 2009), and it has been shown that a shorter decision time reduces the performance in quantity discrimination in fish (Krause et al. 1998). Thus, the reduced decision time for the guppies in experiment 1 might be insufficient for comparing the stimuli and making an appropriate choice. Moreover, fish often show high impulsivity (Danisman et al. 2010; Lucon-Xiccato and Bisazza 2014; Miletto Petrazzini et al. 2015), which might cause guppies to choose the first stimulus they notice without comparing the two options.
The second important difference is the opportunity for fish to correct their first choice whenever it was wrong. Following the original procedure used with zebrafish (Parker et al. 2012a, b; Proulx et al. 2014; Wang et al. 2015), we did not allow correction, while Bisazza et al. (2014) used a correction procedure. A non-correction procedure is believed to lead to greater accuracy (Parducci and Polt 1958), but several studies have found that a correction method can facilitate learning, especially in the early phase (Hull and Spence 1938).
Finally, the third major difference concerns the type of the stimuli used. In previous work, the stimuli were three-dimensional (3D) coloured objects, while here we used 2D images displayed on an LCD monitor. Virtual stimuli have often been used in fish research (for reviews, see Fleishman and Endler 2000; Rosenthal 2000), and it has been shown that fish respond properly to both simple, static stimuli (Agrillo et al. 2014) and complex, moving stimuli such as conspecifics (Nicoletto and Kodric-Brown 1999). However, it is possible that 2D images are less salient than real objects leading guppies to experience difficulty with 2D abstract stimuli and show a reduced discrimination performance (Friedman et al. 2005; O’Hara et al. 2015; Truppa et al. 2015).
An additional potential problem is related to the number of training trials. Warm-blooded vertebrates can perform many training trials per day, and discrimination learning experiments with mammals and birds typically involve, overall, hundreds or even thousands of trials (Roberts and Mitchell 1994; Jaakkola et al. 2005; Cantlon and Brannon 2007). By contrast, many fish species, including guppies, can usually undergo a few reinforced trials per day (Agrillo et al. 2012), and they become easily stressed in prolonged trainings or show reduced motivation in participate to the task (Sovrano et al. 2003; Lucon-Xiccato and Bisazza 2016). For these reasons, discrimination learning experiments with fish normally have a preset limit in the number of training trials, typically one hundred or fewer (Bisazza et al. 2014; Lucon-Xiccato and Bisazza 2016). It is possible that with the setting used here, guppies require a longer training period to achieve the discrimination task. If two training settings lead to the same result but one requires a longer training, predetermining a small number of training trials can exaggerate the difference between the two methods.
In the following four experiments, we tested separately the influence of these four task parameters.
Experiment 2: Extension of decision time
Experiment 2 was similar to experiment 1, except that the fish were kept behind a transparent guillotine door and allowed to observe the stimuli for 20 s before they made their choice.
Materials and methods
Six naïve female guppies were used as subjects, which were different from the individuals used before. The only difference from the apparatus of experiment 1 was the presence of a second transparent guillotine door 25 cm away from the first door (Fig. 1). This second guillotine door formed a start box in the corridor in front of the choice chambers. At the beginning of each trial, the experimenter waited until the subject spontaneously swam into the start box and then closed the guillotine door. After 2 min, the stimuli were presented on the monitor. We released the fish from the start box after 20 s to allow it to observe the stimuli. We chose a decision time longer than the one adopted by Bisazza et al. (2014) because the distance between the guillotine door and the stimuli was greater in our apparatus (34 cm) compared to the previous one (15 cm). Other details of the procedure were identical to experiment 1.
Results
None of the six subjects achieved the primary or the secondary criterion in the 2 versus 3 discrimination (Table 1, column 2). This success rate was significantly lower than the one observed in the Bisazza et al. (2014) experiment (P = 0.005). One subject expressed a significant bias for the left choice chamber, and three subjects expressed a significant bias for the right choice chamber. An overall analysis of the performance of the six subjects failed to reveal a significant acquisition of the 2 versus 3 discrimination (49.98 ± 5.40 %; t(5) = 0.008, P = 0.994). There was no significant improvement in the performance across the 10 days of the 2 versus 3 discrimination (F(9,45) = 0.759, P = 0.654).
