
Abstract
Middle Devonian reefs were widespread on the various synsedimentary tectonic blocks of the Moroccan Meseta, which formed the southern external Variscides. So far, poorly studied biostromes and small to medium-sized bioherms have grown in the southern subtropics, separated by the ca. 3000-km-wide Prototethys from the well-known reefs of the southern Laurussian shelf. Autochthonous, displaced, and reworked Givetian reefal limestones of the Oued Cherrat Zone south of Rabat and from adjacent regions were studied by outcrop facies logging and microfacies analysis. The new data are used to reconstruct their paleoecology, developments in time and space, and contribute to the understanding of reef facies differences along a paleolatitudinal gradient. Eighteen microfacies (sub) types are recognized, described, and illustrated. They represent thick inner-reef platforms (Aïn Khira South, Jennabia), different parts of a synsedimentarily tectonized bioherm (Aïn-Al-Aliliga), a small, open neritic biostrome/low bioherm (Aïn-as-Seffah), the episodically retro-and prograding outer margin of a thick biostrome to bioherm sequence (Sidi Mohamed Smaine), and reworked reef remnants preserved in proximal to distal olistolith units (Aïn Dakhla, Sidi Ahmed Lhemdoun, Biar Setla Conglomerate). Meseta reefs differed from those of stable northern Gondwana (western Anti-Atlas to southern Algeria) but show many similarities with the more distant Rhenish organic buildups. This suggests a very low Givetian paleoclimatic gradient in the southern hemisphere. The lack of facies types dominated by calcimicrobes/calcareous algae is distinctive for the study area.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The Middle and early Upper Devonian (Givetian–Frasnian) is distinguished by the worldwide flourishing of large reef complexes. The Phanerozoic reef database (Kiessling et al. 1999; Copper 2002; Copper and Scotese 2003) summarized the paleogeographic distribution and the number of reefs of this interval. It is seen as a Phanerozoic reef maximum (Kiessling 2008; Flügel and Kiessling 2002b) of a prolonged global greenhouse time (e.g., Copper 2002). Typical Devonian reefs were built by stromatoporoids, tabulate and rugose corals, calcimicrobes (cyanobacteria and green algae), and, to a less extent, by bryozoans. In different regions and paleogeographic settings they exhibit a variable paleoecology. In a wide sense of the term (Flügel and Kiessling 2002a), Devonian reefs include large bioherms, consisting of an initial bank facies growing into differentiated reef slope and protected platform realms (e.g., common in the eastern Rhenish Slate Mountains of Germany, Krebs 1974), smaller-scale atolls sitting on volcanic buildups (Flick and Schmidt 1987), biostromes (e.g., common in the Eifel area of the western Rhenish Slate Mountains, Faber 1980), and mud mounds (e.g., common in the Frasnian of the Ardennes, Boulvain 2007). Most of the large Devonian reefs are situated in (moderately) low paleolatitudinal positions (max. 30°N/S) inside an expanded tropical belt (Kiessling et al. 1999: Fig. 6; Copper 2002: Fig. 7). A smaller number developed outside, reaching paleolatitudes of at least 45°N/S (Kiessling 2002; see plate tectonic map of Golonka 2002).
Of special interest for a deeper understanding of mid-Paleozoic reef ecology and distribution is the comparison of contemporaneous reefal limestones along a paleolatitudinal and paleoclimatic gradient, for example between the southern shelf of Laurussia (Rhenish Slate Mountains) and the northern shelf of Gondwana (Moroccan Meseta). During the Givetian/Frasnian, Moroccan Meseta reefs were separated from the Rhenohercynian Zone roughly from south to north by the western Prototethys, southern European terrains (e.g., Catalonia, Menorca, Sicily, Sardinia, Calabria), the widely flooded Armorican Terrain Assemblage (Iberia to Bohemia), and remnants of the closing Rheic Ocean (e.g., Nance et al. 2012, recent plate tectonic model of Eckelmann et al. 2014). The added terrain width (current distances) and calculations for the Variscan crustal shortening (ca. 50 % in the Rhenohercynian Zone, Oncken et al. 1999) suggest an at least 3000-km-wide (roughly N–S) Middle Devonian seaway between Gondwana and Laurussia (the “Variscan Sea” sensu Neugebauer 1988). Other authors claimed that it was significantly wider. Franke (2007), for example, proposed a width of possibly more than 2000 km for the mid-European Variscides alone. The estimated paleolatitudinal distance in the scale of 3000 km (possibly up to 4000 km) does not consider claims of significant lateral (paleolongitudinal) separation. For example, Cocks and Torsvik (2002) illustrated a highly unlikely extreme eastward shift of North Africa, Simancas et al. (2005) a strong westward shift. Based on paleomagnetic data, some authors (Kent and Van der Voo 1990; Bachtadse et al. 1995; Stampfli and Borel 2002) postulated an even wider western Prototethys, which, however, would place the Moroccan reefs at unrealistic southern paleolatitudes of 50–60°. In any case, the significant N–S distance of reefs at the southern margin of the tropics should have resulted in clear paleoecological differences, an aspect that has not been addressed in previous studies.
The most southern North African Middle Devonian reefs occur on the stable cratonic part of northern Gondwana in the western Tindouf Basin of former Spanish Sahara (e.g., Dumestre and Illing 1967; Königshof and Kershaw 2006; Wendt and Kaufmann 2006; Ernst and Königshof 2008, 2010; May 2008) and in the adjacent Mauritanian Zemmour (e.g., Coen-Aubert 2013). Eastwards, thin and small biostromes and larger mud mounds are developed in the Tafilalt/Maider region (eastern Anti-Atlas; Hollard 1962, 1974; Massa 1965; Töneböhn 1991; Wendt 1993; Kaufmann 1995, 1998; Schröder and Kazmierczak 1999; Kazmierczak and Schröder 1999; Döring and Kazmierczak 2001; Coen-Aubert 2002, 2005; Aboussalam 2003; Fröhlich 2003; Nübel and Becker 2004; Aboussalam and Becker 2011; Becker et al. 2013). Even further onto the craton, mud mounds re-appear in the Ahnet Basin of southern Algeria (e.g., Wendt et al. 1993, 1997).
By comparison, the widespread reef limestones of the Meseta are much less known. Apart from general regional geological overviews that mention specific reef occurrences (e.g., Hollard 1967; Zahraoui 1994; El Hassani and Benfrika 2000), there are only a few, often-short specific publications on Meseta reefs by Gendrot (1973), Cattaneo et al. (1993), El Hassani and El Kamel (2000), and Zahraoui et al. (2000). More details but with limited microfacies data were included in unpublished investigations and PhD thesis (Gendrot et al. 1969; El Kamel 1987; Zahraoui 1991). Hüneke and Krienke (2004) described the sedimentology of a carbonate platform slope of the Rabat-Tiflet Zone, which was rimmed by a Givetian reef. Rytina et al. (2013) and Rytina (2013) recently documented the allochthonous reef facies at the southern margin of the Variscides, preserved in Carboniferous olistostromes of the Tinerhir region.
Our paper presents new data on Givetian to lower Frasnian reefal limestones of the Western Moroccan Meseta, which represent different sedimentary–tectonic settings. As the first part of a larger investigation of all Meseta regions, we present 18 facies types/subtypes for the reef development in the Oued Cherrat Zone, in its southern prolongation (Al Attamna region), and for the adjacent Khatouat area further to the southeast, including reworked reef clasts of eo-Variscan breccias triggered by synsedimentary tectonic movements. These investigations were imbedded in IGCP 596 on “Climate change and biodiversity patterns in the Mid-Paleozoic”.
Geological setting and studied regions
The North African Meseta region is located in the northern part of Morocco and Algeria. It includes rare tectonic windows within the High Atlas and some outcrops with Variscan deformation to the south of it (e.g., Tinerhir region). To the north, the Rif represents a different terrain that was aligned with the Kabylei of northern Algeria in Devonian times (Hoepffner et al. 2005). In the west the Meseta is bordered by the Atlantic Ocean, in the east it continues into western Algeria. In Morocco, the Meseta is split by the Middle Atlas into the Western and the Eastern Meseta. Hollard (1967), Pique and Michard (1981, 1989), El Hassani and Benfrika (2000), and Hoepffner et al. (2005, 2006) characterized the different Paleozoic structural units and summarized the sedimentary and tectonic evolution of Hercynian Morocco. The region was heavily affected by eo-Variscan crust movements, which refers to subsidence changes and widespread block faulting in the Emsian to Tournaisian, the time after the main closure of the Rheic Ocean and before the main Gondwana-Laurussia collision (main Variscan deformation; e.g., Michard et al. 2008; Becker et al. 2015a).
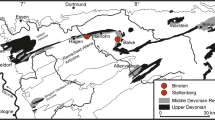
Reefal limestones of the Western Meseta crop out in the Coastal Block (El Kamel 1987), Rabat-Tiflet Zone (El Hassani 1991), West Central Meseta (Oued Cherrat Zone, Al Attamna, western and northeastern Mdakra Massif, Khatouat Massif, e.g., Termier and Termier 1951; Hollard 1967; Zahraoui 1991), East Central Meseta (Oulmes region, Cogney 1967; Azrou and Mrirt regions, Termier et al. 1975; Bohrmann and Fischer 1985), the Middle Atlas basement S of Fes (Cygan et al. 1990), the Rehamna (El Kamel 1987; El Hassani and El Kamel 2000), the western Jebilet (Huvelin 1977; Tahiri 1983), and in the High Atlas (Hollard 1967) (Fig. 1).

Distribution of auto- and allochthonous Givetian reef limestones in the Moroccan Meseta 1 western Jebilet (Jebel Ardouz), 2 Rehamna (Mechra-ben-Abbou), 3 Coastal Block, 4 SW Mdakra Massif (Benahmed region), 5 Oued Cherrat Zone, 6 Al Attamna, 7 NE Mdakra Massif (Chabet-el-Baya), 8 eastern Khataoua Massif (Jennabia), 9 Rabat-Tiflet Zone, 10 Oulmes region (Ain Jemaa), 11 Mrirt region (Dechra Ait Abdallah), 12 Azrou region, 13 Middle Atlas basement (Immouzer-al-Kander)
Oued Cherrat Zone
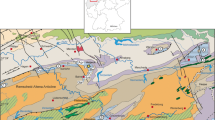
The Oued Cherrat Zone, named after the N–S-running and deeply incised Oued Cherrat valley, is a N–S-running fault zone beginning between Rabat and Casablanca in the western Central Massif (Chalouan 1981; Zahraoui et al. 2000, Fig. 2). It is delimited by two faults, which separate the Famennian-Tournaisian formations of Benslimane to the west and of the Sidi Bettache Basin to the east. Chalouan and Hollard (1979) introduced the current lithostratigraphic subdivision that requires updates because of new biostratigraphic data (Aboussalam et al. 2012, 2013; Fig. 3). The regional succession ranges from the upper Silurian (Ludlow) to the Lower Carboniferous. The first neritic limestones appeared during the upper Pragian/lower Emsian (cherty trilobite limestones in the lower part of the Dhar-es-Smene Formation) and graded into a lower Emsian reef platform that is well exposed on the western side of Oued Cherrat, for example at Ain Dakhla (Zahraoui et al. 1991; Figs. 2, 3, 4). A second massive reef belt of supposed Givetian age east of the valley, for example at Cakhrat-ach-Chleh, turned out to fall also in the Emsian, too (Aboussalam et al. 2012). Based on its type-locality, the Cakhrat-ach-Chleh Formation of Chalouan and Hollard (1967) became a synonym of the Dhar-es-Smene Formation. Herewith we propose to use the first term for the reefal upper member of the latter (Fig. 3). The Emsian reefs (biostromes) are variably overlain by the upper Emsian/Eifelian Ain-Kheneg-an-Nmer (turbiditic dark limestones and marls, western belt, e.g., at Ain-as-Seffah) or Mohamed-Ben-Brahim Formations (argillaceous, eastern belt, e.g., east of Ain-Al-Aliliga, locally followed by un-named neritic limestones, Fig. 3).

Position of sampled reef limestone localities in the Oued Cherrat Zone AKS Aïn Khira South, AD Aïn Dakhla, CAC Cakhrat-ach-Chleh, AAA Aïn-Al-Aliliga, AAS Aïn-as-Seffah (after Zahraoui 1991)

Overview of the lithostratigraphy and position of autochthonous and allochthonous reefal limestones in the Oued Cherrat Valley, Al Attamna, and Khatouat Massive (based on conodont data of ZS Aboussalam)

Geographic position of investigated localities in the Al Attamna, Mdakra, and Khatouat regions (adopted from topographic sheet 1:50,000 Sidi Zemmouri) SAL Sidi Ahmed Lemdoun, SMS Sidi Mohamed Smaine, Jennabia/Biar Setla
True Givetian reefal limestones crop out in a separate, second, more northern belt along the eastern valley margin (Fig. 2). This unit, here assigned to the new Aïn Khira Formation, was previously briefly described by Zahraoui (1991) and Cattaneo et al. (1993) at Aïn Khira. They recognized two reefal sequences. Unfortunately, intense quarrying has destroyed many outcrops in the Aïn Khira area, including the previously figured quarry with large polished walls that was shown during a fieldtrip of the International Subcommission on Devonian Stratigraphy (SDS) in 2000 (Zahraoui et al. 2000). Therefore, our fieldwork concentrated on a new section bordering the large active quarries at Aïn Khira South. Towards the south, at Aïn Dakhla (Fig. 2), reworked Givetian reef limestone clasts (partly rich in alveolitids) occur isolated or in polymictic conglomerates (debris-flow deposits) in the lower part of the supposed Famennian, mostly siliciclastic Al Brijat Formation. Conglomerate olistoliths with mixed Eifelian-Givetian conodonts gave evidence for two separate reworking phases (Aboussalam et al. 2012). Further southwards, at Ain-Al-Aliliga, a similar, thick reworking unit (Ain-Al-Aliga Formation) lies directly on Emsian reef limestone. It consists of intercalating reef limestone and synsedimentary brecciated beds, which grade into coarse upper Givetian to ?basal Frasnian polymictic breccias. We sampled the various reef lithologies preserved within this allochthonous complex, which is overlain by thin Frasnian dolomites. Even further to the south, at Ain-as-Seffah, a tectonically displaced block (large olistolith, Ain-es-Seffah Formation, Members 2–3) consists of an internally pristine Givetian reef succession in disconformable contact with under- (Member 1) and overlying non-reefal carbonates, breccias and shales (Members 4–5; preliminary data in Eichholt et al. 2013).
