
Abstract
Surgical treatment of intramedullary spinal cord tumors (IMSCTs) comes with an inherent risk of postoperative neurologic complications. Intraoperative neurophysiologic monitoring (IONM), including evoked potentials transmitted in the tracts of the spinal cord and the mapping of those tracts, is an essential tool for the surgeon whose goal is thorough and safe resection. Somatosensory evoked potentials (SSEPs), transcranial myogenic and myelogenic motor evoked potentials (TcMEPs and D-waves, respectively) and dorsal column and corticospinal tract (CST) mapping are important adjuncts to the surgical treatment of IMSCTs as each improves sensory and motor functional outcomes and contributes to greater resection. In particular, multimodality monitoring and combining multimodality monitoring with mapping reduce morbidity. SSEPs and TcMEPs serve to protect the spinal cord in general, and also give feedback directly on the function of each respective pathway. D-wave monitoring, an additional test of the motor pathway, adds functional predictive value over TcMEPs alone. Spinal cord mapping aids the surgeon in localizing the sensory and motor tracts and mitigates the risk of injury to them. Dorsal column mapping, physiologically localizing the sulcus separating the left and right dorsal columns, identifies a low-risk trajectory into the spinal cord for access to the tumor. CST mapping aids in differentiating normal from tumor tissue, and allows the surgeon to localize, determine the proximity, or determine specific muscle targets of the CST. In this chapter on neurophysiological applications for IMSCT surgery, the practical aspects of spinal cord monitoring and spinal cord mapping are described to inform the surgical neurophysiologist on how best to assist the surgeon in achieving the safest and most complete surgical resection.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Intramedullary
- Astrocytoma
- Ependymoma
- Hemangioblastoma
- D-Waves
- SSEP
- TcMEP
- Dorsal Columns
- Mapping
- Spinal Cord
- Corticospinal Tracts
- IONM
Surgery for IMSCT
Intramedullary spinal cord tumors (IMSCTs) are rare, often benign, lesions of the spinal cord and surgery continues to be the major option for treatment. The goals of surgery include satisfactory oncological resection to rid the patient of cancer while preserving the delicate neural structural and functional integrity of the spinal cord. Several factors play into the development of the surgical strategy and the choice of selective versus gross total resection. Of these, the tumor type, tumor grade, spinal location, number of spinal levels, and the patient’s preoperative neurologic status are among the most important [1,2,3,4,5]. As such, ependymoma and hemangioblastoma lesions, which comprise 45% and 5%, respectively, of the IMSCTs lend themselves to gross total resection especially if there is a clear plane of resection. Ependymomas occur more often in adults, are found centrally in a cross-section of the spinal cord, are more common in the lower spinal cord, are often encapsulated, are associated with cysts, and appear uniformly dense on imaging. Hemangioblastomas are the least prevalent of the most common IMSCTs. They arise from blood vessels and their resection may have a greater likelihood of neurophysiologic changes than ependymomas [6]. Astrocytomas , which comprise 40% of IMSCTs, are often treated with partial surgical resection because these tumors tend to be infiltrative with associated poorer prognosis. Astrocytomas are more common in children, tend to be eccentric in a cross-section of the spinal cord, can be found at all levels of the spinal cord, and most commonly occur at the thoracic levels. The remaining IMSCT types are rare. The strategy for any IMSCT tumor type may revert to sub-total resection if the intraoperative neurophysiologic monitoring (IONM) data indicate significant risk to spinal cord function.
For IMSCT surgery, a common approach is posteriorly. For this reason, patients will be operated in the prone position. On the occasion where the tumor is lateralized, the surgeon may choose to approach the spine from the side and the patient may be positioned in the lateral oblique position. For patients in the prone position, arms will be in one of two positions. For lesions at lower spinal levels, the “superman” position will be used, the upper arms perpendicular to the plane of the body from shoulder to elbow, and then 90 degrees again from that plane, back to parallel with the body from elbow to hand, with fingers resting palm-down on either side of the patient’s head. For lesions at or above the mid-thoracic level, the arms will be tucked along the plane of the body with the fingers pointing toward the feet and the thumbs down for ergonomic continuity. Extra care should be exercised when securing the electrodes to the arms, wrists, and hands as once the patient is positioned and the arms are tucked, the wrap will limit access to the placed electrodes should the need arise to replace or reposition them.
The spinal canal is exposed by laminectomy, and occasionally the strategy is to reconstitute the spine by laminoplasty. The laminectomy extends just superior and inferior to the expected extent of the solid portion of the lesion and may be extended if necessary. Through the window that is now opened into the spinal canal, the surgeon may choose to employ ultrasound to confirm tumor location.
This is followed by durotomy . Commonly, the dura is pried from the underlying spinal cord using micro forceps. A small hole is made through which the tip of a nerve hook is inserted, lifting the dura away from the spinal cord. Working along the long axis of the spinal cord, a scalpel incision is made and extended using the nerve hook as both a tool for lifting the dura and one against which force can be safely applied with the scalpel, thus protecting the spinal cord beneath it. The dura is opened long-wise and when opened resembles an open eye (where the open dural flaps on either side of the cord are the eyelids, and the spinal cord proper the globe of the eyeball). Traction is applied with dural sutures that are either anchored to the surrounding tissue or to a hemostat clamped to the distal end of the suture and dangled over the side of the patient. Once the dura is opened, the arachnoid is incised.
When D-wave recording is part of the monitoring strategy, after the dura is open and retracted, the surgeon will slide a spinal recording electrode epi- or subdurally under the lamina of spinal segments caudal to the surgical site (Fig. 13.1). Whenever practical, a second spinal recording electrode is worked rostrally in the same fashion. The insertion point of the spinal electrode may be gently irrigated to facilitate placement and to promote good contact. Epidural spinal electrodes can also be placed trans-dermally using a Touhy needle, but this is rarely necessary in open IMSCT surgery.

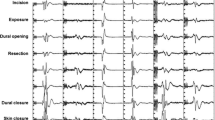
Intraoperative images through the microscope of various stages of surgery for a mid-cervical IMSCT . (a) Placement of the spinal electrode used for recording D-waves. In this figure, the dura is open and retracted using stitches, seen here in black, revealing the vascularized spinal cord in the center of the image. The spinal electrode, seen here with two of three contacts visible, is gently guided by the surgeon into the epidural space inferior to the location of the lesion. (b) Stimulation of the spinal cord with a monopolar probe during dorsal column mapping. (c) Midline dorsal myelotomy via scalpel. (d) Tumor retraction and tumor forceps. (e) Cavitron ultrasonic aspirator (CUSA)
D-waves are notoriously sensitive to electrode position, and spinal electrodes are prone to being displaced and dislodged. Maintaining the electrode in its original position throughout the salient surgical steps is critical for the reliability and interpretation of D-waves. Once the spinal electrode has been placed, its position may be fixed in place in several ways:
-
Stabilize the wire exiting the spinal electrode using the surgical drape clamped around the wire with a hemostat. Be sure to check that the delicate wire of the spinal electrode has not been clamped directly as this may cause interruption of the wire and/or its insulation, resulting in poor recordings.
-
Encourage the surgeon to suture around the epidural electrode as it exits the dura and affix it to the surrounding tissue.
-
Take note of the position and location of the electrode even if with a photograph. This can be aided by putting a fiducial mark on the epidural electrode as it exits the epidural space. Spinal electrodes often have fiducial marks of their own.
We encourage the neurophysiologist to attempt D-waves promptly after the placement and plug-in of the spinal electrodes. This gives time to troubleshoot the contact and placement of the spinal electrode(s) and optimize the stimulation parameters.
Once an optimized D-wave has been obtained and reported, spinal cord mapping may follow. The mechanics of the mapping depend on the method of choice (see below), but they include handoff of a sterile electrode required to stimulate directly or record directly from the spinal cord. Often, access into the spinal cord is at the posterior median sulcus which separates the left and right gracile fasciculi. Anatomically, the posterior median sulcus is at the midpoint between the dorsal root entry zones located at the lateral edges of a normal spinal cord. The visually estimated anatomical midline is compared with what is determined neurophysiologically to better approximate the location of this sulcus.
The spinal cord is opened by posterior midline myelotomy down the posterior median sulcus. This may be initiated directly by scalpel or by coagulation of dorsal midline veins and the superficial and medial aspect of the penetrating/diving fissure. The myelotomy is extended either with a scalpel or by splaying the cord via the outward pressure from the opening of closed micro forceps or the tips of the bipolar electrocautery device, with or without accompanying incising. Once the myelotomy is complete, some surgeons prefer to retract the dorsal columns either manually or with small sutures (such as pial traction sutures). Somatosensory evoked potentials (SSEPs , see below) are the focus of the IONM during these stages of the surgery. SSEP signal changes secondary to retraction forces applied on the dorsal columns indicate adjustments to retraction and/or the myelotomy are warranted. The surgeon should be encouraged to relax the tension on the tissue by either directly reducing retraction or indirectly by extending the myelotomy which allows the dorsal columns to open more freely.
Following the opening of the spinal cord, other types of mapping may ensue including localizing and even mapping the contents of the descending motor tracts through direct spinal cord stimulation. Selective or continuous mapping such as this may be incorporated throughout the resection.
Work on the tumor begins. A tissue specimen for biopsy is obtained using tumor forceps since the pathological identification of the tumor type is integral to the development of the surgical strategy and resection objectives. If there is a cystic component of the lesion, this may aid in identifying the leading and trailing ends of the tumor and help establish the resection plane. A cystic component of the tumor may make it difficult to obtain D-waves. While portions of a tumor that are clear and distinct from normal tissue may be removed by direct excision, tumors are typically debulked in an inside-out fashion. This is accomplished with the use of tumor forceps, electrocautery, or with a Cavitron Ultrasonic Surgical Aspirator (CUSA) . The CUSA uses high-frequency sound waves to morselize the tumor tissue, while preserving healthy tissue. These mobilized tumor fragments are suspended by irrigation fluids and aspirated away through the CUSA.
