
Abstract
Camptothecin (CPT), an indole alkaloid popular for its anticancer property, is considered the third most promising drug after taxol and famous alkaloids from Vinca for the treatment of cancer in humans. Camptothecin was first identified in Camptotheca acuminata followed by several other plant species and endophytic fungi. Increased harvesting driven by rising global demand is depleting the availability of elite plant genotypes, such as Camptotheca acuminata and Nothapodytes nimmoniana, crucial for producing alkaloids used in treating diseases like cancer. Conservation of these genotypes for the future is imperative. Therefore, research on different plant tissue culture techniques such as cell suspension culture, hairy roots, adventitious root culture, elicitation strategies, and endophytic fungi has been adopted for the production of CPT to meet the increasing demand without affecting the source plant’s existence. Currently, another strategy to increase camptothecin yield by genetic manipulation is underway. The present review discusses the plants and endophytes that are employed for camptothecin production and throws light on the plant tissue culture techniques for the regeneration of plants, callus culture, and selection of cell lines for the highest camptothecin production. The review further explains the simple, accurate, and cost-effective extraction and quantification methods. There is enormous potential for the sustainable production of CPT which could be met by culturing of suitable endophytes or plant cell or organ culture in a bioreactor scale production. Also, different gene editing tools provide opportunities for engineering the biosynthetic pathway of CPT, and the overall CPT production can be improved .
Key points
• Camptothecin is a naturally occurring alkaloid with potent anticancer properties, primarily known for its ability to inhibit DNA topoisomerase I.
• Plants and endophytes offer a potential approach for camptothecin production.
• Biotechnology approaches like plant tissue culture techniques enhanced camptothecin production.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Camptothecin (CPT) is a monoterpene indole alkaloid produced by plants as a secondary metabolite (Sadre et al. 2016). CPT has been a propitious chemotherapeutic drug since its discovery in 1966 by Wall and Wani (Malik and Laura 2014). CPT is used in combination with other chemotherapeutic drugs to enhance anti-cancerous properties with the further advantage of lesser side effects when compared to invasive chemotherapy (Nurgali et al. 2018). The drug is used for the treatment of metastatic cancer in organs such as the lung, breast, gastrointestinal tract, liver, gallbladder, spleen, and colon (Kamble et al. 2011). The drug exhibits its anticancer property by inhibiting the DNA topoisomerase I enzyme. DNA topoisomerase enzyme plays a predominant role in DNA replication, transcription, repair, and recombination processes (Ulukan and Swaan 2002). It is the third most promising anti-cancer drugs of the twenty-first century after taxol and vinca alkaloids (Mohinudeen et al. 2021). Figure 1 illustrates the inhibition of topoisomerase I activity by camptothecin.
CPT was first discovered in the Chinese deciduous tree, Camptotheca acuminata Decne. Later, the alkaloid was reported to be distributed among plant species of various families such as Nyssaceae, Icacinaceae, Loganiaceae, Apocynaceae, and Rubiaceae and their presence in endophytes such as Entrophospora infrequens, Fusarium solani, and Neurospora (Pu et al. 2019). The increasing market demand for CPT from these herbal plants has resulted in overharvesting and affected the existence of nativity (Niazian 2019; Greenwell and Rahman 2015). Thus, establishing alternative strategies for camptothecin production using biotechnological approaches provides a viable option. Plant tissue culture offers an alternative method for the conservation of the species. For the enhanced CPT production, different plant tissue culture techniques such as micropropagation, indirect and direct organogenesis, and hairy root culture have been employed from these plants (Malik and Laura 2014). Additionally, an effective strategy of genetic manipulation has been employed recently in enhancing CPT yield (Kai et al. 2015). Moreover, improved extraction methods have helped in isolating the compound efficiently. A simple, accurate, and cost-effective quantification method was developed for the quantification of CPT (Lokesh et al. 2014).
A good number of studies on camptothecin-producing plants and endophytes have been reported. However, there is no extensive documentation on various plant tissue culture methods and optimization strategies for enhanced camptothecin production from different plants and endophytes. There are no substantial reports covering the different extraction and quantification methods of camptothecin and its large-scale production. So, in view of this, the present review provides detailed information on the structure and function of a highly effective anticancer drug, CPT. It reports the plants and endophytes that are utilized for CPT production and discusses the alternative methods for CPT production over conventional methods, which are robust and cost-effective. The alternative approaches include callus, cell suspension, shoot, and root cultures (adventitious root and hairy root). Further, the review discusses the optimization strategies used to enhance the camptothecin production with optimization of media and culture conditions. Elaborative details on the use of bioreactor for the large-scale production of CPT and summary on the current status of metabolic engineering of CPT biosynthetic pathways have been discussed.

Mechanism of inhibition of Topoisomerase I activity by camptothecin
Camptothecin: structure and its derivatives/analogues
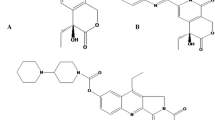
CPT is a pentacyclic alkaloid that was first discovered in the early 1960s (Malik and Laura 2014). The CPT structure comprises a pyrrolo (3,4-β) quinoline moiety, a conjugated pyridone, and an asymmetric center at the 20th position within the α-hydroxy lactone ring with 20 (S) configuration. The pentacyclic ring system consisting of A, B, C, D, and E rings majorly contributes to the topoisomerase inhibition activity of CPT (Kamble et al. 2011). Since its structural elucidation in 1966, CPT has undergone evolution through structural modifications and several CPT derivatives have been generated. These analogues were obtained based on the structural-activity relationships (SAR) which include Topotecan, Irinotecan, Etirinotecan, pegol, Belotecan, Rubitecan, Diflomotecan, Lurtotecan, and Exatecan. Currently, three water soluble anticancer analogues of CPT, Irinotecan, Topotecan, and Belotecan have been approved and are commercially available for cancer treatment (Li et al. 2017). Figure 2 illustrates the structure of camptothecin and its derivatives.
Topotecan
9-Dimethylaminomethyl-10-hydroxycamptothecin (Topotecan) was developed by the National Cancer Institute in collaboration with the University of Florida at Gainesville and SmithKline Beecham headed by Dr. Warren Ross in the 1980s. It was first approved by FDA in 1996 and is now manufactured by GlaxoSmithKline and commercially available as Hycamtin (developmental therapeutics program). It is synthesized from 10-hydroxy-20-(S)-camptothecin or with dimethylamine by aminomethylation in the presence of catalyst trihalomethane (Puri et al. 2003). Topotecan is used in the treatment of ovarian cancer, small cell and non-small cell lung cancers, non-Hodgkin lymphoma, endometrial cancer, oligodendroglioma, and breast cancer.
Irinotecan
7-Ethyl-10-[4-(1-piperidino)-1-piperidino] carbonyl oxy-camptothecin (Irinotecan), commonly available under the brand name Camptosar, is prepared by ethylating 10-[4-(1-piperidino)-1-piperidino] carbonyloxy-camptothecin at the 7th position (Zabudkin 2015). Irinotecan was first developed in 1983 by Yakult Honsha Co., Ltd., Japan. In 1994, it was first approved in Japan for its use (Fukuoka 2001). Irinotecan is used in the treatment of small-cell lung cancer, colon cancer, ovarian cancer, acute and lymphoblastic leukemia, and non-Hodgkin’s lymphoma (Kamble et al. 2011).
Belotecan
(20 S)-7-(2-isopropylamino)-ethyl-camptothecin commercially available as Camtobell marketed by Chong Kun Dang Corp. (Seoul, Korea) was first prepared by Ahn and coworkers of Chong Kun Dang in 1999 (Ahn et al. 2000). It has DNA topoisomerase inhibition activity in cancer cells. Belotecan is synthesized in a two-step reaction: Minisci type reaction, which involves conversion of CPT 7-methylcamptothecin, and Mannich type reaction, which involves conversion of 7-methyl CPT to belotecan (Liew and Yang 2008). Belotecan is used in the treatment of small-cell lung cancer and ovarian cancer (Liew and Yang 2008).

