
Abstract
Agriculture is one of the main factors associated with global biodiversity loss. The present study assesses the impact of horticultural land use on water quality and fish assemblage composition in lowland Pampean streams. Four samplings were conducted with seasonal frequency during 2018 and 2019 in three horticultural stream reaches (HSR) and three grassland reaches (GSR) with low anthropic intervention, two also located in a biosphere reserve area. Environmental parameters were determined, and water and sediment samples were taken for nutrient and pesticide analysis, respectively. Fish assemblages were sampled in two habitat types: open water (OW) and vegetated patches (VP). The concentration of soluble reactive phosphorus and the detection frequency and concentration of pesticides were significatively higher in HSR mainly attributed to the runoff events following application of fertilizers and pesticides to the surrounding crops. Specific richness was significantly higher in GSR for both OW and VP environments. Diversity and evenness in the VP were significantly higher in GSR, while abundance was significantly higher in HSR due to the dominance of the environmental pollution-tolerant taxa of the order Cyprinodontiformes. The multivariate analysis recorded a major contribution of the order Cyprinodontiformes in the HSR, while the GSR species of the order Characiformes, common in environments of low environmental impact, were better represented. Our study demonstrated water quality deterioration associated with horticultural land use resulting in a ruderal fish community. On-going horticulture expansion is expected to cause further deterioration within the region, unless managerial practices are implemented.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Ecosystem disturbances caused by human activity is one of the main drivers of biodiversity decline worldwide (Sala et al. 2000; Albert et al. 2021). Land use plays an important role in the ecological integrity of riverine ecosystems (Brauns et al. 2022). Agriculture has been considered the major cause of water pollution (Davies-Colley et al. 2004) and the main factor linked to loss of global biodiversity worldwide (Dolédec and Statzner 2010; Dudley and Alexander 2017). It affects freshwater environments mainly through non-point sources (Monaghan et al. 2007), changing hydrological characteristics (Allan 2004) and increasing sediments, nutrients and pesticide loads (Nessimian et al. 2008; Thomas et al. 2018; Cornejo et al. 2019; Harrison et al. 2019).
In Argentina, agriculture is the main productive activity; horticulture is developed in the peri-urban areas of large cities. The horticultural area around La Plata city, locally known as “Cinturón Hortícola Platense” (CHP), covers roughly 8600 ha, and crop production is developed 50% in open field and 50% under cover (Baldini et al. 2022). Greenhouse production is intensively managed; it allows several crops per year and requires large amounts of fertilizer and pesticide application. About 170 pesticide active ingredients are being used in the area (DP 2015).
Fish assemblages have been considered sensitive to changes in the environment (Walser and Bart 1999; Teresa and Casatti 2017; Bertora et al. 2024). The decline in their specific richness and diversity due to human impact is well documented (Magurran and Phillip 2001; Helms et al. 2005). The Neotropical ichthyofauna is regarded as the most diverse continental fauna in the world (Lévêque et al. 2007; Albert et al. 2020), and recent reports have suggested that it is severely threatened by human activities (Castro and Polaz 2019; Pelicice et al. 2021).
Several studies have assessed the negative effects of urbanization (Cunico et al. 2011; Paracampo et al. 2020) and industrial (Araújo et al. 2009; Paredes del Puerto et al. 2021) and cattle raising (Matono et al. 2013; Goss and Roper 2018) on fish assemblages. In the USA the biotic integrity of fish assemblages in streams from watersheds with approximately 10% of agriculture in the buffer zone was poor (Fitzpatrick et al. 2001). Effert-Fanta et al. (2019) registered negative effects on biotic integrity in streams from watersheds with a high percentage of agriculture and low riparian forest fringe. By contrast, Wang et al. (1997) reported good habitat quality and biotic integrity of streams in basins where agriculture was over 80%, attributed to environmental features that mitigate the impact of agriculture, such as the presence of high gradients and rocky substrates, and the absence of channelization. Zeni et al. (2017) determined weak correlations between fish community attributes and the environmental change caused by the land use transformation of pasture to sugar cane in Brazilian streams. The authors suggest further monitoring because they argue that the failure to find a correlation between land use change and the fish assemblage attributes is related to a time lag of the impact. The aim of the present study was, therefore, to evaluate the impact of horticultural land use on the water quality, taxonomic structure, and composition of fish assemblages in Pampean lowland streams.
Material and Methods
Study area
The studied streams run roughly parallel through the Pampas plain to end in the Río de la Plata Estuary. The climate is mild and humid, with average monthly temperatures ranging from 10 °C in winter to 22 °C in summer. Mean annual rainfall is 1060 mm with small seasonal variations (Hurtado et al. 2006). The landscape is grassland with small patches of forest. The Pampa plain has a remarkably low slope (< 1%) (Hurtado et al. 2006) and, therefore, the streams are shallow, with silty bottom sediments and abundant macrophytes and riparian vegetation (Giorgi et al. 2005). Water flow does not present variations associated with the seasonality. Changes in the flow occur after heavy rains where flood peaks occur or during periods of exceptional drought where the water level may decrease significantly (Paredes del Puerto et al. 2024; Rodrigues Capítulo et al. 2010).
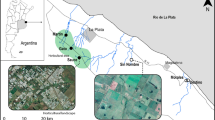
Six wadeable stream reaches were sampled seasonally in winter and spring 2018 and summer and autumn 2019. Three are located along the Sauce stream, running through the CHP, hereafter referred to as horticultural stream reaches (HSR): H1, H2 and H3 (Fig. 1). The other three are located in streams running through less impacted grassland areas, hereafter referred to as grassland stream reaches (GSR): G1, G2 and G3, respectively (Fig. 1). The G1 site is located in the Chubichaminí stream and run through livestock fields with low cattle density. The G2 and G3 sampling sites are in the Morales and Destino streams, running through the Parque Costero Sur, UNESCO Biosphere Reserve (Athor 2009).

Study sites in Pampean streams, Argentina. H1–H3: horticultural sites in Sauce stream. G1–G3: grassland sites. G1: Chubichaminí stream, G2: Morales stream and G3: Destino stream
Environmental variables
At each sampling site, environmental parameters were determined in situ: dissolved oxygen and water temperature with (YSI 51B), pH (Hanna checker), conductivity (Hanna instruments 8733), transparency (Secchi disc) and stream depth with a gage rod. Water samples were taken for nutrient analysis, transported to the laboratory on ice and filtered through 1.2-µm pore size Whatman GF/C filters. The nutrients were then measured in the filtrate: the soluble reactive phosphorus (SRP) was determined by a colorimetric reaction with ascorbic-acid–ammonium-molybdate; the nitrate was calculated by hydrazine reduction followed by diazotation, and the ammonium was quantified by the indophenol-blue method (APHA 2012). Suspended solids were estimated as the weight difference of the filter before and after filtration (APHA 2012).