Working the marginal edges of the tumor and the tumor capsule from the surrounding nervous tissue is a critical stage of the surgery. Transcranial motor evoked potential (TcMEP) and D-wave monitoring is the focus of the IONM at this stage of the surgery. Tumor traction is often applied using the force generated by long-wise sweeps of the suction. Gentle counter traction on the spinal cord accompanied by isolation, division and electrocautery of adhesions and blood vessels are keys to successful excision.
At the completion of the tumor work, the dura is sutured closed. The surgeon will often request a Valsalva maneuver after the dura is closed to verify the integrity of the closure by looking for cerebrospinal fluid leaks during the period of higher intraspinal pressure temporarily created by the Valsalva. The epidural electrode(s) is/are likely to be removed at this point. Monitoring of D-waves should continue until the electrode(s) is/are removed. SSEP and TcMEP monitoring should continue until skin closure [7].
IONM in IMSCT Surgery
There are a number of published studies that represent the important contribution that spinal cord IONM and spinal cord mapping make to the surgical treatment of IMSCTs [7,8,9,10,11,12,13,14,15,16,17,18,19,20]. The monitoring modalities of SSEP and TcMEP, the most common IONM approaches during IMSCT surgery, and electromyography (EMG), a less commonly incorporated modality in IMSCT surgery, have been addressed in detail in other sections of this text. Therefore, the focus here will be on the addition of D-wave recording and on the methods of spinal cord mapping focusing on the dorsal columns and the corticospinal tracts (CSTs).
For IMSCT surgery, IONM including SSEP and TcMEP for monitoring spinal cord function should commence early. Patients with IMSCTs can be challenging neurophysiologically since they often have clinical or subclinical neurologic compromise. Pre-positioning baselines should be part of the IONM strategy for each of these cases. Post-position baselines without pre-position references introduce questions about the reason behind any incomplete or absent IONM data after positioning. Pre-position baselines also permit the surgical neurophysiologist the opportunity to evaluate and discuss the recordings and implement plausible adjustments to the IONM strategy to try to overcome deficiencies in data should they exist. Integrally, the surgical neurophysiologist must provide a thorough and detailed description of the pre- and post-positioning IONM recordings, as they should do throughout the procedure.
Somatosensory Evoked Potentials
For an in-depth view of SSEPs, I recommend the reader review Chap. 6 and the Association guidelines [21,22,23]. An SSEP is the bioelectric activity that originates from the nerves, tracts, and synapses along the ascending dorsal column-medial lemniscal (DCML) pathway . This pathway starts with the axons of dorsal root ganglia neurons that have terminal ends in the periphery. These axons, that carry fine touch, vibration, and proprioceptive information from sensory organs in the skin and from muscle spindles, course up the limb and into the lumbar plexus and lumbosacral plexus for axons of the lower limbs and the brachial plexus for axons of the upper limbs. They traverse the lateral neural foramen and pass into the spinal canal, transitioning from the peripheral to the central nervous systems. Each axon courses superiorly before approaching the spinal cord via the dorsal rootlets. Caudocranially, there is decreasing distance between the spinal level at which each axon enters the spinal column and the level at which it penetrates the spinal cord. Axons of the lower extremity course up the cauda equina, a distribution of nerve roots in the spinal canal ensheathed in the thecal sac, and enter the spinal cord at or just above the conus medullaris around spinal level L1.
After penetrating the spinal cord, the axons enter the posterior tracts, also known as the dorsal columns, and ascend the spinal cord toward the brainstem in distinct, somatotopically arranged, parallel tracts. The axons of the lower extremity course in the gracile fasciculi which bookend the posterior median sulcus. As you move up the spinal cord, axons from the trunk and then from the upper extremity systematically and successively join the dorsal columns laterally. In the cervical spine, the more lateral tracts, which are composed predominantly of the axons of the upper extremity, are the cuneate fasciculi.
In the direct, ascending pathway these axons make their first synapse in the gracile and cuneate dorsal column nuclei of the medulla. Axon collaterals also synapse in the spinal cord, participating in intraspinal neural circuits such as reflex arcs. The gracile and cuneate nuclei are independent regions of the medulla and maintain the separation of these parallel afferent pathways. Axonal projections from neurons in the gracile and cuneate nuclei decussate the brainstem in the internal arcuate of the medulla before ascending in the medial lemniscus toward the thalamus. Of note, a small percentage of patients have a non-decussating sensory pathway.
The ventral posterior nucleus of the thalamus is where the second synapse in the direct DCML pathway is found. The thalamus serves as a processing and relay center, and the pathway continues to the cortex from there. Thalamocortical projections make their way through the posterior limb of the internal capsule until they synapse on cortical pyramidal cells located primarily in the postcentral gyrus of the parietal lobe (the primary sensory cortex). Thalamocortical axons project to homonymous and somatotopically arranged cortical neurons: those axons corresponding to the lower extremity project medially along the interhemispheric medial bank and those axons corresponding to the upper extremity project laterally. The pathway continues through intracortical circuits of higher sensory processing.
SSEPs for spinal cord monitoring are initiated via transdermal electrical activation of peripheral nerves, typically as they come near the surface of the skin in the wrists and ankles. The ulnar nerve in the wrists and the tibial nerve in the ankles are a good choice for IMSCT surgery. In the case of a tumor situated high in a patient’s cervical spinal cord and informed by the patient’s neurologic presentation, adding median nerve SSEPs is recommended as they help to triangulate the location of an SSEP data change should one occur. Be prepared to activate the nerves of the popliteal fossa as well, since lower extremity SSEPs originating from the ankle may be difficult to resolve depending on the amount of neurologic functional compromise imposed by the tumor. Keep in mind this will result in greater patient movement, which can negatively impact the frequency at which they can be run. Popliteal fossa electrodes are also advantageous for recordings of antidromically transmitted action potential volleys elicited through stimulation of the spinal cord during dorsal column mapping (see below) and therefore can serve multiple purposes.
Nerves are activated with surface or, preferentially, with subdermal needle electrodes. At the ankle, the anode is placed behind the medial malleolus for tibial nerve activation. At the wrist, the anode is placed just proximal to the crease of the wrist and lateral to the flexor carpi ulnaris tendon for ulnar nerve activation and between the flexor carpi radialis and palmaris longus tendons for median nerve activation. The cathode is placed 2–3 cm proximal to the anode. For popliteal fossa activation, the anode is placed midway between the tendons of semitendinosus and biceps femoris above the crease behind the knee and the cathode is placed 2–5 cm proximal to the anode and 0–2 cm lateral to midline. Constant current, 200–500 μsec, square pulse, repeating stimuli are applied asymmetrically and alternately. Stimulation frequency, or repetition rate, is typically between 2 and 6 Hz, selecting values that de-harmonize with noise at the electrical line frequency. Higher repetition rates allow for faster acquisition of trials but simultaneously can result in degraded signal. Notwithstanding the longer acquisition time, it may be helpful to reduce the repetition rate, as lower stimulation frequencies may improve the resolution of poor signals. Signal acquisition sweep is typically 100 msec for lower extremity SSEPs and 50 msec for upper extremity SSEPs; however, increasing these time bases should be considered if the lesion has induced a latency delay. The intent of the acquisition sweep is to choose one that collects the major deflections of the response near the middle. Longer acquisition sweeps will consequently limit the high end of employable stimulation frequency as successive recording trials cannot overlap. Stimulus intensity, starting at 20 mA and 30 mA for upper and lower extremity , respectively, is ramped up, if necessary, until a supramaximal response is detected. Employing higher stimulus duration or intensity increases recruitment but consequently increases the likelihood of current spread inadvertently activating a nearby nerve(s), particularly when stimulating at the wrist. Therefore, a possible decrease in specificity must be considered when using higher stimulus intensities and durations.
SSEPs are small potentials and are often obscured by noise. To help resolve these potentials, we employ averaging. Starting at 200–500 trials is recommended, but the effective number of trials depends on the signal-to-noise ratio. At critical times in IMSCT surgery quick feedback is important, so fewer trials per average is advantageous.
Recordings of SSEPs are obtained at select locations on the body so as to maximize the likelihood of sampling the activity in a given portion of the pathway. Integrating information on the nature of the SSEP responses at each location and relative to those recorded earlier in the procedure helps to localize the source of signal change and/or decipher or discount systemic causes.
Peripheral recording sites include the popliteal fossa for lower extremity SSEPs and the supraclavicular fossa for upper extremity SSEPs. These sites capture predominantly high-frequency traveling waves coursing through the underlying nerves, and are intended, in large part, as a checkpoint to confirm activation of the pathway. Popliteal fossa recordings are captured with the reference electrode placed medially in the leg just above the crease behind the knee. In a bipolar montage the active electrode is 2–5 cm proximal and 0–2 cm lateral. In a referential montage the reference electrode is at a distant site. Erb’s point potentials are often captured at supraclavicular fossa recording sites using a referential montage where the left and right electrodes are referred to each other. A starting low-frequency filter (LFF) and high-frequency filter (HFF) of 30 and 1500 Hz, respectively, and an amplifier sensitivity of 20 μV/Div, works well to capture these high-frequency, often large, potentials. The obligate peak of the response obtained at these sites occurs at approximately 7–9 msec and is identified as N9. The latency of N9 and its amplitude relative to the trailing positive trough are tracked.
Recording sites near the neck and in the scalp are included to capture synaptic activity in subcortical structures such as the brainstem and thalamus and the traveling waves of the tracts therein and between. These are often referred to as subcortical potentials and are captured using a “subcort” or cervical electrode placed at any of several possible locations including the inion, at the neck over the C3, C5, or C7 spinous process, at the mastoid or at the chin with a reference electrode in the scalp usually at FPz. The same recording channel is typically used for both upper and lower extremity subcortical SSEPs. Using this montage, the subcortical potentials for lower extremity SSEPs are captured predominantly at the FPz electrode and the convention of negative potentials being upwardly deflected does not apply. Lower neck placement of the subcortical electrode is favored for resolving subcortical potentials particularly for lower extremity SSEPs. Lower neck placement may consequently capture responses from parenchymal generators, such as those that participate in spinal reflex arcs, for upper extremity SSEPs. Therefore, the surgical neurophysiologist must keep this in mind when performing IONM for high cervical IMSCTs and when focusing on the subcortical response as an indicator of spinal cord functional continuity. Placing a subcortical recording electrode in the neck may be prohibited by the surgical site and/or prepped area in which case an alternate recording site, such as the chin, inion or the mastoid, is employed. Other scalp-to-scalp and scalp-to-non-cephalic reference recording montages may be incorporated to elucidate subcortical potentials. A starting LFF of 30 Hz and HFF of 750 Hz and an amplifier sensitivity of 10 μV/Div are suitable for these subcortical recordings. For the lower extremity SSEPs, the obligate peak for the subcortical potentials is the P31, presumed to correspond to activity at the level of the dorsal column nucleus/caudal medial lemniscus. The latency of P31 and its amplitude relative to the trailing N34 are tracked. For the upper extremity SSEPs, the obligate peak for the subcortical potential is N13/P14, also presumed to correspond to activity at the level of the dorsal column nucleus/caudal medial lemniscus. The latency of N13/P14 and its amplitude relative to the trailing P18 are tracked. The N34 and the P18 are likely to originate in or around the thalamus.
