
Abstract
Valorization of solid wastes from palm oil mill based medium for the production of enzyme from Streptomyces philanthi RM-1-138 and the efficacy of its culture filtrate as an antimicrobial agent against plant pathogenic fungi and bacteria were investigated. Solid-state fermentation of S. philanthi RM-1-138 grown in oil palm decanter cake (OPDC), palm pressed fiber (PPF), and OPDC + PPF (1:1) for production of enzymes was investigated after 12 days incubation. OPDC was the best substrate as it gave the highest enzyme activity of xylanase, cellulase, and chitinase (at 5.21 ± 0.26, 3.75 ± 0.38, and 0.48 ± 0.01 U mL−1 respectively) at 10 days incubation. In addition, the production of β-1,3-glucanase was not detected in all raw materials. From our previous studies, the culture filtrate of this strain was found to contain bioactive compounds. In this study, the efficacy of the culture filtrate RM-1-138 produced under the optimal conditions (60% moisture content with the initial pH at 7.0) was evaluated against three strains each of plant pathogenic fungi and bacteria. Its efficacy was most pronounced in Ganoderma boninense (74.33%) followed by Curvularia oryzae (64.67%) and Ceratocystis paradoxa TT1 (58.33%), respectively. Its efficacy against Xanthomonas axonopodis pv. glycines, X. oryzae pv. oryzae, and X. campestris pv. campestris were low and not significantly different (P > 0.05) (19.17, 18.50, and 18.33 mm, respectively). With these results, the OPDC had a high potential to be utilized as substrate for the production of enzymes and antimicrobial agent by S. philanthi RM-1-138.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Palm oil is one of the five major edible oils in the world. In Thailand, the production of palm oil has become one of the most important agro-industries during the last 20 years, mainly in the eastern and southern regions of the country [1, 2]. A typical palm oil mill generates a substantial mass of wastes for every ton of fresh fruit bunches; empty fruit bunches (20–28%), oil palm decanter cake (OPDC) (4%), and palm pressed fiber (PPF) (11-12%) [3]. OPDC is the solid waste generated after processing the oil sludge through the decanter (or 3-phase separator) while PPF is generated after passing the digested oil palm fruits through double screw press, air classifier, and fiber separator, respectively. Basically, the OPDC still contains 30–40% of residual and is currently disposed of directly in landfills without treatment [4], causing severe water and air pollution with a high environmental impact on the sustainability of the palm oil industry.
A current problem in the south of Thailand is to manage the wastes generated during palm oil milling processes. OPDC has high organic carbon (85.9%, volatile solid) and a nutrient-rich composition (2.9%, total Kjeldahl nitrogen) content [5] in the form of oil and fibrous materials that have a high potential feedstock for biogas production. In the past, utilization of OPDC was limited to use in oil palm plantations for mulching and soil conditioning. Recently, many researchers have studied the suitability of OPDC as co-boiler fuel [6, 7], animal feed [8], fish feed [9], composting material and enzymes [10, 11], cellulase and polyoses [12], and as a source of biofuel (biogas, hydrogen, and methane) production [7, 13]. However, its economic value is low and it is, therefore, not considered to possess an attractive potential for use as substrate for bioactive compounds production. As the major components of OPDC are lignin (30.66%), cellulose (21.61%), and hemicellulose (3.94%) [12], it can be utilized as an alternate low-cost substrate for lignocellulosic enzyme production that acts corporately to degrade the components of lignocellulose. Moreover, OPDC contains organic matters and nutrients (nitrogen, carbon sulfur, phosphate, potassium, calcium, magnesium, boron, manganese, copper iron, and zinc) [12] that are essential for the growth of Actinobacteria during ligninolytic enzyme and bioactive compounds production [14].
Actinomycetes play a pivotal role in maintaining a satisfactory biological balance in soil, largely because of their ability to produce antibiotics and other secondary metabolites [15, 16]. Among actinomycetes, Streptomyces species are well known for their essential role in the naturally occurring degradation of lignocellulose, which is enabled by the secretion of extracellular of lignocellulosic (cellulase and xylanase) [17, 18] and ligninolytic (lignin peroxidase, manganese peroxidase, and laccase) [14, 19] enzymes. Moreover, Streptomyces are a major source for production of antibiotics [20] and bioactive compounds [21] with their potential applications in agriculture, medicine, and the food industry. Numerous studies on antimicrobial production by Streptomyces were reported using synthetic media [22,23,24,25,26]. In particular, the glucose yeast-malt extract (GYM) medium has been shown to be a good medium for growth and antifungal production of S. philanthi RM-1-138 [27,28,29,30], but it has never been reported using OPDC as the medium. Therefore, this work contributes to knowledge on an alternative potential of OPDC as a low-cost substrate for bioactive compound production by S. philanthi RM-1-138 using solid-state fermentation (SSF). SSF using agro-industrial wastes is an attractive and cost-effective option because it represents higher productivity with simpler operation when compared with submerged fermentation. This research aimed to investigate the effectiveness of OPDC as a substrate in the SSF system for bioactive compounds production by S. philanthi RM-1-138 and evaluation of its efficacy against both plant pathogenic fungi and bacteria.
2 Materials and methods
2.1 Microorganisms, media, and cultivation conditions
Streptomyces philanthi RM-1-138 was previously isolated and characterized as described by Boukaew et al. [27] and grown on GYM agar at room temperature (28 ± 2 °C) for 10 days before use.
Curvularia oryzae and Ganoderma boninense were oil palm pathogenic fungi that obtained from the Suratthani Oil Palm Research Center, Department of Agriculture, Ministry of Agriculture and Cooperative, Thailand and Biohythane Laboratory at Prince of Songkla University (PSU), Thailand, respectively. Ceratocystis paradoxa TT1 was isolated from the natural fermented oil palm trunk [31] and reported to cause black seed rot disease in oil palm sprouted seeds [32]. They are known to cause significant losses in oil palm crops in Southeast Asia, particularly in Thailand, Malaysia, and Indonesia [33, 34]. The fungal strains were grown on potato dextrose agar (PDA) slants at room temperature (28 ± 2 °C) for 3–5 days and kept at 4 ℃. They were sub-cultured freshly for use in each experiment.