Discussion
In this experiment, we tested the possibility that the poor accuracy of guppies in experiment 1 was due to the reduced time allowed for observing the stimuli before making a decision. Indeed, in experiment 1, the fish observed the two stimuli for a few seconds before making the choice, and this may have contributed to the low performance observed (Chittka et al. 2009; Krause et al. 1998). Our attempt to extend the time before the decision yielded no evident improvement in learning compared to experiment 1. This result suggests that the greater accuracy shown in the previous study was not due to having forced the subjects to observe the task before the choice.
Experiment 3: Correction procedure
This experiment was identical to experiment 1 except that we adopted a correction procedure.
Materials and methods
We observed six different female guppies in this experiment. The apparatus and the procedure were the same as in experiment 1. However, if the subject chose the wrong chamber, we allowed it to switch to the correct chamber to receive the food reward. As in previous experiments, we use the first choice to measure the performance.
Results
None of the six subjects achieved the primary or secondary criterion in the 2 versus 3 discrimination (Table 1, column 3). This proportion of successful guppies was significantly lower (P = 0.005) than the one found by Bisazza et al. (2014). One subject expressed a significant bias for the left choice chamber. An overall analysis of the performance of the six subjects failed to reveal a significant acquisition of the 2 versus 3 discrimination ability (47.36 ± 5.93 %; t(5) = 1.091, P = 0.325). There was no overall significant improvement in the performance across the 10 days of training (F(9,45) = 1.299, P = 0.264).
Discussion
Both correction and non-correction procedures are currently employed in discrimination learning experiments. Because the former procedure is expected to favour fast learning (Hull and Spence 1938), we tested whether guppies’ performance in a numerical discrimination task could be improved by adopting a correction procedure. In experiment 3, we found no evidence of this improved performance. The fish trained using the correction procedure performed as poorly as the fish in experiment 1, which were trained without a correction procedure. Even in this case, we found no evidence that discrimination significantly improved over time, at least within the 120 trials administered. This experiment suggests, therefore, that the correction procedure is not responsible for the differences between this study and the previous one in which the guppies performed better (Bisazza et al. 2014).
Experiment 4: 3D stimuli
In this experiment, we used two sets of 3D objects as stimuli.
Materials and methods
As in the previous experiments, we tested six new female guppies which were experimentally naïve. The stimuli were sets of yellow plastic discs, the same as in Bisazza et al. (2014) experiment. Using a needle, we fixed the discs to two green plastic panels (Fig. 2b). During the trials, the experimenter inserted these panels in the two choice chambers to present the stimuli. Using discs with five different diameters (15, 16, 17, 18, and 19 mm) and varying their position, we corrected the sets separately for cumulative surface area, overall space encompassed, and density as described for experiment 1. We added an additional fluorescent lamp above the choice chambers to light the stimuli (Fig. 1). The guillotine door was covered with opaque plastic to prevent the fish from seeing the stimuli before the experimenter completed the insertion. The monitor was not used in this experiment. Other details of apparatus and procedures were identical to experiment 1.
Results
One out of six subjects achieved the criterion in the 2 versus 3 discrimination. This success rate was, again, significantly lower (P = 0.026) than the one observed by Bisazza et al. (2014). Considering the secondary criterion, an additional subject achieved the 2 versus 3 discrimination (Table 1, column 4). In the following 3 versus 4 discrimination, these two subjects did not reach the primary or the secondary learning criterion (58/120 and 60/120 correct responses; all P > 0.7). One subject expressed a significant bias for the left choice chamber, and three subjects expressed a significant bias for the right choice chamber. An overall analysis of the performance of the six subjects revealed a significant acquisition of the 2 versus 3 discrimination (58.02 ± 4.90 %; t(5) = 3.964, P = 0.011). Even the subjects that did not reach the individual criterion tend to perform above chance level. Indeed, the individual accuracy of these four subjects ranged from 53.33 to 56.67 %, with an mean that was significantly above chance (55.00 ± 1.36 %; t(3) = 7.325, P = 0.005).
Because this was the only experiment in this study in which the subjects, overall, significantly learned the task, we compared the performance of experiment 4 with the pooled data of experiments 1, 2, and 3 in which the stimuli were presented on a monitor (Levene test for homogeneity of variances between the two groups: F(1,24) = 0.910, P = 0.350; Boneau 1960; de Winter 2013). We found that guppies performed significantly better in experiment 4 than in the other three experiments of this work (independent-samples t test: t(24) = 2.727, P = 0.012).