Finally, potentials obtained at scalp recording sites consistent with the international 10–20 system of electrode placement and specific for localization of the postcentral gyrus, help to validate that the SSEP has arrived at the cortex. These are often referred to as cortical potentials although some generators that contribute to these recordings may be subcortical in origin. Cortical potentials are captured at scalp recording sites that are specific to the activated limb. Scalp channels for lower extremity SSEPs include CPz–FPz and CPi–CPc (to account for paradoxical lateralization) while those for upper extremity SSEPs include CPc–FPz and CPc–CPi where the subscripts “i” and “c” denote ipsilateral and contralateral to the stimulated limb, respectively. Depending on the preferences of your practice, additional scalp electrode montages may be included for capturing cortical potentials. A starting LFF of 30 Hz and HFF of 750 Hz and an amplifier sensitivity of 10 μV/Div are suitable for these cortical recordings. For the lower extremity SSEPs, the obligate peak for the cortical potential is P37, presumed to correspond to activity at the level of mesial thalamocortical projections and synapses. The latency of P37 and its amplitude relative to the trailing N45 are tracked. For the upper extremity SSEPs, the obligate peak for the cortical potential is N20, presumed to correspond to activity at the level of lateral thalamocortical projections and synapses. The latency of N20 and its amplitude relative to the trailing P22 or P30 are tracked.
Display parameters are set and adjusted to optimize visibility. The display sweep for SSEPs will depend on the limb that is being stimulated, and on the acquisition sweep required to obtain the signal effectively. Starting sweeps are normally 100 msec for lower extremity SSEPs and 50 msec for upper extremity SSEPs increasing either or both if there appears to be a latency delay. SSEPs are normally small signals, in the range of single digit μV or even fractions of a μV so starting display gains are on the order of 1 μV/Div for lower extremity SSEPs and 3 μV/Div for upper extremity SSEPs and are adjusted according to the qualities of the signal.
Motor Evoked Potentials
For an in-depth view of muscle evoked potentials (MEPs) I recommend the reader review Chap. 7 and the Association guidelines [24, 25]. A MEP is the bioelectric activity that originates from the primary motor pathway, either from the tracts along its descending course or from its target structures, the muscles.
An overview of the primary motor pathway starts with the upper motor neurons which lie predominantly in the pre-central gyrus of the frontal lobe – the primary motor cortex. Axons that contribute to the CST, the portion of the motor pathway that projects down the spinal cord and directly modulates spinal cord motor-neuronal excitability, originate from large, motor-cortical layer V Betz cells. Upper motor neuron axons descend the cortical radiations, course through the posterior limb of the internal capsule down to the lower brainstem. At this level, approximately 85% of the fibers decussate in the pyramids of the medulla and then turn inferiorly to descend predominantly in the lateral CST. Those that do not decussate in the medulla descend predominantly in the ventral CST. The lateral and ventral CSTs modulate activity in limb and axial muscles, respectively. Of note, a small portion of patients naturally have an uncrossed motor pathway. Axons of the CSTs synapse with large, lower motor neurons – the alpha motor neurons – in the anterior horn of the spinal cord motor nuclei. An alpha motor neuron projects its axon out the ventral root of the spinal cord until it terminates at the motor end plates on muscle fibers. This neuron, the axon and all its innervated muscle fibers form the motor unit. Activity in the alpha motor neuron results in excitatory synaptic events at the neuromuscular junction; depolarization of, and initiation of the contractile mechanism in, the corresponding muscle fibers of the motor unit. It is the pre-contraction depolarization of the muscle fibers that we are detecting in our muscle MEP recordings. The activity of the alpha motor neuron, therefore, directly modulates the duration and strength of the force of contraction.
The spinal cord level of a motor nucleus roughly corresponds to the muscle location. For example, high cervical motor nuclei represent proximal upper extremity muscles and lower cervical motor nuclei represent the arm and hand, and correspondingly, upper lumbar motor nuclei represent muscles of the proximal leg whereas lower lumbosacral motor nuclei represent the distal leg and foot and non-limb muscles such as that of the anal sphincter. The spinal level and distribution of motor nuclei in the spinal cord is important to consider when formulating the IONM strategy and the selection of muscle recording sites for muscle MEP in IMSCT surgery.
When the proximal motor pathway is electrically activated either by transcranial stimulation or by direct cortical, subcortical or spinal cord stimulation a coordinated volley of descending action potentials, believed to be originating predominantly from large diameter, fast conducting axons directly responsible for volitional movements within the CSTs, is initiated. This traveling wave of action potentials can be recorded from the spinal cord directly. To distinguish it from other indirect (I) responses, the direct response has been termed the D-wave (Fig. 13.2).

D-waves . (a) D-waves recorded in surgery for a T2 hemangioblastoma from a 3-contact, spinal electrode placed rostral to the lesion (top trace) and caudal to the lesion (bottom trace). D-waves in this panel were obtained with the 1-3 and 3-1 montages which typically produce the largest responses. The latency of the peak of the response is 3.2 and 5.3 msec, and, the amplitude measured from the peak to the base of the trailing trough is 103 μV and 34 μV for the rostral and caudal recordings, respectively. (b) Overlay of 10 consecutive D-waves recorded in the same patient. (c) A selection of the summary of the D-waves. The recording montages are indicated above each column. The three columns on the left are recorded from the rostral electrode while the three columns on the right from the caudal electrode. Display gains were adjusted for clarity and may not be the same across montages
A description of D-wave characteristics and technical aspects of D-wave monitoring and interpretation are described later in the chapter. When the action potentials contributing to the D-wave descending the CST arrive at the respective synaptic targets on the lower motor neurons in the spinal cord they facilitate depolarization. Lower motor neurons are large and have high capacitance; therefore, they require coincident and repetitive excitatory synaptic events to move their membrane potential above the threshold for firing their own action potentials. For this reason, a single electrical stimulus in the proximal motor pathway, while sufficient to initiate a D-wave, is normally insufficient to trans-synaptically generate an action potential in the lower motor neuron. Therefore, to elicit muscle MEPs we employ a train stimulus paradigm. This results in a rapid succession of D-waves, one for each stimulus pulse in the train (and possibly I-waves, described below), arriving at the lower motor neuron, allowing for temporal summation of excitatory inputs.
At an appropriate stimulus train frequency, the corresponding train of D-waves are superior to a single one at raising the membrane potential of the alpha motor neuron above its threshold, driving it to fire its own action potential and activate the respective muscle fibers. The myogenic motor evoked response obtained when a single motor neuron fires an action potential is a motor unit potential (MUP) , whereas that obtained when two or more motor neurons fire is a compound muscle action potential (CMAP) . It is important to note that muscle MEPs (see below) are sometimes difficult to elicit particularly with transcranial stimulation. In general, it is at and above the level of the alpha motor neuron in the spinal cord that we see neurophysiologic complexity in the ability to activate the motor pathway and generate muscle MEP responses.
Transcranial Motor Evoked Potentials
MEPs for intraoperative spinal cord monitoring are initiated by transcranial electrical activation of the motor cortex and descending motor fibers and are therefore called transcranial motor evoked potentials (TcMEPs). Subdermal needle electrodes, often of corkscrew design, are placed in the scalp over the approximate location of the primary motor cortex. This may either be at the 10–20 system-derived positions of C3, C4, C1, or C2 or slightly anterior to each of these positions (the so-called “M” locations). Be prepared to incorporate electrodes along the Z line (down the midline) at the respective positions of 1 cm posterior to, and 6 cm anterior to, CZ, so-called CZ minus and CZ 6 cm, respectively. The stimulus montage of C1–C2 is a good choice as an activation site for TcMEP in IMSCT surgery, and the alternate montages of C3–C4 and CZ minus–CZ 6 cm are incorporated as needed. The more medial the montage, the greater the selectivity of activation of the fibers of the lower extremity, with the midline stimulus montage resulting in the strictest focus on the lower extremities. Placing stimulating electrodes with such a lower-extremity focus may possibly cause optimal upper extremity responses to be sacrificed. The objective of medially derived montages is primarily to reduce otherwise unmanageable patient movement during TcMEP monitoring. It is secondarily to reduce the incidence of bite injuries of the tongue and mouth, an undesirable outcome of the transcranial stimulation. Transcranial stimulation is not specific for the CST and also results in corticobulbar activation and/or local depolarization of the temporalis muscles both of which cause jaw clenching. Always work closely with the anesthesiology team to incorporate an intraoral bite block(s) to protect the patient from bite injuries. Stimulation through C3-C4 appears to impose the greatest risk in this regard.
For TcMEP, activation of the motor pathway favors the side of the brain under the anode and, thus, the muscles of the hemi-body opposite the anode. An optimized stimulus selectively activates the motor pathway for only the hemi-body opposite the anode. For example, employing C3 as the anode, one would expect right limb muscle responses, and vice versa for C4. However, that lateral specificity of muscle responses during IMSCT surgery is less important than obtaining reliable responses in the important distal muscles. Furthermore, I recommend recording from both sides of the body with each stimulus polarity. This helps the surgical neurophysiologist to capture responses even if the cathodic stimulus generates them, and furthermore prevents congenital non-decussating motor pathways from going undetected, which would negatively impact TcMEP interpretation otherwise. Stimulus parameters such as voltage/current intensity, pulse duration, pulses per train and interpulse interval, and stimulation delivery approaches such as double trains and repetitive, “build-up”–style stimulus delivery are all important contributing factors to optimized activation for TcMEP. In general, the “sicker” the spinal cord the higher the required intensity, pulse duration, number of pulses per train and interstimulus interval and the more likely it will require double trains and repetitive stimulation.