At each sampling site, macrophyte community composition was determined following Cabrera and Zardini (1993) and classified according to their life form according to Messetta et al. (2023). In the surveys carried out in spring 2018 and summer and autumn 2019, sediment samples for pesticide determinations were taken in each sampling site from the top two centimeters with a stainless-steel scoop, placed in glass vessels and transported to the laboratory on ice. The samples were extracted following You et al. (2004) with a mixture of acetone and methylene chloride. A clean-up procedure was carried out using Florisil solid-phase-extraction cartridges. Chlorpyrifos, endosulfan, permethrin, λ-cyhalothrin, cypermethrin and deltamethrin were quantified by gas chromatography (HP 6890) using 30 m-length HP1 columns and equipped with electron capture detector. Two microliters were injected on a splitless mode (290 °C) and detector was kept at 290 °C. The oven temperature program was: start at 190 °C, followed by an increase of 10 °C/min up to 260 °C (13 min), then 10 °C/min up to 290 °C (7 min). Quantification was performed using pure standard pesticides (AccuStandard; 4-point calibration curve) and tetrachloro m-xylene (Ultra Scientific) as internal standard. The detection limits in dry sediment ranged between 0.35 ng/g for chlorpyrifos and endosulfan, and 1.0–3.0 ng/g for pyrethroids. Procedural and instrumental blanks (one for every batch of twelve samples) were below the detection limits. Pesticide concentrations below the detection levels were treated as half the detection limits for statistical analysis.
Fish assemblages
Fish were collected in two types of habitats using different fishing gears. In open water (OW) environments, fish were sampled with a sei8ne net with a bag (width 15 m, height 1.1 m and mesh size 10 mm between opposite knots; bag width 2 m and mesh size 5 mm between opposite knots) along a 30 m stretch of stream previously isolated with a block net (width 20 m, height 1.1 m and mesh size 10 × 5 mm). Two people hauled the seine net upstream toward the block net, where both nets were recovered together.
Fish were also collected in vegetated patches (VP). Pampean streams have dense and rich macrophyte communities that increase habitat heterogeneity in relation to a comparatively homogeneous sediment, supporting rich community of consumers that live in association with the vegetation (Giorgi et al. 2005; Feijoó and Lombardo 2007; Cortelezzi et al. 2013). Therefore, two fishing gears were used in the VP with aquatic macrophytes in a 30 m stretch upstream of the seine net-sampled sector: a hand frame (0.6 × 0.6 m, mesh size 2 × 2 mm) and a square frame drop net (frame width 0.48 m and height, 0.48 m; mesh size 1 × 1 mm; rope 5 m) were used. Five fishing attempts were made with each type of gear. Sampling was carried out during base flow conditions to avoid undesired variations associated with the flood flow effect (Junk and Wantzen 2004). The treatment of the specimens was carried out in accordance with the requirement of the National Council of Scientific and Technical Research of Argentina follow the guidelines of its "Comité de Etica" (OCR-RD-20050701–1047.pdf conicet.gov.ar) and its biological sampling guide (https://proyectosinv.conicet.gov.ar/solicitud-colecta-cientifica/). The scientific collection permits were granted by the Subsecretaria de Agricultura, Ganadería y Pesca of the Province of Buenos Aires (exp. DI-2019–488-GDEBA-DAPAYCPMAGP).
Easily identified species were weighed with an Ohaus 0.5-g–precision balance, measured and returned to the stream. All other specimens captured were euthanized with an overdose of anesthetic (benzocaine in excess), conserved with a solution of 10% formalin and later replaced by 70% ethanol for routine laboratory determination. Species were identified following Azpelicueta and Braga (1991), Braga (1993, 1994), Aquino (1997), Reis and Pereira (2000), Casciotta et al. (2005), Miquelarena and Menni (2005), Říčan and Kullander (2008), Almirón et al. (2015) and Rosso et al. (2018). Updates in the taxonomy were reviewed following Mirande and Koerber (2020), Terán et al. (2020) and Fricke et al. (2023). Voucher specimens were stored at the ichthyological collection of the Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" (MACN-Ict), Buenos Aires, Argentina.
Data analysis
Species richness (S), abundance, biomass, Shannon–Wiener diversity index (H’) and evenness (J’) were calculated for each sampling site, separately for OW and VP environments, hereafter referred to as the biotic attributes. The values were calculated with the “Diverse” routine for univariate index treatment performed with Primer-E v6.0 (Clarke and Warwick 2001).
Differences in physicochemical variables and biotic attributes among the different land uses were assessed by t-test or its non-parametric equivalent Mann–Whitney test using Sigmaplot for windows v12. Significance levels were set to α = 0.05. To evaluate differences in the insecticide concentrations, Dixon's test for outliers and one-way ANOVA followed by Tukey HSD comparison were accomplished with XLSTAT software (version 2014.5.03). Significance levels were set to α = 0.1.
Multivariate analyses were performed to assess fish assemblages. Similarity matrices were obtained using the Bray–Curtis index based on previously log10(x + 1) transformed abundance; a Non-Metric Multidimensional Scaling (MDS) analysis was then performed. Significant differences in land use factor were determined by one-way ANOSIM analysis. Finally, the specific contribution to the similarity of each group was assessed by SIMPER analysis. Multivariate analysis was performed with Primer-E v6.0 (Clarke and Warwick 2001).
To avoid overestimation of the specific richness, riverine species entering from the Río de la Plata Estuary (Fig. 1. Almirón et al. 2000; Paracampo et al. 2015, 2020) were not considered in the analyses. Four species were determined as riverine: Megaleporinus obtusidens, Astyanax lacustris, Trachelyopterus galeatus and Pimelodus maculatus; they contributed only to 0.05% of the total abundance of the fish assemblage and were excluded from all the analyses.
Results
Environmental parameters
The environmental variables measured in the streams are summarizes in Table 1. The water was well oxygenated, and the pH was slightly alkaline, showing similar values in the two groups of streams sampled. By contrast, water transparency was significantly higher in HSR than in the GSR (p < 0.001). The maximum value was recorded in H1 sampling site (49 cm) and the minimum in G2, 8 cm. Suspended solids and water conductivity were significantly higher in GSR (p < 0.001 and = 0.003, respectively). The maximum value for suspended solids was registered at site G2 reaching 552 mg/l, while highest value for conductivity was recorded in G3 (910 µS/cm).