Al Attamna
The Al Attamna region is the southern prolongation of the Oued Cherrat fault zone (Fig. 4). The region was studied by Fadli (1990), who distinguished two ca. N–S-striking outcrop areas at Sidi Ahmed Lemdoun (SAL) and Sidi Mohamed Smaine (SMS). Benfrika and Bultynck (2003) presented conodont data for both successions, which are only a few kilometers apart. The Sidi Ahmed Lemdoun Formation consists of four units ranging from the Pragian (Unit A) to a supposed Famennian conglomerate (Unit D) with reworked reefal blocks. The latter consists in fact of two different units. The polymictic lower “red conglomerate”, a debris-flow deposit, is middle Givetian in age and overlain by Givetian marls and nodular limestone. The main, upper conglomerate, a channel-fill consisting of several debris-flow beds, yielded very rare middle Famennian palmatolepids in assemblages dominated by middle Givetian taxa (conodont data of Z.S. Aboussalam). The Sidi Mohamed Smaine Formation consists of three thick limestone units of early middle Givetian age (Po. varcus zone). The regionally abrupt lateral change from a thick reefal platform (SMS), accumulated in a shallow belt with fast subsidence, to a highly condensed but deeper neritic succession (SAL) with very low subsidence, intercalated and overlain by two polymictic breccia levels with reef remnants, proves a small-scale paleotopography that was significantly influenced by synsedimentary (Eovariscan) tectonics. Further to the south, at Chabet el Baya (Fig. 4), the Al Attamna region turns into the northeastern Mdakra Massif, where allochthonous reef blocks were reported from supposed Famennian conglomerates (Fadli 1990, 1994). Unfortunately, we were unsuccessful to retrieve such blocks from the thick, siliciclastic type Chabet El Baya Formation.
Khatouat
The complex depositional history and tectonics of the Khatouat Massif to the SE of the Oued Cherrat Zone have been studied by Fadli (1990, 1994). A poorly preserved, strongly recrystallized reef is exposed in the northeastern succession at Jennabia (Fig. 4; note that the name is marked on the topographic map Sidi Zemmouri at a hill 2.6 km to the east). Conodont dating was unsuccessful but a Givetian age is likely since there is no Lower Devonian (Emsian reef) outcrop in the Khatouat (Zahraoui 1994) and with respect to the dating of reworked clasts. These occur in a valley to the north, within the polymictic Biar Setla Conglomerate (member) of the Fouzir Formation. Parts of the Jennabia carbonate platform were eroded during eo-Variscan block tilting and local uplift and reworked as conglomerates/breccias (debris-flows). In a second (?Famennian) phase, these were re-sedimented along a steep fault scarp as isolated olistoliths into an adjacent siliciclastic basin. Rare conodonts (data of ZS Aboussalam) and phillipsastreid colonies confirm that clasts within reef breccias belong to the lower to upper Givetian while reworked Emsian strata are only represented by non-reefal, crinoidal limestones (Fig. 3).
Methods
Fieldwork took place in autumn 2011, 2012, and spring 2014. At Aïn Khira South (AKS), Aïn-as-Seffah (AAS), and Sidi Mohamed Smaine (SMS) detailed sections were obtained by field logging. Macroscopic facies characteristics were documented every meter and for each facies change representative rock samples were collected for microfacies analysis. At Aïn Dakhla (AD), Aïn-Al-Aliliga (AAA), Jennabia (JEN), Biar Settla (BS), and Sidi Ahmed Lemdoun (SAL) reworked and re-deposited Givetian reef limestones of breccias/conglomerates were collected with the aim to record representative sets of local facies types.
Thin-sections of different size (28 × 48, 50 × 50, 75 × 100, and 100 × 150 mm) were studied with a transmitted light binocular (Leica S4E). Based on the expanded classification of limestones (Dunham 1962; Embry and Klovan 1971), Standard Microfacies Types (SMF) sensu Flügel (1978, 2004) and Facies Zones (FZ) sensu Wilson (1975) were characterized. SMF and FZ are idealized models for carbonate platforms and provided the orientation for the definition of some new facies types. These were interpreted in the light of previous Devonian reef studies and provided a frame for the reconstruction of local reef development through time.
Meseta reef facies and microfacies
Previous work
Gendrot (1973) investigated Devonian reefs of Morocco and described the main reef builders, such as stromatoporoids, tabulate and rugose corals, and their habitats. He classified the regional reef environments into off reef, distal fore reef, fore reef (fore reef breccia), reef core, lagoon (back reef), and inter- and supra-tidal zones. In their study of the Aïn Khira (Oued Cherrat) and Bou Sif sections (Tiliouine area N Oulmes), Cattaneo et al. (1993) recognized eight different Givetian facies types (F1–F8) that were assigned to four environments:
Outer shelf (F1–F3).
F1. Mudstones-wackestones with Tentaculites.
F2. Wackestones-floatstones with small isolates colonies of lamellar stromatoporoids, Favosites, discoid “Tetracorallia” (=Rugosa) and branching Thamnopora.
F3. Peloidal packstones-grainstones with crinoids and bryozoans.
Reef building shelf edge (F4–F5).
F4. Floatstones to coarse rudstones with stromatoporoids and “Tetracorallia”.
F5. Bindstones–framestones with massive colonies of globular, bulbous or lamellar-encrusting stromatoporoids, “Tetracorallia”, and Tabulata.
Inner platform (F6–F7).
F6. Floatstones with Amphipora.
F7. Wackestones/mudstones with parathuramminoids and Amphipora.
Littoral and intertidal (F8).
F8. Pelsparites with birds-eyes, pelmicrites, and laminites.
Our samples enable the recognition of eight lagoonal facies types (MF set A), five types of fore-reef and slope facies, developed as detritus-rich grain-, float- and rudstones (MF set B), and of five types of autochthonous bioherms or biostromes, notably boundstones (MF set C). Facies F1–F8 sensu Cattaneo et al. (1993) are correlated with our extended facies set.
Lagoonal facies set (MF A)
MF A1: Peloidal grainstone
The light grey limestones consist of small micritic lithoclasts within intergranular-peloidal microcrystalline calcite cement. The micritic clasts are well to moderately rounded mud peloids and fecal pellets of equal size. The bioclastic content is low (<10 %); gastropods (Fig. 5c) and brachiopods are the only macrofossils. In thin-section, some parathuramminid foraminifera (Archaesphaera, Bisphaera) were found (Fig. 5b). In some samples, numerous calcite veins and fissures reflect tectonic/late diagenesic stress (Fig. 5a). Pressure solution triggered stylolites and microstylolites.

a–c MF A1 Peloidal grainstone. a Large gastropod within a peloidal grainstone, AKS, 8 m. b Magnification of Fig. 2 showing parathuramminid foraminifera (black arrows), AKS, 8 m. c Field photograph of a large, thick-shelled gastropod, AKS, 8 m. d MF A7 (bioclastic) mud-/wackestone with shell fragments of a gastropod and ostracod (black arrows); burrows are filled with lighter micrite (white arrows), AKS, 24 m. e MF A2 detrital bioclastic grainstone/rudstone; variably sized detritus of stromatoporoids (S), tabulate corals (T) and echinoderms (E) within a peloidal grainstone matrix AKS, 10 m. f MF A3 (detrital) Stringocephalus Amphipora float-/rudstone; Stringocephalus shells (B), Amphipora branches (A) and tabulate corals (T), AKS, 12 m. g MF A4 Stachyodes Thamnopora float-/rudstone; component contacts between Stachyodes (S) and Thamnopora (T) show stylolite seams and grain penetration (black arrows), AKS 21 m. h MF A5 stromatoporoid float-/rudstone; delicate Amphipora (A) branches between bulbous stromatoporoids; clay seams and irregular component contacts create a stylobreccioid structure, AKS, 56 m. i MF A8 Stringocephalus floatstone; Stringocephalus showing median septum within a bioclastic wackestones matrix; only the outer shell shows microborings (black arrows) suggesting a long pre-burial exposure AKS, 62 m. j MF A6 Amphipora bafflestone with stylolitic contact of many branches, AKS, 96 m
Interpretation and comparisons: Lagoonal back reef environments with moderate water agitation and strongly restricted fauna (Fig. 6). As a result of continuous current activity, micrite was winnowed and semi-lithified carbonate mud reworked to peloids (Flügel 2004). The parathuramminids include “calcispheres” of various authors and indicate shallow-water conditions in the back-reef (Flügel and Hötzl 1971). However, Faber (1980) documented that they can occur in the Eifel Mountains both in lagoonal and fore-reef facies. MF A1 is very similar to SMF 16 (Flügel 2004) but occasionally it contains gastropods a well as brachiopods. It belongs to FZ 8 (Wilson 1975) and is recognized for the first time in Givetian reef complexes of Morocco. It is well known from German reefs of the Harz Mountains (Franke 1973: part of facies type Q, Fuchs 1990; Weller 1991).

Simplified facies model for back reef settings within bioherms (Ain Khira, Jennabia) and biostromes to open (conodont-bearing) bioherms (Ain-as-Seffah, Sidi Mohamed Smaine) of the study region. Within lagoons (upper sketch) and open biostromal platforms (lower sketch) more complex lateral facies intergradations than shown are possible
MF A2: detrital, bioclastic grainstone/rudstone (back-reef breccia)
The light-to-middle grey limestones consist of detrital and broken reefal organisms, such as fragmented, dendroid stromatoporoids (Amphipora and Stachyodes) and fragmented, bulbous and laminar stromatoporoids. Secondary are clasts of tabulate corals and small, solitary rugose corals; the latter are often overgrown by stromatoporoids. Further bioclasts are fragmented shells and parathuramminid foraminifera. Fragments are rounded and reach a diameter of a maximum of 10 mm. There are only rare, scattered crinoid fragments. The matrix consists of a peloidal grainstone similar to MF A1, but it is somewhat darker (probably richer in C org). Some shell fragments have distinctive micrite seams. Minor stylolites and fissures indicate pressure solution.
Interpretation and comparisons: Detritus-rich deposits (back reef breccias) within the platform interior (Fig. 6). The broken and rounded parts of Amphipora intermixed with other stromatoporoids indicate phases of increased water agitation by storms, which imported some crinoid debris from the fore reef flanks, the typical crinoid habitat of Devonian reefs. This facies type is found adjacent to MF A1 and MF A3 and it is similar to SMF 16 (Flügel 2004) but in contrast it includes additional storm-derived detritus. Therefore, it better corresponds to SMF 5, which is common in reef flank facies or on the marginal slope, but which also appears in back reef settings adjacent to the reef core or next to patch reefs (MF A5). It reflects within the platform interior open-marine conditions (FZ 7). MF A2 corresponds to F4 of Cattaneo et al. (1993), which was placed within platform interiors adjacent to patch reefs. A similar facies is known from the Rhenish Massive as the detrital stromatoporoid-coral-algal facies (Krebs 1974) or included in facies Ha5-7 of the Elbingerode Reef in Weller (1991).
MF A3 (detrital) Stringocephalus-Amphipora rudstone
This facies consists mainly of detrital and broken very thick brachiopod shells (stringocephalids) and fragmented Amphipora branches, which build densely packed float-/rudstones. Fragmented clasts of thamnoporoids are uncommon. Spaces between components are filled with coarse-grained detritus or peloidal micrite. One sample at AKS (11.5 m above the section base) has a matrix consisting completely of calcite cement. Veins, stylolites, solution seams, and grain penetration indicate a very high rate of pressure solution and tectonic stress during diagenesis.
Interpretation and comparisons: Storm layers within the platform interior. Stringocephalus and dendroid stromatoporoids, especially Amphipora, were typical dwellers of shallow and calm regions of restricted lagoonal settings. Storm events destroyed and disarranged the brachiopods and Amphipora meadows. This facies type is found adjacent to MF A2 and MF A6. It is similar to SMF 12-Brach (Flügel 2004) but the association with dendroid stromatoporoids indicates a transport of broken brachiopods into an Amphipora meadow. It belongs to FZ 7 (Wilson 1975) and has not been distinguished by Cattaneo et al. (1993) within their F4/F6. It is very characteristic at Aïn Khira South and should be distinguished, as is the case in the lagoonal settings of Rhenish reefs (e.g., Jux 1960, Stringocephalus facies of Krebs 1974, lagoonal micrites with fragmented stringocephalids in Schudack 1993; May 2003), including the Harz Mountains (Weller 1991).
MF A4 Stachyodes-Thamnopora float-/rudstone
The middle to dark gray limestones consist mainly of dendroid stromatoporoid (Stachyodes) clasts, with a smaller amount of Amphipora and branching tabulate corals (mainly Thamnopora). Other bioclasts are shell fragments, ostracods, and parathuramminids. The matrix consists of coarse-grained, slightly winnowed micrite and peloids. Especially at AKS the matrix is very dark and components are densely packed with stylolite seams, grain penetration, and irregular grain boundaries, so that the whole facies type has a stylobreccioid structure (Fig. 5g).
Interpretation and comparisons: Back-reef lagoons next to patch reefs (Fig. 6, MF A5; FZ 7, Wilson 1975). The dendroid stromatoporoid Stachyodes was a dweller of niches within the reef and near the reef core in moderately agitated water (Krebs 1974). It was stronger and more resistant against currents and wave action than the delicate Amphipora branches. The partially winnowed micrite indicates repeated current and wave activity. The dark color at AKS is presumably based on a high clay input; this assumption is supported by the stylobreccioid structure with distinct developed clay seams around the components. MF A4 belongs to SMF 8 sensu Flügel (2004). Stachyodes-dominated back-reef limestones also occur in the Rhenish Massif (e.g., Faber 1980; Schudack 1993). Cattaneo et al. (1993) may have included MF A4 in their F5, which, however, contains more dendroid than bulbous or laminar stromatoporoids. In the Harz Mountains, facies type M sensu Franke (1973) is similar but differs in its sparite matrix.
