
Abstract
The function of the tomato K+, Na+/H+ antiporter LeNHX4 has been analyzed using 35S-driven gene construct for overexpressing a histagged LeNHX4 protein in Solanum lycopersicum L. Compared to wild-type plants, the expression of LeNHX4 was enhanced in most of plants transformed with a gene construct for LeNHX4 overexpression although some plants showed a decreased LeNHX4 expression. Overexpression of LeNHX4 was associated to an increased fruit size while silencing of this gene was related to a decreased fruit size. We have investigated the effect of LeNHX4 overexpression on fruit production and quality and we have also evaluated salt tolerance in two different overexpression lines by measuring proline, protein and glucose concentrations in tomato leaves grown either under control (0 mM NaCl) or saline (125 mM NaCl) conditions. Plants overexpressing LeNHX4 showed a higher amount of fruits than WT plants and accumulated higher contents of sugars and cations (Na+ and K+). The application of 125 mM NaCl, affected negatively fruit production and quality of WT plants. However the transgenic lines overexpressing LeNXH4 increased fruit quality and yield. In relation to salt tolerance, overexpression lines showed higher levels of leaf proline, glucose and proteins under NaCl treatment. The overexpression of LeNHX4 in tomato plants, improved salinity tolerance and increased fruit yield and quality under both normal and salinity stress conditions.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Soil salinity is the most severe factor limiting plant growth and crop productivity. This problem increases in arid and semi-arid regions. According to the report of FAO, over 6% of the world’s land is affected by either salinity or sodicity which accounts for more than 800 million ha of land throughout the world [1].
High levels of salinity may cause both ionic and osmotic stress effects, which lead to a decline of turgor, membrane damage, the inhibition of water and essential ions uptake, disordered metabolism, altered levels of growth regulators, enzymatic inhibition and metabolic dysfunction, including photosynthesis which ultimately leads to plant death [2,3,4,5,6]. On the other hand, potassium (K) is an essential element required for various physiological and biochemical processes that influence plant growth and development. In plants K+ is also a key player in the maintenance of osmotic adjustment and cell turgor [7] and plays an important role in enzyme activation, photosynthesis and respiration, assimilate transport, protein metabolism and stomatal regulation [8]. Due to similar physicochemical properties, Na+ and K+ ions have the potential to compete each other for uptake by plant-root cells. Under salt stress, the uptake of Na+ is increased drastically, causing a decrease in the absorption and accumulation of K+ and subsequent deficiency of this element [9,10,11].
Plant cell adaptation to salinity involves avoiding sodium toxicity and potassium loss (CITAS Adecuadas [12, 13]). Therefore, keeping a high K+/Na+ ratio in the cytosol is considered a fundamental process for salinity tolerance [14, 15]. For this purpose, plant cells have developed mechanisms of restriction of Na+ influx, active Na+ efflux, and compartmentalization of Na+ in the vacuole or other intracellular compartments [16]. Both Na+ efflux and Na+ compartmentalization need the operation of Na+/H+ antiporters located at the plasma membrane or intracellular membranes [17].
In previous works we have identified in tomato several Na+, K+/H+ antiporters belonging to the NHX family of proteins and determined their expression level in the salt sensitive cultivated species Solanum lycopersicum L. cv. Volgogradskij and the salt tolerant wild species Solanum pimpinelifolium L. in response to salt stress [18]. This study showed a higher expression of LeNHX4 in the wild tomato and an accumulation of Na+ in aerial parts of these plants. Interestingly, results from these studies showed that unlike other NHXs isoforms, tomato LeNHX4 is highly expressed in reproductive tissues and thus may have a role in fruit production and quality.
Regulation of osmotic adjustment under salt stress is also an important determinant of salt tolerance. In many plant species, the accumulation of compatible osmolytes (i.e., proline and sugars) is an important strategy leading to stress tolerance [19]. These molecules regulate the osmotic adjustment in plants grown under salt stress and enables plants to reestablish the water and osmotic homeostasis [20]. Proline is accumulated preferentially in leaves in order to maintain chlorophyll level and cell turgor to protect photosynthetic activity under salt stress [21]. In addition, soluble sugars are highly accumulated in vacuoles and produce high turgor pressure affecting osmotic potentials [22,23,24,25]. Interestingly, a relationship between NHX gene expression and sugar and proline accumulation has been reported [26,27,28,29,30].
Based on all the above, in this work we have studied whether the overexpression of the vacuolar Na+, K+/H+ antiporter LeNHX4 affect tomato fruit yield and quality under unstressed and NaCl stress cultivation conditions.
Materials and methods
Transformation and molecular characterization of transgenic plants
For stable expression of the LeNHX4 protein in tomato (S. lycopersicum L. cv. MicroTom), the LeNHX4 coding sequence to which the sequence for a C-terminal RGS(H)10 tag was added, was cloned under control of the 35S promoter in the pCAMBIA1303 plant expression vector as described. The plasmid pCAMBIA 1303 carrying the LeNHX4-RGS(H)10 fragment was transferred in the LB4404 Agrobacterium tumefaciens strain [31] and used for tomato cotyledon transformation as has been described by Ellul et al. [32].
