
Abstract
Cassava is a key food security staple and a competitive feedstock for multiple industrial processes and end uses. Farmers grow hybrids which are reproduced vegetatively. Several programs have used the same breeding scheme for the last 40 years. Significant progress has been made, particularly with the first improved varieties released in the 1980s and 1990s. However, gains slowed down since then. Biotechnology tools, after more than two decades, have not yet had impact on increasing yields. Results from ongoing genomic selection show promising results for high-heritability traits, but not for fresh root yield (FRY). Key challenges to increase FRY are the strong influence of nonadditive genetic effects and the heterozygous nature of breeding parents. There is large within-family variation masking the true breeding value of each progenitor. To improve yields, breeding must shift from making crosses where breeders hope to find hybrids that are superior to those already available (a strategy that has made only slow progress in the last decade or two) to develop and improve inbred progenitors that can produce more reliably better hybrids. The use of inbred progenitors and implementation of reciprocal recurrent selection should be an efficient way to exploit heterosis and epistasis, which are large components in the determination of FRY. Induction of flowering would also accelerate genetic gains. In the near-term future, available molecular markers can be used to improve the breeding value of progenitors rather than in selection of segregating progenies. They can also be used to screen germplasm collections in search of useful traits.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
- Cassava
- Manihot esculenta
- Cassava bacterial blight (CBB)
- Cassava brown streak virus (CBSV)
- Cyanogenic glucosides
- Cassava mosaic disease (CMV)
- Harvest index
- Selection index
- Polyploid
- Post-harvest physiological deterioration (PPD)
1 History of Cultivation
Cassava is a crop of Neotropical origin and significant economic relevance, particularly in the lowland tropics. Its main product is the starchy roots that are generally harvested about a year after planting. It has been suggested that the Manihot genus emerged in Mesoamerica and from there diversified to the north and south (Deputié et al. 2011). Endemic Manihot species can be found from southwestern North America to central Argentina (Rogers and Appan 1973; Nassar and Ortiz 2008; Deputié et al. 2011). The taxonomy of the genus has been generally understudied. Rogers and Appan (1973) in their comprehensive monograph described 98 Manihot species. More recent taxonomic updates were done by Allem (1999, 2002), Allem et al. (2001) and Second et al. (1997).
The botanical and geographic origin of cultivated cassava is still unclear. Early publications suggested that cassava had an unknown ancestry but was likely the by-product of indiscriminate introgression among some of the wild relatives in Mexico and Mesoamerica, probably including M. aesculifolia (H.B.K.) Pohl (Rogers and Appan 1973; Bertram 1993; Bertram and Schaal 1993). Renvoize (1973) suggested that cassava was domesticated in Mesoamerica and northern South America. However, today the prevailing hypothesis is that cultivated cassava originated in South America (Allem 2002; Olsen and Schaal 1999, 2001; Nassar and Ortiz 2008). Allem suggested in different articles (Allem 1999, 2002; Allem et al. 2001) that cultivated cassava was domesticated directly from an extant wild species (either Manihot esculenta Crantz ssp. flabellifolia (Pohl) Ciferri or Manihot esculenta Crantz ssp. peruviana (Mueller Argoviensis)). Another contrasting view of the South American origin of cultivated cassava was proposed by Nassar in 1978, suggesting that the origin and domestication of cultivated cassava occurred from a natural hybrid probably between M. pilosa and other species (Nassar 1978, 2000). It is possible that several independent domestication events have taken place in different sites and times. Upon domestication, cassava was disseminated through tribal migrations (particularly the Arawak people) in pre-Columbian times (Nassar 2000).
The timing of domestication has also not been determined. Archaeological evidence of vegetatively propagated crops is generally limited, and cassava is no exception. It is clear, however, that the Mochica culture flourishing in the northern coast of Peru (2200 through 1200 years ago) knew about cassava, as illustrated by a beautiful ceramic artwork (www.museolarco.org). Domestication of cassava, therefore, must have taken place at least 3000 years ago. Cassava was widely grown in pre-Columbian times. The Portuguese introduced the crop into West Africa in the 1500s where it spread quickly thanks to its resilience, flexibility of harvest and diversity of uses. From Africa, cassava spread eastwards and eventually into Asia, where it had also been introduced by the Spanish into the Philippines (Byrne 1984).
More than 20 million ha of cassava are harvested annually, most of it (72%) in Africa, followed by 18% in SE Asia and 12% in the Americas (FAOSTAT, averages from the last three available years – 2012–2014). Worldwide average yield (2012–2014 avg.) is 11.1 t of fresh roots per ha but varies widely (8.4; 12.3 and 21.3 t/ha, respectively, in Africa, America and Asia). Nigeria, Brazil and Thailand are interesting examples, for cassava production and use, in their respective continents. Nigeria is the largest producer of cassava worldwide, with more than 6.7 million ha planted and 51.1 million t of fresh roots harvested (average yield of 7.6 t/ha). Brazil plants about 1.6 million ha with an average yield of 14.2 t/ha. However, large differences in productivity can be observed, for example, between NE and Southern Brazil with average yields of 10.3 and 19.8 t/ha, respectively, for the 2001–2013 period (IBGE 2014). Thailand plants 1.4 million ha of cassava each year, and the average yield is above 22.0 t/ha. It can be postulated that productivity of cassava is closely linked to the strength and stability of markets. SE Asia and Southern Brazil have well-established and reliable markets for cassava which explains the motivation to achieve average productivity above 20 t/ha in those regions. In Africa, on the other hand, markets are not so well developed thus partially explaining the low average productivity of about 8 t/ha. These figures are helpful to understand that proper technologies have been developed for cassava to express its high yield potential, but their adoption by farmers depends on the market perspectives.
Among the key technologies available to farmers are improved varieties. One of the first reports on cassava variety assessment and selection was published in Brazil, in 1899 (Zehntner 1919), reported by Gonçalves Fukuda et al. (2002). Modern breeding programs were initiated during the first half of the last century (Byrne 1984; Jennings and Iglesias 2002) in Brazil (Graner 1935; Lozano et al. 1978; Normanha 1970), Ghana (Hahn et al. 1979), India (Abraham 1957), Indonesia, (Bolhuis 1953), Madagascar, (Cours 1951), and Tanzania (Nichols 1947; Jennings 1957). However, with the exception of Brazil, most of these early efforts were discontinued as a result of the dismantling of the colonial system. The creation of cassava improvement programs at the International Institute of Tropical Agriculture (IITA based in Ibadan, Nigeria) and Centro Internacional de Agricultura Tropical (CIAT in Cali, Colombia) broke the previous isolation that breeding efforts had had in the past (Byrne 1984) and contributed to the establishment of vigorous and successful programs in the most important cassava-growing countries. Most of the information presented in this chapter has been generated by the collaboration between the international centres and national agriculture research programs.
Markets in cassava are diverse. The crop was initially domesticated for the direct use of the roots, which contain little else in addition to starch (e.g. low concentration of proteins, fat/oil and traces of micronutrient mineral and vitamins). Low to very high levels of cyanogenic glucosides (CG) can be found in roots from sweet/cool or bitter cassava cultivars, respectively. CG are eliminated through alternative processing techniques. Different cultures developed diverse ethnic products from cassava roots, for example: gari, kokonte and fufu in Africa; farinha and cassabe in the Americas; and krupuk, gaplek and sago pearls in Asia (Cock 1985; Nweke 2004). Processing roots soon after the harvest is also important to prevent post-harvest physiological deterioration (PPD ) that spoils roots 2–3 days after harvest. Dried chips or silage of cassava roots can be used for animal feeding. Cassava foliage is also used for human consumption, particularly in Africa (Diasolua Ngudi et al. 2003; Lancaster and Brooks 1983; Muoki and Maziya-Dixon 2010), animal feeding (Balagopalan 2002; Buitrago 1990; FAO 2013) and even insect production for human consumption (Caparros Megido et al. 2016).
Globally, in the period 1970–2003, the main uses of cassava roots were for food (54%), followed by feed (30%) and other uses including starch production (4%) (Prakash 2008). During this period, however, there were considerable changes, with a clear trend of increasing industrial processing. Global use of cassava for feed was affected by the reduction of imports from the European Union in the 1980s. Production of starch on the other hand increased considerably in the same period (by 17.5% annually according to Prakash 2008). Today, cassava is the second most important source of starch worldwide (Stapleton 2012). In the 2000s a considerable amount of cassava roots started to be used for the production of fuel ethanol (Anyanwu et al. 2016; Chen et al. 2016; Liang et al. 2016; Nguyen et al. 2007; Sriroth et al. 2010).
The diversity of uses for cassava roots and foliage illustrates the challenges that cassava breeders face. Each end use imposes a set of requirements, sometimes contrasting, that varieties need to meet for them to be acceptable to farmers and processors.
2 Biology
Cassava is a perennial species usually grown as an annual crop. The roots can be harvested from 6 up to 24 months after planting (MAP ). More typically, however, farmers harvest cassava 10–12 MAP, at the end of the dry season (or before the cold season in subtropical regions), when root quality would be optimum, and store the stems only for a few weeks until the arrival of the rains (or several weeks during winter). Commercial multiplication of cassava is achieved by planting stem cuttings. Farmers grow clonal hybrids, so after having access to planting material of a good variety they only have to make sure that health, nutrition and physiological status of the planting material is maintained at optimum levels. There is a genetic component, however, of the capacity of planting material to withstand storage and to sprout quickly and vigorously (Ceballos et al. 2011). This characteristic has become more critical in recent years due to the more erratic arrival of the rains as a result of climate change.
Farmers collect the main stems before harvesting the roots. Depending on the variety and growing conditions, length of harvested stems can range from 50 to 200 cm. There is certain variation in sprouting capacity/vigour, depending on the sections of the stem from which the cuttings come. In a 10–12-month-old cassava, the best cuttings correspond to the middle half section of the stems (Ceballos and Calle 2010). This generates, unavoidably, certain phenotypic variation among plants in the same plot which may even increase the experimental error in evaluation trials. Cuttings are generally 20–30 cm long and have five to seven nodes. Cuttings can be planted horizontally, vertically or at a certain angle. If not planted horizontally, the cuttings are introduced into the ground at least half their length and taking into consideration the orientation of the stem (proximal end down and distal end up) as apical dominance occurs during sprouting. Adventitious roots develop from the underground portion of the cutting, either from the base of the nodal units or from the cut base of the stem. About 2 to 3 MAP, some roots start to swell and become storage roots. Above ground, buds sprout to produce leaves, and below the apical meristems, the stem starts elongating. Usually one or two buds will sprout to generate the respective stems. If cuttings are planted horizontally, many potential stems may emerge, and then some thinning may be advisable. Ideally one or two main stems should develop from each cutting. Planting density is typically 1 × 1 m for a 10,000 plants/ha density. Higher densities are acceptable (up to 20,000 pl/ha, depending on varieties and environmental conditions).
The multiplication rate through stem cuttings in cassava is low. Because of differences in plant architecture, there is large variation in the number of cuttings that a plant can produce. Plants from some genotypes can produce as many as 30 cuttings per plant, whereas in other genotypes plants may barely produce five cuttings. When hundreds or thousands of genotypes need to be screened in uniform conditions, a maximum of eight (at best ten) cuttings per plant can safely be assumed from each and every genotype. This low reproductive rate implies that several years are required until enough planting material is available for multilocation trials. Breeders are chronically short of planting material, which imposes many limitations on the evaluation process. For example, little progress has been made in identifying early bulking germplasm because it would require sacrificing some plants whose stems cannot be used as source of planting material (as they would have been harvested off season). Similarly, it may be desirable to store stems for longer periods of time (to select for capacity to sprout quickly and vigorously, even after long storage) but that would also expose losing some plants when cuttings fail to sprout. In turn, this would complicate the logistics of the following stages in the selection process. Although rapid multiplication schemes based on micro-cuttings or tissue culture approaches can be implemented, they are expensive and have not been routinely used.
Cassava is monoecious, i.e. with separate female and male flowers, occurring in the same inflorescence (raceme or panicle). Male flowers are more numerous and develop in the upper section of the inflorescence (Gonçalves Fukuda et al. 2002; Perera et al. 2012). Female flowers are fewer and are found in the proximal branches of the inflorescence (Fig. 5.1a). Anthesis of female flowers occurs about 10–14 days before that of male flowers (protogynia). Cassava naturally outcrosses (mostly by insects). Self-pollination can occur when male and female flowers on different branches of the same plant are open, or different plants of the same genotypes simultaneously produce male and female flowers.

(a) Illustration of an inflorescence with female flowers in the basal section already open and male flowers in the top still not open. (b) Erect plant architecture. (c) Defoliated plant illustrating the four consecutive flowering (and branching) events. (d) Highly branching plant architecture of a clone planted by farmers in Central Ghana
Inflorescences always develop at the apex of the developing stem. Sprouting of the buds below the inflorescence allows further growth of the plant. Therefore, the plant first flowers and then develops branches (Gonçalves Fukuda et al. 2002). Every flowering event, therefore, results in branching (Fig. 5.1c). Some genotypes flower early and several times (starting from 3 MAP and up to five times) during a growth cycle and others flower little or late (e.g. once at eight MAP). Erect, non-branching types are frequently preferred by farmers because this plant architecture facilitates cultural practices and results in good production of vegetative planting material, and its transport and storage are easier (Fig. 5.1b). Longer stems can withstand a longer storage period and may be a key trait to deal with climate change (Ceballos et al. 2012). In Africa, on the other hand, early branching clones are sometimes preferred by farmers (Fig. 5.1d); an advantage may be early canopy closure to help control weeds.
The general advantages of erect clones which do not branch or branch late in the season result in a dilemma for the breeder because the production of botanical seed from these genotypes is sparse and slow and, ultimately, more expensive. There are indeed many cassava clones with profuse, early and frequent flowering. These materials would facilitate greatly the production of segregating seed. However, as stated above, this flowering behaviour is closely related to an early (i.e. low to the ground) and frequent branching (Fig. 5.1c, d), which is generally undesirable. Since initiation, frequency and prolificacy of flowering are under genetic control, the progenies from early flowering types tend to inherit the trait and, along with it, the undesirable branching plant architecture. Perhaps one of the most important areas of research to accelerate and improve the impact of breeding, therefore, would be the development of a protocol for temporarily inducing flowering in cassava. Conventional breeding would benefit by reducing the costs and time currently required to make crosses and obtain seeds. Induction of flowering would also facilitate the development of inbred progenitors through successive self-pollinations. Accelerated and synchronized flowering in cassava crossing blocks would also be useful for the implementation of genomic selection. There are ongoing approaches to induce flowering through grafting, photoperiod modulation and the use of plant growth regulators.
The development and growth of cassava are rather simplistic; it does not mature from the phenological point of view. After planting the buds sprout to produce leaves and stems. As the leaf area index increases, excess photosynthates are produced by the source (canopy) and are then stored in the sink (roots). The only function of the storage root of cassava is as an energy reserve organ for the mother plant. As the conditions are conducive for photosynthesis (e.g. adequate light, water and temperature), the plant continues growing and partitioning photosynthates into the roots. The root does not reach physiological maturity at any given time. If environmental conditions become limiting for further growth (e.g. a dry season begins or temperatures fall below optimum), the plant ceases growing; starch accumulation will cease or become very slow. In most cassava-growing regions of the world, there will be a “dormant” period in which the plant stops accumulating energy in the root. It is usually at the end of that time when farmers harvest the crop. The roots will have optimum quality (particularly in relation to dry matter content – DMC), and harvested stems will have to be stored for a short period of time before growing conditions improve (e.g. rains arrive). If the plants are left in the ground, growth will restart using the energy stored in the roots. DMC in the root, therefore, falls drastically if plants are allowed to restart growth before they are harvested. Some genotypes quickly recover DMC but not others. The capacity of recovering DMC after a second growth stage is an important characteristic in those systems when harvest is extended to take place during a second year of growth as is done in Southern Brazil and Paraguay.
The fact that cassava roots can be harvested at any time or age of the plant (provided that they are old enough to have had adequate time to accumulate starch) is a great advantage for cassava and explains the remarkable food security role of the crop. Farmers can leave the crop in the field until needed. Cassava does not have a phenological stage during its growth when it is particularly vulnerable to environmental constraints (such as water stress during flowering of cereals), thus its recognized drought tolerance.
3 Genetics
Cassava is frequently considered an historical polyploid species based on the basic chromosome number of species in the Euphorbiaceae family (Westwood 1990). However, cytogenetic analyses during meiosis consistently have found the presence of 18 bivalents which are small and similar in size (Hahn et al. 1990; Umanah and Hartmann 1973; Wang et al. 2011). In some cases, occurrence of univalents/trivalents and late bivalent pairing has been reported. Cassava is therefore a functional diploid (2n = 2x = 36) (Jennings 1963; Westwood 1990; De Carvalho and Guerra-M 2002; Nassar and Ortiz 2008). Magoon and co-workers suggested in 1969 that certain portions of the genome may be duplicated, and, therefore, cassava may be a segmental allotetraploid.
Very few Mendelian genetic studies have been conducted and reported in cassava. Gonçalves Fukuda and co-workers summarized in 2002 a few cases in which inheritance of different traits had been reported. Narrow-lobed leaves are a dominant trait controlled by a single gene, and darker colour of the external root peel is dominant over the light coloration (Graner 1942; Jos and Hrishi 1976). Male sterility is a recessive monogenic trait (Jos and Bai 1981; Jos and Nair 1984). Pale green in the stem collenchyma is dominant over dark green and controlled by a single gene; yellow root parenchyma is partially dominant over white and controlled by two genes (Hershey and Ocampo-N 1989; Iglesias et al. 1997; Morillo-C et al. 2012). The stem in zigzag is a recessive trait, and it has been used as a marker gene in cross identification. However, since some nutrient deficiencies can also cause the zigzag trait, the genetic component may not always be clear. Red is dominant to green in leaf nerves and has also been used to distinguish clones derived from crosses from those of self-pollinations (Kawano et al. 1978). Unfortunately, none of these traits (except parenchyma pigmentation which is linked to higher carotenoids content) have economic relevance. More recently, the inheritance due to a single recessive mutation was confirmed for waxy (amylose-free) starch (Aiemnaka et al. 2012). Resistance to cassava mosaic disease-CMD (a devastating disease present in Africa, India and Sri Lanka) seems to be controlled in some cases by a single dominant gene (Rabbi et al. 2014a).
Quantitative genetic information , on the other hand, has been more widely studied. The vegetative reproduction of cassava allows the quantification of within-family genetic variation, which, in turn, allows a test for epistasis in diallel crosses (Cach et al. 2005; Pérez et al. 2005a, b). These diallel studies covered progenitors adapted to three different agroecological regions of Colombia. The most relevant results from these studies have been summarized in Table 5.1 for two key traits (fresh root yield, FRY, and DMC). Two features are worth highlighting. The use of heterozygous progenitors results in large within-family genetic variation. Nonadditive genetic effects (dominance and epistasis) are clearly relevant for the expression of these two traits but particularly for FRY . This type of information has been useful in the prediction that genomic selection would not be effective for traits such as FRY.