Structure of camptothecin and its derivatives/analogues (Martino et al. 2017)
Camptothecin distribution in plants
Earlier, CPT was limited to only two plants: C. acuminata of the Nyssaceae family and Nothapodytes nimmoniana from the Icacinaceae family. However, the increased CPT demand led to several research studies to identify alternative CPT-producing plants. At present, CPT is found to be distributed among plant species of various plant families such as Nyssaceae, Icacinaceae, Loganiaceae, Apocynaceae, and Rubiaceae (Ulukan and Swaan 2002). The seeds of C. acuminata and N. nimmoniana have reported the presence of 0.110% CPT (Liu and Adams 1996) and 0.179% CPT (Isah and Mujib 2015a), respectively. Some of the Ophiorrhiza species such as O. fucosa, O. plumbea, O. ridleyana, and O. harrisiana have reported the presence of camptothecin in them (Viraporn et al. 2011). The highest CPT content of 1.418% has been reported in the seeds of Miquelia dentata Bedd. (Ramesha et al. 2013). For the production of one ton of camptothecin, nearly 1000–1500 tons of plant material is required. The annual marketing sales of camptothecin and its derivatives have been estimated to be $1 billion (Shrivastava et al. 2021). Table 1 presents the parts of the various plant species from which CPT has been isolated.
Camptothecin distribution in the endophytes
Endophytes are the microorganisms like bacteria or fungi that coexist with a living plant and are reported to be the source for antibiotics, antiviral compounds, anti-diabetic agents, anticancer agents, and many more. In recent times, they serve as an alternative source to produce plant secondary metabolites such as taxol, camptothecin, capsaicin, rohitukine, and several other such compounds (Uzma et al. 2018). The fungal species of Aspergillus, Trichoderma, Fomitopsis. Phomposis, and Fusarium have been reported to produce CPT (Malik and Laura 2014). Three CPT-producing fungi T atroviridae LY357, Aspergillus sp. LY341, and Aspergillus sp. LY355 isolated from barks, twigs, leaves, and fruits of C. acuminata. were 7.93, 42.92, and 197.82 µg L−1, respectively (Pu et al. 2013). F. oxysporum kolhapuriensis from the Nectriaceae family reported the presence of 283 mg L−1 CPT (Bhalkar et al. 2015). Table 2 presents the production of CPT from different endophytes.
Biosynthetic pathway of camptothecin
The biosynthetic pathway for CPT includes three steps: the pre-strictosidine pathway, strictosidine synthesis, and post-strictosidine pathway. Over the past few decades, numerous biochemical investigations have been carried out in CPT for its enhanced production due to its potential anticancer activity. A metabolic engineering approach is currently adopted for the enhanced CPT production wherein the intermediates involved in the biosynthetic pathway are targeted. Thus, it is necessary to have an in-depth understanding of the CPT biosynthetic pathway (Gonçalves and Romano 2018). Figure 3 illustrates the biosynthetic pathway of CPT.
Pre-strictosamide pathway
Tryptophan is initially synthesized from chorismate by the shikimate pathway. The chorismate in the presence of enzyme anthranilate synthase converts into anthranilate, which then combines with 5-phosphoribosyl pyrophosphate to form indole glycerol phosphate. Indole is then formed by the addition of α- subunit of tryptophan synthase (TSA) to indole glycerol phosphate, which is then condensed with β- subunit of tryptophan synthase (TSB) to form tryptophan. The tryptophan is decarboxylated to tryptamine by the tryptophan decarboxylase enzyme. Parallelly, secologanin is synthesized from IPP (isopentenyl diphosphate) and its isomer DMAPP (dimethylallyl diphosphate) both of which are intermediates of the 2 C-methyl-D-erythritol-4-phosphate (MEP) pathway and MVA (mevalonate) pathway. IPP and DMAPP condense to form geranyl diphosphate (GPP), which is then converted to geraniol by geraniol synthase. Geraniol is converted to 10-hydroxygeraniol by geraniol 10-hydroxylase and further converted to loganin. Secologanin is synthesized from loganin by secologanin synthase (SLS) (Sirikantaramas et al. 2013).
Strictosidine synthesis
The tryptamine and secologanin condenses to form strictosidine. This Pictet-Spengler condensation between tryptamine and secologanin is catalyzed by strictosidine synthases (STR) ((Yamazaki et al. 2003).
Post-strictosidine pathway
In this pathway, CPT is synthesized in a multistep reaction. The strictosidine is converted to strictosamide by undergoing intramolecular cyclization. The strictosamide is converted into pulmioside and deoxypulmioside, ultimately forming camptothecin. The conversion of strictosamide to camptothecin involves oxidation and recyclization of the B and C rings, further oxidation of the D ring and removal of C-21 glucose moiety and final oxidation of the E ring forming camptothecin (Sirikantaramas et al. 2013).

The biosynthetic pathway of camptothecin involves the pre-strictosidine, strictosidine synthesis, and post-strictosidine pathways. TIA, monoterpenoid indole alkaloid; MEP, 2 C-methyl-D-erythritol-4-phosphate; STR, strictosidine synthases. The multiple step reaction is represented by an arrow with a dotted shaft (Sirikantaramas et al. 2013)
Production of CPT through plant cell, tissue, and organ culture
Plant tissue culture methods serve as the appropriate alternatives for CPT production to overcome the reduction in the natural population of the plants due to overharvesting. Plant tissue culture techniques involve mass propagation of plants from any part of the mother plant under in vitro conditions without seasonal constraints throughout the year. Moreover, these techniques help in easier isolation and purification of desired secondary metabolites. The secondary metabolites can be produced from an undifferentiated mass of cells, calli, cell suspension cultures and from differentiated shoots, roots (adventitious and hairy roots), or somatic embryos and such plant tissue culture strategies have been adopted for enhancement of camptothecin production (Gonçalves and Romano 2018). Figure 4 demonstrates the production of camptothecin by various tissue culture methods.

Production of CPT from various tissue culture techniques
Production of CPT through callus culture and selection of cell lines
The in vitro production of secondary metabolites occurs in two phases: biomass accumulation and secondary metabolite biosynthesis. Both these phases need to be optimized by utilizing a variety of tissue culture strategies (Chandran et al. 2020). Selection of high secondary metabolite yielding cultivars and screening the high secondary metabolite yielding cell lines are the key strategies for increasing secondary metabolite production. The explants are chosen from the high secondary metabolite yielding genotype of the plant. The callus is induced in such explants by exogenous application of auxin or cytokinin or in a defined ratio. The secondary metabolites can be obtained from callus cultured under optimal culture conditions. Furthermore, the callus can be subjected to cell suspension culture for developing fast-growing, high-yielding cell lines (Schreiner 2005).
The explant may exhibit a wide range of metabolic productivity. The heterogeneity would result in decreased secondary metabolite production. These undesirable variations in the production can be avoided or reduced by selecting potentially high-yielding cell populations from heterogeneous cultures. The cloning of such high-yielding cell lines helps in enhanced secondary metabolite production (Smetanska 2008). The effect of different concentrations and combinations of kinetin (KN) and auxin ranging from 0.1 to 10 mg L−1 on callus induction of C. acuminata has been evaluated and reported by Van Hengel et al. (1994). The highest callus biomass was obtained in the MS (Murashige and Skoog medium, 1962) medium supplemented with B5 vitamins, 0.5 mg L−1 2,4-Dichlorophenoxyacetic acid (2,4-D) and 0.1 mg L−1 KN with a CPT production of 0.000098% (van Hengel et al. 1992). The cell suspension culture of Ophiorrhiza eriantha Wight was established from 16 g friable callus cultured on MS medium with 3% sucrose and 4 mg L−1 indole-3-butyric acid (IBA) showed the highest CPT production of 0.087% (Rani 2011). Induction of callus from hypocotyl explant was favorable when compared to leaf explant. It was observed that 2 mg L−1 TDZ along with 0.5 mg L−1 2,4-D was favorable for induction of callus without further organogenesis. Callus induced on 2 mg L−1 TDZ along with 0.5 mg L−1 2,4-D medium showed two-fold increase when compared to that of callus induced on 1mg L−1 TDZ along with 0.5 mg L−1 2,4-D (Kadam et al. 2023). Table 3 illustrates the CPT production by callus culture and cell suspension cultures of various CPT-producing plants.