Soluble reactive phosphorus was significantly higher in HSR than in the GSR (p < 0.001). The minimum SRP concentration was measured in G2 sampling site (14 µg/l), while the maximum (704 µg/l) was recorded in H1. Nitrate and ammonium concentrations showed no significant differences among the two groups of stream reaches studied (Table 1).
All studied sites showed moderate macrophyte cover year-round (~ 30%). The rooted with floating leaves Ludwigia peploides was present in all sites. In HSR, L. peploides was dominant, together with the emergent Gymnocoronis spilanthoides. In GSR, L. peploides was followed by the submerged Myriophyllum aquaticum. Other submerged species, like Potamogeton gayi and Ceratophyllum demersum, were present in these sites, together with the floating Azolla filiculoides.
Insecticide analysis
Pesticide concentrations in sediment samples are summarized in Table 2. Among the eight pesticides analyzed, six were detected in HSR and four in GSR. Deltamethrin and λ-cyhalothrin were detected only in the HSR. Pesticide detection frequencies in samples from HSR (0.52 ± 0.29), were significantly higher (p = 0.049) than those measured in GRS (0.20 ± 0.26).
Concentrations of chlorpyrifos and endosulfan sulphate were significantly higher (p = 0.089 and = 0.066, respectively) in HSR (2.90 and 1.16 ng/g, respectively) than in GSR (0.54 and 0.26 ng/g, respectively). Total pyrethroids and total pesticide concentrations were significantly higher (p = 0.024 and = 0.041, respectively) in HSR (7.09 and 12.88 ng/g, respectively) than those determined in GSR (2.81 and 3.53 ng/g, respectively).
Chlorpyrifos and cypermethrin were the most abundant pesticides, contributing 9–16% and 33–41%, respectively, to total pesticide in both sampling areas. The maximum concentrations of the four pesticides detected in both areas, chlorpyrifos, endosulfan I, endosulfan sulphate and cypermethrin, were found in HSR (amounting to 11.88; 9.25; 3.50 ng/g and 12.60 ng/g, respectively), while the maximum concentrations measured in GSR showed a lower order of magnitude (amounting 0.73–1.71 ng/g).
Fish assemblages
A total of 25,843 fish were registered corresponding to 33 species, 17 families and five orders (Table S3). Characiformes was the order with the highest number of species in each land use, amounting to 15 species in GSR and 12 for HSR, representing 55.5% and 50% of the total, respectively. Twelve species of Characiformes were recorded in common in both land uses, while the species Diapoma terofali, Hyphessobrycon meridionalis and Oligosarcus oligolepis were recorded exclusively in GSR sampling sites.
The order Siluriformes and Cichliformes recorded the same number of species with 7 and 2, respectively, in both land uses. Three species of Cyprinodontiformes were recorded: Austrolebias belotti were exclusively found in grassland sampling sites, while Cnesterodon decemmaculatus and Phalloceros caudimaculatus were recorded in both land use sampling sites.
Specific richness was significantly higher in GSR than in HSR for both OW and VP, p = 0.015 and p = 0.007, respectively (Table 3). Maximum value in OW was recorded at site G3 with 23 species and for VP patches in G1 and G3 with 12 species, respectively. Minimum values were recorded in H3 with 6 species for OW and 3 species for VP in H1. Diversity and evenness in VP were significantly higher in GSR (Table 3) than in HSR (p < 0.001) with the highest values recorded at sites G2 and G1 with values of 1.4 and 0.7, respectively, while the lowest values were recorded at H3 (0.1). By contrast, abundance in VP was significantly higher in HSR (p < 0.001; Table 3). Taxa of the order Cyprinodontiformes showed a clear dominance and contributed to more than 90% of the total capture in HSR (Fig. 2). Maximum value was recorded in H3 with 1505 individuals, while the minimum was recorded in G1, with 21 individuals.

Relative abundance registered in grassland and horticultural Pampean streams, Buenos Aires province, Argentina. H1–H3: horticultural sites in Sauce stream. G1–G3: grassland sites. G1: Chubichaminí stream, G2: Morales stream and G3: Destino stream
NMDS analysis showed that sampling sites were segregated into two groups according to land use for both OW and VP (Fig. 3). The result of the ANOSIM analysis revealed that the groupings obtained according to each land use were significantly different for both OW and VP, Global R = 0.41; p < 0.001 and Global R = 0.43; p < 0.001, respectively.

Ordination of sampling sites according to NMDS analysis in grassland and horticultural Pampean streams, Buenos Aires province, Argentina. H1–H3: horticultural sites in Sauce stream. G1-G3: grassland sites. G1: Chubichaminí stream, G2: Morales stream and G3: Destino stream. Open water (a), vegetated patches (b). au autumn, wi winter, sp spring, su summer
For OW, SIMPER analysis showed that 11 species contributed to the 89.9% of similarity in GSR (Table 4): four species of Characiformes, with Cheirodon interruptus with the largest contribution followed by Pseudocorynopoma doriae, Psalidodon eigenmanniorum, Hyphessobrycon meridionalis and Siluriformes Corydoras longipinnis, accumulating 56.7% of total contribution (Table 4). By contrast, only 6 species contributed to 82.9% in HSR. Cheirodon interruptus and Cyprinodontiformes Cnesterodon decemmaculatus accumulated 51.9% of similarity.
For VP, five species accumulated 79.2% of similarity in GSR, whereas in HSR, three species accounted for 95.6% of similarity (Table 4). The same first three species in each group were arranged in the same order of contribution with C. decemmaculatus followed by C. interruptus and Characidium rachovii; however, the magnitude of contribution was different in each group. In HSR C. decemmaculatus accumulated twice the percentage of similarity (63.0%) compared to that recorded at GSR (31.6%). Finally, Characiformes P. doriae and H. meridionalis completed the similarity recorded in GSR.