Plant pathogenic bacteria Xanthomonas axonopodis pv. glycines, Xanthomonas oryzae pv. oryzae, and Xanthomonas campestris pv. campestris were obtained from the Department of Plant Pathology, Faculty of Agriculture, at Kasetsart University (KU), Thailand. They are known to cause significant losses in many crop plants worldwide. The bacterial strains were grown on nutrient agar (NA) slants at room temperature (28 ± 2 °C) for 24–48 h and kept at 4 ℃. The inoculum was prepared by culturing the bacterium in nutrient broth (NB) for 24 h on a rotary shaker (at 150 rpm). The concentration of the inoculum was then adjusted to 106 CFU mL−1 before used.
2.2 Oil palm decanter cake and its characteristics
Oil palm decanter cake (OPDC) and palm pressed fiber (PPF) were collected from a palm oil mill (Laff Tavee Palm Co., Ltd.) in Satun province, Southern Thailand. OPDC was dried in an oven at 95 ℃ until the moisture content was lower than 5% [35]. Characteristics of OPDC were determined for chemical oxygen demand (COD), total solid (TS), total suspended solids (TSS), total phosphorous (TP), and phosphate following the Standard Method [36]. Total Kjeldahl nitrogen (TKN) was analyzed using Kjeldahl method [37]. Temperature and pH were measured using thermometer and pH meter, respectively. Glucose, xylose, fructose, and arabinose were determined using high-performance liquid chromatography (HPLC) [38].
2.3 Enzyme assays
2.3.1 Raw materials for enzyme production of S. philanthi RM-1-138
Solid-state fermentation of S. philanthi RM-1-138 grown in oil palm decanter cake (OPDC), palm pressed fiber (PPF), and OPDC + PPF (1:1) as substrates for the production of cellulase, xylanase, chitinase, and β-1,3-glucanase was investigated.
2.3.2 Cellulase and xylanase activity
The cellulase and xylanase activity were determined according to Bailey et al. [39]. Quantitation of reducing sugar released from xylan or carboxymethylcellulose (CMC) prepared in 50 mM acetate buffer pH 5.0 with 0.1 mL of the appropriately diluted enzyme to give an absorbance reading below 0.7. The enzyme-substrate mixture was incubated at 50 ℃ for 5 min. The released reducing sugars were determined by the use of 3,5-dinitrosalicylic acid (DNS) method with xylose or glucose used as standards for xylanase and cellulase activities, respectively [40]. One unit of xylanase or cellulase is defined as the amount of enzyme that liberates 1 µmol of xylose or glucose equivalents per minute, respectively.
2.3.3 Chitinase and β-1,3-glucanase activity
Chitinase and β-1,3-glucanase activity were measured using colloidal chitin and laminarin as the substrates, respectively following the DNS method [40]. The reaction mixture of chitinase and β-1,3-glucanase activity was prepared according to the method of Chairin and Petcharat [41] with some modification. Briefly, the reaction mixture contained 250 μL of crude sample and 1% (w/v) substrate in 250 μL of 50 mM potassium phosphate buffer (KPB) pH of 7.0 for colloidal chitin and 50 mM acetate buffer pH of 6.0 for laminarin and incubated for 30 min in 50 °C water bath. Reducing sugar released in the mixtures was determined by recording the absorbance at 570 nm for chitinase and 550 nm for β-1,3-glucanase. One unit (U) of chitinase activity was defined as releasing 1 μmol of N-acetyl-D-glucosamine from the substrate per minute and one unit of β-1,3-glucanase activity was defined as releasing 1 μmol of glucose from the laminarin per minute.
2.4 Optimization of raw materials and environmental condition for enzymes production of S. philanthi RM-1-138
2.4.1 Effect of raw materials
The effect of raw materials, OPDC, PPF, and OPDC mixed with PPF (1:1), on enzyme production of S. philanthi RM-1-138 was investigated. Five milliliter aliquots seed culture (107 spore mL−1) of the strain RM-1-138 was transferred into 50 g each of the sterilized (121 ℃/15 min) raw materials (with moisture content of 60%; initial pH adjusted to 7.0) and incubated at room temperature (28 ± 2 °C). After 12 days incubation, the enzymes were extracted from the cultivated materials by adding 100 mL of 0.1 M phosphate buffer (pH 7.0) and placed on a rotary shaker (at 150 rpm) for 2 h [42, 43]. The suspension was centrifuged (at 8880 × g for 20 min), then filtered through a 0.45 µm Millipore membrane to obtain the supernatant. The samples were taken every 2 days to measure the enzyme activities. The supernatant was determined for enzyme activity. The raw materials that exhibited the highest enzyme activity were selected for further studies.
2.4.2 Effect of moisture content
The effect of moisture contents (30, 40, 50, 60, and 70%) of the selected raw material (results from previous experiment) on enzyme production from S. philanthi RM-1-138 was studied. Five milliliter aliquots seed culture of the strain RM-1-138 was transferred into 50 g of each sterilized selected raw materials with pH adjusted to 7.0 and incubated at room temperature (28 ± 2 °C) for 10 days. The moisture content that exhibited the highest enzyme activity was selected for further studies.
2.4.3 Effect of initial pH
The effect of initial pH (6.0, 7.0, and 8.0) on production of enzymes by S. philanthi RM-1-138 was investigated. Five milliliter aliquots seed culture of the strain RM-1-138 was transferred into 50 g of the sterilized optimum raw materials (result from the previous experiment) and incubated at room temperature (28 ± 2 °C) for 10 days. The initial pH that exhibited the highest enzyme activity was selected for further studies.