Organ culture for CPT production
Secondary metabolite synthesis is often higher in differentiated tissues and organ cultures, and this has been developed as an alternative method for secondary metabolite production. The organ culture method involves the culture of organs such as root, shoot, and embryo and is the most stable method (Gonçalves and Romano 2018). In medicinal plants, shoot cultures have been established for a higher accumulation of secondary metabolites. Similarly, root cultures also serve as an alternative for root-derived secondary metabolites, which are otherwise difficult to harvest from a root system that exhibits slow growth. On the other hand, hairy root cultures have shown to be a viable method for secondary metabolite synthesis in vitro. The hairy roots can be induced by co-culturing the explant with Agrobacterium rhizogenes. The T-DNA from the plasmid of A. rhizogenes transferred to host tissue induces hairy root formation on account of auxin synthesis genes coded by bacterial DNA. Thus, it avoids the need for an external supply of auxins (Rao and Ravishankar 2002). In the study conducted by Vineesh et al. (2007), a maximum number of shoots were initiated from the leaf of O. rugosa, and the highest amount of CPT (0.039%) in multiple shoots was obtained in media with 3% sucrose, 5 mg L−1 BAP, and 0.5 mg L−1 NAA (Vineesh et al. 2007). It has been reported that the MS media with IBA induces 4–6 adventitious roots/shoot and the media with IBA or NAA alone or NAA combination with BAP or KN produced the wound in the shoot with A. rhizogenes 15834 and cultured on B5 medium with 2% sucrose, 0.2% of gellan gum and 200 mg L−1 cefotaxime for 7 weeks (Isah and Mujib 2015b). The highest CPT of 471 µg was obtained in hairy roots of Ophiorrhiza liukiuensis Hayata (Asano et al. 2004). Table 4 illustrates the total CPT content produced in root and shoot cultures of various CPT-producing plants.
Optimization strategies employed for improved production of CPT
The increasing market demand for secondary metabolites has led to the implementation of traditional and advanced metabolic strategies for incessant high yield production of secondary metabolites (Hussain et al. 2012). The optimization for secondary metabolites can be done in bioaccumulation stages and secondary metabolite synthesis stages. The important parameters such as pH, temperature, light intensity, carbon, and nitrogen source can be optimized at the bioaccumulation stage, and strategies like elicitation, precursor feeding, and immobilization can be achieved at the secondary metabolite biosynthetic stage (Gonçalves and Romano 2018).
Optimization of culture conditions
The supply of media with carbon, nitrogen, and phosphate sources at optimum concentration influences the secondary metabolite production. These nutrient sources play an important role in signal transduction, thereby regulating the expression of secondary metabolite genes as well as in biomass accumulation. The physical factors like pH, light intensity, temperature, and agitation speed also influence the biomass and metabolites productivity. Thus, choosing the right culture medium formulation is a vital step (Gonçalves and Romano 2018). The effect of sugar concentration on CPT production in cell suspension culture of C. acuminata has been studied using different concentrations (2, 4, 6, 8, and 10%) of sucrose, glucose, and fructose in the source media. The highest camptothecin (0.0029 mg L−1) was obtained at 6% sucrose concentration (Kim et al. 1999). The effect of nitrogen source on CPT production was studied in N. nimmoniana by supplementing the medium with different concentrations of potassium nitrate and ammonium chloride ranging from 60/0 to 0/60 mM NO3−/NH4+ balance. The best combination was found to be 50/1 mM NH4+/NO3− balance, with 0.5 mM phosphate on day 15 yielding 0.00474% CPT (Karwasara and Dixit 2013). The cell suspension culture of C. acuminata was subjected to pH varying from 4 to 7.5. It was also subjected to two different temperatures 26 and 30 °C with shaking rates of 148 and 184 rpm. The highest biomass was achieved at pH of 4.5, at 30 °C and 184 rpm (Sakato and Misawa 1974). The callus of C. acuminata when subjected to different light intensities of 400 W dysprosium lamps ranging from 0 to 100% irradiance, it was observed that highest CPT production of 3.56 mg g−1 was achieved at 50% light intensity (Hu et al. 2016). Table 5 tabulates the different optimization strategies that have been adopted for enhanced CPT production.
Elicitation
Elicitation is another strategy that can be adopted to improve camptothecin production. Elicitors mimic biotic/abiotic attacks, thus eliciting defense mechanisms in plants. The elicitors induce gene upregulation in CPT-producing plants which govern a wide range of cellular activities at the molecular and biochemical levels. Elicitors are classified as abiotic or biotic elicitors. The cell wall fragments of bacteria virus, fungus, enzymes, and molecules such as jasmonic acid (JA) and salicylic acid (SA) are biotic elicitors, and inorganic salts, heavy metals, and physical factors like UV radiation are types of abiotic elicitors (Gonçalves and Romano 2018). CPT can be elicited by elicitors like ferulic acid, methyl jasmonate (MeJA), jasmonic acid (JA), gamma irradiation, and yeast extract (YE) (Song and Byun 1998). The cell suspension culture O. mungos was amended with different concentrations of YE ranging from 25 to 200 mg L−1 and AgNO3 ranging from 2.0 to 7.5 µM. The highest CPT yield of 0.8 mg L−1 and 0.52 mg L−1 was achieved at 50 mg L−1 of YE and 2.5 µM of AgNO3, respectively (Deepthi and Satheeshkumar 2016). The effect of gamma radiation elicitation on CPT enhancement in callus cultures of N. nimmoniana was studied by irradiating with gamma radiation ranging from 5 to 30 Gy. The enhanced production of CPT in the callus culture was irradiated with 20 Gy radiation yielding 0.098% CPT (Fulzele et al. 2015). Recent reports also suggest that supplementation of yeast extract and glycine helped in increased production of camptothecin from callus cultures of Chonemorpha fragrans (Ambujakshi et al. 2022). Table 6 shows the different types of elicitation methods for CPT enhancement in plants.
Precursor feeding
Precursor feeding has been a well-known and widely used method for increasing secondary metabolite production in plant cells. The basic working principle of precursor feeding is that the intermediate compounds involved in the biosynthetic pathway of secondary metabolites have a fair possibility of improving the yield of the final product. Thus, attempts have been made to enhance secondary metabolites by identifying and supplementing the culture media with precursors. The concentration, time of addition, and type of the precursor should be taken into account for precursor feeding (Rao and Ravishankar 2002). CPT production can be enhanced by supplementing precursors such as tryptamine, loganin, and secologanin which are the intermediate compounds involved in biosynthetic pathways (Silvestrini et al. 2002). E. infrequens, the endophytic fungus of N. nimmoniana, has been treated with different precursors such as tryptophan, tryptamine, citral, geraniol, leucine, and mevalonic acid either alone or in combination with tryptophan. The highest CPT content of 0.000503% was observed in the sabouraud medium with tryptophan and leucine (Amna et al. 2012). The effect of precursors such as tryptamine and secologanine of varying concentrations of 10, 50, and 100 µM in cell suspension culture of O. eriantha has been studied. The enhanced CPT production of 0.00914% and 0.00843% DW was achieved at 50 µM concentration of secologanine and tryptamine, respectively (Rani 2011). Entrophospora infrequens, a fungus isolated from N. nimmoniana, produced the CPT content 0.8−1 mg g−1 when fed with combination of two precursors- tryptophan + leucine (Amna et al. 2012). Table S1 shows the different types of precursors used for CPT enhancement in plants.
Immobilization
Immobilization of plant cells is a new strategy achieved by encapsulating the plant cell within a solid support. This technique involves the use of hydro colloidal gels such as agarose, calcium alginate, carrageenin, gelatin, and polyacrylamide which entraps the plant material within it. Immobilization plays a crucial role in enhanced production of high value secondary metabolites. It makes it possible for a group of cells to work together at the same time and ensures continual production of the desired metabolites without cell washout, thus increasing the cell’s productivity. The low yield of CPT in the culture medium of large-scale bioreactors has been a major concern. Thus, immobilization of plant cells is employed for the increased production of camptothecin in large scale bioreactors. Mamkulathil Devasia et al. (2021) reported the callus of O. mungos has been immobilized for continuous production and to achieve high yield of CPT. It was found that the immobilized callus of O. mungos produced 420 µg L−1 CPT (Mamkulathil Devasia et al. 2021).