Discussion
Water quality was different in each land use; higher SRP concentrations in HSR suggest contributions from applications of fertilizer to the surrounding crops. The SRP concentrations measured in HSR in the present study were consistent with high concentrations (200–900 µg/l) reported by Mugni (2008), Solis et al. (2016, 2017), Arias et al. (2020, 2023) and Cunha et al. (2020) in stream draining agricultural basins in Argentina and Brazil. Higher pesticide detection frequencies and concentrations in sediments of HSR are mainly contributed by runoff events following application to the surrounding crops (Jergentz et al. 2005; Mugni et al. 2012; Paracampo et al. 2012). Concentrations measured in the present study were consistent with higher concentrations reported in sediment samples from stream draining basins with agricultural and horticultural land use (Liess 1998; Arias et al. 2020, 2023; Mac Loughlin et al. 2022).
The occasional pesticide detection at low concentrations in GSR points the long-range atmospheric transport from the application sites (Weber et al. 2010). Pesticides are also detected in protected areas, but detection frequencies and concentrations are lower than those in agricultural areas (Wolfram et al. 2023). The pesticide concentration detected in the grassland streams might also be attributed to applications associated with cattle-pest control within the livestock areas surrounding the reserve. Cypermethrin is used for such a purpose (Ferré et al. 2018) to combat the horn-fly pest in cows (Oyarzún et al. 2008).
Horticultural stream reaches sustain lower specific richness than the less impacted Pampean GSR. Our results agree with several studies reporting decreased fish diversity and biotic integrity in agricultural basins in the USA (Walser and Bart 1999; Pyron and Lauer 2004; Diana et al. 2006). These studies linked decreased diversity to increased water turbidity and substrate homogenization. Similarly, Casatti et al. (2015) and Dala-Corte et al. (2016) reported lower richness and functional diversity in streams with high sedimentation in agricultural basins in Neotropical areas in Brazil. None of these papers measured pesticide concentrations in the streams studied. Unlike the quoted studies increased siltation was not the cause of decreased species richness in the present study because transparency was higher and suspended matter was lower in the horticultural stretches. It seems likely that increased greenhouse coverage decreased soil erosion loss in adjacent streams. Furthermore, irrigation channels provide streams with transparent water with low suspended matter. Fish assemblages in Pampean streams are adapted to high water turbidity and siltation because these characteristics are naturally found in regional streams (Giorgi et al. 2005; Feijoó and Lombardo 2007; Rodrigues Capítulo et al. 2010). The life histories of local communities inhabiting sites with high proportions of fine sediments are adapted to siltation (Crowe and Hay 2004).
Mugni et al. (2012) and Paracampo et al. (2012) simulated rain events in an experimental crop, measured pesticide concentrations and assessed fish toxicity in runoff at different times following application of cypermethrin, endosulfan and chlorpyrifos. Runoff events were toxic to fish for the pesticides assayed, endosulfan persistence being remarkably long, several months since application. The decrease observed in species richness is consistent with the expected effect of repeated toxicity pulses on assemblages composed of species with different pesticide sensitivity.
Characiformes was the most important order in terms of richness and abundance in the grassland streams, as in most not contaminated Neotropical streams (Pinto and Araujo 2007; Gonino et al. 2020). Characiformes are water-column feeders sensitive to water quality degradation (Casatti et al. 2003; Pelicice and Agostinho 2006). In neotropical freshwater environments with abundant vegetation coverage, Characiformes may reach up to 88% of the total abundance and feed mainly on invertebrates, with low interspecific overlapping (Pelicice et al. 2008). In the present study the Cyprinodontiformes Cnesterodon decemmaculatus became dominant in HSR, contributions to similarity being much higher than those in GSR. Cnesterodon decemmaculatus is tolerant to environmental pollution, occasionally the only taxa present in heavily contaminated water bodies (Paracampo et al. 2020; Paredes del Puerto et al. 2021). The dominance of C. decemmaculatus was evident in lower diversity and evenness in the VP of HSR.
Solis et al. (2018) reported lower invertebrate densities in Pampeanstreams of agricultural basins. Arias et al. (2020) recorded lower taxonomic richness, density and absence of sensitive taxa to insecticide exposure in Pampean streams of horticultural basins, including the stream sampled in the present study. Thus, decreased trophic resources might have contributed to the impoverished fish assemblage observed in the HSR of the present study. C. decemmaculatus in polluted environments feeds mostly on detritus (Quintans et al. 2009). Once the community was impoverished by sensitive species at low density, tolerant ones increased abundance due to the lower competence for resources available. The species of Characiformes Cheirodon interruptus and Characidium rachovii that contributed more to similarity in GSR than in HSR and the species that only contributed in GSR Pseudocorynopoma doriae and Hyphessobrycon meridionalis are commonly abundant in Pampean streams with low anthropogenic impact (Paracampo et al. 2020; Paredes del Puerto et al. 2021, 2022) and prey mainly on invertebrates (Fernandez et al. 2012; Brancolini et al. 2015).
For the assemblages sampled in OW environments, only significant differences in specific richness were recorded. Nevertheless, multivariate analysis showed different groupings among land use and the different contribution of species to each group. Eleven species contributed to similarity in the GSR sampling sites while only five did so in HSR. In addition, the tolerant species C. decemmaculatus was last in the list of contributors in GSR, whereas it ranked second in HSR. An increase in dissimilarity seen in OW environments between the group of sampling sites may result from an environmental impact of the toxicity pulses caused by insecticides. Disturbances do not always translate into changes in diversity or biotic homogenization (Hawkins et al. 2015).
Conclusion
Our study showed deterioration of water quality associated with horticultural land use in Neotropical lowland streams. Pesticides were found in sediments of the horticultural basins. Detection frequencies and concentrations were significantly higher than those in grassland basins with lower impact. Deterioration of water quality resulted in lower richness of species in the horticultural streams. Lower diversity and evenness and higher abundance were evidenced in the vegetated stretches of horticultural basins.
The on-going horticulture expansion leads to further deterioration resulting in ruderal fish community within the region, unless managerial practices are implemented. Monitoring surveys are needed to assess the effect on fish biodiversity and evaluate pollution mitigation practices.
Data availability
The datasets generated during and/or analyzed in the current study are available from the corresponding author on reasonable request.