Additional quantitative genetic information has been produced in Africa and Asia. These studies can be grouped as diallel analyses and North Carolina II designs (Bueno 1991; Chipeta et al. 2013; Easwari Amma et al. 1995; Kamau et al. 2010; Lokko et al. 2006a; Njenga et al. 2014; Owolade et al. 2006; Parkes et al. 2013; Were et al. 2012; Zacarias and Labuschagne 2010). In general, all these papers support the information presented in Table 5.1 regarding the importance of nonadditive genetic effects for FRY.
Cassava, as most outcrossing species, shows considerable inbreeding depression for FRY (Gonçalves Fukuda et al. 2002; Kawuki et al. 2011b; Rojas-C et al. 2009). When inbreeding depression is important in a crop, it is often the case that heterosis will also be prevalent. The relative importance of nonadditive genetic effects for FRY suggested by diallel and North Carolina II Designs agree with the inbreeding depression observed for this trait by several studies.
4 Germplasm: Intra- and Interspecific
The following section draws heavily from an extensive survey and review of cassava genetic resources supported by the Crop Trust and published as part of their series on Crop Conservation Strategies (Hershey 2008).
One of the challenges in Manihot taxonomy is that interspecific hybridization between cultivated cassava and wild relatives, and among some of the wild relatives, occurs readily. It is feasible therefore that many wild species may have experienced important introgression of cultivated cassava alleles and vice versa. This has been suggested, for example, in the case of M. glaziovii (Nassar 2000; Bredeson et al. 2016). Ideally, wild species should be collected in isolated areas where no cassava is cultivated. This is critically important. The first source of resistance to cassava mosaic disease (CMD ), a devastating virus disease in Africa, was first reported in M. glaziovii by the East African Agriculture and Forestry Research Organization, based at Amani (then Tanganyika). That program then developed interspecific hybrids to generate the first cassava clones with resistance to CMD, sources which are still widely used in breeding in both East and West Africa.
As for any crop, the future potential of cassava to contribute to the sustainable benefit of humankind will rely fundamentally on safe long-term conservation of broad-based genetic resources and their use in effective breeding programs. These resources are basically the landrace varieties that evolved for centuries under farmer and natural selection and about 100 wild species of the genus Manihot. The genus is native to the Americas, and most of the genetic diversification has occurred here. Traders first introduced cassava into Africa and later into Asia. Both continents have become important secondary centres of genetic diversity, especially Africa.
Cassava is a vegetatively propagated crop, while all the wild Manihot species are seed-propagated in their natural environments. In order to preserve the genetic integrity of a landrace, cassava must be conserved in vegetative form. The most common forms of conservation are as field-grown plants or as plantlets started from meristem tips, cultured on sterile artificial media, under light, temperature and media conditions that induce slow growth. For either field or in vitro conservation, expensive periodic regeneration is required, at a much higher frequency (typically every 12–24 months) than is typical for seed conservation.
Many cassava-growing countries have established a gene bank of local landraces, managed by government organizations. Most of these collections have been established since the 1970s, but some much more recently. Up until the 1970s, and through the 1980s, relatively few landraces had been lost due to broad-scale replacement by new, bred varieties. However, the risks are now much higher for genetic erosion of landraces, due to multiple factors, including success of new varieties, replacement of cassava by other crops in some regions (e.g. southern cone of South America), crop intensification and the associated trends towards less on-farm variety diversity and, possibly, climate change.
Most countries note collection gaps (less so for Asia), due to lack of funding, losses of diversity due to natural disasters and social conflict, difficult access to areas for collecting and inadequate collecting techniques of the past. Nearly all programs rely primarily on field-grown plants but may have part of their collection in vitro as well. In vitro collections have had a mixed success except in the few institutions where they are well funded for the long-term such as Embrapa, Brazil; CIAT, Colombia; and IITA, Nigeria. Globally, only about one-quarter of accessions held by national programs appear to be conserved in vitro. Two international centres (CIAT and IITA) maintain regional collections for the Americas and Asia (CIAT) and for Africa (IITA). There are very few national gene banks that have the capacity to carry out safe international exchange in situations where viruses, of quarantine significance, are present. Most international exchange is facilitated via the international centres.
About two-thirds of cassava is currently grown in Africa, but probably well over half the landraces occur in the Americas. This is to be expected in view of origin of the species in the Americas. A study in 2008 (Table 5.2) estimates that some 27,000 distinct landraces of cassava are conserved in situ and about 10,000 maintained in gene banks. Hershey (2008) proposed that a total of about 15,000 landrace varieties should be conserved ex situ in order to represent the complete genetic diversity of the species. However, these estimates are based on very tentative results from genetic diversity studies, and much more information from molecular diversity studies will be needed to develop more precise information.
Currently CIAT curates the gene bank with the largest number of cassava landrace varieties (about 5500 accessions), sourced from throughout the crop’s origin in the Americas and from Asia. IITA holds an extensive collection from West Africa and is developing plans to incorporate a larger representation from East Africa. A conservation strategy should consider security, cost and efficiency in its design (Epperson et al. 1997). Security is a function of both the number of replications of a gene bank (in different sites or in different forms) and the management level of each. Field gene banks are the least secure, followed by in vitro slow growth and, finally, cryo-conservation. Although cryo-conservation has been researched for more than two decades for cassava, and with relatively good success, there are as yet no cassava gene banks which rely on it as a main form of conservation.
Hershey (2008), based on survey and workshop results, proposed a comprehensive conservation strategy consisting of the following elements:
-
Collecting in priority areas is carried out to fill gaps, with the aid of genetic diversity studies and GIS.
-
National program gene banks and international centre gene banks are systematically compared for matching and non-matching accessions, based on passport, morphological and molecular information. This would evolve into a common cassava registry at a global level.
-
CIAT and IITA duplicate all the landraces of national program collections, in their respective regions of responsibility (CIAT: Americas and Asia; IITA: Africa). Currently they appear to maintain about 50–60% of these accessions.
-
National programs commit to at least one working gene bank that serves the purposes both of conservation at a moderate level of security and evaluation.
-
CIAT and IITA maintain at least two forms of each accession. Currently this may be an in vitro active gene bank plus a black box duplicate kept in another centre. In the future, cryopreserved accessions will be either the main or the backup gene bank.
-
CIAT and IITA commit to making the material they maintain available to national program gene banks, when requested.
-
CIAT and IITA commit to meeting the demands and phytosanitary requirements for international exchange of cassava landrace varieties under terms of the International Treaty. Along with this, it is urgent to develop protocols for the safe movement of vegetative germplasm between the Americas and Africa.
-
There is a mechanism developed for periodic interaction among stakeholders. Most notably this will be between the international centres and the national programs. Each will have a formal responsibility to periodically inform the other of the status of collections.
Duplicate identification, further improvements for in vitro slow growth techniques, improving cryo-conservation, and flower induction for seed conservation are all research areas outside the funding stream for routine conservation, but which will contribute to greater conservation and use efficiencies in the long term. Cryo-conservation is clearly an option for effective, inexpensive, secure long-term conservation, but work remains to be done on achieving an adequate recovery level for about one-third of accessions (based on results from CIAT’s core collection). Research should continue on improving recovery of these recalcitrant types before committing to large-scale cryo-conservation of any gene bank.
As a future alternative to vegetative cassava gene banks , the seed from self-pollinated accessions could be a less expensive and more efficient conservation method and would be equally or more effective for breeding programs. Since many cassava accessions do not readily flower, there is a need for research on the induction of flowering in order to have a broadly applicable strategy for seed conservation. Long-term, a conservation strategy that consists of a combination of cryo-conserved meristem shoots and seed maintained in conventional cold storage might be envisioned. This would combine the advantages of both seed and vegetative conservation in a low-cost, secure system.
The wild Manihot species present a situation that is simpler in some aspects but more complex in others, compared to cassava. It is simpler in that only a handful of institutions are involved in conservation – mainly Embrapa and the University of Brasilia in Brazil, CIAT and IITA. It is more complex in that:
-
The taxonomy of species is still poorly defined.
-
The highest concentration of species is native to threatened habitats. This is especially true in the Campo Cerrado of South-Central Brazil, where the expansion of agriculture and urbanization are rapidly encroaching on the wild species habitats.
-
A secondary centre of diversity, with a distinct set of species, exists in Mesoamerica. Here, and especially in Mexico, cassava is a relatively unimportant crop, and it is difficult for these governments to justify investment in Manihot conservation in terms of value addition to the crop per se.
-
Fewer than half the species are conserved in vitro, and very few are protected in national or regional reserves, in their native habitat.
-
Wild species conservation presents many challenges, especially with regard to regeneration. Progress is being made both in seed and in vitro propagation, but much remains to be done.
-
The value of the wild species is continually becoming more evident as new characters are identified with potential for transfer to cassava, and the techniques for efficient transfer and selection of specific genes are developed.
-
Genes from cultivated cassava may have been introgressed into wild relatives.
Cassava is unusual among major crops in that there is relatively little differentiation between landrace varieties and “modern”-bred varieties. This is due in large part to the late and relatively low investment that cassava breeding has had and to the fact that the large majority of clients for new varieties are the small-holder farmers who grow the crop with minimal inputs for traditional markets (especially in Africa). This relatively low differentiation between landraces and bred materials has allowed breeders to continue to make wide use of gene banks to introduce new traits into breeding populations and to derive superior individuals without the need for extensive further crossing to eliminate unfavourable genes. However, this is changing, and breeders will ultimately broaden the gap between what is available in gene banks or in situ, compared to new varieties for modern management and markets.
Cassava gene banks have been moderately to well evaluated for basic agronomic traits like plant architecture, yield in multiple agroecosystems, DMC, cyanogenic glucosides (CG ) and reaction to a range of pests and diseases. In addition, significant numbers of accessions have been evaluated for nutrient use efficiency, multiple root quality traits such as amylose/amylopectin ratio, sugar, and carotenoid content and tolerance to PPD. Systematic efforts to screen for useful recessive traits, such as amylose-free starch (Morante et al. 2016), need to be conducted. As we face new challenges such as climate change and the spread of pests and diseases, and new opportunities with new markets, novel genetic diversity will become ever more important, such as capacity to withstand long storage periods of the stems. From the standpoint of ease of use by breeders, the first step should be the full exploration of the M. esculenta gene pools. There is little advantage – and many disadvantages – to the extraction of genes from wild species if the same genes are available in cassava landraces. For example, the use of M. glaziovii as a source of resistance to cassava mosaic disease required decades of breeding in order to restore agronomic performance in genotypes carrying the CMD resistance. Initially it was believed that there were no sources of resistance within the cultivated species, but more recently it has been shown that variation for resistance does exist. Molecular markers will ultimately allow more efficient identification and extraction of genes from wild species (Duitama 2017). There is an urgent need to collect, conserve and evaluate these species as a resource for future breeding.
5 Crossing Approaches
Controlled pollinations in cassava result in the production of full-sib families (Fig. 5.2a). Several publications illustrate crossing techniques (Kawano 1980; Byrne 1984; Jennings and Iglesias 2002; Gonçalves Fukuda et al. 2002). There is no evidence of incompatibility, but there is variation in the success of seed set among different crosses (Njoku et al. 2015a). A maximum of three seeds per pollination can be obtained, although averages in controlled pollinations are considerably lower (ranging from one to two seeds). Making controlled pollinations in cassava is not difficult. Male flowers, ready for anthesis, are collected in the morning hours before they open. At the same time, on the day of anthesis, female flowers are covered (before stigmas are exposed). Field workers can easily recognize those flowers that will open each day, and they do these operations efficiently. Collecting or covering flowers before anthesis prevents contamination with undesired pollen.

(a) Illustration of female (top left) and male (mid-left) flowers and procedure used in controlled pollinations (bottom left) which take place in the afternoon. Female flowers are covered with bags before they open and the bags are removed 2–3 days after pollination (centre top). Bags are placed again over near-mature fruits to collect seeds after dehiscence. Male flowers are collected in the morning and kept in plastic containers until needed (centre bottom). (b) Example of “asparagus” phenotype (top) and aerial view of a trial where it was compared with cassava genotypes with normal phenotype
Open pollinations can also be used as source of botanical seed. Polycross nurseries are planted following a special field design to favour pollinations of different genotypes (Wright 1965). At CIAT all the seeds collected from a given female progenitor in a polycross define a unique half-sib family. The female parent is known, since the seeds are collected from identified genotypes. The source of pollen that generated each seed is unknown, but a set of potential male progenitors is restricted to the genotypes that make up the polycross nursery each year.
The major bottleneck in the production of seed in cassava breeding programs is the scarcity of flowering and/or problems of synchronizing the flowering of particular pairs of genotypes that the breeder wants to cross. Efforts to develop a protocol for pollen conservation several decades ago were unfortunately fruitless (Orrego and Hershey 1984). The preference in many regions of the world for erect plant architecture aggravates matters further because this phenotype implies late flowering and few flowering events that the breeder can exploit. Certain environmental conditions may favour flowering. Longer photoperiods and cooler temperatures have been reported to stimulate flowering (De Bruijn 1977; Keating 1982). Lack of balance in the number of crosses representing each progenitor has implications for breeding. Indirect information suggests that at least 200–300 genotypes would be required to properly assess the breeding value of a given progenitor (Ceballos et al. 2016a). This figure is often difficult to attain and breeders may have to wait for 2–3 years until enough crosses from a given set of progenitors have been made and the resulting seed harvested.
The late flowering habit of erect plant types implies that breeders typically need 18–24 months to produce an acceptable number of seeds (e.g. at least 20–30) from any planned cross. This time frame was not a problem for ordinary breeding projects where the main objective was developing high-yielding materials adapted to a particular agroecological zone. If seed from a given cross could not be evaluated in one year, it would be included in the following year. However, as breeding projects sought to develop clones with special attributes (e.g. high carotenoids or special starches), the slow rate in the production of botanical seed became a logistic problem as crosses were usually concentrated during 12–18 months. Special breeding projects are usually supported by time-constrained research grants that impose a restriction in the time allocated to make crosses. The Next-Generation Cassava Breeding Project (www.nextgencassava.org) is evaluating the potential of genomic selection in cassava (de Oliveira et al. 2012; Ly et al. 2013; Rabbi et al. 2014b; Wolfe et al. 2016a, b). It recognized, early on, that the induction of flowering was a key requirement because it would allow the achievement of a more balanced number of progenies from each progenitor and shorten the length of each recurrent selection cycle.
Another example of difficulties in making crosses in cassava can be given with the so-called “asparagus cassava ” (Fig. 5.2b). This particular phenotype is characterized by leaves without petiole and absence of flowering within the first 8–10 MAP (no or very late branching). This very particular phenotype could be a new plant type as it is well adapted for high planting densities (e.g. 40,000 pl ha−1 instead of the normal 10,000). Preliminary results have demonstrated that “asparagus cassava” responds better to high densities than ordinary phenotypes (CIAT, unpublished results). However, breeding this type of cassava would be nearly impossible due to the current difficulties producing segregating seeds.
Because of the reasons described above, the induction of flowering in cassava has become an important research objective. Grafting of stems from non-flowering genotypes into a rootstock from an early and frequently flowering genotype has induced flowering in some genotypes, but not in others (Ceballos et al. 2017). There are ongoing efforts to induce earlier flowering and stimulate number of flowers produced and seed set through modulation of photoperiod or application of plant growth regulators. These efforts have been successful in some genotypes but not in others. Preliminary results indicate that the application of certain growth regulators has been successful for inducing flowering in the “asparagus cassava ” (CIAT, unpublished data). Genetic transformation modulating the FT locus has also been successfully achieved (Adeyemo et al. 2008).
6 Breeding Schemes
Several reviews on cassava breeding have been made over the years (Byrne 1984; CIAT 1991; Gonçalves Fukuda et al. 2002; Jennings and Hershey 1985; Jennings and Iglesias 2002; Ceballos et al. 2004, 2007a, 2010, 2012; Kawano and Cock 2005; Kawuki et al. 2011a). These reviews provide new information regarding discoveries of new sources for relevant traits, consolidated information regarding the relative importance of genotype , environment and their interaction for productivity and, more recently, on the potential of molecular markers. However, basically the breeding scheme has remained unchanged since the inception of modern cassava breeding in the late 1960s. Below is a brief description of the scheme used at CIAT. All programs apply some variation of a scheme which begins with selection of parents; crossing to produce a segregating population; seedling nurseries where each individual is genetically distinct; then a series of clonal trials of increasing plot size; number of replications and number of testing sites, ending in regional and on-farm trials; and finally official release. Typically, the time frame from selection of parents to release is at least 10 years and often longer. Under highly accelerated schemes, release could take place in as few as 6 or 7 years.
Figure 5.3 illustrates a general scheme of the different trials used in cassava breeding. In the case of cassava, botanical seeds from a cross between two heterozygous parents are normally referred to as the F1 generation, in contrast to common use of the term to refer to progeny from two homozygous parents. Botanical seeds from full- or half-sib families are germinated and the resulting seedlings grown in a screenhouse for 1–2 months. Some programs, however, germinate the seeds directly in the field, a possibility especially under high soil temperature conditions. Vigorous and healthy seedlings are then transplanted to the field and grown for 10–11 months. The size of seedling nurseries varies considerably depending on seed availability, regions, objectives and program resources to manage nurseries. If selection can be effectively made on a one-plant plot, i.e. for traits with high heritability, then the F1 nurseries tend to be relatively large (>15,000 plants). Strong selection pressure is used at the seedling plant stage in few specific cases where the breeder aims at selecting for a high-heritability trait: resistance to CMD (Rabbi et al. 2014a; Jennings and Iglesias 2002), bio-fortification for enhanced carotenoids content (Belalcazar et al. 2016; Ceballos et al. 2013), waxy starch (Aiemnaka et al. 2012) or white vs. brown root surface.

Illustration of the different stages of evaluation used at CIAT in the selection of clones eventually released for commercial growth by farmers. The number of plants per plot, replications and locations for each stage is indicated at the bottom
When the goal is to generate clones with good agronomic performance and high productivity, in the absence of a limiting factor such as CMD, selection based on a single plant plot is not reliable. In these cases, therefore, selection pressure cannot be high at the F1 stage, and the nursery size tends to be smaller, such that the large majority of genotypes can be cloned for more complete evaluation in larger plots. Seedling nurseries at CIAT for a particular target environment are typically <10,000 plants. Genotypes with undesirable phenotypes (e.g. poor vigour, susceptibility to thrips, chlorosis, very low or high harvest index , etc.) are also eliminated by default (Fig. 5.4a). The seedling plants (F1 in Figs. 5.3 and 5.4a) are used as source of planting material for the next stage in the selection process and must, therefore, provide six to eight good quality cuttings. At CIAT, the seedling nursery is planted at Palmira Experimental Station, which offers nearly ideal growing conditions (including irrigation) to guarantee excellent nutritional and sanitary conditions of the resulting planting material. The next stage in the selection process is the single-row trial (SRT ) which is planted in the target environment. In Colombia, CIAT has four main targets: subhumid, acid-soil savannas, mid-altitude valleys and the highland environments.