MF A5 stromatoporoid float-/rud-/boundstone (patch reef facies)
The middle to dark gray limestones consist of tabular and bulbous stromatoporoids. The dominant bulbous forms (ca. 50 % of the whole limestone) reach a diameter of up to 20 cm. Secondary are dendroid stromatoporoids, especially Stachyodes and Amphipora, tabulate corals (Thamnopora), and solitary Rugosa. Additional fossils are parathuramminids and ostracods. The matrix consists of coarse-grained detritus, slightly winnowed micrite, and peloids. Clay seams, grain penetration, and microstylolites are common, based on compaction and pressure solution, which creates a very coarse-grained appearance similar to MF A4.
Interpretation and comparisons: Small patch reefs within the open-marine platform interior (Fig. 6), FZ 7 (Wilson 1975). The micrite content indicates low-energy conditions. The wave-resistant bulbous stromatoporoids sheltered niches that were settled by Stachyodes, Thamnopora, and delicate Amphipora. MF A5 belongs to SMF 7 (Flügel 2004) and F5 (Cattaneo et al. 1993). In Rhenish reefs, it is widely known, e.g., from the inner platform of the Brilon, Balve, Dornap, and Elbingerode reef complexes (Krebs 1974; Malmsheimer et al. 1991; Fuchs 1990; Weller 1991; Schudack 1993) or from lagoons of small atolls (Flick and Schmidt 1987). Krebs (1974) described it as dark bulbous stromatoporoid-Amphipora facies.
MF A6 Amphipora float-/rud-/boundstone
This facies type consists mainly of thin dendroid stromatoporoids (Amphipora), secondarily of Stachyodes and Thamnopora. Occasionally, the stromatoporoids are preserved in situ, but in most cases they were broken and re-sedimented as float- or rudstones. The additional fossil content is very low (small brachiopods, ostracodes, and parathuramminids) and similar to MF A4. The matrix consists of dark micrite. Diagenetic alteration played a major role; the bioclasts are very densely packed and show grain penetration, solution seams, and stylolites. Even the micrite has a re-crystallized, coarse-grained stylobreccioid habitus.
Interpretation and comparisons: Quiet and calm facies realm within the protected platform interior, with ideal growth conditions for dendroid stromatoporoids. Amphipora was a typical dweller of shallow and calm, restricted lagoonal settings of European reefs (Krebs 1974: dark Amphipora facies), e.g., of the Brilon (Malmsheimer et al. 1991), Balve (Schudack 1993), and Elbingerode reefs (Weller 1991). After Wilson (1975), this facies type belongs to facies zone 7 and it corresponds to SMF 7 (Flügel 2004). It is F6 of Cattaneo et al. (1993).
MF A7 (bioclastic) mud-/wackestone
This facies type consists of middle-gray micrite and very fine grained peloidal micrite. Bioturbation created a common, cloudy appearance. The macrofossil content is restricted; there are only some gastropods and fragments of dendroid stromatoporoids and tabulate corals in specific samples. In thin-sections, small ostracods, rare parathuramminids, and gastropods may occur. Some samples show birdseye structures filled with calcite cement. Burrows are variably filled with micrite, peloids, or calcite cement. Some of them and the inner space of ostracods are filled with geopetals.
Interpretation and comparisons: Restricted platform interiors unfavorable for a diverse benthos. The micritic matrix indicates quiet and calm environments, without significant current or water agitation (Fig. 6). The decrease of biodiversity and the presence of birdseyes suggest a transition to protected intertidal environments (Shinn 1968), possibly with occasional freshwater influx by heavy rain (Faber 1980). The (bioclastic) mud-/wackestones are adjacent to and intercalated with MF A4, MF A5, and MF A6. This facies type is most similar to SMF 19 (Flügel 2004), but without its significant lamination. After Wilson (1975), it belongs to facies zone 8 and it corresponds with F7 and reaches into F8 of Cattaneo et al. (1993). In Rhenish reefs, Krebs (1974) described it as “dark fossil-poor micrite facies”, Koch-Früchtl and Früchtl (1993) as “Foraminifera-Calcisphere Wackestone”. Faber (1980) included it in his Calcisphere Mudstone. MF A7 also occurs in sheltered lagoons of small-sized atolls (Flick and Schmidt 1987), in larger platforms of the Harz Mountains on top of volcanic (Elbingerode Reef, Fuchs 1990; Weller 1991) or structural elevations (Iberg Reef, Franke 1973), or in protected lagoonal settings within Frasnian mud mounds of the Ardennes (Boulvain 2007).
MF A8 Stringocephalus floatstone
This facies type is similar to MF A7. It consists of a higher amount of macrofauna, mainly of thick-shelled brachiopods (Stringocephalus) and secondarily of dendroid and laminar stromatoporoids, as well as small solitary rugose corals. Additional bioclasts are small ostracods, parathuramminids, and small shell fragments. The matrix consists of bioclastic mud-/wackestone as in MF A7. Bioturbation fabrics within the matrix are filled with peloids and pelmicrite. The inner space of shells shows geopetals, the outer shell often beginning micritization by microborings.
Interpretation and comparisons: Protected, wide, shallow, open-marine carbonate platform. Stringocephalus is a thick-shelled brachiopod with a distinctive median septum and indicates a Givetian age. It is a typical fossil in lagoonal settings (Struve 1989) and also well known from European back reef environments (Krebs 1974: “Stringocephalus Facies”; Schudack 1993). The increasing diversity as well as the higher amount of bioturbation than in the former facies type indicate an open-marine influence with better living conditions. After Wilson (1975) this facies type belongs to facies zone seven and it is similar to SMF 9. The micritization suggests a long exposure of brachiopod shells on the seafloor to cyanobacteria, such as Girvanella (Faber 1980). Stringocephalus has previously been noted in the Oued Cherrat area (Roch 1950; El Hassani and Benfrika 2000) but not been recognized in biofacies analysis. The matrix composition of MF A8 is similar to F7 of Cattaneo et al. (1993).
Fore reef facies set MF B
MF B1 crinoid facies
MF B1a crinoid grainstone
The light to middle gray limestones consist of crinoid debris, peloids, and small bioclasts within an intergranular, sparry calcite matrix. The crinoids are the main components; secondary are peloids, cortoids, and smaller shell fragments. Small fragmented clasts of stromatoporoids and corals (diameter max. 5 mm, less than 10 %) are rare. Micrite is completely winnowed. In some samples, crinoids have a poor state of preservation; especially at the base of Aïn-as-Seffah they have been grinded. In all samples they show a distinctive syntaxial calcite overgrowth. Some lithoclasts have micrite envelopes, which may form cortoids.
Interpretation and comparisons: Middle part of marginal slope of bioherms, where crinoid forests were exposed to wave and current action (Fig. 6). MF B1a belongs to FZ 4 and equals SMF 12. Water agitation was strong enough to keep fine micrite in suspension, to rework semi-lithified carbonate mud, and to form small mud peloids. Small clasts of corals and stromatoporoids derived from the upper slope. Alternatively, crinoid mass accumulation may have taken place outside, around and at the base of biostromes, which may represent the initial stadium of bioherms. Krebs (1974) and Faber (1980) recognized crinoid limestones from within platform successions; such occurrences probably reflect times of reef drowning and increased open-marine circulation, supported by conodont occurrences. The very bad state of preservation of MF B1a at the base of Aïn-as-Seffah is a tectonic aspect based on the transport of the whole reefal block as a big olistolith. Cattaneo et al. (1993) recognized MF B1a as F2 but did not record it from the Oued Cherrat area. MF B1 occurs widely on the slopes of German reefs (e.g., Franke 1973; Krebs 1974; Weller 1991; Gischler and Erkoҫ 2012).
MF B1b bioclastic crinoid-brachiopod grainstone
The middle gray limestones consist of dominant crinoid debris and abundant fragmented brachiopods within a sparry calcite matrix. In addition, there are smaller shell fragments and calcareous algae (Girvanella). The predominant cement type is the syntaxial calcite overgrowth of crinoids. Dolomite crystals may be present, especially along stylolites. Compared to MF B1a, this subtype contains larger crinoid debris (up to 5 mm diameter) and the brachiopods while corals or stromatoporoids are completely lacking.
Interpretation and comparisons: Non-reefal, deeper neritic carbonate platforms (part of the brachiopodetum sensu Struve 1963) around biostromes and on the distal marginal slope of bioherms (e.g., Krebs 1974, subordinate brachiopod coquinas in the arenitic crinoid debris facies of Faber 1980). They belong to facies zone 4 and SMF 12, the main habitat of crinoids and brachiopods. Water agitation is strong enough to keep fine micrite in suspension and winnow it. This facies type is similar to MF B1a, but the lack of real fauna and the deposition on the top of the Aïn-as-Seffah Reef suggest deposition after extinction of the reef by drowning. It has not been recognized previously in the Meseta and is not a common post-reef facies type elsewhere.
MF B2 detrital coral-stromatoporoid floatstone/rudstone (fore reef breccia)
This facies type is very diverse and widespread in the whole Meseta. It is possible to define three subtypes, which have either been recognized in different outcrops (a versus b, c) or which show a different spatial distribution and different dominant bioclasts (b versus c).
a: Aïn-as-Seffah type (Fig. 7b, h)

a MF B1a well-sorted crinoid grainstone with syntaxial calcite overgrowth, AAA. b MF B2a; Aïn-as-Seffah type; clasts of laminar stromatoporoids (S) and Thamnopora (T) within a peloidal crinoid grainstone, AAS 10 m. c MF B2c, dendroid type; Thamnopora with dark radiaxial fibrous calcite cement (black arrow), inner spaces are filled with brighter calcite (white arrow), SMS, 1 m. d MF C2a, Phillipsastrea boundstone: Phillipsastrea (P) and tabulate coral (T) overgrow a laminar stromatoporoid (S), AAS, 20 m. e MF B3, Brachiopod-rugosa floatstone, field photograph of silicified slab with bivalved brachiopods (black arrows) and solitary rugose corals (white arrows), AAA. f MF C3, coral-stromatoporoid floatstone/rudstone (biostrome facies). h Weathered surface of silicified thamnoporid (T), alveolitid (A), and solitary rugose corals (R), AAA. g Patch reef facies (MF A5) containing of laminar stromatoporoid (S), thamnoporoid coral (T), solitary rugose corals (R), and small Amphipora branches, AKS, 56 m. h Fore reef breccia (MF B2a); small clast of an alveolitid coral (A) and laminar stromatoporoid (S), AAS, 8 m
b: reef breccia (Fig. 8a, g–h)

a Fore reef breccia (MF B2b); breccia with laminar stromatoporoids (S) and alveolitid corals (A) SMS, ca. 70 m. b Stromatoporoid coral boundstone (MF C2) consisting of bulbous stromatoporoids (S) and alveolitid corals (A), SMS, ca. 160 m. c Dendroid stromatoporoid coral float-/boundstone (MF B2c) consisting of thamnoporid corals (T) and dendroid stromatoporoids, SMS, ca. 150 m. d Upper conglomerate consisting also of unsorted debris flow deposits, SAL. e Isolated crinoid debris (white arrows) and branching tabulate corals (black arrows) floating between bigger clasts of crinoid grainstone, SAL. f Amphipora boundstone (MF A6), note the poor preservation of fine structures due to dolomitization, thin-section showing the stylolitic, teconically overprinted contact of branches, Jennabia. g Olistolith consisting of reef breccia (MF B2b) with bulbous stromatoporoids (black arrow), Phillipsastrea, and clasts of crinoid grainstones (white arrows) in a matrix consists of coarse crinoid fragments and smaller rugose and tabulate corals, Biar Settla Conglomerate. h Reef breccia olistolith (MF B2b) with numerous branching tabulate corals (black arrows) and various lithoclasts in a bioclastic matrix, Biar Settla Conglomerate
c: dendroid type (Figs. 7c, 8c)
Subtype a was found at Aïn-as-Seffah and is similar to MFT B1a. The color is the same but it contains an increasing content of stromatoporoids and corals (50 %) within a bioclastic crinoid pack-/grainstone matrix. In this subtype, the dominant reefal organisms are laminar to tabular stromatoporoids, colonial Rugosa (Phillipsastrea), and branching, rounded or laminar tabulate corals (Thamnopora, Pachyfavosites, Alveolites). These bioclasts are smaller than in the other subtypes and reach diameters of a maximum of 10 cm. Intercalated are small patches of in situ Alveolites colonies or laminar stromatoporoids (Fig. 7b). Additional bioclasts within the partially peloidal to arenitic or sparitic matrix are crinoid remains, small shell fragments of brachiopods, and dendroid bryozoans. As in MF B1, most crinoids show syntaxial calcite overgrowth.
Subtype b is represented by bigger and more numerous stromatoporoid and coral clasts. The color is light to middle gray. The bioclasts compose often more than 50 % of the limestones and build floatstones and rudstones. Most bioclasts are bulbous or tabular stromatoporoids, tabulate corals (Favosites, Alveolites), or colonial Rugosa (Phillipsastrea). Associated but uncommon are solitary rugose corals, thamnoporoids, and dendroid stromatoporoids (Stachyodes). The matrix consists of bioclastic grainstones (calcarenites) containing crinoid debris, brachiopod and bryozoan fragments, ostracods, and calcareous algae (Girvanella). As in subtype a, crinoids show syntaxial calcite overgrowth. Micrite is uncommon; instead spaces between components are mostly filled with sparite.
Subtype c is also light to middle gray. It is dominated by dendroid stromatoporoids (Stachyodes) and tabulate corals (Thamnopora), which compose more than 50 % of the framework. Less abundant, and not present in every sample, are solitary rugose corals, laminar tabulate corals, and stromatoporoids. Some dendroid stromatoporoids and corals are overgrown by laminar forms. Their branches often show filling and are surrounded by a dark, radiaxial fibrous calcite cement; interspaces are filled with micrite or sparite. This subtype is similar to the Stachyodes-Thamnopora floatstones (MF A4) of the lagoonal facies but it contains detrital crinoid remains and consists of more fragmented components. The amount of calcite cement is much higher.
In all three subtypes the inner space of corallites are often filled with geopetals and component contacts have stylolite seams and sutured grain contacts, giving a stylobreccioid fabric. Common fissures and veins are filled with calcite.