The presence of the LeNHX4-RGS(H)10 construction in selected tomato transgenic lines was assessed by polymerase chain reaction (PCR) analysis using specific primers to amplify a 468 bp fragment of the hygromicin resistance gene harboured in the plant expression cassette (Forward: 5′-GATGTTGGCGACCTCGTATT-3′, Reverse: 5′-GTGCTTGACATTGGGGAGTT-3′) and DNA obtained from tomato leaves following a method by Edwards et al. [33].
The level of expression of LeNHX4 in leaves of untransformed and transgenic plants was assessed by real time PCR using gene-specific primers as previously described [18].
Plant material and growth conditions
Plants used for molecular characterization and fruit yield and quality determinations were cultivated in hydroponics for 4 weeks in 1/4 Hoagland nutrient solution [34]. For this purpose, seed were surface sterilized in ethanol 70% for 1 min followed by 50% commercial bleach for 5 min and 3 washes with sterile distilled water. Seeds were then cultivated in polystyrene boxes containing quartz sand, watered for 1 week with one-tenth strength Hoagland nutrient solution [34] and another 2 weeks with one-fourth strength of the same solution. Seedlings with four leaves were then transferred to pots for hydroponic cultivation. Typically, plants were grown for 4 weeks in a 2.5 L pot containing an aerated one-fourth strength Hoagland nutrient solution. Hydroponic cultivation was performed in a growth chamber at 24/20 °C day/night, under an illumination of 140 μmol/m2/s (photoperiod of 16 h light and 8 h darkness) and 40–50% relative humidity.
Alternatively, and in order to extend the growing period, tomato seeds were sown in seedbeds containing peat-moss. The seedbeds were kept in a greenhouse and irrigated with tap water for 7 weeks. 50-day-old tomato plants were transferred to 1.2 L pot (1 plant per pot) containing peat-moss and kept in a greenhouse. These plants were irrigated with tap water for 1 week and then with either tap water or 125 mM NaCl three times ever week for 16 weeks.
Determination of fruit quality parameterss
Ion content was measured in untransformed and transgenic tomato plants grown in hydroponics for 4 weeks in growth chamber under the conditions described above. Fruits were dried for 48 h at 80 °C, milled to powder and digested in a concentrated HNO3:HClO4 (2:1, v/v) solution. K+ and Na+ concentrations were determined in the digested material by inductively coupled plasma spectrometry (Varian ICP 720-ES).
Sugar analysis by GC–MS was carried out by a method modified from Schauer et al. [35]. Five tomato fruits were taken at postharvest stage to perform the GC–MS determinations.
Fruits were harvested and the fleshy, edible tissue was frozen in liquid nitrogen and crushed to powder with a grindomixer (Retsch, Haan, Germany). Samples were stored at − 80 °C until further analysis. For each individual fruit, frozen tomato tissue (150 mg) was extracted with 700 µL MeOH solution containing ribitol (9 µg ribitol/mL MeOH) as an internal standard. The mixture was extracted for 15 min at 70 °C and mixed vigorously with 700 µL of distilled water. To separate polar and non-polar metabolites, 325 µL chloroform was added to the mixtures. After centrifugation at 14,000 rpm for 5 min, the upper methanol/water phase (500 µL) was taken and dried overnight under vacuum.
The residue was re-dissolved in 40 µL of 20 mg/mL methoxyamine hydrochloride in pyridine and derivatized at 37 °C for 90 min followed by a 30 min treatment with 60 µL MSTFA (N-methyl-N-(trimethylsilyl)trifluoroacetamine) at 37 °C. Sample volumes of 1 µL were then injected into the GC column.
Total soluble solid (TSS) was measured as °Brix in a few drops of the juice using a digital refractometer (ATAGO Co., Ltd., Tokyo, Japan).
Tests for pH were performed on fruit homogenate using a portable pH meter.
Glucose, proline and protein determination in leaves
For glucose and proline determination samples of 0.5 g of plant materials were used by following the methods described by Irigoyen et al. [36] and Paquin and Lechasseur [37], respectively.
Fresh leaves were used for the determination of soluble proteins using Bradford G-250 reagent [38], and expressed as mg/g of fresh weight.
Statistics
All data in this report were obtained from at least three independent experiments with three or four replicates each. Analysis of variance was used to assess difference between treatments and plant genotypes were considered as significant when P ≤ 0.05. Significant differences according to the Duncan’s multiple range test (DMRT) are indicated with different letters in the figures and tables.
Results
Molecular characterization of transgenic plants
The cDNA corresponding to LeNHX4 gene was expressed under the cauliflower mosaic virus 35S gene promoter in tomato (S. lycopersicum L. cv MicroTom). T3 homozygous plants with a single insertion of the transgene were selected on the basis of PCR amplification of hygromicin resistance gene and the analysis of hygromicin resistance segregation after seed germination in selective media containing the antibiotic (Supporting Information Fig. S1). From these plants, three independent lines, L-2, L-3 and L-5, were further characterized. Real-time RT-PCR analysis showed a higher expression level of LeNHX4 in leaves of L-3 and L-5 transgenic lines than in those of untransformed controls (Fig. 1a). Surprisingly, the expression level of LeNHX4 in plants from line L-2 was half of that in WT plants, which indicate the silencing of LeNHX4 in this line (Fig. 1a).

