
Abstract
This paper reports the effect of high hydrostatic pressure (HHP) on wines contaminated with Dekkera/Brettanomyces populations of 104 and 106 cfu mL−1 growing at either pH 3.2 or 3.6 (both normal pHs for red wine) and at room temperature (25 °C). HHP (100 MPa for 24 h) was highly effective at controlling the growth of all combinations of starting yeast population and pH, yet caused scant modification of thermosensitive wine molecules such as pigments and volatile compounds (molecules that greatly influence wine quality). The present results support the potential use of HHP as a means of cold-pasteurising wines to control Dekkera/Brettanomyces.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The yeast genera Dekkera and Brettanomyces (in fact sporulating and non-sporulating forms of the same organism) cause problems for winemakers owing to their ability to transform grape hydroxycinnamic acids (HCAs) into vinylphenols, and then the latter into ethylphenols. The metabolic pathway leading to the formation of ethylphenols involves the enzymes hydroxycinnamate decarboxylase [10] and vinylphenol reductase [11, 29]. Dekkera members catabolise ferulic, caffeic and p-coumaric acids and possess inducible enzymes with similar pH and temperature optima [12]. Dekkera/Brettanomyces can transform some 90 % of HCA into ethylphenols [8] under oenological conditions, especially those encountered during ageing. For a review of the main physiological, morphological and metabolic characteristics of Dekkera/Brettanomyces, see [28].
Ethylphenols are molecules with low sensorial thresholds; for example, 4-ethylphenol has a sensorial threshold of just 230 μg L−1 [4]. Unfortunately, they have a negative impact on wine quality. The perception of a ‘Brett character’ (i.e. ‘animal’, ‘old leather’ or ‘stable’ aromas that reduce the value of aged wines), however, depends on the type of grape used. Higher sensorial thresholds have been reported in monovarietal Cabernet Sauvignon wines than Tempranillo wines [24]. Other authors report wine 4-ethylphenol concentrations of 3.0, 1.74 and 0.68 mg L−1 to be associated with strong, medium and absent Brett character, respectively [17]. Although wineries do not like to admit Dekkera/Brettanomyces problems, the presence of these yeasts (or their ethylphenol product) has been detected in the cellars of many winemaking regions [2, 23, 26].
Dekkera/Brettanomyces can be controlled by flash pasteurisation and membrane filtration [3], but these techniques also have negative impacts on the sensorial quality of wines. The addition of SO2 can also control these yeasts [9], but it is difficult to guarantees a stable molecular SO2 concentration in wines ageing in wooden barrels. Other additives, such as dimethyl dicarbonate [6], are also effective, but deciding on an adequate dose can be complicated.
The food industry is currently tending towards the use of non-thermal food preservation techniques. One of these, subjection to high hydrostatic pressure (HHP), can be regarded as a form of cold pasteurisation. Pressures of 100–700 MPa are commonly used, which damage the cell walls and membranes of spoilage organisms [20]. High hydrostatic pressure has the advantage that it produces fewer sensorial modifications to foods than do physical techniques such as conventional pasteurisation or filtration. Neither does it have any of the disadvantages of adding chemical agents. Subjecting wines to high pressures has been proposed as a means of microbiologically and biochemically stabilising wines [7, 25] with minimum sensorial repercussions [19].
The aim of the present work was to test the use of HHP as a means of controlling Dekkera/Brettanomyces in red wines and to determine the effects of such treatment on wine colour and aromatic profile.
Materials and methods
Yeast strain used and the preparation of contaminated wines
The wine used was DO Mancha, which is made from Tempranillo, Merlot and Syrah grapes (V. vinifera L.) (alcohol content, 13 % v/v; pH, 3.6; total SO2, 35 mg L−1; free SO2, 11 mg L−1). To provide a wine sample with a second pH, a fraction of the original was acidified with tartaric acid (Panreac, Barcelona, Spain) to pH 3.2.
The spoilage yeast used was Dekkera bruxellensis D37 (a member of the Instituto de Fermentaciones Industriales collection, CSIC, Madrid, Spain), regarded here as a model member of Dekkera/Brettanomyces.
