
Abstract
Main conclusion
Open-Top Chambers should be more used in tropical ecosystems to study climate change effects in plants as they are still insufficient to extract plant response patterns in these ecosystems.
Abstract
Understanding flora response to climate change (CC) is critical for predicting future ecosystem dynamics. Open-Top Chambers (OTCs) have been widely used to study the effects of CC on plants and are very popular in temperate ecosystems but are still underused in tropical regions. In this systematic review, we aimed to discuss the use of OTCs in the study of the effects of different agents of climate change on tropical flora by presenting scientometric data, discussing the technical aspects of its use and enumerating some observations on plant response patterns to climatic alterations in the tropics. Our analysis indicated that the bottleneck in choosing an OTC shape is not strictly related to its purpose or the type of parameter modulated; instead, passive or active approaches seem to be a more sensitive point. The common critical point in using this technique in warmer regions is overheating and decoupling, but it can be overcome with simple adaptations and extra features. The most frequently parameter modulated was CO2, followed by O3 and temperature. The plant families with more representatives in the studies analyzed were Fabaceae, Myrtaceae, and Poaceae, and the most represented biome was tropical and subtropical moist broadleaf forests. In conclusion, OTCs are a valuable and feasible tool to study CC effects on various tropical ecosystems, regardless of structure, active/passive approach, or other technical features. One of the primary advantages of this methodology is its applicability for in situ use, eliminating the need for plant transplantation. We encourage studies using OTC experimental design for plant conservation in the tropics.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Anthropic activities exacerbatedly releases greenhouse gases, leading to Earth’s increased surface temperature and accelerated climate change (Min et al. 2011). Besides other effects, predictions indicate an average temperature rise of 1 °C to 5.7 °C and an increased frequency of extreme events (IPCC 2022). Alongside other anthropogenic pressures, these imbalances can have both local and global effects, such as decreased rainfall in specific regions (Lawrence and Vandecar 2015) and alternations in the carbon cycle (Mitchard 2018).
Tropical ecosystems are crucial for global nutrient and climate dynamics, storing over half of the available carbon (Bai and Cotrufo 2022; Lewis 2006; Pan et al. 2011; Stocker et al. 2014). They also contribute to one-third of terrestrial primary productivity (Beer et al. 2010) and regulate the Earth's energy balance and water cycle (Duran-Quesada et al. 2009). Disturbances in this regulation lead to habitat fragmentation, altered vegetation distribution (Debb et al. 2018), loss of plant diversity (Bitencout et al. 2016), and reduced primary productivity (Lyra et al. 2017). Climate change impacts can also affect crop species (native or not) cultivated in the tropics, resulting in lower yields and agricultural production losses (El-Sharkawy 2014). In contrast, there is extensive discussion in the scientific literature about the positive effects of climate change on plant species. Those with C3 metabolism, for example, would benefit from higher atmospheric CO2 concentration due to increased photosynthetic rates and reduced photorespiration (Munné-Bosch et al. 2013).
Understanding species’ vulnerability to the effects of climate change is fundamental for building focused strategies to preserve more sensitive species and also predict further extinctions. Vulnerability assessments are complex, requiring information from the individual level, to community levels and socio-ecological evaluations (Foden et al. 2019). One of the ways to evaluate the response at the individual level is by measuring functional traits of the species (Andrew et al. 2022), which has already been reported in the literature to reflect adaptation of species to the site’s climate and soil properties (Wright et al. 2017). Also, these individual responses can summarize plant life history strategies in different environments (Diaz et al. 2016). Hence, to address these and other related questions, there is urgent need for further exploration of climate change experiments, especially in tropical regions (Cavaleri et al. 2015).
Several techniques are available for climatic abiotic changes simulation studies, such as FACE (free-air CO2 enrichment system) and FC (Fully enclosed chamber). However, the Open-Top Chambers (OTCs) are one of the most widely used (Heyneke et al. 2012). First described by Heagle et al. (1973), OTCs are transparent chambers used for in situ climatic experiments with an upper opening to maintain airflow with the external environment. They can have a lower opening too. The chambers isolate their internal environment from external conditions laterally, which allows them to modulate microclimatic conditions inside them. One of the advantages of this methodology is the possibility of abiotic modulation, either actively or passively, directly impacting the project's cost and feasibility. The active modulation of parameters is controlled using an equipment attached to the OTC, for example, ventilating pre-heated air into the chamber. In contrast, passive modulation is a consequence of the chamber structure itself, inducing natural internal climatic changes. Although not fully controlled, they do not need any other equipment keeping, the project cost low.
The OTCs allow the modulation of several abiotic conditions, such as temperature, concentration of atmospheric compounds, precipitation, and even biotic effects, e.g., herbivory. Since this device can be assembled virtually anywhere, it allows studies on plant response to several environmental conditions in situ, and it was already used in several ecosystems (e.g., De Frenne et al. 2010; Heyneke et al. 2012; Hollister et al. 2022; Samol et al. 2015). An OTC experiment does not need transplantation of individuals, which could bias the plant response results. Furthermore, compared to other techniques, OTCs are more stable in maintaining atmospheric parameters, such as induced CO2 concentrations (Kumar et al. 2017).
Interestingly, the OTCs are not very popular in the tropics, despite their great potential for tropical climates. They are broadly used in field research on temperate and polar zones, where they are well established for studying flora traits response to diverse microclimatic conditions as a climate change consequence. In this systematic review, we collected data from OTC experiments based solely in the tropical region to understand and disseminate its use in this diverse and variable environment. The choice of not adding data from temperate or polar region was to avoid the interference of the difference on the well-stablished experimental design for these regions. We understand that divergence on biotic and abiotic conditions between polar, temperate, and tropical regions’ ecosystems may dictate several OTC design adaptations. Therefore, we posed three main questions for insights in the regular, but still scarce, use of OTCs in the tropics:
-
Where OTCs are being used?
-
How are climate change effects investigated using OTCs?
-
What organisms and traits are being looked at?
With this aim, we registered and discussed scientometric data, assessing technical aspects, benefits, limitations, and challenges of OTCs experiments done in the tropics. Then, we enumerated the most frequent observations on patterns of plant response to climatic alterations caused by OTC experiments. In conclusion, OTCs are powerful tools for understanding consequences caused by environmental changes in the tropical flora although they are still underutilized in the tropics. As OTCs allow the measurement of plant response and functional traits, this is an essential and accessible methodology to evaluate the species vulnerability to climate change. Hopefully, this review will bring valuable information to new users and foment a broad use of OTCs in tropical ecosystems.
Methodology
Systematic review
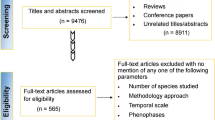
For the selection of scientific articles, a search was conducted in the Scopus database on May 15, 2023, with the words: (TITLE-ABS-KEY (otc* OR open-top chamber* OR open top chamber*) AND TITLE-ABS-KEY (plant* OR vegetation* OR tree* OR shrub* OR herbaceous*) AND ALL (tropic*) AND NOT TITLE-ABS-KEY (outdoor thermal comfort), without time restriction, which returned 697 articles. The first screening followed the following exclusion criteria: C1: full articles in English; C2: Use of Open-Top Chambers as a method; C3: experiments conducted in tropical environments; and C4: terrestrial vascular plants evaluated. After the first screening, 117 articles remained.