a Transcript levels of LeNHX4 in leaves of WT and transgenic plants. Plants were cultivated in hydroponics for 4 weeks. The results show the expression of LeNHX4, quantified by real-time qPCR and expressed as increase in transcript levels relative to the level in leaves of WT plants, to which value 1 is assigned. Values are means ± standard deviation of three independent experiments with three replicates each. Means that have different letters at the top of each bar are significantly different at P ≤ 0.05. b Image of the fruits obtained from WT and transgenic plants (lines L-3 and L-2). It can be observed that relative to WT plants, the low expression of LeNHX4 in L2 causes a lower fruit size while, while the high expression of the gene in L3 line provokes an increase of fruit size
Characterisation of fruits
Since LeNHX4 is highly expressed in fruits [18] we tested whether overexpression of LeNHX4 have an effect on fruit yield. Fruit size was related to the level of expression of LeNHX4 in the transgenic lines (Fig. 1b). Plants ovexpressing LeNHX4 (lines 3 and 5) showed the highest fruit size, while plants in which LeNHX4 (line 2) was silenced showed the lowest fruit size. Fruit production was measured in untransformed and transgenic tomato plants overexpressing LeNHX4 grown in hydroponics for 4 weeks in Hoagland nutrient solution (Fig. 2a). Our data demonstrate that overexpression of LeNHX4 lead to a significant increase in fruit production, while the lowest production was reached in WT plants. Fruit fresh weight (Fig. 2b) showed trends similar to those of fruit production, reaching high values in transgenic lines L-3 and L-5, exceeding WT by 61% and 88% respectively.

Effect of LeNHX4 overexpression on fruit production (a), fruit weight (b), water content (c) and soluble sugars (d) of tomato plants. WT and LeNHX4 plants were cultivated in hydroponics for 4 weeks. Values are means ± standard deviation of three independent experiments with three replicates each. Means that have different letters at the top of each bar are significantly different at P ≤ 0.05
The overexpression of LeNHX4 increased the fruit water content respect to the value in WT plants (Fig. 2c). The concentrations of glucose, fructose and sucrose were determined in fruits of WT plants and plants overexpressing LeNHX4 in order to study the effect of the overexpression of LeNHX4 on the quality of tomato fruits (Fig. 2d). Fructose and glucose levels were strongly increased in transgenic plants in comparison to WT plants. However, the concentration of sucrose was not significantly different between WT and transgenic plants.
Figure 3 shows the contents of Na+ and K+ in fruits of WT plants and plants overexpressing LeNHX4. The highest Na+ contents were found in the LeNHX4 overexpressing lines, with values of 0.10 and 0.06% of dry weight, respectively, while the WT fruits reached the lowest value. Similar results were obtained for fruit K+ content, with transgenic lines L-3 and L-5 showing 14 and 18% higher K+ content than WT fruits.

K+ (a) and Na+ (b) contents in fruit of WT and transgenic tomato plants overexpressing LeNHX4. Plants were cultivated in hydroponics for 4 weeks. Values are means ± standard deviation of three independent experiments with three replicates each. Means that have different letters at the top of each bar are significantly different at P ≤ 0.05
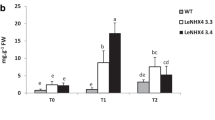
The effect of LeNHX4 overexpression in fruit yield and quality was also studied in plants grown in the presence or the absence of NaCl for longer time periods. Our results demonstrate that plants overexpressing LeNHX4 showed higher fruit production that WT plants when both were irrigated either with or without NaCl (Fig. 4a). Compared to untreated plants salt-treated wild type plants reduced fruit production by 33%, while fruit production by LeNHX4 overexpressing plants was slightly increased as a result of salt treatment (Fig. 4a).

Effect of LeNHX4 overexpression on fruit production (a) and glucose (b), proline (c) and protein concentrations (d) of tomato plants. WT and LeNHX4 plants were cultivated in peat-moss for 16 weeks in the absence (empty bars) or the presence (black bars) of 125 mM NaCl. Values are means ± standard deviation of three independent experiments with three replicates each. Means that have different letters at the top of each bar are significantly different at P ≤ 0.05
Fruit quality characteristics such as total soluble solids (TSS% or °Brix), and pH are shown in Table 1. Under normal conditions (without 125 mM NaCl), transgenic lines overexpressing LeNHX4 showed a significant reduction in TSS. In relation to pH, we have not found significant differences between WT and both transgenic lines. However, the addition of 125 mM NaCl to the irrigation solution significantly increased total soluble solids in LeNHX4 overexpressing lines and decreases it in WT plants. Similarly, the pH decreased in WT and increased in transgenic lines under salinity conditions.