Discussion
In experiment 4, we used two sets of 3D objects as stimuli to test the possibility that the reduced performance shown by guppies in experiment 1 was due to difficulty with the 2D abstract stimuli, which in some cases may negatively affect discrimination performance compared to the use of 3D stimuli (Friedman et al. 2005; O’Hara et al. 2015; Truppa et al. 2015). As we expected, the use of real 3D objects improved the numerical discrimination performance of the guppies. Although this improvement was limited and only one subject reached the primary learning criterion, it seems clear that the performance of most fish tended to be above chance. Similar effects have been reported also in other species. For example, Truppa et al. (2015) trained capuchin monkeys (Sapajus spp.) to select either the larger between two objects or the larger between two images presented with a computer monitor, finding a better discrimination performance with the former stimuli. The format of the stimuli (2D versus 3D) appears to be an important source of variation for animal discrimination performance. This could be explained by the fact that 3D stimuli resemble the stimuli that animals normally experience and are therefore more salient (Mustafar et al. 2015). The use of 2D images is often preferred in discrimination learning experiments because it offers a better control of stimulus features. However, our results, together with those of previous works, suggest that this choice might bear the cost of reduced discrimination performance.
Despite the increased accuracy due to the 3D stimuli, the overall performance of our guppies remained very low compared with the previous study (Bisazza et al. 2014). Only two fish learned to discriminate between 2 versus 3 objects, and none of these learned to discriminate between 3 versus 4 objects. Therefore, even if the format of the stimuli appears to be a parameter that affects the outcome of the experiment, it can only partially explain the difference between the performances observed in our experiment 1 and in the previous experiment.
Experiment 5: Extended training
In this experiment, we tested whether guppies could learn the numerical discrimination if we extended the training from 10 days to 30 (360 trials overall).
Materials and methods
We used three subjects from experiment 2 and three subjects from experiment 3. None of these subjects reached the learning criterion within the predetermined 120 trials. The prolonged training was performed using the modality and the apparatus described for experiments 2 and 3, respectively.
Results
Only one of the six subjects (originally from experiment 3) reached the secondary learning criterion after extending the training from 120 trials to 360. An overall analysis of the performance of the six subjects did not reveal a significant acquisition of the 2 versus 3 discrimination ability (50.83 ± 3.54 %; t(5) = 0.577, P = 0.589; Table 1, column 5), nor was there a significant improvement in the performance over time (LMM: F(1,4) = 0.112, P = 0.754).
Discussion
Increasing the number of training trials from 120 to 360 did not substantially improve the performance of the guppies. Only one fish succeeded in reaching the learning criterion after extending the training to three times the original length. The overall performance of the six fish remained non-significant. Therefore, the length of the training does not seem to be the key parameter in explaining why guppies obtained a much better record in the numerical task used in the previous study (Bisazza et al. 2014) compared with the procedure used in this study.
General comparison between the experiments
We performed a tentative analysis to compare our experiments with the previous one (Bisazza et al. 2014). Overall, the performance of the 26 guppies of our experiments pooled together was significantly worse than the one of the guppies in the previous work (2 versus 3 discrimination; independent-samples t test: t(32) = 16.316, P < 0.001). We also ran a one-way ANOVA on the pooled data of the proportion of correct responses in the 2 versus 3 discrimination with experiment as a five-level factor (previous experiment, our experiment 1, 2, 3, and 4). We found a significant effect of the factor experiment (F(4,29) = 13.977, P < 0.001). A Tukey’s post hoc test revealed that the performance was significantly lower in our four experiments compared to the previous experiment (experiment 1: P < 0.001; experiment 2: P < 0.001; experiment 3: P < 0.001; experiment 4: P = 0.018; Fig. 3). We performed a further comparison using the pooled data of our experiments without the ten subjects that showed a preference bias for one of the two chambers and we found a similar result. The performance in our experiments (52.00 ± 6.51 %) was significantly lower compared with that of the previous experiment (t(22) = 5.803, P < 0.001).