2.5 Testing for the presence of bioactive compounds in the culture filtrate of S. philanthi RM-1-138 against oil palm pathogenic fungi and bacteria
The culture filtrate of S. philanthi RM-1-138 (culture filtrate RM-1-138) grown in the optimum raw materials and condition (results from the optimization studies) was tested for its efficacy against both oil palm pathogenic fungi (C. oryzae, G. boninense, and C. paradoxa TT1) and bacteria (X. axonopodis pv glycines, X. oryzae pv. oryzae, and X. campestris pv. campestris). The experiment was carried out as above and the culture filtrate was extracted from the cultivated medium (as described above) at the optimum incubation time and used for the antimicrobial assay. The positive results on antifungal or antibacterial assay would indicate the presence of bioactive compounds in the culture filtrate RM-1-138.
2.5.1 Antifungal assay
The culture filtrate RM-1-138 (1 mL) was mixed with 9 mL melted sterile PDA (at 60 °C) and poured onto a 9 cm diameter culture plate, while distilled sterilized water (1 mL) mixed with 9 mL melted sterile PDA at an equivalent amount was used as a control. A 5 mm diameter plug of mycelia was cut from a 3-day old in each oil palm pathogens colony and transferred onto the center of the test agar plates. The cultures were further incubated at room temperature (28 ± 2 °C) for 2 days of C. paradoxa TT1, 4 days of C. oryzae, and 7 days of G. boninense. The experiment was conducted in three replications. The colony size of each treatment was recorded and the percentage inhibition of hyphal growth was calculated using the equation: percentage of inhibition = [{(control-treatment) / control} × 100] [29].
2.5.2 Antibacterial assay
Agar well diffusion assay was used for the detection of antibacterial activity. NA plates containing 106 CFU per mL of bacteria pathogen were prepared. A well with a diameter of 6 mm was then cut into the agar using a sterile cork-borer. A droplet of agar was added to the well in order to seal it to avoid leakage. Then, 100 µL of the culture filtrate RM-1-138 was added to the well and allowed to diffuse into the agar during a 5-h pre-incubation period at room temperature, followed by incubation at room temperature (28 ± 2 °C) for 48 h. The antibacterial zone was recorded.
2.6 Statistical analysis
The data were submitted to analyses of variance using Statistical Package for the Social Sciences (SPSS) ver. 26 (IBM Corp; IBM SPSS Statistics for Windows, ver. 26.0, Armonk, NY). A P value < 0.05 was considered significant.
3 Results
3.1 Characteristics of oil palm decanter cake (OPDC)
Raw OPDC had an acidic pH (5.33) and black color. The characteristics of OPDC (Table 1) indicated that the high values of organic matter (9828 mg kg−1, COD) showed the possibility to be used as a carbon source for the growth of microorganisms. Total solids (TS) and suspended solids (TSS) (21,762 and 18,330 mg kg−1, respectively). It also contained nitrogen (TKN of 1443 mg kg−1 and NH4-N of 358.8 mg kg−1) as well as various sugars: glucose, xylose, fructose, and arabinose (1396, 1154, 319, and 324 mg kg−1, respectively). These chemical composition showed the possibility that OPDC could be used as substrate for the growth of microorganisms.
3.2 Optimization of raw materials and environmental condition for enzymes production of S. philanthi RM-1-138
3.2.1 Effect of raw materials
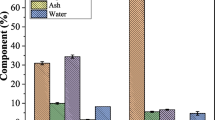
Three sets of raw materials, OPDC, PPF, and OPDC + PPF (1:1), were used to study their effects on xylanase, cellulase, chitinase, and β-1,3-glucanase production of S. philanthi RM-1-138. They showed significant differences in enzyme production (Fig. 1) with the highest values from OPDC followed by PPF and OPDC + PPF (1:1) after 12 days incubation. OPDC as the substrate gave the higher enzyme activity of xylanase (Fig. 1A), cellulase (Fig. 1B), and chitinase (Fig. 1C) (5.21 ± 0.26, 3.75 ± 0.38, and 0.48 ± 0.01 U mL−1, respectively) at 10 days incubation. In addition, the production of β-1,3-glucanase was not detected in all raw materials. Therefore, the OPDC was the selected raw material for further studies.

Time course on the production of xylanase (A), cellulase (B), and chitinase (C) from Streptomyces philanthi RM-1-138 in oil palm decanter cake (OPDC) (square), palm pressed fiber (PPF) (circle), and OPDC + PPF(triangle) and adjusted moisture content to 60% after cultivation for 12 days using solid-state fermentation. Data are the mean of three replicates ± standard deviation (SD)
3.2.2 Effect of moisture content and initial pH
The enzyme assay was employed to study the effect of moisture contents (30–70%) and initial pH (6–8) in OPDC culture of S. philanthi RM-1-138 on the production of xylanase, cellulase, chitinase, and β-1,3-glucanase after incubating for 10 days (Fig. 2). The increasing amounts of moisture content from 30 to 60% resulted in a significant increase in the production of xylanase and cellulase by 2.34 and 2.45 fold (from 2.53 ± 0.12 to 5.91 ± 0.26 U mL−1 and 1.61 ± 0.38 to 3.95 ± 0.38 U mL−1, respectively). The optimal moisture content for enzyme production was found to be 60%. Substrate moistened at this level afforded high xylanase, cellulase, and chitinase activity values of 5.91 ± 0.26, 3.95 ± 0.38, and 0.56 ± 0.01 U mL−1, respectively (Fig. 2A), indicating this moisture content being favorable for S. philanthi RM-1-138 growth in OPDC medium. However, β-1,3-glucanase activity was not detected at all in every initial moisture content tested.