Interpretation and comparisons: Fore-reef slope adjacent to or intercalated with MF B1 and MF B3. It ranges from the distal fore reef slope (transition of FZ3–FZ4 sensu Wilson 1975) to the outer reef edge (FZ 5, Fig. 6). High amounts of large stromatoporoid and coral clasts derived from reef margins, which were prone to storms, tidal waves, and currents. This coarse detritus was shed into the debris of adjacent lower slope crinoid forests, which provided the bioclastic crinoid grainstones. The same facies occurs in German reefs (e.g., facies type C of Franke 1973, detrital stromatoporoid-echinoderm facies of Krebs 1974; Weller 1991). MF B2a represents detritus of the initial stage of a small-scale bioherm, whereas the bigger clasts of MF B2b suggest deposition on the proximal fore-reef flank adjacent to a reef core (see also facies type D of Franke 1973). MFT B2c occurs in combination with MFT 2b and reaches the reef edge, where it deposited adjacent to MF C2b, but it can also be found within back-reef settings, where it deposited in combination with patch reef facies (MF A5). Significant amounts of micrite within some samples indicate protected fore-reef environments, such as channels or depressions. This is also known from the Rhenish Massif and Harz Mountains (Franke 1973; Krebs 1974: fore-reef Stachyodes facies; Weller 1991). The stylolite seams and sutured grain contacts result from pressure solution during diagenesis. MF B2 corresponds to F2 and F4 sensu Cattaneo et al. (1993). After Flügel (2004) it equals SMF 6 with transitions to SMF 5.
Reef facies set (MF C)
MF C1 brachiopod-Rugosa floatstone
This dark gray limestone is rich in bioclastic components. The main components, often more than 50 % of the rock, are variably dominant solitary rugose corals and bivalved brachiopods. There are subordinate small tabulate corals (Alveolites), which indicate a transition to the adjacent MF C3. The matrix contains crinoids with syntaxial calcite overgrowth and small fragments of trilobites, small brachiopod shells, and some foraminifera (Nanicella). It represents a crinoid-dominated bioclastic pack-/grainstone with some lime mud. In some samples, it is completely dolomitized and shows an inequigranular fabric with non-rhombic crystals (Fig. 7e). Silicification led in some cases to excellent preservation of atrypid brachiopods on bedding surfaces.
Interpretation and comparisons: The neritic crinoid-solitary rugose corals-brachiopod association indicates deposition in the open-marine outer area of a biostrome (FZ 4 of Wilson 1975), possibly also on the gentle toe of slope of a low bioherm (Fig. 6). The lime mud and often-complete preservation of brachiopods, suggesting a calm deposition below the wave base. Nanicella has previously been recorded from the Moroccan Meseta (Mrirt region), where it occurs in Givetian/Frasnian open-marine crinoidal to biostromal brachiopod-stromatoporoid-coral limestones (Termier et al. 1975).
MF C2 coral stromatoporoid boundstone
Three subtypes:
a: Phillipsastrea boundstone
b: Thamnopora boundstone
c: Alveolites-stromatoporoid boundstone
This is a variable facies type of light to middle grey limestones. The main builders are colonial rugose corals (Phillipsastrea) and stromatoporoids in association with Tabulata (Alveolites, Favosites). These build almost 70 % of the rock and colonize in layers, and overgrow or encrust each other. Phillipsastrea boundstones (MF C2a) have niches settled by dendroid tabulate corals (Thamnopora), dendroid stromatoporoids (Stachyodes), and solitary rugose corals. The second subtype with very dominant thamnoporids in growth position gives Thamnopora boundstones (MF C2b). The third subtype is built by patches of Alveolites and laminar stromatoporoids, whereas Phillipsastrea and Thamnopora are secondary. Arenitic detritus, composed of shell fragments and crinoid debris (similar to MF B2a), accumulated between the coral framework. The matrix consists of micrite, mud peloids, or sparry calcite. The corallite lumina and open-space structures are often filled with geopetals. Stylolites form many component contacts. In some samples, dolomitization proceeded along stylolites.
Interpretation and comparisons: Biostromes or low-relief bioherms on a neritic carbonate platform (FZ 4 sensu Wilson 1975). Productive conodont samples prove an influx of open-marine biota (data of ZS Aboussalam). The laminar to tabular stromatoporoids and colonial corals constructed a rigid, potentially wave resistant, flat framework from which some detritus was shed. In protected niches and interspaces, micrite accumulated, and more delicate, branching tabulate corals found a habitat. Constantly agitated water transported crinoid detritus from the flanks (MF B1 and B2a) into such niches. The Thamnopora boundstone must have formed in a lower hydrodynamic regime. It falls within F5 of (Cattaneo et al. 1993) and SMF 7 of Flügel (2004). Krebs (1974) distinguished in Rhenish reefs two dominant core facies: “massive stromatoporoid biolithe”, consisting of large bulbous stromatoporoids, and a “tabular stromatoporoid facies”. Both are not preserved in the studied outcrops. Phillipsastreids occur in Givetian Rhenish sections rather occasionally in proximal margin slope debrites (Errenst 1993) but they were more significant tabular constructers in the last phase of the Frasnian Iberg Reef of the Harz Mountains (Franke 1973: facies type I). In the Frasnian of the Ardennes, they form biostromes within the fair weather wave base, above alveolitid-stromatoporoid biostromes (Poty and Chevalier 2007).
MF C3 coral-stromatoporoid floatstone/rudstone (biostrome facies)
This facies type was found at Aïn-Al-Aliliga as allochthonous lithoclasts. It consists of dark grey to black limestones with solitary rugose corals, tabulate corals, and small, laminar stromatoporoids. Solitary rugose corals and Thamnopora are the main components, followed by alveolitids and laminar stromatoporoids. The matrix is bioclastic pack-/grainstone, similar to MF C1. However, there is less crinoid debris and trilobites are present. Some samples are strongly dolomitized; in other cases, corals are silicified and weather out well (Fig. 7f).
Interpretation and comparisons: The fragmented or dislocated faunal assemblage with mixed reefal (corals, stromatoporoids) and open-marine neritic groups (crinoids, trilobites) suggest deposition on an open, biostromal carbonate platform above the storm wave base, adjacent to but shallower than MF C1. Trilobites preferred quiet and calm habitats, without strong current activity but frequently they also occur disarticulated in crinoidal limestones. MF C3 shows similarities with MF B2. It matches mostly F2 of Cattaneo et al. (1993). In the Rhenish Massive, it belongs to the “stromatoporoid-coral-brachiopod facies” of biostromes within the “Schwelm facies” (Krebs 1974). Similar initial reef facies, but with more algae and cyanobacteria, occur widely in the Eifel Mountains and Sauerland (Malmsheimer et al. 1996).
Local Oued Cherrat reef developments
Aïn Khira South
The Aïn Khira South section of the northern Oued Cherrat Zone exposes between an active quarry margin and adjacent forest almost 100 m of reefal limestone of the (new) Aïn Khira Formation (Fig. 2, GPS coordinates N33°42′56.9′′ W007°01′12.8′′). Conodont dating of the succession was unsuccessful but the index brachiopod Stringocephalus (MF A3 and MF A7) from 12 and 59 m above base proves a lower to middle Givetian age (Figs. 2, 3, 5, 7g, 9).

Field facies log and microfacies succession of the reefal Aïn Khira Formation at Aïn Khira South
Supported by the lack of any conodonts, which are long known to have avoided back reef settings, the whole outcrop represents a protected platform interior with characteristic lagoonal facies types; fore reef and reef core facies are missing. At the base, the limestone consists of middle to dark gray peloidal grainstones (MF A1) containing gastropods and brachiopods (Fig. 5a–c). At 7 m, there are intercalations of unfossiliferous, late diagenetic dolomite banks (15-30 cm in thickness) followed by recurring MF A1. This is a typical facies of restricted lagoonal settings with moderate water circulation. At 9 m begins a short interval (2 m) with bioclastic grainstone (MF A2) containing fragmented stromatoporoids, corals, and crinoids. Subordinate back-reef crinoid accumulations are also known from German reefs (Krebs 1974; Faber 1980). We suspect that crinoid debris was brought in from the reef edge by strong current and storm activity and through channels. This interval indicates an open-marine input during episodic transgression or proximity to the fore reef. An Amphipora boundstone (MF A6) at 11 m documents the return to a restricted quiet and shallow lagoonal setting. It is overlain by a characteristic storm event layer (MF A3: Stringocephalus-Amphipora float-/rudstone, at 11.5–15), with dolomite in the next meter. Above, an interval of peloidal grainstone (MF A1, 16–20 m) returns, which is overlain by Stachyodes-Thamnopora float-/rudstones (MF A4, 20–23) with a transition to even more restricted, deeper conditions, indicated by subsequent bioclastic mud-/wackestones at 23 m (MF A7). Their very poor fossil record is visible in thin-sections (ostracodes, gastropods, parathuramminids). Associated and intercalated are Amphipora boundstones (MF A6) at 44–46 m and a patch reef facies (MF A4 and MF A5, Fig. 7g) at 48–59 m, consisting of small bulbous and laminar stromatoporoids, solitary rugose, and tabulate corals. Dendroid stromatoporoids and corals settled between them. At 59 m, an interval of Stringocephalus floatstones (MF A8; Fig. 5i) starts. These are also known from the Rhenish Massive, where they occur within the protected wide, shallow platform, but the massive brachiopod shells were often fragmented during storm events. Within this succession, a thin-bedded Amphipora boundstone (MF A6) occurs at 64–65 m. At 66 m, the Stringocephalus floatstone interval is overlain by 2 m of patch reef facies (MF A5). Above, an interval (27 m) of alternating Amphipora boundstone (A6, at 70), Stachyodes-Thamnopora float-/rudstones (MF A4, at 75, and 80 m), and bioclastic mud-/wackestone returns, which indicates an episodic deepening, with calm and restricted water conditions. At the top of the section, at 95 m, Amphipora boundstones give evidence for a persisting lagoonal setting, with some evidence for shallowing upward.
Unlike as in contemporaneous German (Schudack 1993) or Australian (Brownlaw et al. 1996) reef platforms, there is no evidence for distinctive cyclicity at Aïn Khira South. In agreement with Cattaneo et al. (1993), two back-reef deepening phases can be recognized, separated by a patch reef phase that can alternatively be interpreted as a shift towards an expanding inner reef margin. Our section does not include at the boundary of both “sequences” a shallowing-upwards interval of birdseye (fenestral) limestone and microbial laminites (F8 of Cattaneo et al. 1993). Unfortunately, our section exposes neither the initial stage nor the final reef phase and extinction level. The previous, now destroyed (quarried) outcrop contained a true reefal succession with massive stromatoporoids near the top of the succession, overlain by thin wacke- and floatstones. Currently, we are unable to identify the reason for the extinction of the main Givetian reef belt of the Oued Cherrat Zone.
Aïn-Al-Aliliga
Aïn-Al-Aliliga (Aïn Aliliga) is a ca. W-E-running secondary valley located between Aïn Khira South in the north and Aïn-as-Seffah in the south, running into the Oued Cherrat (Fig. 2, GPS coordinates N33°37′16.3′′ W007°00′48.9′′). A thick, hardly studied Emsian reef (Cakhrat-ach-Chleh Member of Dhar-es-Smene Formation) overlies phacopid-rich neritic limestones that are exposed at the western and eastern ends of a reef hill (GPS N33°37′17.0′′ W007°01′09.2′′). As shown in the cross section of Chalouan (1981), the Eifelian is locally missing in the succession of the main hill (right part of column in Fig. 3). Due to folding, the trilobite limestones re-appear on the eastern slope and are faulted against the Givetian. In the lower part of the latter, bedded reefal limestones alternate with strongly brecciated units (debris-flow and slump deposits) and a few thin, crinoidal limestones. All yielded upper Givetian conodonts. Sampled individual reef and breccia blocks yielded variably lower/middle Givetian or mixed middle/upper Givetian conodonts (data of ZS Aboussalam). This suggests the local synsedimentary destruction of an originally long-lasting (all of the Givetian) reef interval. At the top, the conglomerate/breccia becomes polymictic, with up-to- 3-m-large clasts of Ordovician quartzite and volcanites in thick debris-flow beds. These are well exposed near the local spring. Closely adjacent to the current outcrop, uplift-related continuing erosion must have penetrated eventually the complete Devonian succession and reached the Lower Paleozoic. Conodonts from this part of the breccia and from its top contain, again, a diverse, top-Givetian conodont assemblage. Overlying, yellowish, thin-bedded dolomites yielded only belodellids, which prove a post-reefal/post-reworking calm, very condensed and very shallow sedimentation that was no younger than the Frasnian. Contrary to conodont data of D. Kergomard (quoted in Chalouan 1981), we found no evidence that the type Aïn-Al-Aliliga Formation contains any Famennian conodonts (Figs. 2, 3, 7a, e–f).
Lithoclasts range in diameter from a few centimeters to several meters. Typical are well-sorted crinoid grainstones (MF B1a, Fig. 7a) containing large, slumped clasts of favositids and colonial rugose corals, such as Phillipsastra. In addition, there are brachiopod-Rugosa floatstones (MF C1) with silicified brachiopods as well as solitary rugose corals. Very similar in color and variably silicified or dolomitized are float-/boundstones with weathered-out reef organisms. These clasts belong to the MF C3 biostrome facies and contain solitary rugose corals, tabulate corals (Thamnopora and Alveolites), and laminar stromatoporoids. All reef builders, except Thamnopora, form flat and laminar patches. The matrix is a bioclastic pack-/grainstone consisting of crinoid debris, brachiopod shells, and foraminifera. An additional characteristic and dominant facies type is a reef breccia (MF B2b) with colonies of Heliolites, Alveolites, and big bulbous and laminar stromatoporoids (up to more than 20 cm in diameter). Interspaces include small clasts of Thamnopora. Other important facies types are Stachyodes-Thamnopora floatstones of the back reef environment (MF A4) and adjacent mudstones with few gastropods (MF A7). The recognized biofacies array suggests an original reef zonation developing on or adjacent to a neritic brachiopod-rich carbonate bank. The coarse fore reef breccia (MF B2b) and the big bulbous stromatoporoid clasts suggest that a reef core existed originally near-by. Lagoonal depositions with dendroid stromatoporoids and corals confirm this assumption.