Omics approaches
Omics approaches such as genomics, transcriptomics, proteomics, and metabolomics can be employed to study and understand the biosynthetic pathway of camptothecin production and the reactions of enzymes that take part in the pathway at gene and protein levels. Genomics helps in the identification and characterization of the candidate genes involved in camptothecin biosynthesis by comparing the genomes of camptothecin-producing plants with non-producing ones. Kang et al. (2021) have obtained a high-quality genome assembly of C. acuminata using single-molecule real-time long reads technique (Pacific Biosciences (PacBio) Sequel platform and high-throughput chromosome conformation capture (Hi-C), with which they have further investigated the evolution of camptothecin biosynthesis. It was discovered that C. acuminata underwent a whole-genome duplication event, resulting in the emergence of genes involved in camptothecin production. Notably, it was observed that C. acuminata lacks a specific enzyme called loganic acid O-methyltransferase (LAMT), but instead has two secologanic acid synthases (SLASs) that convert loganic acid to secologanic acid. The functional divergence of the LAMT gene and positive evolution of two SLAS genes, therefore, contributed to C. acuminata’s effective production of camptothecin (Kang et al. 2021). Parallely, in O. pumila, metabolite profiling revealed that 3α-(S)-strictosidine, rather than 3-(S), 21-(S)-strictosidinic acid, is the exclusive intermediate involved in CPT biosynthesis (Yang et al. 2021).
In another study by Natarajan et al. (2023), the genomic DNA of Alternaria burnsii NCIM 1409 was isolated and sequenced on an Illumina NextSeq500, while the RNA was isolated and the transcriptome analysis was performed with RNA seq. The genome assembly and annotation revealed the presence of candidate genes involved in camptothecin biosynthesis. Comparative genomics analyses with related fungi were further conducted. The study concluded that there was no evidence of horizontal gene transfer from the host plant to the endophyte (Natarajan et al. 2023). In C. acuminata treated with elicitors such as MeJa, AgNO3, and PEG, 32 genes involved in CPT biosynthesis and 12 CYP450 genes that play a crucial role in the previously unexplored oxidation steps of CPT synthesis were explored (Pu et al. 2022). Genome-wide identification was employed to identify 8 out of 198 APETALA2/ethylene-responsive factor (AP2/ERF) transcription factor genes have been identified to be involved in CPT synthesis regulation with higher level of expression in immature bark and upper stem (Hu et al. 2020). The proteomics and transcriptomic studies in C. acuminata have discovered three O-methyltransferases and five cytochrome P450s that involve in camptothecin biosynthesis and 15 transcription factors that regulate CPT biosynthesis (Zhang et al. 2023).
Metabolic engineering of CPT biosynthesis
Biosynthesis and biotechnological production of CPT have made much progress in recent times. Metabolic engineering is one such approach that has enhanced CPT production. It is a biotechnological discipline that deals with the manipulation of the genes that code enzymes which take part in the biosynthetic pathways. The biosynthetic genes involved in the synthesis of strictosidinic acid and CPT-derivatives have been partially resolved and identified. These genes include CaG10H, Ca10HGO, CaIS, CaSLAS, CaTDC, CaSTRAS, and Ca10OMT. Understanding the functions of these genes allows researchers to precisely manipulate the biosynthetic pathway and enhance CPT production (Fan et al. 2022). Through metabolic engineering, CPT production can be enhanced either by overexpressing the genes that encode the enzymes that are involved in the biosynthesis of CPT or by inhibiting the competitive pathways in turn enhancing the metabolic flux of targeted biosynthetic pathways. Several metabolic engineering studies have been conducted for CPT enhancement in the past decade.
In the study by Cui et al. (2015), the co-overexpression of strictosidine synthase (STR) and geraniol 10-hydroxylase (G10H) genes from C. roseus introduced in O. pumila yielded 1.77 mg g−1 of CPT, i.e., 56% increase (Cui et al. 2015). Furthermore, study by Van der Fits and Memelink (2000) showed that overexpression of ORCA3 (Octadecanoid-derivative Responsive Catharanthus AP2-domain) from C. roseus in C. acuminata hairy roots enhance CPT production by 1.5-fold compared by up-regulating the expression key genes involved in terpenoid indole alkaloid (TIA) biosynthetic pathway (van der Fits and Memelink 2000). Metabolic engineering of CrORCA3 genes in C acuminata yielded 0.112% of CPT (Ni et al. 2011). The OpWRKY2 gene, OpSLS (secologanin synthase), OpG10H, STR genes of O. pumila, and CrG10H (genes from Catharanthus roseus inserted in O. pumila) on overexpression yielded 0.00248% (Hao et al. 2021), 0.328%, 0.240% (Shi et al. 2020), 0.177, and 0.128%, respectively (Cui et al. 2015). Furthermore, the overexpression of the NfSTR gene of O. rugosa yielded 0.213% of CPT (Singh et al. 2020). In O. pumila, it has been discovered that the transcription factor OpWRKY6 plays a key role in regulating camptothecin biosynthesis. Overexpression of OpWRKY6 reduced camptothecin levels, while its knockout resulted in increased camptothecin production, providing valuable insights for enhancing camptothecin production in plants (Wang et al. 2022). In O. pumila, the knockout of OpLAMT1 expression led to camptothecin expression, and further OpNAC1 (NAC transcription factor) was demonstrated to suppress the expression of OpLAMT1 and identified as a candidate gene for CPT production (Hao et al. 2023).
Extraction and quantification of camptothecin
Extraction
The extraction method is one of the most imperative methods for the recovery of CPT from its source. It plays a crucial role in the estimation of CPT obtained from various sources. For the extraction of CPT, the development of simple, faster, accurate extraction methods that require a lesser amount of solvents is important. Various extraction methods such as stirring extraction, soxhlet extraction, and microwave extraction have been developed for the extraction of camptothecin (Fulzele and Satdive 2005). Sonication is one of the most commonly used extraction methods. Camptothecin ranging from 0.85 to 3.6% and 0.15 to 0.23% has been extracted by sonication method from callus cultures of C. acuminata (van Hengel et al. 1994) using 18 mL water and 20 mL dichloromethane solvent, and from O. mungos (Nagesha et al. 2018) using 50 mL of methanol, respectively. 0.014% and 0.008–0.0096% of CPT were extracted from cultures of O. mungos (Krishnan et al. 2018) and rugosa var. decumbens (Roja 2006) using methanol solvent by soxhlet extraction method, respectively. 0.51% of CPT has been extracted from I. coccinea by water bath extraction method using 61% of methanol by incubating for 3 h at 45 °C (Saravanan and Boopalan 2011). The cultured endophytes have been subjected to different extraction methods such as solvent extraction, water bath extraction, and ultrasonication. The highest CPT content of 283 mg L−1 has been extracted by ultrasound-assisted extraction from F. oxysporum kolhapuriensis isolated from N. nimmoniana (Bhalkar et al. 2015). Table S2 illustrates the different extraction methods for CPT from different plant cultures and endophytes.
Quantification methods
Quantification of CPT in different plant sources using advanced techniques provides a viable option to determine the possible accurate amounts of CPT production in the plant species, microbial cultures, and in vitro cells. Among these, HPLC is one of the most commonly used methods for CPT estimation. Studies have been conducted on quantification of CPT by HPTLC and TLC methods (Lokesh et al. 2014; Hashim et al. 2016). The CPT content of 0.85–3.6% and 0.70–2.62% was quantified using HPLC-Waters model 510 with acetonitrile: water (25:75) solvent system at 1 mL min−1 flow rate (van Hengel et al. 1994), and 0.024–0.030% of CPT has been quantified by using HPLC Jasco PU 2080 in C. acuminata (Namdeo and Sharma 2012). About 0.010–0.084% of CPT has been quantified in N. nimmoniana using TLC plate coated with silica gel 60F254 pre-coated (20 × 20 cm) using chloroform: ethyl acetate (1: 1) as solvent system (Lokesh et al. 2014). Recently, a new RP HPLC method (90:10; Acetonitrile: Water as the mobile phase, 1 mL/min flow rate at 30 °C) has been developed for estimation of camptothecin in mixed micelles (CPT, PF108, and TPGS). The recovery of camptothecin was determined to be between 98 and 102%, showing that the method proposed is reliable (Patil et al. 2022). Table S2 illustrates the different quantification methods of CPT in different plant cultures and endophytes.