To be selected in the second screening, the articles should present the following information: C5: information on the parameter modulated in the OTCs (temperature, humidity, gases, light, etc.); C6: formats/dimensions of the OTC; C7: duration of the OTC experiment (classified as very short (any amount of time up to 12 months), short (between 13 and 24 months), medium (between 25 and 60 months), and long terms (more than 60 months); C8: evaluate the plant response to the applied treatment (classified in morphological, phenological, physiological, biochemical, agronomic, and genetic responses, adapted from Griffin-Nolan et al. (2018)—Table S1); and C9: inform the species used. If one or more of the above information items was missing or not clearly stated, the paper was removed from the screening.
All the screening process is illustrated in Fig. 1. In addition, to check if groups are being prioritized in these studies, species were consulted regarding their degree of threat following the International Union for Conservation of Nature (IUCN) classification. Species not classified by the IUCN will be listed as "unclassified." The parameters modulated in the OTCs were classified into atmospheric, soil, and biological and could simultaneously meet multiple classifications. To determine the biomes in which the experiments were conducted, sites were classified using Olson et al.'s (2001) biome delimitations.

Flowchart of the screening process on the review of the use of Open-Top Chambers in the tropics. Hexagons represent the first (S1—blue) and second (S2—green) screening steps. Sharp-edge rectangles represent each exclusion criterion (C) used. Rounded-edged rectangles are the classifications applied. *: classifications adapted from Griffin-Nolan et al. (2018). ▲: species were checked for threat classification according to the International Union for Conservation of Nature (IUCN)
To examine whether the shape of the OTCs varied based on the experimental design, a PCA was performed. This analysis considered the influence of modulated parameters and active/passive approaches. OTC shapes were categorized as variables, and the count of experiments employing specific parameters (e.g., CO2, O3, active temperature, etc.) in each OTC shape was recorded. Additionally, the number of publications that utilized active or passive approaches in experiments within each shape was calculated. Data were transformed using cube root, which can be applied to zero values and showed the best skewness reduction compared to other transformation methods. The PCA was conducted using dudi.pca of the Ade4 package (Dray and Dufour 2007) and visualization used fviz_pca_var of fact extra package (Kassambara and Mundt 2020) in the R Studio software (Rstudio Team 2020).
Results and discussion
Where OTCs are being used?
After the screenings, the analysis included 80 studies, with approximately 85% of them published after 2010 (Table S2, Figure. S1). These studies were distributed across various journals, including Flora, Journal of Plant Ecology, Photosynthetica, and WSEAS, accounting for 20% of the total (Figure S2, Table S2).
Although, at first, the total number of studies analyzed herein seems low compared to all OTC studies ever published, we opted to analyze exclusively those from the tropical region. This avoided statistical bias due to differences in the number of experiments and type of experimental design used in other climatic zones. Moreover, the tropical ecosystems are more diverse and have different environmental characteristics compared to temperate and polar ecosystems, what can affect the work results. Therefore, adaptations on the OTC design for the tropical region may be employed. Our aim in this review is to register the current used methods and applications solely in the tropics. One of the primary criteria leading to the exclusion of several papers from the analysis was their lack of relevance to tropical areas, despite containing the word "tropic*." This discovery suggests that, despite the potential of OTCs to simulate climate change scenarios, their utilization in the tropics remains underexplored compared to temperate or polar regions. Two potential factors contributing to this are the unstable climate and possible overheating, which will be discussed further in the text. For a comprehensive list of results from the literature survey, including author names and paper titles, please refer to Supplementary Table S2.
Regarding the distribution of publications by countries, the study areas that were most prominently represented were India (27.8%), followed by China (19%), and Brazil (19%). Additionally, there were articles from several other countries, although in smaller proportions, including South Africa, Argentina, Mexico, Malaysia, Colombia, Panama, Thailand, Sri Lanka, Venezuela, the Philippines, and the United States (Hawaii). We registered 43 experiments conducted in tropical and subtropical moist broadleaf forests (green in Fig. 2). The second most common biome was subtropical grasslands savannas and shrublands (n = 14; orange), followed by tropical and subtropical dry broadleaf forests (n = 12; dark blue), deserts and xeric shrublands (n = 6; pink), tropical and montane grasslands and shrublands (n = 4; brown), and mangroves (n = 2; yellow) (Fig. 2, Table S2).

Map illustrating the site experiments by biome from the papers published that conducted experiments using Open-Top Chambers in the tropics. Each color represents a biome that followed the classification given by Olson et al. (2001): tropical and subtropical moist broadleaf forests (green), tropical and subtropical dry broadleaf forests (dark blue), deserts and xeric shrublands (pink), tropical and subtropical grasslands savannas and shrublands (orange), montane grasslands and shrublands (brown), and mangroves (yellow)
How are climate change effects investigated using OTCs?
In this topic, we summarized and discussed trends in technical information on tropical OTC experiments. Unfortunately, some technical details are still unclear in the literature. The tropics are a diverse region with several types of biomes and ecoregions; therefore, we would expect different shapes and sizes of OTCs adapted to each specific environment. Nevertheless, we obtained valuable information on general trends in terms of technical details on this methodology fundamental for planning effective experimental designs in the tropics.
The active vs. passive modulation of microclimate and other conditions
The choice between active and passive approaches for OTC experiments is mainly determined by the parameters that will be modulated and the technical support available on-site. First, our analysis observed that gases, soil compounds, and precipitation were always modulated actively, representing 87.1% of the experiments. Compared to other methodologies such as FACE, active OTC systems have lower costs per experiment due to reduced carbon dioxide consumption, being more cost-effective. While OTCs may have chamber effects on microclimate, such as airflow and shading (which will be explored afterward), they offer a closer approximation of natural field conditions than greenhouse or laboratory experiments (Macháčová, 2010).
When comparing passive and active OTCs, the latter can be labor intensive, so in remote locations or without technical support, passive OTCs become a great option (Aragón-Gastélum et al. 2021; Hollister and Webber 2000). Passive OTC studies were broadly distributed in tropical and subtropical moist broadleaf forests, tropical and subtropical dry broadleaf forests, montane grasslands and shrublands, and mangrove areas, showing the possibility of using passive OTCs in several tropical environments. In our results, all experiments that used passive OTC reported significant changes in temperature and humidity (of air and/or soil), indicating the efficiency of the technique from potentially shaded (broadleaf forests) to open areas (grasslands and mangroves). Despite the lack of direct control over the imposed conditions, passive OTCs provide a uniform microclimate like a natural environment (Samol et al. 2015). Furthermore, microclimatic conditions in passive OTC experiments depend on natural daily fluctuations of environmental conditions (Bokhorst et al. 2013) and will vary accordingly; hence, constant monitoring of environmental variables is crucial.
Format of the OTCs and number of replicates per treatment
The original OTCs format, described by Heagle et al (1973), corresponds to a cylindrical structure of 3 m in diameter and 2.7 m in height. According to our results, the shapes and dimensions of the OTCs diversified a lot, allowing them to meet the needs of different experimental designs and local characteristics. Among the shapes cited, 50% of the studies used cylindrical OTCs, followed by hexagonal (14.1%), cubic (12.8%), octagonal (9%), rectangular (2.6%), and decagonal (1.3%) formats. About 12% of the studies did not specify the shape of the OTC used, informing only its dimensions such as diameter or internal area.