A second conglomeratic unit with some isolated, twice reworked reefal limestone and Pragian “griotte” limestones occurs ca. 500 m eastwards on the slope of the main Oued Cherrat within the Al Brijat Formation. This level corresponds to the Aïn Dakhla olistoliths.
Aïn-as-Seffah
The isolated Givetian “reef” of Aïn-as-Seffah (Fig. 2) in the southern Oued Cherrat Zone (GPS coordinates N33°34′30.8′′ W006°59′18.7′′) is constituted by more than 40 m of massive limestone, named by Chalouan and Hollard (1979) as Aïn-as-Seffah Formation. Its conodont stratigraphy has been studied by Aboussalam et al. (2013), who introduced a member subdivision. Based on new data, the “reef” base (base of Member 2) falls in the Schmidtognathus hermanni Zone, which begins at the middle/upper Givetian boundary. Below and laterally it ends abruptly at a succession of older (lower Givetian) thin-bedded marls and flaser limestones (Member 1). At ca. 10 m above the base, opposite to higher limestones of Member 1, conodont fauna prove a slightly younger upper Givetian age (Po. cristatus ectypus Zone). On the southern flank, the “reef” grades into lower Frasnian brachiopod limestone (Member 3, MN 1–4 Zone, Ancyrodella rotundiloba pristina—Palmatolepis transitans Zones), followed disconformably by upper Frasnian flaser limestone (Member 4, MN 10 Zone) and a distinctive, upper Frasnian, partly slumped Manticoceras Bed (Member 5, basal MN 13 Zone). In the center, above the marker tree, the reef growth also ends with the upper Givetian (Skeletognathus norrisi Zone) and the top is covered by top middle to upper Frasnian flaser limestones (higher Member 4, MN 10, and 12 Zones). These lie morphologically higher in the outcrop than lateral, supposedly lower Famennian red shales, which form a locally preserved Member 5 below the silty/sandy Al Brijat Formation. The whole complex seems to represent a big olistolith. Strong tectonization resulted in stylolites, solution seams, elongated components within the limestones, and many deformed conodonts. However, the internal fabric and stratification is still intact. In comparison with other reefal limestones of the Meseta, the Aïn-as-Seffah “reef” represents a small, isolated bioherm/patch reef that developed in the upper Givetian on a deeper neritic, argillaceous carbonate platform (Member 1). A distinctive zonation with fore reef, reef core, and back reef never developed (Figs. 2, 3, 7b–d, h, 10).

Lithostratigraphy, field facies logging, and microfacies succession of Member 2 of the Aïn-as-Seffah Formation at Aïn-as-Seffah
At the base, Member 2 consists of crinoid grainstone (MF B1a) with minor detritus of biostromal organisms (a few small fragments of stromatoporoids and Alveolites). One meter above base, fragments become larger and more diverse, which suggests initial biostrome growth (or a “parabiostrome”, see Álvaro et al. 2007). Especially the coral fauna increases and the grainstone facies changes to detrital coral-stromatoporoid float-/rudstone (MF B2a: 1 m). Typical are floatstones with fragments of Thamnopora, Alveolites and, especially between 8 and 16 m, solitary Rugosa; laminar stromatoporoids occur at 4 and 8 m (Fig. 7h). Intercalated are small in situ patches of laminar stromatoporoids at 17–18 m. These forms stabilized the detritus-rich float-/rudstones and mark the transition to coral-stromatoporoid boundstones (MF C2a and MF C2b). Phillipsastrea boundstones appear 19 m above the base and are overlain by an interval of detrital coral-stromatoporoid float-/rudstone (MF B2a 22–28 m). Intercalated is a crinoid grainstone at 24–26 m, which marks a relative deepening. At 28 m, coral-stromatoporoid boundstones reappear, either with Phillipsastrea (MF C2a) or Thamnopora and Stachyodes (MF C2b: Thamnopora boundstones). A strongly brecciated coral-stromatoporoid float-/rudstone at 37 m suggests a deepening phase. This assumption is supported by a bioclastic crinoid-brachiopod grainstone at 42 m, which indicates a rapid drowning of the reef. Laterally, around the marker tree, the reef growth continued. The brecciated interval ends at 46 m and is overlain by Philipsastrea boundstones (MF C2a). This facies reaches the top of the succession at 52 m.
The succession indicates that a laminar pioneer fauna settled on a crinoidal bank. It could first keep up with subsidence but eventually it drowned and was overlain by detritus-rich floatstones. This interval is represented by MF B2a and MF C2b. The intervals of coral stromatoporoid boundstone (MF C2) indicate three shallowing phases, which are intercalated by detrital coral-stromatoporoid float-/rudstones (MF B2). The laterally overlying, basal Frasnian crinoid brachiopod floatstones (MFT B1b) suggest final extinction by drowning during the main transgressive phase of the global Frasnes Event (Aboussalam et al. 2013). The central reef top ends with a correlative drowning unconformity, followed by a long episode (MN 1–11 Zones) of non-deposition. The persisting conodont faunas throughout the reefal succession prove permanent open-marine conditions.
Sidi Mohamed Smaine
The lower middle Givetian (Po. varcus Zone, Benfrika and Bultynck 2003) Sidi Mohamed Smaine Formation is located in the northern Al Attamna area close (west) to a cemetery and small mosque (Fig. 4, GPS coordinates N33°29′21.5′′ W006°59′56.5′′). It consists of more than 170 m of light gray limestone beds with reefal organisms in most parts. The occurring facies types are similar to AAS and AAA. The rather continuous presence of conodonts shows that the whole succession was of open-marine type (Figs. 3, 4, 8a–c, 11).

Field facies logging of the upper part of the Sidi Mohamed Smaine succession (Al Attamna region)
The base of our section is marked by several ca. 50-cm-thick Stachyodes-Thamnopora Floatstones (MF B2c), which may represent the storm-generated debris of a biostrome. This interpretation is in accord with its belodellid conodont biofacies. Reef growth kept up with subsidence since the same facies developed in an episodically exposed interval of at least 40 m (base of Fig. 11; Unit A of Benfrika and Bultynck 2003). Above follows a ca. 30-m interval of bioclastic crinoid grainstone (MF B1a), which reflects biostrome retrogradation and local deepening (lower part of Unit B in (Benfrika and Bultynck 2003), with an onset of polygnathids). With relative sea-level fall and progradation, this facies turns into a thick fore-reef breccia (MF B2b) dominated by laminar stromatoporoids and Thamnopora within a bioclastic grainstone matrix (Fig. 8a). Brachiopods occur occasionally in this reef margin succession. At 80 m above base, dendroid builders decrease while laminar stromatoporoids and corals increase within breccia beds. This is an interval of almost 15 m, then swinging back into dendroid stromatoporoid-coral floatstones that resemble the lower phase. At 100 m above base, the floatstones are covered by 2 m of bioclastic crinoid grainstone (MF B1a) with brachiopods. This short interval of maximum flooding is overlain by massive, 1-3-m-thick stromatoporoid-coral float-/rud-/boundstones (MF B2b), which mark a second reef progradation episode (base of Unit C of Benfrika and Bultynck 2003). Large reefal bioclasts, mainly alveolitids and laminar or bulbous stromatoporoids, increase (Fig. 8b). The matrix becomes more micritic, crinoid and other debris becomes less important. Additionally, there are some solitary rugose corals. The dominance of proximal reefal blocks reaches to the top of the succession: In the upper part, at 150 m, there are intercalated less massive banks (max. 1 m thick) of dendroid stromatoporoid-coral float-/boundstones (Fig. 8c), consisting of Thamnopora and Stachyodes, sometimes encrusted by stromatoporoid layers. The last 10 m of the succession is partly dolomitized.
The section top is a reddish polymictic “conglomerate” (here assigned as unit D). It represents a coarse, proximal debris flow with mixed components, such as large clasts of tabulate corals (Alveolites, up to 20 cm), solitary and colonial rugose corals (Phillipsastrea, up to 20 cm), bulbous and laminar stromatoporoids, some thamnoporids, rare brachiopods, reworked goethite crusts, and some platy slate slabs. The matrix is a crinoid grainstone (MFT B1a). Unlike at Aïn-Al-Aliliga, there are no quartzites or volcanic clasts.
In summary, the higher Sidi Mohamed Smaine Formation (Unit C) consists primarily of large reef breccias (floatstones and rudstones), intercalated especially in the upper part with some boundstones. These and the grainstone matrix suggest a turbulent, current, and wave-influenced upper fore reef slope reaching into the reef edge. Support comes from the presence of polygnathid conodont faunas (Benfrika and Bultynck 2003). The reef may have had a low relief. The boundstones contain micritic matrix, which indicates short-term decreased water energy or deposition in niches or channels. This pattern is well known from the Rhenish massive (Krebs 1974). The local reef extinction is difficult to determine since the autochthonous sequence was cut off after deposition by a violent, erosive mass flow event. It is not likely that tectonic uplift ended the local reef growth since there are no cements that indicate subaerial exposure.
Sidi Ahmed Lemdoun
Sidi Ahmed Lemdoun is located ca. 1 km south of Sidi Mohamed Smaine directly east (downhill) of the main road to the south (P 3333, Fig. 4, GPS coordinate N33°28′57.8′′ W006°59′05.5′′). Reefal organisms occur only as reworked components in the variably 6–12-m-thick unit D of the Sidi Ahmed Lemdoun Formation (Benfrika and Bultynck 2003). There are two massive conglomerate units separated by marls and thin-bedded, nodular crinoidal limestone (Fig. 3). The lower conglomerate (Bed 13, 30–50 cm thick) has partially a reddish matrix and conodonts of the basal middle Givetian varcus Zone. It correlates clearly with our unit D of Sidi Mohamed Smaine, which is supported by the conodont dating. The upper conglomerate (Bed 23, ca. 1.8 m, laterally = Bed 23′, up to 8 m) consists of massive boulders and mostly yielded middle Givetian conodonts (ansatus Zone, data of ZS Aboussalam) but at the base also a single middle Famennian Palmatolepis. This proves that a first reworking and re-sedimentation took place in the Givetian, a second much later in the Famennian. Unit D is abruptly terminated by the silty-sandy, unfossiliferous Al Brijat Formation (Figs. 3, 4, 8d–e).
The conglomerates consist of rounded to well-rounded limestone clasts, which range in diameter from 0.5 to 20 cm. The lower conglomerate contains lithoclasts of a Peloidal Grainstone (MF A1a) with small ostracods and parathuramminids; some of these clasts are partly dolomitized. Associated are isolated stromatoporoids and corals (Fig. 8d–e). Dominant are bulbous stromatoporoids up to 15 cm in diameter, tabulate (Alveolites and Favosites up to 10–15 cm in diameter), and small solitary rugose corals. The upper conglomerate differs and contains small branches of Thamnopora and stromatoporoid or coral clasts reach a diameter of 10 cm. The matrix consists in both intervals of crinoid debris within bioclastic mud-/wackestones. The reefal material derived both from fore reef/reef core and internal platform (MF A1a) environments of the Sidi Mohamed Smaine bioherm to the west. Re-deposition of the mixed remnants took place late in the middle Givetian as channeled debris flows on a deeper neritic slope. This explains the laterally highly variable thicknesses and the rather small amount of material.
Jennabia/Biar Setla
The Jebel Jennabia lies in the northern Khatouat region southeast of the Oued Cherrat Zone (Fig. 4, GPS N33°25′25.7′′ W006°48′37.6′′), 16 km ESE of Sidi Ahmed Lemdoun and ca. 3 km SE of Al-Haddada. It exposes a thick succession of vertically bedded, bluish, solid to massive limestones that still lack a formation name. Unfortunately, its reefal fauna is very poorly preserved; all limestones are strongly dolomitized and lost most fine structures. Recognizable reef builders are thamnoporids, solitary rugose corals, and dendroid stromatoporoids. Better preserved is an Amphipora boundstone (MF A6; Fig. 8f), typical of a back reef environment. It seems that the succession represents a strongly tectonized and altered inner platform, which explains why conodont samples were barren. The limestone is separated by a mineralized fault zone from quartzite of the probably Famennian Jennabia-Babot Member of the Fouzir Formation (Fadli 1994), which crops out near the crest of the adjacent steep northern slope (Figs. 3, 4, 8f–h).
On the steep southern slope of the next hill, reefal fauna occurs within isolated blocks of the Biar Settla Conglomerate, the middle member of the Fouzir Formation (see geological cross section in Fadli 1994: Fig. 6). GPS coordinates are N33°25′44.2′′ W006°48′17.3′′. The up-to-3-m-thick allochthonous limestone blocks consist of stromatoporoid coral float-/rudstones (MF B2b) and are developed as massive reef breccias (Fig. 8g–h). They resemble the olistoliths of Aïn Dakhla, which include twice re-sedimented reefal clasts of different provenance. Well after the first brecciation and mixing of clasts, the second gravitative embedding as olistoliths within a basinal shale setting took place in the middle Famennian (see review of regional biostratigraphy in Kaiser et al. 2007).
The breccias contain bulbous stromatoporoids, colonial Rugosa (Phillipsastrea), and various tabulate corals (Heliolites, alveolitids, thamnoporoids). Phillipsastrea colonies reach 45 cm in diameter and confirm an original Givetian age. Other reefal bioclast reach 20 cm in size (Fig. 8g). Secondary components are variable lithoclasts of micrite mudstones and crinoid grainstones (MFT B1a). The matrix consists of smaller clasts of fragmented dendroid tabulate corals (Thamnopora, Fig. 8g), dendroid stromatoporoids, and solitary rugose corals, often overgrown by stromatoporoids and alveolitids, within a peloidal crinoid grainstone.
The rather variable reef clasts but without any typical lagoonal facies types suggest that the initial reworking mostly affected a biostromal to biohermal marginal slope succession. This contrasts with the poorly preserved autochthonous platform facies of Jennabia nearby, which probably represents the back reef platform of the same complex.
