
Abstract
Agriculture is undergoing a paradigm shift as it moves away from relying only on agrochemicals toward natural-based product to enhance plant growth and productivity while sustainably maintaining soil quality and productivity. In this sense, microalgae and bacteria offer a unique potential due to the growing use of novel and eco-friendly products such as biofertilizers, biostimulants, and biopesticides. Microalgae improve crop growth and health by fixing nitrogen, releasing soil trace elements, solubilizing potassium, and phosphorus, producing exopolysaccharides, and converting organic matter into utilizable nutrients. They also release bioactive substances including, carbohydrates, proteins, enzymes, vitamins, and hormones, to promote plant growth, control pests, and mitigate plant stress responses. Even though it has long been known that microalgae produce various bioactive and signaling molecules (like phytohormones, polysaccharides, lipids, carotenoids, phycobilins, and amino acids) which are effective in crop production, the targeted applications of these molecules in plant science are still in the very early stages of development. Microalgae are beneficial to bacteria because they produce oxygen and extracellular chemicals, and bacteria, in turn, provide microalgae with carbon dioxide, vitamins, and other nutrients in exchange. This review discusses the possible role of microalgae in increasing crop yield, protecting crops, and maintaining soil fertility and stability, and it points out that interactions of microalgae and bacteria may have a better enhancement of crop production in a sustainable way than using either of them alone.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
There has been a global tension between ever-growing demand for food, water, and energy sources, which calls for novel and sustainable approaches to increase agricultural productivity and maintain the environment. It is generally believed that sustainable agricultural intensification should be considered the issues of increasing production and reducing environmental damage. However, the current crop production system has become strongly dependent on agrochemicals, which have caused considerable damage to global ecological security such as acidification and hardening, decreasing beneficial soil microorganisms, and increasing disease incidence (Chandini et al. 2019; (Meena et al. 2020b). Modern agriculture must assess its methods by integrating new systems to produce food sustainably. A novel and eco-friendly approach to addressing these challenges involves the development of microalgae-bacteria based products such as biofertilizers, biostimulants, and biopesticides, which reduce reliance on agrochemicals and achieve higher production and sustainable value in modern agriculture with minimalised the negative effects on agroecosystem. Microalgae and beneficial bacteria can be used alone or in consortiums as an alternative source of chemical fertilizers to enhance plant growth, nutrient cycling, plant protection, productivity, and soil fertility (Garcia-Gonzalez and Sommerfeld 2016; Holajjer et al. 2013; Niu et al. 2020; Singh et al. 2011).
The use of microalgae in agriculture is becoming increasingly implementing due to the wide adaptability, includes capacity for heterotrophic growth, high photosynthetic efficiency, capability of adapting to both industrial and domestic wastewater, and production of biomolecules (Chiaiese et al. 2018). The utilization of microalgae biomass can be beneficial because it has a highest nitrogenase activity (Prasanna et al. 2014, 2016b; Ranjan et al. 2016), carbon and nitrogen fixation (Song et al. 2022), enhancing stability of soil aggregates and soil ecological health (Nichols et al. 2020; Song et al. 2022), prevention of nutrient losses (Jimenez et al. 2020), and improving soil fertility (Alvarez et al. 2021). Studies in these areas have increased our understanding of the positive roles of biomolecules produced by microalgae, including carbohydrates, proteins, enzymes, fatty acids, organic acids, vitamins, and hormones, which improve soil health, aid in plant disease management, and mitigate the effects of climatic stresses (Alvarez et al. 2021; Nichols et al. 2020; Prasanna et al. 2013). Plant nutrient access is dependent on soil microbes, which stimulate plant growth and improve crop yield through phosphate solubilization, potassium solubilization, biocontrol activity, exopolysaccharide secretion, and organic matter decomposition (Naik et al. 2019; Souza et al. 2015).
Microalgae and bacteria can interact with one another in a broad variety of ways, ranging from mutualism and commensalism to competition and parasitism ((Fuentes et al. 2016b). Microalgae improve soil health, root growth, and shoot growth, as well as crop productivity by functioning independently or in association with beneficial bacteria (Kang et al. 2021). It is evident that microalgae and bacteria have enormous potential and present a significant chance to improve agriculture’s sustainability and resilience when considered alongside the rising costs of synthetic fertilizers, the effect of climate change, and pesticide resistance. However, the interaction processes among microalgae and bacteria are still poorly understood, especially how to integrate this information for agricultural purpose, renewable energy production, and other biotechnological fields. We give a detailed review highlighting the most recent investigation on potential microalgae and their interaction with bacteria for sustainable agricultural production.
Potential of microalgae in crop production
Algae are photosynthetic organisms that can be found in a different variety of water and soil environments. Algae are generally classified as macroalgae and microalgae, with macroalgae being referred to as seaweeds, which are multicellular large-size algae that can grow up to 65 m. However, microalgae are microscopic, single-celled organisms with small size, from 1 to 900 μm. Microalgae are composed of eukaryotic organisms and prokaryotic cyanobacteria (blue-green algae) that have found widespread application as a biological source across a variety of industries, including the agriculture, food, pharmaceutical, and biofuel (Khan et al. 2018; Kusvuran and Kusvuran 2019; Renuka et al. 2018). In recent years, microalgae have become a sustainable agricultural product due to increasing the availability of nutrients, enhancing plant growth and crop yields, and maintaining the organic carbon and fertility of soil by boosting microbial activity in the soil. The ability of the photoautotrophic microalgae to produce high-value compounds (like pigments, polyunsaturated fatty acids, and vitamins), alternative energy sources and natural processes for environmental protection (such as CO2 mitigation, biofuel production, and wastewater treatment) has led to a large market demand (Prasanna et al. 2016a; Renuka et al. 2018; Touloupakis et al. 2021). Microalgae are potential components of products that are biologically active metabolites such as biofertilizers, biostimulants, and biopesticides, which can be used in crop production, protection, and soil improvement (Gonçalves 2021; Marks et al. 2019; Pathak et al. 2018; Plaza et al. 2018). The most common species of algae include Spirulina, Chlorella, Nostoc spp., Dunaliella, Scenedesmus, Isochrysis, Tetraselmis, Skeletonema, Pavlova, Chaetoceros, Phaeodactylum, Nitzschia, and Thalassiosira (Beal et al. 2018; Han et al. 2019).
Mechanisms of microalga to promote plant growth and development
Several studies demonstrate that microalgae containing products can stimulate plant growth and yield either in a single (Table 1) or a consortium with bacteria (Table 2); and have the potential to reduce synthetic fertilizer and can defend against plant pathogens. These are due to a large variety of bioactive compounds producing excellent sources of chemicals such as phytohormones, carotenoids, phycobilins and amino acids. Microalgae enhance crop productivity by promoting plant growth, nutrient availability, and pathogens biocontrol (Michalak and Chojnacka 2015b; Stirk et al. 2013). These products have diverse functional characteristics in crop production that promote an improvement in soil quality, nutrient uptake, enhancing crop performance, tolerance to biotic and abiotic stress conditions, and plant growth stimulation (Gonçalves 2021; Kusvuran and Kusvuran 2019; Renuka et al. 2018). This section describes the details of the potential development of the effect of biofertilizers, biostimulants, and biopesticides on agricultural production.