It is a fact that OTCs induce climatic changes, but does the shape vary depending on their application and goals? Our survey noted that (i) the same parameter could be modulated in different formats and (ii) different parameters could be modulated simultaneously within the same OTC. These results indicate that the nature of the parameter modulated does not necessarily require a specific format of the OTC. Studies aiming at gas elevations got significant alterations using five different OTC shapes in the active approach. Also, the same structure used for active gas modulation was used for active temperature modulation.
Through a PCA (Fig. 3, Table S3), we noted that variables such as CO2, O3, and active modulation of temperature were prevalent in cylindrical OTCs. This result shows a tendency of using cylindrical OTCs for active modulation experiments (126 active- and 3 passive-approach experiments). Also, most experiments analyzing soil nutrient, which are actively modulated, utilize cylindrical OTCs. On the contrary, passive temperature and humidity modulations are preferably made with hexagonal OTCs, such as overall passively modulated experiments (24 passive- and 9 active-approach experiments). Other shapes are much less used in the tropics. Only one study used octagonal OTC for passive approach, and 16 used for active approach. Cubic shape had 24 active-approach studies, while decagonal had 32 and rectangular had 2.

Principal Component Analysis of the contribution of variables (modulated parameters and active and passive approaches) in the choice of Open-Top Chamber shape used in tropical environments. 1: cubic; 2: cylindrical; 3: decagonal; 4: hexagonal; 5: octagonal; 6: rectangular
Our result indicates that the determination of the OTC shape is mainly based on the type of modulation as active or passive, instead of the analyzed parameters. Therefore, the choice of a passive or an active approach is a critical point to define the OTC structure. Hexagonal OTCs were designed for maximizing temperature increase in passive approaches, trapping long wave radiation and avoiding advection flux by wind (Hollister et al 2022). The cylindrical shape facilitates active air circulation; therefore, mostly used for active approaches. Other features, such as the size of the experimental site and the species, have a strong role in OTC shape too (Frei et al. 2020).
To identify and minimize biased effects of the OTC’s structure on the experiment, such as limitations in luminosity and airflow, many studies used the so-called absolute controls, consisting of sample groups with no OTC and control groups with OTC but no treatment or control conditions. Moreover, to avoid these limiting factors, care with the material and angulation of its faces is imperative, despite the minimal shading effects of the OTC (Chiba and Terao 2014). The material of the OTC must allow a good light transmission to avoid impairment of the photosynthetic active radiation (PAR). Thus, polycarbonate, polyvinyl chloride (PVC), acrylic, and glass are viable options to maintain more than 80% of the PAR (Godfree et al. 2011; Heyneke et al. 2012; Samol et al. 2015). Hence, all these materials were already used in tropical experiments with success.
Especially in passive OTC structures, in which the maintenance of airflow depends solely on environmental dynamics, mostly wind, the angle of the frustum is another relevant factor (Fig. 4). Frustum angles between 45° and 60° help maintain airflow into the OTC (D'Andrea and Rinaldi 2010; Godfree et al. 2011; Samol et al. 2015). If too small, it restricts the airflow and might cause overheating. It is important to consider airflow in the positioning of sensors for measuring the internal conditions, so that the revolving air of the upper layer inside the chamber does not influence it.

Diagram of the frustum angle (purple dashed circles) on passive (A) and active (B) OTCs, which might influence on air flow, especially on passive approaches. Frustum angles determine the aperture size of the chamber. Blue arrows indicate airflow near the soil when OTCs are not fully in contact with the ground
Duration of the on-site experiments
There was great variation in the duration of the experiments among the analyzed studies. Considering our classification on the duration of the experiments, 63.5% of the experiments were very short-term, followed by medium-term (23%), short-term (12.2%), and long-term (1.4%) experiments. Although the OTCs are an on-site methodology that allows performing in situ experiments for longer periods of time, annual or interannual analysis was rare. Interestingly, previous studies reported that the effect of modulated parameters with OTCs can be time dependent due to interannual variations (Ganjurjav et al. 2015; Yu et al. 2019; Wang et al. 2013; Zheng et al. 2013); therefore, the use of year-round approaches must be considered when planning OTC’s experiments in the tropics.
Modulated environmental parameters and replicas
Among the parameters, 57.7% of the treatments modulated only one or more atmospheric factors, followed by atmospheric factors + soil compounds (35.2%). In smaller proportion, the modulation of atmospheric with biological factors and soil-only changes accounted for 7% of the treatments.
Most studies (67.4%) used up to three replicas of OTC per treatment. Complex OTC systems, requiring high costs using equipment such as heaters or CO2 pumps, usually had one OTC per treatment, with no replica (17.5%). The number of individuals monitored within the OTCs varied greatly, mainly according to the studied species size. Trees are usually represented by one or a few individuals, while grasses can be quantified in the density of seeds planted per square meter and have several biological replicates (Padhy et al. 2020). This observed pattern is similar to that used in non-tropical environments (D'Andrea and Rinaldi 2010). Below we discuss the importance of the most frequently modulated environmental parameters in relation to climatic change studies using OTCs in the tropics.
Modulated atmospheric gases
Most publications modulated atmospheric parameters (82.7%). The two greenhouse gas representatives of the selected articles were CO2 (76.2%) and tropospheric O3 (6.4%). About 70% of the studies that performed gas modulation opted for active modulation in cylindrical shapes, although hexagonal and rectangular shapes were also reported in one single work each. Clarifying the effects of these greenhouse agents on the tropical flora is crucial not only for biodiversity conservation but also to the world’s food security, as the tropical zone is a major food producer. This is why we can directly correlate the use of OTC methodology with climatic change studies.
The concentrations of CO2 used for control groups were between 350 and 400 ppm, depending on each experimental site, which was the value published in the IPCC AR6 report (IPCC 2022) and the highest in the last two million years. The active approach maintained the CO2 concentration inside the control groups’ OTC equal to the external environment by a ventilation system pulling the external air into the OTC, without CO2 enhancement. For the treatment groups, inducted CO2 concentrations ranged from 500 to 800 ppm, which are in accordance to the IPCCs predictions for the year 2100 (IPCC 2014, 2022).
It is known that increased CO2 will cause impacts on the flora in several ways. Literature indicates changes in mineral content (Martins et al. 2014), growth parameters (Esmail and Oelbermann 2011), and reproductive abnormalities (Alzate-Marin et al. 2021). Some positive interactions, such as reduced abiotic stress (AbdElgawad et al. 2016) and efficient nutrient use (Baligar et al. 2021), might happen if conditions are adequate (Raubenheimer and Ripley 2022). Therefore, it is important to note that responses may differ between plant groups (AbdElgawad et al. 2016).
In the studies that performed atmospheric modification with O3, the control concentration ranged from 10 to 30 ppb, depending on the location of the experiment. For agricultural production security, concentrations above 40 ppb are considered critical levels in several localities (Rai and Agrawal 2012). The most common concentrations for the groups subjected to O3 elevation were 50, 110, and 250 ppb. High concentrations of tropospheric ozone can cause various impacts on plants on different levels, from macroscales, such as negatively influencing the production of agricultural species (Singh and Agrawal 2011; Tiwari et al. 2008), to microscales, causing stomatal damage (Mills et al. 2007), production of reactive oxygen species within tissues (Broberg et al. 2015), damage to the tissues, and cellular malfunction (Saxena et al. 2019).