Comparison of the five experiments of the present study and the results of Bisazza et al. (2014)’s study. Bars represent M ± SD percentage of correct choices in the 2 versus 3 discrimination. Horizontal line represents chance performance
Conclusions
In this study, we provided evidence that the numerical acuity of guppies is radically different according to the training setting adopted. With the setting of the previous study by Bisazza et al. (2014), five out of eight subjects achieved a 4 versus 5 discrimination, while with the more conventional two-choice discrimination setting adopted in the present work, most of the guppies failed even an ‘easy’ 2 versus 3 discrimination. To understand whether the task parameters could account for the difference, we varied the extension of decision time, whether or not correction was allowed, and the length of the training period. None of these variants enhanced guppies’ performance. By contrast, the use of 3D stimuli slightly improved guppies’ accuracy in numerical discrimination, but this improvement was still not enough to approximate the performance observed in the previous study. A possible limitation of our study is that we used a small number of subjects in each experiment (6–8 fish). Although our sample size was in line with the ones typically used in discrimination learning experiments (Schluessel et al. 2014; Prétôt et al. 2016), it is possible that this factor limited the likelihood to evidence subtle differences among the different experiments of this study. It is, however, unlikely that the sample size affected the general conclusion that the methods used in the two studies yield quite different results, as shown also by the overall analyses in which we pooled all our subjects (N = 26) to increase statistical power.
We detected another difference between the two studies. With the setting adopted in this work, some guppies (10 out of 26) exhibited a side bias, with four choosing the left chamber and six choosing the right chamber significantly more often. With the setting previously used by Bisazza et al. (2014), none of the subjects showed side bias. There are at least two possible explanations for the side bias observed in the present study. The first explanation is that the choice between the right and the left chambers was affected by the presence of spontaneous side preferences due to cerebral lateralization. Cerebral lateralization and the resulting motor and sensory asymmetries have been demonstrated in virtually all vertebrates (reviewed in Rogers and Andrew 2002), including guppies and many other fish (reviewed in Bisazza and Brown 2011). In most of the discrimination learning studies on fish showing lateralization, motorial asymmetries have not been reported to prevent learning (e.g. cichlids: Schluessel et al. 2014; guppies: Lucon-Xiccato and Bisazza 2016; zebrafish: Colwill et al. 2005). However, in few cases, there is evidence that these motorial asymmetries may hamper learning in discrimination tasks. For example, rats showing spontaneous side bias in a T-maze showed poor learning, or even the absence of learning, when trained to choose the contralateral arm of the maze (Andrade et al. 2001). Concerning fish, Brown and Braithwaite (2005) found lower spatial learning performance in a lateralized population of the poeciliid Brachyrhaphis episcopi and suggested that spontaneous turning preferences interfered with the choice of the correct option. The second possible explanation is that the side bias was acquired during the training. Animals often develop side biases in discrimination tasks when the two stimuli are presented side-by-side in two fixed locations (rodents: Prusky et al. 2000; primates: Genty et al. 2004; birds: Smith and Zentall 2016). A tendency to rely on the spatial position of the stimuli rather than on other cues may be present in guppies too (Miletto Petrazzini et al. 2015) and might have hampered learning.
The absence of side bias in the study of Bisazza et al. (2014) might be due to the fact that their setting minimized any precise left–right localization of the two stimuli by presenting them on a large plate on the bottom of the tank and changing their position every trial. Although side bias might be an important factor in our setting, it is unlikely that it alone explains the difference between the two studies. Indeed, excluding the guppies with side bias did not increase the level of the performance in our experiments, which remained, on average, at a correct choice rate of 52 %. In addition, a previous study employing a training procedure that prevented any left–right localization of the stimuli (Agrillo et al. 2014) did not obtain a numerical performance comparable to that of Bisazza et al. (2014).
In sum, none of the five factors examined above appear to explain the huge difference in the performance between our study and the previous one. Such difference is likely to arise from Bisazza and colleagues’ use of a setting that mimics guppies’ natural foraging habitat. An interesting parallel to our results comes from works on pigeons, which often forage by displacing leaves or gravel on the ground. Pigeons trained on concept learning using gravel of different colours hiding food—a setting that resembles their natural foraging habit—reached learning performances that were unattainable with conventional training in the Skinner box (Wright and Delius 1994). Remarkably, in pigeons, the position of the stimuli (vertical versus horizontal presentation) was not the key factor explaining the different performance with the two methods (Wright et al. 1988) and it would be interesting to investigate whether the same occurs in guppies.