Bioreactor studies for the production of CPT
The tremendous market demand requires increased production of CPT. Owing to this, efforts have been made to study the potentiality of its production at a large scale. Thus, the adoption of bioreactor technology has served to enhance CPT production. However, it is one of the most challenging possibilities because of the unstable productivity, slow growth rate, high shear sensitivity, and low oxygen requirement in the plant cells. Bioreactors are highly efficient, predictable, and enable the easy harvest of metabolites from biomass or cultivation media. At present, bioreactors are specially designed for plant tissue cultures which are different from conventional bioreactors. The high shear-stress-sensitive cells are cultured in wave reactors, slug bubble reactors, and undertow reactors, while less shear-sensitive cells are cultured in airlift bioreactors. The optimization of culture conditions and the measurement of biomass productions are the factors that influence secondary metabolite production in these bioreactors (Gonçalves and Romano 2018). About 16.5% camptothecin has been produced on a large scale from hairy roots culture of C. acuminata in a 3 L bioreactor (length 235 mm; diameter, 140 mm) with 5-mm stainless steel mesh at 25 °C; 0.25/min aeration rate (Sudo et al. 2002). 0.0045% of CPT has been produced from Nodulisporium isolated from N. nimmoniana cultured in the bioreactor with 18 L working volume, maintained at an aeration rate of 1 vvm, 0.2 kg/cm pressure, 28 °C, and an agitation rate of 220 rpm (Rehman et al. 2009). Table S3 illustrates the bioreactor studies for large-scale production of CPT by plants and endophytes.
Conclusion and prospects
The major sources of camptothecin are the plants, for which a wide variety of valuable plants have been overexploited to meet the increased market demand, and that has resulted in their depletion. Clonal propagation does not yield satisfactory amounts of CPT. Thus, plant tissue culture techniques are the alternative strategies for the sustainable production of CPT rather than overharvesting of the plants, and these techniques serve as an alternative system for sustainable and economical production of camptothecin throughout the year irrespective of climatic conditions. The current review has made attempts to investigate the various plants which are the sources for CPT alkaloid, various endophytes that can serve as a new source for CPT production, plant tissue culture strategies, and the optimization of the culture conditions for both the plants and endophytes. During this exploration, it is evident that there is a crucial need to develop novel techniques for plant tissue culture and refined extraction techniques to increase the production and extraction of such metabolites that are produced in minute quantities in plant parts. Currently, CPT production can be enhanced by using bioreactors in addition to the use of elicitors and precursors. However, in some cases, CPT is produced at very low yields due to limited information about the biosynthetic pathways and the enzymes and genes involved in these pathways. Hence, identifying and understanding the functions of candidate genes involved in the biosynthetic pathways and their engineering using modern biotechnological approaches provides a viable option to enhance CPT production. CRISPR/Cas9 system can be used in the manipulation of the genes that control overexpression of enzymes involved in biosynthetic pathways of CPT and the knockout of genes that are involved in competing pathways.
References
Ahn SK, Choi NS, Jeong BS, Kim KK, Journ DJ, Kim JK, Lee SJ, Kim JW, Hong C II, Jew S-S (2000) Practical synthesis of (S)-7-(2-isopropylamino)ethylcamptothecin hydrochloride, potent topoisomerase I inhibitor. J Heterocycl Chem 37:1141–1144
Ambujakshi NP, Ravikiran T, Raveesha HR (2022) Influence of elicitors on the enhancement of camptothecin accumulation and antioxidant potential in callus cultures of Chonemorpha fragrans. S Afr J Bot 150:225–232
Amna T, Puri SC, Verma V, Sharma JP, Khajuria RK, Musarrat J, Spiteller M, Qazi GN (2006) Bioreactor studies on the endophytic fungus Entrophospora infrequens for the production of an anticancer alkaloid camptothecin. Can J Microbiol 52:189–196
Amna T, Amina M, Sharma PR, Puri SC, Al-Youssef HM, Al-Taweel AM, Qazi GN (2012) Effect of precursors feeding and media manipulation on production of novel anticancer pro-drug camptothecin from endophytic fungus. Braz J Microbiol 43:1476–1490
Arbain D, Putra DP, Sargent MV (1993) The alkaloids of Ophiorrhiza filistipula. Aust J Chem 46:977–985
Arisawa M, Gunasekera SP, Cordell GA, Farnsworth NR (1981) Plant anticancer agents XXI. Constituents of Merrilliodendron megacarpum. Planta Med 43:404–407
Asano T, Watase I, Sudo H, Kitajima M, Takayama H, Aimi N, Yamazaki M, Saito K (2004) Camptothecin production by in vitro cultures of Ophiorrhiza Liukiuensis and O. Kuroiwai. Plant Biotechnol 21:275–281
Bhalkar BN, Bedekar PA, Patil SM, Patil SA, Govindwar SP (2015) Production of camptothecine using whey by an endophytic fungus: standardization using response surface methodology. RSC Adv 5:62828–62835
Chandran H, Meena M, Barupal T, Sharma K (2020) Plant tissue culture as a perpetual source for production of industrially important bioactive compounds. Biotechnol Rep (Amst) 26:e00450
Ciddi V, Shuler ML (2000) Camptothecine from callus cultures of Nothapodytes Foetida. Biotechnol Lett 22:129–132
Cui L, Ni X, Ji Q, Teng X, Yang Y, Wu C, Zekria D, Zhang D, Kai G (2015) Co-overexpression of geraniol-10-hydroxylase and strictosidine synthase improves anti-cancer drug camptothecin accumulation in Ophiorrhiza Pumila. Sci Rep 5:8227
Dai J-R, Hallock YF, Cardellina JH, Boyd MR (1999) 20-O-β-Glucopyranosyl camptothecin from Mostuea brunonis: a potential camptothecin pro-drug with improved solubility. J Nat Prod 62:1427–1429
Dandin VS, Murthy HN (2012) Enhanced in vitro multiplication of Nothapodytes Nimmoniana Graham using semisolid and liquid cultures and estimation of camptothecin in the regenerated plants. Acta Physiol Plant 34:1381–1386
Deepthi S, Satheeshkumar K (2016) Enhanced camptothecin production induced by elicitors in the cell suspension cultures of Ophiorrhiza mungos Linn. Plant Cell Tiss Org Cult 124:483–493
Fan X, Lin X, Ruan Q, Wang J, Yang Y, Sheng M, Zhou W, Kai G, Hao X (2022) Research progress on the biosynthesis and metabolic engineering of the anti-cancer drug camptothecin in Camptotheca acuminate. Ind Crops Prod 186:115270
Fukuoka M (2001) Current status of irinotecan in lung cancer. Oncology 15:6–7
Fulzele DP, Satdive RK (2005) Comparison of techniques for the extraction of the anti-cancer drug camptothecin from Nothapodytes Foetida. J Chromatogr A 1063:9–13
Fulzele DP, Satdive RK, Pol BB (2001) Growth and production of camptothecin by cell suspension cultures of Nothapodytes Foetida. Planta Med 67:150–152
Fulzele DP, Satdive R, Kamble S, Singh S, Singh S (2015) Improvement of anticancer drug camptothecin production by gamma irradiation on callus cultures of Nothapodytes Foetida. Int J Pharm Res Alli Sci 4:19–27
Gharpure G, Chavan B, Lele U, Hastak A, Bhave A, Malpure N, Vasudeva R, Patwardhan A (2010) Camptothecin accumulation in Ophiorrhiza rugosa var. Prostrata from northern Western Ghats. Curr Sci 98:302–304