Modulated air temperature
About 53% of the papers showed air temperature increases by passive heating. All these works used hexagonal chambers, whereas the average heating increase was around 2 °C. The minimum heating increase reported was 1.3 °C and the maximum was 8 °C among these studies. Moreover, other chamber shapes have been used in the tropics for passive temperature induction, including models adapted for specific locations, such as flooded areas (Chiba and Terao 2014). The passive heating effect obtained with OTCs is mostly the result of the wind dam because their upper opening allows direct solar irradiation (Hollister et al. 1998). Furthermore, variation in temperature increase is also associated with other factors such as time of the day (Aragón-Gastélum et al. 2020), time of experiment, period of the year/season (Pérez-Ruiz et al. 2018), or both (Samol et al. 2015). In some papers in which other factors were actively modulated, temperature increase occurred as a side effect.
Despite the advantages of the passive compared to the active approach, a limiting factor is the impossibility of temperature control and the eventual decoupling of internal and external conditions. This may cause opposition to some new users to apply the methodology in areas with less predictable environmental conditions, such as in the tropics. Tropical seasons are more distinguished between one dry and one wet season than 4-season year; therefore, they have high variability (including interannual) in the precipitation regime (Feng et al. 2013) influencing chamber performance. This fact is another limitation to the use of OTCs in these areas compared to temperate or polar regions. However, some strategies can cope with these limitations, making the passive approach a good cost–benefit methodology (Heyneke et al. 2012).
In the context of OTC experiments, decoupling can occur in tropical and temperate areas. It is characterized when the induced microclimate inside the chamber is too different from the external environmental conditions and no longer reflects it. Decoupling can happen with any climatic factor, but it is prevalent with temperature. One of the solutions to avoid decoupling is to have a space between the OTCs and the ground for small air flow. This improves air circulation, avoiding excessive heating (Hollister et al. 2000). Also, the samples should be centralized inside the OTCs to avoid overheating due to proximity to the faces of the structure (Aragón-Gastélum et al. 2014) or perform rotation cycles of sample positioning when grown in pots (Chiba and Terao 2014). Another alternative is using height-adjustable chambers, preventing overheating during summer (Godfree et al. 2011). For locations with large daily temperature variations and overcooling during the night, the passive increase of nighttime temperatures could be obtained using thermal masses around the chambers (Godfree et al. 2011). The thermal masses are narrow transparent cylinders filled with liquid that, attached to the walls of the OTCs, retain part of the day’s heat and release it progressively during the night. Even with the lack of air temperature control, passive OTCs are very efficient in mimicking the temperature elevation due to global warming, according to the IPCC predictions.
Modulated soil parameters
Nitrogen (N) is one of the most used compounds (58.6%) in OTCs soil treatments, with the application at concentrations ranging from 6.25 g m−2 year−1 to 20 g m−2 year−1. In the second place, it is Phosphorus (P) (12.9%), administered alone or in association with N. The modulation of other substances, such as sodium chloride, cadmium, commercial fertilizers (NPK 10:10:10 type), and high-nutrition soils, was less represented. The influence of these compounds has been tested alone or in conjunction with atmospheric conditions, such as air temperature increase. Alterations in soil compound dynamics, especially N, can unshorten nitrogen cycle in tropical forests under warming (Lie et al. 2021). Another common association observed in the selected articles was between N and/or P and CO2 or O3 alterations. The soil nutrient balance directly affects plant response under different scenarios of atmospheric modifications. Higher N concentrations in O3 stress situations may contribute to leaf senescence in Brassica campestris (Singh et al. 2012). Conversely, in some plant species, no effect was observed between N modulation and CO2 (Liu et al. 2011).
Complete data regarding the modulated parameters, OTC technical details, and other information presented in this section are shown in Table S4. It is noted that some modulations were less representative within studies, such as precipitation modulation (Avila et al. 2020; Kumar et al. 2017; Oliveira et al. 2013; Sreeharsha et al. 2020, Pérez-Ruiz 2018; Pillay and Ward 2022), and the effects of air pollutants on vegetation (Nakazato et al. 2016; Esposito and Domingos 2016; Rai and Agrawal 2014, Ishii et al. 2004), but they are also listed in Table S4
What organisms and traits are being looked at?
In total, 47 plant families are represented in the selected articles, with Fabaceae (17.6%), Myrtaceae (13%), and Poaceae (14.1%) as the most abundant. Crop species accounted for 26.5% of the 92 studied species. The most studied species in all papers was Oryza sativa (8.8%), followed by the non-agricultural Acmena acuminatissima (6.1%) (Table S5). Of the countries with the most publications, India had 95.2% of the studied species as cultivated varieties, of which 33.3% was Oryza sativa. On the contrary, studies in China and Brazil evaluated mostly non-agronomic species (98.7% and 64.7%, respectively).
Excluding cultivated varieties, most species (72.3%) were not classified by the IUCN risk. Of those evaluated by IUCN, 22.9% were classified as LC (least concern), while EN (endangered), VU (vulnerable), and NT (nearly threatened) species corresponded to 1.6% each. Trees were more represented (63.6%) in the studies, followed by shrubs (11.7%), grasses (11.3%), lianas (6.1%), herbaceous plants (3.2%), cacti (0.8%), and bromeliads (0.4%).
Our results indicate that most of the attention has been given to species not classified yet by IUCN, although they could have local conservation importance. The motivation for choosing the study species may be influenced by several factors, such as economic/medicinal importance, abundance, access to individuals, and availability of plant material for cultivation (e.g., seeds). As threatened species are often more difficult to find and eventually cannot be cultivated ex situ, OTCs are an excellent methodological alternative for increasing the studies about climate change effect on endangered species by allowing in situ experiments. In addition, since OTCs have highly adaptable dimensions and shapes, they can be adjusted to different species size and terrains for in situ installation, which is a great advantage of this methodology.
Regarding plant responses to treatments with OTCs, the metrics evaluated were classified as physiological (45%), morphological (26.4%), biochemical (15.9%), phenological (5.2%), agronomic (4%), anatomical (2.3%), and genetic (0.5%) (Fig. 5). The physiological category had the highest number of metrics evaluated, with a total of 118 traits being analyzed in all papers. This category was followed by the biochemical (51), morphological (47), phenological (27), agronomic (15), anatomical (7), and genetic (1) (Table S6).

Percentage of publications using Open-Top Chambers in tropical environments that evaluated plant traits classified as: physiological, morphological, biochemical, phenological, agronomic, anatomical, and genetic
The most frequently studied metrics from the most represented classifications will be discussed below. Overall, we observed that the most studied plant traits are direct plant responses to alterations in CO2 levels. This topic is widely discussed in the literature of climatic changes effects on plants, regarding whether it would benefit plant species or not. Additionally, the responses observed, especially in stomatal conductance and biomass, are related to the significant carbon storage capacity in tropical environments and underscore the fundamental role of rainfall regimes in this process. This emphasizes the efficiency of OTCs in studies of plant responses to climate change, regardless of their structure, active/passive approach, or other technical features.
Stomatal conductance (gs)
Stomatal conductance (gs) was measured in 15 studies, with the majority focusing on its response to elevated CO2 levels (eCO2) (76.2%). The sensitivity of gs in tropical flora varies in response to different abiotic factors. Although it is generally considered a highly sensitive plant trait (Slot and Winter 2017), it does not respond uniformly to all abiotic factors. Interestingly, most studied species (71% of 29) did not exhibit a gs response to eCO2 levels. This suggests that tropical flora may exhibit a greater resistance to the effects of eCO2 on gs. This resilience could be attributed to the predominant influence of seasonality (related to precipitation and temperature) and the carbon store capacity of tropical flora. Similar findings were previously observed in Amazonian species, which showed a greater gs response to temperature and light availability (Fauset et al. 2019; Marenco et al. 2014).