Illustration of different types of trials in the cassava breeding scheme. (a) Seedling nursery (F1) in which adequate plant architecture, vigour, health and harvest index are key selection criteria; (b) single-row trials (SRT) are usually very large (up to 2 ha); (c) preliminary yield trials (PYT) follow a special design that leaves one empty row between plots to reduce plant competition among different genotypes. Notice the difference in plant height in two neighbouring clones; (d) plots in advanced yield trials (AYT) and uniform yield trials (UYT) are larger, and only the six or nine central plants are harvested, to eliminate effects of inter-genotypic competition. The surrounding plants in the plot periphery are left in the field and used as source of planting material
In SRT each genotype is planted in a single row with six to eight plants per row (the number may change from one year to another, but it is uniform for each trial). These are large experiments with 1500–2500 genotypes (1–2 ha) and therefore prone to large experimental errors (Fig. 5.4b). Target environments and evaluation plots lack uniformity which reduces the precision of evaluations and reliability of data. At CIAT, therefore, the SRTs are split in three to four blocks (strata), and selection is made within each stratum (Ceballos et al. 2007a). This strategy reduces the environmental variation among strata in the selection process (Gardner 1961). A selection index (SIN, described in next section) is used to facilitate selection of genotypes evaluated at SRT.
The following stage in the selection process is the preliminary yield trails (PYT ) in which each genotype is planted in three replications with ten-plant plots (two rows of five plants). The six to eight plants from SRT are used as source of planting material for the PYTs. Because of limitation in the amount of planting material available, a special design is often used for PYTs . In cassava there is still considerable variation in the farmer-preferred plant architecture, ranging from short and bushy to tall and erect (Fig. 5.1). To minimize inter-genotypic competition arising from differences in plant height and architecture, an empty row is left between neighbouring plots (Fig. 5.4c). Plant spacing is set to 0.8 × 0.8 m within plots and 1.6 m between plots, for a final plant density of 9600 pl ha−1. The number of genotypes evaluated in PYTs is still relatively large (200–500), and therefore they are split into three to five separate trials with 50–100 clones each. Selection is conducted within each trial but is not as stringent as for SRT. About 50–120 genotypes are selected for the following stage (advanced yield trials or AYT).
Plots in AYT have four (or five) rows with five plants each. The six (or nine) central plants are harvested and the resulting data used for selection. The surrounding plants in the periphery are left in the field and used as source of planting material when needed (Fig. 5.4d). Occasionally AYT may be planted for two consecutive years or at two or more locations. The same selection index is used throughout the different stages of the selection process, but as the number of genotypes is gradually reduced, more information is taken in later stages (e.g. boiling time, cooking quality, CG etc.).
The last stage in the breeding scheme is the uniform yield trials (UYT ) which are conducted in several locations and at least two consecutive years. About 20–25 experimental clones and 5–8 commercial checks/controls are involved in these trials, which have three replications and plots similar to those used in AYT. Genotypes reaching UYT are usually incorporated as progenitors in crossing blocks and the best two to three may be considered for release as official varieties.
There are many variations that can be introduced to this generalized scheme, to achieve specific goals for any given breeding program or to adapt to local conditions and resources. For example, CIAT uses a disease “hotspot” in the eastern plains region (Llanos Orientales) to screen all material that has reached intermediate selection stage. For example, at the same time a set of genotypes is evaluated in the PYT in the principal target environment of the Atlantic coast region (subhumid tropics), a set of the same genotypes is evaluated in a SRT in the Llanos for resistance to bacterial blight and super-elongation disease.
7 Traits
For many years, the main objective of cassava breeding programs was to produce high-yielding clones that were adapted to the target environments and tolerant to their main biotic and abiotic stresses. However, several traits, in addition to high and stable productivity, are required in order for clones to be adopted by farmers. Some of these additional characteristics are common across regions (e.g. capacity of stems to withstand long storage period or fast and vigorous sprouting of the cuttings). But others may be contrasting and region specific (e.g. erect plant architecture is preferred in many Asian and Latin American countries, whereas bushy types may be preferred in certain regions of Africa).
Depending on the target region, different biotic stresses have to be considered by breeders. Bacterial blight (CBB) is found widely across the three continents. Across Africa, in India and Sri Lanka, different strains of CMV are a major threat to cassava. Cassava brown streak virus (CBSV) originated in East Africa but is spreading westward. In Latin America, cassava frogskin disease (CFSD) (whose aetiology is not clearly established) is a problem that can be managed with simple cultural practices (e.g. confirm that roots are symptomless before collecting the stems as source of planting material an approach that is also useful for CBSV) but may also be a target for resistance breeding. Thrips can have a devastating effect on cassava. Fortunately, pubescence of newly emerging leaves at the shoot tips offers an excellent and stable source of resistance. Whiteflies can cause direct damage to cassava, but their most serious impact is transmitting viruses such as CMV and CBSV. An excellent source of resistance has been found in landraces from Ecuador and Peru (Bellotti and Arias 2001). Mites can have detrimental effects on cassava, particularly during the dry seasons. There is an interesting example of interaction between genotype and biological control in relation to mites. In Africa, the management of the cassava green mite problem is achieved through biological control by exotic predatory mites. However, cassava morphological traits can be improved to provide better shelter and ensure continuous survival of the natural enemies of the cassava green mite (Chalwe et al. 2015; Molo et al. 2016).
End uses of cassava drastically define the traits that breeders have to take into consideration. For starch, ethanol or dried chips for animal feeding the key traits will be DMC and FRY. White parenchyma is preferred by the starch industry, but the enhanced nutritional quality of yellow roots would be preferable for animal feeding. The great diversity of food uses of cassava was thoroughly described by Balagopalan (2002). Some regional and ethnic uses of cassava such as farinha and casabe (Amazon basin), kokonte (Ghana), gaplek and krupuk (Indonesia) or tapioca pearls (India) would also benefit from adequate DMC and FRY. However other ethnic uses require additional traits. Boiled cassava roots require low levels of CG, reduced boiling time and consumer-preferred texture. On the other hand, African products such as fufu and gari require proper poundability or mealiness. It is not clear which are the anatomical or biochemical characteristics that define cooking quality, mealiness or poundability, and there is ongoing research to elucidate these characteristics to facilitate the selection process made by breeders.
As breeding techniques and cassava utilization evolved, however, there has been a growing tendency to shift from breeding to develop general-purpose cultivars towards more specialized, market-oriented products. This new trend is a result of the confluence of different circumstances briefly described below.
The first modern cassava varieties started to be released in the early 1980s through early 1990s. These varieties fulfilled the basic requirements of adequate levels and stability of productivity, acceptable levels of DMC and, depending on the end uses, root quality traits. This early group of varieties includes, for example, the very successful variety KU50 released in Thailand in 1992 (FAO 2013; Fisher et al. 2014; Kawano 2003; Kawano and Cock 2005; Kawano et al. 1998). A second wave of improved clones was released during the 2000s, and a third wave is being released in the 2010s. The varieties released were very successful in increasing productivity (particularly in SE Asia where it basically doubled in the last 30 years). However, genetic gains in the second and third waves of new varieties seem to have plateaued (Ceballos and Hershey 2016). The breeders’ hopes of producing new clones with higher productivity than that of already-released varieties face increasing challenges. It has been postulated that the difficulties of increasing productivity in cassava arise from the heterozygous nature of the progenitors used and the strong influence of nonadditive genetic effects already described in Table 5.1 (Ceballos et al. 2015, 2016a). These problems require change(s) in the current breeding methods, which will be discussed later in this chapter.
At the same time that breeders recognized the need for alternative breeding approaches for increasing productivity, their attention has also shifted towards other traits with commercial relevance and with higher heritabilities. This is the case of breeding for increased carotenoids content (Ceballos et al. 2013; Esuma et al. 2016; Morillo-C et al. 2012; Njenga et al. 2014; Njoku et al. 2015b; Ssemakula and Dixon 2007) or cassava with special starch functional properties (Carvalho et al. 2004; Ceballos et al. 2007b, 2008; Morante et al. 2016).
The identification of traits with high commercial value (such as starch mutants) or the development of clones with desirable nutritional traits (e.g. high carotenoids or low levels of CG) highlight the importance of a thorough and complete screening of gene banks as indicated in the section of germplasm resources. The development of reverse genetic molecular approaches (such as Eco-TILLING) or similar technologies (such as pooled targeted resequencing of DNA to detect rare SNPs in specific genes) would make the identification of useful traits much easier (Duitama et al. 2017). As we expand our understanding of the specific needs for the ever-widening uses of cassava, these molecular tools offer a very appealing alternative for the identification of useful (recessive) traits.
Cassava roots spoil quickly due to PPD. The short shelf life of roots imposes many limitations to post-harvest handing, transport and processing of cassava roots (Beeching et al. 1993; Reilly et al. 2007; Vlaar et al. 2007). Sources of tolerance to PPD have been reported (Morante et al. 2010; Moyib et al. 2015), but changes during root storage, including starch losses, limit the shelf life after harvest to less than 2 weeks (Sánchez et al. 2013). Little progress can be achieved through conventional breeding to reduce the negative impact of PPD until an adequate protocol for proper screening is developed. The experimental errors associated with PPD are huge and the effect of genotype-by-environment interaction unacceptably large (CIAT unpublished data).
Increase in levels of atmospheric CO2 is one of the most certain expectations of climate change and the environmental conditions for the next few decades. It is expected that increased CO2 will have a positive effect on cassava productivity by making photosynthesis (specifically CO2 fixation) more efficient. This is an interesting situation because it would result in reducing the physiological advantages that C4 crops such as maize, sugar cane and sorghum have over C3 plants such as cassava, wheat or soybean (Long et al. 2004, 2006). Free-Air CO2 Enrichment (FACE) methods allow field evaluation of crops under elevated CO2 concentrations that simulate the predicted levels for the decades to come (El-Sharkawy 2009). Modelling and FACE results could be very useful guides in the molecular optimization of the photosynthetic apparatus to maximize carbon gains without increasing crop inputs (Zhu et al. 2007, 2010). Preliminary results indicate that increases in productivity under elevated CO2 concentrations failed to meet theoretical expectations. If this is confirmed, it can be hypothesized that some bottlenecks in the complex metabolic processes following CO2 fixation during photosynthesis prevent the theoretical expectations. Overcoming these potential bottlenecks may allow breeding to maximize the enhanced productivity that elevated CO2 concentrations will have on cassava in years to come.
8 Field Trialling and Selection Approaches
The experimental design used for the different stages of breeding trials is basically a randomized block. Excel spreadsheets satisfy the needs and allow for the use of tablets or small portable computers for direct data uploading in the field. Statistical analyses (e.g. ANOVA) are not used routinely as statistical significances are not as relevant as properly ranking the different genotypes according to their performance – although error variances do give a good indication of what reliance one can place on the trial results in general. A method for the adjustment for missing plants has been developed (Pérez et al. 2010). However, correcting for missing plants may have the negative effect of rewarding genotypes with weak sprouting capacity after a normal storage period of the stems. A critical concept in the implementation of evaluation trials is to stratify them when they are large and the field is variable (Gardner 1961). Alternatively, large trials can be split into few smaller ones.
A few traits are broadly accepted across breeding programs as common key goals for improvement: high FRY, high and stable DMC, suitable plant architecture, resistance to locally important pests and diseases and harvest index (HIN). At CIAT, in addition to individual ratings, breeders integrate plant architecture and resistance to biotic/abiotic stresses into a single score indicating overall desirability of the above-ground plant appearance (plant type score or PTS) where 1 is very good and 5 is very poor. This is essentially a subjective answer to the question: “How much do I like the overall appearance of this genotype, based on the above-ground parts of the plant?” It is a sort of subjective ideotype, based on the breeder’s experience and skill, and may be included among the more objective “select” or “reject” criteria.
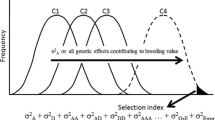
Because of the low heritability of FRY in early stages of selection, cassava breeders for many years have applied indirect selection for yield by using correlated traits with higher heritabilities, such as HIN (Kawano et al. 1998). Since 2000, CIAT has used a selection index (SIN) that integrates four high-priority variables, assigning them best judgement weight (in italics in the formula below). These weights have been established by the breeder’s experience (Ceballos et al. 2012) and can be modified with time:
In the case of PTS, the desired target is a lower score. Therefore, a negative sign is assigned to the respective term in the SIN equation. The variables used in SIN are measured in units that differ drastically in magnitude. To overcome this problem, standardized values are used (x i – X/σ, where x i is the individual observation, X is the average across genotypes and σ is the standard deviation) in the estimation of SIN . About 15% of clones evaluated in SRT are selected. Other traits, such as cooking quality, may be incorporated in the selection process but in later stages (AYT or UYT). Selection pressure is gradually relaxed through the scheme (e.g. 30–40% of clones evaluated in AYT may be selected for evaluation in UYT).
The selection process and criteria described above are ideally suited for a goal of high productivity. However, as stated earlier, other traits may be required for varietal adoption, particularly where cassava plays an important role in food security. Participatory breeding approaches (Gonçalvez Fukuda and Saad 2001; Gonçalves Fukuda et al. 2002; Grüneberg et al. 2009; Kamau et al. 2011; Manu-Aduening et al. 2006) are ideally suited for these conditions. Participatory breeding allows for much broader selection criteria, e.g. taking into account traits such as “maturity” period, suitability for intercropping, leaf production, taste, bitterness, processing amenability and cooking quality and even some traits that may just have a role as morphological markers such as petiole or shoot colour, leaf lobule shape or branching characteristics (Benesi et al. 2010). Participatory breeding emerged as a response to failures observed in some breeding efforts that limited the benefits of the green revolution in many developing countries for different crops. Many of the participatory breeding principles were initially developed in cassava (Ashby et al. 1987; Prudencio et al. 1992). Because of the size of the early trials (SRT and PYT), participation by farmers in the selection process may start in AYT or UYT.
Participatory approaches are based on the farmers’ ability to select what is best for his/her conditions, a process which proved its power most significantly through the domestication of crops. However, it must also take advantage of the scientific knowledge generated in the past century. A major issue that is particularly acute in participatory schemes relates to the negative impact of genotype-by-environment effects (Grüneberg et al. 2009). Local selection of germplasm may fail to take advantage of selection based on multilocation evaluations where stability of performance is a key objective. If possible, the same set of genotypes should be grown in different environments by farmers that have common interests. Farmers may select the materials of their preference, but breeders can then select for the best across-environment genotypes which should offer a more stable performance.
A critical decision that breeders constantly take and debate about is the progenitors to be included in the crossing blocks. More often than not, progenitors used in cassava breeding are just clones with outstanding performance or carrying a desirable trait (e.g. resistance to CMD or CBB, amylose-free starch or high carotenoids, etc.). Alternatively, the use of breeding value or general combining ability (Falconer 1981) as a criterion for choosing progenitors in cassava breeding has been proposed (Ceballos et al. 2004). This initial idea was renewed (in a more sophisticated way) with the implementation of genomic selection (de Oliveira et al. 2012; Rabbi et al. 2014a, b). The usefulness of selection of progenitors based on breeding values is reduced by their heterozygous nature and the resulting within-family genetic variation (Ceballos et al. 2016a). Additionally, at least for fresh root yield, there is a strong influence of nonadditive genetic effects, further reducing the predictive value of general combining ability (Table 5.1, Ceballos et al. 2015, 2016a).
Another important factor affecting the selection process is the relationship between different characteristics that can be positively or negatively correlated. For many years, for example, selection in early stages such as SRT was based preferably on HIN rather than FRY (Kawano 2003). The rationale was that these two variables are closely associated, but HIN has higher heritability than FRY (at least in unreplicated trials). An analysis of the selection process after 14 years of continuous evaluations in the subhumid environment of Colombia was recently published (Joaqui et al. 2016). In this study, the benefits of using HIN as an indirect selection criterion for enhanced productivity in SRT were questioned. HIN will still be a key criterion for selection at the seedling plant stage (F1). Another interesting observation was that the relationship between FRY and DMC was weakly but positively associated in SRT (r = 0.21) a result similar to the one reported by Kawano et al. in 1987. However, that relationship gradually becomes negative in successive stages of the selection process. In UYT the correlation was r = −0.42. This clear and consistent trend suggests that cassava genotypes attain high dry matter productivity by maximizing DMC or FRY, or else through a compromise for “acceptable” levels in both variables simultaneously. But it is difficult for a given genotype to express simultaneously maximum levels of DMC and FRY. The nature of the association between DMC and FRY cannot be properly stated without a clarification in which stage of the selection process this association is being considered. Similar conclusions can be drawn from Kawano et al. (1998).
New technologies are likely to have a positive impact on cassava breeding, and one of these is high-throughput phenotyping. The way that near infrared spectroscopy (NIR) benefited breeding for high carotenoid content has already been reported (Belalcazar et al. 2016; Davrieux et al. 2016; Sánchez et al. 2014). The use of drones has become a standard strategy to analyse canopy size and health. However, little progress has been made for monitoring root growth in cassava until now. The use of ground penetrating radar offers promising results that would allow non-destructive monitoring of root growth through the growing season (Delgado et al. 2017). Early bulking has been among the traits sought after by cassava breeders, particularly in Africa (Kamau et al. 2011; Okechukwu and Dixon 2009; Olasanmi et al. 2014; Tumuhimbise et al. 2014). But the resources required to managing multiple harvests for many genotypes, and chronic lack of planting material during the selection process, prevent breeders from sacrificing some plants to assess FRY before the usual harvesting time (11–12 MAP). This new technology offers promising advantages not only for selection of genotypes but also adequate cultural practices, for example, in the type and timing of fertilizers.