Biofertilizers
Biofertilizers are environmentally friendly containing living microbes or natural materials that improve soil fertility, crop development, and productivity by colonizing the plant’s rhizosphere and increasing the plant ability to absorb nitrogen, phosphorus, potassium, and minerals when applied to soil, plant, or seed (Mahanty et al. 2017; Ronga et al. 2019). Considerable research studies on biofertilizers have demonstrated their ability to supply the required nutrients to the crop in enough amounts that result in the improvement of crop growth and yield. Biofertilizers are living microbes that increase crop productivity by mobilizing or increasing nutrient availability in soil, in an economically feasible and environmentally friendly manner (Singh et al. 2011), and they are an substitute to chemical fertilizers. Biofertilizers are cost-effective; they minimize the side effect of environmental stress to a great extent and enhances soil fertility (Singh et al. 2011). It was testified that the application of biofertilizers improve crop yield by about 10–40% by increasing the contents of amino acids, proteins, nitrogen fixation, and vitamins (Bhardwaj et al. 2014; Prasanna et al. 2017). When microalgae were utilized as a source of biofertilizer, several research found a correlation between increased crop yields, increased nutrient uptake, and increased biomass accumulation (Hajnal-Jafari et al. 2020; Ronga et al. 2019; Shaaban 2001).
Nitrogen fixation
Soil serves as a medium for plant growth and is a crucial resource that must be continually resupplied with nutrients. Among the different features of biofertilizers, formulations based on oxygenic photosynthesis, including cyanobacteria and eukaryotic microalgae, are of increasing benefit in nutrient cycling, crop productivity, soil fertility and reducing chemical fertilizer application (G et al., 2016; Li et al. 2017). Certain cyanobacteria (free-living blue-green algae) can efficiently transform atmospheric nitrogen (N2) into organic nitrogen forms, which is one of the vital nutrients for plant growth (Dey et al. 2017; Gonçalves 2021; Renuka et al. 2018). Cyanobacteria has specialized cells known as heterocysts, that can fix atmospheric nitrogen and, as a result, are able to meet the needs of soil macro and micro fauna as well as flora and plants (Babu et al. 2015; Karthikeyan et al. 2007). Several researchers have investigated that inoculation with cyanobacteria proved to boost the yield and microbial activity by 5-25%, enhance plant growth, and seed germination in a wide variety of cereal and vegetable crops (Dey et al. 2017; (Prasanna et al. 2016b, 2017); can contribute to savings of 25-50% on chemical nitrogen fertilizers (G et al., 2016; Nain et al. 2010; Prasanna et al. 2016a). Due to their abundance in soil and their ability to fix atmospheric N, cyanobacteria like Nostoc and Anabaena strains are frequently used as biofertilizers (Renuka et al. 2018).
Leaching of biologically fixed N may be an environmental hazard, but the extent may be minimal compared to leaching caused by synthetic fertilizers. Research reports revealed that only 7% of total nitrogen is leached away when microalgae are applied to soil, whereas 50% of total nitrogen is leached when synthetic fertilizer is applied (Jimenez et al. 2020). Exopolysaccharide-producing cyanobacteria generate biological soil crusts and are also said to immobilize access to nitrogen (Mager and Thomas 2011), which inhabit nitrogen from leaching out of the soil. Generally, cyanobacteria inoculation improves the yield, microbial activity by 5-25%, and provides 25-40 kg N/ha, which results in substantial fertilizer cost savings and reduces environmental pollution load by preventing nitrogen leaching from the soil. Microalgae fertilization increased plant growth rate (shoot + root) by 32% in tomato plants, paralleling an increase in chlorophyll-a content (Jimenez et al. 2020); and it may also enhanced yield and microbial activity by 12–25% (Prasanna et al. 2014).
Improve soil structure
As a result of intensive agricultural methods, agricultural land is continuously degraded. Soil erosion, tilling and using heavy equipment too often can affects the soil’s structure, water holding capacity, fertility, nutrients movement, and productivity of agricultural soil. For agriculture to be sustainable, it is important to keep the soil’s organic matter and structure at the appropriate levels. Numerous varieties of green algae and cyanobacteria that are capable of producing extracellular polymeric substances (EPS) and releasing them into the environment (Xiao and Zheng 2016). The extracellular polymeric substances (EPS) have adhesive properties that contribute to enhancing soil organic carbon, aggregation of soil particles, enhancing soil structure, and preventing soil erosion to a large degree (Weiss et al. 2012; Xiao and Zheng 2016). The study found that inoculating cyanobacteria in soil resulted in the formation of organo-mineral soil aggregates composed of filaments and EPS, which increased aggregate stability six weeks after inoculation compared to the uninoculated control (Malam Issa et al. 2007).
In desert and semiarid soils, which are often highly compacted, low in fertility, saline or sodic, poorly aerated, and retain less water, microalgae make physio-chemical contributions to the health of the soil by supporting to form and stabilize soil aggregates, which increase pore space and continuity (Nichols et al. 2020). This enhances aeration, nutrient cycling, seed germination, water holding capacity, and water infiltration. Following the application of cyanobacteria (Nostoc and Anabaena) to a loam, silty clay loam, and sandy loam, there was an increase in soil aggregation of 85%, 130%, and 160%, respectively (Kaushik 2014). Hence, using algal biomass as a biofertilizer could improve the soil’s structure, water-holding capacity, and soil aeration.
Biostimulants
Plant biostimulants are derived from microorganisms or organic substances, when applied to the plant in a small quantity, they increase nutrient use efficiency, promotes plant growth, resistance to abiotic and biotic stress, and quality traits, regardless of their essential nutrient content for plant (García-Sánchez et al. 2022). Algae, both macroalgae (seaweeds) and microalgae, have long been viewed as a potentially profitable commercial prospect in the field agronomy and agro-industries due to their high concentrations of plant biostimulants (Kapoore et al. 2021). Hence, microalgal extracts are becoming promising natural resources for plant biostimulation (Romanenko et al. 2016). It is essential to keep in mind that biostimulants are not the same thing as biofertilizers because they do not directly supply the crops with the nutrients that they require; rather, they enhance the uptake of the nutrient by altering the rhizosphere and the metabolic processes of the plant (Drobek et al. 2019).
The biostimulatory effect of microalgae-based biostimulants under normal and stress situation can modulate microbial community inhabiting in the phyllosphere and rhizosphere areas of plants (Ranjan et al. 2016; Renuka et al. 2018). Recent experimental studies of biostimulatory microalga extracts have been shown to improve vegetative growth, absorption and distribution of nutrients, biomass and yield, resilience to biotic and abiotic stress, and water uptake in many crops under open and greenhouse settings (El Arroussi et al. 2018; Garcia-Gonzalez and Sommerfeld 2016; García-Sánchez et al. 2022; Prasanna et al. 2017). Biostimulants can also alter root formation, which influences plant health, nutritional composition, and growth by improving water and nutrient uptake (Garcia-Gonzalez and Sommerfeld 2016). The biostimulant activity of cyanobacteria, Arthrospira platensis, root and foliar applications on papaya has been tested. After integrating the findings into a surface model for plant height, stem diameter, leaf number, and leaf area, it was determined that a root treatment of 1.08% (w/v) was ideal for papaya seedling biomass production, whereas foliar spraying had no effect (Guedes et al. 2018). When the cyanobacteria (Nostoc calcicole or Anabaena vaginicola) were sprayed on the leaves of tomato, squash, and cucumber plants, compared to the controls, there were substantial increases in fresh weigh, dry weight, hight, root length, and leaf number (Shariatmadari et al. 2013). Therefore, algal biomass can be applied directly to plant leaves or roots to boost plant growth and yield. The identified potential algal biostimulant metabolites include phytohormones, humic substance, polysaccharides, amino acid, vitamins etc.