Total biomass
Most of the 27 articles that measured biomass as a morphological parameter analyzed aerial and root biomass. A significant increase in plant biomass was observed in 85.7% of the studies, either due to the modulation of CO2 alone or in association with other factors such as N, P, or air temperature.
The boost in plant biomass resulting from eCO2 levels retains nutrients, as it enhances photosynthetic rates and resource demand. However, the availability of nitrogen (N) can become a limiting factor in some ecosystems (Luo 2004; Dieleman et al. 2012). Unless changes in nutrient cycling increase N availability (Luo et al. 2004, 2006), growth in response to eCO2 will eventually stabilize. Corroborating this, the most substantial increase in plant biomass in the studies analyzed occurred when the rise in CO2 was accompanied with an increase in N availability in the soil, as opposed to when they were modulated separately.
Concerning air temperature, two papers that solely manipulated this parameter showed an increase in plant biomass, either in the root system or in both root and aerial systems. Air temperature can also stimulate alterations in nutrient availability (Hyvönen et al. 2007; Pendall et al. 2004), increasing N availability, as observed in heating experiments (Way and Oren 2010). Consequently, this change promotes an increase in biomass. However, other crucial factors, such as precipitation, should be considered in the context of tropical flora, although they have received limited attention in OTC experiments. In tropical forest trees and lianas, it was observed that only the root system exhibited an increase in biomass in response to eCO2, while aboveground biomass increased later, following the onset of the rainy season (Marvin et al. 2015).
Starch content
Starch content was the most frequently studied trait in the biochemical category. Interestingly, although Oryza sativa was the most studied species, starch content was primarily investigated in native species. Among the publications analyzed, all species exhibited a significant increase in foliar starch content in response to eCO2 levels, as expected in stress responses (Thalmann and Santélia 2017). The rise in starch content can significantly enhance photosynthetic activity and biomass (Arenque et al. 2014) under various conditions, such as high salinity (Amirjani 2011) and drought (Parida et al. 2007). It has been associated with resistance to flooding in some species, although starch content typically responds more to drought than to flooding (Li et al. 2018).
One study showed an increase in leaf starch without a concurrent increase in biomass (Coley et al. 2002), suggesting that starch content and biomass may not always be directly correlated. The increase in starch may also be linked to the successional stage of the species. Plants in early and mid-successional stages exhibited a greater increase than those in late successional stages (Lovelock et al. 1998; Winter et al. 2000), possibly due to differences in growth rate and physiological processes (Li et al. 2018).
Final considerations
Briefly, the most frequent parameters modulated were CO2, tropospheric O3, and air temperature in cylindrical active-approach OTC experiments that lasted up to 12 months. Interestingly, we observed that the shape of the OTC does not vary based on the parameter modulated in the tropics. A more critical defining point to the OTC structure is how these parameters are modulated, passively or actively, where hexagonal and cylindrical shapes were most used respectively.
In our view, OTCs have great potential for studying climate change effects on tropical flora; however, they are still extremely underused in tropical ecosystems. The lack of OTCs experiments in the tropics makes it still difficult to precisely determine if there is an optimal chamber for each scenario/objective in such a variable environment as the tropics, although some insights were obtained in this review. Despite the high potential of this methodology for transferability between different environments, the peculiarities and needs of each environment are reflected by the diversity of chambers developed over the years. Therefore, simply relying on experimental conditions that are being used in temperate areas to be applied in the tropics may bias the development of the technique. As OTC experimental research is promoted in the tropics, more data and results will be disseminated. Some limitations on using OTCs are the decoupling phenomenon and overheating inside chambers, which can be overcome with simple modifications by changing OTCs' structure, positioning, or using thermal masses.
Most plant response evaluated by OTC experiments in the tropics were physiological, morphological, and biochemical plant traits. We highlighted stomatal conductance, whole plant biomass, and starch content, which were more frequent. Surprisingly, Oryza sativa was the most studied species in the tropics, which indicates that OTC experiments are being conducted on crops due to their economic value instead of focusing on the conservation of tropical species and ecosystems. We could not observe a common tendency for plant response to climate change in the tropics probably due to the influence of several environmental factors together with the lack of studies with tropical wild species. However, we highlight a high capacity of tropical flora to deal with climate change and carbon stock noted with this review.
Tropical environments have highly complex dynamics, in which other factors, such as precipitation and nutrient availability, should also be considered to study plant response to climate change. So, we believe that it is necessary to expand the number of studies in the tropics for climate change doing in situ OTC experiments, especially in other tropical biomes less represented in this review, such as mangroves, grasslands savannas, and shrublands. Also, we urgently need to understand the response of endangered tropical species to climate predicted conditions for improvement of conservation status and management actions. Open-Top Chambers are an excellent experimental approach for in situ climatic alteration simulations since it does not require plant transplantation with low cost.
Conclusion
The tropics have unique environments, endowed with immense biotic and abiotic diversity. Preventing and mitigating the effects of climate change in these biomes require specific strategies and management due to their particularities. OTCs can be a valuable support tool to obtain the data needed so these objectives are achieved. They perform well in different tropical locations; in addition, they are versatile and accessible. Considering the advancement of research with the technique compared to temperate regions, more research is needed to explore the potential of OTCs in tropical ecosystems.
Data availability
All data supporting the findings of this study are available within the paper and its Supplementary Information.