Cassava production in many regions of the world is seasonal. It is very common to have a large peak of availability of roots for periods of only 3–6 months. This is a major problem for large processing facilities that can operate only a few months of the year. An alternative solution to this problem is to delay harvest for a few months. This is how the cassava starch sector operates in Southern Brazil and Paraguay. Cassava is planted in early spring (e.g. August) and harvest can be split in two batches. Farmers may harvest part of the field early the following year in April–June and leave the rest for a delayed harvest in November–February. This approach benefits not only processors that can have their facilities operational for most of the year but also for farmers. Delayed harvests imply a sharp increase in the productivity (almost doubled, without a major increase in the costs). A key requirement for this approach, however, is that DMC at harvest time is at an optimum. In the system described above, DMC drops when plants reinitiate growth in August. This is a well-known phenomenon related to starch hydrolysis in the root (van Oirschot et al. 2000). When a similar strategy was attempted in tropical regions, however, DMC in available clones did not recover after 1 or 2 months after re-initiation of growth. There are ongoing efforts to select for a rapid recovery of DMC in delayed harvests. However, to do so, special trial arrangements are required: in the same locations, plots need to be harvested at the standard age (10–11 MAP) and then in delayed harvests (15–16 and 18–19 MAP).
9 Tissue Culture and Clonal Production
Tissue culture plays several fundamental roles related indirectly or directly to cassava breeding. The applications of tissue culture techniques are similar in cassava to those in other vegetatively propagated crops. Tissue culture has been used in cassava for rapid multiplication of clean planting material (Wasswa et al. 2010; Wongtiem et al. 2011), conservation/exchange of germplasm (Angel et al. 1996; Escobar et al. 1997; Roca 1984), embryo rescue approaches in interspecific crosses (Akinbo et al. 2010; Biggs et al. 1986; Fregene et al. 1999) and as a required system for genetic transformation (Bull et al. 2009; Ibrahim et al. 2008; Liu et al. 2011; Mongomake et al. 2015; Raemakers et al. 2001; Taylor et al. 2001, 2012). These are just representative reports that can be found in the literature for this broad range of applications of tissue culture. The conservation and exchange of germplasm have direct impact on breeding activities. Rapid multiplication and cleaning of planting material from diseases (particularly viruses) are important for the spreading and fast adoption of new varieties.
Tissue culture systems have been developed and used in cassava since the 1970s and are quite refined for routine use, such as for slow growth plantlet culture in sterile media for germplasm conservation and international exchange. For these systems, essentially any genotype can be successfully cultured in vitro with a single media formulation and light/temperature conditions. The wild species are more complex, and different media and growth conditions are required for different species. The development of friable embryogenic calli for transformation or gene editing is also rather genotype-sensitive, though many genotypes have now been successfully cultured for transformation.
Cassava is one of the few vegetatively propagated crops where important tissue culture efforts have been made for the production of doubled haploids. Ceballos et al. (2015) have listed the advantages that the use of inbred progenitors would have for the genetic enhancement of cassava:
-
Implementing the back-cross scheme and trait introgression. The deployment and impact of desirable traits (resistance to diseases and pests, special starch quality traits, herbicide tolerance, etc.) is slow and limited because their introgression requires breeding for a new variety de novo. Back-crossing is a highly successful breeding scheme used in many crops (Xu and Crouch 2008), but it cannot be applied to cassava because of the heterozygous nature of the progenitors currently used. The relevance of the homozygote advantage has been recently highlighted by the efforts to deploy commercial cassava varieties with the waxy (amylose-free) starch. Introgressing a single recessive gene implied developing a new variety. The best varieties from the first batch of waxy clones had a productivity level similar to normal (non-waxy) clones released 30 years ago. Although the second generation of waxy varieties are expected to quickly catch up (Karlström et al. 2016), the cost of introgressing a simple trait is unacceptably high.
-
Doubling of breeding value: Self-pollinating a heterozygous source of resistance to a given pest or disease and selecting a homozygous descendent would double the breeding value of the material when used as progenitor. Whereas 50% of the gametes produced by the heterozygous source carry the resistance gene, 100% of gametes from the homozygous version do (Ceballos et al. 2016a; Kaweesi et al. 2016).
-
Reduction of genetic load: Inbreeding exposes undesirable recessive alleles and allows a rapid reduction in their frequency (which tends to be relatively high in heterozygous populations).
-
Discovery of useful recessive traits: There are many examples in the literature of useful recessive traits including in cassava (Ceballos et al. 2007b, 2008).
-
Facilitated germplasm exchange and conservation: When inbred progenitors are available, their conservation and exchange could be through true-breeding botanical seed.
-
Development of superior hybrids by design, not by trial and error: Hybrid vigour (e.g. nonadditive genetic effects) can be progressively improved, but only through reciprocal recurrent selection methods (Bernardo 2014; Hallauer and Miranda Fo 1981) or through inbred line development within heterotic groups. Improving heterosis would be slow if no inbreeding were employed. The impact that the use of inbred progenitors has had in maize is unquestionable (Troyer 2006). The use of inbred progenitors offers the chance to maintain favourable gene combinations at different loci controlling the small, but critically relevant, nonadditive genetic effects as demonstrated in the case of maize (Crow 2000).
-
Facilitated conventional and molecular genetic studies: The availability of homozygous progenitors would facilitate greatly the logistics of conventional and molecular genetic studies (Gallais and Bordes 2007; Tuvesson et al. 2007).
-
Shortening the length of breeding cycles: The starting point for the selection process in Fig. 5.3 could be, for example, 30 plants (as currently required for PYT).
-
True collaboration and synergies among the few cassava breeding programs: Development and sharing of inbred lines with known characteristics and combining abilities by land granted US universities were the backbone of maize research during a significant part of the last century. Today, the collaboration between cassava programs is limited because of problems related to sharing germplasm (in vitro) and because the materials shared are finished products whose main characteristic is an outstanding performance in certain environments. Ideally, however, breeding programs should exchange good progenitors that can be used by other programs in crosses with local germplasm.
Inbred lines could be produced after successive self-pollinations. A few programs, especially CTCRI in India and IITA in Nigeria, have had medium- or long-term inbreeding programs. The first reported sequenced genome in cassava was on an S3 line developed at CIAT (Prochnik et al. 2012), which demonstrates that this is a feasible approach for cassava. However, successive self-pollinations are not practical because unavoidably it favours the production of early flowering types which are not desirable for breeding purposes and would require too many years (12–15 years). Instant homozygosity can be achieved through the production of doubled haploids, an advantage that was recognized many years ago (Woodward and Puonti-Kaerlas 2001). There are important ongoing efforts to develop a protocol for the production of doubled haploids through different strategies based on anther and microspore culture (androgenesis), ovary and ovule culture (gynogenesis) and wide crosses with Ricinus communis or irradiated pollen (parthenogenesis). There has been considerable progress towards inducing cell division in gametic tissue during the past few years (Perera et al. 2012, 2013), and research is now focusing on regenerating plants. Protocols for early embryo rescue (7–14 days after pollination) have also been developed, and plants were successfully regenerated. Early embryo rescue is fundamental in gynogenesis and parthenogenesis because the absence or abnormal fertilization in which these technologies are based results in weak embryos, poor or absent endosperm development and a strong tendency for fruit abortion.
10 Seed Production and Marketing
This chapter will mostly refer to seed as the botanical seed resulting from hybridization and used in the initial stages of selection of breeding programs. In this section, however, seed refers to the stem pieces or other clonal propagules used for planting the crop in other experimental or commercial production conditions. For the large majority of cassava production around the world, seed systems used by farmers have changed little over the past centuries. Because it is a vegetatively propagated crop, farmers are able to save planting material from their own production fields or trade informally with their neighbours, without any perceived need for a formal seed system to provide certified disease-free, high-quality and true-to-type seed. Farmers are generally underinformed about the potential improvements that can be made to seed quality, and scientists generally have inadequate research evidence to convince farmers of better options. This is due to the inadequate research on the subject, to the poor dissemination of results in easily accessed publications and to the generally poor extension systems to disseminate available information.
The basic feature of this traditional seed system is that farmers either store stems over a short period of time (from several days to a few months but typically a few weeks) or plant immediately after harvest of the previous crop without any significant storage period. Usually there is little or no discrimination between plants that are used for seed and those that are used strictly for root production. There are, however, efforts to promote the idea that part of the field should be targeted as the source of planting material for the following season. This system is described below.
This traditional system will adequately supply grower needs under stable production conditions and where no new varieties are being introduced. But obviously the goal of breeders is to see dissemination and impact from new varieties as efficiently and as quickly as possible. Multiplication and dissemination through traditional systems are very slow processes. This has the advantage of minimizing risk, in that any new variety will be well-proven over many years before it is grown on a large area, making dramatic failure unlikely. Cassava’s traditional slow multiplication rate is thus a built-in risk-management system to give varieties a chance to progressively prove themselves in real-life farm and market situations. On the other hand, for varieties that are truly superior, the impacts of their advantages and benefits may only be felt slowly and for a limited number of end users.
This informal system has obvious advantages and a proven track record of success. It has allowed cassava to succeed and expand as a crop over many centuries. Nonetheless, there are a number of changes impacting current and future cassava production which will drive changes in the way seed is produced and marketed. This section will review the drivers of change in cassava seed production and the opportunities afforded by new technologies and new systems.
Traditional seed systems work just fine where there are few production constraints and where farmers have no need or motivation to adopt new varieties at a higher pace. But these conditions are increasingly rare in modern times. Cassava production has continued to expand rapidly in the past 50 years, with much of this expansion in areas of stress such as poor soils and periodic drought, conditions which affect seed quality. In addition, pests and diseases are spreading, especially in Asia and Africa, and one of their primary modes of dispersion is through infested or infected stems used as planting material. Climate change includes enhanced uncertainty in the initiation of the rainy season. Delayed arrival of the rains implies extended storage period of the stems and (eventually) the possibility of sharp reduction in their sprouting capacity.
Consequently, improved seed systems have two main functions, which may be combined or independent in any given situation. First, seed systems are needed to improve access to high-quality seed even where current varieties are grown and used on-farm. Second, seed systems are needed to accelerate access to new varieties by farmers. The CGIAR Research Program on Roots, Tubers and Bananas (CRP-RTB ) has developed a framework for intervening in seed systems, specifically aimed at the major vegetatively propagated crops (RTB 2016).
10.1 Improving On-Farm Seed Production of Current Varieties
Typically, under stable production systems, about 10% of the plants in a cassava field need to be used for seed for the next planting season, i.e. a reproduction rate of about 1:10. This varies with variety and growing conditions. Typically, farmers do not make a distinction between managing plants that will be used for seed and those that will not. In other words, the planting material may be chosen on the basis of various factors, such as harvest date (i.e. material harvested closest to next planting season will be used for seed) or convenience for transportation of seed to next field for planting. There is generally little perception that different management practices can have significant effect on the quality of seed and consequently yield of the subsequent crop. Nonetheless, some broad guidelines can be provided that will allow farmers to maximize seed quality and the resulting contributions to yield.
A basic tenet of producing high-quality seed on-farm is that production management specifically oriented towards seed quality will be different from management aimed at maximizing returns from harvesting and selling roots. At the same time, since the commercial product (the roots) are not propagative material, there may be no, or only a small, trade-off between maximizing income from roots and producing highest-quality seed. A concept promoted at CIAT and with the Colombian national research agency, Corpoica, is the “corner of prosperity ” for seed management. For maintaining the same area of the same varieties from year to year, a farmer will need to use about 10% of the plants from each variety to supply the next season’s planting material. CIAT and Corpoica suggest that 10% of the production area (the “corner of prosperity”) be dedicated to management that will optimize quality of stem cuttings. This strategy will involve, in broad terms:
-
Selection of the best part of the field (well-drained; most fertile soil; protected against mammalian invaders such as deer, wild pigs, etc.; easiest access to irrigation if available).
-
Fertilizer to optimize plant health (which may be at levels that are higher than those that maximize net returns for root production).
-
Irrigation where extreme drought stress will significantly impact plant growth and development.
-
Excellent weed management.
-
Periodic inspection for preventive management of any negative impacts.
-
Control of pests and diseases, including roguing if virus symptoms appear.
-
Harvest of stems near to next season planting, to minimize seed deterioration during storage.
-
Harvest stems from plants whose roots have been inspected to be symptom-free from diseases such as CFSD or CBSD.
Unfortunately, there is little quantitative information to indicate specific returns on these various management practices, and there is a high level of need for well-designed research to obtain such data. While there are many publications recommending practices to produce and select high-quality planting material, the quantification of benefits that farmers should expect by specific management of a small part of their production field for high-quality seed is still not well established.
Even if a farmer does not manage a “corner of prosperity ” specifically for quality seed production, he or she can adopt several practices to improve seed quality. These have been well-documented in several publications (Ceballos and Calle 2010); Howeler and Maung Aye 2014). One of the key practices will be “positive selection” at harvest time. This concept involves inspection of both tops and roots in order to identify best sources of seed. In many cases, a healthy-looking plant above ground will correspond to healthy roots, high yield, and high-quality seed. However, there are some key seed-disseminated pathogens that may not have visible leaf or stem symptoms. For example, cassava frogskin disease (CFSD) and cassava brown streak disease (CBSD) may both be symptomless above ground while having moderate or even severe root symptoms. In these cases, the stems would carry the pathogen into the next production cycle and initiate a series of cycles of degeneration of seed quality. While there are sophisticated tools such as real-time PCR to detect the virus or phytoplasma, farmers can make good progress in reducing inoculum pressure by harvesting plants before cutting stems for seed and only selecting those plants without root symptoms of CFSD or CBSD . Empirical results have demonstrated that positive selection can efficiently keep CFSD under control in farmers’ fields.
10.2 Sustainable Systems for Supplying High-Quality Seed of New Varieties
The goal of most breeding programs is to periodically introduce new varieties to farmers – quickly and at scale. The traditional farmer-to-farmer distribution system will not adequately achieve this, and other, more intensive systems of intervention will need to be developed. Unlike the situation for seed crops, and especially hybrid varieties, the private seed sector for cassava is poorly developed and, in fact, non-existent in most countries due to generally low and variable demand. Most commonly, national research and extension agencies work together (sometimes in collaboration with processing enterprises such as starch factories) before and after varietal release with a system of multiplication and distribution to farmers. However, beyond the initial release and small-scale distribution, most countries do not have in place an effective continuing system to provide quality seed.
The majority of the research on developing high-quality seed at mass scale has focused on tissue culture systems and other forms of intensive rapid propagation. However, to date, these intensive systems have had little practical success outside the laboratory and experiment station. Exceptions are primarily where tissue culture has been applied at the very earliest stages of variety multiplication, followed by conventional multiplication through stem cuttings in later stages, up to farmer acquisition and use. Current thinking among most donors and the CGIAR is to promote public/private sector alliances which rely on some public support for start-up costs but ultimately are driven mainly by profits in the private sector to sustain seed systems that provide additional profits to farmers and have zero or minimal ongoing public sector costs. Two prominent examples are seed systems in Uganda and Nigeria. In both cases, the emphasis is on developing viable private sectors. This is a major challenge which has never been achieved on a significant scale. While seed traders are common in major producing countries, there is typically little management for seed quality. In SE Asia, for example, large-scale seed exchange occurs across the Thai-Vietnam-Cambodia borders but with little inspection or control for varietal purity, physiological quality or phytosanitary status. As cassava production intensifies and new superior varieties are more in demand, likewise the incentives for good seed systems will increase and should provide opportunities for sustainable public/private initiatives.
In 2016, a major cassava seed systems’ initiative was launched in Nigeria, funded by the Bill and Melinda Gates Foundation and implemented by CRP-RTB and IITA. This initiative pulls together the key lessons from many years of (often-failed) seed systems work, to try to positively affect productivity and profitability in the world’s top cassava producer. It is based, first and foremost, on the principles of sustainability, i.e. the mutual long-term profit advantages that will accrue to both private and public/private sector seed producers and farmers. In Nigeria, demand for seed is expected to be driven both by the need to provide seed of varieties degenerated from accumulated effects of CMD and by the demand for new, higher-yielding varieties to supply dynamic cassava markets.
10.3 Rapid Multiplication
10.3.1 In Vitro Systems to Support Seed Production
When a cassava plant is infected with a virus or phytoplasma , there may be few options to eliminate that pathogen other than tissue culture systems that involve some combination of small apical meristem culturing and thermo- or chemotherapy. Such systems have proven effective for viruses such as CMD and CBSV and for the virus-/phytoplasma-induced CFSD. It is also effective for most bacterial and fungal pathogens and for all insects and mites.
A system of maintaining breeder’s seed in an in vitro, pathogen-free environment is fundamental to assuring long-term access to highest-quality, true-to-type seed. Such systems typically need to be managed by only one or a very few laboratories in a given country, due to costs and expertise involved. Theoretically, tissue culture systems can also achieve very high multiplication rates – almost unlimited. At the high end of these possibilities is somatic embryogenesis, where individual cells can be induced to develop into somatic embryos and subsequently, through “artificial seeds,” into plants. In this case, millions of plants could be produced from a small amount of leaf tissue and the hardening of plantlets quickly become a major bottleneck. There are intermediate systems, such as micropropagation from in vitro plantlets, where nodal pieces are cut and propagated into new media, successively until the desired number of plants is derived to harden and taken to the field. In vitro systems have even been proposed and tested for use at the village and farm level in order to cut costs and improve efficiency, but these systems have been largely unsuccessful.
10.4 Rapid Multiplication from Sprouted Shoots and from Mini-Stakes
Intermediate-level technologies (between the traditional stem cuttings and sophisticated tissue culture systems) have been developed and used to some degree in applied, on-farm systems. If stems are planted horizontally in beds, the growing shoots can be harvested periodically over several months, rooted in water, hardened in containers (e.g. pots or bags) and transplanted to the field to produce conventional seed (mature stems) for further seed production or for commercial root production. In this system, a single plant in the field can theoretically produce several thousand plants within a year’s time. However, as with tissue culture systems, the practical use has been quite limited due to costs and the feasibility of covering those costs through added value compared to more conventional multiplication.
A very low-level technology to increase rate of reproduction for cassava is simply to reduce the length of the stem used for propagation. For example, if an average plant can produce ten 20 cm stakes, it could produce forty 5 cm stakes and quadruple the standard multiplication rate. The constraint to this system is that the plants resulting from the shorter stem pieces are likely to be less vigorous and robust, at least initially, which will require additional management inputs such as more fertilizer, water and weed control. There are numerous successful examples of the mini-stake system applied at the experimental level and some cases where it is adopted by progressive farmers.
10.4.1 The Way Forward for Cassava Seed Management
There have probably been more failures than successes in investments aimed at developing sustainable improved seed systems for cassava. While there is considerable research describing optimum seed management, there have been relatively few cases where this knowledge has translated to consistent farmer demand for improved seed. This demand is essential to sustain private sector investment. It now seems clear that in the past, donor or publicly funded investments in seed systems have overemphasized the more sophisticated approaches of tissue culture and other technologies that have been difficult to implement in ways that provide clear profit both for seed producers and for farmer-customers. The most difficult challenge for profit-based systems is in situations where the demand for new varieties is low. This can be the case where high-yielding, high-quality varieties are already available, and there is little pressure for farmers to change varieties.