Wines with D. bruxellensis D37 populations of either 106 or 104 cfu mL−1 were prepared by adding, to 50 mL of wine, either 312 or 3 μL of a yeast suspension (population 1.6 × 108 cfu mL−1) prepared in YEPD medium [15]. Four types of contaminated wines were finally produced, that is, with populations of either 106 or 104 cfu mL−1 and at pH 3.2 or 3.6. Non-contaminated wine samples were also prepared at both pHs.
High hydrostatic pressure
The contaminated wines were pressurised to eliminate D. bruxellensis D37 cells using a TFS-2L HHP apparatus (Mayekawa, Tokyo, Japan) (treatment vessel volume 2 L; pressurisation range 0–100 MPa). This was performed at the maximum pressure produced by this apparatus (100 MPa) for 24 h at 25 °C in Falcon tubes in a vacuum-sealed plastic bag. All four contaminated wines (i.e. both population sizes at both pHs) were subjected to HHP. Control wines with populations of 106 cfu mL−1 at pH 3.2 and 3.6, not subjected to HHP, were used to confirm normal yeast development. All assays performed in triplicate.
Non-contaminated wine at pH 3.6 was also subjected to HHP at 100 MPa to determine whether this treatment had any effect on wine sensorial features.
Selective/differential media for D. bruxellensis D37 detection
The growth of D. bruxellensis D37 in the wines was confirmed using DBDM selective/differential medium [27].
Determination of anthocyanins
The following anthocyanins and pyranoanthocyanins were analysed using an Agilent Technologies (Palo Alto, CA) HPLC chromatograph series 1100, equipped with a diode array detector and a quadrupole mass spectrometer with an electrospray interface: delphinidin-3-O-glucoside (D3G), petunidin-3-O-glucoside (Pt3G), peonidin-3-O-glucoside (Pn3G), malvidin-3-O-glucoside (M3G), malvidin-3-O-glucoside-pyruvate adduct (vitisin A; VITA), malvidin-3-O-(6″-p- acetylglucoside)-pyruvate adduct (AcVITA), malvidin-3-O-(6″-p-coumaroylglucoside)-pyruvate adduct (CmVITA), petunidin-3-O-(6″-acetylglucoside) (Pt3G6Ac), peonidin-3-O-(6″-acetylglucoside) (Pn3G6Ac), malvidin-3-O-(6″-acetylglucoside) (M3G6Ac), peonidin -3-O-(6″-p-coumaroylglucoside) (Pn3GCm), malvidin-3-O-(6″-p-coumaroylglucoside) (M3GCm), malvidin-3-O-glucoside-4-vinylcatechol (M3GVCath), malvidin-3-O-glucoside-4-vinylepicatequine (M3GVEpiCat), malvidin-3-O-glucoside-4-vinylphenol (M3GVPh), malvidin-3-O-glucoside-4-vinylguaiacol (M3GVG) and malvidin-3-O-(6″-p-coumaroylglucoside)-vinylphenol (M3GCmVPh). Gradients of solvent A (water/formic acid, 90:10, v/v) and B (methanol/formic acid, 90:10, v/v) were used in a reverse-phase Kinetex C18 column (Phenomenex, Torrance, CA, USA) (100 × 4.6 mm; particle size 2.6 μm) as follows: 10 % B (0.8 ml min−1) from 0 to 3 min, 10–50 % B linear (0.8 ml min−1) from 3 to 18 min, 50 % B from 18 to 20 min, 50–10 % B linear (0.8 ml min−1) from 20 to 21 min, and re-equilibration of the column from 21 to 23 min. Detection was performed by scanning in the 500–600 nm range. Quantification was performed by comparison against an external standard at 525 nm and expressed as a function of the concentration of malvidin-3-O-glucoside (Extrasynthèse, Genay, France). The different anthocyanins were identified by their retention times with respect to the majority anthocyanin malvidin-3-O-glucoside, and by comparing the UV–visible and mass spectra with data in the literature [13, 22]. The ESI parameters were drying gas (N2) flow, 10 ml min−1; temperature, 350 °C; nebuliser pressure, 380 Pa (55 psi); and capillary voltage, 4,000 V. Mass spectrometry was performed in positive mode scanning, from 100 to m/z 1,000, using as a fragmenter voltage of 150 V from 0 to 23 min. One-hundred-microlitre samples of previously filtered (0.45-μm membrane filters made of cellulose methylic esters) (Tecknokroma, Barcelona, Spain) wines were injected into the HPLC apparatus. The detection limit was 0.1 mg L−1.