Gonçalves S, Romano A (2018) Production of plant secondary metabolites by using biotechnological tools. In: Vijayakumar R, Raja SSS (eds) Secondary Metabolites. IntechOpen, pp 81–99
Greenwell M, Rahman PKSM (2015) Medicinal plants: their use in anticancer treatment. Int J Life Sci Pharma Res 6:4103–4112
Gunasekera SP, Badawi MM, Cordell GA, Farnsworth NR, Chitnis M (1979) Plant anticancer agents X. isolation of camptothecin and 9-methoxycamptothecin from Ervatamia heyneaya. J Nat Prod 42:475–477
Guo-yin KAI (2009) Distribution of anticancer drug in Ophiorrhiza Japonica. J Shanghai Univ Nat Univ 38:59–62
Hao X, Xie C, Ruan Q, Zhang X, Wu C, Han B, Qian J, Zhou W, Nützmann H-W, Kai G (2021) The transcription factor OpWRKY2 positively regulates the biosynthesis of the anticancer drug camptothecin in Ophiorrhiza Pumila. Hortic Res 8:7
Hao X, Wang C, Zhou W, Ruan Q, Xie C, Yang Y, Xiao C, Cai Y, Wang J, Wang Y, Zhang X, Maoz I, Kai G (2023) OpNAC1 transcription factor regulates the biosynthesis of the anticancer drug camptothecin by targeting loganic acid O-methyltransferase in Ophiorrhiza Pumila. J Integr Plant Biol 65:133–149
Hashim S, Beh HK, Hamil MSR, Ismail Z, Majid AMSA (2016) High-performance thin-layer chromatography method development, validation, and simultaneous quantification of four compounds identified in standardized extracts of Orthosiphon Stamineus. Pharmacog Res 8:238–243
Hu Y, Yu W, Song L, Du XH, Ma X, Liu Y, Wu J, Ying Y (2016) Effects of light on production of camptothecin and expression of key enzyme genes in seedlings of Camptotheca acuminate Decne. Acta Physiol Plant 38:65
Hu Y-T, Xu Z-C, Tian Y, Gao R-R, Ji A-J, Pu X-D, Wang Y, Liu X, Song J-Y (2020) Genome-wide identification and analysis of AP2/ERF transcription factors related to camptothecin biosynthesis in Camptotheca acuminata. Chin J Nat Med 18:582–593
Hussain MS, Fareed S, Ansari S, Rahman MA, Ahmad IZ, Saeed M (2012) Current approaches toward production of secondary plant metabolites. J Pharm Bioallied Sci 4:10–20
Isah T (2017) Production of camptothecin in the elicited callus cultures of Nothapodytes Nimmoniana (J. Graham) Mabberly. Chem Pap 71:1091–1106
Isah T, Mujib A (2015a) Camptothecin from Nothapodytes Nimmoniana: review on biotechnology applications. Acta Physiol Plant 37:106
Isah T, Mujib A (2015b) In vitro propagation and camptothecin production in Nothapodytes Nimmoniana. Plant Cell Tiss Org Cult 121:1–10
Isah T, Umar S (2019) Influence of plant growth regulators on morphogenic response, biomass and camptothecin production in the callus cultures of Chonemorpha fragrans. Moon) Alston. Research Square
Isah T, Masood S, Umar S (2022) Biomass and camptothecin production in the calcium chloride elicited and liquid medium overlayed Nothapodytes Nimmoniana (J. Graham) Mabberly callus cultures. Vegetos 35:104–114
Jain AK, Nessler CL (1996) Clonal propagation of Camptotheca acuminata through shoot bud culture. Plant Cell Tiss Org Cult 44:229–233
Jisha KG (2006) A study on the production of camptothecin from Ophiorrhiza mungos and Nolhapodytes foelida using cell and tissue culture. http://hdl.handle.net/10603/466
Kadam SB, Godbole RC, Pable AA, Singh S, Barvkar VT (2023) Thidiazuron-mediated callogenesis and biosynthesis of anti-cancerous monoterpene indole alkaloid camptothecin in Nothapodytes Nimmoniana (J.Graham) Mabb. Callus culture. S Afr J Bot 156:411–419
Kai G-Y, Dai L-M, Mei X-Y, Zheng J-G, Wang W, Lu Y, Qian Z-Y, Zhou G-Y (2008) In vitro plant regeneration from leaf explants of Ophiorrhiza Japonica. Biol Plant 52:557–560
Kai G, Wu C, Gen L, Zhang L, Cui L, Ni X (2015) Biosynthesis and biotechnological production of anti-cancer drug Camptothecin. Phytochem Rev 14:525–539
Kamble S, Gopalakrishnan R, Eapen S (2011) Production of camptothecin by hairy roots and regenerated transformed shoots of Ophiorrhiza rugosa var. Decumbens. Nat Prod Res 25:1762–1765
Kang M, Fu R, Zhang P, Lou S, Yang X, Chen Y, Ma T, Zhang Y, Xi Z, Liu J (2021) A chromosome-level Camptotheca acuminata genome assembly provides insights into the evolutionary origin of camptothecin biosynthesis. Nat Commun 12:3531
Karwasara VS, Dixit VK (2013) Culture medium optimization for camptothecin production in cell suspension cultures of Nothapodytes Nimmoniana (J. Grah.) Mabberley. Plant Biotechnol Rep 7:357–369
Kim SH, Hur BK, Byun SY (1999) Effect of sugar concentration on camptothecin production in cell suspension cultures of Camptotheca acuminata. Biotechnol Bioprocess Eng 4:277–280
Krishna Kumar G, Muhammed Fayad A, Jayakumaran Nair A (2018) Ophiorrhiza mungos var. Angustifolia – estimation of camptothecin and pharmacological screening. Plant Sci Today 5:113–120
Krishnakumar G, Dintu KP, Varghese SC, Nair DS, Gopinath G, Rameshkumar KB, Satheeshkumar K, Krishnan PN (2020) Ophiorrhiza, a promising herbaceous source of the anticancer compound camptothecin. Plant Sci Today 7:240–250
Krishnan JJ, Gangaprasad A, Satheeshkumar K (2018) In vitro mass multiplication and estimation of camptothecin (CPT) in Ophiorrhiza mungos L. var. Angustifolia (Thw.) Hook. F. Ind Crops Prod 119:64–72
Kulkarni AV (2008) In vitro studies and secondary metabolite production in Tabernaemontana alternifolia L and Chonemorpha fragran (Moon) alston [dissertation] Pune (India): University of Pune; 2008
Kulkarni AV, Patwardhan AA, Lele U, Malpathak NP (2010) Production of camptothecin in cultures of Chonemorpha Grandiflora. Pharmacog Res 2:296–299
Kusari S, Zühlke S, Spiteller M (2009) An endophytic fungus from Camptotheca acuminata that produces camptothecin and analogues. J Nat Prod 72:2–7
Lakshmanan P, Lee CL, Goh CJ (1997) An efficient in vitro method for mass propagation of a woody ornamental Ixora coccinea L. Plant Cell Rep 16:572–577
Lee J-Y, Goto E, Yoshida H, Hikosaka S (2022) Optimal harvest-time to maximize the annual camptothecin production by Ophiorrhiza pumila in a plant factory with artificial light. J Nat Med 76:865–872
Lekshmi GM (2011) In vitro culture of Ophiorrhiza pectinata Am for propagation exploitation of active compounds and pharmacological evaluation. Dissertation, University of Kerala
Li S, Yi Y, Wang Y, Zhang Z, Beasley RS (2002) Camptothecin accumulation and variations in camptotheca. Planta Med 68:1010–1016
Li F, Jiang T, Li Q, Ling X (2017) Camptothecin (CPT) and its derivatives are known to target topoisomerase I (Top1) as their mechanism of action: did we miss something in CPT analogue molecular targets for treating human disease such as cancer. Am J Cancer Res 7:2350–2394
Liew ST, Yang L-X (2008) Design, synthesis and development of novel camptothecin drugs. Curr Pharm Des 14:1078–1097
Liu Z, Adams J (1996) Camptothecin yield and distribution within Camptotheca acuminata trees cultivated in Louisiana. Can J Bot 74:360–365
Lokesh P, Balasubramanya S, Anuradha M (2014) Cost effective quantification of camptothecin and a comparative study of its content in Nothapodytes foetida and ophiorrhiza mungos sourced from selected geographical locations. Orient Pharm Exp Med 14:87–92
Malik SS, Laura JS (2014) Distribution of camptothecin through the plant kingdom. Int J Curr Res Rev 6:6497–6507