Glucose, proline and proteins in leaves of WT and transgenic plants overexpressing LeNHX4
Figure 4b shows the glucose concentration in leaves of WT and the transgenic lines overexpressing LeNHX4 (lines L-3 and L-5). The NaCl treatment increases the glucose level in all plants, above all in LeNHX4 overexpressing plants.
Our results show that NaCl treatment significantly increased the proline content in WT and LeNHX4 overexpressing plants, being the increase more pronounced in the transgenic plants (Fig. 4c).
Figure 4d shows the effect of NaCl treatment on protein contents. Significant differences were found in the protein content between untreated and treated plants. Under NaCl stress, transgenic plants showed higher protein content than untreated plants, while the protein level in WT plants was negatively affected.
Discussion
Salt stress is a major abiotic factor limiting plant growth and productivity. Plants exhibit various physiological and biochemical responses to salt stress. An important mechanism to overcome salt stress is the exclusion of Na+ from the cytoplasm, by the operation of Na+/H+ antiporters at the plasma membrane or tonoplast. Plant NHX antiporters play a key role in NaCl tolerance by the extrusion of Na+ out of cytosol [18, 39].
In tomato, four LeNHX proteins were reported by Gálvez et al. [18]. From these proteins, LeNHX2 was studied in relation to its role in salt tolerance. Indeed, Huertas et al. [40] showed that tomato plants overexpressing LeNHX2 grew better in the presence of 120 mM NaCl than untransformed controls and Baghour et al. [14] demonstrated that the joint overexpression of LeNHX2 and SlSOS2 increased even more the plant salt tolerance. In addition to the increased salt tolerance, an increased fruit production was found in plants overexpressing LeNHX2, SlSOS2 or both [14, 41]. In relation to this, Bassil et al. [42] reported that AtNHX1 and AtNHX2 are required for growth and floral development in Arabidopsis, while AtNHX3 and AtNHX4 play role in seed development [43, 44]. More recently, Sharma et al. [45], showed that in Triticum aestivum the TaNHX4 group genes were highly expressed in later development stages of leaf, spike and grain.
In previous work, we have demonstrated that LeNHX4 is highly expressed in flowers and fruits of tomato plants, indicating specific roles of this isoform in the reproductive tissues [18]. In this work, data of fruit production of tomato plants cultivated in media lacking NaCl indicate an improvement of fruit yield in transgenic lines overexpressing the LeNHX4 antiporter (Fig. 2a, b), suggesting a role of LeNHX4 in fruit production. Fruit of tomato plants overexpressing LeNHX4 showed a higher water content than those of WT plants (Fig. 2c), which together with the higher accumulation of sugars (Fig. 2d), K+ and Na+ (Fig. 3) in fruits of these plants under NaCl cultivation conditions could suggest that plants overexpressing LeNHX4 withstand better a shortage of water under salinity conditions. In agreement with our results, Li et al. [46] showed that Arabidopsis transgenic plants overexpressing RtNHX1 increased their capacity for osmotic adjustment and improved salinity tolerance due to a high relative water content and accumulation of solutes allowing osmoregulation.
The total soluble solids (TSS) and pH are important quality parameters, which plays a significant role in fruit selection. Reina-Sánchez et al. [47] recorded that an increase in fruit quality represented an increase of total soluble solids and titratable acidity. Salinity positively affect the levels of TSS, including sugars, organic acids, and amino acids in fruits [48,49,50,51,52,53,54]. Salinity stress enhances gluconeogenesis, as well as metabolic flow in ripe fruit, resulting in high-Brix fruit [54]. In relation to the role of NHX ion transporters on fruit quality Hanana et al. [55] reported that the NHX antiporters play a critical role in plant growth and development, and improve organoleptic characteristics of fruits as well as those related to fruit ripening [41]. Contrary to what was expected, TSS levels are lower in overexpressing lines than in wild type plants cultivate in the absence of NaCl. However the overexpression of LeNHX4 gene improved the fruit quality in tomato by increasing the solid solute content and pH under salt stress (Table 1). As opposed to our results, Zhang and Blumwald [56] reported that overexpression in tomato of the Arabidopsis antiporter AtNHX1 did not affect the total soluble solids content in plants grown under 200 mM NaCl.
The increase in the concentration of soluble sugars such as glucose enhances plant tolerance to several abiotic stresses, such as drought, salinity and cold [57] because of their involvement in osmotic adjustment. Our results in tomato showed an increase of glucose and fructose content in fruits of transgenic plants under normal conditions (Fig. 2d) and suggest that the increased expression of LeNHX4 could have a positive effect on glucose synthesis under non stressful conditions. Similar results were reported by Leidi et al. [26], who reported that transgenic tomato plants overexpressing AtNHX1 accumulated higher level of free sugars even if they were not transferred to salt. NaCl treatment significantly increased the concentration of glucose in plants overexpressing LeNHX4 in relation to WT plants (Fig. 4b). Lieu et al. [58] did not find any significant differences on the sugar contents of WT and transgenic Beta vulgaris plants overexpressing AtNHX3. However, with the application of high NaCl treatment (300 or 500 mM), a greater sugar accumulation was observed in transgenic plants, suggesting that the expression of AtNHX3 in sugar beet improved sugar synthesis in transgenic plants and increased the plants salt tolerance by influencing the transcription of genes involved in the sugar synthesis pathways.