References
Albert JS, Tagliacollo VA, Dagosta F (2020) Diversification of neotropical freshwater fishes. Annu Rev Ecol Evol Syst 51:27–53. https://doi.org/10.1146/annurev-ecolsys-011620-031032
Albert JS, Destouni G, Duke-Sylvester SM, Magurran AE, Oberdorff T, Reis RE, Winnemiller KO, Ripple WJ (2021) Scientists’ warning to humanity on the freshwater biodiversity crisis. Ambio 50(1):85–94. https://doi.org/10.1007/s13280-020-01318-8
Allan JD (2004) Landscapes and riverscapes: the influence of land use on stream ecosystems. Annu Rev Ecol Evol Syst 35:257–284. https://doi.org/10.1146/annurev.ecolsys.35.120202.110122
Almirón AE, García ML, Menni RC, Protogino L, Solari LC (2000) Fish ecology of a seasonal lowland stream in temperate South America. Mar Freshw Res 51(3):265–274
Almirón AE, Casciotta JR, Ciotek L, Giorgis P (2015) Guía de los peces del Parque Nacional Pre-Delta. Administración de Parque Nacionales, Buenos Aires
APHA (2012) Standard methods for the examination of water and wastewater, 22nd edn. American Public Health Association, Washington DC, p 1360
Aquino AE (1997) Las especies de Hypoptomatinae (Pisces, Siluriformes, Loricariidae) en la Argentina. Revista De Ictiología 5:5–21
Arias M, Scalise A, Solis M, Paracampo A, Indaco M, Fanelli S, Mugni H, Bonetto C (2020) Horticulture affects macroinvertebrate assemblages in adjacent streams (Buenos Aires, Argentina). Knowl Manag Aquat Ecosyst 421(5):1–10. https://doi.org/10.1051/kmae/20190481
Arias M, Bonetto C, Fanelli SL, Scenna L, Miglioranza KSB, Mugni H (2023) Macroinvertebrate assemblages in lowland streams under horticultural impact (Buenos Aires, Argentina). Hydrobiologia 850:399–416. https://doi.org/10.1007/s10750-022-05081-7
Athor J (2009) Parque Costero del Sur Naturaleza, conservación y patrimonio cultural. Fundación de Historia Natural “Félix de Azara”, Buenos Aires, pp 562
Azpelicueta MM, Braga L (1991) Los curimátidos en Argentina. Fauna De Agua Dulce De La República Argentina 40(1):1–53
Baldini C, Marasas ME, Tittonell P, Drozd AA (2022) Urban, periurban and horticultural landscapes–Conflict and sustainable planning in La Plata district, Argentina. Land Use Policy 117:106120. https://doi.org/10.1016/j.landusepol.2022.106120
Bertora A, Grosman F, Sanzano P, Rosso JJ (2024) Impoverished fish assemblages of temperate Neotropical streams respond to environmental degradation and support a sensitive Index of Biotic Integrity. Hydrobiologia 851(2):383–408
Braga L (1993) Los Anostomidae (Pisces, Characiformes) de Argentina. In: Castellanos ZA (ed) Fauna de agua dulce de la República Argentina. Profadu, CONICET, La Plata, pp 5–45.
Braga L (1994) Los Characidae de Argentina de las subfamilias Cynopotaminae y Ancestrorhynchinae. In: Castellanos ZA (ed) Fauna de Agua Dulce de la República Argentina. Profadu, CONICET, La Plata, pp 21–29.
Brancolini F, Maroñas ME, Sendra ED (2015) Dieta de Pseudocorynopoma doriae (Characiformes: Characidae) en el arroyo de La Choza, Buenos Aires, Argentina. Biología Acuática 30:259–655. https://revistas.unlp.edu.ar/bacuatica/article/view/7726. Accessed March 2023
Brauns M, Allen DC, Boëchat IG et al (2022) A global synthesis of human impacts on the multifunctionality of streams and rivers. Glob Change Biol 28:4783–4793. https://doi.org/10.1111/gcb.16210
Cabrera AL, Zardini EM (1993) Manual de la Flora de los alrededores de Buenos Aires. ACME, segunda edición aumentada y actualizada, Buenos Aires, Argentina, pp 755
Casatti L, Mendes HF, Ferreira KM (2003) Aquatic macrophytes as feeding site for small fishes in the Rosana Reservoir, Paranapanema River, Southeastern Brazil. Braz J Biol 63:213–222. https://doi.org/10.1590/S1519-69842003000200006
Casatti L, Teresa FB, Zeni JDO, Ribeiro MD, Brejao GL, Ceneviva-Bastos M (2015) More of the same: high functional redundancy in stream fish assemblages from tropical agroecosystems. Environ Manag 55:1300–1314. https://doi.org/10.1007/s00267-015-0461-9
Casciotta JR, Almirón A, Bechara JA (2005) Peces del Iberá: hábitat y diversidad. Grafikar, La Plata.
Castro R, Polaz CN (2019) Small-sized fish: the largest and most threatened portion of the megadiverse neotropical freshwater fish fauna. Biota Neotrop. https://doi.org/10.1590/1676-0611-BN-2018-0683
Clarke KR, Warwick RM (2001) Change in Marine Communities. An Approach to Statistical Analysis and Interpretation. 2nd Edition, PRIMER-E, Ltd., Plymouth Marine Laboratory, Plymouth.
Cornejo A, Tonin AM, Checa B, Tuñon AR, Pérez D et al (2019) Effects of multiple stressors associated with agriculture on stream macroinvertebrate communities in a tropical catchment. PLoS ONE 14(8):e0220528. https://doi.org/10.1371/journal.pone.0220528
Cortelezzi A, Sierra MV, Gómez N, Marinelli C, Rodrigues Capítulo A (2013) Macrophytes, epipelic biofilm, and invertebrates as biotic indicators of physical habitat degradation of lowland streams (Argentina). Environ Monit Assess 185:5801–5815. https://doi.org/10.1007/s10661-012-2985-2
Crowe A, Hay J (2004) Effects of fine sediments on river biota. Motueka Integrated Catchment Management Programme 951, pp 38
Cunha DGF, Finkler NR, Gómez N, Cochero J, Donadelli JL, Saltarelli WA, Calijuri MDC, Miwa PAC, Tromboni F, Dodds WK, Gonçalves Boëchat L, Gücker B, Thomas SA (2020) Agriculture influences ammonium and soluble reactive phosphorus retention in South American headwater streams. Ecohydrology 13:e2184. https://doi.org/10.1002/eco.2184
Cunico AM, Allan JD, Agostinho AA (2011) Functional convergence of fish assemblages in urban streams of Brazil and the United States. Ecol Ind 11(5):1354–1359. https://doi.org/10.1016/j.ecolind.2011.02.009
Dala-Corte RB, Giam X, Olden JD, Becker FG, Guimarães TDF, Melo AS (2016) Revealing the pathways by which agricultural land-use affects stream fish communities in South Brazilian grasslands. Freshw Biol 61(11):1921–1934. https://doi.org/10.1111/fwb.12825
Davies-Colley RJ, Nagels JW, Smith RA, Young RG, Phillips CJ (2004) Water quality impact of a dairy cow herd crossing a stream. N Z J Mar Freshw Res 38(4):569–576. https://doi.org/10.1080/00288330.2004.9517262
Diana M, Allan JD, Infante D (2006) The influence of physical habitat and land use on stream fish assemblages in southeastern Michigan. Am Fish Soc Sympos 48:359–374
Dolédec S, Statzner B (2010) Responses of freshwater biota to human disturbances: contribution of J-NABS to developments in ecological integrity assessments. J N Am Benthol Soc 29(1):286–311. https://doi.org/10.1899/08-090.1
DP 2015. Defensoría del Pueblo de la Provincia de Buenos Aires. Relevamiento de la utilización de Agroquímicos en la Provincia de Buenos Aires – Mapa de Situación e incidencias sobre la salud 497–532.