A possible explanation of pigeons and guppy data calls into play innate predispositions to learn. Natural selection can favour the evolution of learning predispositions in one species to solve specific ecological problems (Shettleworth 1972). As a consequence, the species shows enhanced cognitive performances in that specific context compared to other situations, and it may often outperform other species that lack the same ecological specialization. For example, food-storing birds can memorize the location of thousands of food items, but in other circumstances their mnemonic performance is similar to that of other birds (Shettleworth and Hampton 1998). Pigeons recognize rotated objects better than humans do, which might be due to the fact that objects’ orientations change continuously during flight (Hollard and Delius 1982). Based on observational studies (Dussault and Kramer 1981; Rodd et al. 2002), we suggest that guppies might be predisposed to choose patches with a large number of objects that could potentially hide food, since the likelihood of finding food is increased in such patches.
A similar but distinct explanation for the difference between the two studies is that the naturalistic setting used by Bisazza et al. (2014) measured the real cognitive repertoire of the species, while the artificial conditions of the present learning setting prevented somehow guppies from fully displaying their cognitive abilities. The experimental setting of this study resembled the classical setting of the Skinner box adopted in rodents, primates, and birds (Skinner 1933; Douglas and Whitty 1941; Wright et al. 1988), and that is gaining popularity in fish studies (Gierszewski et al. 2013; Manabe et al. 2013; Proulx et al. 2014). The fish is placed in a bare environment, and artificial bi-dimensional stimuli are displayed on a computer screen near the tank. To obtain the food reward, the fish is required to approach the side of the tank where the correct stimulus is presented. Since the position of the correct stimulus switches in the different trials, the fish is also required to inhibit the natural predisposition to associate food to the place where it has been previously found. All these features might favour comparison with works on other species, but might hamper learning. Both in nature and in the semi-natural conditions of the maintenance aquaria, guppies have hardly the opportunity to experience such conditions. Conversely, guppies spend considerable time moving objects on the bottom to search for hidden food, a situation similar to the setting that favoured learning in guppies (Bisazza et al. 2014). Both explanations—the naturalistic setting favours learning and the artificial setting hampers learning—appear plausible, and our work cannot unfortunately provide direct support to either of them. Testing these ideas will require careful experimental investigation both in the laboratory and in the field.
The main message of this work is that the choice of the training setting sometimes affects the results of the experiment and, consequently, the estimation of the numerical discrimination abilities of the species. Although it was already known that different tests may lead to different results, such as in the case of spontaneous choice versus training tests (Agrillo and Bisazza 2014), our work is one of the very few studies showing that within the same type of test (training), different settings may also lead to quite different results. For fish, a similar conclusion has been recently drawn by a study of guppies’ spontaneous preference for larger social groups (Lucon-Xiccato et al. unpublished result). In that study, guppies showed a higher accuracy in numerical discrimination when the experimental setting and the procedure were modified to remedy some flaws of the classical method, such as favouring the switching of the subject between the two shoals to compare them, and using stimulus fish that were well acclimated to the apparatus and thus behaved more naturally.
The problem of the experimental setting may be very relevant for studies that compare the performance of different species in the same task in order to understand the evolution of cognitive abilities. For example, one study found lower performance in numerical discrimination in zebrafish compared to that of four other fish species; however, a control experiment revealed that the zebrafish was simply less efficient in learning with the specific setting adopted in that study (Agrillo et al. 2012). In another study, primates were thought to be unable to perform a task that cleaner fish (Labroides dimidiatus) perform quite well (Salwiczek et al. 2012). The setting adopted in that study matched the fish’s ecology, but not that of the primates. The adoption of an experimental setting more appropriate for primates showed that they, too, were able to solve the task (Prétôt et al. 2016). To obtain an estimation of cognitive abilities that can be used in comparative research, it is therefore paramount to use a setting adequate to each species studied and, possibly, to perform multiple comparisons for the same cognitive ability .