A large number of studies show a positive correlation between proline accumulation and plant stress and proline concentration has been shown to be generally higher in salt tolerant than in salt sensitive plants. The accumulation of osmotic compounds such as proline is a common response of plants to high salinity to combat osmotic stress [59, 60]. Proline is accumulated preferentially in leaves in order to maintain chlorophyll level and cell turgor to protect photosynthetic activity under salt stress [21]. Plants overexpressing NHXs show an enhanced capacity for osmotic adjustment and improved salinity tolerance due to a better relative water content and accumulation of proline [26,27,28,29,30]. Our data showed that under control conditions, the proline content in leaves was slightly increased in the transgenic lines (Fig. 4c). However, when exposed to saline conditions, the level of this aminoacid dramatically increased in plants overexpressing LeNHX4. Recently, Wang et al. [61] reported that the overexpression of Ipomoea batatas IbNHX2 significantly increased proline content, superoxide dismutase (SOD) activity, and photosynthetic rate in sweet potato. Similarly, transgenic Arabidopsis plants overexpressing the Na+/H+ antiporter RtNHX1 from Reaumuria trigyna accumulated more proline than WT and nhx1 mutant plants under 200 mM NaCl treatment [46]. The increase of proline during salt stress has been associated to the up-regulation of proline biosynthesis genes (AtP5CS1 and AtP5CS2) in transgenic plants [46, 61].
There are many evidences that proteins could have a role in osmotic adjustment [62,63,64]. Plants are also able to respond and adapt to salt stress through the synthesis of specific proteins, and the synthesis of stress-induced proteins is part of that stress tolerance mechanism [65]. Our results show that the NaCl treatment affected negatively the protein content in WT plants (Fig. 4d). However, the overexpression of LeNHX4 increased the protein content in transgenic lines L-3 and L-5. These results are consistent with those reported by Tester and Davenport [66], who suggested that the disruption in protein synthesis appears to be an important cause of damage caused by Na+. Comparing the total soluble protein content between the salt-tolerant cultivar (Rio Grande) and the salt-sensitive one (Heinz-2274), Kahlaoui et al. [67] found that the protein levels significantly increased more in the salt-tolerant tomato cultivar when irrigated with saline water. The accumulation of these proteins under saline conditions may provide a storage of nitrogen that is re-utilized when stress is over [68] and may play a role in osmotic adjustment [67]. However, there are also studies reporting no differences in protein content as a result of NaCl tolerance. Ashraf and Fatima [69] found that salt-tolerant and salt-sensitive safflower accessions did not differ significantly in soluble leaf proteins.
In conclusion, the results reported in this study indicate that overexpression of LeNHX4 in tomato increases size, sugar (glucose and fructose) and ion (Na+ and K+) contents of fruits. Moreover, relative to WT plants, plants overexpressing LeNHX4 show an enhanced fruit yield under both non-stress and salinity stress conditions as well as higher levels of osmoregulatory compounds (proline and proteins) in leaves.
References
FAO (2008) Land and plant nutrition management service. http://www.fao.org/ag/agl/agll/spush
Zhu JK (2001) Plant salt tolerance. Trends Plant Sci 6:66–71
Mahajan S, Tuteja N (2005) Cold, salinity and drought stresses: an overview. Arch Biochem Biophys 444:139–158
Kim JK, Bamba T, Harada K, Fukusaki E, Kobayashi A (2007) Time-course metabolic profiling in Arabidopsis thaliana cell cultures after salt stress treatment. J Exp Bot 58:415–424
Munns R, Tester M (2008) Mechanisms of salinity tolerance. Annu Rev Plant Biol 59:651–668
Hasanuzzaman M, Hossain MA, da Silva JAT, Fujita M (2012) Plant responses and tolerance to abiotic oxidative stress: antioxidant defenses is a key factors. In: Bandi V, Shanker AK, Shanker C, Mandapaka M (eds) Crop stress and its management: perspectives and strategies. Springer, Berlin, pp 261–316
Osakabe Y, Yamaguchi-Shinozaki K, Shinozaki K, Tran LS (2014) ABA control of plant macroelement membrane transport systems in response to water deficit and high salinity. N Phytol 202:35–49