Comparison with contemporaneous reefs of the western Prototethys
The described microfacies sets gave numerous references to occurrences of identical facies types in reefs of the Rhenohercynian Zone on the southern Laurussian shelf. A full comparison should await the description of other Meseta Givetian reefs limestones but the overall similarity is striking and in contrast to the wide paleolatitudinal separation at the southern margin of the (sub)tropics. A plate tectonic reconstruction with the position of Rhenish, Meseta, and cratonic Gondwana reefs (Western Sahara, Anti-Atlas, Ahnet) is given in Fig. 12. The lack of some reef facies in the Meseta, such as dasycladacean-, codiacean- and cyanobacteria-rich limestones, is probably meaningful and could reflect water temperature differences, but the similarity favors an only moderately wide Prototethys Ocean at the time and suggests a rather low paleoclimatic gradient from ca. 15/20° to 35/40° S in the Givetian. This is surprising because the oxygen isotope data of Joachimski et al. (2009) suggested that the lower/middle Givetian was the coolest period of all of the Devonian, which should have promoted a latitudinal (N–S) ecological zonation. Unfortunately, there are so far no conodont-based Givetian water temperature calculations for Morocco or across a Prototethys/Rheic Ocean transect. Joachimski et al. (2004) mentioned typical tropical seawater temperatures calculated from Anti-Atlas brachiopod calcite but such isotope data are thought to be less reliable than conodont phosphate values. The still-limited taxonomic knowledge of Givetian/Frasnian reef builders (tabulate and colonial rugose corals) and dwellers (shallow-water solitary rugose corals) of the Anti-Atlas gives strong paleobiogeographic links with crustal units north of the Prototethys (northern Spain, Armorican Massive) and, beyond, to the Ardennes-Rhenish Massive and even to Russian regions (Urals, Kuznetsk Basin) taxa (Pedder 1999; Schröder and Kazmierczak 1999; Coen-Aubert 2002, 2005, 2013; May 2008). At the species level, the southern Moroccan (cratonic Gondwana) rugose coral faunas are mixtures of endemic and European taxa, including a range of species that are well known from the Rhenish Shelf. This probably expresses the combined effect of ecological differences and biogeographic barriers. However, there are so far no endemic taxa among the main reef builders (stromatoporoids and tabulate corals, e.g., Aboussalam 2003; May 2008) and, therefore, no evidence that the same reef facies of northern Gondwana and the Rhenish Shelf were regionally built by different taxa, but the database is very small for the Anti-Atlas/Western Sahara and even more unreliable for the Meseta.

Position of the reefs of the Rhenish Massive, Western Sahara, Anti-Atlas (Maider, Tafilalt) and southern Algeria in a plate tectonic reconstruction with a moderately wide “Variscan Sea” (Prototethys to southern Laurussian shelf) and without significant lateral (paleolongitudinal) displacement of northern Gondwana (altered from Dopieralska et al. 2015; Fig. 1)
The lack of clear reef cyclicity, as prominent in Givetia/Frasnian reefs of the Rhenish Massive (Schudack 1993), Ardennes (e.g., Preat 1989, Boulvain et al. 1995), Moravia (Hladil 1986), Canada (McLean and Mountjoy 1994), South China (Chen et al. 2001, 2002), and Australia (Brownlaw et al. 1996), all from lower latitudes than Morocco, may indicate a position of the Meseta reefs outside of orbitally controlled sedimentation, for example outside significant monsoonal shifts.
Baidder et al. (2008) confirmed that the Meseta formed the Variscan foreland adjacent to the stable cratonic Anti-Atlas region. A comparison of Meseta reefs with contemporaneous reefs of the Tindouf Basin and eastern Anti-Atlas has to consider their different, more stable synsedimentary tectonic setting and an originally larger spatial separation. Gomez et al. (2000) calculated a post-Paleozoic crustal shortening for the dividing Atlas chains in the scale of 17–45 %.
Western Sahara
The spectacular reefs of the western Tindouf Basin and Mauritanian Zemmour were described by Dumestre and Illing (1967), Wendt and Kaufmann (2006), and Ernst and Königshof (2008, 2010). During the Givetian, six reef cycles separated by neritic limestones and marls were distinguished. The first reef cycle is dominated by mud mounds with micritic boundstones (Stromatactis), auloporids (incrusting tabular tabulates), and microbial mats. The second cycle is only weekly developed and lies directly on the second limestone. The most spectacular reefs occur in the third cycle and consist of tabular reef bodies, built of bioclastic grainstones with crinoids, brachiopods, stromatoporoids, and trilobite remains. Its biostrome builders are bulbous and tabular stromatoporoids, tabulate and rugose corals. The fourth reef cycle overlies oolitic grainstones and is characterized by a stromatoporoid biostrome (bulbous and tabular), colonial rugose (Phillipsastrea), solitary rugose and tabulate corals. The fifth reef cycle consists of a coquina with corals, brachiopods, and bryozoans as well as patch reefs. The sixth reef cycle is a large coral patch reef or biostrome with rugose corals, Alveolites, Coenites, and Aulopora; stromatoporoids are less common and very large rugose corals, Mesophyllum, characterize the base (Königshof and Kershaw 2006), as in the initial stadium of some Rhenish reefs (Becker et al. 2015b). The lack of microfacies studies prevents a detailed comparison with the Oued Cherrat reefs but the cyclic nature of biostromal growth is distinctive in the western Tindouf Basin, also the significance of calcimicrobes, bryozoans, and auloporids. Mud mounds (first reef cycle) are lacking completely in the Oued Cherrat area but the faunal composition of the third and fourth reef cycle resemble Aïn-as-Seffah, Aïn-Al-Aliliga, and the middle to upper part of Sidi Mohamed Smaine. Corresponding facies types are MF B1b, MF B2a, and MF C2a. Conspicuous is the missing of delicate, dendroid stromatoporoids (e.g., Amphipora or Stachyodes) in the Western Sahara reefs. This shows the absence of protected and calm environments behind a barrier, which may reflect a less strongly oscillating relative sea level (Wendt and Kaufmann 2006). In the Meseta, reefs had to keep up with a stronger subsidence.
Maider
Following initial data by Hollard (1974), Schröder and Kazmierczak (1999) described the fringing reef of Ouihlane at the northern margin of the Maider (eastern Anti Atlas). It consists of a brecciated biostrome with tabulate and rugose corals and some stromatoporoids; the reef growth ended with a “coralgal” limestone. Its destruction and re-sedimentation in polymictic mass flows was caused by synsedimentary tectonism (block faulting). The same mass flow succession extends on strike west- (Boultam South) and eastwards (Tahrerhat, Stichling 2013). A notable feature, which resembles the biostromes of the western Tindouf Basin, is the abundance of partly large heliolitids. There are also marl units (soft-bottom pioneer communities) with many isolated thamnoporids and favositids. At the Jebel Rheris 10 km to the East, Fröhlich (2003) investigated Givetian coral-stromatoporoid biostromes that alternate with crinoidal grainstone facies. They were mostly built by alveolitids, followed in abundance by phillipsastreids, favositids, solitary Rugosa, and heliolitids.
Towards the SW, a several-hundred-meters-large biostrome at Taboumakhlouf was first described by Hollard (1974) and later investigated by Nübel and Becker (2004). It is by far dominated by beautifully preserved, very large-sized, flat phillipsastreid colonies, which often sit isolated in marl, creating very low cryptic habitats occupied by micromorphic polychaetes (Spirorbis). Thamnoporids, brachiopods, and scutelluid trilobites are accessory while alveolitids, which were obviously well adapted to muddy substrate, dominate the local initial stadium. The dominance of phillipsastreids is stronger than at any of the Meseta biostromes but a similar facies is known from northern Spain (Méndez-Bedia et al. 1994).
Kaufmann (1998) investigated biostromes and mud-mounds of the southern Maider. He established a carbonate ramp model with five facies zones: shallow-ramp facies (stromatoporoid-coral-cyanobacteria boundstones) built by domal stromatoporoids, solitary rugose and tabulate corals (thamnoporids, alveolitids, heliolitids, favositids, and auloporids), mid-ramp facies (burrowed, skeletal wackestones), outer-ramp facies (burrowed, argillaceous skeletal wackestones), transitional outer-ramp to basinal facies (blue-grey, laminated mudstones), and basinal facies (monotonous, poorly fossiliferous shales). The shallow-ramp facies differs from Meseta reef types by its significant calcimicrobial content.
Döring and Kazmierczak (2001) described Givetian biostromes from the southwestern Maider. These were built by float-/boundstones and consist mainly of stromatoporoids, rugose corals, favositids, heliolitids, and crinoids. This biofacies strongly resembles the northern Maider.
Rytina et al. (2013) and Rytina (2013) investigated Givetian reefal lithoclasts re-deposited in Lower Carboniferous olistostromes in the Tinerhir region (NW Maider). They were transported northwards from the uplifted Ougnate block between the Maider and southern Atlas margin. Common are coral floatstones with Thamnopora, Alveolites, Phillipsastrea, and solitary Rugosa. Conodonts prove an open setting for Upper Givetian biostromes, supported by other blocks with fragmented crinoids, brachiopods, and gastropods. A second locally common facies type is a coral float-/rudstone with stromatoporoids and significant content of crinoids.
The so-far-recorded reef facies types of the Maider region are rather similar to the Western Sahara types. Many biostromes are dominated by a flat to bulbous pioneer fauna, which allowed the settlement in an agitated environment, for example on a lower ramp. This resembles Aïn-as-Seffah reef (MF C2a) and Aïn-Al-Aliliga (MF C3) but cyanobacteria and algae are not represented within the Oued Cherrat facies sets.
Tafilalt
From the Amessoui Syncline in the southern Tafilalt, Massa (1965) and Hollard (1974) described very small sized patch reefs, which represent biostromes to meter-scale mini-bioherms. Coen-Aubert (2002, 2005) described the coral taxonomy of this area. Aboussalam (2003), Aboussalam and Becker (2011), and Becker et al. (2013) provided more recent data. Locally dominant are tabulate corals (Favosites, Alveolites, Thamnopora, Heliolites), colonial Rugosa (Phillipsastrea), and rather large solitary Rugosa, while stromatoporoids and bryozoans are subordinate. There are mostly reworked, tempestitic coral float- and rudstones. Small biostrome or bioherms may consist of alveolitid and stromatoporoid bindstones. To the south, there was a more extensive but poorly studied carbonate platform with only subordinate tabulate and rugose corals.
Töneböhn (1991) described from the eastern end of the Hamar Laghdad region (eastern Tafilalt) the Givetian “Hollard Mound”, characterized by 13 facies types/subtypes ranging from hemipelagic platform to reef mound facies. The latter include beds with Favosites or Syringopora (type 7), “Rasenriffe” consisting of branching rugose corals (Amplexocariina; type 9), float-/rudstones with bioclastic debris (type 10), mud-/floatstones with rugose and tabulate corals, crinoids, and brachiopods (types 12 and 13), and Stromatactis mudstones (type 11). Töneböhn’s types 10, 12, and 13 resemble the fore reef facies set of the Meseta (MF B). Apart from the microbialites (type 11), the Amplexocariina patch reefs have not yet been found in the Meseta.
In summary, the Tafilalt reefs consist either of very small patch reefs (biostromes to mini- bioherms) or mud mounds, which both are not known from the Oued Cherrat Zone. Distinctive are also the relatively large solitary rugose corals, the more common heliolitids, which are tolerant to muddy substrate, and the absence of lagoonal facies.
Southern Algeria
Spectacular Middle Devonian reefs of the Ahnet (southern Algeria) are mud mounds, mud ridges, and mud atolls (Wendt et al. 1997). They are constructed by Stromatactis-bearing boundstones. The accessory reefal fauna consists of tiny tabulates (Thamnopora, Auloporacea) and small solitary rugose corals. Conspicuous is the lack of dendroid stromatoporoids; other faunal content are brachiopods, mollusk debris, trilobites, and sponge spicules. Similar mounds do not occur in the Meseta.
Conclusions
During the Givetian, the tectonically very active northern shelf of Gondwana (Moroccan Meseta) included many shallow structural highs, for example the Oued Cherrat Zone, its southern prolongation, the Al Attamna region, and parts of the Khatouat Massif. On these, stromatoporoids and tabulate corals developed small biostromes up to large bioherm complexes with a distinct zonation in fore reef, reef, and back reef. Due to the widespread synsedimentary Givetian tectonics (Becker et al. 2015a, compare Fig. 3), each reef shows individual developments with episodes of deepening and shallowing that are not correlated and without clear cyclicity.
The Aïn Khira Formation section represents a wide, protected platform interior (Fig. 6) with ecologically diverse back reef (lagoonal) facies types that fluctuate episodically with minor relative sea-level change. The lack of precise stratigraphic data within the platform prevents correlation of the latter beyond the outcrop. The previously exposed reef margin (Cattaneo et al. 1993; Zahraoui et al. 2000) does not extend to the currently available outcrop. Southwards, a Givetian reef at Ain Dakhla was completely destroyed by local eo-Variscan uplift (block tilting); its remnants are found as olistoliths in the Famennian Al Brijat Formation. Further to the south, at Ain-Al-Aliliga, different parts of a thick, at least partially zoned reef complex were brecciated and re-sedimented due to eo-Variscan block tectonics (tilting), starting during middle Givetian reef growth and ending at the top of the Givetian. Just to the east, some reefal olistoliths occur in the Al Brijat Formation, as a continuation on strike of the Ain Dakhla occurrence. On the deeper neritic carbonate shelf to the south, a small biostrome to low bioherm with permanent open-marine conditions developed in the upper Givetian at Aïn-as-Seffah. It shows three main episodes of reefal framework construction and drowned in the course of the global Frasnes Event. Subsequently, it was dislocated as a megaclast in the higher Famennian. In the Al Attamna area a thick early middle Givetian biostrome grew into an open bioherm (Fig. 6, Sidi Mohamed Smaine Formation), with a continuous influx of open-water organisms (conodonts), evidence of a retrogradation episode, and a later, minor deepening interval. It was capped by a tectonically triggered mass flow event, which mixed reef blocks, large colonial corals, and reworked iron crusts with uplifted and eroded Lower Paleozoic clastics. It can be followed to Sidi Ahmed Lemdoun nearby, where a second, thicker debris flow channel shows a second Al Attamna reef erosion in the middle Famennian. In the northern Khatouat Massif, an original reef platform is very poorly preserved at Jennabia. Its reef margin was eroded and finally re-sedimented as olistoliths in the middle Famennian Biar Setla Member of the Fouzir Formation, which correlates with the Al Brijat Formation to the northwest (Fig. 3).