This may be the situation in large areas of SE Asia, where varieties such as KU50 are already widespread and new varieties with clear superior performance are not common. The same is true of areas of Colombia (North Coast and Llanos) and Brazil (South) where new varieties are already widely used. While there is clearly continuing interest in testing and adopting new varieties in these areas, there are not compelling forces for change. On the other hand, in much of Africa, disease pressure, especially CMD and CBSD, is forcing farmers either to look for varieties that are resistant or to purchase clean seed of current varieties.
One of the major lessons from seed systems research in cassava is that clean seed by itself is not sufficient to control seed-borne diseases. For example, the Great Lakes Cassava Initiative (CRS 2010) attempted to produce and distribute seed free of CMV as a means of controlling the disease. However, the reinfection rate was too high, even during the seed multiplication process, such that the farmer had only a small or a short-term advantage to clean seed if the varieties were susceptible. It now seems clear that, in the case of some of the key pathogens, a strategy combining resistance and clean seed systems is essential to success.
11 Biotechnology Tools and Their Use in Cassava
A wide array of biotechnology tools has been developed during the past three decades. To facilitate a description of their impact and potential in cassava breeding, they will be grouped into five types and discussed separately below. The applications and huge potential of tissue culture protocols have already been described earlier.
11.1 Molecular Markers for Diversity and Identity Studies
Isozymes were the first type of molecular markers used in cassava. Typically, the first applications focused on analysing genetic diversity or for identification purposes (Ramírez et al. 1987; Hussain et al. 1987; Lefèvre and Charrier 1993). During the 1990s, different types of (DNA or RNA) molecular markers were gradually developed and used for these same initial purposes (Asante and Offei 2003; Carmo et al. 2015; Carvalho and Schaal 2001; Chepkoech et al. 2015; Kawuki et al. 2009; Maredia et al. 2016; Marmey et al. 1993; Moyib et al. 2007; Rabbi et al. 2015; Zacarias et al. 2004). Markers have also been used to distinguish hybrids from self-pollinations in breeding nurseries (Otti et al. 2011). These applications for molecular marker technologies offer clear advantages that become even more evident with their constant reduction of costs and enhancement of discriminating capacity.
The use of markers to identify clones grown by farmers is a powerful tool for impact assessment studies which are always relevant for cassava, considering the informal or weak seed systems. More often than not, there is no reliable way to know the area planted to different cassava varieties (with the exception of Thailand). Diversity studies based on molecular markers have been particularly useful to assess the relationship among different Manihot species and the evolution of this genus (Deputié et al. 2011; Olsen and Schaal 2001; Roa et al. 1997; Second et al. 1997).
There is a growing interest and need to conduct diversity studies aiming at identifying potential heterotic groups in cassava. One of the proposed strategies to be able to resume strong genetic gains for productivity would be the implementation of reciprocal recurrent selection (Ceballos et al. 2015) as already done in sweetpotato. This approach relies on two (perhaps three) breeding populations which exhibit high heterosis when crossed with each other (Hallauer and Miranda Fo 1981; Bernardo 2014). Unfortunately, genetic distances , based on molecular markers, do not seem to be good predictors for identifying potential heterotic groups in cassava (Ceballos et al. 2016b). A strategic effort needs to be made in cassava to develop a population structure that would facilitate the creation or identification of heterotic groups. One way to achieve this would be to focus on diverse gene pools that have evolved isolated from each other over a long period of time. CIAT has been working on the definition of diverse gene pools from its large germplasm collection using SNPs markers. Eight subpopulations have emerged from this diversity study (Becerra Lopez-Lavalle 2015). Representatives of each pool could be used initially as a proto-heterotic grouping.
11.2 Molecular Markers for Diagnostic Tools and Plant Health
Molecular diagnostic tools can be used for diseases of complex aetiology, such as frogskin disease (Alvarez et al. 2009; Calvert et al. 2008), detection and quantification of viral diseases (Monger et al. 2001; Kaweesi et al. 2014) and analysis of their genetic diversity (Calvert et al. 2008; Legg et al. 2011; Monger et al. 2001), in the characterization and diversity studies of fungal and bacterial diseases (Restrepo and Verdier 1997; Álvarez et al. 2003; Wydra et al. 2004) as well as in gene expression studies in host-pathogen interactions (Kemp et al. 2004, 2005; Fregene et al. 2004; Maruthi et al. 2014). An interesting application of molecular markers has been for the dissection of the pathway leading to post-harvest physiological deterioration (PPD) in cassava roots (Reilly et al. 2007). This is not a comprehensive list of publications on the subject.
The availability of tools for understanding genetic diversity of pests and diseases is an important asset for developing strategies for durable and efficient resistance in cassava. Reliable and affordable diagnostic tools are fundamental for the safe exchange of cassava germplasm.
11.3 Marker-Assisted Selection (MAS )
The first molecular map for cassava was published nearly two decades ago (Fregene et al. 1997). Since these pioneering days, a large number of research articles have been published and will not be exhaustively listed here. QTL maps for a broad range of traits (reaction to anthracnose, CMD, CBB, whiteflies, HCN or CG, DMC, early bulking, PPD, plant architecture, carotenoids and protein contents in roots, FRY, etc.) and based on different type of markers (RFLP, AFLP, SSR, DArT, RAPD, etc.) are available (Akinbo et al. 2007, 2011, 2012; Blair et al., 2007; Chen et al. 2012; Ferguson et al. 2012; Kizito et al. 2007; Kunkeaw et al. 2011; Okogbenin and Fregene 2002, 2003; Mkumbira et al. 2003; Whankaew et al. 2011 to list just a few). A comprehensive summary of applications of molecular markers in cassava and progress so far achieved was published by Ferguson and co-workers in 2011. In the last two decades, the cost of genotyping has been drastically reduced. The latest technology of genotyping by sequencing based on SNPs has also been applied to cassava (Rabbi et al. 2014b) and opens up the possibility of implementing genomic selection and marker-assisted recurrent selection as described in the following section.
CMD is not present in Latin America, and therefore nearly all cassava germplasm from this region appears to be susceptible to the disease (Okogbenin et al. 1998). Resistance was reported to be controlled by a single dominant gene, designated as CMD2 (Akano et al. 2002; Fregene et al. 2000) although evidence for different sources has also been reported (Lokko et al. 2006a, b; Okogbenin et al. 2012). CIAT, in collaboration with cassava breeding programs in the African National Agriculture Research System and International Institute of Tropical Agriculture (IITA), has exploited CMD2 for the development of CMD-resistant Latin American germplasm in the centre of origin using MAS (Akano et al. 2002; Okogbenin et al. 2007, 2012). This is the first report of MAS in cassava. The advantage of the application is obvious: because CMD is absent in the Americas, the availability of molecular markers was the only alternative for selection.
To this day, the selection for resistance to CMD is the only example of MAS applied to cassava (Ferguson et al. 2011). Although markers are available for other traits (e.g. waxy starch, yellow root parenchyma, etc.), their use is economically not competitive. For these high-heritability traits, selection could be made as early as 6 months after planting a seedling nursery (Belalcazar et al. 2016), with the advantage that along with the key trait, the breeder can select for other characteristics (vigour, plant architecture, resistance to thrips, etc.). Typically, a seedling nursery for these types of traits has around 15,000–20,000 genotypes. Growing 1–2 ha of seedling plants is still much less expensive than extracting and analysing DNA samples for so many genotypes, but costs for the latter continue to decline.
The use of markers for selection purposes in a large segregating population has so far failed to deliver the expected benefits. MAS, however, could have interesting applications for the selection of progenitors (Ceballos et al. 2016a). For example, progenitors that are homozygous for CMD2 would have twice the breeding value (for this trait) compared with heterozygous progenitors. Markers could be used to identify genotypes that are homozygous for CMD2. In the case of carotenoids, it may be desirable to reduce the activity of carotenoid β-hydroxylase, controlling the conversion of β-carotene into other molecules. Molecular markers could be used to identify genotypes heterozygous for this gene, self-pollinate them and select progenies that are homozygous recessive (Ceballos et al. 2013; Morillo-C et al. 2012). Again, by this process, the breeding value of such self-pollinated genotypes would be better than that of its progenitor. As is often the case, application of new technologies to cassava requires special adaptation. Most efforts to apply MAS in cassava have been for selection in segregating progenies, which has until now had negligible impact. However, no effort has been made so far, in the selection of better progenitors.
The genome of cassava has been sequenced (Bredeson et al. 2016; Prochnik et al. 2012). The information generated by these studies and the public access to it is very relevant. For example, it has been used for determining the sequence in cassava of key genes related to starch biosynthesis or herbicide tolerance. With that information, reverse genetic approaches to screen germplasm collections in search of sources of useful alleles is an alternative that has been already been initiated (Duitama et al. 2017) as described earlier.
11.4 Genomic Selection
Genomic selection (GS ) simultaneously tags many loci across the entire genome to estimate genomic estimated breeding values or GEBV. It offers several advantages and overcomes key problems of MAS based on QTLs. Key among the different advantages is that several different traits can be improved simultaneously through a selection index , similar to those based on phenotypic traits and economic values. de Olviera and co-workers suggested the potential of GS for cassava in 2012. GS was also proposed for other crops (Heslot et al. 2012) characterized by the fact that breeding is based on the use of inbred progenitors. Different articles emphasize that GS would maximize genetic gains by unit of time (De Oliveira et al. 2012; Heffner et al. 2009). Like conventional breeding, GS has serious limitations for the selection of low narrow-sense heritability traits because of their low additive genetic effects in relation to the phenotypic variance. Genotype-by-environment interactions also affect the precision of GS estimates as well as the relatedness among genotypes. Ceballos et al. (2015) predicted that GS would be effective in high-heritability traits such as plant architecture, pest and disease resistance or DMC. However, GS was not expected to be efficient in improving FRY because of the relatively high influence of nonadditive genetic effects and the large within-family genetic variation generated by the heterozygous progenitors used in cassava (Table 5.1). Other factors may limit further the usefulness of breeding value in cassava (Ceballos et al. 2016a; Joaqui et al. 2016).
There is an ongoing project (NextGen Cassava Project ) to test GS in cassava led by Cornell University with field work in Uganda and Nigeria (Rabbi et al. 2015). As expected, GS is proving to be efficient improving traits such as resistance to CMD (Wolfe et al. 2016b; Rabbi et al. 2014a). Whereas DMC was also increased considerably, progress increasing FRY was, as predicted, not satisfactory (Wolfe et al. 2016a). An honest assessment of cost/benefit for technologies such as GS will have to be made in the years to come. It should be pointed out that improving resistance to CMD or increasing DMC is not enough for such an expensive approach that has the additional restriction of the need of “closed” breeding populations. The use of inbred progenitors would overcome some of the problems that implementing GS in cassava has. The induction of flowering would also benefit greatly the success of GS. Alternatives of GS combined with reciprocal recurrent selection would also be an interesting approach to overcome, at least partially, the problems of nonadditive genetic effects influencing FRY, which is ultimately the most important trait to improve.
11.5 Genetic Transformation and Gene Editing
The first published reports of genetic transformation in cassava date back from two decades ago (Li et al. 1996; Raemakers et al. 1996; Schöpke et al. 1996; Sarria et al. 1995). The technology offers a great potential considering the problems related to trait introgression discussed earlier, challenges to improve the productivity after successful clones such as KU50 were released and limited knowledge of genetic variability available in cassava germplasm. As it is often the case, genetic transformation initially suffered from genotypic dependency: initially a single genotype could be efficiently transformed (60444). However, protocols were improved quickly, and many genotypes have now been transformed (Chauhan et al. 2015; Liu et al. 2011; Raemakers et al. 2001; Taylor et al. 2004, 2012; Zainuddin et al. 2012). The commonly used methods for the genetic transformation of cassava include Agrobacterium-mediated gene delivery and particle bombardment. The explants used for transformation include somatic cotyledons and friable embryogenic calli.
A broad range of traits have been considered for genetic transformation: (a) Resistance to CMD and CBSD (Bi et al. 2010; Chellappan et al. 2004; Ntui et al. 2015; Patil et al. 2011; Vanderschuren et al. 2007; Zhang et al. 2005; Yadav et al. 2011); (b) Enhanced nutritional quality of the roots including high carotenoids, Fe, Zn, proteins as well as reduction in cyanogenic glucosides (Jørgensen et al. 2005; Leyva-Guerrero et al. 2012; Sayre et al. 2011; Welsch et al. 2010); (c) Quantity and quality of starches (Ihemere et al. 2006; Koehorst-van Putten et al. 2012; Zhao et al. 2011); (d) Reduction of PPD (Zidenga et al. 2012) and physiological traits such as leaf retention (Zhang et al. 2010); (e) Herbicide tolerance (Sarria et al. 2000, initially reported in 1995); (f) Induction of flowering (Adeyemo et al. 2008).
Genetic transformation for new starch types (e.g. waxy starch) has been successful and the phenotype of transgenic material fulfilled expectations regarding amylose content in the starch (Koehorst-van Putten et al. 2012; Zhao et al. 2011). The discovery of a spontaneous mutation for waxy starch may have reduced the appeal of waxy transgenic cassava. However, the complications of introgressing a single recessive gene into successful commercial varieties should be taken into consideration, at least for countries where the growth and commercialization of transgenic crops/products are more relaxed. Improving AGPase activity can contribute to the conversion of sugar to starch and subsequently increase the starch quantity. Early work (Ihemere et al. 2006) suggested that this is indeed the case. However, no further research on the subject has been published since then. Smith (2008) pointed out some of the potential stumbling blocks that may explain why no further efforts in this area have been published.
With the support of the Bill and Melinda Gates Foundation and several years of global cooperation, the BioCassava Plus project team has developed a number of transgenic cassava lines with value-added traits, such as improved protein content and increased vitamin A, iron and zinc contents. In spite of the promising results forecasted by Sayre and co-workers in 2011 - genetic transformation regarding enhanced nutritional quality, not a single product has shown promising results in the field. In the case of high carotenoids, transgenic materials show a drastic reduction of DMC. Early claims for enhanced protein content in the root had to be retracted. In addition, attempts to elevate protein levels in roots had impacts on overall nitrogen allocation between leaves and roots reflecting the strong leaf sink strength for reduced nitrogen (Leyva-Guerrero et al. 2012). Recent efforts by BioCassava Plus initiative using a new construct for enhanced Fe and Zn in the roots have yielded promising results (Gaitán-Solís et al. 2015).
The alternative to develop virus resistant transgenic cassava is also a justified and relevant objective. Although there is genetic resistance to CMD, apparently it is based on a single source. Chance of the resistance breaking down is a feasible threat, thus justifying the development of a new source of resistance. In the case of CBSD, available natural resistance is not as effective as in the case of CMD2, and, therefore, efforts have concentrated on resistance to CBSD. Transformed materials have shown excellent levels of resistance to CBSD (Chauhan et al. 2015; Odipio et al. 2014; Vanderschuren et al. 2012). However, transgenic material resistant to CBSD would not be useful if they are susceptible to CMD. Therefore, farmers preferred varieties carrying the CMD2 resistance were transformed to incorporate the proven resistance to CBSD. Unfortunately, the process of somatic embryogenesis used to regenerate cassava caused the resulting plants to become susceptible to CMD, even though the original clones carried the CMD2 resistance (Beyene et al. 2016). This is a systematic process observed by two independent research groups.
Genetic transformation remains a promising technology. Early experiences have been humbling and have helped expose the complexities in plant physiology and metabolism. Hopes to increase productivity or enhanced levels of proteins in the roots may need a long time to materialize. But for simpler goals such as starch variants, virus resistance (in spite of the current drawback of loss of CMD2 resistance), herbicide tolerance and (perhaps) tolerance to PPD, the technology may have a positive impact. Advances have also been made regarding the legal framework for field evaluation of transgenic materials in different African countries, Colombia, Indonesia and China. No commercial exploitation of transgenic cassava has been requested or authorized yet.
Associated with the potential of genetic transformation is the emerging alternative of gene editing, for example, based on CRISPR (Odipio et al. 2016). Gene editing can overcome some of the regulatory problems that genetic transformation has. Regeneration of protoplasts is feasible in cassava (Sofiari et al. 1998). This is important because regulatory issues are less limiting when regeneration is from edited protoplasts. Four copies of the GUS gene have already been edited in cassava (P. Chavarriaga, personal communication), and it is expected that soon field evaluations of edited cassava will be reported. Editing inbred progenitors would also allow overcoming the regulatory issues, and this is yet another reason justifying the need to develop inbred cassava.
12 Future Prospects and Outlook
Cassava is today a key food security staple and a competitive commodity for different industries. It will be even more important in the next few decades. Population growth will be particularly high in Africa, where cassava is a fundamental source of calories for the diet of millions of people. Increases in the concentration of CO2 in the air will improve its productivity, thus reducing the physiological advantage that C4 crops such as maize have as a feedstock for industrial processes. There is, however, an urgent need to fundamentally change the way cassava is bred, so that major advances in productivity can be attained again and new technologies can finally have a positive impact on the crop.
Cassava is the only diploid species, among the major staple crops, whose breeding is based on the use of heterozygous progenitors. Breeding should shift from the current system in which elite progenitors are crossed – hoping to find an outstanding hybrid by a process that is largely trial and error – to improving progenitors that would more predictably produce better clones. Initially, this approach is more complex because a pre-breeding step (e.g. parental development) needs to be added. However, the added complexity is only temporary. Once a group of a few elite inbred progenitors, producing outstanding hybrids, is identified, the system becomes much simpler and more efficient. There are several advantages that justify the use of inbred progenitors: genetic variability would be conveniently partitioned in ways that breeders can more easily exploit; dominance and epistatic effects can be efficiently and predictably exploited when inbred progenitors are used; heterosis can gradually and systematically be enhanced; trait introgression would not result in yield penalties as is happening today and back-crossing could be implemented; storage and exchange of germplasm (as botanical seed that breeds true) would be facilitated; and collaboration among the few cassava breeding programs will be greatly facilitated allowing true synergies among them.
Genomic selection offers a great potential. It allows identifying genotypes carrying a set of desirable alleles that can reliably be identified as such. However, genomic selection only allows “seeing” the good and bad alleles that each genotype has. Combining desirable alleles and maintaining them together are nearly impossible if heterozygous progenitors are used. On the other hand, if inbred progenitors are used, desirable alleles can gradually and systematically be assembled in a given genotype. Every step accumulating desirable alleles is maintained. In the current system, every meiotic event recombines randomly the alleles, essentially erasing a considerable proportion of the genetic progress made in generating the genotype where it takes place. Current applications of GS in maize breeding could be readily implemented for cassava, if inbred progenitors were available.