Pettigrew WT (2008) Potassium influences on yield and quality production for maize, wheat, soybean and cotton. Physiol Plant 133:670–681
Miransari M, Smith DL (2007) Overcoming the stressful effects of salinity and acidity on soybean nodulation and yields using signal molecule genistein under field conditions. J Plant Nutr 30:1967–1992
Wakeel A, Farooq M, Qadir M, Schubert S (2011) Potassium substitution by sodium in plants. Crit Rev Plant Sci 30:401–413
Wakeel A (2013) Potassium–sodium interactions in soil and plant under saline-sodic conditions. J Plant Nutr Soil Sci 176:344–354
Serrano R, Gaxiola R (1994) Microbial models and salt stress tolerance in plants. Crit Rev Plant Sci 13:121–138
Hasegawa PM, Bresan RA, Zhu JK, Bohnert HJ (2000) Plant cellular and molecular responses to high salinity. Annu Rev Plant Physiol Plant Mol Biol 51:463–499
Baghour M, Gálvez FJ, Sánchez ME, Aranda MN, Venema K, Rodríguez-Rosales MP (2019) Overexpression of LeNHX2 and SlSOS2 increases salt tolerance and fruit production in double transgenic tomato plants. Plant Physiol Biochem 135:77–86
Cagnac O, Baghour M, Jaime-Pérez N, Aranda MN, Sánchez ME, Rodríguez-Rosales MP, Venema K (2020) Deletion of the N-terminal domain of the yeast vacuolar (Na+, K+)/H+ antiporter Vnx1p improves salt tolerance in yeast and transgenic Arabidopsis. Yeast 37:173–185
Niu X, Bressan RA, Hasegawa PM, Pardo JM (1995) Ion homeostasis in NaCl stress environments. Plant Physiol 109:735–742
Baghour M, Ben Chekroun K, Rodríguez-Rosales MP, Venema K (2010) Antiporters: role in salinity tolerance (a review). Moroc J Biol 6–7:16–22
Gálvez FJ, Baghour M, Hao G, Cagnac O, Rodríguez-Rosales MP, Venema K (2012) Expression of LeNHX isoforms in response to salt stress in salt sensitive and salt tolerant tomato species. Plant Physiol Biochem 51:109–115
Huang Z, Zhao L, Chen D, Liang M, Liu Z et al (2013) Salt stress encourages proline accumulation by regulating proline biosynthesis and degradation in Jerusalem Artichoke plantlets. PLoS ONE 8:e62085. https://doi.org/10.1371/journal.pone.0062085
Zhu JK (2001) Cell signaling under salt, water and cold stresses. Curr Opin Plant Biol 4:401–406
Silva-Ortega CO, Ochoa-Alfaro AE, Reyes-Agüerob JA, Aguado-Santacruz GA, Jimenez-Bremont JF (2008) Salt stress increases the expression of P5CS gene and induces proline accumulation in Cactus pear. Plant Physiol Biochem 46:82–92
Moustakas M, Sperdouli I, Kouna T, Antonopoulou CI, Therios I (2011) Exogenous proline induces soluble sugar accumulation and alleviates drought stress effects on photosystem II functioning of Arabidopsis thaliana leaves. Plant Growth Regul 65:315–325
Rasheed R, Wahid A, Farooq M, Hussain I, Basra SM (2011) Role of proline and glycine betaine pretreatments in improving heat tolerance of sprouting sugarcane (Saccharum sp.) buds. Plant Growth Regul 65:35–45
Gibson SI (2005) Control of plant development and gene expression by sugar signaling. Curr Opin Plant Biol 8:93–102
Ma QJ, Sun MH, Lu J, Liu YJ, Hu DG, Hao YJ (2017) Transcription factor AREB2 is involved in soluble sugar accumulation by activating sugar transporter and amylase genes. Plant Physiol 174:2348–2362
Leidi EO, Barragan V, Rubio L, El-Hamdaoui A, Ruiz T, Cubero B, Fernández JA, Bressan RA, Hasegawa PM, Quintero FJ, Pardo JM (2010) The AtNHX1 exchanger mediates potassium compartmentation in vacuoles of transgenic tomato. Plant J 61:495–506
Mishra S, Alavilli H, Lee B, Panda SK, Sahoo L (2014) Cloning and functional characterization of a vacuolar Na+/H+ antiporter gene from Mungbean (VrNHX1) and its ectopic expression enhanced salt tolerance in Arabidopsis thaliana. PLoS ONE 9:e106678. https://doi.org/10.1371/journal.pone.0106678
Mishra S, Alavilli H, Lee B, Panda SK, Sahoo L (2015) Cloning and characterization of a novel vacuolar Na+/H+ antiporter gene (VuNHX1) from drought hardy legume, cowpea for salt tolerance. Plant Cell Tissue Organ Cult 120:19–33
Metwali EMR, Soliman HIA, Fuller MP, Al-Zahrani HS, Howladar SM (2015) Molecular cloning and expression of a vacuolar Na+/H+, antiporter gene (AgNHX1) in fig (Ficus carica L.) under salt stress. Plant Cell Tissue Organ Cult 123(2):377–387
Pehlivan N, Sun L, Philip J, Yang X, Mishra N, Chen L, Kadioglu A, Shen G, Zhang H (2016) Co-overexpressing a plasma membrane and a vacuolar membrane sodium/proton antiporter significantly improves salt tolerance in transgenic Arabidopsis plants. Plant Cell Physiol 57:1069–1084