References
AbdElgawad H, Zinta G, Beemster GT, Janssens IA, Asard H (2016) Future climate CO2 levels mitigate stress impact on plants: increased defense or decreased challenge? Front Plant Sci 7:556. https://doi.org/10.3389/fpls.2016.00556
Alzate-Marin AL, Teixeira SP, da Rocha-Filho LC, Bonifácio-Anacleto F, Rivas PMS, San Martin JA, Martinez CA (2021) Elevated CO2 and warming affect pollen development in a tropical legume forage species. Flora 283:151904. https://doi.org/10.1016/j.flora.2021.151904
Amirjani MR (2011) Effect of salinity stress on growth, sugar content, pigments and enzyme activity of rice. Int J Bot 7(1):73–81. https://doi.org/10.3923/ijb.2011.73.81
Andrew SC, Gallagher RV, Wright IJ, Mokany K (2022) Assessing the vulnerability of plant functional trait strategies to climate change. Glob Ecol Biogeogr 31(6):1194–1206
Aragon-Gastelum JL, Flores J, Yanez-Espinosa L, Badano E, Ramirez-Tobias HM, Rodas-Ortiz JP, Gonzalez-Salvatierra C (2014) Induced climate change impairs photosynthetic performance in Echinocactus platyacanthus, an especially protected Mexican cactus species. Flora-Morphology, Distribution, Functional Ecology of Plants 209(9):499–503. https://doi.org/10.1016/j.flora.2014.06.004
Aragón-Gastélum JL, Yáñez-Espinosa L, Ramírez-Albores JE, González-Salvatierra C, Flores J (2021) Seasonal ecophysiological variations of Echinocactus platyacanthus, a specially protected cactus species: Effect of induced climate warming. Glob Ecol Conserv. https://doi.org/10.1016/j.gecco.2021.e01919
Arenque BC, Grandis A, Pocius O, de Souza AP, Buckeridge MS (2014) Responses of Senna reticulata, a legume tree from the Amazonian floodplains, to elevated atmospheric CO2 concentration and waterlogging. Trees 28:1021–1034. https://doi.org/10.1007/s00468-014-1040-5
Avila RT, Cardoso AA, de Almeida WL et al (2020) Coffee plants respond to drought and elevated [CO2] through changes in stomatal function, plant hydraulic conductance, and aquaporin expression. Environ Exp Bot 177:104148
Bai Y, Cotrufo MF (2022) Grassland soil carbon sequestration: Current understanding, challenges, and solutions. Science 377(6606):603–608. https://doi.org/10.1126/science.abg5364
Baligar VC, Elson MK, He Z, Li Y, Paiva ADQ, Almeida AAF, Ahnert D (2021) Impact of Ambient and Elevated [CO2] in Low Light Levels on Growth, Physiology and Nutrient Uptake of Tropical Perennial Legume Cover Crops. Plants 10:193. https://doi.org/10.3390/plants10020193
Beer C, Reichstein M, Tomelleri E et al (2010) Terrestrial gross carbon dioxide uptake: global distribution and covariation with climate. Science 329:834–838. https://doi.org/10.1126/science.1184984
Bitencourt C, Rapini A, Damascena LS, Junior PDM (2016) The worrying future of the endemic flora of a tropical mountain range under climate change. Flora Morphol Distribut Funct Ecol Plants 218:1–10. https://doi.org/10.1016/j.flora.2015.11.003
Bokhorst S, Huiskes AD, Aerts R, Convey P, Cooper EJ, Dalen L et al (2013) Variable temperature effects of Open Top Chambers at polar and alpine sites explained by irradiance and snow depth. Glob Change Biol 19(1):64–74. https://doi.org/10.1111/gcb.12038
Broberg MC, Feng Z, Xin Y, Pleijel H (2015) Ozone effects on wheat grain quality–A summary. Environ Pollut 197:203–213. https://doi.org/10.1016/j.envpol.2014.12.032
Cavaleri MA, Reed SC, Smith WK, Wood TE (2015) Urgent need for warming experiments in tropical forests. Glob Change Biol 21:2111–2121. https://doi.org/10.1111/gcb.12860
Chiba M, Terao T (2014) Open-top chambers with solar-heated air introduction tunnels for the high-temperature treatment of paddy fields. Plant Prod Sci 17(2):152–165
Coley P, Massa M, Lovelock C, Winter K (2002) Effects of elevated CO2 on foliar chemistry of saplings of nine species of tropical tree. Oecologia 133:62–69. https://doi.org/10.1007/s00442-002-1020-7
D’Andrea L, Rinaldi M (2010) Systems to evaluate the effects of atmospheric CO2 concentration on field crops: a review of open top chambers. Ita J of Agrometeorology 1:23–34
De Frenne P, De Schrijver A, Graae BJ, Gruwez R, Tack W, Vandelook F, Hermy M, Verheyen K (2010) The use of open-top chambers in forests for evaluating warming effects on herbaceous understorey plants. Ecol Res 25:163–171. https://doi.org/10.1007/s11284-009-0640-3
Debb JC, Phinn S, Butt N, McAlpine CA (2018) Climate change impacts on tropical forests: identifying risks for tropical Asia. J Trop for Sci 30(2):182–194. https://doi.org/10.26525/jtfs2018.30.2.182194
Díaz S, Kattge J, Cornelissen JHC, Wright IJ, Lavorel S, Dray S, Reu B, Kleyer M, Wirth C, Colin Prentice I, Garnier E, Bönisch G, Westoby M, Poorter H, Reich PB, Moles AT, Dickie J, Gillison AN, Zanne AE, Gorné LD (2016) The global spectrum of plant form and function. Nature 529:167–171. https://doi.org/10.1038/nature16489
Dieleman WIJ, Vicca S, Dijkstra FA, Hagedorn F, Hovenden MJ, Larsen KS, Janssens IA (2012) Simple additive effects are rare: a quantitative review of plant biomass and soil process responses to combined manipulations of CO2 and temperature. Glob Chang Biol 18(9):2681–2693. https://doi.org/10.1111/j.1365-2486.2012.02745.x
Dray S, Dufour A (2007) The ade4 Package: Implementing the Duality Diagram for Ecologists. J Stat Softw 22(4):1–20. https://doi.org/10.18637/jss.v022.i04
Duran-Quesada AM, Reboita M, Gimeno L, Nieto R (2009) The role of the tropics in the global water cycle: Precipitation and moisture transport in Tropical America. Earth Observat Water Cycle Sci 674:34. https://doi.org/10.5194/essd-12-674-2020
El-Sharkawy MA (2014) Global warming: causes and impacts on agroecosystems productivity and food security with emphasis on cassava comparative advantage in the tropics/subtropics. Photosynthetica 52:161–178. https://doi.org/10.1007/s11099-014-0028-7
Esmail S, Oelbermann M (2011) The impact of climate change on the growth of tropical agroforestry tree seedlings. Agroforest Syst 83:235. https://doi.org/10.1007/s10457-011-9424-1
Esposito MP, Domingos M (2014) Establishing the redox potential of Tibouchina pulchra (Cham.) Cogn., a native tree species from the Atlantic Rain Forest, in the vicinity of an oil refinery in SE Brazil. Environ Sci Pollut Res 21(8):5484–5495. https://doi.org/10.1007/s11356-013-2453-8
Fauset S, Oliveira L, Buckeridge MS, Foyer CH, Galbraith D, Tiwari R, Gloor M (2019) Contrasting responses of stomatal conductance and photosynthetic capacity to warming and elevated CO2 in the tropical tree species Alchornea glandulosa under heatwave conditions. Environ Exp Bot 158:28–39. https://doi.org/10.1016/j.envexpbot.2018.10.030
Feng X, Porporato A, Rodriguez-Iturbe I (2013) Changes in rainfall seasonality in the tropics. Nature Clim Change 3:811–815. https://doi.org/10.1038/nclimate1907
Foden WB, Young BE, Akçakaya HR, Garcia RA, Hoffmann AA, Stein BA, Thomas CD, Wheatley CJ, Bickford D, Carr JA, Hole DG, Martin TG, Pacifici M, Pearce-Higgins JW, Platts PJ, Visconti P, Watson JEM, Huntley B (2019) Climate change vulnerability assessment of species. Wiley Interdiscipl Rev Clim Change 10:e551