Phytohormones
The growth and development of plants are influenced to a significant level by phytohormones. In agriculture, the practice of exogenously supplementing plants with plant hormones (either natural or synthetic) has been a common method for increasing crop production and productivity (Aliyu et al. 2011). Extract of microalgal may contain phytohormones like auxin, cytokinins, ethylene, abscisic acid (ABA), and gibberellins, which can be used as biostimulant in agriculture (Stirk et al. 2002; Tarakhovskaya et al. 2007). New evidence reveals that phytohormones in microalgae have similar regulatory actions to those found in higher plants, but their precise role in these organisms remains unclear (Lu and Xu 2015). The two dominant kinds of auxin such as indole-3-acetic acid (IAA) and indole-3-butyric acid (IBA) are regulating growth and development including cell division and expansion (Hashtroudi et al. 2012). Cytokinins influences many traits of plant growth and physiology such as seed germination, shoot, and root development, and leaf senescence (Ha et al. 2012). While gibberellins play an important role mostly involves in elongation and expansion of the cell (Salazar-Cerezo et al. 2018). Ethylene is a gaseous phytohormone that plays an important role in physiological activities of plant, like growth and development, as well as resistance to biotic and abiotic stressor (Pierik et al. 2006). Abscisic acid is important in regulating several biological processes such as stomatal closure, seed maturation and improves resistance to temperature stress (Sagar and Singh 2019).
The phytohormones (such as auxin and gibberellins) are found in Chlorella kessleriwhen extact, when applied to Vicia faba, it increased leaf area, seedling growth parameters, germination, pigment content, and sodium and potassium accumulation in roots and shoots (El-Naggar et al. 2005). (Hussain and Hasnain 2011) investigated the efficacy of hormone-secreting cyanobacterial strains (cytokinin and auxin) in boosting growth both in axenic and natural environments. As a result, an approach to agronomic techniques that uses prospective phytohormone-excreting cyanobacterial strains as a biostimulant would be an environmentally acceptable way to stimulate plant development. However, research on the evaluation of algal hormone application at a field scale is limited, so this area needs further research.
Amino acids and protein hydrolysates
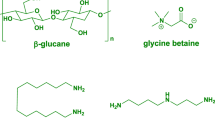
Amino acids and protein hydrolysates (PHs), together constitute a significant portion of the category within the broader field of plant biostimulants and find widespread application within environmentally responsible agriculture practices (Bulgari et al. 2019). Amino acids contain a minor amount of lipids, phytohormones, polysaccharides, and elements that are both macro and micronutrients, as well as protein hydrolysates, which may also consist primarily of short peptides (polypeptides and oligopeptides) (Calvo et al. 2014; Kapoore et al. 2021). The overall concentration of free amino acid and peptides (including arginine, alanine, proline, glycine, glutamate, valine, leucine and glutamine, among others) can range from 2 to 18% (w/w) to 1 to 85% (w/w), respectively (Calvo et al. 2014). Glycine betaine and proline are known to help to the mobility and uptake of micronutrients, as well as mitigation of environmental stress and antioxidant activity including heat, cold, drought, salinity, oxidative, and heavy metals through chelating actions (du Jardin 2015; Paul et al. 2019). Bioactive peptides, on the other hand, function in plants similarly to auxin and gibberellin, which improves overall plant growth and productivity (Colla et al. 2017). Certain microalgal strains contain more than 40% dry weight amino acid, including C. saccharophila, Chlorella sp., A. maxima, and A. platensis, and which contains 42.4%, 44.3%, 44.9%, and 46.8% respectively (Hempel et al. 2012), making them appropriate for biostimulants products.
Polysaccharides
The polysaccharides found in microalgae have the potential to be used as a bioresource in agriculture, both for the protection and improvement of crops. Polysaccharides are involved in many plants’ metabolic pathways and can act as biostimulants to increase crop quality and protect against biotic and abiotic challenges (Rachidi et al. 2020).
The highest concentration of polysaccharides found in the microalgae (Porphyridium cruentum and Chaetoceros gracilis species) and Dunaliella salina at the range of 40–57% and 199.8%, respectively (Kapoore et al. 2021). Studies shows that application of 1 mg/mL microalgae polysaccharides from D. Salina, Porphorydium sp., and A. platensis on tomato plants significantly improved the shoot dry weight, shoot length, and nodes number, shoot dry weight, and shoot length by 46.6%, 25.26%, and 75%, respectively, compared to control (Rachidi et al. 2020). Further example, foliar application of microalgae polysaccharide extract from A. platensis at concentration of 3 g/L (w/v) increased plant growth and development of leaf area size by 57% and 100%, size of nodes by 33% and 57%, and root weight by 67% and 230%, for pepper and tomato plants, respectively (Elarroussia et al. 2016).
Humic substances
Humic compounds are generally included in categories of biostimulants; but their algal origin has not been well studied (Kapoore et al. 2021). Humic substances are naturally occurring components that make up around 60% of the organic matter in the soil. They can be produced either by breaking down microbial, animal, and plant residues or through the metabolism of soil microbes that use these materials (du Jardin 2015). Humic substance are divided into humic acids, fulvic acids, and humins according to their molecular weights and solubilities (du Jardin 2015). When applied to crops, humic material in digestate may bind to algal cells and act as a biostimulant. On the other hand, the biostimulant effect of humic compounds isolated from agro-industrial waste on S. quadricauda and C. vulgaris showed that there was a significant increase in chlorophyll, lipids, carbohydrate content and biomass (Puglisi et al. 2018).
Biopesticides
Utilizing chemical pesticides for the control of pests and pathogens in agricultural activities poses a threat to the sustainability of agroecosystems. Sustainable crop protection against pests and pathogens by using modern technologies allows to keep plants healthy and achieve stable high yield. Biopesticides are naturally known substances that are obtained from microorganisms, plants, or animals, primarily for insect and plant disease control. These substances or materials, including antioxidant, antimicrobial, antifungal, or antiviral properties, help crop growth by defending plants against harmful effect of pathogens. Some bacteria and fungi are among the most often discovered organisms that can be used for biocontrol (Spadaro and Gullino 2005). Microalgae, particularly cyanobacteria, have gained attention in recent decades as possible biocontrol agents against pests and diseases (Hernández-Carlos and Gamboa-Angulo 2011).
Phytohormones are essential for the controlling a growth and development of plants as well as its defense against biotic and abiotic stress via interacting among them (Checker et al. 2018). Many investigations were carried out to evaluate microalgae, as potential biocontrol substances that have demonstrated antagonistic effects against many plant pathogens, like nematodes, fungi, and bacteria, mainly due to the produce hydrolytic enzymes and biocidal compounds, like benzoic acid and majusculonic acid (Chaudhary et al. 2012; Gupta et al. 2013; Renuka et al. 2018). These antimicrobial substances can suppress microorganisms by either disrupting the cytoplasmic membrane or inhibiting protein synthesis (Swain et al. 2017).
Roles in crop stress tolerance
Crops are stationary organisms that are constantly stressed by biotic and abiotic causes. The influence of abiotic factors such as low and/or high temperatures, salinity, drought, alkalinity, and other factors, can lead to low productivity and yield quality because of the reductions in respiration, photosynthesis, and protein synthesis (Dwivedi et al. 2015; Sharma et al. 2012). The biotic factors caused by pathogenic bacteria, viruses, fungi, weeds, etc., affect the plant’s host cell and modify the plant’s genetic code, which takes to leads to the death of the plant (Suzuki et al. 2014). Research reports showed that around 30% of world’s crop production is lost because of abiotic stress (Goswami et al. 2016). One possible way to reducing the effects of abiotic stress is the application of microalga which can play a substantial role in minimizing this loss by induced systemic tolerance (IST), which is stimulating various types of biochemical and physiological tolerance systems in plants (Sharma et al. 2012). Microbial biostimulants have been used in a sustainable approach for enhancing plant growth, productivity, and nutrition, even in the climate-stress situation (Fadiji et al. 2022). Some bacteria species (Azospirillum brasilense, Pseudomonas sp., and Bacillus lentus) have been used alone or in microbial associations that could minimize drought stress impact in crops (Sangiorgio et al. 2020).