Analysis of volatile compounds by gas chromatography with flame ionisation detection
Volatile compounds were determined using an Agilent Technologies 6850 gas chromatograph (Network GC System) equipped with an integrated flame ionisation detector, as described by [1]. A DB-624 column (60 m × 250 μm × 1.40 μm) was employed, calibrated with the following compounds as external standards: acetaldehyde, methanol, 1-propanol, 1-butanol, 2-butanol, isobutanol, 2-methyl-1-butanol, 3-methyl-1-butanol, 2-phenylethyl acetate, 2-phenylethyl alcohol, diacetyl, ethyl acetate, isoamyl acetate, isobutyl acetate, ethyl butyrate, ethyl lactate and hexanol. 4-Methyl-2-pentanol was used as an internal standard (all compounds from Fluka, Sigma–Aldrich Corp., Buchs SG, Switzerland). The injector temperature was 250 °C, and the detector temperature was 300 °C. The column temperature was 40 °C for the first 5 min, rising linearly by 10 °C min−1 until reaching 250 °C; this temperature was then maintained for 5 min. Hydrogen was used as the carrier gas; this was provided by a hydrogen generator (LNI Schmidlin SA, Geneva, Switzerland). The flow rate was 22.1 L min−1, the injection split ratio was 1:10, and the detection limit was 0.1 mg L−1. One hundred microlitres of internal standard (concentration 500 mg L−1) were added to 1mL test samples and filtered through syringe membrane filters (0.45 μm pore size) (Teknokroma, Barcelona, Spain). They were then placed in 2-mL glass vials sealed using a PTFE/silicon septum. One microlitre of this filtrate was injected into the GC apparatus.
Analysis of methylphenol by gas chromatography-mass spectrometry
Ethylphenol was analysed using an Agilent Technologies 6890 N-MSD-5973 N gas chromatography-mass spectrometer (GC–MS) in single-ion mode (SIM) detection after liquid extraction with dichloromethane (Merck, Hohenbrunn, Germany). Ten millilitres of wine was mixed with 1 mL of dichloromethane, 100 μL of a 10 mg L−1 solution of 3,4-dimethylphenol (Merck, Hohenbrunn, Germany) as an internal standard, and 1.5 g of NaCl to produce a salting out effect. The mixture was vortex-stirred for 5 min and then centrifuged at 7,000 rpm for 10 min at 4 °C. The dichloromethane phase was filtered through 0.45-μm syringe membrane filters, and 1 μL injected into the GC–MS apparatus. A DB-WAX column (30 m × 0.25 mm internal diameter ×0.25 μm film thickness) (J & W Scientific, Folsom, CA, USA) was used for chromatographic separation; the split ratio used was 20:1. The oven temperature programme was 60 °C for 1 min, followed by a 3 °C min−1 ramp until 150 °C, followed by a 10 °C min−1 ramp to 210 °C, which was maintained for 2 min. The helium flow rate was 1 mL min−1. Detection was performed using SIM mode MS (target ion for 4-ethylphenol = 107). Calibration patterns were produced using reference compounds (Alfa Aesar, Karlsruhe, Germany).
Statistical analysis
Means and standard deviations were calculated, and ANOVA and least significant difference (LSD) tests performed using PC Statgraphics v.5 software (Graphics Software Systems, Rockville, MD, USA). Significance was set at p < 0.05 for the ANOVA matrix F value. The LSD test was used to detect significant differences between means. Multivariate statistical analyses of data were performed using principal components analysis (PCA).