Frei ER, Schnell L, Vitasse Y, Wohlgemuth T, Moser B (2020) Assessing the Effectiveness of in-situ Active Warming Combined With Open Top Chambers to Study Plant Responses to Climate Change. Front Plant Sci. https://doi.org/10.3389/fpls.2020.539584
Ganjurjav H, Gao Q, Zhang W, Liang Y, Li Y, Cao X et al (2015) Effects of warming on CO2 fluxes in an alpine meadow ecosystem on the central Qinghai-Tibetan Plateau. PLoS ONE. https://doi.org/10.1371/journal.pone.0132044
Godfree R, Robertson B, Bolger T, Carnegie M, Young A (2011) An improved hexagon open-top chamber system for stable diurnal and nocturnal warming and atmospheric carbon dioxide enrichment. Glob Change Biol 17(1):439–451. https://doi.org/10.1111/j.1365-2486.2010.02250.x
Griffin-Nolan RJ, Bushey JA, Carroll CJW et al (2018) Trait selection and community weighting are key to understanding ecosystem responses to changing precipitation regimes. Funct Ecol 32:1746–1756. https://doi.org/10.1111/1365-2435.13135
Heagle AS, Body DE, Heck WW (1973) An open-top field chamber to assess the impact of air pollution on plants. Am Soc Agron Crop Sci Soc Am Soil Sci Soc Am 2(3):365–368
Heyneke E et al (2012) Open-top chambers to study air pollution impacts in South Africa Part I: microclimate in open-top chambers. South Afr J Plant Soil 29(1):1–7
Hollister RD, Webber PJ (2000) Biotic validation of small open-top chambers in a tundra ecosystem. Glob Change Biol 6:835–842. https://doi.org/10.1046/j.1365-2486.2000.00363.x
Hollister RD et al (2022) A review of open top chamber (OTC) performance across the ITEX Network. Arct Sci First. https://doi.org/10.1139/as-2022-0030
Hollister, R.D. 1998. Response of Wet Meadow Tundra to Interannual and Manipulated Temperature Variation: Implications for Climate Change Research. Master’s Thesis, Michigan State University
Hyvönen R, Ågren GI, Linder S et al (2007) Tansley Review. The likely impact of elevated [CO2], nitrogen deposition, increased temperature and management on carbon sequestration in temperate and boreal forest ecosystems: a literature review. New Phytol 173:463–480. https://doi.org/10.1111/j.1469-8137.2006.01914.x
IPCC (2014) Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. IPCC, Geneva, Switzerland. https://www.ipcc.ch/site/assets/uploads/2018/02/SYR_AR5_FINAL_full.pdf. Accessed 4 of January of 2023
IPCC 2022. Climate Change 2022: Impacts, Adaptation, and Vulnerability. Contribution of 687 Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press
Ishii S, Marshall FM, Bell JNB, Abdullah AM (2004) Impact of ambient air pollution on locally grown rice cultivars (Oryza sativa L.) in Malaysia. Wat Air Soil Poll 154(1):187–201. https://doi.org/10.1023/B:WATE.0000022964.55434.05
Kassambara A, Mundt F (2020) Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. R Package Version 1:7
Kumar A, Nayak AK, Sah RP, Sanghamitra P, Das BS (2017) Effects of elevated CO2 concentration on water productivity and antioxidant enzyme activities of rice (Oryza sativa L.) under water deficit stress. Field Crop Res 212:61–72. https://doi.org/10.1016/j.fcr.2017.06.020
Lawrence D, Vandecar K (2015) Effects of tropical deforestation on climate and agriculture. Nature Clim Change 5:27–36. https://doi.org/10.1038/nclimate2430
Lewis SL (2006) Tropical forests and the changing earth system. Philos. Trans R Soc Lond B Biol Sci 361:195–210. https://doi.org/10.1098/rstb.2005.1711
Li W, Hartmann H, Adams HD, Zhang H, Jin C, Zhao C, Guan D, Wang A, Yuan F, Wu J, Tissue DT (2018) The sweet side of global change – dynamic responses of non-structural carbohydrates to drought, elevated CO2 and nitrogen fertilization in tree species. Tree Physiol 38:1706–1723. https://doi.org/10.1093/treephys/tpy059
Lie Z, Huang W, Liu X et al (2021) Warming leads to more closed nitrogen cycling in nitrogen-rich tropical forests. Glob Chang Biol 27(3):664–674. https://doi.org/10.1111/gcb.15453
Liu J, Zhou G, Xu Z et al (2011) Photosynthesis acclimation, leaf nitrogen concentration, and growth of four tree species over 3 years in response to elevated carbon dioxide and nitrogen treatment in subtropical China. J Soils Sediments 11(7):1155. https://doi.org/10.1007/s11368-011-0398-4
Lovelock CE, Winter K, Mersits R, Popp M (1998) Responses of Communities of Tropical Tree Species to Elevated CO2 in a Forest Clearing. Oecologia. https://doi.org/10.1007/s004420050502
Luo Y, Su B, Currie WS et al (2004) Progressive nitrogen limitation of ecosystem responses to rising atmospheric carbon dioxide. Bioscience 54(8):731–739. https://doi.org/10.1641/0006-3568(2004)054[0731:PNLOER]2.0.CO;2
Luo YQ, Hui DF, Zhang DQ (2006) Elevated CO2 stimulates net accumulations of carbon and nitrogen in land ecosystems: a meta-analysis. Ecology 87(1):53–63. https://doi.org/10.1890/04-1724
Lyra A, Imbach P, Rodriguez D et al (2017) Projections of climate change impacts on central America tropical rainforest. Clim Change 141:93–105. https://doi.org/10.1007/s10584-016-1790-2
Macháčová K (2010) Open top chamber and free air CO2 enrichment - approaches to investigate tree responses to elevated CO2. iForest 3:102–105. https://doi.org/10.3832/ifor0544-003
Marenco RA, Nascimento HCS, Magalhães NDS (2014) Stomatal conductance in Amazonian tree saplings in response to variations in the physical environment. Photosynthetica 52(4):493–500. https://doi.org/10.1007/s11099-014-0056-3
Martins LD, Tomaz MA, Lidon FC et al (2014) Combined effects of elevated [CO2] and high temperature on leaf mineral balance in Coffea spp. plants. Clim Change 126:365–379. https://doi.org/10.1007/s10584-014-1236-7
Marvin DC, Winter K, Burnham RJ, Schnitzer SA (2015) No evidence that elevated CO2 gives tropical lianas an advantage over tropical trees. Glob Change Biol 21(5):2055–2069. https://doi.org/10.1111/gcb.12820
Mills G, Buse A, Gimeno B, Bermejo V, Holland M, Emberson L, Pleijel H (2007) A synthesis of AOT40-based response functions and critical levels of ozone for agricultural and horticultural crops. Atmos Environ 41(12):2630–2643. https://doi.org/10.1016/j.atmosenv.2006.11.034
Min SK, Zhang X, Zwiers FW, Hegerl GC (2011) Human contribution to more-intense precipitation extremes. Nature 470(7334):378–381. https://doi.org/10.1038/nature09763
Mitchard ETA (2018) The tropical forest carbon cycle and climate change. Nature 559:527–534. https://doi.org/10.1038/s41586-018-0300-2
Munné-Bosch S, Queval G, Foyer CH (2013) The impact of global change factors on redox signaling underpinning stress tolerance. Plant Physiol 161(1):5–19. https://doi.org/10.1104/pp.112.205690
Nakazato RK, Rinaldi MC, Domingos M (2016) Tropical trees: Are they good alternatives for biomonitoring the atmospheric level of potential toxic elements near to the Brazilian Atlantic Rainforest? Ecotoxicol Environ Saf 134:72–79. https://doi.org/10.1016/j.ecoenv.2016.08.013
Oliveira VF, Silva EA, Zaidan LBP, Carvalho MAM (2013) Effects of elevated CO2 concentration and water deficit on fructan metabolism in Viguiera discolor Baker. Plant Biol 15(3):471–482. https://doi.org/10.1111/j.1438-8677.2012.00654.x