One of the promising example observed for mitigating salt stress during seed germination process of bell pepper is application of microalgea extracts from Phaeodactylum spp and Dunaliella spp (Guzmán-Murillo et al. 2013). According to (Abd El-Baky et al. 2010), it has been advised that the addition of microalgal extracts to wheat (Triticum aestivum L.) that are irrigated with seawater could be beneficial in increasing wheat’s resistance to salty environments. Similarly, Chlorella spp. and Spirulina spp. boosted the antioxidant capacity and protein content of whole grains, as well as improved wheat’s resistance to salinity (Abd El-Baky et al. 2010).
Impact of microalga-bacteria interaction on crop production
In nature, not only plant-microbe interactions, but also microbe-microbe associations are vital assemblages affecting plant growth, development, health, and productivity. In natural ecosystems or industrial processes, microalgae and bacteria live together, demonstrating both beneficial and harmful relationships (Unnithan et al. 2014). Under industry settings, bacteria are considered pollutants in algae research, but most recent investigation have demonstrated that most algal symbionts not only stimulate algal growth but also extend benefits in downstream processing (Lian et al. 2018). There have been several studies that have brought attention to the potential of bacteria include nitrogen fixation (Azotobacter vinelandii, Azospirillum brasilens, Rhizobium etli, and Mesorhizobium loti), phosphate solubilization (Azospirillum spp., Pseudomonas spp., Arbuscular mycorrhizal fungi, and Bacillus spp.), cellulolytic activity (Bacillus spp, Aspergillus spp., Penicillium spp., and Trichoderma spp.), and the production of siderophores are recognized as plant growth promoting rhizobacteria (PGPR) (Bhattacharyya and Jha 2012; (Meena et al. 2020a; Woo and Pepe 2018). Plant growth promoting bacteria (PGPB) are soil bacteria that can promote plant growth, suppress pathogens, promote nutrient availability to plants, and increase abiotic stress resistance mechanisms (Kumar et al. 2017).
A combined application of the microalgae-bacteria can improve plant growth and control plant disease, which is much more efficient than a sole application (Spadaro and Gullino 2005). This is due to the microbes in the consortia/combination system having the capability to improve plant growth and development and/or control pathogens by different mechanisms. (Trivedi et al. 2017) reported that the beneficial microbiome could form relations with other microbiomes, reproduce highly structured systems in the rhizosphere soils, and may have a greater likelihood to assistances the host than a single culture. One of the promising examples showed that the combined application of the cyanobacterial (Nostoc muscorum and Anabaena flos-aquae) and bacterial suspensions (Azotobacter brasilense and Azotobacter chroococcum) was substantially improved germination rate of Lupinus termis seeds by 53.13%, 211.48%, 129.04%, and 104.1%, respectively, when compared to control (Tantawy and Atef 2010).
In interaction systems, microalgae and bacteria can be symbiotic to competition (mutualism to antagonism). The relationships between microalgal and bacterial communities are based on signal transduction, gene transfer, and nutrient exchange (Aditya et al. 2022). Microalgae and bacteria engage in a dynamic exchange of carbon, energy, and essential molecules, which is seen in Fig. 1. In a synergetic association, microalgae stimulate bacteria grow by supplying oxygen (through photosynthesis) and dissolved organic matter, such as calcium carbonate and organic carbon, that become accessible to bacteria (Cooper and Smith 2015). On the other side, the bacteria produce carbon dioxide (CO2) and remineralize nitrogen, phosphorus, and sulphur to maintain further microalgae growth (Yao et al. 2019). Moreover, bacteria supply vitamin B as organic cofactors (Yao et al. 2019), amino acids (Palacios et al. 2016), and hormones (De-Bashan et al. 2008), which become bioavailable for microalgae.

Mechanism of a possible symbiotic interaction of microalgae and bacteria and their potential role in the agricultural production. In normal interactions, microalgae exude oxygen (O2), organic carbon, and calcium carbonate (CaCO3), which bacteria can use. In exchange, the bacteria remineralize nitrogen (N) and phosphorus (P), growth promoter and produces carbon dioxide (CO2) to assist the growth of the microalgae. In specialized interactions, the bacteria supply B vitamins as organic cofactors or create siderophores to bind iron, making it bioavailable to the microalgae. The interaction of microalgae and bacteria offers a unique potential for eco-friendly products such as biofertilizers, biostimulants, biopesticides, and soil conditioners, which reduce reliance on agrochemicals and sustainably increase crop production and productivity
In a pot experiment with rice varieties, the combined treatment of cyanobacteria strains (Anabaena sp., Anabaena oscillarioides, and Anabaena laxa) and bacteria strains (Brevundimonas sp., Ochrobactrum sp., and Providencia sp.,) was examined. In this trial, the authors evaluated that a significantly increase in the growth, grain yield by 19.02%, nitrogen fixing potential of rice, and improving soil fertility by nitrogen savings of 40-80 kg/ha, especially with Ochrobacterium and Anabaena species (Prasanna et al. 2012).
During the spring and summer, the combined biostimulant properties of freshwater algae (Chlorella vulgaris) and bacteria (Azospirillum sp., Azotobacter sp., Herbaspirillum sp., Bacillus licheniformis, and Bacillus megatherium) significantly influenced the weight of the romaine and leaf lettuce crops (Kopta et al. 2018). Moreover, the research also suggested that the photosynthetic substances produced by algae, like carotenoids, could boost the quality and productivity of crops and give support during times of stress. Similarly, the positive result was observed by combined application of algae and bacteria to crops promoted growth, productivity, and quality in common bean, maize, and onion (Gavilanes et al. 2020; Geries and Elsadany 2021; Horácio et al. 2020). This consortium reduces synthetic pesticide use, making it essential to sustainable agriculture and food safety (Niu et al. 2020). A suitable microalgae-bacteria consortium is necessary to boost the potential of strains to enhance growth and development and to inhibit pathogen attack (Yanti et al., 2021). Considering these factors, it is feasible to assume that microalgae-bacteria consortium can be successful at increasing soil microbial activities, crop productivity, and plant disease resistance. However, more investigation needs on the molecular mechanisms underlaying the influence of microalgae and bacteria association to help plants development and disease prevention, so that they can be used in agriculture in a safer and more widespread way.
Table 2 highlighted valuable practical reports on the microalgae–bacteria combined treatments in different crop cultivation. Some studies revealed a promise of the microalgae-bacteria association, examined in the field and greenhouse conditions that can promote seedling growth, germination, and biomass in plants (Kang et al. 2021). An association between microalgae and bacteria can increase plant growth and development by the production of phytohormones (like auxin, cytokinin, and so on) and polysaccharides; it can also stimulate nutrient uptake by regulating a variety of biochemical and physiological processes; and it can reduce the risk of pathogen infection (Fuentes et al. 2016a; (Michalak and Chojnacka 2015a).
Challenges of microalgae-bacteria interactions
The field of microbial consortia is still in its initial stages, and there are still a lot of problems to solve when it comes to how cells communicate to each other and how to make systems that are stable and easy to control. The main challenge of the association of microbes, soil, and climate in an agricultural setting is to understand their specific structural function activities on plants. A lot of study has demonstrated a promising result in the greenhouse trials, but it fails to confirm in field trials. The microalgae-bacteria interaction can also be affected by environmental factors like pH, temperature, and light intensity (Quijano et al. 2017). It is difficult to depict their ecosystem-wide processes such as metabolic pathways and nutrient cycling because the majority of heterotrophic bacteria and photosynthetic microalgae have not yet been cultured (Zhang et al. 2020). Furthermore, the amount of nutrient present in the growing media has a major impact on the dynamic between microalgae and bacteria (Liu et al. 2012).