The effect of initial moisture content (A) and initial pH (B) in oil palm decanter cake (OPDC) medium on the production of xylanase, cellulase, and chitinase from Streptomyces philanthi RM-1-138 after cultivation for 10 days. Data are the mean of three replicates ± standard deviation (SD). Values with the same letter are not significantly different (ANOVA, P > 0.05; Duncan’s multiple range test)
The impact of the initial pH (6–8) of OPDC on the production of xylanase, cellulase, chitinase, and β-1,3-glucanase from S. philanthi RM-1-138 was studied in the optimum OPDC culture (Fig. 2B). The highest xylanase, cellulase, and chitinase activities were obtained at an initial pH of 7.0 (6.03 ± 0.25, 4.79 ± 0.76, and 0.76 ± 0.02 U mL−1, respectively) but no significant differences (P > 0.05) of xylanase (5.87 ± 0.10 U mL−1) and chitinase (0.61 ± 0.02 U mL−1) activities at the initial pH of 8.0.
3.3 Testing for the presence of bioactive compounds in the culture filtrate of S. philanthi RM-1-138 against oil palm pathogenic fungi and bacteria
The presence of bioactive compounds in the culture filtrate of S. philanthi RM-1-138 produced in the optimal OPDC medium on the growth of the three oil palm pathogenic fungi (C. oryzae, C. paradoxa TT1, and G. boninense) and bacteria (X. axonopodis pv. glycines, X. oryzae pv. oryzae, and X. campestris pv. campestris) was investigated (Table 2). The mycelial growth of the three oil palm pathogenic fungi was inhibited with the most pronounced inhibitory effect (74.33%) was against G. boninense followed by against C. oryzae (64.67%) and C. paradoxa TT1 (58.33%) (Table 2A). For agar well diffusion assay on pathogenic bacteria tested, no significant differences (P > 0.05) on pathogenic bacteria were observed. The bioactive compounds were shown to inhibit the growth of each plant pathogenic bacteria in the inhibition zone range of 18.33 to 19.17 mm (Table 2B). The most pronounced inhibitory effect was against X. axonopodis pv. glycines (19.17 mm) followed by X. oryzae pv. oryzae (18.50 mm), and X. campestris pv. campestris (18.33 mm).
4 Discussion
The antimicrobial metabolites produced by Streptomyces species are known to be efficient against plant pathogenic fungi [44, 45] and bacteria [46,47,48]. To substitute the mostly used expensive synthetic medium, oil palm decanter cake (OPDC) was proposed to be used as substrate for production of enzymes under solid-state fermentation. In this study, the efficacy of the antimicrobial metabolites produced in OPDC of S. philanthi RM-1-138 was evaluated against three strains of oil palm pathogenic fungi (Curvularia oryzae, Ganoderma boninense, and Ceratocystis paradoxa TT1) and bacteria (Xanthomonas axonopodis pv. glycines, X. oryzae pv. oryzae, and X. campestris pv. campestris).
Initial moisture contents and pH of the substrate are known to critically influence microorganism growth and bioactive compounds production in solid-state fermentation (SSF) [49, 50]. The highest enzymes activity was noted in OPDC followed by PPF after 12 days incubation. OPDC was high in organic matters (9828 mg kg−1 COD, 21,762 mg kg−1 TS and 18,330 mg kg−1 TSS). The concentration of nitrogen (1443 mg kg−1 TKN), glucose and xylose (1396 and 1154 mg kg−1, respectively) were similar while the amount of fructose and arabinose were much lower (319, and 324 mg kg−1, respectively). The characteristics of OPDC obtained are expected to deviate highly from the literature as it varies with the quality of palm fruits, processing techniques, quality control of individual mills, crop seasons, and other factors. Besides, TKN, COD, TS, and TSS of OPDC are observed to be relatively lower than those reported elsewhere [12, 51,52,53] as it depends on the sampling point of the substrate where the settling of solids occurred. With its high TS content, OPDC is a promising material for use as a renewable energy source through the SSF process for production of lignocellulosic enzymes and bioactive compounds. Although the weight substrates (50 g) content of the two raw materials was similar, OPDC is more easily digested than PPF. This probably explains the better bioactive compounds production in OPDC. The results indicated that the differences in compositions of raw materials had influenced bioactive compounds production by S. philanthi RM-1-138.
OPDC used as a sole substrate afforded the highest levels of xylanase (5.21 ± 0.26 U mL−1), cellulase (3.75 ± 0.38 U mL−1), and chitinase (0.48 ± 0.01 U mL−1) production than the PPF and the mixed substrates. It is worth of noting that the strain RM-1-138 showed multiple (endo- and exo-cellulase as well as xylanase) enzymatic activities that lends them interesting as sources of biocatalysts for lignocellulose conversion. Very few works investigated so far cellulase, xylanase, and chitinase production in S. philanthi RM-1-138, even if cellulolytic activity has been reported in different Actinobacteria, such as Streptomyces, Thermoactinomyces, and Cellulomonas. The highest xylanase activity (5.21 ± 0.26 U mL−1) achieved by S. philanthi RM-1-138 in the present work was higher than the activity from S. flavogriseus AE63X (0.95 U mL−1) [54], Streptomyces strain C1-3 (0.6 U mL−1) [55], and Streptomyces strain AMT-3 (0.17 ± 0.02 U mL−1) [56]. In addition, the cellulase activity (3.75 ± 0.38 U mL−1) was much higher than the cellulase activity (0.87 ± 0.19 U mL−1) from S. flavogriseus AE64X [54]. However, the chitinase activity (0.48 ± 0.01 U mL−1) using OPDC medium was lower than previously reported with cultivated in a basal medium containing 0.05% (w/v) MgSO4, 0.02% (w/v) K2HPO4, 0.03% (w/v) KH2PO4, and 0.001% (w/v) each of FeSO4, ZnSO4, and MnCl2 (0.53 U mL−1) [57], than that of S. thermocarboxydus TKU045 (52.985 U mL−1) [58], and that of S. vinaceusdrappus S5MW2 (14.5 ± 0.4 U mL−1) [59]. It is worth of noting that the values of these enzyme activities may be obtained from the differences in the analytical methods such as the substrate and incubation condition.