Results and discussion
Viability of D. bruxellensis D37 following HHP treatment
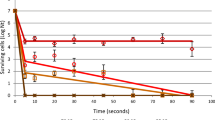
Following HHP treatment, viable D. bruxellensis D37 cells were sought by plating on DBDM medium (one of the most selective media available for these yeasts [27]. None of the inoculated wines (i.e. irrespective of the inoculum size or pH) subjected to HHP showed any viable cells. Further, 4-ethylphenol was only detected in the non-HHP-treated inoculated controls (at pH 3.2 and 3.6). In these non-pressurised control wines, populations of >104 cfu mL−1 were detected and 4-ethylphenol concentrations of >400 μg L−1 (Table 1).
Effect of pressurisation on wine pigment content
Irrespective of pH, the HHP-treated wines, contaminated or not, showed a smaller total anthocyanin content (by about 15 %) than the non-HHP-treated controls (Table 2). Indeed, these controls showed larger contents of all anthocyanin monomers, especially of malvidin-3-O-glucoside (Table 2). Thus, the final anthocyanin content was affected by HHP treatment. These results agree with those reported by [5]. According to these ANOVA results, the anthocyanin content is not affected by the presence of D. bruxellensis D37. However, when a PCA is performed considering all treatments (depending on: pH, HHP, inoculation and number of cells) regarding the contents of monomeric anthocyanins, the treatments can be separated in isolated groups (Fig. 1). Therefore, the inoculation of D. bruxellensis D37 influences the anthocyanin profile of the wine. The two principal components explain 83.9 % of the variance in this analysis.

Principal components analyses for different samples treatments (pH, HHP, Dekkera inoculation and amount of cells inoculated) regarding with the monomeric anthocyanin concentration
Some authors have reported Dekkera/Brettanomyces to reduce strongly wine anthocyanin content (for a review see [28], probably the manifestation of β-glucosidase activity [18]. This discrepancy with the present results may be explained in that the present work was performed with wines recently contaminated with D. bruxellensis D37, while these other authors made observations on wines that had been contaminated for much longer.
Irrespective of pH, the HHP-treated wines, contaminated or not, also showed a smaller vitisin A content than the non-HHP-treated controls (Table 3). Corrales et al. [5] reported the formation of certain types of pyranoanthocyanin (vitisin A-type) via condensation reactions during pressurisation but depending on temperature. In the present work, no such effect was seen probably because of the much less extreme pressure and temperature conditions employed.
Effect of pressurisation on wine volatile compound content
The main molecules studied were higher alcohols and esters. Only small differences (perhaps unnoticeable to consumers even though in some cases significant) were seen between the differently treated wines and controls (Table 4).
GC has not been used before to study the modification of the volatile compound profile of wines treated by HHP. However, the present results confirm those of other authors who pressurised wine at 400 MPa at 20 °C for 2 min [5], at 400–500 MPa at 4–20 °C for 5–15 min [25], and at 350 MPa at 25 °C for 20 min [19], and who used wine tasters to determine their effects.
Wine ester contents are very sensitive to thermal stabilisation treatments and freezing [16]. These molecules, which have fruity aromas and strongly influence the olfactory quality of wines, generally have low sensorial thresholds and are normally found in equilibrium with their hydrolysis products. They can therefore be understood as indicators of the ‘gentleness’ of a microbiological stabilisation treatment. In the present work, independent of the pH, no significant differences were seen in the ethyl acetate content of the HHP-treated controls and the HHP-treated wines inoculated with 104 cfu mL−1; in addition, the 106 cfu mL−1-inoculated HHP-treated wines showed only a <10 % lower ethyl acetate concentration than the same controls (Table 4). The results for ethyl lactate behaved in the same way. A PCA statistical analysis was performed to evaluate using a multivariate technique the repercussion of HHP treatment in aromatic quality. This analysis shows that 66 % of variance can be expressed by two components and also that all controls either pressurised or not and at the two studied pH are grouped in the centre of the figure (Fig. 2). Only the treatments inoculated with Dekkera were separated in the left and right of the figure depending on inoculation size. HHP treatment can therefore be considered gentle.