TcMEPs are normally large amplitude, high-frequency, bi- or multiphasic responses and their complexity is defined by the number of undulations as well as the overall duration of the waveform. The complexity and duration are in part related to the number of pulses in the transcranial stimulus, such that longer stimulus trains typically result in more complex and longer TcMEP responses. TcMEP response amplitude is determined as the absolute difference between the largest negative deflection and the largest positive deflection (peak to trough, or vice versa). In general, the latency of the TcMEP response, measured at the point of take-off of the first deflection, depends on the distance from the stimulus site and the distance from the spinal cord, so distal limb muscles have longer latencies than proximal muscles of the same limb. Determining the absolute latency of TcMEP responses during IONM is difficult because it is not known which pulse in the stimulus train is responsible for initiating the response unless the number of pulses in the train is decreased to the point at which the response disappears. This is not a common practice in the operating room, since the absolute latency of TcMEP responses is much less critical to their application and interpretation. TcMEP responses are usually obtained from a one stimulus–one response approach and not normally averaged. In the case of irreconcilable noise, TcMEP responses may be averaged over a few consecutive trials. It is important to keep in mind that TcMEP responses inherently vary from trial to trial.
TcMEP responses are obtained from intramuscular or subdermal needle electrodes placed in or near the belly of the muscles of interest. Since TcMEPs are large, high-frequency responses, a LFF of 10–100 Hz, a HFF of 1500–3000 Hz are appropriate, and a high amplifier sensitivity of 500–3000 μV/Div is necessary to avoid clipping of the waveform which would make amplitudes uninterpretable. Start with an acquisition sweep of 100 msec and consider stretching this acquisition to be able to capture lesion induced delayed responses, particularly for distal muscle groups. Display gain should be initially set at 50–100 μV/Div in order to be able to resolve small responses and adjustments to optimize visibility are often warranted. Tracking the display gains is important particularly when TcMEP data changes occur.
A wide distribution of muscle recording sites is recommended. Proximal and distal muscle groups of the lower extremity such as quadriceps, tibialis anterior and foot muscles are recommended in all cases. Anal sphincter recordings may be included as well. For cervical lesions, segmental and suprasegmental muscles should be included. Proximal “control” muscles should be included whenever possible.
Myelogenic Motor Evoked Potentials: D-Waves
D-waves may be elicited by electrical stimulation delivered directly to the descending CST fibers in the spinal cord or in the brain [26,27,28]. In general, direct stimulation like this is used for mapping and localization. For monitoring D-waves during IMSCT surgery the most common method of activation is transcranial stimulation similar to that employed for TcMEP: electrical pulses delivered to scalp electrodes strategically placed near the motor cortex.
There are important differences between transcranial activation of the motor pathway for TcMEP recording and that for D-wave recording. To obtain D-waves we employ a stimulus of 50–500 μsec (or more) as a single pulse, rather than a train of pulses, since the lower motor neuron and its dependence on temporal summation are excluded during D-wave recording. As a result, patient movement is reduced or eliminated, permitting nearly continuous sampling of D-waves throughout the time the spinal electrode(s) is/are in place. The stimulus for eliciting a D-wave is delivered in such a manner as to obtain responses that correspond to simultaneous activation of both the left and right CST to capture the entire motor axis. To increase the likelihood of this, the C3–C4 scalp electrode positions are favorable, although the other transcranial stimulating electrode positions are also useful and maintain the benefits of focal activation. These more medially located stimulation sites are of greater use in thoracic IMSCT surgery.
For D-wave monitoring, the intensity of the stimulus must be appropriate for simultaneous activation of bilateral CSTs. The stimulus intensity is ramped until the observation of a maximum amplitude response, which suggests bilateral pathways are recruited. The intensity is further increased until a leftward shift in D-wave latency is detected (Fig. 13.3). The intensity just below that which generated the shift in latency at maximum response amplitude is employed for D-wave monitoring. The leftward shift results from deeper activation of the pathway. As the transcranial stimulus intensity is increased, the stimulus-dependent electric field generated under the anode extends across the brain to recruit the contralateral CST. Additional increases in the stimulus intensity then drive the current/electric field deeper into the brain. Consequently, the leading edge of the electric field generated by the stimulus and, therefore, the site of activation of the CSTs, is now more distal and closer to the recording electrode in the spine. This results in the shorter latency described above. Thus, at a stimulus intensity just below that which produces this leftward shift and a maximal response amplitude, it is presumed that bilateral CSTs are maximally activated. Independent left and right D-waves elicited by anodal stimulation of the right and left scalp, respectively, are possible but methods to validate confinement of the activation to one side of the motor axis are lacking.

D-waves at increasing stimulus intensity. D-waves obtained during surgery for C3-6 IMSCT at increasing stimulus intensity from low at the top to higher at the bottom. A time marker is indicated for reference. Note, the D-wave amplitude increases, and the latency decreases as the stimulus is increased until reaching a maximum amplitude at a similar latency (overlaid traces). Additional increases in stimulus result in no further increase in amplitude but a decrease in latency
Unlike TcMEPs which are typically stored after a single trial, D-waves may be averaged. Averaging can help to resolve a poorly formed, small or artifact or noise-contaminated D-wave. The spinal electrode contacts are often of moderate impedance and consequently invite noise, so averaging is an important tool for the surgical neurophysiologist. The number of trials per average depends on the signal-to-noise ratio, with better ratios favoring fewer trials per average, possibly as few as one or two trials. D-wave recordings may also be confounded by large stimulus artifacts. With an optimized stimulus (see above) either stimulus polarity should produce monitorable D-waves. Nevertheless, alternating the stimulus polarity between successive trials for an equal number of trials per polarity before collecting an averaged response is an effective way of canceling the stimulus artifact and of improving the measurability of the collected D-wave.
D-waves are high-frequency, large amplitude waveforms. A LFF of 0.2–2 Hz, a HFF of 1500–3000 Hz, and a starting amplifier sensitivity of 20 μV/Div are appropriate. Pinching the LFF to 30–100 may help manage artifact but phase shifting must be considered. D-waves are short latency waveforms requiring concomitantly brief 10–30 msec acquisition sweeps. The display gain is set to 20 μV/Div and the display sweep matches or is longer than the acquisition sweep. When determining the appropriate stimulus intensity, the display sweep may temporarily be decreased to see greater detail of the small latency shift described above.
D-waves are recorded as near field potentials using a spinal electrode that is inserted in the epidural or subdural space by the surgeon (see Fig. 13.1). To facilitate the placement of the electrode, and to optimize contact between the electrode and the tissue, it is recommended that the surgeon irrigates with warm saline as the spinal electrode is inserted. One spinal electrode is placed caudal (distal) to the site of the lesion as the source of the monitored D-wave, and one rostral (proximal) to the site of the lesion as a control whenever possible.
The recording channels for these electrodes depend on their orientation and the number of contacts. Bipolar montages are preferred to take advantage of the differential amplifier’s ability to reject common mode noise. Commercially available spinal electrodes come with either two or three 1.3 mm contacts approximately 1.5 cm apart center to center. Two to three centimeter interelectrode distance is reported to be optimal. The electrode contacts are numbered distal to proximal. In order to have the negativity of the D-wave represented as an upward deflection, the active recording channel must be the more proximal contact. Therefore, if using a three-contact spinal electrode, the caudal electrode recording montages are (active input to reference input) 3-1, 3-2, and 2-1, whereas, the rostral electrode recording montages are 1-3, 2-3, and 1-2. Since the difference in electrophysiological potentials is being amplified, the montages that include contacts that are farther from each other on the spinal electrode (e.g., 3-1 or 1-3) produce the largest responses due to a reduction in the common mode cancellation of signal that tends to occur between the more closely spaced recording sites (3-2, 2-1 or 1-2, 2-3, Fig. 13.4).

Varying D-wave characteristics at epidural spinal electrode contacts. D-waves obtained from a three-contact epidural spinal electrode using three different bipolar recording montages. Each montage produces a D-wave of different amplitude and latency. A schematic of the epidural electrode with the three contacts is presented on the right. The montages used for recording these D-waves are 3-2 (top trace), 2-1 (middle trace) and 3-1 (lower trace)
In cases where one of the contacts demonstrates poor impedance, then a referential montage may be employed by adding an electrode outside the epidural space as the reference. This montage is unfavorable as it lends to poorly resolved and noise-contaminated recordings since the recording and reference electrodes have different noise detection. Additionally, the shape of the observed D-wave would not match that observed by using a bipolar montage making it difficult to reconcile with the pattern we expect for D-waves (we are, after all, pattern recognition machines).
The D-wave can be recognized by its characteristic shape and latency. D-wave morphology, when recorded extracellularly using a bipolar montage like that described above, is a large negativity (upward deflection) that is bounded by smaller positivity (see Fig. 13.2). This is common for traveling waves recorded from nerves or tracts. The current loops that allow for axonal conduction and repolarization create these brief periods of preceding and trailing positivity surrounding the large negative deflection of the D-wave. Note, at high stimulus intensities the D-wave may bifurcate or trifurcate.
As a traveling wave, the latency of the D-wave depends on the distance from the activation site. Thus, even the small distance between contacts in the epidural spinal electrode is sufficient, and necessary, to see differences in the recorded latencies of the D-wave (see Fig. 13.4). Furthermore, D-waves at lower spinal levels will have relatively longer latency and D-wave latency increases with near-linearity from the mid-cervical levels down. Following transcranial stimulation, D-waves recorded in the cervical spine may appear at a latency of only a few msec whereas those recorded at the lower thoracic spine may appear at 10’s of milliseconds. The short latency of cervical D-waves makes them technically challenging to resolve, as they are often obscured or confounded by an artifact from the stimulus. Averaging responses obtained from alternate stimulus polarities helps to mathematically decrease the stimulus artifact as described above.