Olson DM, Dinerstein E, Wikramanayake ED, Burgess ND, Powell GV, Underwood EC, Kassem KR (2001) Terrestrial Ecoregions of the World: A New Map of Life on EarthA new global map of terrestrial ecoregions provides an innovative tool for conserving biodiversity. Bioscience 51(11):933–938
Padhy SR, Bhattacharyya P, Dash PK, Roy KS, Neogi S, Baig MJ, Mohapatra T (2020) Enhanced labile carbon flow in soil-microbes-plant-atmospheric continuum in rice under elevated CO2 and temperature leads to positive climate change feedback. Appl Soil Ecol. https://doi.org/10.1016/j.apsoil.2020.103657
Pan Y, Birdsey RA, Fang J et al (2011) A large and persistent carbon sink in the world’s forests. Science 333:988–993. https://doi.org/10.1126/science.1201609
Parida A, Das AB, Mittra B, Mohanty P (2007) Alterations in photosynthetic pigments, protein and osmotic components in cotton genotypes subjected to short-term drought stress followed by recovery. Plant Biotechnol Rep 1:37–48
Pendall E, Bridgham S, Hanson PJ et al (2004) Below-ground process responses to elevated CO2 and temperature: a discussion of observations, measurement methods, and models. New Phytol 162(2):311–322. https://doi.org/10.1111/j.1469-8137.2004.01053.x
Pérez-Ruiz CL, Badano EI, Rodas-Ortiz JP, Delgado-Sánchez P, Flores J, Douterlungne D, Flores-Cano JA (2018) Climate change in forest ecosystems: a field experiment addressing the effects of raising temperature and reduced rainfall on early life cycle stages of oaks. Acta Oecologica 92:35–43. https://doi.org/10.1016/j.actao.2018.08.006
Pillay T, Ward D (2022) Experimental drought suppresses grass productivity and passive warming promotes tree sapling performance: Insights from African savanna species. Acta Oecol. https://doi.org/10.1016/j.actao.2022.103813
Rai R, Agrawal M (2012) Impact of tropospheric ozone on crop plants. Proc Nat Acad Sci Ind Sect B Biol Sci 82(2):241–257. https://doi.org/10.1007/s40011-012-0002-7
Rai R, Agrawal M (2014) Assessment of competitive ability of two Indian wheat cultivars under ambient O3 at different developmental stages. Environ Sci Pollut Res 21(2):1039–1053. https://doi.org/10.1007/s11356-013-1981-6
Raubenheimer SL, Ripley BS (2022) CO2-stimulation of savanna tree seedling growth depends on interactions with local drivers. J Ecol 110:1090–1101. https://doi.org/10.1111/1365-2745.13863
RStudio Team (2020). RStudio: Integrated Development for R. RStudio, PBC, Boston, MA URL http://www.rstudio.com/
Samol P, Umponstira C, Klomjek P, Thongsanit P (2015) Responses of rice yield and grain quality to high temperature in open-top chamber to predict impact of future global warming in Thailand. Aust J Crop Sci 9(9):886–894
Saxena P, Srivastava A, Tyagi M, Kaur S (2019) Impact of tropospheric ozone on plant metabolism–a review. Pollut Res 38(1):175–180. https://doi.org/10.36468/pollut.2019.41
Singh S, Agrawal SB (2011) Cultivar-specific response of soybean (Glycine max L.) to ambient and elevated concentrations of ozone under open top chambers. Water Air Soil Pollut 217:283–302. https://doi.org/10.1007/s11270-010-0566-7
Singh P, Singh S, Agrawal SB et al (2012) Assessment of the interactive effects of ambient O3 and NPK levels on two tropical mustard varieties (Brassica campestris L) using open-top chambers. Environ Monit Assess 184(10):5863–5874. https://doi.org/10.1007/s10661-011-2386-y
Slot M, Winter K (2017) In situ temperature relationships of biochemical and stomatal controls of photosynthesis in four lowland tropical tree species. Plant Cell Env 40:3055–3068. https://doi.org/10.1111/pce.13071
Sreeharsha RV, Mudalkar S, Sengupta D, Unnikrishnan DK, Reddy AR (2020) Mitigation of drought-induced oxidative damage by enhanced carbon assimilation and an efficient antioxidative metabolism under high CO2 environment in pigeonpea (Cajanus cajan L.). Photosynth Res 139(1):425–439. https://doi.org/10.1007/s11120-018-0517-6
Stocker TF, Qin D, Plattner GK et al. (Eds.) (2014) Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of IPCC the Intergovernmental Panel on Climate Change. (Cambridge: Cambridge University Press). https://doi.org/10.1017/CBO9781107415324
Thalmann M, Santelia D (2017) Starch as a determinant of plant fitness under abiotic stress. New Phytol 214:943–951. https://doi.org/10.1111/nph.14491
Tiwari S, Rai R, Agrawal M (2008) Annual and seasonal variations in tropospheric ozone concentrations around Varanasi. Int J Remote Sens 29(15):4499–4514. https://doi.org/10.1080/01431160802036598
Wang J, Zhang L, Xie Y, Han X, Wang Y, Gao S et al (2013) Effects of elevated CO2 and nitrogen deposition on ecosystem carbon fluxes on the Sanjiang plain wetland in Northeast China. PLoS ONE. https://doi.org/10.1371/journal.pone.0066563
Way DA, Oren R (2010) Differential responses to changes in growth temperature between trees from different functional groups and biomes: a review and synthesis of data. Tree Physiol 30:669–688. https://doi.org/10.1093/treephys/tpq015
Winter K, Garcia M, Lovelock CE, Gottsberger R, Popp M (2000) Responses of model communities of two tropical tree species to elevated atmospheric CO2: growth on unfertilized soil. Flora 195:289–302. https://doi.org/10.1016/S0367-2530(17)30709-2
Wright IJ, Dong N, Maire V, Prentice IC, Westoby M, Díaz S, Gallagher RV, Jacobs BF, Kooyman R, Law EA, Leishman MR, Niinemets Ü, Reich PB, Sack L, Villar R, Wang H, Wilf P (2017) Global climatic drivers of leaf size. Science 357:917–921. https://doi.org/10.1126/science.aal4760
Yu X, Wu L, Zhang Y, Lin Q, Zhang Y, Li W et al (2019) Comparative analyses of wetland plant biomass accumulation and litter decomposition subject to in situ warming and nitrogen addition. Sci Total Environ 691:769–778
Zheng F, Wang X, Zhang W, Hou P, Lu F, Du K et al (2013) Effects of elevated O3 exposure on nutrient elements and quality of winter wheat and rice grain in Yangtze River Delta. China Environ Pollut 179:19–26. https://doi.org/10.1016/j.envpol.2013.04.010
Acknowledgements
The Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) has supported this work. The authors also thank Vitor G. Cunha for the map’s graphic production. They thank the assertive comments and suggestions of anonymous journal reviewers and the editor.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Material preparation, data collection were performed by Amanda Santos de Alencar, and Catarina da Fonseca Lira. Data analysis was performed by Amanda Santos de Alencar, Catarina da F. Lira, and reviewed by Bruno H. P. Rosado. The first draft of the manuscript was written by Amanda Santos de Alencar. Careful revisions on the manuscript were made by Vidal de F Mansanoand. All authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Communicated by Gerhard Leubner.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

{kind=link}
{kind=link}
Cite this article
de Alencar, A.S., da F Lira, C., Rosado, B.H.P. et al. Twenty-five years of Open-Top Chambers in tropical environments: where, how, and what are we looking at regarding flora response to climate change?. Planta 259, 82 (2024). https://doi.org/10.1007/s00425-024-04356-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00425-024-04356-8