Dudley N, Alexander S (2017) Agriculture and biodiversity: a review. Biodiversity 18(2–3):45–49. https://doi.org/10.1080/14888386.2017.1351892
Effert-Fanta EL, Fischer RU, Wahl DH (2019) Effects of riparian forest buffers and agricultural land use on macroinvertebrate and fish community structure. Hydrobiologia 841:45–64. https://doi.org/10.1007/s10750-019-04006-1
Feijoó CS, Lombardo RJ (2007) Baseline water quality and macrophyte assemblages in Pampean streams: a regional approach. Water Res 41(7):1399–1410. https://doi.org/10.1016/j.watres.2006.08.026
Fernández EM, Ferriz RA, Bentos CA, López GR (2012) Dieta y ecomorfología de la ictiofauna del arroyo Manantiales, provincia de Buenos Aires, Argentina. Revista Del Museo Argentino De Ciencias Naturales 14(1):1–13
Ferré DM, Quero AAM, Hernández AF, Hynes V, Tornello MJ, Lüders C, Gorla NBM (2018) Potential risks of dietary exposure to chlorpyrifos and cypermethrin from their use in fruit/vegetable crops and beef cattle productions. Environ Monit Assess 190(5):292. https://doi.org/10.1007/s10661-018-6647-x
Fitzpatrick FA, Scudder BC, Lenz BN, Sullivan DJ (2001) Effects of multi-scale environmental characteristics on agricultural stream biota in eastern Wisconsin. J Am Water Resour Assoc 37(6):1489–1507. https://doi.org/10.1111/j.1752-1688.2001.tb03655.x
Fricke R, Eschmeyer WN, Van der Laan R (2023) Eschmeyer’s catalog of fishes: genera, species, references [Internet]. San Francisco: California Academy of Science. Available from: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp.. Accessed March 2023
Giorgi A, Feijoó C, Tell G (2005) Primary producers in a Pampean stream: temporal variation and structuring role. Biodivers Conserv 14:1699–1718. https://doi.org/10.1007/s10531-004-0694-z
Gonino G, Benedito E, Cionek VDM, Ferreira MT, Oliveira JM (2020) A fish-based index of biotic integrity for neotropical rainforest sandy soil streams—Southern Brazil. Water 12(4):1215. https://doi.org/10.3390/w12041215
Goss LM, Roper BB (2018) The relationship between measures of annual livestock disturbance in Western riparian areas and stream conditions important to trout, salmon, and char. West N Am Nat 78(1):76–91. https://doi.org/10.3398/064.078.0108
Harrison S, McAree C, Mulville W, Sullivan T (2019) The problem of agricultural ‘diffuse’ pollution: getting to the point. Sci Total Environ 677:700–717. https://doi.org/10.1016/j.scitotenv.2019.04.169
Hawkins CP, Mykrä H, Oksanen J, Vander Laan JJ (2015) Environmental disturbance can increase beta diversity of stream macroinvertebrate assemblages. Glob Ecol Biogeogr 24(4):483–494. https://doi.org/10.1111/geb.12254
Helms BS, Feminella JW, Pan S (2005) Detection of biotic responses to urbanization using fish assemblages from small streams of western Georgia, USA. Urban Ecosyst 8:39–57. https://doi.org/10.1007/s11252-005-1418-1
Hurtado MA, Giménez JE, Cabral MG, da Silva M, Martínez OR, Camilión MC, Sánchez CA, Muntz D, Gebhard JA, Forte, LM, Boff LD, Crincoli A, Lucesoli H (2006) Análisis ambiental del partido de La Plata. Aportes al ordenamiento territorial. Buenos Aires: Consejo Federal de Inversiones pp 124
Jergentz S, Mugni H, Bonetto C, Schulz R (2005) Assessment of insecticide contamination in runoff and stream water of small agricultural streams in the main soybean area of Argentina. Chemosphere 61(6):817–826. https://doi.org/10.1016/j.chemosphere.2005.04.036
Junk WJ, Wantzen KM (2004) The flood pulse concept: new aspects, approaches and applications - an update. In: Welcomme RL, Petr T (eds) Proceedings of the Second International Symposium on the Management of Large Rivers for Fisheries, pp 117–149. Bangkok: Food and Agriculture Organization and Mekong River Commission, FAO Regional Office for Asia and the Pacific. https://hdl.handle.net/11858/00-001M-0000-000F-DB40-5.. Accessed April 2023
Lévêque C, Oberdorff T, Paugy D, Stiassny MLJ, Tedesco PA (2007) Global diversity of fish (Pisces) in freshwater. In: Balian EV, Lévêque C, Segers H, Martens K (eds) Freshwater animal diversity assessment. Developments in hydrobiology, vol 198. Springer, Dordrecht, pp 545–567. https://doi.org/10.1007/978-1-4020-8259-7_53
Liess M (1998) Significance of agricultural pesticides on stream macroinvertebrate communities. Internationale Vereinigung Für Theoretische Und Angewandte Limnologie: Verhandlungen 26(3):1245–1249. https://doi.org/10.1080/03680770.1995.11900921
Mac Loughlin TM, Peluso ML, Marino DJ (2022) Multiple pesticides occurrence, fate, and environmental risk assessment in a small horticultural stream of Argentina. Sci Total Environ 802:149893. https://doi.org/10.1016/j.scitotenv.2021.149893
Magurran, AE, Phillip DA (2001) Implications of species loss in freshwater fish assemblages. Ecography 24(6):645–650. https://doi.org/10.1111/j.1600-0587.2001.tb00526.x
Matono P, Sousa D, Ilhéu M (2013) Effects of land use intensification on fish assemblages in Mediterranean climate streams. Environ Manag 52:1213–1229. https://doi.org/10.1007/s00267-013-0152-3
Messetta ML, Anselmo JH, Gantes P, Pérez B, Feijoó CS (2023) Plantas acuáticas de arroyos pampeanos: una guía de campo: revalorizando nuestro ambiente. Luján, Libros del INEDES, pp 76
Miquelarena AM, Menni RC (2005) Astyanax tumbayaensis, a new species from northwestern Argentina highlands (Characiformes: Characidae) with a key to the Argentinean species of the genus and comments on their distribution. Rev Suisse Zool 112:661–676. https://doi.org/10.5962/bhl.part.80319
Mirande JM, Koerber S, 2020 Checklist of the Freshwater Fishes of Argentina. 2nd edition (CLOFFAR-2). Ichthyological Contributions of Peces Criollos 72: 1–81. https://usercontent.one/wp/pecescriollos.de/wp-content/uploads/2020/12/ICP-72-Mirande-Koerber-2020-CLOFFAR-2.pdf