The amplitude of the D-wave is directly related to the number of contributing axons. D-waves recorded at higher spinal levels tend to have larger amplitudes than those recorded at lower levels in the same patient since the number of CST fibers is highest in the cervical spinal cord and decreases progressively craniocaudally down the spinal cord. This defines thoracic spinal level 10 as the lower limit of the spine at and above which D-waves can reliably be recorded. In most patients, D-waves are not reliably obtained at and below the conus, because the number of CST fibers there is below a recordable number.
D-waves are typically brief potentials lasting only a few msec in duration. Complicating their identification, occasionally D-waves spread or disperse due to variability in the conduction velocities and desynchronization of the action potentials across the population of the contributing fibers, particularly when there is an impediment to conduction such as a lesion. Some patients will exhibit dispersion that renders D-waves unrecordable; nevertheless, conduction in the CST is sufficient to permit recording of TcMEP following train stimulation. For example, TcMEP responses are obtainable in infants 18 months and younger but they typically have unrecordable D-waves presumably because immature myelination causes too much dispersion.
When stimulating the proximal motor pathway at high intensities or in an awake or lightly anesthetized patient, a single or a series of waves with D-wave-like morphology may follow the D-wave. These are the so-called indirect, or I-waves, which are believed to derive from the activation of intracortical motor circuits. Their activation results in additional coordinated volleys of action potentials descending the CSTs. I-waves are not a good indicator of functional continuity of the motor pathway and are not used in monitoring. The confounding contribution of I-waves may be reduced by increasing anesthesia or decreasing the stimulus.
Despite D-waves being initiated by a single pulse, muscles may still respond and cause muscle artifact at the spinal electrode. Muscle artifact, which is discernible by its later appearance, broader profile, and often large amplitude, may be seen trailing the D-wave (Fig. 13.5). Since the D-wave potential is neurogenic, suspicion of muscle artifact can be validated by giving a small dose of relaxant. Taking appropriate steps like decreasing the stimulus, increasing anesthesia, or adding a low-dose infusion of relaxant may be helpful if resolving the D-wave from muscle artifact is difficult. Keep in mind that, while D-waves are highly resistant to anesthetic type and concentration, including relaxants, intraoperative changes in anesthesia will have secondary, and potentially profound, effects on the other evoked potentials such as SSEP and TcMEP.

Epidurally recorded muscle artifact . Overlay of 10 consecutive trials showing a poorly formed D-wave (time marker) at a latency of approximately 8 msec, with a large trailing negative deflection likely to be muscle artifact, recorded just below spinal level T8 during surgery for ependymoma
Electromyography
For an in-depth view of EMG, I recommend the reader review Chap. 8. EMG is the recording of the bioelectric activity of muscles as an indicator of neuromuscular activation. EMG is normally quiescent and deflections from the low-level background activity are indicators of perturbation of the innervating nerve, root, or tract. Intramuscular or subdermal needle electrodes are placed in or near the belly of the muscles of interest. Bipolar montages are preferred to optimize common mode cancellation of noise but referential montages are applicable as well. Acquisition sweep is set to 50–500 msec/Div with the objectives being to simultaneously sample enough time per sweep while also being able to resolve details of the EMG responses. Recordings are free-running sweeps taken with filters set at 10–3000/5000 Hz and an amplifier sensitivity of 500–2000 μV/Div.
While the sensory and motor evoked potentials emphasized above serve as the focus of the IONM regime applied for protection of the long tracts of the spinal cord during IMSCT surgery, it is important to include EMG as well. In the case of IMSCT surgery, the descending CSTs are at risk of direct mechanical perturbation or trauma. Mechanical perturbation of spinal tracts can initiate volleys of action potentials in the CST that trans-synaptically activate lower motor neurons. Their activation is subsequently detected as sustained discharges or trains of free-running EMG. Whether this EMG activity is an indication of impending decrement in TcMEP or motor function remains to be elucidated. Skinner et al. [8] reported that EMG from tibialis anterior and abductor hallucis recorded at amplifier sensitivity of 20 μV/Div and filters set from 30 Hz–2000 Hz was beneficial in monitoring during IMSCT surgery, and even that EMG events precede and provide warning signs of changes in TcMEPs, findings also recognized in the study by Baeesa et al. [29]. I encourage surgical neurophysiologists to include EMG for the same muscle groups being monitored by TcMEP in their strategy for monitoring during IMSCT surgery as personal experience (unpublished observations) corroborates that of Skinner et al. and Baeesa et al. [8, 29].
Spinal Cord Mapping
Spinal cord mapping includes methods for minimizing neural injury caused by both the surgical entry into the spinal cord and while developing the trajectory to the tumor. Others aim to localize, characterize, or determine the approximate distance from the CSTs [9, 20, 30]. Spinal cord mapping may be incorporated into the IONM strategy during surgery for non-tumor spinal cord lesions and malformations as well. For IMSCTs, the mass effect of space-occupying spinal cord lesions distorts the structure of the spinal cord and/or obscures normal anatomical landmarks, making it difficult for the surgeon to determine the safest entry point into the spinal cord using visual cues alone. Once inside the cord, the margins between a lesion, particularly those of infiltrative subtypes, and normal tissue are difficult to distinguish visually. To this end, mapping of the surface of the dorsal columns and intramedullary mapping of the CSTs has been incorporated in the IONM strategy during IMSCT surgery. It is important to note that intraoperative spinal cord mapping methods, in general, are not a replacement for spinal cord monitoring methods.
Mapping the surface of the spinal cord during IMSCT surgery reduces the incidence of postoperative neurologic deficits [9, 10] and, as such, has emerged as an important addition to the IONM regime therein. The physiologic map determined intraoperatively is combined with imaging and/or navigation results, and with the appearance of the tissue to identify the safest entry point and trajectory into the spinal cord. Access to the inside of the spinal cord during IMSCT surgery is commonly through midline myelotomy at the posterior median septum down the sulcus between the left and right gracile fasciculi. Identifying this septum is critical in this stage of the surgery.
Dorsal Column Mapping
Dorsal column mapping, as the method has been aptly named, has benefitted from some recent methodologic refinements. There are essentially two approaches to dorsal column mapping. In the first, a peripheral nerve is stimulated and potentials in the spinal cord are recorded directly. In the second, the spinal cord is stimulated directly and potentials in distal nerves or the brain via scalp electrodes are recorded. The methods are, nevertheless, based on the same principle. The left and right gracile fasciculi are discrete, parallel (normally) spinal cord pathways that can be readily distinguished physiologically.
Recording from the Spinal Cord
In one method of dorsal column mapping, a specialized, micro-grid electrode is placed on the dorsal surface of the spinal cord [10, 13, 31, 32]. Tibial nerve spinal responses are elicited by standard SSEP stimulation techniques. Responses resulting from independent stimulation of each of the tibial nerves (the median nerves can be used as well, although less commonly and for different mapping information [31]) are recorded directly from the spinal cord, and the gradient of response amplitudes across grid contacts is compared. The grid is laid perpendicular to the long axis of the spinal cord and each contact of the grid contributes to a separate referential montage with the reference electrode placed in the surrounding tissue preferentially at a location equidistant from the micro-grid contacts. Responses are obtained at filters set to LFF of 20–50 Hz and HFF of 1700–2000 Hz. The latency and absolute amplitude of the responses depends on the level of the spinal cord at which the recordings are taken. The grid contact producing the most robust/largest response is localized over the respective gracile fasciculus closest to the midline. The rationale being that the dorsal median septum lies between the contacts that produce the largest responses following stimulation of the left and then the right tibial nerves. Admittedly, the difference between the maximal responses and the others is often subtle and a trained eye is required to interpret them. Furthermore, they are often small responses, inherently variable from trial to trial, tend to be easily contaminated by noise and therefore take time to average the high number of trials required to overcome the poor signal-to-noise ratio. Finally, the micro-grids used for recording are normally developed in house or are expensive if purchased commercially and are challenging to keep stably in place on the spinal cord throughout the required recording period.
Stimulating the Spinal Cord
Stimulation of the spinal cord via two adjacent contacts of a micro-grid placed perpendicular to the long axis of the spinal cord elicits sensory responses at scalp electrodes [33]. Recording from the CP3–CP4 montage, as you would for SSEPs generated from nerves in the limbs, produces averaged scalp potentials of opposite polarity when the right versus the left side of the spinal cord is stimulated. This approach is aptly named the phase reversal method. The stimulus employed is 300 μsec, 0.2 mA constant current pulses delivered at 3.17 Hz at two adjacent contacts on the micro-grid. It was postulated that a “null point” between the electrodes that activated the left and right sides of the cord, corresponding to the anatomical location of the dorsal median septum, would produce cancellation of the potentials at the CP3–CP4 channel as there would be equal activation of the right and left gracile fasciculi simultaneously. The method was later adapted further, employing the same stimulus and recording parameters, but including a handheld bipolar stimulation probe rather than the micro-grid [34] (Figs. 13.6 and 13.7).

Dorsal column mapping by the phase reversal method . Left panels are intraoperative photographs of the spinal cord showing the simulation method incorporating a side-by-side bipolar probe on the right (top), at the midline (middle) and on the left (bottom) of the spinal cord. The right panel shows corresponding scalp recordings at CP3-CP4 that are opposite polarity for the right and left sides and flattened at the midline. (Modified with permission from Nair et al. [34]; by permission of Oxford University Press)

Dorsal column mapping by the phase reversal method . The spinal cord was stimulated at 1 mA, 200 μsec pulse duration, 2.79 Hz through a handheld monopolar probe. Recording channels are indicated at the top of each column. From top to bottom, the scalp potentials at CP4-CP3 indicate that left cord, midline, right cord, midline, left cord and then two trials at the midline stimulation occurred
These authors demonstrate phase reversal of scalp potentials as the stimulus is moved across the spinal cord and also a point at which the scalp potentials flatten, which they describe as corresponding to the location of the dorsal median septum.