The presence of water in the substrate makes the nutrients more easily accessible for bacterial growth. Moreover, water has an impact on physico-chemical properties of the substrate, which in turn affect enzyme production [60]. Too much water adversely affects oxygen diffusion in the substrate [49]. In this work, the initial moisture contents of the substrate were adjusted to 30–70% in separate experiments before inoculation with the strain RM-1-138. Clearly, the optimum initial moisture level was 60%. Substrate moistened at this level afforded a high cellulase, xylanase, and chitinase activity (5.91 ± 0.26, 3.95 ± 0.38, and 0.56 ± 0.01 U mL−1, respectively) at 10 days. This result was lower than that for xylanase and cellulase production by A. niger (65%) [61], (77.67%) [62], (75%) [63], Penicillium canescens (83%) [64], Melanocarpus albomyces IIS-68 (67%) [65], and Thermoascus aurantiacus (81%) [66], but higher than by Chaetomium globosum (50%) [67]. This may be due to the different culture media and microorganisms. Moisture levels above 60% reduced enzyme production as the substrate became water logged [68]. High moisture content is known to reduce voidage or porosity of substrates, causes particles to stick together, and adversely impacts oxygen transfer to the microorganisms [46, 69]. This explains the effects of elevated levels of moisture.
As a general practice, the pH in SSF is almost never controlled during fermentation, and only the initial pH of the substrate is adjusted before inoculation. Most substrates such as wheat bran, rice husk, rice bran, spent brewing grain, coconut oil cake, palm kernel cake, sesame oil cake, jackfruit seed powder, and olive oil cake used in SSF are known to possess an excellent buffering capacity [46, 69]. The results revealed that the optimum pH of the medium for enzyme production was 7.0. It could be speculated that SSF contributed to a better buffering capacity. Medium pH is very important in nutrient absorption and growth of bacteria, stimulation of enzyme production via signaling pathways, and release of extracellular enzymes [70].
In the in vitro assay, S. philanthi RM-1-138 was able to produce both antifungal and antibacterial metabolites compounds using OPDC as substrate medium, which greatly inhibited the plant pathogenic growth of three strains each of fungi (C. oryzae, C. paradoxa TT1, and G. boninense) and bacteria (X. axonopodis pv. glycines, X. oryzae pv. oryzae, and X. campestris pv. campestris) tested. In this study, we demonstrated that the antifungal compounds of S. philanthi RM-1-138 can significantly (P < 0.05) affect against the three strains of oil palm pathogen. Among the three strains of oil palm fungal pathogen tested, antifungal compounds exhibit the strongest inhibition against G. boninense (74.33%). It is much higher than the antifungal compounds produced in the synthetic medium (56.64% inhibition) reported by Shariffah-Muzaimah et al. [71] but lower than those against G. boninense (92.04% inhibition) and C. oryzae (84.60% inhibition) reported by Pithakkit et al. [33]. Among the three bacterial strains tested, the antibacterial compounds was not significantly difference (P > 0.05) against X. axonopodis pv. glycines (19.17 mm), X. oryzae pv. oryzae (18.50 mm), and X. campestris pv. campestris (18.33 mm). It can be seen that the antibacterial compounds obtained in OPDC medium is much higher than the efficacy reported by Okudoh and Wallis [72], Jaivel et al. [73], Kovácsová et al. [74], and He et al. [75] (15–16 mm) but lower than those against X. oryzae pv. oryzae (25 mm) reported by Hastuti et al. [76]. The results from antifungal and antibacterial actives indicated that OPDC has high potential for utilization as an alternative low-cost SSF medium for the production of both antimicrobial compounds by S. philanthi RM-1-138. This strain was reported to have several mechanisms such as bioactive and volatile compounds by which it inhibits plant pathogens as reported for many Streptomyces species, such as, S. globisporus JK-1 [22], S. philanti RM-1-138 [27,28,29,30], and S. mycarofaciens SS-2-243 [30]. However, to our knowledge, this study is the first one revealing the utilization of OPDC, a waste product of palm oil processing, as an alternative low-cost SSF medium for the production of the bioactive compound by S. philanthi RM-1-138. Based on the in vitro experiment, the S. philanthi RM-1-138 could grow in OPDC and secreted both antifungal and antibacterial compounds into the medium.
In conclusion, the results obtained in this study clearly indicated that OPDC had high potential for utilization as an alternative low-cost SSF medium for the production of both antifungal and antibacterial compounds by S. philanthi RM-1-138. The antimicrobial compounds have high efficacy against three strains of oil palm pathogenic fungi and bacteria.
References
Rongwang C, Polprasert S, Kanchanasuta S (2017) Effect of partial ozonation and thermal pretreatment on biogas production from palm oil decanter cake. Chem Eng Trans 57:1987–1992
Thamvithayakorn P, Phosri C, Pisutpaisal N, Krajangsang S, Whalley AJS, Suwannasai N (2019) Utilization of oil palm decanter cake for valuable laccase and manganese peroxidase enzyme production from a novel white-rot fungus, Pseudolagarobasidium sp. PP17-33. 3 Biot 9:417
Prasertsan P, Prasertsan S (2004) Bioconversion of agro industrial residues for bioprocesses. Palm oil industry residues. in Concise Encyclopedia of Bioresource Technology. (Pandey, A., Editor). The Haworth Press. pp. 460–465.