The growth phase is another significant aspect that plays a role in the interactions that take place between microalgae and bacteria. Expensive harvesting of biomass, insufficient biomass production, and extraction technologies that need a lot of energy are also main constraints that are preventing their large-scale development. Subsequently, it is challenging to distinguish the individual metabolites that microalgae and bacteria produce in a consortium due to the complexity of their interactions, which are either naturally occurring or artificially engineered for a specific goal (Zhang et al. 2020). The result of microalgae-bacteria interactions is often varied in different studies under different climatic, or soil conditions, which is the main problem in the implementation of the technology. Species and environmental circumstances are the primary variables detrimental to the microalgae-bacteria relationship (Lauritano et al. 2020; Mujtaba and Lee 2016). Which highlighted the importance of choosing a suitable combination of microalgae strains versus bacteria strains for the efficient application in the agricultural production. To overcome these restrictions, researchers have focused on enhancing microalgal-bacterial consortia, which offer several economic, energy, and environmental benefits due to their mutual interactions.
Conclusion and future perspectives
As eco-friendly tools for sustainable agriculture, microalgae and bacteria have drawn a lot of attention as biofertilizers, biostimulants, and biopesticides. Numerous investigations have indicated that microalgae and bacteria association synergistically impact each other’s metabolism and physiology. Combinations of microalgae and bacteria that have been carefully engineered are anticipated to significantly improve crop productivity, efficiency, and agricultural sustainability. Different combinations/association of microalgae and bacteria will probably exhibit district activities, so it is necessary to selecting the optimal consortia and investigating all relevant factors is crucial for maximizing the benefits of algae-bacterial interactions. Generally, based on different studies there is an agreement on the potential advantages of the microalgae-bacteria association on the crop, but still, there is insufficient scientific evidence to support this interaction in a wide range of crop species and environments. To overcome such limitations, researchers should focus on the basic effects of microalgae–bacteria co-culture/combination on different variety of crop species and environmental conditions to understand the biofertilizer, biogeochemical cycling, biostimulant, and biopesticides effects, which are potentially used for sustainable agriculture practices. Hence, researchers should focus on the following area to understand their associations: (i) sorting out the different compatible strains of microalgae-bacteria to select ideal combinations; (ii) application methods (e.g., timing, soil, inoculation, or foliar) and rate of application; (iii) identifying potentially synergistic microalgal-bacterial interaction providing supportive traits, like the nitrogen fixation, production of phytohormones, and biocontrol; and iv) refine and characterize plant genotype-environment-microbiome-management relationships. Meeting these aims should speed up our ability to design and build successful associations of microalgae-bacteria with specific biofertilizers, biostimulants, and biopesticides properties, which will benefit both consumers and food producers.
References
Abd El-Baky HH, El-Baz FK, Baroty E, G. S (2010) Enhancing antioxidant availability in wheat grains from plants grown under seawater stress in response to microalgae extract treatments. J Sci Food Agric 90:299–303
Aditya L, Mahlia TMI, Nguyen LN, Vu HP, Nghiem LD (2022) Microalgae-bacteria consortium for wastewater treatment and biomass production. Sci Total Environ 838:155871
Aliyu OM, Adeigbe OO, Awopetu JA (2011) Foliar application of the exogenous plant hormones at pre-blooming stage improves flowering and fruiting in cashew (Anacardium occidentale L). J Crop Sci Biotechnol 14:143–150
Alvarez AL, Weyers SL, Goemann HM, Peyton BM, Gardner RD (2021) Microalgae, soil and plants: a critical review of microalgae as renewable resources for agriculture. Algal Res 54:102200
Babu S, Prasanna R, Bidyarani N, Singh R (2015) Analysing the colonisation of inoculated cyanobacteria in wheat plants using biochemical and molecular tools. J Appl Phycol 27:327–338
Beal CM, Gerber LN, Thongrod S, Phromkunthong W, Kiron V, Granados J, Archibald I, Greene CH, Huntley ME (2018) Marine microalgae commercial production improves sustainability of global fisheries and aquaculture. Sci Rep 8:15064
Bhardwaj D, Ansari MW, Sahoo RK, Tuteja NK (2014) Biofertilizers function as key player in sustainable agriculture by improving soil fertility, plant tolerance and crop productivity. Microb Cell Fact 13:66–66
Bhattacharyya PN, Jha DK (2012) Plant growth-promoting rhizobacteria (PGPR): emergence in agriculture. World J Microbiol Biotechnol 28:1327–1350
Bulgari R, Franzoni G, Ferrante A (2019) Biostimulants Application in Horticultural crops under abiotic stress conditions. Agronomy 9:306
Calvo P, Nelson L, Kloepper JW (2014) Agricultural uses of plant biostimulants. Plant Soil 383:3–41
Chandini R, Kumar R, Kumar, Prakash O (2019) In: Sharma P (ed) The impact of Chemical fertilizers on our environment and Ecosystem.,“ in Research Trends in Environmental Sciences. AkiNik Publications, New Delhi
Chaudhary V, Prasanna R, Nain L, Dubey SC, Gupta V, Singh R, Jaggi S, Bhatnagar AK (2012) Bioefficacy of novel cyanobacteria-amended formulations in suppressing damping off disease in tomato seedlings. World J Microbiol Biotechnol 28:3301–3310
Checker VG, Kushwaha HR, Kumari P, Yadav S (2018) Role of Phytohormones in Plant Defense: signaling and Cross Talk. In: Singh A, Singh IK (eds) Molecular Aspects of Plant-Pathogen Interaction. Springer Singapore, Singapore, pp 159–184
Chiaiese P, Corrado G, Colla G, Kyriacou MC, Rouphael Y (2018) Renewable sources of Plant Biostimulation: Microalgae as a sustainable means to improve crop performance. Front Plant Sci 9:1782
Colla G, Hoagland L, Ruzzi M, Cardarelli M, Bonini P, Canaguier R, Rouphael Y (2017) Biostimulant Action of protein hydrolysates: unraveling their Effects on Plant Physiology and Microbiome. Front Plant Sci 8:2202
Cooper MB, Smith AG (2015) Exploring mutualistic interactions between microalgae and bacteria in the omics age. Curr Opin Plant Biol 26:147–153
De-Bashan LE, Antoun H, Bashan Y (2008) Involvement of Indole-3-Acetic acid produced by the growth-promoting bacterium Azospirillum Spp. In promoting growth of Chlorella Vulgaris1. J Phycol 44:938–947
Dey SK, Chakrabarti B, Prasanna R, Pratap D, Singh SD, Purakayastha TJ, Pathak H (2017) Elevated carbon dioxide level along with phosphorus application and cyanobacterial inoculation enhances nitrogen fixation and uptake in cowpea crop. Arch Agron Soil Sci 63:1927–1937