The dorsal median septum can be further elucidated by the inclusion of peripheral recordings such as at Erb’s point, the median and ulnar nerves, the popliteal fossa and/or the tibial nerve [9, 35]. In one method of dorsal column mapping focused on recording from the tibial nerve at the ankle, the stimulus consists of 3–8 mA, 200 μsec constant current pulses delivered at 9.1 Hz through a handheld bipolar stimulator with 2–3 mm separation of the tips [35]. The cathode is oriented inferiorly. Subdermal needle electrodes placed behind the medial malleolus over the distal tibial nerve capture antidromic potentials generated at the spinal cord using LFF and HFF filters set to 30 and 300 Hz, respectively. Methodologic advances include recording from Erb’s point, the median and ulnar nerves at the wrist and elbow in addition to the popliteal fossa, with stimuli of 2 mA, 100 μsec constant current pulses delivered at 2.1 Hz through a concentric bipolar handheld stimulator [9]. Again, antidromic potentials elicited at the spinal cord are captured using subdermal needle electrodes placed over the targets using LFF of 30 Hz and HFF of 500 Hz. These researchers suggest, for cervical lesions, proximal recordings of the median and ulnar nerves at the elbow are preferred over recordings at the wrist and that in all cases they met with limited success obtaining potentials at the popliteal fossa. For each of these methods , the laterality of the responses corresponds to the side of the spinal cord stimulated. Stimulation near the dorsal median septum results in either bilateral peripheral responses or no responses, depending on whether the stimulus delivered at the dorsal midline recruits dorsal column fibers on both sides of the cord or is lost in the sulcus.
Peripheral recordings during dorsal column mapping are particularly helpful in patients who have problematic or unresolvable tibial nerve cortical SSEPs. Furthermore, combining dorsal column mapping methods may provide enhanced applicability, accuracy, and reliability. Figure 13.8 depicts dorsal column mapping results obtained from a 6-year old with a mid-cervical ependymoma. Stimulation consisted of 200 μsec, 1 mA constant current pulses delivered at 2.67 Hz through a handheld, side-by-side bipolar probe with the tips separated by 0.3–0.5 cm. The tips of the stimulator were oriented long-wise, but the polarity of the stimulator tips was not tracked; therefore, it is unknown if the cathode was superior or inferior. Figure 13.8 depicts simultaneous responses at scalp, popliteal fossa and muscles recording sites obtained from direct stimulation of the spinal cord and shows that (1) the phase of the scalp response is consistent with the laterality of the responses at popliteal fossa and muscle recording sites and (2) the “null point” scalp responses are consistent with bilateral responses at the popliteal fossa and muscle recording sites. This combination of methods resulted in quick and accurate localization of the dorsal median septum in this patient. Future research regarding refinements and outcomes of this method will be beneficial. It is important to note that, in this case, the responses obtained at muscle recording sites were small and many of them, particularly of the upper arm, were short latency and duration. Furthermore, the most robust responses appeared at the popliteal fossa recording sites and the most robust responses at muscle recording sites appeared at the foot and secondarily in the proximal arm. Despite the stimulator being placed on the dorsal columns, the precise activation site of the observed muscle responses may either be the dorsal columns or the CST directly. A method for distinguishing between the activation site is described below.

Dorsal column mapping combining phase reversal at the scalp with peripheral nerve and muscle recordings obtained from a 6-year old with a mid-cervical ependymoma. The spinal cord was stimulated at 1 mA, 200 μsec pulse duration, 2.79 Hz, with a handheld, side-by-side, bipolar probe at approximately spinal level C6. The scalp montages and recording sites are indicated at the top of each column. The left two columns show scalp recordings at CPz-FPz and CP4-CP3. The remaining columns show alternating left and right side recordings. Columns three through 20 show muscle recordings. The two columns on the far right show popliteal fossa recordings. From top to bottom, the responses in each row indicate left side, midline then right side spinal cord stimulation
CST Mapping
The CST can be localized via “collision studies”— recording a caudal D-wave elicited by transcranial stimulation of the proximal motor pathway and pairing that with direct spinal cord stimulation of the distal motor pathway [32]. Distal stimulation using a handheld probe delivering 2 mA cathodal stimulus pulses in proximity to fibers of the CST results in their recruitment and retrogradely transmitted action potentials in the CST fibers. These retrograde action potentials interfere, or “collide,” with the transcranially elicited action potentials anterogradely descending the CST. According to these methods, a reduction in amplitude of the D-wave recorded caudally indicates a collision has taken place and is interpreted as meaning the stimulation is localized to or near the CST.
Responses in muscles are elicited by direct CST stimulation when employing stimulus trains [35, 36]. Free-running EMG responses of the tibialis anterior and abductor hallucis muscles are recorded at filters set to LFF of 30 Hz and HFF of 1000 Hz. Responses are elicited with constant current, 1 mA and higher, 1 msec pulses delivered at 60 Hz through a handheld bipolar probe the tips of which are separated by 0.5 cm. Stimuli are delivered for 1 sec. In another method, the location of the tumor-tissue interface and the precise proximity of specific contents of the CST can be elucidated in greater detail by micro-stimulation, wherein a handheld concentric bipolar stimulator is employed to deliver 1 msec, biphasic current pulses of 0.1–1.0 mA at 60.11 Hz [37]. Recordings are obtained using filters of 30 and 500 Hz for LFF and HFF, respectively, and an amplifier sensitivity of 200 μV/Div at the thenar-hypothenar hand muscles, the tibialis anterior, extensor hallucis longus, medial gastrocnemius, and the abductor hallucis muscles of the lower extremity. The data from this micro-stimulation study emphasize the need for a broad distribution of muscle recording sites when monitoring for IMSCT surgery.
Recently, Bazilar et al. [38] described a method of transforming the tip of a CUSA into an electrical stimulator delivering 0.5–2 mA, cathodal, constant current, stimulus trains of three 200 μsec long pulses per train, repeated at 1.2 Hz. They report continuous “mapping” of the CST by observing MEPs while working on the tumor since they are able to stimulate repeatedly as they are resecting with the electrified CUSA. While similar to mapping methods, what these authors describe may lead to a new monitoring method during IMSCT surgery. Additional data on the applicability and interpretation of this method are needed.
In methods that employ direct stimulation of the CST with detection of muscle responses, the site of activation of the pathway is unclear. Muscle responses are known to be elicited from stimulation of CST fibers directly, but also indirectly from activation of the dorsal columns. Activating the spinal cord with a double train stimulation protocol with a 60 msec intertrain interval can distinguish the source of activation of muscle responses [39]. Trains of 0.3–5.0 mA constant current, cathodal stimuli of three to five 500 μsec pulses per train, with an interpulse interval of 2–4 msec, are delivered twice consecutively with 60 msec between each train through a concentric bipolar handheld stimulator. Muscle responses are recorded at filters of 10 Hz and 2000 Hz for the LFF and HFF, respectively, in the deltoid, biceps, triceps, extensor digitorum, abductor pollicis brevis, quadriceps, tibialis anterior, gastrocnemius, and abductor hallucis. Since the CST fibers have a shorter refractory period than sensory fibers (such as those of the dorsal columns) direct activation of the CST results in similar muscle responses from each of the two stimulus trains but disparate muscle responses when the dorsal column is activated (Fig. 13.9).

Spinal cord mapping using double trains to distinguish the CST from the dorsal columns as the activation site of muscle response. Left panels are intraoperative photographs of the spinal cord showing the double train simulation method incorporating a concentric bipolar probe on the right corticospinal tract (top) and on the dorsal columns (bottom) of the spinal cord. Right panels depict corresponding muscle recordings from train one (left column) and train two (right column) showing consistent muscle responses (arrows) elicited by both of the trains when activating the corticospinal tract (top) in contrast to disparate responses between the two trains when activating the dorsal columns (bottom). (Modified with permission from Deletis et al. [39], copyright 2018, with permission from BMJ Publishing Group Ltd)
Anesthesia
Anesthesia for monitoring SSEPs, TcMEPs, and EMG has been described in detail in other sections of this text (Chap. 5). Constant, stable anesthesia is critical for the most effective delivery of IONM. This is achieved through steady-state infusion at a constant rate avoiding bolus administration or limiting boluses to induction and select stages of the surgery (like exposure). The surgical neurophysiologist should track closely the anesthetics being used, the infusion rates and boluses administered and incorporate electroencephalography (EEG) since it is an indispensable tool for evaluating the relative depth of the patient throughout the procedure. Optimal anesthesia is delivered intravenously with moderate infusion rates of propofol at 50–200 μg/kg/min and narcotics such as fentanyl around 1 μg/kg/hr. or remifentanyl at 0.2–0.5 μg/kg/min. Gas anesthesia should be avoided, as it reduces the “activatability” of the tissue, particularly, motor tissue of TcMEP. In some instances, particularly for neurologically intact patients, halogenated agents administered at ½ MAC or less or nitrous oxide administered at less than 50% is acceptable but not advised and, in instances of mixed anesthesia, would be paired with lower infusion rates of propofol.
In general, low doses of short-acting muscle relaxants are restricted to induction for cases involving TcMEP and EMG, and clearance of the relaxant must be validated through a train of four evaluation of the neuromuscular junction before baselines are obtained. Relaxants block muscle activation aggressively, such that even with 3 of 4 retained twitches up to 75% of the activatable tissue is still chemically blocked. These neuromuscular blocking agents must be incorporated with great care in order for TcMEP and EMG monitoring to be accurate and reliable.
The anesthetic approaches described above are appropriate for D-wave monitoring and spinal cord mapping. D-waves are tolerant of anesthetics, including relaxants, at doses normally administered for IONM. Anesthesia for spinal cord mapping involving scalp recordings and muscle responses should follow standard approaches for anesthesia appropriate for SSEP and EMG monitoring, respectively. Dorsal column mapping methods that incorporate peripheral nerve recordings do not have specific anesthesia requirements, as peripheral responses are largely insensitive to anesthetics including muscle relaxants.
The surgical neurophysiologist should always track closely systemic physiologic variables such as blood pressure, mean arterial pressure, heart rate, oxygenation, and temperature, since these variables directly impact IONM responses and neural responsiveness. This is particularly true during IONM for IMSCT surgery since the interpretation of SSEPs and TcMEPs is intimately related to the state of these variables.