Noparat P, Maneerat S, Saimmai A (2014) Utilization of palm oil decanter cake as a novel substrate for biosurfactant production from a new and promising strain of Ochrobactrum anthropi 2/3. World J Microbiol Biotechnol 30:865–877
Eko H, Chaiprasert P (2020) Enhancement of methane production from high solid anaerobic digestion of pretreated palm oil decanter cake using a modified solid inclined reactor. J Chem Technol Biotechnol 95:781–790
Maniam GP, Hindryawati N, Nurfitri I, Jose R, Ab. Rahim MH, Dahalan FA, Yusoff MM (2013) Decanter cake as a feedstock for biodiesel production: a first report. Energy Convers Manag 76:527–532
Kanchanasuta S, Pisutpaisal N (2016) Waste utilization of palm oil decanter cake on biogas fermentation. Int J Hydrog Energy 41:15661–15666
Gafar AA, Alimon AR, Sazili AQ, Man YC, Abubakar AR (2013) Effect of varying levels of palm oil decanter cake on feed intake, growth performance and carcass characteristics of Kacang goats. IOSR J Agricult Vet Sci (IOSR-JAVS) 3:24–29
Chang YP, Chuah JH (2015) Evaluation of palm decanter cake as an alternative fish feed. In: Ab. Hamid K, Ono O, Bostamam A, Poh Ai Ling A (eds) The Malaysia-Japan model on technology partnership. Springer, Tokyo, pp 321–326
Nutongkaew T, Duangsuwan W, Prasertsan S, Prasertsan P (2014) Effect of inoculum size on production of compost and enzymes from palm oil mill biogas sludge mixed with shredded palm empty fruit bunches and decanter cake. Songklanakarin J Sci Technol 36:275–281
Nutongkaew T, Duangsuwan W, Prasertsan S, Prasertsan P (2014) Physicochemical and biochemical changes during composting of different mixing ratios of biogas sludge with palm oil mill wastes and biogas effluent. J Mater Cycles Waste Manag 16:131–140
Razak MNA, Ibrahim MF, Yee PL, Hassan MA, Abd-Aziz S (2012) Utilization of oil palm decanter cake for cellulase and polyoses production. Biotechnol Bioprocess Eng 17:547–555
Kanchanasuta S, Sillaparassamee O (2017) Enhancement of hydrogen and methane production from co-digestion of palm oil decanter cake and crude glycerol using two stage thermophilic and mesophilic fermentation. Int J Hydrog Energy 42:3440–3446
Riyadi FA, Tahir AA, Yusof N, Sabri NSA, Noor MJMM, Akhir FNMD, Othman N, Zakaria Z, Hara H (2020) Enzymatic and genetic characterization of lignin depolymerization by Streptomyces sp. S6 isolated from a tropical environment. Sci Rep 10:7813
El-Tarabily KA (2008) Promotion of tomato (Lycopersicon esculentum Mill.) plant growth by rhizosphere-competent 1-aminocyclopropane-1-carboxylic acid deaminase- producing streptomycete actinomycetes. Plant Soil 308:161–174
El-Tarabily KA, AlKhajeh AS, Ayyash MM, Alnuaimi LH, Sham A, ElBaghdady KZ, Tariq S, AbuQamar SF (2019) Growth promotion of Salicornia bigelovii by Micromonospora chalcea UAE1, an endophytic 1-aminocyclopropane-1-carboxylic acid deaminase-producing actinobacterial isolate. Front Microbiol 10:1694
Poornima S, Divya P, Karmegam N, Karthik V, Subbaiya R (2020) Aqueous two-phase partitioning and characterization of xylanase produced by Streptomyces geysiriensis from low cost lignocellulosic substrates. J Biosci Bioeng 130:571–576
Marasinghe SD, Jo E, Hettiarachchi SA, Lee Y, Eom TY, Gang Y, Kang YH, Oh C (2021) Characterization of glycoside hydrolase family 11 xylanase from Streptomyces sp. strain J103; its synergetic effect with acetyl xylan esterase and enhancement of enzymatic hydrolysis of lignocellulosic biomass. Microb Cell Fact 20:129
Devi P, Kandasamy S, Chendrayan K, Uthandi S (2016) Laccase producing Streptomyces bikiniensis CSC12 isolated from compost. Microbiol Biotech Food Sci 6:794–798
Strohl WR (2004) Antimicrobials. In: Bull AT (ed) Microbial diversity and bioprocessing. American Society for Microbiology, Washington, D.C, pp 336–355
Berdy J (2005) Bioactive microbial metabolites. J Antibiot 58:1–26
Li Q, Jiang Y, Ning P, Zheng L, Huang J, Li G, Jiang D, Hsiang T (2011) Suppression of Magnaporthe oryzae by culture filtrates of Streptomyces globisporus JK-1. Biol Control 58:139–148
Shakeel Q, Lyu A, Zhang J, Wu M, Chen S, Chen W, Li G, Yang L (2016) Optimization of the cultural medium and conditions for production of antifungal substances by Streptomyces platensis 3–10 and evaluation of its efficacy in suppression of clubroot disease (Plasmodiophora brassicae) of oilseed rape. Biol Control 101:59–68
Xu T, Cao L, Zeng J, Franco CMM, Yang Y, Hu X, Liu Y, Wang X, Gao Y, Bu Z, Shi L, Zhou G, Zhou Q, Liu X, Zhu Y (2019) The antifungal action mode of the rice endophyte Streptomyces hygroscopicus OsiSh-2 as a potential biocontrol agent against the rice blast pathogen. Pestic Biochem Physiol 160:58–69
Sharma P, Thakur D (2020) Antimicrobial biosynthetic potential and diversity of culturable soil actinobacteria from forest ecosystems of Northeast India. Sci Rep 10:4104
Duan Y, Chen J, Pang Z, Ye X, Zhang C, Hu H, Xie J (2020) Antifungal mechanism of Streptomyces ma. FS-4 on fusarium wilt of banana. J Appl Microbiol 130:196–207
Boukaew S, Chuenchit S, Petcharat V (2011) Evaluation of Streptomyces spp. for biological control of Sclerotium root and stem rot and Ralstonia wilt of chili. BioControl 56:365–347
Boukaew S, Klinmanee C, Prasertsan P (2013) Potential for the integration of biological and chemical control of sheath blight disease caused by Rhizoctonia solani on rice. World J Microbiol Biotechnol 29:1885–1893