Dineshkumar R, Subramanian J, Gopalsamy J, Jayasingam P, Arumugam A, Kannadasan S, Sampathkumar P (2017) The impact of using Microalgae as Biofertilizer in Maize (Zea mays L). Waste Biomass Valoriz 10:1101–1110
Drobek M, Frąc M, Cybulska J (2019) Plant biostimulants: importance of the Quality and Yield of Horticultural crops and the improvement of plant tolerance to abiotic Stress—A review. Agronomy 9:335
du Jardin P (2015) Plant biostimulants: definition, concept, main categories and regulation. Sci Hort 196:3–14
Dwivedi SL, Sahrawat KL, Upadhyaya HD, Mengoni A, Galardini M, Bazzicalupo M, Biondi EG, Hungria M, Kaschuk G, Blair MW, Ortiz R (2015) Chapter One - Advances in Host Plant and Rhizobium Genomics to Enhance Symbiotic Nitrogen Fixation in Grain Legumes. In “Advances in Agronomy” (D. L. Sparks, ed.), Vol. 129, pp. 1-116. Academic Press
El Arroussi H, Benhima R, Elbaouchi A, Sijilmassi B, El Mernissi N, Aafsar A, Meftah-Kadmiri I, Bendaou N, Smouni A (2018) Dunaliella salina exopolysaccharides: a promising biostimulant for salt stress tolerance in tomato (Solanum lycopersicum). J Appl Phycol 30:2929–2941
El-Naggar AH, Osman MEHE-S, Gheda SF (2005) Influence of the aqueous extracts of Ulva lactuca and Chlorella kessleri on growth and yield of Vicia faba. Algol Studies/Archiv für Hydrobiol Supplement Volumes 116:213–229
Elarroussia H, Elmernissia N, Benhimaa R, Kadmiria E, Bendaou IM, Smouni N, A., and, Wahbya I (2016) Microalgae polysaccharides a promising plant growth biostimulant. J Algal Biomass Utln 7:55–63
Fadiji AE, Babalola OO, Santoyo G, Perazzolli M (2022) The potential role of Microbial Biostimulants in the amelioration of Climate Change-Associated Abiotic stresses on crops. Front Microbiol 12
Fuentes JL, Garbayo I, Cuaresma M, Montero Z, Gonzalez-Del-Valle M, Vilchez C (2016a) Impact of Microalgae-Bacteria Interactions on the Production of Algal Biomass and Associated Compounds. Mar Drugs 14
Fuentes JL, Garbayo I, Cuaresma M, Montero Z, González-del-Valle M, Vílchez C (2016b) Impact of Microalgae-Bacteria interactions on the production of Algal Biomass and Associated Compounds. Mar Drugs 14:100
G T, D.J, B., and M.S, R (2016) Selected microbial consortia developed for chilly reduces application of chemical fertilizers by 50% under field conditions. Sci Hort 198:27–35
Garcia-Gonzalez J, Sommerfeld M (2016) Biofertilizer and biostimulant properties of the microalga Acutodesmus dimorphus. J Appl Phycol 28:1051–1061
García-Sánchez F, Simón-Grao S, Navarro-Pérez V, Alfosea-Simón M (2022) Scientific advances in Biostimulation reported in the 5th Biostimulant World Congress. Horticulturae 8:665
Gavilanes FZ, Souza Andrade D, Zucareli C, Horácio EH, Yunes S, Barbosa J, Alves AP, Cruzatty LAR, Maddela LG, N. R., and, Guimarães M (2020) Co-inoculation of Anabaena cylindrica with Azospirillum brasilense increases grain yield of maize hybrids. Rhizosphere 15:100224 d. F
Geries LSM, Elsadany AY (2021) Maximizing growth and productivity of onion (Allium cepa L.) by Spirulina platensis extract and nitrogen-fixing endophyte Pseudomonas stutzeri. Arch Microbiol 203:169–181
Gonçalves AL (2021) The Use of Microalgae and Cyanobacteria in the improvement of Agricultural Practices: a review on their Biofertilising, Biostimulating and Biopesticide Roles. Appl Sci 11:871
Goswami D, Thakker JN, Dhandhukia PC (2016) Portraying mechanics of plant growth promoting rhizobacteria (PGPR): a review. Cogent Food & Agriculture 2:1127500
Guedes WA, Araújo RHCR, Rocha JLA, Lima JFd, Dias GA, Oliveira, Á. M. F. d., Lima RF d., and, Oliveira LM (2018) Production of Papaya Seedlings Using Spirulina platensis as a Biostimulant Applied on Leaf and Root. Journal of Experimental Agriculture International 28, 1–9
Gupta V, Ratha SK, Sood A, Chaudhary V, Prasanna R (2013) New insights into the biodiversity and applications of cyanobacteria (blue-green algae)—Prospects and challenges. Algal Res 2:79–97
Guzmán-Murillo MA, Ascencio F, Larrinaga-Mayoral JA (2013) Germination and ROS detoxification in bell pepper (Capsicum annuum L.) under NaCl stress and treatment with microalgae extracts. Protoplasma 250:33–42
Ha S, Vankova R, Yamaguchi-Shinozaki K, Shinozaki K, Tran L-SP (2012) Cytokinins: metabolism and function in plant adaptation to environmental stresses. Trends Plant Sci 17:172–179
Hajnal-Jafari T, Seman V, Stamenov D, Đurić S (2020) Effect of Chlorella vulgaris on growth and photosynthetic pigment content in swiss chard (Beta vulgaris L. subsp. cicla). Pol J Microbiol 69:1–4
Han P, Lu Q, Fan L, Zhou W (2019) A review on the Use of Microalgae for sustainable aquaculture. Appl Sci 9:2377
Hashtroudi MS, Ghassempour A, Riahi H, Shariatmadari Z, Khanjir M (2012) Endogenous auxins in plant growth-promoting Cyanobacteria—Anabaena vaginicola and Nostoc calcicola. J Appl Phycol 25:379–386
Hempel N, Petrick I, Behrendt F (2012) Biomass productivity and productivity of fatty acids and amino acids of microalgae strains as key characteristics of suitability for biodiesel production. J Appl Phycol 24:1407–1418
Hernández-Carlos B, Gamboa-Angulo MM (2011) Metabolites from freshwater aquatic microalgae and fungi as potential natural pesticides. Phytochem Rev 10:261–286
Holajjer P, Kamra A, Gaur HS, Manjunath M (2013) Potential of cyanobacteria for biorational management of plant parasitic nematodes: a review. Crop Prot 53:147–151
Horácio EH, Zucareli C, Gavilanes FZ, Yunes JS, Sanzovo AW d. S., and, Andrade DS (2020) Co-inoculation of rhizobia, azospirilla and cyanobacteria for increasing common bean production. Semina: Ciências Agrárias, 2015–2028
Hussain A, Hasnain S (2011) Phytostimulation and biofertilization in wheat by cyanobacteria. J Ind Microbiol Biotechnol 38:85–92
Jimenez R, Markou G, Tayibi S, Barakat A, Chapsal C, Monlau F (2020) Production of Microalgal slow-release fertilizer by valorizing Liquid Agricultural Digestate: growth experiments with tomatoes. Appl Sci 10:3890
Kang Y, Kim M, Shim C, Bae S, Jang S (2021) Potential of algae-Bacteria synergistic Effects on Vegetable Production. Front Plant Sci 12:656662
Kapoore RV, Wood EE, Llewellyn CA (2021) Algae biostimulants: a critical look at microalgal biostimulants for sustainable agricultural practices. Biotechnol Adv 49:107754
Karthikeyan N, Prasanna R, Nain L, Kaushik BD (2007) Evaluating the potential of plant growth promoting cyanobacteria as inoculants for wheat. Eur J Soil Biol 43:23–30
Kaushik B (2014) Developments in cyanobacterial biofertilizer. In “Proc Indian Nat Sci Acad” 80:379–388
Khan MI, Shin JH, Kim JD (2018) The promising future of microalgae: current status, challenges, and optimization of a sustainable and renewable industry for biofuels, feed, and other products. Microb Cell Fact 17:36
Kopta T, PavlÍKovÁ M, SĘKara A, Pokluda R, MarŠÁLek B (2018) Effect of bacterial-algal Biostimulant on the yield and Internal Quality of Lettuce (Lactuca sativa L.) produced for spring and summer crop. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 46:615–621
Kumar H, Dubey RC, Maheshwari DK (2017) Seed-coating fenugreek with Burkholderia rhizobacteria enhances yield in field trials and can combat Fusarium wilt. Rhizosphere 3:92–99