Monaghan RM, Wilcock RJ, Smith LC, Tikkisetty B, Thorrold BS, Costall D (2007) Linkages between land management activities and water quality in an intensively farmed catchment in southern New Zealand. Agric Ecosyst Environ 118(1–4):211–222. https://doi.org/10.1016/j.agee.2006.05.016
Mugni H, Demetrio P, Paracampo A, Pardi M, Bulus G, Bonetto C (2012) Toxicity persistence in runoff water and soil in experimental soybean plots following chlorpyrifos application. Bull Environ Contam Toxicol 89:208–212. https://doi.org/10.1007/s00128-012-0643-6
Mugni HD (2008) Concentración de nutrientes y toxicidad de pesticidas en aguas superficiales de cuencas rurales. Tesis Doctoral. Universidad Nacional de La Plata. https://doi.org/10.35537/10915/4410
Nessimian JL, Venticinque EM, Zuanon J, De Marco P, Gordo M, Fidelis L, Juen L (2008) Land use, habitat integrity, and aquatic insect assemblages in Central Amazonian streams. Hydrobiologia 614:117–131. https://doi.org/10.1007/s10750-008-9441-x
Oyarzún MP, Quiroz A, Birkett MA (2008) Insecticide resistance in the horn fly: alternative control strategies. Med Vet Entomol 22(3):188–202. https://doi.org/10.1111/j.1365-2915.2008.00733.x
Paracampo A, Mugni HD, Demetrio PM, Pardi MH, Bulus GD, Asborno MD, Bonetto CA (2012) Toxicity persistence in runoff and soil from experimental soybean plots following insecticide applications. J Environ Sci Health B 47(8):761–768
Paracampo A, García I, Mugni H, Marrochi N, Carriquiriborde P, Bonetto C (2015) Fish assemblage of a Pampasic stream (Buenos Aires, Argentina): temporal variations and relationships with environmental variables. Stud Neotrop Fauna Environ 50:145–153. https://doi.org/10.1080/01650521.2015.1065658
Paracampo A, Marrochi N, García I, Maiztegui T, Carriquiriborde P, Bonetto C, Mugni H (2020) Fish assemblages in Pampean streams (Buenos Aires, Argentina): relationship to abiotic and anthropic variables. An Acad Bras Ciênc 92(2):e20190476. https://doi.org/10.1590/0001-3765202020190476
Paredes del Puerto JM, Paracampo AH, García ID, Maiztegui T, Garcia de Souza JR, Maroñas ME, Colautti DC (2021) Fish assemblages and water quality in Pampean streams (Argentina) along an urbanization gradient. Hydrobiologia 848(19):4493–4510. https://doi.org/10.1007/s10750-021-04657-z
Paredes del Puerto JM, García ID, Maiztegui T, Paracampo AH, Rodrigues Capítulo L, Garcia de Souza JR, Colautti DC (2022) Impacts of land use and hydrological alterations on water quality and fish assemblage structure in headwater Pampean streams (Argentina). Aquat Sci 84(1):6. https://doi.org/10.1007/s00027-021-00836-1
Paredes del Puerto JM, Sathicq MB, Altieri P, Nicolosi Gelis MM, Paracampo A, Pazos RS, Tarda AS, Gómez N, Colautti D (2024) Extreme drought conditions interact with urbanisation, affecting hydrological regimes and water quality in temperate lowland streams. Aquat Sci 86(1):13
Pelicice FM, Agostinho AA (2006) Feeding ecology of fishes associated with Egeria spp. patches in a tropical reservoir, Brazil. Ecol Freshw Fish 15(1):10–19. https://doi.org/10.1111/j.1600-0633.2005.00121.x
Pelicice FM, Thomaz SM, Agostinho AA (2008) Simple relationships to predict attributes of fish assemblages in patches of submerged macrophytes. Neotrop Ichthyol 6(4):543–550. https://doi.org/10.1590/S1679-62252008000400001
Pelicice FM, Bialetzki A, Camelier P, Carvalho FR, García-Berthou E, Pompeu PS, Teixeira de Mello FT, Pavanelli CS (2021) Human impacts and the loss of Neotropical freshwater fish diversity. Neotrop Ichthyol 19(3):e210134. https://doi.org/10.1590/1982-0224-2021-0134
Pinto BCT, Araújo FG (2007) Assessing of biotic integrity of the fish community in a heavily impacted segment of a tropical river in Brazil. Braz Arch Biol Technol 50(3):489–502. https://doi.org/10.1590/S1516-89132007000300015
Pyron M, Lauer TE (2004) Hydrological variation and fish assemblage structure in the middle Wabash River. Hydrobiologia 525:203–213. https://doi.org/10.1023/B:HYDR.0000038867.28271.45
Quintans F, Scasso F, Loureiro M, Yafe A (2009) Diet of Cnesterodon decemmaculatus (Poeciliidae) and Jenynsia multidentata (Anablepidae) in a hypertrophic shallow lake of Uruguay. Iheringia Série Zoologia 99(1):99–105. https://doi.org/10.1590/S0073-47212009000100014
Reis RE, Pereira EHL (2000) Three new species of the loricariid catfish genus Loricariichthys (Teleostei: Siluriformes) from southern South America. Copeia 4:1029–1047. https://doi.org/10.1643/0045-8511(2000)000[1029:TNSOTL]2.0.CO;2
Říčan O, Kullander SO (2008) The Australoheros (Teleostei: Cichlidae) species of the Uruguay and Paraná River drainages. Zootaxa 1724:1–51. https://doi.org/10.5281/zenodo.181173
Rodrigues Capítulo A, Gómez N, Giorgi A, Feijoó C (2010) Global changes in Pampean lowland streams (Argentina): implications for biodiversity and functioning. In: Stevenson RJ, Sabater S (eds) Global change and river ecosystems—implications for structure, function and ecosystem services. Developments in Hydrobiology 215. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-0608-8_5
Rosso JJ, González-Castro M, Bogan S, Cardoso YP, Mabragaña E, Delpiani M, Díaz Astarloa JM (2018) Integrative taxonomy reveals a new species of the Hoplias malabaricus species complex (Teleostei: Erythrinidae). Ichthyol Explor Freshw 28:235–252. https://doi.org/10.23788/IEF-1076
Sala OE, Chapin FS III, Armesto JJ, Berlow E, Bloomfield J, Dirzo R, Huber-Sanwald E, Huenneke LF, Jackson RB, Kinzig A, Leemans R, Lodge DM, Mooney HA, Oesterheld M, Poff L, Sykes MT, Walker BH, Walker M, Wall DH (2000) Global Biodiversity Scenarios for the Year 2100. Science 287(5479):1770–1774. https://doi.org/10.1126/science.287.5459.1770
Sigmaplot version 12 for Windows (Systat Software, Inc., U.S.A.)