For IMSCTs at high thoracic levels particularly, special consideration should be given relative to blood pressure and heart rate. Autonomic dysreflexia resulting in rapidly spiking blood pressure and/or heart rate caused by manipulation of the spinal cord at these spinal levels may be misconstrued as an emerging patient [40]. It is critical to have a discussion with the anesthesia team prior to surgery to prepare them to treat these events with non-anesthetic drugs and not address them with an additional anesthetic, an increase in infusion rate or a bolus of anesthetic such as propofol, as this will adversely impact the IONM data with the additional consequence of slow recovery.
IONM Data Changes: Warning Criteria
Evaluation of IONM data changes in IMSCT surgery includes several factors:
-
The quality of the signal(s) to start
-
The pattern of the change(s)
-
The circumstances surrounding the change(s) including:
-
Surgical stage
-
Anesthesia composition and levels
-
Physiologic variables
-
The condition of other IONM signals
-
Evidence of signal fade
-
-
The spinal location, spinal levels, and type of the tumor
With these in mind, the neurophysiologist can identify with reasonable accuracy the site of the IONM data change and can discriminate the source as surgical versus positional versus systemic. Additionally, warning criteria for alerting the surgeon to IONM signal changes, based largely on comparison of the state of the current signals to the parameters of baseline signals strategically set at an earlier stage of the surgery, exist. Nevertheless, the surgical neurophysiologist should be attentive to deteriorating IONM signals, as they can happen rapidly in IMSCT surgery, and notify the surgeon at the outset to have the best chance of employing appropriate tissue- and function-preserving interventions. False negatives may result in unexpected postoperative neurologic outcomes, whereas false positives may lead to an abandonment of the tumor resection. IONM interpretation in IMSCT surgery must include a balance of sensitivity and specificity that limits both false negatives, minimizing unanticipated iatrogenic sequelae as well as false positives, thereby preventing suboptimal or incomplete tumor resection.
SSEPs are evaluated at each recording point along the pathway to localize the site of an SSEP data change. Responses are evaluated throughout the procedure for decreases in amplitude or increases in latency relative to a baseline strategically set earlier in the procedure. SSEPs should also be evaluated against a brief recent history of acquired SSEPs to reduce false positives that may arise from the comparison of data acquired long after the setting of an opening baseline. In general, precipitous changes in SSEPs are more concerning than gradual ones. A decrease in amplitude of 50% and/or an increase in latency of 10% have long been the standard warning criteria for SSEP cortical potentials (although there are new perspectives on the effective warning criteria) [21,22,23]. For many surgeons, it is rare to abort the IMSCT surgery on the basis of isolated SSEP signal changes in the absence of concomitant TcMEP and/or D-wave signal changes.
Like SSEPs, abrupt changes in TcMEP are a greater concern than gradual changes. It is important to note that TcMEP changes often happen rapidly in IMSCT surgery; therefore, TcMEPs need to be sampled frequently in order to anticipate motor changes and to make TcMEPs as effective a tool as possible in both protection as well as prognostication. There are varying views regarding the most effective warning criteria for TcMEP responses, and a body of work suggests the best way to evaluate the risk to motor function in IMSCT surgery is to relate TcMEP responses to the status of the D-wave, whenever possible [41]. Warning criteria for TcMEP fall into three main categories: the absence or presence criteria, activation threshold criteria, and the morphology (particularly percent-reduction) criteria. When evaluating TcMEP using the absence or presence criteria, which is the most documented alarm criteria for TcMEP during IMSCT surgery, an alarm is raised when a previously responsive muscle is now unresponsive [17, 42]. When evaluating TcMEP using the activation threshold criteria, an alarm is raised when there is a 100 V or more increase in the stimulus required to generate TcMEP responses [14, 43]. There are several aspects of TcMEP morphology that are evaluated, including amplitude, which is the most common method, complexity, and duration. When evaluating the TcMEP using an amplitude reduction criterion, an alarm is raised when the TcMEP responses are reduced by 50–80% of the baseline amplitude [6, 44]. When evaluating TcMEP responses using the complexity criterion an alarm is raised when responses become less complex, for example, from multiphasic to biphasic [14]. When evaluating TcMEP responses using a duration criterion an alarm is raised when the duration decreases by approximately 50% [14]. In order to optimally apply complexity and duration criteria to TcMEP the stimulus used to elicit responses should consist of a high number of pulses per train. The results of the complexity criterion and duration criterion methods have not been replicated and more research is required to validate them. In any case, an alarm based on the absence or presence criteria is more likely to warrant a major intervention whereas an alarm based on the other criteria may suitably be addressed with a minor intervention. Finally, improvement of responses that have changed from baseline and the duration of these signal changes are related to the likelihood of functional preservation and/or the length of time required for functional recovery – the closer the improved responses approximate baseline and the shorter the period during which the responses were altered the better the outcomes. The most effective TcMEP alarm criteria in IMSCT surgery that does not include D-wave monitoring is likely to be one that combines these methods and includes consensus regarding the acceptable postoperative neurological status – is no weakness or transient weakness acceptable? Studies on alarm criteria for TcMEPs in IMSCT typically include permissible levels of transient postoperative weakness that recovers in the hours to months following surgery since the major objective of surgery is clearing the patient of cancer. Some investigators extend the understanding of TcMEP warning criteria and report that the appropriate warning criteria depend on the tumor type or the spinal level [6, 18].
D-waves are normally very stable as long as the stimulus and electrode position remain constant, and there is little evidence that they exhibit fade like that commonly seen for other evoked potentials such as SSEPs and TcMEPs. The most common interpretive parameter of D-waves is amplitude. The important threshold for decreases in D-wave amplitude is 50%, and a D-wave that has decreased below this threshold indicates there will be motor functional changes postoperatively, with full loss of D-wave equating with permanent paralysis [17, 24,25,26,27]. The best way of evaluating D-waves is in the context of the results of TcMEPs. If there is loss of TcMEPs with coincident full preservation of the D-wave amplitude, or if there is fully preserved TcMEPs with coincident loss of D-wave, then the motor function is preserved or recovers postoperatively. In the case of loss of TcMEPs with preservation of the D-wave to at least 50% of the baseline amplitude, there will be transient motor functional changes. In the case of loss of TcMEPs and loss or decrease of the D-wave to below 50% of the baseline amplitude there will be long-term or permanent paralysis. Note, in a case of intraoperative decrease of or loss of D-wave with fully preserved TcMEP responses a technical issue should be considered.
The warning criteria for EMG during IMSCT surgery are primarily based on similar warning criteria established for other surgery types, since there has not been a large-scale study of the relationship between patterns of EMG and IMSCT surgical interventions or patient outcomes. Based on one study with a small sample size but an apparent relationship of EMG patterns to motor functional outcomes, the warning criteria include observation of EMG bursts that are coincident with surgical manipulation, prolonged EMG discharges lasting longer than a few seconds and EMG signal that abruptly silences [8]. In all instances of an alarm being raised, TcMEP should be run and muscle responses evaluated based on TcMEP warning criteria and the observed pattern of EMG activity.
Strategies Following IONM Data Changes
In the event an IONM data change exceeding the warning criteria arises during surgery for IMSCT, there are several corrective interventions the surgeon and surgical team may deploy, and these strategies may be (and often are) combined. The first is to pause the surgery. In theory, this allows the nervous system to reset and should last as long as it takes for the IONM data to improve, even as long as 30 minutes.
Irrigation with warm saline is also recommended. A consequence of perturbation of the nervous system is the pooling of ions, particularly potassium, in the extracellular space and the degradation of the normal ion concentration gradients. An abnormal concentration gradient of potassium blocks action potential conduction and results in loss of IONM signals. Irrigation helps to reestablish the appropriate gradients and axonal conduction. At the same time, irrigation helps to clear blood products that are also known to impede nerve conduction.
When the suspected sources of IONM signal loss, usually seen in SSEPs, is retraction on the dorsal surface of the spinal cord, then the surgeon should consider releasing or relaxing the retraction as mentioned above. The surgeon may opt to simply pause with the reduced retraction, and then reapply retraction after the signals have improved. Improvement in SSEPs in this circumstance often occurs rapidly. Alternatively, the surgeon can release the retraction and extend the myelotomy. This is thought to allow the spinal cord to open more freely, decreasing the retractor-induced mechanical stretch or compression on the dorsal columns when the retractors are reapplied.
Maybe the most common strategy to address IONM data change is to take measures to increase perfusion. This is achieved most simply by pharmacologically increasing the blood pressure/mean arterial pressure, presumably resulting in improved perfusion of the spinal cord. Additionally, the surgeon may directly apply papaverine, a smooth muscle relaxant, that results in vasodilation and consequently improves blood volume to the spinal cord.
Persistent IONM data changes may be addressed with the administration of steroids such as an intravenous dose of 30 mg/kg of methylprednisolone.
Lastly, the surgery may be aborted or deferred until after the patient has recovered and has shown improvement in their neurologic function. If surgery is still indicated, they may return to the operating room at that point.
Conclusion
The goals of surgery for IMSCT are extensive if not complete tumor resection in order to not only limit the progression of the disease, but to rid the patient of cancer, its associated symptoms and risks, and to reduce recurrence. These are paralleled by the goals of preserving the patient’s neural function and quality of life. IONM incorporating SSEPs, TcMEP, and D-waves of spinal cord sensory and motor function, and neurophysiologic mapping of spinal cord structures such as the dorsal columns and the CSTs helps the surgeon achieve these goals. Recent advances in the methodology and application of these neurophysiologic tools have made IONM indispensable in the practice of surgery for IMSCTs.
References
Jallo GI, Kothbauer KF, Epstein FJ. Intrinsic spinal cord tumor resection. Neurosurgery. 2001;49(5):1124–8.
Takami T, Naito K, Yamagata T, Ohata K. Surgical management of spinal intramedullary tumors: radical and safe strategy for benign tumors. Neurol Med Chir (Tokyo). 2015;55:317–27.
Hobbs JG, Desai B, Young JS, Polster SP, Tobin MK, Geraghty JR, et al. Intramedullary spinal cord tumors: a review and discussion of surgical rationale. WScJ. 2016;2:65–83.
Sun JJ, Teo M, Wang ZY, Li ZD, Wu HB, Zheng M, et al. Characteristic and surgical results of multisegment intramedullary cervical spinal cord tumors. Interdiscip Neurosurg (Advanced Techniques and Case Management). 2017;7:29–43.