Kusvuran A, Kusvuran S (2019) Using of Microbial Fertilizer as Biostimulant alleviates damage from Drought stress in Guar (Cyamopsis Tetragonoloba (L.) Taub.) Seedlings. Int Lett Nat Sci 76:147–157
Lauritano C, Rizzo C, Giudice L, A., and, Saggiomo M (2020) Physiological and Molecular Responses to Main Environmental Stressors of Microalgae and Bacteria in Polar Marine Environments. Microorganisms 8, 1957
Li R, Tao R, Ling N, Chu G (2017) Chemical, organic and bio-fertilizer management practices effect on soil physicochemical property and antagonistic bacteria abundance of a cotton field: implications for soil biological quality. Soil Tillage Res 167:30–38
Lian J, Wijffels RH, Smidt H, Sipkema D (2018) The effect of the algal microbiome on industrial production of microalgae. Microb Biotechnol 11:806–818
Liu H, Zhou Y, Xiao W, Ji L, Cao X, Song C (2012) Shifting nutrient-mediated interactions between algae and bacteria in a microcosm: evidence from alkaline phosphatase assay. Microbiol Res 167:292–298
Lu Y, Xu J (2015) Phytohormones in microalgae: a new opportunity for microalgal biotechnology? Trends Plant Sci 20:273–282
Mager DM, Thomas AD (2011) Extracellular polysaccharides from cyanobacterial soil crusts: a review of their role in dryland soil processes. J Arid Environ 75:91–97
Mahanty T, Bhattacharjee S, Goswami M, Bhattacharyya P, Das B, Ghosh A, Tribedi P (2017) Biofertilizers: a potential approach for sustainable agriculture development. Environ Sci Pollut Res Int 24:3315–3335
Malam Issa O, Défarge C, Le Bissonnais Y, Marin B, Duval O, Bruand A, D’Acqui LP, Nordenberg S, Annerman M (2007) Effects of the inoculation of cyanobacteria on the microstructure and the structural stability of a tropical soil. Plant Soil 290:209–219
Marks EAN, Montero O, Rad C (2019) The biostimulating effects of viable microalgal cells applied to a calcareous soil: increases in bacterial biomass, phosphorus scavenging, and precipitation of carbonates. Sci Total Environ 692:784–790
Matsuo O, Zucareli C, Horácio EH, Alves LAR, Saab OJGA (2022) Co-inoculation of Anabaena cylindrica and Azospirillum brasilense during initial growth and chloroplast pigments of corn. Revista Brasileira de Engenharia Agricola e Ambiental 26:97–102
Meena M, Swapnil P, Divyanshu K, Kumar S, Harish, Tripathi YN, Zehra A, Marwal A, Upadhyay RS (2020a) PGPR-mediated induction of systemic resistance and physiochemical alterations in plants against the pathogens: current perspectives. J Basic Microbiol 60:828–861
Meena RS, Kumar S, Datta R, Lal R, Vijayakumar V, Brtnicky M, Sharma MP, Yadav GS, Jhariya MK, Jangir CK, Pathan SI, Dokulilova T, Pecina V, Marfo TD (2020b) Impact of Agrochemicals on Soil Microbiota and Management: A Review. Land 9, 34
Michalak I, Chojnacka K (2015a) Algae as production systems of bioactive compounds. Eng Life Sci 15
Michalak I, Chojnacka KJ (2015b) Algae as production systems of bioactive compounds. Eng Life Sci 15
Mujtaba G, Lee K (2016) Advanced Treatment of Wastewater using Symbiotic co-culture of microalgae and Bacteria. Appl Chem Eng 27:1–9
Naik K, Mishra S, Srichandan H, Singh PK, Sarangi PK (2019) Plant growth promoting microbes: potential link to sustainable agriculture and environment. Biocatal Agric Biotechnol 21:101326
Nain L, Rana A, Joshi M, Jadhav SD, Kumar D, Shivay YS, Paul S, Prasanna R (2010) Evaluation of synergistic effects of bacterial and cyanobacterial strains as biofertilizers for wheat. Plant Soil 331:217–230
Nichols K, Olson M, Ayers AD (2020) Microalgae as a beneficial soil amendment. Arizona: MyLand Compani LLC
Niu B, Wang W, Yuan Z, Sederoff RR, Sederoff H, Chiang VL, Borriss R (2020) Microbial interactions within multiple-strain Biological Control Agents Impact Soil-Borne Plant Disease. Front Microbiol 11
Ördög V, Stirk WA, Takács G, Pőthe P, Illés Á, Bojtor C, Széles A, Tóth B, van Staden J, Nagy J (2021) Plant biostimulating effects of the cyanobacterium Nostoc piscinale on maize (Zea mays L.) in field experiments. South Afr J Bot 140:153–160
Palacios OA, Gomez-Anduro G, Bashan Y, de-Bashan LE (2016) Tryptophan, thiamine and indole-3-acetic acid exchange between Chlorella sorokiniana and the plant growth-promoting bacterium Azospirillum brasilense. FEMS Microbiol Ecol 92:fiw077
Pathak J, Rajneesh, Maurya PK, Singh SP, Häder D-P, Sinha RP (2018) Cyanobacterial farming for Environment Friendly Sustainable Agriculture Practices: innovations and perspectives. Front Environ Sci 6
Paul K, Sorrentino M, Lucini L, Rouphael Y, Cardarelli M, Bonini P, Miras Moreno MB, Reynaud H, Canaguier R, Trtilek M, Panzarova K, Colla G (2019) A combined phenotypic and Metabolomic Approach for elucidating the Biostimulant Action of a plant-derived protein hydrolysate on Tomato grown under Limited Water availability. Front Plant Sci 10:493
Pierik R, Tholen D, Poorter H, Visser EJW, Voesenek LACJ (2006) The Janus face of ethylene: growth inhibition and stimulation. Trends Plant Sci 11:176–183
Plaza BM, Gómez-Serrano C, Acién-Fernández FG, Jimenez-Becker S (2018) Effect of microalgae hydrolysate foliar application (Arthrospira platensis and Scenedesmus sp.) on Petunia x hybrida growth. J Appl Phycol 30:2359–2365
Prasanna R, Joshi M, Rana A, Shivay YS, Nain L (2012) Influence of co-inoculation of bacteria-cyanobacteria on crop yield and C–N sequestration in soil under rice crop. World J Microbiol Biotechnol 28:1223–1235
Prasanna R, Chaudhary V, Gupta V, Babu S, Kumar A, Singh R, Shivay YS, Nain L (2013) Cyanobacteria mediated plant growth promotion and bioprotection against Fusarium wilt in tomato. Eur J Plant Pathol 136:337–353
Prasanna R, Triveni S, Bidyarani N, Babu S, Yadav K, Adak A, Khetarpal S, Pal M, Shivay YS, Saxena AK (2014) Evaluating the efficacy of cyanobacterial formulations and biofilmed inoculants for leguminous crops. Arch Agron Soil Sci 60:349–366
Prasanna R, Kanchan A, Kaur S, Ramakrishnan B, Ranjan K, Singh MC, Hasan M, Saxena AK, Shivay YS (2016a) Chrysanthemum Growth gains from beneficial microbial interactions and fertility improvements in Soil under protected cultivation. Hortic Plant J 2:229–239
Prasanna R, Kanchan A, Ramakrishnan B, Ranjan K, Venkatachalam S, Hossain F, Shivay YS, Krishnan P, Nain L (2016b) Cyanobacteria-based bioinoculants influence growth and yields by modulating the microbial communities favourably in the rhizospheres of maize hybrids. Eur J Soil Biol 75:15–23
Prasanna R, Ramakrishnan B, Simranjit K, Ranjan K, Kanchan A, Hossain F, Nain L (2017) Cyanobacterial and rhizobial inoculation modulates the plant physiological attributes and nodule microbial communities of chickpea. Arch Microbiol 199:1311–1323
Puglisi I, Barone V, Sidella S, Coppa M, Broccanello C, Gennari M, Baglieri A (2018) Biostimulant activity of humic-like substances from agro-industrial waste on Chlorella vulgaris and Scenedesmus quadricauda. Eur J Phycol 53:433–442
Quijano G, Arcila JS, Buitrón G (2017) Microalgal-bacterial aggregates: applications and perspectives for wastewater treatment. Biotechnol Adv 35:772–781