Solis M, Mugni H, Hunt L, Marrochi N, Fanelli S, Bonetto C (2016) Land use effect on invertebrate assemblages in Pampasic streams (Buenos Aires, Argentina). Environ Monit Assess 188:1–12. https://doi.org/10.1007/s10661-016-5545-3
Solis M, Mugni HD, Fanelli SL, Bonetto CA (2017) Effect of agrochemicals on macroinvertebrate assemblages in Pampasic streams, Buenos Aires, Argentina. Environ Earth Sci 76(180):1–13. https://doi.org/10.1007/s12665-017-6476-1
Solis M, Bonetto C, Marrochi N, Paracampo A, Mugni H (2018) Aquatic macroinvertebrate assemblages are affected by insecticide applications on the Argentine Pampas. Ecotoxicol Environ Saf 148:11–16. https://doi.org/10.1016/j.ecoenv.2017.10.017
Terán GE, Benitez MF, Mirande JM (2020) Opening the Trojan horse: phylogeny of Astyanax, two new genera and resurrection of Psalidodon (Teleostei: Characidae). Zool J Linn Soc 190(4):1217–1234
Teresa FB, Casatti L (2017) Trait-based metrics as bioindicators: responses of stream fish assemblages to a gradient of environmental degradation. Ecol Ind 75:249–258
Thomas KE, Lazor R, Chambers PA, Yates AG (2018) Land-use practices influence nutrient concentrations of southwestern Ontario streams. Can Water Resour J/revue Canadienne Des Ressources Hydriques 43(1):2–17. https://doi.org/10.1080/07011784.2017.1411211
Walser CA, Bart HL (1999) Influence of agriculture on in-stream habitat and fish community structure in Piedmont watersheds of the Chattahoochee River System. Ecol Freshw Fish 8(4):237–246. https://doi.org/10.1111/j.1600-0633.1999.tb00075.x
Wang L, Lyons J, Kanehl P, Gatti R (1997) Influences of watershed land use on habitat quality and biotic integrity in Wisconsin streams. Fisheries 22(6):6–12. https://doi.org/10.1577/1548-8446(1997)022%3C0006:IOWLUO%3E2.0.CO;2
Weber J, Halsall CJ, Muir D, Teixeira C, Small J, Solomon K, Hermanson M, Hung H, Bidleman T (2010) Endosulfan, a global pesticide: a review of its fate in the environment and occurrence in the Arctic. Sci Total Environ 408(15):2966–2984. https://doi.org/10.1016/j.scitotenv.2009.10.077
Wolfram J, Bub S, Petschick LL, Schemmer A, Stehle S, Schulz R (2023) Pesticide occurrence in protected surface waters in nature conservation areas of Germany. Sci Total Environ 858(3):160074. https://doi.org/10.1016/j.scitotenv.2022.160074
XLSTAT Version 2014.5.03 Copyright Addinsoft 1995–2014 (2014) XLSTAT and Addinsoft are Registered Trademarks of Addinsoft. https://www.xlstat.com.. Accessed March 2023
You J, Weston D, Lydy MA (2004) Sonication extraction method for the analysis of pyrethroid, organophosphate, and organochlorine pesticides from sediment by gas chromatography with electron-capture detection. Arch Environ Contam Toxicol 47:141–147. https://doi.org/10.1007/s00244-003-3165-8
Zeni JO, Hoeinghaus DJ, Casatti L (2017) Effects of pasture conversion to sugarcane for biofuel production on stream fish assemblages in tropical agroecosystems. Freshw Biol 62(12):2026–2038. https://doi.org/10.1111/fwb.13047
Acknowledgements
The authors thank the Argentine National Scientific and Technical Research Council (CONICET) and the financial support of the Argentine National Agency for the Promotion of Science and Technology (ANPCyT; PICT 2016-0902). We are also grateful to Sergio Bogan, curator of the ichthyology collection, for receiving and storing the vouchers at the Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" (MACN-CONICET). This paper is Scientific Contribution N° 1259 of the Institute of Limnology “Dr. Raúl A. Ringuelet” (ILPLA, CCT-La Plata CONICET, UNLP).
Author information
Authors and Affiliations
Contributions
J.M.P.d.P investigation, conceptualization, field samplings, formal analysis, data curation, writing of the original draft, editing and visualization. H.M. and N.C. methodology, data processing, investigation, laboratory measurements, writing and editing. M.A. and S.F. field samplings, laboratory measurements, editing. C.B. conceptualization writing, review and editing. A.P. designed the study, field samplings, investigation, conceptualization, methodology, resources, data processing, writing of the original draft, writing and editing, funding acquisition, project administration and supervision. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that may have influenced the work reported in this paper.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Paredes del Puerto, J., Mugni, H., Cappelletti, N. et al. Horticultural land use effect on fish assemblages in Neotropical lowland streams, Buenos Aires, Argentina. Aquat Sci 86, 62 (2024). https://doi.org/10.1007/s00027-024-01076-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00027-024-01076-9