References
Agrillo C, Bisazza A (2014) Spontaneous versus trained numerical abilities. A comparison between the two main tools to study numerical competence in non-human animals. J Neurosci Methods 234:82–91
Agrillo C, Miletto Petrazzini ME, Tagliapietra C, Bisazza A (2012) Inter-specific differences in numerical abilities among teleost fish. Front Psychol 3:483
Agrillo C, Miletto Petrazzini ME, Bisazza A (2014) Numerical acuity of fish is improved in the presence of moving targets, but only in the subitizing range. Anim Cogn 17:307–316
Al Aïn S, Giret N, Grand M, Kreutzer M, Bovet D (2009) The discrimination of discrete and continuous amounts in African grey parrots (Psittacus erithacus). Anim Cogn 12:145–154
Andrade C, Alwarshetty M, Sudha S, Chandra JS (2001) Effect of innate direction bias on T-maze learning in rats: implications for research. J Neurosci Methods 110:31–35
Bánszegi O, Urrutia A, Szenczi P, Hudson R (2016) More or less: spontaneous quantity discrimination in the domestic cat. Anim Cogn. doi:10.1007/s10071-016-0985-2
Benson-Amram S, Heinen VK, Dryer SL, Holekamp KE (2011) Numerical assessment and individual call discrimination by wild spotted hyaenas, Crocuta crocuta. Anim Behav 82:743–752
Beran MJ (2008) Monkeys (Macaca mulatta and Cebus apella) track, enumerate, and compare multiple sets of moving items. J Exp Psych Anim Behav Proc 34:63–74
Bisazza A, Brown C (2011) Lateralization of cognitive functions in fish. In: Brown C, Krause J, Laland KN (eds) Fish cognition and behaviour. Wiley, Oxford, pp 298–324
Bisazza A, Agrillo C, Lucon-Xiccato T (2014) Extensive training extends numerical abilities of guppies. Anim Cogn 17:1413–1419
Boneau CA (1960) The effects of violations of assumptions underlying the t test. Psychol Bull 57:49–64
Brown C, Braithwaite VA (2005) Effects of predation pressure on the cognitive ability of the poeciliid Brachyraphis episcopi. Beahv Ecol 16:482–487
Cantlon JF, Brannon EM (2007) How much does number matter to a monkey (Macaca mulatta)? J Exp Psych Anim Behav Proc 33:32–41
Chittka L, Skorupski P, Raine NE (2009) Speed–accuracy tradeoffs in animal decision making. Trends Ecol Evol 24:400–407
Colwill RM, Raymond MP, Ferreira L, Escudero H (2005) Visual discrimination learning in zebrafish (Danio rerio). Behav Process 70:19–31
Danisman E, Bshary R, Bergmüller R (2010) Do cleaner fish learn to feed against their preference in a reverse reward contingency task? Anim Cogn 13:41–49
de Winter JCF (2013) Using the Student’s t test with extremely small sample sizes. Pract Assess Res Eval 18:1–12
Deaner RO, Isler K, Burkart J, Van Schaik C (2007) Overall brain size, and not encephalization quotient, best predicts cognitive ability across non-human primates. Brain Behav Evol 70:115–124
Douglas JWB, Whitty CWM (1941) An investigation of number appreciation in some sub-human primates. J Comp Psychol 31:129–143
Dussault GV, Kramer DL (1981) Food and feeding behavior of the guppy, Poecilia reticulata (Pisces: Poeciliidae). Can J Zool 59:684–701
Emmerton J, Renner JC (2006) Scalar effects in the visual discrimination of numerosity by pigeons. Learn Behav 34:176–192
Feigenson L, Dehaene S, Spelke ES (2004) Core systems of number. Trends Cogn Sci 8:307–314
Fleishman LJ, Endler JA (2000) Some comments on visual perception and the use of video playback in animal behavior studies. Acta Ethol 3:15–27
Friedman A, Spetch ML, Ferrey A (2005) Recognition by humans and pigeons of novel views of 3-D objects and their photographs. J Exp Psychol Gen 134:149–162
Genty E, Palmier C, Roeder JJ (2004) Learning to suppress responses to the larger of two rewards in two species of lemurs, Eulemur fulvus and E. macaco. Anim Behav 67:925–932
Gierszewski S, Bleckmann H, Schluessel V (2013) Cognitive abilities in Malawi cichlids (Pseudotropheus sp.): matching-to-sample and image/mirror-image discriminations. PLoS One 8:e57363