Rachidi F, Benhima R, Sbabou L, Arroussi E, H (2020) Microalgae polysaccharides bio-stimulating effect on tomato plants: growth and metabolic distribution. Biotechnol Rep 25:e00426
Ranjan K, Priya H, Ramakrishnan B, Prasanna R, Venkatachalam S, Thapa S, Tiwari R, Nain L, Singh R, Shivay YS (2016) Cyanobacterial inoculation modifies the rhizosphere microbiome of rice planted to a tropical alluvial soil. Appl Soil Ecol 108:195–203
Renuka N, Guldhe A, Prasanna R, Singh P, Bux F (2018) Microalgae as multi-functional options in modern agriculture: current trends, prospects and challenges. Biotechnol Adv 36:1255–1273
Romanenko KO, Kosakovskaya IV, Romanenko PO (2016) Phytohormones of Microalgae: Biological role and involvement in the regulation of physiological processes. 18:179–201
Ronga D, Biazzi E, Parati K, Carminati D, Carminati E, Tava A (2019) Microalgal Biostimulants and Biofertilisers in Crop Productions. Agronomy 9:192
Sagar S, Singh A (2019) Chapter 22 - abscisic acid, a principal Regulator of Plant Abiotic stress responses. In: Khan MIR, Reddy PS, Ferrante A, Khan NA (eds) Plant Signaling Molecules. Woodhead Publishing, pp 341–353
Salazar-Cerezo S, Martínez-Montiel N, García-Sánchez J, Pérez-y-Terrón R, Martínez-Contreras RD (2018) Gibberellin biosynthesis and metabolism: a convergent route for plants, fungi and bacteria. Microbiol Res 208:85–98
Sangiorgio D, Cellini A, Donati I, Pastore C, Onofrietti C, Spinelli F (2020) Facing Climate Change: application of Microbial Biostimulants to mitigate stress in Horticultural crops. Agronomy 10:794
Shaaban MM (2001) Green microalgae water extract as foliar feeding to wheat plants
Shariatmadari Z, Riahi H, Seyed Hashtroudi M, Ghassempour A, Aghashariatmadary Z (2013) Plant growth promoting cyanobacteria and their distribution in terrestrial habitats of Iran. Soil Sci Plant Nutr 59:535–547
Sharma P, Jha AB, Dubey RS, Pessarakli M (2012) Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. Journal of Botany 2012, 217037
Singh JS, Pandey VC, Singh DP (2011) Efficient soil microorganisms: a new dimension for sustainable agriculture and environmental development. Agric Ecosyst Environ 140:339–353
Song X, Bo Y, Feng Y, Tan Y, Zhou C, Yan X, Ruan R, Xu Q, Cheng P (2022) Potential applications for multifunctional microalgae in soil improvement. Front Environ Sci 10
Souza R, Ambrosini A, Passaglia LM (2015) Plant growth-promoting bacteria as inoculants in agricultural soils. Genet Mol Biol 38:401–419
Spadaro D, Gullino ML (2005) Improving the efficacy of biocontrol agents against soilborne pathogens. Crop Prot 24:601–613
Stirk WA, Ördög V, Van Staden J, Jäger K (2002) Cytokinin- and auxin-like activity in Cyanophyta and microalgae. J Appl Phycol 14:215–221
Stirk WA, Ördög V, Novák O, Rolčík J, Strnad M, Bálint P, van Staden J (2013) Auxin and cytokinin relationships in 24 microalgal strains(1). J Phycol 49:459–467
Suzuki N, Rivero RM, Shulaev V, Blumwald E, Mittler R (2014) Abiotic and biotic stress combinations. New Phytol 203:32–43
Swain SS, Paidesetty SK, Padhy RN (2017) Antibacterial, antifungal and antimycobacterial compounds from cyanobacteria. Biomed Pharmacother 90:760–776
Swarnalakshmi K, Prasanna R, Kumar A, Pattnaik S, Chakravarty K, Shivay YS, Singh R, Saxena AK (2013) Evaluating the influence of novel cyanobacterial biofilmed biofertilizers on soil fertility and plant nutrition in wheat. Eur J Soil Biol 55:107–116
Tantawy SA, Atef N (2010) Growth responses of Lupinus termis to some plant growth promoting cyanobacteria and bacteria as biofertilizers. J Food Agric Environ 8:1178–1183
Tarakhovskaya ER, Maslov YI, Shishova MF (2007) Phytohormones in algae. Russ J Plant Physiol 54:163–170
Toribio AJ, Suárez-Estrella F, Jurado MM, López-González JA, Martínez-Gallardo MR, López MJ (2022) Design and validation of cyanobacteria-rhizobacteria consortia for tomato seedlings growth promotion. Sci Rep 12:13150
Touloupakis E, Faraloni C, Silva Benavides AM, Masojídek J, Torzillo G (2021) Sustained photobiological hydrogen production by Chlorella vulgaris without nutrient starvation. Int J Hydrog Energy 46:3684–3694
Trivedi P, Schenk PM, Wallenstein MD, Singh BK (2017) Tiny microbes, big yields: enhancing food crop production with biological solutions. Microb Biotechnol 10:999–1003
Unnithan VV, Unc A, Smith GB (2014) Mini-review: a priori considerations for bacteria–algae interactions in algal biofuel systems receiving municipal wastewaters. Algal Res 4:35–40
Weiss TL, Roth R, Goodson C, Vitha S, Black I, Azadi P, Rusch J, Holzenburg A, Devarenne TP, Goodenough U (2012) Colony organization in the green alga Botryococcus braunii (race B) is specified by a complex extracellular matrix. Eukaryot Cell 11:1424–1440
Woo SL, Pepe O (2018) Microbial consortia: promising Probiotics as Plant Biostimulants for sustainable agriculture. Front Plant Sci 9:1801
Xiao R, Zheng Y (2016) Overview of microalgal extracellular polymeric substances (EPS) and their applications. Biotechnol Adv 34:1225–1244
Yanti Y, Hamid H (2021) and Reflin Development of the PGPR and Cyanobacteria Consortium for Growth Promotion and Control Ralstonia syzigii subsp. indonesiensis of Tomato. IOP Conference Series: Earth and Environmental Science 709, 012085
Yao S, Lyu S, An Y, Lu J, Gjermansen C, Schramm A (2019) Microalgae-bacteria symbiosis in microalgal growth and biofuel production: a review. J Appl Microbiol 126:359–368
Yilmaz E, Sönmez M (2017) The role of organic/bio–fertilizer amendment on aggregate stability and organic carbon content in different aggregate scales. Soil Tillage Res 168:118–124
Zhang B, Li W, Guo Y, Zhang Z, Shi W, Cui F, Lens PNL, Tay JH (2020) Microalgal-bacterial consortia: from interspecies interactions to biotechnological applications. Renew Sustain Energy Rev 118:109563
Funding
This research did not receive any funding from outside sources.
Open access funding provided by Széchenyi István University (SZE).
Author information
Authors and Affiliations
Contributions
Wogene Solomon: conceptualization (principal); first draft writing (principal); data curation (equal); editing (equal); and investigation (equal). Lamnganbi Mutum: conceptualization (helping); editing (equal); and data curation (equal). Tibor Janda: conceptualization (helping); supervision (helping); editing (equal); data curation (equal); and investigation (equal). Zoltán Molnár: supervision and rectification (principal); conceptualization (helping); editing (equal); data curation (equal); and investigation (equal). The final manuscript was contributed by all authors.
Corresponding author
Ethics declarations
Conflict of interest
The authors affirm that they do not have any conflicts of interest.
Additional information
Communicated by Vijay Pratap Singh.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Solomon, W., Mutum, L., Janda, T. et al. Potential benefit of microalgae and their interaction with bacteria to sustainable crop production. Plant Growth Regul 101, 53–65 (2023). https://doi.org/10.1007/s10725-023-01019-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10725-023-01019-8