Virdi G. Intramedullary spinal cord tumors: a review of current insights and future strategies. Spine Res. 2017;3(2):13.
Kim DG, Son YR, Park YS, Hyun SJ, Kim KJ, Jahng TA, et al. Differences in multimodality intraoperative neurophysiological monitoring changes between spinal intramedullary ependymoma and hemangioblastoma. J Clin Neurophysiol. 2016;33:120–6.
Foster M, Marquardt G, Seifert V, Szelenyi A. Spinal cord tumor surgery – importance of continuous intraoperative neurophysiological monitoring after tumor resection. Spine. 2012;37:E1001–8.
Skinner S, Nagib M, Bergman T, Maxwell R, Msangi G. The initial use of free-running electromyography to detect early motor tract injury during resection of intramedullary spinal cord lesions. Neurosurgery. 2005;56(4):299–314.
Mehta AI, Mohrhaus CA, Husain AM, Karikari IO, Hughes B, Hodges T, et al. Dorsal column mapping for intramedullary spinal cord tumor resection decreases dorsal column dysfunction. J Spinal Disord Tech. 2012;25(4):205–9.
Yanni DS, Ulkatan S, Deletis V, Barrenechea IJ, Sen C, Perin NI. Utility of neurophysiological monitoring using dorsal column mapping in intramedullary spinal cord surgery. J Neurosurg Spine. 2010;12:623–8.
Choi I, Hyun SJ, Kang JK, Rhim SC. Combined muscle motor and somatosensory evoked potentials for intramedullary spinal cord tumor surgery. Yonsi Med J. 2014;55(4):1063–71.
Verla T, Fridley JS, Khan AB, Mayer RR, Omeis I. Neuromonitoring for intramedullary spinal cord tumor surgery. World Neurosurg. 2016;95:108–16.
Scibilia A, Terranova C, Rizzo V, Raffa G, Morelli A, Esposito F, et al. Intraoperative neurophysiological mapping and monitoring in spinal tumor surgery: sirens or indispensable tools? Neurosurg Focus. 2016;41(2):E18.
Quinones-Hinojosa A, Lyon R, Zada G, Lamborn KR, Gupta N, Parsa AT, et al. Changes in transcranial motor evoked potentials during intramedullary spinal cord tumor resection correlate with postoperative motor function. Neurosurgery. 2005;56:982–93.
Verla T, Fridley JS, Omeis I. Intramedullary spinal cord tumor surgery: the utility of multimodal intraoperative neurophysiologic monitoring. WScJ. 2016;2:111–20.
Siller S, Szelenyi A, Herlitz L, Tonn JC, Zausinger S. Spinal cord hemangioblastomas: significance of intraoperative neurophysiological monitoring for resection and long-term outcome. J Neurosurg Spine. 2017;26:483–93.
Kothbauer K, Deletis V, Epstein F. Motor-evoked potential monitoring for intramedullary spinal cord tumor surgery: correlation of clinical and neurophysiological data in a series of 100 consecutive procedures. Neurosurg Focus. 1998;4(5):Article 1.
Lakomkin N, Mistry A, Zuckerman S, Ladner T, Kothari P, Lee N, et al. Utility of intraoperative monitoring in the resection of spinal cord tumors. Spine. 2018;43(4):287–94.
Sala F, Bricolo A, Faccioli F, Lanteri P, Gerosa M. Surgery for intramedullary spinal cord tumors: the role of intraoperative (neurophysiological) monitoring. Eur Spine J. 2007;16(Suppl 2):S130–9.
Cheng J, Ivan M, Stapleton C, Quinones-Hinojosa A, Gupta N, Auguste K. Intraoperative changes in transcranial motor evoked potentials and somatosensory evoked potentials predicting outcome in children with intramedullary spinal cord tumors. J Neurosurg Pediatr. 2014;13(6):591–9.
Toleikis R. Intraoperative monitoring using somatosensory evoked potentials: a position statement by the American Society of Neurophysiological Monitoring. J Clin Monit Comput. 2005;19(3):241–58.
American Clinical Neurophysiology Society. Guideline 11B: Recommended Standards for Neurophysiologic Intraoperative Monitoring – Principles [Internet]. Milwaukee: American Clinical Neurophysiology Society: 2009. p. 1–10. . Available from: https://www.acns.org/pdf/guidelines/Guideline-11B.pdf.
MacDonald D, Dong C, Quatrale R, Sala F, Skinner S, Soto F, et al. Recommendations of the International Society of Intraoperative Neurophysiology for intraoperative somatosensory evoked potentials. Clin Neurophysiol. 2019;130:161–79.
MacDonald DB, Skinner S, Shils J, Yingling C. Intraoperative motor evoked potential monitoring – a position statement by the American Society of Neurophysiological Monitoring. Clin Neurophysiol. 2013;124:2291–316.
Legatt AD, Emerson RG, Epstein CM, MacDonald DB, Deletis V, Bravo RJ, et al. ACNS Guideline: Transcranial electrical stimulation motor evoked potential monitoring. J Clin Neurophysiol. 2016;33:42–50.
Deletis V, Rodi Z, Amassian V. Neurophysiological mechanisms underlying motor evoked potentials in anesthetized humans. Part 2. Relationship between epidurally and muscle recorded MEPs in man. Clin Neurophysiol. 2001;112(3):445–52.
Deletis V, Sala F. Intraoperative neurophysiological monitoring of the spinal cord during spinal cord and spine surgery: a review focus on the corticospinal tracts. Clin Neurophysiol. 2008;119:248–64.
Fukaya C, Sumi K, Otaka T, Shijo K, Nagaoaka T, Kobayashi K, et al. Corticospinal descending direct wave elicited by subcortical stimulation. J Clin Neurophysiol. 2011;28:297–301.
Baeesa S, Labram E, Mahfoodh M, Khalid M, Al-Said Y. Evolution and role of intraoperative neurophysiological monitoring in intramedullary spinal cord surgery: a 2-year series from Saudi Arabia. World J Neurosci. 2014;4:326–33.
Gonzalez AA, Shilian P, Hsieh P. Spinal cord mapping. J Clin Neurophysiol. 2013;30:604–12.
Krzan M. Intraoperative neurophysiological mapping of the spinal cord’s dorsal columns. In: Deletis V, Shils J, editors. Neurophysiology in neurosurgery: a modern intraoperative approach. New York: Academic Press; 2002. p. 153–65.
Deletis V, Bueno De Camargo A. Interventional neurophysiological mapping during spinal cord perfusions. Stereotact Funct Neurosurg. 2001;77:25–8.
Simon MV, Chiappa KH, Borges LF. Phase reversal of somatosensory evoked potentials triggered by gracilis tract stimulation: case report of a new technique for neurophysiologic dorsal column mapping. Neurosurgery. 2012;70(3):E783–8.
Nair D, Kumaraswamy VM, Braver D, Kilbride RD, Borges LF, Simon MV. Dorsal Column mapping via phase reversal method: the refined technique and clinical applications. Neurosurgery. 2014;74(4):437–46.
Quinones-Hinojosa A, Gulati M, Lyon R, Gupta N, Yingling C. Spinal cord mapping as an adjunct for resection of intramedullary tumors: surgical technique with case illustrations. Neurosurgery. 2002;51:1199–207.
Duffau H, Campelle L, Sichez J. Direct spinal cord electrical stimulation during surgery of intramedullary tumoral and vascular lesions. Stereotact Funct Neurosurg. 1998;71:180–9.
Ghandi R, Curtis CM, Cohen-Gadol AA. High-resolution direct microstimulation mapping of spinal cord motor pathways during resection of an intramedullary tumor. J Neurosurg Spine. 2015;22:205–10.
Barzilai O, Lidar Z, Constantini S, Salame K, Bitan-Talmor Y, Korn A. Continuous mapping of the corticospinal tracts in intramedullary spinal cord tumor surgery using an electrified ultrasonic aspirator. J Neurosurg Spine. 2017;27:161–8.
Deletis V, Seidel K, Sala F, Raabe A, Chudy D, Beck J, Kothbauer KF. Intraoperative identification of the corticospinal tract and dorsal column of the spinal cord by electrical stimulation. J Neurol Neurosurg Psychiatry. 2018;89:754–61.
Lyford T, Borowczyk K, Danieletto S, Vlok R. Recognition and management of intraoperative autonomic dysreflexia. J Surgery Emerg Med. 2016;1:1e102.
Costa P, Peretta P, Faccani G. Relevance of intraoperative D wave in spine and spinal cord surgeries. Eur Spine J. 2013;22:840–8.
Kothbauer KF. The interpretation of muscle Motor Evoked Potentials for spinal cord monitoring. J Clin Neurophysiol. 2017;34:32–7.
Calancie B. Intraoperative Neuromonitoring and alarm criteria for judging MEP responses to transcranial electric stimulation: The threshold-level method. J Clin Neurophysiol. 2017;34:12–22.
Journee HL, Berends HI, Kruytt MC. The percentage of amplitude decrease warning criteria for transcranial MEP monitoring. J Clin Neurophysiol. 2017;34:22–31.
Acknowledgments
The author thanks Christopher Martin CNIM, Rose Tash CNIM, Christopher Solotaroff CNIM, Gregory Jason CNIM and Debbie Davy PhD, for their contributions to this manuscript and for their assistance with its preparation.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Review Questions
Review Questions
-
1.
What advantages does monitoring D-waves offer for intramedullary spinal cord surgery?
-
2.
How do D-wave and TcMEP monitoring work together to improve surgical outcome?
-
3.
Describe two techniques for dorsal column mapping and why dorsal column mapping may be performed.
-
4.
How might surgeons respond to changes in spinal cord mapping and monitoring data?
Rights and permissions
Copyright information
© 2020 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Pace, C.J. (2020). Monitoring and Mapping of the Spinal Cord. In: Davis, S., Kaye, A. (eds) Principles of Neurophysiological Assessment, Mapping, and Monitoring. Springer, Cham. https://doi.org/10.1007/978-3-030-22400-4_13
Download citation
DOI: https://doi.org/10.1007/978-3-030-22400-4_13
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-22399-1
Online ISBN: 978-3-030-22400-4
eBook Packages: MedicineMedicine (R0)