Hoekema A, Hirsch PR, Hooykaas PJJ, Schilperoort RA (1983) Binary vector strategy based on separation of vir- and T-region of the Agrobacterium tumefaciens Ti-plasmid. Nature 303:179–180
Ellul P, García-Sogo B, Pineda B, Ríos G, Roig LA, Moreno V (2003) The ploidy level of transgenic plants in Agrobacterium-mediated transformation of tomato cotyledons (Lycopersicum esculentum Mill.) is genotype and procedure dependent. Theor Appl Genet 106:231–238
Edwards K, Johnstone C, Thompson C (1991) A simple and rapid method for the preparation of plant genomic DNA for PCR analysis. Nucleic Acid Res 19:1349
Hoagland DR, Arnon DI (1950) The water-culture method for growing plants without soil. Calif Agric Exp Circ 347:1–32
Schauer N, Zamir D, Fernie AR (2005) Metabolic profiling of leaves and fruit of wild species tomato: a survey of the Solanum lycopersicum complex. J Exp Bot 56:297–307
Irigoyen JJ, Emerich DW, Sánchez-Díaz M (1992) Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativa) plants. Physiol Plant 84:55–60
Paquin R, Lechasseur P (1979) Observations sur une méthode de dosage de la proline libre dans les extraits de plantes. Can J Bot 57:1851–1854
Bradford MM (1976) A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Rodriguez-Rosales MP, Galvez FJ, Huertas R, Aranda MN, Baghour M, Cagnac O, Venema K (2009) Plant NHX cation/proton antiporters. Plant Signal Behav 4(4):265–276
Huertas R, Rubio L, Cagnac O, García-Sánchez MJ, Alché JD, Venema K, Fernández JA, Rodríguez-Rosales MP (2013) The K+/H+ antiporter LeNHX2 increases salt tolerance by improving K+ homeostasis in transgenic tomato. Plant Cell Environ 36:2135–2149
Huertas R, Olías R, Eljakaoui Z, Gálvez FJ, Li J, De Morales PA, Belver A, Rodríguez-Rosales MP (2012) Overexpression of SlSOS2 (SlCIPK24) confers salt tolerance to transgenic tomato. Plant Cell Environ 35:1467–1482
Bassil E, Ohto M, Esumi T, Tajima H, Zhu Z, Cagnac O, Belmonte M, Peleg Z, Yamaguchi T, Blumwald E (2011) The Arabidopsis intracellular Na+/H+ antiporters NHX5 and NHX6 are endosome associated and necessary for plant growth and development. Plant Cell 23:224–239
Li HT, Liu H, Gao XS, Zhang H (2009) Knock-out of Arabidopsis AtNHX4 gene enhances tolerance to salt stress. Biochem Biophys Res Commun 382:637–641
Liu H, Tang R, Zhang Y, Wang C, Lv Q, Gao X, Li W, Zhang H (2010) AtNHX3 is a vacuolar K+/H+ antiporter required for low-potassium tolerance in Arabidopsis thaliana. Plant Cell Environ 33:1989–1999
Sharma H, Taneja M, Upadhyay SK (2019) Identification, characterization and expression profiling of cation-proton antiporter superfamily in Triticum aestivum L. and functional analysis of TaNHX4-B. Genomics. https://doi.org/10.1016/j.ygeno.2019.02.015
Li N, Wang X, Ma B, Du C, Zheng L, Wang Y (2017) Expression of a Na+/H+ antiporter RtNHX1 from a recretohalophyte Reaumuria trigyna improved salt tolerance of transgenic Arabidopsis thaliana. J Plant Physiol 218:109–120
Reina-Sánchez A, Romero-Aranda R, Quartero J (2005) Plant water uptake and water use efficiency of greenhouse tomato cultivars irrigated with saline water. Agric Water Manag 78:54–66
Tal M, Katz A, Heikin H, Dehan K (1979) Salt tolerance in the wild relatives of the cultivated tomato: proline accumulation in Lycopersicon esculentum Mill., L. peruvianum Mill. and Solanum pennellii Cor. treated with NaCl and polyethyleneglycol. N Phytol 82:349–355
Ho LC, Grange RI, Picken AJ (1987) An analysis of the accumulation of water and dry matter in tomato fruit. Plant Cell Environ 10:157–162
Adams P (1991) Effects of increasing the salinity of the nutrient solution with major nutrients or sodium chloride on the yield, quality and composition of tomatoes grown in rockwool. J Hortic Sci 66:201–207
Balibrea M, Martinez-Andújar C, Cuartero J, Bolarín M, Pérez-Alfocea F (2006) The high fruit soluble sugar content in wild Lycopersicon species and their hybrids with cultivars depends on sucrose import during ripening rather than on sucrose metabolism. Funct Plant Biol 33:279–288
Gao Z, Sagi M, Lips SH (1998) Carbohydrate metabolism in leaves and assimilate partitioning in fruits of tomato (Lycopersicon esculentum L.) as affected by salinity. Plant Sci 135:149–159