Other applications of molecular markers can have an impact in cassava breeding as well. However, cost/benefits should be taken into account. MAS for the sake of using it should be avoided. There are examples where molecular markers offer appealing advantages that have not been exploited. Improving the breeding value of progenitors could have an immediate and obvious impact. Molecular markers could be used to screen germplasm collections in search of sources of useful mutations. The discovery of the waxy starch mutation in cassava illustrates the wealth of genetic diversity that has remained hidden in germplasm collections and needs to be exposed and exploited. Screening germplasm collections through conventional phenotyping methods is cumbersome, time consuming and expensive. Molecular tools offer clear advantages that have not yet been considered. These applications of molecular markers are helpful to support the idea that new technologies in cassava need to be applied but in ways that differ from the conventional uses for other crops.
Genetic transformation in cassava has evolved considerably in the last two decades. Protocols of genetic transformation in cassava have improved considerably in the last two decades. Protocols are efficient and many cultivars can be transformed. A broad set of traits have been addressed by several laboratories that have succeeded transforming cassava in Africa, Asia, Latin America, Europe and the USA. Although there have been unexpected drawbacks, they have contributed to a better understanding of the complexities of gene regulation and expression, and some of them should be quickly overcome. It would be very useful to have at least one example of commercial exploitation of transgenic technologies in cassava. Surprisingly no effort has been made to release herbicide-tolerant transgenic cassava. Weed management is labour-intensive (requiring one to two person-months each year per hectare) and is often carried out by women. Herbicide tolerance would also allow direct planting, with further protection of the environment, reducing costs and allowing more mechanized operations. This would also be fundamental for the future of cassava as a competitive feedstock for industrial uses.
Finally, as different emerging technologies are developed and applied, there is a tension between the old and the new approaches. Young scientists, trained in novel technologies, have the legitimate interest in applying them to cassava. Cassava research now is a multipronged effort with limited coordination. Scientists unavoidably promote the use of the technologies with which they work. This is a reasonable attitude. However, farmers’ needs require that the best and most efficient technologies are used to maximize the chances of developing superior hybrids. There is also a tension between centres that specialize in a particular technology and scan for opportunities to apply them. The commitment of these centres is for these technologies, not to cassava. Ideally, resources should be allocated to institutions and programs that have clear and long-standing commitment to cassava research.
References
Abraham A (1957) Breeding of tuber crops in India. Indian J Gen Plant Breed 17:212–217
Adeyemo OS, Davis SJ, Chavarriaga P, Tohme J, Ceballos H, Fregene M (2008) Strategies for enhancing flowering in cassava using molecular tools: towards a more efficient breeding program. First Scientific Meeting of the Global Cassava Partnership GCP-1. Ghent. p 51
Aiemnaka P, Wongkaew A, Chanthaworn J et al (2012) Molecular characterization of a spontaneous waxy starch mutation in cassava (Manihot esculenta Crantz). Crop Sci 52:2121–2130
Akano AO, Dixon AGO, Mba C, Barrera E, Fregene M (2002) Genetic mapping of a dominant gene conferring resistance to cassava mosaic disease. Theor Appl Genet 105:521–525
Akinbo O, Gedil M, Ekpo EJA, Oladele J, Dixon AGO (2007) Detection of RAPD markers-linked to resistance to cassava anthracnose disease. Afr J Biotechnol 6:677–682
Akinbo O, Labuschagne M, Fregene M (2010) Embryo rescue as a method to develop and multiply a backcross population of cassava (Manihot esculenta Crantz) from an interspecific cross of Manihot esculenta ssp. flabellifolia. Afr J Biotechnol 9:7058–7062
Akinbo O, Labuschagne M, Fregene M (2011) Introgression of whitefly (Aleurotrachelus socialis) resistance gene from F1 inter-specific hybrids into commercial cassava. Euphytica 183:19–26
Akinbo O, Labuschagne M, Marin J et al (2012) QTL analysis for root protein in a backcross family of cassava derived from Manihot esculenta ssp flabellifolia. Trop Plant Biol 5:1–12
Allem AC (1999) The closest wild relatives of cassava (Manihot esculenta Crantz). Euphytica 107:123–133
Allem AC (2002) The origins and taxonomy of cassava. In: Hillocks RJ, Tresh JM, Bellotti AC (eds) Cassava: biology, production and utilization. CABI Publishing, Wallingford, pp 1–16
Allem AC, Mendes RA, Salomão AN, Burle ML (2001) The primary gene pool of cassava (Manihot esculenta Crantz subspecies escultenta, Euphorbiaceae). Euphytica 120:127–132
Alvarez E, Mejía JF, Llano GA, Loke JB, Calari A, Duduk B, Bertaccini A (2009) Characterization of a phytoplasma associated with frogskin disease in cassava. Plant Dis 93:1139–1145
Álvarez E, Mejía JF, Valle T (2003) Molecular and pathogenicity characterization of Sphaceloma manihoticola isolates from south-central Brazil. Plant Dis 87:1322–1328
Angel F, Barney VE, Tohme J, Roca WM (1996) Stability of cassava plants at the DNA level after retrieval from 10 years of in vitro storage. Euphytica 90:307–313
Anyanwu CN, Ibeto CN, Ezeoha SL, Ogbuagu NJ (2016) Sustainability of cassava (Manihot esculenta Crantz) as industrial feedstock, energy and food crop in Nigeria. Renew Energy 81:745–752
Asante IK, Offei SK (2003) RAPD-based genetic diversity study of fifty cassava (Manihot esculenta Crantz) genotypes. Euphytica 131:113–119
Ashby JA, Quirós CA, Rivera YM (1987) Farmer participation in on-farm variety trials. Agricultural Administration (Research and Extention) Network Discussion Paper No. 22. London. Overseas Development Institute
Balagopalan C (2002) Cassava utilization in food, feed and industry. In: Hillocks RJ, Tresh JM, Bellotti AC (eds) Cassava: biology, production and utilization. CABI Publishing, Wallingford, pp 301–318
Becerra Lopez-Lavalle LA (2015) Revisiting cassava genetic diversity reveals eco-geographic signature of the crop’s domestication. Plant and Animal Genome XXIII, San Diego
Beeching JR, Marmey P, Gavalda MC, Noirot M, Haysom HR, Hughes MA, Charrier A (1993) An assessment of genetic diversity within a collection of cassava (Manihot esculenta Cranz) germplasm using molecular markers. Ann Bot 72:515–520
Belalcazar J, Dufour D, Pizarro M et al (2016) High-throughput phenotyping and improvement in breeding cassava for increased carotenoids in the roots. Crop Sci 56:2916–2925
Bellotti AC, Arias B (2001) Host plant resistance to whiteflies with emphasis on cassava as a case study. Crop Prot 20:813–823
Benesi IRM, Labuschagne MT, Herselman L, Mahungu N (2010) Ethnobotany, morphology and genotyping of cassava germplasm from Malawi. J Biol Sci 10:616–623
Bernardo R (2014) Essentials of plant breeding. Stemma Press, Woodbury
Bertram RB (1993) Application of molecular techniques to genetic resources of cassava (Manihot esculenta Crantz, Euphorbiaceae): interspecific evolutionary relationships and intraspecific characterization. Ph.D. dissertation. University of Mariland. 465 pp
Bertram RB, Schaal BA (1993) Phylogeny of Manihot and the evolution of cassava. CBN Newsl 1:4–6
Beyene G, Chauhan RJ, Wagaba H et al (2016) Loss of CMD2-mediated resistance to cassava mosaic disease in plants regenerated through somatic embryogenesis. Mol Plant Pathol 17:1095–1110
Bi H, Aileni M, Zhang P (2010) Evaluation of cassava varieties for cassava mosaic disease resistance jointly by agro-inoculation screening and molecular markers. African J Plant Sci 4(9):330–338
Biggs BJ, Smith MK, Scott KJ (1986) The use of embryo culture for the recovery of plants from cassava (Manihot esculenta Crantz) seeds. Plant Cell Tissue Organ Cult 6:229–234
Blair MW, Fregene MA, Beebe SE, Ceballos H (2007) Marker assisted selection in common Beans and Cassava. In: Guimaraes EP, Ruane J, Scherf BD, Sonnino A, Dargie JD (eds) Marker-Assisted Selection (MAS) in crops, livestock, forestry and fish: current status and the way forward. Food and Agriculture Organization of the United Nations (FAO), Rome, pp 81–115
Bolhuis GG (1953) A survey of some attempts to breed cassava varieties with a high content of proteins in the roots. Euphytica 2:107–112
Bredeson JV, Lyons JB, Prochnik SE et al (2016) Sequencing wild and cultivated cassava and related species reveals extensive interspecific hybridization and genetic diversity. Nat Biotechnol. doi:10.1038/nbt.3535
Bueno A (1991) Estimación de los parámetros genéticos en la yuca. In: Hershey CH (ed) Mejoramiento genético de la yuca en América Latina. CIAT Publication, Cali, pp 197–220
Buitrago AJA (1990) La yuca en la alimentación animal. Centro Internacional de Agricultura Tropical. CIAT, Cali. 446 p
Bull SE, Owiti JA, Niklaus M, Beeching JR, Gruissem W, Vanderschuren H (2009) Agrobacterium-mediated transformation of friable embryogenic calli and regeneration of transgenic cassava. Nat Protoc 4:1845–1854
Byrne D (1984) Breeding cassava. Plant Breed Rev 2:73–134
Cach NT, Perez JC, Lenis JI, Calle F, Morante N, Ceballos H (2005) Epistasis in the expression of relevant traits in cassava (Manihot esculenta Crantz) for subhumid conditions. J Hered 96(5):586–592
Calvert LA, Cuervo M, Lozano I, Villareal N, Arroyave J (2008) Identification of three strains of a virus associated with cassava plants affected by frogskin disease. J Phytopathol 156:647–653
Caparros Megido R, Alabi T, Nieus C et al (2016) Optimisation of a cheap and residential small-scale production of edible crickets with local by products as an alternative protein-rich human food source in Ratanakiri Province, Cambodia. J Sci Food Agric 96:627–632
Carmo CD, Santos DB, Alves LB, Oliveira GAF, Oliveira EJ (2015) Development of TRAP (Target Region Amplification Polymorphism) as new tool for molecular genetic analysis in cassava. Plant Mol Biol Rep 33:1953–1966
Carvalho C-BLJ, Schaal BA (2001) Assessing genetic diversity in the cassava (Manihot esculenta Crantz) germplasm collection in Brazil using PCR-based markers. Euphytica 120:133–142
Carvalho LJCB, de Souza CRB, Cascardo JCM, Junior CB, Campos L (2004) Identification and characterization of a novel cassava (Manihot esculenta Crantz) clone with high free sugar content and novel starch. Plant Mol Biol 56:643–659
Ceballos H, Becerra LA, Calle F, Morante N, Ovalle T, Pérez JC, Hershey C (2016) Genetic distance and heterosis in cassava. Euphytica 210:79–92. doi:10.1007/s10681-016-1701-7
Ceballos H, Calle F (2010) Cassava. In: Fajardo J, NeBambi L, Larinde MI, Rosell C, Barker I, Roca W, Chujoy E (eds) Quality declared planting material. Protocols and standards for vegetatively propagated crops. FAO Plant Production and Protection Paper 195. p 33–40. Food and Agriculture Organization (FAO) of the United Nations, Rome
Ceballos H, Fregene M, Pérez JC, Morante N, Calle F (2007) Cassava genetic improvement. In: Kang MS, Priyadarshan PM (eds) Breeding major food Staples. Blackwell Publishing, Ames, pp 365–391
Ceballos, Hershey (2016) Road map for cassava genetic improvement. Proceedings of the World Congress on Root and Tuber Crops and Third Scientific Conference of the Global Cassava Partnership for the 21st Century. Nanning, China. January 2016
Ceballos H, Hershey C, Becerra-López-Lavalle LA (2012) New approaches to cassava breeding. Plant Breed Rev 36:427–504. Hoboken, Wiley-Blackwell
Ceballos H, Iglesias CA, Pérez JC, Dixon AGO (2004) Cassava breeding: opportunities and challenges. Plant Mol Biol 56:503–515
Ceballos H, Jaramillo JJ, Salazar S, Pineda LM, Calle F, Setter T (2017) Induction of flowering in cassava through grafting. J Plant Breed Crop Sci 9:19–29
Ceballos H, Kawuki RS, Gracen VE, Yencho GC, Hershey CH (2015) Conventional breeding, marker assisted selection, genomic selection and inbreeding in clonally propagated crops: a case study for cassava. Theor Appl Genet 128:1647–1667
Ceballos H, Morante N, Sánchez T, Ortiz D, Aragón I, Chávez AL, Pizarro M, Calle F, Dufour D (2013) Rapid cycling recurrent selection for increased carotenoids content in cassava roots. Crop Sci 53:2342–2351
Ceballos H, Okogbenin E, Pérez JC, Becerra LA, Debouck D (2010) Cassava. In: Bradshaw J (ed) Root and tuber crops. Springer Publishers, New York, pp 53–96
Ceballos H, Pérez JC, Joaqui-B O, Lenis JI, Morante N, Calle F, Hershey H (2016) Cassava breeding I: the value of breeding value. Front Plant Sci. doi:10.3389/fpls.2016.01227
Ceballos H, Ramirez J, Bellotti AC, Jarvis A, Alvarez E (2011) Adaptation of cassava to changing climates. In: Yadav S, Redden B, Hatfield JL, Lotze-Campen H (eds) Crop adaptation to climate change. Wiley-Blackwell Publishers. pp 411–425
Ceballos H, Sánchez T, Morante N et al (2007) Discovery of an amylose-free starch mutant in cassava (Manihot esculenta Crantz). J Agric Food Chem 55:7469–7476
Ceballos H, Sánchez T, Tofiño AP et al (2008) Induction and identification of a small-granule, high-amylose mutant in cassava (Manihot esculenta Crantz). J Agric Food Chem 56:7215–7222
Chalwe A, Melis R, Shanahan P, Chiona M (2015) Inheritance of resistance to cassava green mite and other useful agronomic traits in cassava grown in Zambia. Euphytica 205:103–119
Chauhan RD, Beyene G, Kalyaeva M, Fauquet CM, Taylor N (2015) Improvements in Agrobacterium-mediated transformation of cassava (Manihot esculenta Crantz) for large-scale production of transgenic plants. Plant Cell Tiss Org Cult 121:591–603
Chellappan P, Masona MV, Vanitharani R, Taylor NJ, Fauquet CM (2004) Broad spectrum resistance to ssDNA viruses associated with transgene-induced gene silencing in cassava. Plant Mol Biol 56:601–611
Chen X, Fu Y, Xia Z, Jie L, Wang H, Lu C, Wang W (2012) Analysis of QTL for yield-related traits in cassava using an F1 population from non-inbred parents. Euphytica 187:227–234
Chen H, Xu M-l, Guo Q, Yang L, Ma Y (2016) A review on present situation and development of biofuels in China. J Energy Inst 89:248–255
Chepkoech E, Kinyua M, Kiplagat O, Arunga EE, Kimno S (2015) Genetic diversity of cassava mutants, hybrids and landraces using simple sequence repeat markers. Am J Exp Agric 5(4):287–294
Chipeta MM, Bokosi JM, Saka VW, Benesi IRM (2013) Combining ability and mode of gene action in cassava for resistance to cassava green mite and cassava mealy bug in Malawi. J Plant Breed Crop Sci 5:195–202
CIAT (1991) In: Hershey CH (ed). Mejoramiento genético de la yuca en América Latina. CIAT, Cali p 426. (ISBN 958-9183-16-6)
Cock J (1985) Cassava. New potential for a neglected crop. Westview Press, Boulder
Cours G (1951) Le manioc a Madagascar. Memoires de L’Institut Scientifique de Madagascar Serie B 3:203–400
Crow JF (2000) The rise and fall of overdominance. Plant Breed Rev 17:225–257
CRS (2010) Final report on the Great Lakes Cassava initiative. 137p. http://www.crs.org/sites/default/files/tools-research/final-report-great-lakes-cassava-initiative.pdf
Davrieux F, Dufour D, Dardenne P et al (2016) LOCAL regression algorithm improves NIRS predictions when the target constituent evolves in breeding populations. JNIRS 24:109–117
De Bruijn GH (1977) Influence of day length on the flowering of cassava. Trop Root Tuber Crops Newsl 10:1–3
De Carvalho RD, Guerra-M M (2002) Cytogenetics of Manihot esculenta Crantz (cassava) and eight related species. Hereditas 136:159–168
De Oliveira EJ, de Resende V, da Silva Santos V et al (2012) Genome-wide selection in cassava. Euphytica 187:263–276
Delgado A, Hays DB, Bruton RK, Ceballos H, Novo A, Boi E, Gomez Selvaraj M (2017) Ground penetrating radar: a case study for estimating root bulking rate in cassava (Manihot esculenta Crantz). Plant Methods 13:65. https://doi.org/10.1186/s13007-017-0216-0
Deputié A, Salick J, McKey D (2011) Evolutionary biogeography of Manihot (Euphorbiaceae), a rapidly radiating Neotropical genus restricted to dry environments. J Biogeogr 38(6):1033–1043
Diasolua Ngudi D, Kuo Y-H, Lambein F (2003) Amino acid profiles and protein quality of cooked cassava leaves or ‘saka-saka’. J Sci Food Agric 83:529–534
Duitama J, Kafuri L, Tello D, Leiva AM, Hofinger B, Datta S, Lentini Z, Aranzales E, Till B, Ceballos H (2017) Deep assessment of genomic diversity in cassava for herbicide tolerance and starch biosynthesis. Comput Struct Biotechnol J 15:185–194
Easwari Amma CS, Sheela N, Thankamma Pillai PK (1995) Combining ability analysis in cassava. J Root Crops 21(2):65–71
El-Sharkawy MA (2009) Pioneering research on C4 leaf anatomical, physiological, and agronomic characteristics of tropical monocot and dicot plant species: implications for crop water relations and productivity in comparison with C3 cropping systems. Photosynthetica 47:163–183
Epperson JE, Pachico DH, Guevara CL (1997) A cost analysis of maintaining cassava plant genetic resources. Crop Sci 37:1641–1649