Mamkulathil Devasia R, Altaf M, Fahad Alrefaei A, Manoharadas S (2021) Enhanced production of camptothecin by immobilized callus of Ophiorrhiza mungos and a bioinformatic insight into its potential antiviral effect against SARS-CoV-2. J King Saud Univ Sci 33:101344
Martin KP, Zhang C-L, Hembrom ME, Slater A, Madassery J (2008) Adventitious root induction in Ophiorrhiza prostrata: a tool for the production of camptothecin (an anticancer drug) and rapid propagation. Plant Biotechnol Rep 2:163–169
Martino E, Della Volpe S, Terribile E, Benetti E, Sakaj M, Centamore A, Sala A, Collina S (2017) The long story of camptothecin: from traditional medicine to drugs. Bioorg Med Chem Lett 27:701–707
Mithun PR, Xavier J, Jayarama R, Praveen N (2017) Production of camptothecin from Nothapodytes Nimmoniana: an overview. Int J Life Sci Res 3:1476–1483
Mohinudeen IAHK, Kanumuri R, Soujanya KN, Shaanker RU, Rayala SK, Srivastava S (2021) Sustainable production of camptothecin from an Alternaria sp. isolated from Nothapodytes Nimmoniana. Sci Rep 11:1478
Murthy HN, Dalawai D, Park S-Y, Paek K-Y (2019) Endophytes of Nothapodytes Nimmoniana (J. Graham) Mabb. In: Jha S (ed) Endophytes and secondary metabolites. Springer International Publishing, Cham, pp 145–164
Musavi SF, Dhavale A, Balakrishnan RM (2015) Optimization and kinetic modeling of cell-associated camptothecin production from an endophytic Fusarium oxysporum NFX06. Prep Biochem Biotech 45:158–172
Nagesha BV, Amilineni U, Gudasalamani R, Nataraja Karaba N, Ramanan US (2018) Elicitors act as a signal transducer in the enhancement of camptothecine production from in vitro cultures of Ophiorrhiza mungos L. Ann Phytomed 7:46–54
Namdeo AG, Sharma A (2012) HPLC analysis of camptothecin content in various parts of Nothapodytes Foetida collected on different periods. Asian Pac J Trop Biomed 2:389–393
Namdeo AG, Priya T, Bhosale BB (2012) Micropropagation and production of camptothecin form in vitro plants of Ophiorrhiza mungos. Asian Pac J Trop Biomed 2:S662–S666
Natarajan S, Pucker B, Srivastava S (2023) Genomic and transcriptomic analysis of camptothecin producing novel fungal endophyte - Alternaria Burnsii NCIM 1409. Sci Rep 13:14614
Ni X, Wen S, Wang W, Wang X, Xu H, Kai G (2011) Enhancement of camptothecin production in Camptotheca acuminata hairy roots by overexpressing ORCA3 gene. J Basic Appl Pharm Sci 1:85
Niazian M (2019) Application of genetics and biotechnology for improving medicinal plants. Planta 249:953–973
Nishitha IK, Martix KP, Ligimol, Shahanaz Beegum A, Madhusoodanan PV (2006) Micropropagation and encapsulation of medicinally important Chonemorpha grandiflora. Vitro Cell Dev Bio - Plant 42:385–388
Nurgali K, Thomas Jagoe R, Abalo R (2018) Adverse effects of cancer chemotherapy: anything new to improve tolerance and reduce sequelae? Front Pharm 9:245
Pan X-W, Xu H-H, Liu X, Gao X, Lu Y-T (2004) Improvement of growth and camptothecin yield by altering nitrogen source supply in cell suspension cultures of Camptotheca acuminata. Biotechnol Lett 26:1745–1748
Park YG, Kim MH, Yang JK, Chung YG, Choi MS (2003) Light-susceptibility of camptothecin production from in vitro cultures of Camptotheca acuminata Decne. Biotechnol Bioprocess Eng 8:32–36
Patil S, Chougale KD, Hajare RA (2022) Development and validation of RP-HPLC method for estimation of camptothecin in mixed micelle formulation. Res J Pharm Technol 15:4248–4252
Pu X, Qu X, Chen F, Bao J, Zhang G, Luo Y (2013) Camptothecin-producing endophytic fungus Trichoderma atroviride LY357: isolation, identification, and fermentation conditions optimization for camptothecin production. Appl Microbiol Biotechnol 97:9365–9375
Pu X, Zhang C-R, Zhu L, Li Q-L, Huang Q-M, Zhang L, Luo Y-G (2019) Possible clues for camptothecin biosynthesis from the metabolites in camptothecin-producing plants. Fitoterapia 134:113–128
Pu X, Gao H-C, Wang M-J, Zhang J-H, Shan J-H, Chen M-H, Zhang L, Wang H-G, Wen A-X, Luo Y-G, Huang Q-M (2022) Integrative analysis of elicitor-induced camptothecin biosynthesis in Camptotheca acuminata plantlets through a combined omics approach. Front Plant Sci 13:851077
Puri SC, Handa G, Dhar KL, Suri OP, Qazi GN (2003) Process for preparing topotecan from 10-hydroxy-4-(S) camptothecin. US Patent
Rajan R, Varghese SC, Kurup R, Gopalakrishnan R, Venkataraman R, Satheeshkumar K, Baby S (2013) Search for camptothecin-yielding Ophiorrhiza species from southern western ghats in India: a HPTLC-densitometry study. Ind Crops Prod 43:472–476
Ramesha BT, Suma HK, Senthilkumar U, Priti V, Ravikanth G, Vasudeva R, Kumar TRS, Ganeshaiah KN, Shaanker RU (2013) New plant sources of the anti-cancer alkaloid, camptothecine from the Icacinaceae taxa, India. Phytomedicine 20:521–527
Rani VKJ (2011) Study on the production of camptothecin and its derivatives from selected Ophiorrhiza species and its in vitro culture. [Ph.D]. Padikkala J, editor. Amala Cancer Research Centre; 2011. https://shodhganga.inflibnet.ac.in/handle/10603/25802
Rao SR, Ravishankar GA (2002) Plant cell cultures: chemical factories of secondary metabolites. Biotechnol Adv 20:101–153
Rehman S, Shawl AS, Verma V, Kour A, Athar M, Andrabi R, Sultan P, Qazi GN (2008) An endophytic Neurospora sp. from Nothapodytes foetida producing camptothecin. Prikl Biokhim Mikrobiol 44:225–231
Rehman S, Shawl AS, Kour A, Sultan P, Ahmad K, Khajuria R, Qazi GN (2009) Comparative studies and identification of camptothecin produced by an endophyte at shake flask and bioreactor. Nat Prod Res 23:1050–1057
Roja G (2006) Comparative studies on the camptothecin content from Nothapodytes Foetida and Ophiorrhiza species. Nat Prod Res 20:85–88
Roja G (2008) Micropropagation and production of camptothecin from in vitro plants of Ophiorrhiza rugosa var. decumbens. Nat Prod Res 22:1017–1023
Sadre R, Magallanes-Lundback M, Pradhan S, Salim V, Mesberg A, Jones AD, DellaPenna D (2016) Metabolite diversity in alkaloid biosynthesis: a multilane (diastereomer) highway for camptothecin synthesis in Camptotheca acuminata. Plant Cell 28:1926–1944
Saito K, Sudo H, Yamazaki M, Koseki-Nakamura M, Kitajima M, Takayama H, Aimi N (2001) Feasible production of camptothecin by hairy root culture of Ophiorrhiza Pumila. Plant Cell Rep 20:267–271
Sakato K, Misawa M (1974) Effects of chemical and physical conditions on growth of Camptotheca acuminata cell cultures. Agri Biol Chem 38:491–497
Sankar-Thomas YD, Saare-Surminski K, Lieberei R (2008) Plant regeneration via somatic embryogenesis of Camptotheca acuminata in temporary immersion system (TIS). Plant Cell Tiss Org Cult 95:163–173
Saravanan P, Boopalan E (2011) Occurrence of camptothecin an anticancer drug from Ixora coccinea Linn. Int J Appl Biology 2:30–34
Sasidharan H, Chembrakuniyil M, Krishnan S (2023) Enhancement of camptothecin biosynthesis in the green root cultures of Pyrenacantha Volubilis Hook. Under varied concentrations of ammonium nitrate and sucrose along with exogenous elicitors. Ind Crops Prod 192:116000
Schreiner M (2005) Vegetable crop management strategies to increase the quantity of phytochemicals. Eur J Nutr 44:85–94
Shahanaz Beegum A, Poulose Martin K (2007) Organogenesis from leaf and internode explants of Ophiorrhiza prostrata, an anticancer drug (camptothecin) producing plant. Electron J Biotechol 10:114–123