Principal components analyses for different samples treatments (pH, HHP, Dekkera inoculation and amount of cells inoculated) regarding with the concentration of aromatic compounds
Influence of HHP treatment on wine colour
Although HHP treatment (irrespective of pH or contamination with D. bruxellensis D37) led to some significant differences in colour intensity compared to the appropriate controls, no clear trend was seen with regard to the effects of inoculation or HHP treatment (Table 5). This agrees with that reported by other authors [7, 25]. However, lower pH (3.2) was associated with a reduction in the tonality (degree of ‘yellow’) of the controls (irrespective of HHP treatment or contamination with D. bruxellensis D37). Increasing tonality has been associated both with oxidation and ageing. Thus, tonality protection looks more related with low pH and not dependant on HHP treatment.
Industrial scale-up
HHP technique is enough mature to be used at industrial level in fact it is applied by many companies working in juice, meat, ham, dairy and other products. The cost of an industrial device to process by HHP ranges between 500,000 € and 1.5 million € [21]. The operational costs including amortisation in HHP processing can range between 0.045 €/kg to 0.087 €/kg depending on pressure, apparatus and production [14].
Conclusions
Traditional techniques for microbiologically stabilising wines, such as filtration, pasteurisation and flash pasteurisation at 60–70 °C all have a negative effect on their sensorial quality and reduce their polyphenolic compound, pigment and volatile compound contents. HHP treatment, however, seems to control D. bruxellensis D37 very well while largely avoiding these negative effects. The results therefore suggest that HHP might be a good alternative for controlling Dekkera/Brettanomyces in red wines.
References
Abalos D, Vejarano R, Morata A, González C, Suárez-Lepe JA (2011) The use of furfural as a metabolic inhibitor for reducing the alcohol content of model wines. Eur Food Res Technol 232:663–669
Benito S, Palomero F, Morata A, Calderón F, Suárez-Lepe JA (2009) A method for estimating Dekkera/Brettanomyces populations in wines. J Appl Microbiol 106:1743–1751
Calderón F, Morata A, Uthurry C, Suárez JA (2004) Aplicaciones de la ultrafiltración en la industria enológica. Últimos avances tecnológicos. Tecnología del Vino 16:49–54
Chatonnet P, Boidron J, Pons M (1990) Elevage des vins rouges en fûts de chêne: évolution de certains composés volatils et de leur impact arômatique. Science des Aliments 10:587–656
Corrales M, Butz P, Tauscher B (2008) Anthocyanin condensation reactions under high hydrostatic pressure. Food Chem 110:627–635
Delfidini C, Gaia P, Schellino R, Strano M, Pagliara A, Ambro S (2002) Fermentability of grape must after inhibition with dimethyl dicarbonate (DMDC). J Agric Food Chem 50:5605–5611
Delfini C, Conterno L, Carpi G, Rovere P, Tabusso A, Cocito C, Amati A (1995) Microbiological stabilisation of grape musts and wines by high hydrostatic pressures. J Wine Res 6:143–151
Dias L, Pereira-da-Silva S, Tavares M, Malfeito-Ferreira M, Loureiro V (2003) Factors affecting the production of 4-ethylphenol by the yeast Dekkera bruxellensis in enological conditions. Food Microbiol 20:377–384
Du Toit WJ, Pretorious IS, Lonvaud-Funel A (2005) The effect of sulphur dioxide and oxygen on the viability and culturability of a strain of Acetobacter pasterianus and a strain of Brettanomyces bruxellensis isolated from wine. J Appl Microbiol 98:862–871
Edlin DAN, Narbad A, Gasson MJ, Dickinson JR, Lloyd D (1998) Purification and characterization of hydroxycinnamate decarboxylase from Brettanomyces anomalus. Enzyme Microb Technol 22:232–239
Godoy L, Martínez C, Carrasco N, Ganga MA (2008) Purification and characterization of a p-coumarate decarboxylase and a vinylphenol reductase from Brettanomyces bruxellensis. Int J Food Microbiol 127:6–11