Hager MC, Helfman GS (1991) Safety in numbers: shoal size choice by minnows under predatory threat. Behav Ecol Sociobiol 29:271–276
Hanus D, Call J (2007) Discrete quantity judgments in the great apes (Pan paniscus, Pan troglodytes, Gorilla gorilla, Pongo pygmaeus): the effect of presenting whole sets versus item-by-item. J Comp Psychol 121:241–249
Hinde RA (1973) Constraints on learning: an introduction to the problems. In: Hinde RA, Stevenson-Hinde J (eds) Constraints on learning: limitations and predispositions. Academic Press, New York
Hollard VD, Delius JD (1982) Rotational invariance in visual pattern recognition by pigeons and humans. Science 218:804–806
Hull CL, Spence KW (1938) “ Correction” versus” non-correction” method of trial-and-error learning in rats. J Comp Psychol 25:127–145
Jaakkola K, Fellner W, Erb L, Rodriguez M, Guarino E (2005) Understanding of the concept of numerically” less” by bottlenose dolphins (Tursiops truncatus). J Comp Psychol 119:296–303
Krause J, Godin JGJ, Rubenstein D (1998) Group choice as a function of group size differences and assessment time in fish: the influence of species vulnerability to predation. Ethology 104:68–74
Lefebvre L, Reader SM, Sol D (2004) Brains, innovations and evolution in birds and primates. Brain Behav Evolut 63:233–246
Lucon-Xiccato T, Bisazza A (2014) Discrimination reversal learning reveals greater female behavioural flexibility in guppies. Biol Lett 10:20140206
Lucon-Xiccato T, Bisazza A (2016) Male and female guppies differ in speed but not in accuracy in visual discrimination learning. Anim Cogn 19:733–744
Lucon-Xiccato T, Miletto Petrazzini ME, Agrillo C, Bisazza A (2015) Guppies discriminate between two quantities of food items but prioritize item size over total amount. Anim Behav 107:183–191
Lucon-Xiccato T, Dadda M, Bisazza A (2016) Sex differences in discrimination of shoal size in the guppy (Poecilia reticulata). Ethology 122:481–491
Manabe K, Dooling RJ, Takaku S (2013) An automated device for appetitive conditioning in zebrafish (Danio rerio). Zebrafish 10:518–523
Miletto Petrazzini ME, Wynne CD (2016) What counts for dogs (Canis lupus familiaris) in a quantity discrimination task? Behav Proc 122:90–97
Miletto Petrazzini ME, Lucon-Xiccato T, Agrillo C, Bisazza A (2015) Use of ordinal information by fish. Sci Rep 5:15497
Mustafar F, De Luna P, Rainer G (2015) Enhanced visual exploration for real objects compared to pictures during free viewing in the macaque monkey. Behav Process 118:8–20
Nicoletto PF, Kodric-Brown A (1999) The use of digitally-modified videos to study the function of ornamentation and courtship in the guppy, Poecilia reticulata. Environ Biol Fish 56:333–342
Normand E, Ban SD, Boesch C (2009) Forest chimpanzees (Pan troglodytes verus) remember the location of numerous fruit trees. Anim Cogn 12:797–807
O’Hara M, Huber L, Gajdon GK (2015) The advantage of objects over images in discrimination and reversal learning by kea, Nestor notabilis. Anim Behav 101:51–60
Olson DJ, Kamil AC, Balda RP, Nims PJ (1995) Performance of four-seed caching corvid species in operant tests of nonspatial and spatial memory. J Comp Psychol 109:173–181
Parducci A, Polt J (1958) Correction versus noncorrection with changing reinforcement schedules. J Comp Physiol Psychol 51:492–495
Parker MO, Gaviria J, Haigh A, Millington ME, Brown VJ, Combe FJ, Brennan CH (2012a) Discrimination reversal and attentional sets in zebrafish (Danio rerio). Behav Brain Res 232:264–268
Parker MO, Millington ME, Combe FJ, Brennan CH (2012b) Development and implementation of a three-choice serial reaction time task for zebrafish (Danio rerio). Behav Brain Res 227:73–80
Pepperberg IM (2006) Grey parrot numerical competence: a review. Anim Cogn 9:377–391
Prétôt L, Bshary R, Brosnan SF (2016) Comparing species decisions in a dichotomous choice task: adjusting task parameters improves performance in monkeys. Anim Cogn 19:819–834