Boukaew S, Prasertsan P (2014) Suppression of rice sheath blight disease using heat stable culture filtrate of Streptomyces philanthi RM-1-138. Crop protect 61:1–10
Boukaew S, Prasertsan P, Troulet C, Bardin M (2017) Biological control of tomato gray mold caused by Botrytis cinerea by using Streptomyces spp. BioControl 62:793–803
Nutongkaew T, Prasertsan P, O-Thong S, Chanthong S, Suyotha W (2020) Improved methane production using lignocellulolytic enzymes from Trichoderma koningiopsis TM3 through co-digestion of palm oil mill effluent and oil palm trunk residues. Waste Biomass Valorization 11:5123–5136
Eziashi EI, Omamor IB, Odigie EE (2006) Antagonism of Trichoderma viride and effects of extracted water soluble compounds from Trichoderma species and benlate solution on Ceratocystis paradoxa. Afr J Biotechnol 6:388–392
Pithakkit S, Petcharat V, Chuenchit S, Pornsuriya C, Sunpapao A (2015) Isolation of antagonistic actinomycetes species from rhizosphere as effective biocontrol against oil palm fungal diseases. Walailak J Sci Tech 12:481–490
Sunpapao A, Kittimorakul J, Pornsuriya C (2014) Disease note: identification of Curvularia oryzae as cause of leaf spot disease on oil palm seedlings in nurseries of Thailand. Phytoparasitica 42:529–533
Mamimin C, Jehlee A, Saelor S, Prasertsan P, O-Thong S (2016) Thermophilic hydrogen production from co-fermentation of palm oil mill effluent and decanter cake by Thermoanaerobacterium thermosaccharolyticum PSU-2. Int J Hydrog Energy 41:21692–21701
APHA, Awwa, WEF (1999) Standard methods for the examination of water and wastewater, 21st edn. American Public Health Association, Washington, DC
Singh R, Pradhan K (1981) Determination of nitrogen and protein by Kjeldahl method. In: Forage Evaluation Science, p. 23, Pvt. Publishers Ltd., New Delhi
Eyéghé-Bickong HA, Alexandersson EO, Gouws LM, Young PR, Vivier MA (2012) Optimisation of an HPLC method for the simultaneous quantification of the major sugars and organic acids in grapevine berries. J Chromatogr B 885–886:43–49
Bailey MJ, Biely P, Poutanen K (1992) Interlaboratory testing of methods for assay of xylanase activity. J Biotechnol 23:257–270
Miller GL (1959) Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal Biochem 31:426–428
Chairin T, Petcharat V (2017) Induction of defense responses in longkong fruit (Aglaia dookkoo Griff.) against fruit rot fungi by Metarhizium guizhouense. Biol Control 111:40–44
Ali CH, Mbadinga SM, Liu JF, Yang SZ, Gu JD, Mu BZ (2015) Significant enhancement of Pseudomonas aeruginosa FW_SH-1 lipase production using response surface methodology and analysis of its hydrolysis capability. J Taiwan Inst Chem Eng 52:7–13
Neethu CS, Rahiman KMM, Rosmine E, Saramma AV, Mohamed AA (2015) Utilization agro-industrial wastes for the production of lipase from Stenotrophomonas maltophilia isolated from arctic and optimization of physical parameters. Biocatal Agric Biotechnol 4:703–709
Shariffah-Muzaimah SA, Idris AS, Madihah AZ, Dzolkhifli O, Kamaruzzaman S, Maizatul-Suriza M (2018) Characterization of Streptomyces spp. isolated from the rhizosphere of oil palm and evaluation of their ability to suppress basal stem rot disease in oil palm seedlings when applied as powder formulations in a glasshouse trial. World J Microbiol Biotechnol 34:15
Sujarit K, Pathom-aree W, Mori M, Dobashi K, Shiomi K, Lumyong S (2020) Streptomyces palmae CMU-AB204T, an antifungal producing-actinomycete, as a potential biocontrol agent to protect palm oil producing trees from basal stem rot disease fungus. Ganoderma boninense. Biol Control 148:104307
Tan HM, Cao LX, He ZF, Su GJ, Lin B, Zhou SN (2006) Isolation of endophytic actinomycetes from different cultivars of tomato and their activities against Ralstonia solanacearum in vitro. World J Microbiol Biotechnol 22:1275–1280
Zhao J, Han L, Yu M, Cao P, Li D, Guo X, Liu Y, Wang X, Xiang W (2019) Characterization of Streptomyces sporangiiformans sp. nov., a novel soil actinomycete with antibacterial activity against Ralstonia solanacearum. Microorganisms 7:360
Promnuan Y, Promsai S, Meelai S (2020) Antimicrobial activity of Streptomyces spp. isolated from Apis dorsata combs against some phytopathogenic bacteria. PeerJ 18:e10512
Chisti Y (1999) Solid substrate fermentations, enzyme production, food enrichment. In: Flickinger MC, Drew SW (eds) Encyclopedia of bioprocess technology: fermentation, biocatalysis, and bioseparation, vol 5. Wiley, New York, pp 2446–2462
Hamidi-Esfahani Z, Shojaosadati SA, Rinzema A (2004) Modelling of simultaneous effect of moisture and temperature on A. niger growth in solid-state fermentation. Biochem Eng J 21:265–272
Lim YF, Chan YJ, Hue FS, Ng SC, Hashma H (2021) Anaerobic co-digestion of palm oil mill effluent (POME) with decanter cake (DC): effect of mixing ratio and kinetic study. Bioresour Technol Rep 15:100736
Kanchanasuta S, Pisutpaisal N (2017) Improvement of glycerol waste utilization by co-feedstock with palm oil decanter cake on biohydrogen fermentation. Int J Hydrog. Energy 42:3447–3453
Chaikitkaew S, Kongjan P, O-Thong S (2015) Biogas production from biomass residues of palm oil mill by solid state anaerobic digestion. In: Energy Procedia pp. 838–844