The Givetian Meseta reefs show distinctive differences to those of stable northern Gondwana (western Anti-Atlas to southern Algeria). In the Oued Cherrat region, there are no mud mounds, very few bryozoans, so far no chaetetids and large rugose corals, and only minor alveolitid and phillipsastreid biostromes. The latter do not occur as many small, individual bioconstructions. There are more similarities with the rather distant Rhenish Shelf. All 18 microfacies (sub) types occur in the Rhenohercynian Zone (Ardennes to Harz Mountains). This suggests a rather low Givetian paleoclimatic gradient in the southern hemisphere. Therefore, a more conservative estimate of ca. 3000-km distance for the Prototethys/Rheic Ocean system “Variscan Sea” between the Rhenish Massive and the Meseta seems more likely than higher values (Fig. 12). The lack of facies dominated by calcimicrobes and green algae is distinctive for the Meseta and possibly paleoclimatically relevant. Future studies have to address the question whether there are taxonomic differences between the main Moroccan and Rhenish reef constructors.
References
Aboussalam ZS (2003) Das “Taghanic-Event” im höheren Mittel-Devon von West-Europa und Marokko. Münst Forsch Geol Paläont 97:1–332
Aboussalam ZS, Becker RT (2011) The global Taghanic Biocrisis (Givetian) in the eastern Anti-Atlas, Morocco. Palaeogeogr Palaeoclim Palaeoec 304:136–164
Aboussalam ZS, Becker RT, Eichholt S (2012) Conodont dating of reefs and carbonate platforms in the Middle and Upper Devonian of the Moroccan Meseta. Terra Nostra 3:27–28
Aboussalam ZS, Becker RT, Eichholt S, El Hassani A (2013) Conodont biostratigraphy and the timing of facies changes at Ain-as-Seffah (Oued Cherat Zone, Moroccan Meseta). In: Becker RT, El Hassani A, Tahiri A (eds) International Field Symposium “The Devonian and Lower Carboniferous of northern Gondwana”. Doc Inst Sci Rabat 27:11–13
Álvaro JJ, Aretz M, Boulvain F, Munnecke A, Vachard D, Vennin E (2007) Fabric transitions from shell accumulations to reefs: an introduction with Palaeozoic examples. In: Álvaro JJ, Aretz M, Boulvain F, Munnecke A, Vachard D, Vennin E (eds) Palaeozoic Reefs and Bioaccumulations: Climatic and Evolutionary Controls. Geol Soc London Spec Publ 275:1–16
Bachtadse V, Torsvik TH, Tait JA, Soffel HC (1995) Paleomagnetic Constraints on the Paleogeographic Evolution of Europe During the Paleozoic. In: Dallmeyer RD, Franke W, Weber K (eds) Pre-Permian geology of central and Eastern Europe: IGCP 233 International Conference, Göttingen, Federal Republic of Germany. Springer, Berlin, pp 567–578
Baidder L, Raddi Y, Tahiri M, Michard A (2008) Devonian extension of the Pan-African crust north of the West African craton, and its bearing on the Variscan foreland deformation: evidence from eastern Anti-Atlasw (Morocco). In: Ennih N, Liégeois J-P (eds) The Boundaries of the West African Craton. Geol Soc London Spec Publ 297:453–465
Becker RT, Aboussalam ZS, Hartenfels S, El Hassani A, Fischer T (2013) The Givetian—Famennian at Oum el Jerane (Amessoui Syncline, southern Tafilalt). In: Becker RT, El Hassani A, Tahiri A (eds) International Field Symposium “The Devonian and Lower Carboniferous of northern Gondwana”. Doc Inst Sci Rabat 27:61–76
Becker RT, Aboussalam ZS, El Hassani A, Hartenfels S, Baidder L (2015a) The timing of Eovariscan block faulting, reworking and re-deposition in the Moroccan Meseta. Strata, Sér 1(16):14–15
Becker RT, Aboussalam ZS, Stichling S, Eichholt S (2015b) The Givetian–Frasnian Hönne Valley reef complex (northern Sauerland)—a brief outline of stratigraphy and facies development. Münster Forsch Geol Paläont (pre)108: 95–106
Benfrika EM, Bultynck P (2003) Lower to Middle Devonian conodonts from the Oued Cherrat area and its southern extension (north-western Meseta, Morocco). Cour Forsch Inst Senck 242:209–215
Bohrmann G, Fischer G (1985) Stratigraphie und Fazies des Paläozoikums nördlich Azrou. Ein Beitrag zur Geologie der nordöstlichen marokkanischen Meseta. Geol et Paläont 19:15–37
Boulvain F (2007) Frasnian carbonate mounds from Belgium: sedimentology and palaeoceanography. In: Álvaro JJ, Aretz M, Boulvain F, Munnecke A, Vachard D, Vennin E (eds) Palaeozoic Reefs and Bioaccumulations: Climatic and Evolutionary Controls. Geol Soc London Spec Publ 275:125–142
Boulvain F, Coen-Aubert M, Mansy J-L, Proust J-N, Tourneur F (1995) Le Givetien en Avesnois (nord de la France): paléo-environment et implications paléogeographiques. Bull Soc belge Géol 103(1–2):171–203
Brownlaw RLS, Hocking RM, Jell JS (1996) High frequency sea-level fluctuations in the Pillara Limestone, Guppy Hills, Lennard Shelf, northwestern Australia. Hist Biol 11:187–212
Cattaneo G, Tahiri A, Zahraoui M, Vachard D (1993) La sédemintation récifale du Givétien dans la Meseta Marocaine nord-occidentale. C R Acad Sci Paris sér II 1993:73–80
Chalouan A (1981) Stratigraphie et structure du Paléozoique de l´Oued Cherrat: un segment du couloir de cisaillement de Meseta occidentale (Maroc). Notes Serv géol Maroc 42(308):33–100
Chalouan A, Hollard H (1979) Stratigraphie des terrains du Paléozoique moyen de l´oued Cherrat (Meseta occiderntale marocaine). C R Acad Sci Paris Sér D 288:199–202
Chen D, Tucker ME, Jiang M, Zhu J (2001) Long-distance correlation between tectonic-controlled, isolated carbonate platforms by cyclostratigraphy and sequence stratigraphy in the Devonian of South China. Sedimentology 48:57–78
Chen D, Tucker ME, Zhu J, Jiang M (2002) Carbonate platform evolution: from a bioconstructed platform margin to a sand-shoal system (Devonian, Guilin, South China). Sedimentology 49:737–764
Cocks LRM, Torsvik TH (2002) Earth geography from 500 to 400 million years ago: a faunal and palaeomagnetic review. J Geol Soc London 159:631–644
Coen-Aubert M (2002) Nouvelles espèces du genre Phillipsastrea d´Orbigny, 1849 près de la limite Givetien-Frasnien dans le Tafilalt et le Ma´der au Maroc et notes sur des types espagnols. Coral Res Bull 7:21–37
Coen-Aubert M (2005) Rugueux fasciculés et solitaires du Givetien supérieur dans le Tafilalt et le Ma´der (Maroc). Bull Inst Roy Sci Nat Belg, Sci d. l. Terre 75:67–85
Coen-Aubert M (2013) Phillipsastreids and Ptenophyllids (Rugosa) from the Givetian of Mauritania and northwestern Spain. Geol Belg 16(3):164–178
Cogney G (1967) Le Dévonien de la region d´Oulmes (maroc central). C R Somn Soc géol Fr 7:283–284
Copper P (2002) Silurian and Devonian reefs: 80 million years of global greenhouse between two ice ages. In: Kiessling W, Flügel E, Golonka J (eds) Phanerozoic Reef Patterns. SEPM Spec Publ 72:181–238
Copper P, Scotese CR (2003) Megareefs in Middle Devonian supergreenhouse climates. In: Chan MA, Archer AW (eds) Extreme depositional environments: Mega end members in geologic time. Geol Soc Am Spec Pap 370:209–230
Cygan C, Charriere A, Regnault S (1990) Datation par Conodontes du Dévonien (Emsien-Frasnien basal) dans le substratum du Moyen Atlas au Sud de Fès (Maroc); implications paléogéographiques. Géol Méditer 3–4:321–330
Dopieralska J, Belka Z, Walczak A (2015) Nd isotope composition of conodonts: an accurate proxy of sea-level fluctuations. Gondwana Res. doi:10.1016/j.gr.2015.02.022
Döring S, Kazmierczak M (2001) Stratigraphy, geometry, and facies of a Middle Devonian ramp-to-basin transect (western Anti-Atlas, SE Morocco). Facies 44:137–150
Dumestre A, Illing LV (1967) Middle Devonian reefs in Spanish Sahara. In: Oswald DH (ed) International Symposium on the Devonian System, vol 2. Alberta Society of Petroleum Geologists, Calgary, pp 333–350
Dunham RJ (1962) Classification of carbonate rocks according to depositional texture. Am Assoc Pet Geol Mem 1:108–121
Eckelmann K, Nesbor H-D, Königshof P, Linnemann U, Hofmann M, Lange J-M, Sagawe A (2014) Plate interactions of Laurussia and Gondwana during the formation of Pangea—constraints from U-Pb LA-S-ICP-MS detrital zircon ages of Devonian and Early Carboniferous siliciclastics of the Rhenohercynian zone. Central European Variscides. Gondw Res 25(4):1484–1500
Eichholt S, Becker RT, Stichling S (2013) Microfacies and Devonian reef development in the Oued Cherrat Zone (Aïn Khira South and Aïn-as-Seffah), Moroccan Meseta. In: Becker RT, El Hassani A, Tahiri A (eds) International Field Symposium “The Devonian and Lower Carboniferous of northern Gondwana”. Doc Inst Sci Rabat 27:37–39
El Hassani A (1991) La Zone de Rabat-Tiflet: bordure nord de la chaîne calédono-hercynienne du Maroc. Bull Inst Sci 15:1–134
El Hassani A, Benfrika EM (2000) The Devonian of the Moroccan Meseta: biostratigraphy and correlations. Cour Forsch-Inst Senck 225:195–209
El Hassani A, El Kamel F (2000) Tectonic control of Devonian reef building in Mechra ben Abbou area (northern Rehamna, Morocco). Trav Inst Sci Rabat Sér Géol Géogr Phys 20:25–30
El Kamel F (1987) Géologie du Paléozoique des Rehamna nord orientaux, Maroc. Evolution sédimentaire et structuration hercynienne d´un basin dévono-carbonifère. Sédimentation et deformation des molasses post-orogénique. Unpubl. PhD Thesis, Universite Aix-Marseille, p 298
Embry AF, Klovan EJ (1971) A late devonian reef tract on Northeastern Banks Island, NWT. Can Soc Petrol Geol 19:730–781
Ernst A, Königshof P (2008) The role of bryozoans in fossil reefs—an example from the Middle Devonian of the Western Sahara. Facies 54:613–620
Ernst A, Königshof P (2010) Bryozoan fauna and microfacies from a Middle Devonian reef complex (Western Sahara, Morocco). Abh Senck Ges Naturf 568:1–91
Errenst C (1993) Koloniebildende Phillipsastreidae und Hexagonariinae aus dem Givetium des Messinghäuser Sattels und vom Südrand des Briloner Massenkalkes (nordöstliches Sauerland). Geol Paläont Westf 26:7–45
Faber P (1980) Fazies-Gliederung und Entwicklung im Mittel-Devon der Eifel. Mainz geow Mitt 8:83–149
Fadli D (1990) Evolution sédimentaire et structural des massifs de Mdakra et du Khatouat; deux segments hercyniens de la Meseta marocaine nord-occidentale. Unpubl PhD Thesis Univ Rabat, p 272
Fadli D (1994) Le Famenno-Tournaisien. Bull Inst Sci Rabat 18:57–70
Flick H, Schmidt J (1987) Eine Vulkaninsel mit Saumriff im Devon des südlichen Rheinischen Schiefergebirges. Facies 17:67–72
Flügel E (1978) Mikrofazielle Untersuchungsmethoden von Kalken. Springer-Verlag, Berlin
Flügel E (2004) Microfacies of Carbonate rocks. Analysis Interpretation and Application. Springer-Verlag, Berlin
Flügel E, Hötzl H (1971) Foraminiferen, Calcisphaeren und Kalkalgen aus dem Schwelmer Kalk (Givet) von Letmathe im Sauerland. N Jb Geol Paläont Abh 137:358–395
Flügel E, Kiessling W (2002a) A new look at ancient reefs. In: Kiessling W, Flügel E, Golonka J (eds) Phanerozoic reef patterns. SEPM Spec Publ 72:3–10
Flügel E, Kiessling W (2002b) Patterns of Phanerozoic reef crisis. In: Kiessling W, Flügel E, Golonka J (eds) Phanerozoic reef patterns. SEPM Spec Publ 72:691–733
Franke W (1973) Fazies, Bau- und Entwicklungsgeschichte des Iberger Riffes (Mitteldevon und Unterkarbon III, NW-Harz, W-Deutschland). Geol Jb A11:3–127
Franke W (2007) Rheno-Hercynian belt of Central Europe: review of recent findings and comparisons with south-west England. Geosci south-west Engl 11:263–272
Fröhlich S (2003) Facies pattern and genesis of the Jebel Rheris Biostromes (Givetian, Eastern Anti-Atlas, Morocco). Facies 49:209–220
Fuchs A (1990) Charakter und Ende der devonischen Riffentwicklung im Elbingeröder Komplex (Harz). Facies 23:97–108
Gendrot C (1973) Environnement du Dévonien rècifal au Maroc. Notes Mem Serv géol Maroc 254:55–86
Gendrot C, Kergomard D, Rabate J (1969) Etude des formations récifales du Dévonien de la Meseta occidentale. Unpubl Rep BRPM, No 40476
Gischler E, Erkoҫ (2012 online) Facies of Devonian fore reef limestones: a quantitative study (Iberg Reef, Harz Mts., Germany). Palaeobio Palaeoenv 93:91–101