Shi M, Gong H, Cui L, Wang Q, Wang C, Wang Y, Kai G (2020) Targeted metabolic engineering of committed steps improves anti-cancer drug camptothecin production in Ophiorrhiza Pumila hairy roots. Ind Crops Prod 148:112277
Shrivastava V, Sharma N, Shrivastava V, Sharma A (2021) Review on camptothecin producing medicinal plant: Nothapodytes Nimmoniana. Biomed Pharmacol J 14:1799–1813
Shweta S, Zuehlke S, Ramesha BT, Priti V, Mohana Kumar P, Ravikanth G, Spiteller M, Vasudeva R, Uma Shaanker R (2010) Endophytic fungal strains of Fusarium solani, from Apodytes Dimidiata E. Mey. Ex Arn (Icacinaceae) produce camptothecin, 10-hydroxycamptothecin and 9-methoxycamptothecin. Phytochem 71:117–122
Shweta S, Gurumurthy BR, Ravikanth G, Ramanan US, Shivanna MB (2013) Endophytic fungi from Miquelia Dentata Bedd., produce the anti-cancer alkaloid, camptothecine. Phytomed 20:337–342
Silvestrini A, Pasqua G, Botta B, Monacelli B, van der Heijden R, Verpoorte R (2002) Effects of alkaloid precursor feeding on a Camptotheca acuminata cell line. Plant Physiol Biochem 40:749–753
Singh S, Kamble SN, Satdive RK, Fulzele DP (2020) Heterologous overexpression of Nothapodytes Foetida strictosidine synthase enhances levels of anti-cancer compound camptothecin in Ophiorrhiza rugosa. Plant Cell Tiss Org Cult 141:67–76
Sirikantaramas S, Yamazaki M, Saito K (2013) Camptothecin: biosynthesis, biotechnological production and resistance mechanism(s). In: Giglioli-Guivarc’h N (ed) Advances in Botanical Research, Elsevier Ltd, Amsterdam, pp 139–161
Smetanska I (2008) Production of secondary metabolites using plant cell cultures. Adv Biochem Eng Biotechnol 111:187–228
Song SH, Byun SY (1998) Elicitation of camptothecin production in cell cultures of Camptotheca acuminata. Biotechnol Bioprocess Eng 3:91
Sudo H, Yamakawa T, Yamazaki M, Aimi N, Saito K (2002) Bioreactor production of camptothecin by hairy root cultures of Ophiorrhiza Pumila. Biotechnol Lett 24:359–363
Thengane SR, Kulkarni DK, Shrikhande VA, Joshi SP, Sonawane KB, Krishnamurthy KV (2003) Influence of medium composition on callus induction and camptothecin(s) accumulation in Nothapodytes Foetida. Plant Cell Tiss Org Cult 72:247–251
Ulukan H, Swaan PW (2002) Camptothecins: a review of their chemotherapeutic potential. Drugs 62:2039–2057
Uzma F, Mohan CD, Hashem A, Konappa NM, Rangappa S, Kamath PV, Singh BP, Mudili V, Gupta VK, Siddaiah CN, Chowdappa S, Alqarawi AA, Abd Allah EF (2018) Endophytic fungi-alternative sources of cytotoxic compounds: a review. Front Pharmacol 9:309
van der Fits L, Memelink J (2000) ORCA3, a jasmonate-responsive transcriptional regulator of plant primary and secondary metabolism. Science 289:295–297
van Hengel AJ, Harkes MP, Wichers HJ, Hesselink PGM, Buitelaar RM (1992) Characterization of callus formation and camptothecin production by cell lines of Camptotheca acuminata. Plant Cell Tiss Org Cult 28:11–18
van Hengel AJ, Buitelaar RM, Wichers HJ (1994) Camptotheca acuminata Decne: in vitro culture and the production of camptothecin. In: Bajaj YPS (ed) Medicinal and aromatic plants VII. Springer, Berlin Heidelberg, Berlin, Heidelberg, pp 98–112
Varghese C (ed) (2017) In vitro studies on Ophiorrhiza trichocarpos blume for camptothecin production. [Ph.D.]. Satheeshkumar K, editor. Manonmaniam Sundaranar University; 2017. https://shodhganga.inflibnet.ac.in/handle/10603/203106
Vineesh VR, Jelly CL, Fijesh PV, Jaimsha VK, Padikkala J (2007) Effect of benzyl adenine and naphathalene acetic acid on the production of camptothecin in multiple shoot and root cultures of Ophiorrhiza rugosa var. decumbens. Nat Prod Rad 6:405–409
Viraporn V, Yamazaki M, Saito K, Denduangboripant J, Chayamarit K, Chuanasa T, Sukrong S (2011) Correlation of camptothecin-producing ability and phylogenetic relationship in the genus Ophiorrhiza. Planta Med 77:759–764
Wang C, Hao X, Wang Y, Maoz I, Zhou W, Zhou Z, Kai G (2022) Identification of WRKY transcription factors involved in regulating the biosynthesis of the anti-cancer drug camptothecin in Ophiorrhiza Pumila. Hortic Res 9:uhac099
Wetterauer B, Hummel E, Walczak S, Distl M, Langhans M, Wetterauer P, Sporer F, Wildi E, Wink M (2021) Physiology of camptothecin synthesis in plants and root organ cultures of Ophiorrhiza mungos L. and its production in root fermenters. In: Malik S (ed) Exploring Plant cells for the production of compounds of interest. Springer International Publishing, Cham, pp 291–338
Ya-ut P, Chareonsap P, Sukrong S (2011) Micropropagation and hairy root culture of Ophiorrhiza Alata Craib for camptothecin production. Biotechnol Lett 33:2519–2526
Yamazaki Y, Urano A, Sudo H, Kitajima M, Takayama H, Yamazaki M, Aimi N, Saito K (2003) Metabolite profiling of alkaloids and strictosidine synthase activity in camptothecin producing plants. Phytochemistry 62:461–470
Yang M, Wang Q, Liu Y, Hao X, Wang C, Liang Y, Chen J, Xiao Y, Kai G (2021) Divergent camptothecin biosynthetic pathway in Ophiorrhiza Pumila. BMC Biol 19:122
Yuan Y, Liu Z, Yang Y, Wu H (2008) Studies on the production of comptothecin in in vitro cultured roots of Camptotheca acuminata. 2nd International Conference on Bioinformatics and Biomedical Engineering, Shanghai. IEEE, Newyork, pp.1169–1172
Zabudkin A (2015) Method for the synthesis of irinotecan. US Patent 20160264590
Zeng X-H, Li Y-H, Wu S-S, Hao R-L, Li H, Ni H, Han H-B, Li H-H (2013) New and highly efficient column chromatographic extraction and simple purification of camptothecin from Camptotheca acuminata and Nothapodytes pittosporoides. Phytochem Anal 24:623–630
Zhang H, Shen X, Sun S, Li Y, Wang S, Wei J, Guo B, Sun C (2023) Integrated transcriptome and proteome analysis provides new insights into camptothecin biosynthesis and regulation in Camptotheca acuminata. Physiol Plant 175:e13916
Zhou BN, Hoch JM, Johnson RK, Mattern MR, Eng WK, Ma J, Hecht SM, Newman DJ, Kingston DG (2000) Use of COMPARE analysis to discover new natural product drugs: isolation of camptothecin and 9-methoxycamptothecin from a new source. J Nat Prod 63:1273–1276
Acknowledgements
The authors express their appreciation for the support provided by the Deanship of Scientific Research, Vice Presidency for Graduate Studies and Scientific Research, King Faisal University, Saudi Arabia (Project No. GRANTA341).
Funding
The author Praveen Nagella is thankful to Center for Research, CHRIST (Deemed to be University), Bangalore, for financial support in the form of Major Research Project (MRP-DSC1414).
Author information
Authors and Affiliations
Contributions
Conceptualization—P.N., A.R.V., K.M., R.S.V., J.M.A.-K.; methodology—A.B., W.N.S.; resources—A.B., W.N.S., V.S.D., P.M.N.; writing—original draft preparation—A.B., W.N.S., V.S.D., P.M.N.; writing—review and editing—P.N., A.R.V., K.M., M.T.; supervision—P.N., A.R.V., K.M., R.S.V., J.M.A.-K.; funding acquisition—P.N. All authors have read and agreed to the published version of the manuscript.
Corresponding authors
Ethics declarations
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Banadka, A., Narasimha, S.W., Dandin, V.S. et al. Biotechnological approaches for the production of camptothecin. Appl Microbiol Biotechnol 108, 382 (2024). https://doi.org/10.1007/s00253-024-13187-2
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00253-024-13187-2