Pennacchio A, Ventorino V, Cimini D, Pepe O, Schiraldi C, Inverso M, Faraco V (2018) Isolation of new cellulase and xylanase producing strains and application to lignocellulosic biomasses hydrolysis and succinic acid production. Bioresour Technol 259:325–333
Meryandini A (2007) Characterization of xylanase from Streptomyces strain C1–3. Hayati J Biosci 3:115
Nascimiento RP, Coelho RRR, Marques S, Alves L, Gírio FM, Bon EPS, Amaral-Collaço MT (2002) Production and partial characterization of xylanase from Streptomyces sp. strain AMT-3 isolated from Brazilian Cerrado soil. Enzyme Microb Technol 31:349–555
Boukaew S, Petlamul W, Suyotha W, Prasertsan P (2016) Simultaneous fermentative chitinase and β-1,3 glucanase production from Streptomyces philanthi RM-1-1-38 and their antifungal activity against rice sheath blight disease. BioTechnologia 97:271–284
Tran TN, Doan CT, Nguyen VB, Nguyen AD, Wang SL (2019) The isolation of chitinase from Streptomyces thermocarboxydus and its application in the preparation of chitin oligomers. Res Chem Intermed 45:727–742
Yandigeri MS, Malviya N, Solanki MK, Shrivastava P, Sivakumar G (2015) Chitinolytic Streptomyces vinaceusdrappus S5MW2 isolated from Chilika lake, India enhances plant growth and biocontrol efficacy through chitin supplementation against Rhizoctonia solani. World J Microbiol Biotechnol 31:1217–1225
Pandey A, Selvakumar P, Soccol CR, Nigam P (1999) Solid state fermentation for the production of industrial enzymes. Curr Sci 77:149–162
Park YS, Kang SW, Lee JS, Hong SI, Kim SW (2002) Xylanase production in solid state fermentation by Aspergillus niger mutant using statistical experimental designs. Appl Microbiol Biotechnol 58:761–766
Tian M, Wai A, Guha TK, Hausner G, Yuan Q (2018) Production of endoglucanase and xylanase using food waste by solid-state fermentation. Waste Biomass Valorization 9:2391–23983
Tai WY, Tan JS, Lim V, Lee CK (2019) Comprehensive studies on optimization of cellulase and xylanase production by a local indigenous fungus strain via solid state fermentation using oil palm frond as substrate. Biotechnol Prog 35:e2781
Bakri Y, Jacques P, Thonart P (2003) Xylanase production by Penicillium canescens 10–10c in solid-state fermentation. In: Davison B.H., Lee J.W., Finkelstein M., McMillan J.D. (eds) Biotechnology for Fuels and Chemicals. Applied Biochemistry and Biotechnology. Humana Press: Totowa
Jain A (1995) Production of xylanase by thermophilic Melanocarpus albomyces IIS-68. Process Biochem 30:705–709
Souza MCO, Roberto IC, Milagres AMF (1999) Solid state fermentation for xylanase production by Thermoascus aurantiacus using response surface methodology. Appl Microbiol Biotechnol 52:768–772
Wiacek-Zychlinska A, Czakaj J, Jedrychowska B, Sawicka- Zukowska R (1992) Production of xylanases by Chaetomium globosum. Prog Biotechnol 7:493–496
Chutmanop J, Chuichulcherm S, Chisti Y, Srinophakun P (2008) Protease production by Aspergillus oryzae in solid-state fermentation using agro industrial substrates. J Chem Technol Biotechnol 83:1012–1018
Sandhya C, Sumantha A, Szakacs G, Pandey A (2005) Comparative evaluation of neutral protease production by Aspergillus oryzae in submerged and solid-state fermentation. Process Biochem 40:2689–2694
Palekar AA, Vasudevan PT, Yan S (2000) Purification of lipase: a review. Biocatal Biotransform 18:177–200
Shariffah-Muzaimah SA, Idris AS, Madihah AZ, Dzolkhifli O, Kamaruzzaman S, Cheong PCH (2015) Isolation of actinomycetes from isolation of rhizosphere of oil palm (Elaeis guineensis Jacq.) for antagonism against Ganoderma boninense. J Oil Palm Res 27:19–29
Okudoh VI, Wallis FM (2007) Antimicrobial activity of rare actinomycetes isolated from natural habitats in KwaZulu-Natal, South Africa. S Afr J sci 103:216–221
Jaivel N, Rajesh R, Marimuthu P (2014) Evaluation of antimicrobial activity against bacterial leaf blight pathogen Xanthomonas oryze pv. oryzae and antioxidant activities of Streptomyces sp. TC1. Afr J Microbiol Res 8:3558–3564
Kovácsová S, Javoreková S, Medo J, Charousová I, Elbl J, Plošek L (2015) Characteristic of Streptomyces species with antimicrobial activity against selected phytopathogenic bacteria and fungi. J Microbiol Biotech Food Sci 5:55–59
He H, Hao X, Zhou W, Shi N, Feng J, Han L (2019) Identification of antimicrobial metabolites produced by a potential biocontrol Actinomycete strain A217. J Appl Microbiol 128:1143–1152
Hastuti RD, Lestari Y, Suwanto A, Saraswati R (2012) Endophytic Streptomyces spp. as biocontrol agents of rice bacterial leaf blight pathogen (Xanthomonas oryzae pv. oryzae). Hayati J Biosci 19:155–162
Acknowledgements
This research work was financially supported by the Agricultural Research Development Agency (Public Organization) (CRP6205012110) and Thailand Research Fund (RTA6280014).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Human and animals rights
No human and/or animal participants were involved in this research.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Boukaew, S., Prasertsan, P., Petlamul, W. et al. Palm oil decanter cake wastes as alternative nutrient sources for production of enzymes from Streptomyces philanthi RM-1-138 and the efficacy of its culture filtrate as an antimicrobial agent against plant pathogenic fungi and bacteria. Biomass Conv. Bioref. 14, 1895–1904 (2024). https://doi.org/10.1007/s13399-022-02448-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13399-022-02448-7