Golonka J (2002) Plate-tectonic maps of the Phanerozoic. In: Kiessling W, Flügel E, Golonka J (eds) Phanerozoic reef patterns. SEPM Spec Publ 72:21–75
Gomez F, Beauchamp W, Barazangi M (2000) Role of the Atlas Mountains (northwest Africa) within the African-Eurasian plate–boundary zone. Geology 28(9):775–778
Hladil J (1986) Trends in the development and cyclic patterns of middle and upper Devonian buildups. Facies 15:1–34
Hoepffner C, Soulaimani A, Pique A (2005) The Moroccan Hercynides. J Afr Earth Sci 43:144–165
Hoepffner C, Houari MR, Bouabdelli M (2006) Tectonics of the North African Variscides (Morocco, western Algeria): an outline. C R Geosci 338:25–40
Hollard H (1962) Attribution du Givetien des calcaires coralligènes du Maider (Maroc présaharien). C R Somn Soc Géol France 6:175–177
Hollard H (1967) Le Dévonien du Maroc et du sahara nord-occidental. In: Oswald DH (ed) International Symposium on the Devonian System, vol 1. Alberta Society of Petroleum Geologists, Calgary, pp 203–244
Hollard H (1974) Recherches sur la Stratigraphie des Formations du Dévonien Moyen, de l´Emsien Supérieur au Frasnien, dans le Sud du Tafilalt et dans de Ma`der (Anti-Atlas Oriental). Notes Serv géol Maroc 36(264):7–68
Hüneke H, Krienke K (2004) Toe-of-slope deposits of a Givetian reef-rimmed platform: provenance of calcareous density-flow deposits (Rabat-Tiflet-Zone, Morocco). Facies 50:327–346
Huvelin P (1977) Etude géologique et gîtologique du Massif hercynien des Jebilet (Maroc occidental). Notes Mém Serv géol Maroc 232(bis):1–308
Joachimski MM, van Geldern R, Breisig S, Buggisch W, Day J (2004) Oxygen isotope evolution of biogenic calcite and apatite during the Middle and Late Devonian. Int J Earth Sci 93:542–553
Joachimski MM, Breisig S, Buggisch W, Talent JA, Mawson R, Gereke M, Morrow JR, Day J, Weddige K (2009) Devonian climate and reef evolution: insights from oxygen isotopes in apatite. Earth Planet Sci Lett 284:599–609
Jux U (1960) Die devonischen Riffe im Rheinischen Schiefergebirge. N Jb Geol Paläont, Abh 110(2):186–258
Kaiser SI, Becker RT, El Hassani A (2007) Middle to Late Famennian successions at Ain Jemaa (Moroccan Meseta)—implications for regional correlation, event stratigraphy and synsedimentary tectonics of NW Gondwana. In: Becker RT, Kirchgasse WT (eds) Devonian Events and Correlations. Geol Soc London Spec Publ.,vol. 278. Geological Society of London, pp 237–260
Kaufmann B (1995) Middle Devonian mud mounds of the Ma´der Basin in the eastern Anti-Atlas, Morocco. Facies 32:49–57
Kaufmann B (1998) Facies, stratigraphy and diagenesis of Middle Devonian reef- and mud-mounds in the Mader (eastern Anti-Atlas, Morocco). Acta Geol Polon 48(1):43–106
Kazmierczak M, Schröder S (1999) Faziesentwicklung, relative Meeresspiegelschwankungen und die Migration von Korallenfaunen im Mitteldevon des östlichen Anti-Atlas, Marokko. Zbl Geol Paläont, Teil 1 1997(7–9):1177–1188
Kent DV, Van der Voo R (1990) Palaeozoic palaeogeography from palaeomagnetism of the Atlantic-bordering continents. In: McKerrow WS, Scotese CR (eds) Palaeozoic palaeogeography and biogeography. Geol Soc Mem 12:49–56
Kiessling W (2002) Secular variations in the Phanerozoic Reef Ecosystems. In: Kiessling W, Flügel E, Golonka J (eds) Phanerozoic Reef Patterns. SEPM Spec Publ 72:625–690
Kiessling W (2008) Sampling-standardized expansion and collapse of reef building in the Phanerozoic. Foss Rec 11(1):7–18
Kiessling W, Flügel E, Golonka J (1999) Paleoreef maps: evaluation of a comprehensive database on Phanerozoic reefs. AAPG Bull 83(10):1552–1587
Koch-Früchtl U, Früchtl M (1993) Stratigraphie und Faziesanalyse einer mitteldevonischen Karbonatabfolge im Remscheid-Altenaer Sattel (Sauerland). Geol Paläont Westf 26:47–75
Königshof P, Kershaw S (2006) Growth forms and palaeoenvironmental interpretation of stromatoporoids in a Middle Devonian reef, southern Morocco (west Sahara). Facies 52(2):299–306
Krebs W (1974) Devonian carbonate complexes of Central Europe. SEPM Spec Publ 18:155–208
Malmsheimer KW, Mensink H, Stritzke R (1991) Gesteinsvielfalt im Riffgebiet um Brilon. Geol Paläont Westf 18:67–83
Malmsheimer KW, Flajs G, Koch-Früchtl U (1996) Middle Devonian initial reef-Facies from the Rhenish Schiefergebirge (Sauerland and Eifel), Western Germany. Gött Arb Geol Paläont Sb2:371–375
Massa D (1965) Observations sur les series Siluro-Dévoniennes des confins Algéro-Marocains du Sud. Notes Mem Comp Franc Pétrol 8:1–187
May A (2003) Die Fossilführung des Mitteldevons im Raum Attendorn-Olpe (West-Sauerland; Rechtsrheinisches Schiefergebirge). Geol Paläont Westf 60:47–79
May A (2008) Corals (Anthozoa, Tabulata and Rugosa) and chaetetids (Porifera) from the Devonian of the Semara area (Morocco) at the Museo Geominero (Madrid, Spain), and their biogeographic significance. Bull Inst Sci Rabat, sect Sci de la Terre 30:1–12
McLean DJ, Mountjoy EW (1994) Allocyclic control on Late Devonian buildup development, southern Canadian Rocky Mountains. J Sed Res 64(3):326–340
Méndez-Bedia I, Soto F, Fernández-Martínez E (1994) Devonian reef types in the Cantabrian Mountains (NW Spain) and their faunal composition. Courier Forschungsinstitut Senckenberg 172:161–183
Michard A, Hoepffner C, Soulaimani A, Baidder L (2008) The Variscan Belt. In: Michard A, Saddiqi O, Chalouan A, Frizon de Lamotte D (eds) Continental evolution: the geology of Morocco. Lect Notes Earth Sci 116:65–132
Nance RD, Gutiérrez-Alonso G, Keppie JD, Linnemann U, Murphy JB, Quesada C, Strachan RA, Woodcock NH (2012) A brief history of the Rheic Ocean. Geosci Front 3(2):125–135
Neugebauer J (1988) The Variscan plate tectonic evolution: an improved “Iapetus model”. Schweiz Mineral Petrogr Mitt 68:313–333
Nübel H, Becker RT (2004) A Phillipsastreid Biostrome in the Late Givetian of the Maider (SE Anti-Atlas, Morocco). In: Devonian neritic-pelagic correlation and events, International Meeting on Stratigraphy, Rabat, Morocco, March1-10, 2004, p 79–80
Oncken O, von Winterfeld C, Dittmar U (1999) Accretion of a rifted passive margin: the Late Paleozoic Rhenohercynian fold and thrust belt (Middle European Variscides). Tectonics 18(1):75–91
Pedder AEH (1999) Paleogeographic implications of a Devonian (Givetian, Lower Varcus Subzone) rugose coral fauna from the Ma´der Basin (Morocco). Abh Geol B.-A. 54:385–434
Pique A, Michard A (1981) Les zones structurales du Maroc hercynien. Sci Geol Bull 34(2):135–146
Pique A, Michard A (1989) Moroccan Hercynides: a synopsis. The Paleozoic sedimentary and tectonic evolution at the northern margin of West Africa. Am J Sci 289:286–330
Poty E, Chevalier E (2007) Late Frasnian phillipsastreid biostromes in Belgium. In: Álvaro JJ, Aretz M, Boulvain F, Munnecke A, Vachard D, Vennin E (eds) Palaeozoic Reefs and Bioaccumulations: Climatic and Evolutionary Controls. Geol Soc London Spec Publ 275:143–161
Preat A (1989) Sedimentology, facies and depositional environment of the Hanonet (upper Eifelian) and Trois-Fontaines (lower Givetian) formations in Couvin area (Dinant Basin, Belgium). Bull Soc belge Géol 98(2):149–154
Roch E (1950) Histoire stratigraphique du Maroc. Notes et Mém Div Mines Géol Serv Géol 80:1–435
Rytina M-C (2013) Faziesentwicklung des mittleren Palaeozoikums am Südrand der Varisziden (Tinerhir-Region, SE Marokko). Unpubl. MSc Thesis WWU Münster, p 1–91
Rytina M-C, Becker RT, Aboussalam ZS, Hartenfels S, Helling S, Stichling S, Ward D (2013) The allochthonous Silurian-Devonian in olistostromes at „the Southern Variscan Front “(Tinerhir region, SE Morocco)—preliminary data. In: Becker RT, El Hassani A, Tahiri A (eds) International Field Symposium “The Devonian and Lower Carboniferous of northern Gondwana”. Doc Inst Sci Rabat 27:11–21
Schröder S, Kazmierczak M (1999) The Middle Devonian “coral reef” of Ouihlane (Morocco)—new data on the geology and rugose coral fauna. Geol et Paläont 33:93–115
Schudack M (1993) Karbonatzyklen in Riff- und Lagunenbereichen des devonischen Massenkalkvorkommens von Asbeck (Hönnetal, Rheinisches Schiefergebirge) Geol Paläont Westf 26:77–106
Shinn EA (1968) Practical significance of birdseye structures in carbonate rocks. J Sed Petrol 38:215–223
Simancas JF, Tahiri A, Azor A, Gonzáles Lodeiro F, Martínez Poyatos DJ, El Hadi H (2005) The tectonic frame of the Variscan-Alleghanian orogeny in southern Europe and northern Africa. Tectonophysics 398:181–198
Stampfli GM, Borel GD (2002) A plate tectonic model for the Paleozoic and Mesozoic constrained by dynamic plate boundaries and restored synthetic oceanic isochrons. Earth Plan Sci Lett 196:17–33
Stichling S (2013) Biostrom-Abfolgen am Nordrand des Maider (S-Marokko): Mikrofazies, Paläoökologie und Stratigraphie. Unpubl MSc Thesis WWU Münster, p 1–87
Struve W (1963) Das Korallen-Meer der Eifel vor 300 Millionen Jahren. Funde, Deutungen, Probleme. Nat u Mus 93:237–276
Struve W (1989) Zur Lebensweise von Schalentieren auf mitteldevonischen Karbonat-Plattformen. Nat u Mus 119(4):128–139
Tahiri A (1983) Lithostratigraphie et structure du Jebel Ardouz—Maroc hercynien. Bull Inst Sci Rabat 7:1–16
Termier H, Termier G (1951) Stratigraphie et Paléobiologie des Terrains Primaires de Benahmed (Chaouia sud, Maroc). Notes et Mem, Div Mines et Géol, Serv Géol 85:47–104
Termier H, Termier G, Vachard D (1975) Recherches micropaléontologiques dans le Paléozoique supérieur du Maroc central. Cahiers Micropaléont 4:1-99, 10 pls
Töneböhn R (1991) Bildungsbedingungen epikontinentaler Cephalopodenkalke (Devon, SE-Marokko). Gött Arb Geol Paläont 47:1–114
Weller H (1991) Facies and Development of the Devonian (Givetian/Frasnian) Elberingerode Reef Complex in the Harz Area (Germany). Facies 25:1–50
Wendt J (1993) Steep-sided carbonate mud mounds in the Middle Devonian of the eastern Anti-Atlas, Morocco. Geol Mag 130:69–83
Wendt J, Kaufmann B (2006) Middle Devonian (Givetian) coral-stromatoporoid reefs in West Sahara. J Afr Earth Sci 44:339–350
Wendt J, Belka Z, Moussine-Pouchkine A (1993) New architecture of deep-water carbonate buildups: evolution of mounds into mud ridges (Middle Devonian, Algerian Sahara). Geology 21:723–726
Wendt J, Belka Z, Kaufmann B, Kostrewa A, Hayer J (1997) The world’s most spectacular carbonate mud mounds (Middle Devonian, Algerian Sahara). J Sed Res 67(3):424–436
Wilson JL (1975) Carbonate facies in geologic history. Springer, New York, p 1–471
Zahraoui M (1991) La plateforme carbonate dévonienne du Maroc occidental et sa dislocation hercynienne. Unpubl PhD Thesis, Universite Bretagne Occidentale, p 261
Zahraoui M (1994) Le Dévonien inférieur et moyen. Bull Inst Sci Rabat 18:43–56
Zahraoui M, El Hassani A, Tahiri A (2000) Devonian outcrops of the Oued Cherrat shear zone. Notes et Mem Serv géol Maroc 399:123–128
Acknowledgments
This study was funded by the German Research Foundation (DFG) within project Be 1367/11-1, which includes formal co-operation with Ahmed El Hassani (Rabat) and fieldwork co-funding by the Centre National pour la Recherche Scientifique et Technique (CNRST) Maroc. Z. Sarah Aboussalam (Münster) provided within the joint DFG programme the conodont data that enabled the reef dating. Sören Stichling (Münster), Sven Hartenfels (Münster), Ahmed El Hassani, L. Baidder, and M. Benfrika (Casablanca) participated in parts of the field work. We are especially thankful to our very helpful driver Alaoui, G. Schreiber and the Münster preparation team assisted in the production of polished slabs and many thin-sections, T. Fährenkemper (Münster) produced some of the diagrams. We thank Peter Königshof (Frankfurt a. M.) and Heiko Hüneke (Greifswald) for their helpful reviews.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Eichholt, S., Becker, R.T. Middle Devonian reef facies and development in the Oued Cherrat Zone and adjacent regions (Moroccan Meseta). Facies 62, 7 (2016). https://doi.org/10.1007/s10347-015-0459-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10347-015-0459-z