Escobar RH, Mafla G, Roca WM (1997) A methodology for recovering cassava plants from shoot tips maintained in liquid nitrogen. Plant Cell Rep 16:474–478
Esuma W, Herselman L, Labuschagne MT et al (2016) Genome-wide association mapping of provitamin A carotenoid content in cassava. Euphytica. doi:10.1007/s10681-016-1772-5
Falconer DS (1981) Introduction to quantitative genetics, 2nd edn. Longman, New York
FAO (2013) In: Howeler R, NeBambi L, Thomas G (eds) Save and grow: cassava. A guide to sustainable production intensification. Food and Agriculture Organization (FAO) of the United Nations, Rome, p 129 p. ISBN 978-92-5-107641-5
Ferguson ME, Hearne SJ, Close TJ et al (2012) Identification, validation and high-throughput genotyping of transcribed gene SNPs in cassava. Theor Appl Genet 124(4):685–695
Ferguson ME, Rabbi IY, Kim DJ, Gedil M, Becerra Lopez-Lavalle LA, Okogbenin E (2011) Molecular markers and their application to cassava breeding: past, present and future. Trop Plant Biol 5:95–109
Fisher T, Buerlee D, Edmeades G (2014) Crop yields and global food security. Will yield increase continue to feed the world? ACIAR Monograph, vol 158. Australian Centre for International Agricultural Research, Canberra, pp 285–289
Fregene M, Angel F, Gomez R, Rodríguez F, Chavarriaga P, Roca W, Tohme J (1997) A molecular genetic map of cassava (Manihot esculenta Crantz). Theor Appl Genet 95:431–441
Fregene M, Bernal A, Duque M, Dixon A, Tohme J (2000) AFLP analysis of African cassava (Manihot esculenta Crantz) germplasm resistant to the cassava mosaic disease (CMD). Theor Appl Genet 100:678–685
Fregene M, Matsumura H, Akano A, Dixon A, Terauchi R (2004) Serial analysis of gene expression (SAGE) of host–plant resistance to the cassava mosaic disease (CMD). Plant Mol Biol 56(4):563–571
Fregene M, Ospina JA, Roca W (1999) Recovery of cassava (Manihot esculenta Crantz) plants from culture of immature zygotic embryos. Plant Cell Tissue Organ Cult 55:39–43
Gaitán-Solís E, Taylor NJ, Siritunga D, Stevens W, Schachtman DP (2015) Overexpression of the transporters AtZIP1 and AtMTP1 in cassava changes zinc accumulation and partitioning. Front Plant Sci 6:492. https://doi.org/10.3389/fpls.2015.00492
Gallais A, Bordes J (2007) The use of doubled haploids in recurrent selection and hybrid development in maize. Crop Sci 47:S191–S200
Gardner CO (1961) An evaluation of effects of mass selection and seed irradiation with thermal neutrons on yields of corn. Crop Sci 1:241–245
Gonçalves Fukuda WM, de Oliveira, Silva S, Iglesias C (2002) Cassava breeding. Crop Breed Appl Biotech 2(4):617–638
Gonçalvez Fukuda WM, Saad N (2001) Participatory research in cassava breeding with farmers in Northeastern Brazil. Document CNPMF 99. EMBRAPA, Cruz das Almas. Bahia
Graner EA (1935) Contribuição para o estudo citológico da mandioca. Escola Superior de Agricultura Luiz de Queiroz, Piracicaba, pp 1–28
Graner EA (1942) Genética da Manihot. I Hereditariedade da forma da folha e da coloração da película externa das Raízes em Manihot utilíssima. Pohl Bragantia 2:13–22
Grüneberg W, Mwanga R, Andrade M, Espinoza J (2009) Breeding clonally propagated crops. In: Ceccarelli S, Guimarães EP, Weltzien E (eds) Plant breeding and farmer participation. Food and Agriculture Organization of the United Nations, Rome, pp 175–322
Hahn SK, Bai KV, Asiedu R (1990) Tetraploids, triploids, and 2n pollen from diploid interspecific crosses with cassava. Theor Appl Genet 79:433–439
Hahn SK, Terry ER, Leuschner K, Akobundu IO, Okali C, Lal R (1979) Cassava improvement in Africa. Field Crops Res 2:193–226
Hallauer AR, Miranda Fo JB (1981) Quantitative genetics in maize breeding, 2nd edn. Iowa State University Press, Ames
Heffner EL, Sorrells ME, Jannink J-L (2009) Genomic selection for crop improvement. Crop Sci 49:1–12
Hershey CH. 2008. A global conservation strategy for cassava (Manihot esculenta) and wild Manihot species (https://www.croptrust.org/wp-content/uploads/2014/12/cassava-strategy.pdf)
Hershey CH, Ocampo-N CH (1989) New marker genes found in cassava. Cassava Newsl 13(1):1–5
Heslot N, Yang H-P, Sorrells ME, Jannink J-L (2012) Genomic selection in plant breeding: a comparison of models. Crop Sci 52:146–160
Howeler RH, Maung Aye T (2014) Sustainable management of cassava in Asia: from research to practice. International Centre for Tropical Agriculture (CIAT); The Nippon Foundation, Vietnam. 168 p. (http://hdl.handle.net/10568/65172)
Hussain A, Brushuk W, Ramírez H, Roca W (1987) Identification of cassava (Manihot esculenta Crantz) cultivars by eletrophoretic patterns of esterase isozymes. Seed Sci Technol 15:19–22
IBGE (2014) Instituto Brasileiro de Geografia e Estatística (http://www.ibge.gov.br/). Produção Agrícola Municipal
Ibrahim AB, Heredia FF, Pinheiro CB, Aragao FJL, Campos FAP (2008) Optimization of somatic embryogenesis and selection regimes for particle bombardment of friable embryogenic callus and somatic cotyledons of cassava (Manihot esculenta Crantz). Afr J Biotechnol 7:2790–2797
Iglesias C, Mayer J, Chávez AL, Calle F (1997) Genetic potential and stability of carotene content in cassava roots. Euphytica 94:367–373
Ihemere U, Arias-Garzon D, Lawrence S, Sayre R (2006) Genetic modification of cassava for enhanced starch production. Plant Biotechnol J 4:453–465
Jennings DL (1957) Further studies in breeding cassava for virus resistance. E Afr Agric J 22:213–219
Jennings DL (1963) Variation in pollen and ovule fertility in varieties of cassava, and the effect of interspecific crossing on fertility. Euphytica 12:69–76
Jennings DL, Hershey CH (1985) Cassava breeding: a decade of progress from international programmes. In: Russell GE (ed) Progress in plant breeding-1. Butterworths, London, pp 89–116
Jennings DL, Iglesias CA (2002) Breeding for crop improvement. In: Hillocks RJ, Tresh JM, Bellotti AC (eds) Cassava: biology, production and utilization. CABI Publishing, Wallingford, pp 149–166
Joaqui BO, Lenis JI, Calle F et al (2016) Cassava breeding II: phenotypic correlations through the different stages of selection. Front Plant Sci 7:1649. (in press)
Jørgensen K, Bak S, Busk PK, Sørensen C, Olsen CE, Puonti- Kaerlas J, Møller BL (2005) Cassava plants with a depleted cyanogenic glucoside content in leaves and tubers. Distribution of cyanogenic glucosides, their site of synthesis and transport, and blockage of the biosynthesis by RNA interference technology. Plant Physiol 139:363–374
Jos JS, Bai KV (1981) Functional male sterility in cassava. Curr Sci (Banglare) 50:1035–1036
Jos JS, Hrishi N (1976) Inheritance of leaf shape in cassava. J Root Crop 2(2):10–12
Jos, Nair (1984) Genetics of male sterility in a genotype of cassava. Curr Sci (Bangalore) 53:494–496
Kamau J, Melis R, Laing M, Derera J, Shanahan P, Ngugi E (2010) Combining the yield ability and secondary traits of selected cassava genotypes in the semi-arid areas of Eastern Kenya. J Plant Breed Crop Sci 2(7):181–191
Kamau J, Melis R, Laing M, Derera J, Shanahan P, Ngugi ECK (2011) Farmers’ participatory selection for early bulking cassava genotypes in semi-arid Eastern Kenya. J Plant Breed Crop Sci 3:44–52
Karlström A, Calle F, Salazar S, Morante N, Dufour D, Ceballos H (2016) Biological implications in cassava for the production of amylose-free starch: impact on root yield and related traits. Front Plant Sci 7:604. doi:10.3389/fpls.2016.00604
Kawano K (1980) Cassava. In: Fehr WR, Hadley HH (eds) Hybridization of crop plants. ASA, CSSA, Madison, pp 225–233
Kawano K (2003) Thirty years of cassava breeding for productivity – biological and social factors for success. Crop Sci 43:1325–1335
Kawano K, Amaya A, Daza P, Ríos M (1978) Factors affecting efficiency of hybridization and selection in cassava (Manihot esculenta Crantz). Crop Sci 18:373–376
Kawano K, Cock JH (2005) Breeding cassava for underprivileged: institutional, socio-economic and biological factors for success. J Crop Improv 14:197–219
Kawano K, Goncalves Fukuda WM, Cenpukdee U (1987) Genetic and environmental effects on dry matter content of cassava root. Crop Sci 27:69–74
Kawano K, Narintaraporn K, Narintaraporn P et al (1998) Yield improvement in a multistage breeding program for cassava. Crop Sci 38:325–332
Kaweesi T, Kawuki R, Kyaligonza V, Baguma Y, Tusiime G, Ferguson ME (2014) Field evaluation of selected cassava genotypes for cassava brown streak disease based on symptom expression and virus load. Virol J 11:216. doi:10.1186/s12985-014-0216-x
Kaweesi T, Kyaligonza V, Baguma Y, Kawuki R, Ferguson M (2016) Inbreeding enhances field resistance to cassava brown streak viruses. J Plant Breed Crop Sci 8:138–149
Kawuki R, Ferguson M, Labuschagne M, Herselman L, Kim DJ (2009) Identification, characterisation and application of single nucleotide polymorphisms for diversity assessment in cassava (Manihot esculenta Crantz). Mol Breed 23:669–684
Kawuki RS, Nuwamanya E, Labuschagne MT, Herselman L, Ferguson ME (2011) Segregation of selected agronomic traits in six S1 cassava families. J Plant Breed Crop Sci 3:154–160
Kawuki RS, Pariyo A, Amuge T et al (2011) A breeding scheme for local adoption of cassava (Manihot esculenta Crantz). J Plant Breed Crop Sci 3:120–130
Keating B (1982) Environmental effects on growth and development of cassava (Manihot esculenta Crantz) with special reference to photoperiod and temperature. Cassava News 1(10):10–12
Kemp BP, Beeching JR, Cooper RM (2005) cDNA-AFLP reveals genes differentially expressed during the hypersensitive response of cassava. Mol Plant Pathol 6:113–123
Kemp BP, Horne J, Bryant A, Cooper RM (2004) Xanthomonas axonopodis gumD gene is essential for EPS production and pathogenicity and enhances epiphytic survival on cassava (Manihot esculenta). Physiol Mol Plant Pathol 64:209–218
Kizito E, Rönnberg-Wästljung AC, Egwang T, Gullberg U, Fregene M, Westerbergh A (2007) Quantitative trait loci controlling cyanogenic glucoside and dry matter content in cassava (Manihot esculenta Crantz) roots. Hereditas 144:129–136
Koehorst-van Putten HJJ, Sudarmonowati E, Herman M, Pereira-Bertram IJ, Wolters AMA, Meima H et al (2012) Field testing and exploitation of genetically modified cassava with low-amylose or amylose-free starch in Indonesia. Transgenic Res 21:39e50
Kunkeaw S, Yoocha T, Sraphet S et al (2011) Construction of a genetic linkage map using simple sequence repeat markers from expressed sequence tags for cassava (Manihot esculenta Crantz). Mol Breed 27:67–75
Lancaster PA, Brooks JE (1983) Cassava leaves as human food. Econ Bot 37:331–348
Lefèvre E, Charrier A (1993) Heredity of seventeen isozyme loci in cassava (Manihot esculenta Crantz). Euphytica 66:171–178
Legg JP, Jeremiah SC, Obiero HM et al (2011) Comparing the regional epidemiology of the cassava mosaic and cassava brown streak virus pandemics in Africa. Virus Res 159:161–170
Leyva-Guerrero E, Narayanan NN, Ihemere U, Sayre RT (2012) Iron and protein biofortification of cassava: lessons learned. Curr Opin Biotechnol 23:257–264
Li HQ, Sautter C, Potrykus I, Puonti-Kaerlas J (1996) Genetic transformation of cassava (Manihot esculenta Crantz). Nat Biotechnol 14:736–740
Liang H, Ren J, Gao Z, Gao S, Luo X, Dong L, Scipioni A (2016) Identification of critical success factors for sustainable development of biofuel industry in China based on grey decision-making trial and evaluation laboratory (DEMATEL). J Clean Prod 131:500–508
Liu J, Zheng Q, Ma Q, Gadidasu KK, Zhang P (2011) Cassava genetic transformation and its application in breeding. J Integr Plant Biol 53:552–569
Lokko Y, Dixon AGO, Offei SK, Danquah EY (2006) Combining ability analysis of resistance to mosaic virus disease in cassava. Afr Crop Sci J 14(3):221–230
Lokko Y, Dixon A, Offei S, Danquah E, Fregene M (2006) Assessment of genetic diversity among African cassava Manihot esculenta Crantz accessions resistant to the cassava mosaic virus disease using SSR markers. Genet Resour Crop Evol 53:1441–1453
Long SP, Ainsworth EA, Leakey ADB, Nösberger J, Ort DR (2006) Food for thought: lower-than-expected crop yield stimulation with rising CO2 concentrations. Science 312:1918–1921
Long SP, Ainsworth EA, Rogers A, Ort DR (2004) Rising atmospheric carbon dioxide: plants face the future. Annu Rev Plant Biol 55:591–628
Lozano JC, Cock JH, Castaño J (1978) New developments in cassava storage. In: Brekelbaum T, Bellotti A, Lozano JC (eds) Proceedings of cassava protection workshop. CIAT, Cali, pp 135–141
Ly D, Hamblin M, Rabbi I et al (2013) Relatedness and genotype x environment interaction affect prediction accuracies in genomic selection: a study in cassava. Crop Sci 53:1312–1325
Magoon ML, Krishnan R, Bai KV (1969) Morphology of the pachytene chromosomes and meiosis in Manihot esculenta Crantz. Cytologia 34:612–626
Manu-Aduening JA, Lamboll RI, Ampong Mensah G, Lamptey JN, Moses E, Dankyi AA, Gibson RW (2006) Development of superior cassava cultivars in Ghana by farmers and scientists: the process adopted, outcomes and contributions and changed roles of different stakeholders. Euphytica 150:47–61
Maredia MK, Reyes BA, Manu-Aduening J et al (2016) Testing alternative methods of varietal identification using DNA fingerprinting: results of pilot studies in Ghana and Zambia. MSU International Development Working Paper No. 149. 36 p
Marmey P, Beeching J, Hamon S, Charrier A (1993) Evaluation of cassava (Manihot esculenta Crantz.) germplasm using RAPD markers. Euphytica 74:203–209
Maruthi MN, Bouvaine S, Tufan HA, Mohammed IU, Hillocks RJ (2014) Transcriptional response of virus-infected cassava and identification of putative sources of resistance for cassava brown streak disease. PLoS One. doi:10.1371/journal.pone.0096642
Mkumbira J, Chiwona-Karltun L, Lagercrantz U et al (2003) Classification of cassava into ‘bitter’ and ‘cool’ in Malawi: from farmers’ perception to characterisation by molecular markers. Euphytica 132:7–22
Molo R, Aool W, Adumo S, Mutisya DL (2016) Integrating cassava varieties and Typhlodramulus aripo to sustain biological control of cassava green mite. Afr Crop Sci J 24(s1):117–126
Monger WA, Seal S, Cotton S, Foster GD (2001) Identification of different isolates of Cassava brown
Mongomake K, Doungous O, Khatabi B, Fondong VN (2015) Somatic embryogenesis and plant regeneration of cassava (Manihot esculenta Crantz) landraces from Cameroon. SpringerPlus 4:477. doi:10.1186/s40064-015-1272-4
Morante N, Ceballos C, Sánchez T et al (2016) Discovery of new spontaneous sources of amylose-free cassava starch and analysis of their structure and techno-functional properties. Food Hydrocoll 56:383–395
Morante N, Sánchez T, Ceballos H et al (2010) Tolerance to post-harvest physiological deterioration in cassava roots. Crop Sci 50:1333–1338
Morillo-C Y, Sánchez T, Morante N, Chávez AL, Morillo-C AC, Bolaños A, Ceballos H (2012) Estudio preliminar de herencia del contenido de carotenoides en raíces de poblaciones segregantes de yuca (Manihot esculenta Crantz). Acta Agron 61(3):253–264
Moyib KO, Mkumbira J, Odunola OA, Dixon AG, Akoroda MO, Kulakow P (2015) Genetic variation of postharvest physiological deterioration susceptibility in a cassava germplasm. Crop Sci 55:2701–2711
Moyib OK, Odunola OA, Dixon AGO (2007) SSR markers reveal genetic variation between improved cassava cultivars and landraces within a collection of Nigerian cassava germplasm. Afr J Biotechnol 6:2666–2674
Muoki PN, Maziya-Dixon B (2010) Household utilization of manioc (Manihot esculenta Crantz) in northern Mozambique. Ecol Food Nutr 49:337–356
Nassar NMA (1978) Conservation of the genetic resources of cassava (Manihot esculenta): determination of wild species localitities with emphasis on probably origin. Econ Bot 32:311–320
Nassar NMA (2000) Cytogenetics and evolution of cassava (Manihot esculenta Crantz). Genet Mol Biol 23:1003–1014
Nassar NMA, Ortiz R (2008) Cassava genetic resources: manipulation for crop improvement. Plant Breed Rev 31:247–275
Nguyen TLT, Gheewala SH, Garivait S (2007) Full chain energy analysis of fuel ethanol from cassava in Thailand. Environ Sci Technol 41:4135–4142
Nichols RFW (1947) Breeding cassava for virus resistance. East Afr Agric J 12:184–194
Njenga P, Edema R, Kamay J (2014) Combining ability for beta-carotene and important quantitative traits in a cassava F1 population. J Plant Breed Crop Sci 6(2):24–30
Njoku DN, Gracen VE, Offei SK, Asante IK, Egesi C, Kulakow P, Ceballos H (2015) Parent-offspring regression analysis for total carotenoids and some agronomic traits in cassava. Euphytica. doi:10.1007/s10681-015-1482-4
Njoku DN, Ikeogu UN, Ewa F, Egesi C (2015) Crossability and germinability potentials of some cassava (Manihot esculenta Crantz) progenitors for selection. J Plant Breed Crop Sci 7(3):61–66
Normanha ES (1970) Cassava breeding work at the São Paulo State. Agronomic Institute, Campinas, Brazil. In: Trabalhos do Encontro de Engenheiros Agrônomos do Estado de São Paulo, Brazil. pp 40–47
Ntui VO, Kong K, Khan RS, Igawa T, Janavi GJ, Rabindran R et al (2015) Resistance to Sri Lankan cassava mosaic virus (SLCMV) in genetically engineered cassava cv. KU50 through RNA silencing. PLoS One 10(4):e0120551. doi:10.1371/journal.pone.0120551