Harris V, Ford CM, Jiranek V, Grbin PR (2009) Survey of enzyme activity responsible for phenolic off-flavour production by Dekkera and Brettanomyces yeast. Appl Microbiol Biotechnol 81:1117–1127
Heier A, Blaas W, Drob A, Wittkowski R (2002) Anthocyanin analysis by HPLC/ESI–MS. Am J Enol Vitic 53:78–86
Jung S, Tonello Samson C, de Lamballerie M (2011) High hydrostatic pressure food processing. In: Proctor A (ed) Alternatives to conventional food processing. RSC Publishing, UK
Kurtzman CP, Fell JW (1998) Methods for the isolation, maintenance and identification of yeasts. In the yeast, a taxonomic study. Elsevier, Amsterdam, pp 79–80
Lambert Y, Demazeau G, Largeteau A, Bouvier J-M (1999) Changes in aromatic volatile composition of strawberry after high pressure treatment. Food Chem 67:7–16
Licker JL, Acree TE, Henick-Kling T (1999) What is ‘‘Brett’’ (Brettanomyces) flavour? A preliminary investigation. In: Waterhouse AL, Ebeler SE (eds) Chemistry of wine flavour. ACS symposium series., 714American Chemical Society, Washington, pp 96–115
Mansfield AK, Zoecklein BW, Whiton RS (2002) Quantification of glycosidase activity in selected strains of Brettanomyces bruxellensis and oenococcus oeni. Am J Enol Vitic 53:303–307
Mok C, Song K-T, Park Y-S, Lim S, Ruan R, Chen P (2006) High hydrostatic pressure pasteurization of red wine. J Food Sci 71:M265–M269
Morata A (2010a) Presiones hidrostáticas elevadas. In: Madrid Vicente A (ed) Nuevas tecnologías de conservación de alimentos. Antonio Madrid Vicente Ediciones, Madrid, pp 16–17
Morata A (2010b) Presiones hidrostáticas elevadas. In: Madrid Vicente A (ed) Nuevas tecnologías de conservación de alimentos. Antonio Madrid Vicente Ediciones, Madrid, p 32
Morata A, González MC, Suárez JA (2007) Formation of vinylphenolic pyranoanthocyanins by selected yeasts fermenting red grape musts supplemented with hydroxycinnamic acids. Int J Food Microbiol 116:144–152
Oelofse A, Lonvaud-Funel A, du Toit M (2009) Molecular identification of Brettanomyces bruxellensis strains isolated from red wines and volatile phenol production. Food Microbiol 26:377–385
Physter TG, Mills DA (2004) Novel methods to detect Brettanomyces (Dekkera) in wine. In Technical abstracts, 55th annual meeting, San Diego, CA, American Society for Enology and Viticulture, Davis, CA, p 30
Puig A, Vilavella M, Daoudi L, Guamis B, Minguez S (2003) Microbiological and biochemical stabilization of wines using the high pressure technique. Bulletin del l’OIV 76:596–617
Rayne S, Eggers NJ (2008) 4-ethylphenol and 4-ethylguaiacol concentrations in barreled red wines from the okanagan valley appellation, British Columbia. Am J Enol Vitic 59:92–97
Rodrigues N, Goncalves G, Pereira-da-Silva S, Malfeito-Ferreira M, Loureiro V (2001) Development and use of a new medium to detect yeast of the genera Dekkera/Brettanomyces ssp. J Appl Microbiol 90:588–599
Suárez R, Suárez-Lepe JA, Morata A, Calderón F (2007) The production of ethylphenols in wine by yeasts of the genera Brettanomyces and Dekkera: a review. Food Chem 102:10–21
Tchobanov L, Gal T, Nardi M, Guilloux-Benatier F, Remize J, Guzzo V, Serpaggi H, Alexandre H (2008) Partial vinylphenol reductase purification and characterization from Brettanomyces bruxellensis. FEMS Microbiol Lett 284:213–217
Acknowledgments
This work was funded by the Ministerio de Ciencia e Innovación (MICINN) (Project AGL-2008-05603-C02-01/AGR). The authors thank Mayekawa (Tokyo, Japan) for the use of the HHP apparatus in the pressurisation assays. The authors thank S. Somolinos and J. A. Sánchez (Depto. Tecnología de Alimentos, ETSI Agrónomos, UPM) for excellent technical assistance.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Morata, A., Benito, S., González, M.C. et al. Cold pasteurisation of red wines with high hydrostatic pressure to control Dekkera/Brettanomyces: effect on both aromatic and chromatic quality of wine. Eur Food Res Technol 235, 147–154 (2012). https://doi.org/10.1007/s00217-012-1742-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00217-012-1742-7