Krauss SW, Schnitzler H, Grassmann J, Woitke M (2006) The influence of different electrical conductivity values in a simplified recalculating soilless system on inner and outer fruit quality characteristics of tomato. J Agric Food Chem 54:441–448
Saito T, Matsukura C, Ban Y, Shoji K, Sugiyama M, Fukuda N, Nishimura S (2008) Salinity stress affects assimilate metabolism at the gene-expression level during fruit development and improves fruit quality in tomato (Solanum lycopersicum L.). J Jpn Soc Hortic Sci 77:61–68
Hanana M, Cagnac O, Zarrouk M, Blumwald E (2009) Rôles biologiques des antiports vacuolaires NHX : acquis et perspectives d’amélioration génétique des plantes. Botanique 87:1023–1035
Zhang HX, Blumwald E (2001) Transgenic salt-tolerant tomato plants accumulate salt in foliage but not in fruit. Nat Biotechnol 19:765–768
Rathinasabapathi B (2000) Metabolic engineering for stress tolerance: installing osmoprotectant synthesis pathways. Ann Bot 86:709–716
Liu H, Wang Q, Yu M, Zhang Y, Wu Y, Zhang H (2008) Transgenic salt-tolerant sugar beet (Beta vulgaris L.) constitutively expressing an Arabidopsis thaliana vacuolar Na+/H+ antiporter gene, AtNHX3, accumulates more soluble sugar but less salt in storage roots. Plant Cell Environ 31:1325–1334
Hayat S, Hayat Q, Alyemeni MN, Wani AS, Pichtel J, Ahmad A (2012) Role of proline under changing environments: a review. Plant Signal Behav 7:1456–1466
Gharsallah C, Fakhfakh H, Grubb D, Gorsane F (2016) Effect of salt stress on ion concentration, proline content, antioxidant enzyme activities and gene expression in tomato cultivars. AoB Plants. https://doi.org/10.1093/aobpla/plw055
Wang B, Zhai H, He S, Zhang H, Ren Z, Zhang D, Liu Q (2016) A vacuolar Na+/H+, antiporter gene, IbNHX2, enhances salt and drought tolerance in transgenic sweet potato. Sci Hortic 201:153–166
Mansour M (2000) Nitrogen containing compounds and adaptation of plants to salinity stress. Biol Plant 43:491–500
Ashraf M, Harris PGC (2004) Biochemical indicators of salinity tolerance in plant. Plant Sci 166:3–16
Parvaiz A, Satyawati S (2008) Salt stress and phyto-biochemical responses of plants—a review. Plant Soil Environ 54:89–99
Veeranagamallaiah G, Chandraobulreddy P, Jyothsnakumari G, Sudhakar C (2007) Glutamine synthetase expression and pyrroline-5-carboxylate reductase activity influence proline accumulation in two cultivars of foxtail millet (Setaria italica L.) with differential salt sensitivity. Environ Exp Bot 60:239–244
Tester M, Davenport R (2003) Na+ tolerance and Na+ transport in higher plants. Ann Bot 91:503–507
Kahlaoui B, Hachicha M, Misle E, Fidalgo F, Teixeira J (2018) Physiological and biochemical responses to the exogenous application of proline of tomato plants. J Saudi Soc Agric Sci 17:17–23
Singh NK, Bracken CA, Hasegawa PM, Handa AK, Buckel S, Hermodson MA, Pfankoch F, Regnier FE, Bressan RA (1987) Characterization of osmotin. A thaumatin-like protein associated with osmotic adjustment in plant cells. Plant Physiol 85:529–536
Ashraf M, Fatima H (1995) Responses of some salt tolerant and salt sensitive lines of safflower (Carthamustin ctorius L.). Acta Physiol Plant 17:61–71
Acknowledgements
This work was supported by Grants from Consejería de Economía, Innovación, Ciencia y Empresa, Junta de Andalucía, Spain (CVI-7558 to MPRR), Spanish Ministry of Economy and Competitiveness and Agencia Estatal de Investigación (BIO2015-65056-P, BIO2016-81957-REDT/AEI and Programa I-COOPB+2013 Ref. COOPB20053). National Centre for Scientific and Technical Research and Minister for Higher Education, Scientific Research and Executive Training (Morocco).
Author information
Authors and Affiliations
Contributions
Study concepts: MB, KV, MPRR. Study design: MB, KV, MPRR. Literature research: MM, MB, MPRR. Experimental studies: MM, FJG, MES, MNA. Data analysis/interpretation: MM, MB, MA, FJG, KV, MPRR. Statistical analysis: MM, MB. Manuscript preparation: MM, MB, MPRR. Manuscript revision: MB, MPRR.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
This manuscript does not imply human participants or studies on animals.
Informed consent
Not required.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Maach, M., Baghour, M., Akodad, M. et al. Overexpression of LeNHX4 improved yield, fruit quality and salt tolerance in tomato plants (Solanum lycopersicum L.). Mol Biol Rep 47, 4145–4153 (2020). https://doi.org/10.1007/s11033-020-05499-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11033-020-05499-z