Nweke F (2004) New challenges in the cassava transformation in Nigeria and Ghana, EPT Discussion Paper, vol 118. International Food Policy Research Institute (IFPRI), Washington, DC
Odipio J, Alicai T, Nusinow D, Bart R, Ingelbrecht I, Taylor N (2016) CRISPR/CAS9 genome editing of putative cassava flowering genes. Proceedings of the World Congress on Root and Tuber Crops and Third Scientific Conference of the Global Cassava Partnership for the 21st Century. Nanning, China. Jan 2016
Odipio J, Ogwok E, Taylor NJ, Halsey M, Bua A, Fauquet CM, Alicai T (2014) RNAi-derived field resistance to cassava brown streak disease persists across the vegetative cropping cycle. GM Crops Food 5:16–19
Okechukwu RU, Dixon AGO (2009) Performance of improved cassava genotypes for early bulking, disease resistance, and culinary qualities in an Inland Valley ecosystem. Agron J 101:1258–1265
Okogbenin E, Egesi CN, Olasanmi B et al (2012) Molecular marker analysis and validation of resistance to cassava mosaic disease in elite cassava genotypes in Nigeria. Crop Sci 52:2576–2586
Okogbenin E, Fregene M (2002) Genetic analysis and QTL mapping of early bulking in an F1 segregating population from non-inbred parents in cassava (Manihot esculenta Crantz). Theor Appl Genet 106:58–66
Okogbenin E, Fregene M (2003) Genetic mapping of QTL affecting productivity and plant architecture in a full-sib cross from non-inbred parents in cassava (Manihot esculenta Crantz). Theor Appl Genet 107:1452–1462
Okogbenin E, Porto MCM, Dixon AGO (1998) Influence of planting season on incidence and severity of African cassava mosaic disease in the subhumic zone of Nigeria. In: Akoroda MO, Ekanayake IJ (eds) Root crops and poverty alleviation. International Institute of Tropical Agriculture (IITA), Ibadan, pp 388–392
Okogbenin E, Porto MCM, Egesi C et al (2007) Marker aided introgression of CMD resistance in Latin American germplasm for genetic improvement of cassava in Africa. Crop Sci 47:1895–1904
Olasanmi B, Akoroda MO, Okogbenin E et al (2014) Bulked segregant analysis identifies molecular markers associated with early bulking in cassava (Manihot esculenta Crantz). Euphytica 195(2):235–244
Olsen KM, Schaal BA (1999) Evidence on the origin of cassava: phylogeography of Manihot esculenta. Proc Natl Acad Sci USA 96:5586–5591
Olsen KM, Schaal BA (2001) Microsatellite variation in cassava (Manihot esculenta, Euphorbiaceae) and its wild relatives: further evidence for a southern Amazonian origin of domestication. Am J Bot 88(1):131–142
Orrego JI, Hershey CH (1984) Almacenamiento del pólen de yuca (Manihot esculenta Crantz) por medio de liofilización y varios regímenes de humedad y temperatura. Acta Agron 34(1):21–25
Otti G, Fakoya A, Andrew I, Gedil M (2011) Development of genomic tools for verification of hybrids and selfed progenies in cassava (Manihot esculenta). Afr J Biotechnol 10:17400–17408
Owolade OF, Dixon AGO, Adeoti AYA (2006) Diallel analysis of cassava genotypes to anthracnose disease. World J Agric Sci 2:98–104
Parkes EY, Fregene M, Dixon A, Peprah BB, Labuschagne MT (2013) Combining ability of cassava genotypes for cassava mosaic disease and cassava bacterial blight, yield and its related components in two ecological zones in Ghana. Euphytica 194:13–24
Patil BL, Ogwok E, Wagaba H et al (2011) RNAi-mediated resistance to diverse isolates belonging to two virus species involved in cassava brown streak disease. Mol Plant Pathol 12(1):31–41
Perera PIP, Ordoñez CA, Becerra López-Lavalle LA, Dedicova B (2013) A milestone in the doubled haploid pathway of cassava. Protoplasma 251:233–246
Perera PIP, Dedicova B, Ordoñez C, Kularatne JDJS, Quintero M, Ceballos H (2012) Recent advances in androgenesis induction of cassava (Manihot esculenta Crantz). Acta Hortic 961:319–325
Pérez JC, Ceballos H, Calle F, Morante N, Gaitán W, Llano G, Alvarez E (2005) Within-family genetic variation and epistasis in cassava (Manihot esculenta Crantz) adapted to the acid-soils environment. Euphytica 145(1–2):77–85
Pérez JC, Ceballos H, Jaramillo G, Morante N, Calle F, Arias B, Bellotti AC (2005) Epistasis in cassava adapted to mid-altitude valley environments. Crop Sci 45:1491–1496
Pérez JC, Ceballos H, Ramirez IC, Lenis JI, Calle F, Morante N, Lentini M. del C. (2010) Adjustment for missing plants in cassava evaluation trials. Euphytica 172(1):59–65
Prakash A (2008) Cassava: international market profile. Background paper for the Competitive Commercial Agriculture in Sub–Saharan Africa (CCAA) Study. Trade and Markets Division. Food and Agriculture Organisation of the United Nations, Rome
Prochnik S, Marri PR, Desany B et al (2012) The cassava genome: current progress, future directions. Trop Plant Biol 5:88–94
Prudencio CY, Orkwor GC, Kissiedu AFK (1992) The relationships between cassava variety set characteristics, farmers’ food security objectives, environmental and socio-economic conditions in Africa. Agric Syst 39:387–408
Rabbi I, Hamblin M, Gedil M, Kulakow P, Ferguson M, Ikpan AS, Ly D, Jannink J-L (2014) Genetic mapping using genotyping-by-sequencing in the clonally propagated cassava. Crop Sci 54:1384–1396
Rabbi IY, Hamblin MT, Lava Kumar P, Gedil MA, Ikpan AS, Jannink J-L, Kulakow PA (2014) High-resolution mapping of resistance to cassava mosaic geminiviruses in cassava using genotyping by sequencing and its implications for breeding. Virus Res 186:87–96
Rabbi I, Kulakow P, Manu-Aduening J et al (2015) Tracking crop varieties using genotyping-by-sequencing markers: a case study using cassava (Manihot esculenta Crantz). BMC Genet 16:115. doi:10.1186/s12863-015-0273-1
Raemakers K, Schreuder M, Pereira I, Munyikwa T, Jacobsen E, Visser R (2001) Progress made in FEC transformation of cassava. Euphytica 120:15–24
Raemakers CJJM, Sofiari E, Taylor NJ, Henshaw GG, Jacobsen E, Visser RGF (1996) Production of transgenic cassava plants by particle bombardment using luciferase activity as the selection marker. Mol Breed 2:339–349
Ramírez H, Hussain A, Roca W, Bushuk W (1987) Isozyme electrophoregrams of sixteen enzynes in five tissues of cassava (Manihot esculenta Crantz) varieties. Euphytica 36:39–48
Reilly K, Bernal D, Cortes DF, Gomez-Vasquez R, Tohme J, Beeching JR (2007) Towards identifying the full set of genes expressed during cassava post-harvest physiological deterioration. Plant Mol Biol 64:187–203
Renvioze BS (1973) The area of origin of Manihot esculenta as a crop plant: a review of the evidence. Econ Bot 26:352–360
Restrepo S, Verdier V (1997) Geographical differentiation of the population of Xanthomonas axonopodis pv. manihotis in Colombia. Appl Environ Microbiol 63:4427–4434
Roa AC, Maya MM, Duque MC, Tohme J, Allem AC, Bonierbale MW (1997) AFLP analysis of relationships among cassava and other Manihot species. Theor Appl Genet 95:741–750
Roca WM (1984) Cassava. In: Sharp WR, Evans DA, Ammirato PV, Yamada Y (eds) Handbook of plant cell culture. Crop species. Macmillan, New York, pp 269–301
Rogers DJ, Appan SG (1973) Manihot and Manihotoides (Euphorbiaceae), a computer-assisted study, Flora Neotropica, monograph, vol 13. Hafner Press, New York. 272 p
Rojas-C M, Pérez JC, Ceballos H, Baena D, Morante N, Calle F (2009) Introduction of inbreeding and analysis of inbreeding depression in eight S1 cassava families. Crop Sci 49:543–548
RTB (2016) The CGIAR Research Program on Roots, Tubers and Bananas (RTB) 2016 Annual Report. www.cgiar-rtb-2016-annual-report
Sánchez T, Ceballos H, Dufour D et al (2014) Carotenoids and dry matter prediction by NIRS and hunter color in fresh cassava roots. Food Chem 151:444–451
Sánchez T, Dufour D, Moreno JL, Pizarro M, Arango I, Domínquez M, Ceballos H (2013) Changes in extended shelf life of cassava roots during storage in ambient conditions. Postharvest Biol Technol 86:520–528
Sarria R, Torres E, Angel F, Chavarriaga P, Roca WM (2000) Transgenic plants of cassava (Manihot esculenta) with resistance to Basta obtained by Agrobacterium-mediated transformation. Plant Cell Rep 19:339–344
Sarria R, Torres E, Balcazar, M, Destafano-Beltran L, Roca WM (1995) Progress in Agrobacterium-mediated transformation of cassava (Manihot esculenta Crantz). In: Proceedings of the Second International Scientific Meeting Cassava Biotechnology Network (22–26 August 1994, Bogor, Indonesia) (Working document 150), CIAT, Cali. pp 241–244
Sayre R, Beeching J, Cahoon E et al (2011) The BioCassava plus program: Biofortification of cassava for sub-Saharan Africa. Annu Rev Plant Biol 62:251–272
Schöpke C, Taylor NJ, Carcamo R, Konan NK, Marmey P, Henshaw GG, Beachy RN, Fauquet CM (1996) Regeneration of transgenic cassava plants (Manihot esculenta Crantz) from microbombarded embryogenic suspension cultures. Nat Biotech 14:731–735
Second G, Allem A, Emperaire L et al (1997) AFLP based Manihot and cassava numerical taxanomy and genetic structure analysis in progress: implications for dynamic conservation and genetic mapping. Afr J Root Tuber Crops 2:140–147
Smith AM (2008) Prospects for increasing starch and sucrose yields for bioethanol production. Plant J 54:546–558
Sofiari E, Raemakers CJJM, Bergervoet JEM, Jacobsen E, RGF V (1998) Plant regeneration from protoplasts isolated from friable embryogenic callus of cassava. Plant Cell Rep 18:159–165
Sriroth K, Piyachomkwan K, Wanlapatit S, Nivitchanyong S (2010) The promise of a technology revolution in cassava bioethanol: from Thai practice to the world practice. Fuel 89:1333–1338
Ssemakula G, Dixon A (2007) Genotype x environment interaction, stability and agronomic performance of carotenoid-rich cassava clones. Sci Res Essay 2(9):390–399
Stapleton G (2012) Global starch market outlook and competing starch raw materials for starches by product segment and region. Cassava Starch World 2010. Centre for Management Technology (CMT), Cambodia
Taylor N, Chavarriaga P, Raemakers K, Siritunga D, Zhang P (2004) Development and application of transgenic technologies in cassava. Plant Mol Biol 56:671–688
Taylor N, Gaitán-Solís E, Moll T et al (2012) A high-throughput platform for the production and analysis of transgenic cassava (Manihot esculenta) plants. Trop Plant Biol 5:127–139
Taylor NJ, Masona MV, Carcamo R, Ho T, Schöpke C, Fauquet CM (2001) Production of embryogenic tissues and regeneration of transgenic plants in cassava (Manihot esculenta Crantz). Euphytica 120:25–34
Troyer AF (2006) Adaptedness and heterosis in corn and mule hybrids. Crop Sci 46:528–543
Tumuhimbise R, Shanahan P, Melis R, Kawuki R (2014) Genetic variation and association among factors influencing storage root bulking in cassava. J Agric Sci 153:1267–1280
Tuvesson S, Dayteg C, Hagberg P et al (2007) Molecular markers and doubled haploids in European plant breeding programs. Euphytica 158:305–312
Umanah EE, Hartmann CRW (1973) Chromosome numbers and karyotypes of some Manihot species. J Am Soc Hortic Sci 98(3):272–274
van Oirschot QEA, O’Brien GM, Dufour D, El-Sharkawy MA, Mesa E (2000) The effect of pre-harvest pruning of cassava upon root deterioration and quality characteristics. J Sci Food Agr 80:1866–1873
Vanderschuren H, Moreno I, Anjanappa RB, Zainuddin IM, Gruissem W (2012) Exploiting the combination of natural and genetically engineered resistance to cassava mosaic and cassava brown streak viruses impacting cassava production in Africa. PLoS One 7:e45277
Vanderschuren H, Stupak M, Fütterer J, Gruissem W, Zhang P (2007) Engineering resistance to geminiviruses––review and perspectives. Plant Biotechnol J 5:207–220
Vlaar PWL, van Beek P, RGF V (2007) Genetic modification and its impact on industry structure and performance: post-harvest deterioration of cassava in Thailand. J Chain Netw Sci 7:133–142
Wang C, Lentini Z, Tabares E et al (2011) Microsporogenesis and pollen formation in cassava (Manihot esculenta Crantz). Biol Plant 55(3):469–478
Wasswa P, Alicai T, Mukasa SB (2010) Optimisation of in vitro techniques for cassava brown streak virus elimination from infected cassava clones. Afr Crop Sci J 18:235–241
Welsch R, Arango J, Bär C et al (2010) Provitamin a – accumulation in cassava (Manihot esculenta) roots driven by a single nucleotide polymorphism in a phytoene synthase gene. Plant Cell 22:3348–3356
Were WV, Shanahan P, Melis R, Omari OO (2012) Gene action controlling farmer preferred traits in cassava varieties adapted to mid-altitude tropical climatic conditions of western Kenya. Field Crop Res 133:113–118
Westwood NN (1990) Maintenance and storage: clonal germplasm. Plant Breed Rev 7:111–128
Whankaew S, Poopear S, Kanjanawattanawong S, Tangphatsornruang S, Boonseng O, Lightfoot DA, Triwitayakorn K (2011) A genome scan for quantitative trait loci affecting cyanogenic potential of cassava root in an outbred population. BMC Genomics 12:266. http://www.biomedcentral.com/1471-2164/12/266
Wolfe MD, Egesi C, Kawuki R, Kulakow P, Rabbi IY, Jannink J-L (2016) Next generation cassava breeding: progress implementing genomic selection” in Proceedings of the World Congress on Root and Tuber Crops and Third Scientific Conference of the Global Cassava Partnership for the 21st Century (Nanning). Available online at: http://www.gcp21.org/wcrtc/PS08/
Wolfe MD, Rabbi IY, Egesi C et al (2016) Genomewide association and prediction reveals genetic architecture of cassava mosaic disease resistance and prospects for rapid genetic improvement. Plant Genome 9. doi:10.3835/plantgenome2015.11.0118
Wongtiem P, Courtois D, Florin B, Juchaux M, Peltier D, Broun P, Ducos JP (2011) Effects of cytokinins on secondary somatic embryogenesis of selected clone Rayong 9 of Manihot esculenta Crantz for ethanol production. Afr J Biotechnol 10:1600–1608
Woodward B, Puonti-Kaerlas J (2001) Somatic embryogenesis from floral tissue of cassava (Manihot esculenta Crantz). Euphytica 120:1–6
Wright CE (1965) Field plans for a systematically designed polycross. Rec Agric Res 14:31–41
Wydra K, Zinsou V, Jorge V, Verdier V (2004) Identification of pathotypes of Xanthomonas axonopodis pv. manihotis in Africa and detection of quantitative trait loci and markers for resistance to bacterial blight of cassava. Phytopathology 94:1084–1093
Xu Y, Crouch JH (2008) Marker-assisted selection in plant breeding: from publications to practice. Crop Sci 48:391–407
Yadav JS, Ogwok E, Wagaba H et al (2011) RNAi-mediated resistance to cassava brown streak Uganda virus in transgenic cassava. Mol Plant Pathol. doi:10.1111/j.1364-3703.2010.00700.x
Zacarias AM, Botha A-M, Labuschagne MT, Benesi IRM (2004) Characterization and genetic distance analysis of cassava (Manihot esculenta Crantz) germplasm from Mozambique using RAPD fingerprinting. Euphytica 138:49–53
Zacarias AM, Labuschagne MT (2010) Diallel analysis of cassava brown streak disease, yield and yield related characteristics in Mozambique. Euphytica 176:309–320
Zainuddin IM, Schlegel K, Gruissem W, Vanderschuren H (2012) Robust transformation procedure for the production of transgenic farmer-preferred cassava landraces. Plant Methods 8:24. http://www.plantmethods.com/content/8/1/24
Zehntner L (1919) Estudo sobre algumas variedades de mandiocas Brasileiras. Sociedad Nacionale de Agricultura, Impresa Ingleza-Camerino 61, Rio de Janeiro
Zhang P, Vanderschuren H, Fütterer J, Gruissem W (2005) Resistance to cassava mosaic disease in transgenic cassava expressing antisense RNAs targeting virus replication genes. Plant Biotechnol J 3:385–397
Zhang P, Wang WQ, Zhang GL, Kaminek M, Dobrev P, Xu J, Gruissem W (2010) Senescence-inducible expression of isopentenyl transferase extends leaf life, increases drought stress resistance and alters cytokinin metabolism in cassava. J Integr Plant Biol 52:653–669
Zhao SS, Dufour D, Sánchez T, Ceballos H, Zhang P (2011) Development of waxy cassava with different biological and physico-chemical characteristics of starches for industrial applications. Biotechnol Bioeng. doi:10.1002/bit.23120
Zhu X-G, de Sturler E, Long SP (2007) Optimizing the distribution of resources between enzymes of carbon metabolism can dramatically increase photosynthetic rate: a numerical simulation using an evolutionary algorithm. Plant Physiol 145:513–526
Zhu X-G, Long SP, Ort DR (2010) Improving photosynthetic efficiency for greater yield. Annu Rev Plant Biol 61:235–261
Zidenga T, Leyva-Guerrero E, Moon H, Siritunga D, Sayre R (2012) Extending cassava root shelf life via reduction of reactive oxygen species production1[c][w][OA]. Plant Physiol 4:1396–1407
Acknowledgments
This research was undertaken as part of the CGIAR Research Program on Roots, Tubers and Banana (RTB) and HarvestPlus, part of the CGIAR Research Program on Agriculture for Nutrition and Health (A4NH). It has also received financial support from the Bill and Melinda Gates Foundation and USAID.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Copyright information
© 2017 Springer International Publishing AG
About this chapter

Cite this chapter
Ceballos, H., Hershey, C.H. (2017). Cassava (Manihot esculenta Crantz). In: Genetic Improvement of Tropical Crops. Springer, Cham. https://doi.org/10.1007/978-3-319-59819-2_5
Download citation
DOI: https://doi.org/10.1007/978-3-319-59819-2_5
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-59817-8
Online ISBN: 978-3-319-59819-2
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)