Proulx MJ, Parker MO, Tahir Y, Brennan CH (2014) Parallel mechanisms for visual search in zebrafish. PLoS One 9:e111540
Prusky GT, West PW, Douglas RM (2000) Behavioral assessment of visual acuity in mice and rats. Vision Res 40:2201–2209
Reebs SG (1999) Time–place learning based on food but not on predation risk in a fish, the inanga (Galaxias maculatus). Ethology 105:361–371
Roberts WA, Mitchell S (1994) Can a pigeon simultaneously process temporal and numerical information? J Exp Psych Anim Behav 20:66–78
Rodd FH, Hughes KA, Grether GF, Baril CT (2002) A possible non-sexual origin of mate preference: are male guppies mimicking fruit? Proc R Soc Lond B Bio Sci 269:475–481
Rogers LJ, Andrew R (2002) Comparative vertebrate lateralization. Cambridge University Press, Cambridge
Rosenthal GG (2000) Design considerations and techniques for constructing video stimuli. Acta Ethol 3:49–54
Salwiczek LH, Prétôt L, Demarta L, Proctor D, Essler J, Pinto AI, Wismer S, Stoinski T, Brosnan SF, Bshary R (2012) Adult cleaner wrasse outperform capuchin monkeys, chimpanzees and orang-utans in a complex foraging task derived from cleaner–client reef fish cooperation. PLoS One 7:e49068
Schluessel V, Kraniotakes H, Bleckmann H (2014) Visual discrimination of rotated 3D objects in Malawi Cichlids (Pseudotropheus sp.): a first indication for form constancy in fishes. Anim Cogn 17:359–371
Sherry DF (2006) Neuroecology. Ann Rev Psychol 57:167–197
Shettleworth SJ (1972) Constraints on learning. In: Lehrman DS, Hinde RA, Shaw E (eds) Advances in the study of behaviour: IV. England, Oxford, pp 1–68
Shettleworth SJ, Hampton RR (1998) Adaptive specializations of spatial cognition in food storing birds? In: Balda RP, Pepperberg IM, Kamil AC (eds) Approaches to testing a comparative hypothesis. Animal cognition in nature, San Diego, pp 65–98
Skinner BF (1933) The rate of establishment of a discrimination. J Gen Psychol 9:302–350
Smith AP, Zentall TR (2016) Suboptimal choice in pigeons: choice is primarily based on the value of the conditioned reinforcer rather than overall reinforcement rate. J Exp Psychol Anim Learn Cogn 42:212
Sokal RR, Rohlf FJ (1995) Biometry: the principals and practice of statistics in biological research. WH Freeman and Company, New York
Sovrano VA, Bisazza A, Vallortigara G (2003) Modularity as a fish (Xenotoca eiseni) views it: conjoining geometric and nongeometric information for spatial reorientation. J Exp Psychol Anim Behav Proc 29:199–210
Truppa V, Carducci P, Trapanese C, Hanus D (2015) Does presentation format influence visual size discrimination in tufted capuchin monkeys (Sapajus spp.)? PLoS One 10:e0126001
Wang MY, Brennan CH, Lachlan RF, Chittka L (2015) Speed–accuracy trade-offs and individually consistent decision making by individuals and dyads of zebrafish in a colour discrimination task. Anim Behav 103:277–283
Wright AA, Delius JD (1994) Scratch and match: pigeons learn matching and oddity with gravel stimuli. J Exp Psychol Anim Behav Processes 20:108–112
Wright AA, Cook RG, Rivera JJ, Sands SF, Delius JD (1988) Concept learning by pigeons: matching-to-sample with trial-unique video picture stimuli. Anim Learn Behav 16:436–444
Acknowledgments
Data from Bisazza et al. (2014) have been used with the permission of the authors. We would like to thank Matteo Rigato, Jessica Montanari, Camilla Giacobbi, Elisa Monastero, and Giulia Goldin for their help in testing the animals.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interests.
Ethical standards
The experiments comply with the law of the country (Italy) in which they were performed (Decreto legislativo 4 marzo 2014, n. 26).
Rights and permissions
About this article

Cite this article
Gatto, E., Lucon-Xiccato, T., Savaşçı, B.B. et al. Experimental setting affects the performance of guppies in a numerical discrimination task. Anim Cogn 20, 187–198 (2017). https://doi.org/10.1007/s10071-016-1037-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10071-016-1037-7