
Abstract
Miocene coral-bearing limestones, distributed in the western Makran Basin, are attributed to Aquitanian and Burdigalian. This investigation is focused on three Early Miocene coral-bearing limestone sections in western Makran. Remarkably, well-preserved scleractinian corals and other components could be very definitive to reconstruct paleoenvironmental conditions. Lithologically, there are some differences between the studied sections. Furthermore, zooxanthellate corals are diverse and abundant in all sections. Based on different components and richness of zooxanthellate corals, it seems that the studied carbonate corals are precipitated in a subtropical condition, between 19 and 20 °C in all sections. In light of the dominancy of corals, the presence of symbiont-bearing larger benthic foraminifera (LBF), lithological features, bioeroding features, and encrusting organisms, the oligotrophic to slightly mesotrophic conditions are considered for Tejek section and the mesotrophic condition had been prevailed in Kermestan and Irer sections. According to light intensity in water column and coral morphotypes, Tejek section is considered to precipitate under euphotic to slightly mesophotic condition, while Kermestan and Irer sections were deposited under mesophotic to euphotic light conditions. In transparent water, photic zones continue to deeper depths, while in less transparent water, these zones are limited to shallower parts. A defined depth in the photic zone may represent euphotic, mesophotic, or oligophotic zone. Based on the water transparency, a taxon in a defined photic zone can occur in various depths. On the basis of trophic-light intensity-depth chart, the estimated depth ranges are 12–85 m for Tejek section and 5–62 m for Kermestan and Irer sections. Water energy as another important factor in environmental condition is acquired from coral morphotypes. Accordingly, Tejek section is precipitated under moderate-high energy and Kermestan and Irer sections are deposited in low to moderate-high energy. The presence of encrusting coralline algae, corals, and other constituents is indicative of different substrates in the studied areas. Corals favorably develop in normal salinity waters. The existence of colonial corals and occurrence of benthic foraminifera with hyaline wall indicate normal seawater conditions.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Because of high tectonic activity, the Makran Basin is very tectonized (e.g. McCall and Kidd 1982; Dolati 2010; Burg et al. 2011) so that different kinds of sediments were deposited in each time interval. Among them, Miocene coral-bearing limestones are in the center of our attention. Such Miocene coral-bearing limestones are present in Nikshahr, Fannuj, Taherui, and Minab quadrangle maps (Peterson and Rudzinskas 1982). These carbonate deposits may be a part of Dehirdan, Sabz, Roksha, Ghasr Ghand, Band-e-Chaker and Darkhunish units which are Aquitanian/Burdigalian in age (McCall et al. 1994).
Well-preserved scleractinian corals as main components as well as other minor constituents in studied sections (Tejek, Irer and Kermestan) could be very definitive to reconstruct paleoenvironmental circumstances. Many of scleractinian corals host light-dependant symbionts which are so sensitive to different environmental conditions; for that reason, they are directly affected by light penetration in the water and other factors such as nutrient availability, salinity, temperature, depth, substrate, and water energy extremely influence the corals life.
Most of the investigations carried out in Makran Basin are restricted to studies of tectonics and seismicity (e.g., Falcon 1947; Farhoudi and Karig 1977; Jacob and Quittmeyer 1979; Niazi et al. 1980; Sondhi 1947; McCall 1997; McCall and Kidd 1982; Dolati 2010; Burg et al. 2011), while few paleontological studies have been accomplished during the past decades (e.g., McCall et al. 1994; Crimes and McCall 1995; Abbasi 2000).
McCall et al. (1994) investigated on Early Miocene coral limestones of Iranian Makran; later, Crimes and McCall (1995) and Abbasi (2000) recorded diverse trace fossils of Eocene–Miocene and Miocene rocks of the Makran Basin, respectively. That is why investigation on coral-bearing limestone is very valuable for us. This study aims to consider the paleoecological and environmental factors of Early Miocene coral-bearing limestones of the Band-e-Chaker Unit in the western Makran (Tejek, Kermestan and Irer sections (Plate 1)).

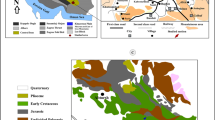
The structural divisions of Iranian Makran Basin (after McCall et al. 1994) and road map of investigated sections (red circles are studied locations)
Geological setting
The Makran Basin extends from Oman sea coasts in south to Jazmurian depression in north. Its western limit is Minab fault which separates the Makran and Zagros basins; the Makran Basin continues to the Las Bela axis of Pakistani Balouchestan in east and totally is 160,000 km2 (Aghanabati 2004). Based on McCall and Kidd (1982) and McCall (1985), eight geotectonic zones of the Makran Basin are as follow: (1) Jazmurian Depression, (2) Spreading Zone, (3) Carbonate Fore-arc Zone, (4) Trench Zone (colored mélange), (5) Eocene–Oligocene Flysch Zone, (6) Oligocene–Miocene Flysch Zone, (7) Miocene Neritic Sediment Zone, and (8) Miocene–Pliocene Neritic-Continental Sediment Coastal Zone (Fig. 1). The studied sections are situated in Miocene Neritic Sediment Zone.

The map of studied area around Band-e-chaker Syncline in western Makran (modified after Peterson and Rudzinskas 1982); the studied sections are as follows: I Irer, K Kermestan, T Tejek. The blue lines are Miocene coral limestones mapped in Peterson and Rudzinskas (1982), and red lines are faults positions
There are no formal lithostratigraphic divisions in the Makran Basin; thus, the Makran successions are informally divided into different units. Lithological subdivisions of the Makran Miocene successions and their main properties are summarized in Table 1.
As an informal unit, the single name “Vaziri Unit” was initially allocated to all Miocene coral-bearing limestone units in Taherui, Minab, Fannuj, and Nikshahr quadrangles, but this was perhaps premature because it was later found more satisfactory to classify them as intervals within these larger units; they also proved to be of two ages, namely Aquitanian and Burdigalian (McCall et al. 1994). Therefore, the previously named Vaziri Unit could be a limestone part in Dehirdan, Sabz, Roksha, Ghasr Ghand, Band-e-Chaker, and Darkhunish units (McCall et al. 1994).
In the Early Miocene, analysis of deposits shows that shallow-water conditions and shelf deposition were common and they were influenced by tides; on the other hand, the Middle Miocene was characterized by high tectonic activity and highly variable sedimentary facies (Burg et al. 2011). Accordingly, it seems that the Early Miocene deposits of western Makran (which our study sections are among them) are precipitated in a rather more stable tectonic condition.
This investigation is focused on three Early Miocene coral limestone sections in western Makran. Based on McCall et al. (1994), Miocene limestones in Makran Basin occur in Dehirdan Unit, Sabz Unit, Roksha Unit, Ghasr Ghand Unit, and Band-e-Chaker Unit. Besides, considering Peterson and Rudzinskas (1982) and our field observation, coral limestone units in the studied area are sandwiched between “Dehirdan Unit” and “shaly, siltstones, and sandstone layers of Band-e-Chaker Unit”; therefore, it may be difficult to attribute coral limestones to the mentioned unites, because the coral limestones may be related to the end of Dehirdan Unit or beginning of Band-e-Chaker Unit. It is worth noting that McCall et al. (1994) differentiated ages of units and limestone members of the Makran Basin (Table 2). Based on these authors, the coral-bearing limestone in Dehirdan Unit is Aquitanian in age, while in Band-e-Chaker Unit, it is Burdigalian in age. To confirm the age of the studied sections, the index corals in the Makran Basin (McCall et al. 1994) and distribution of LBF in Indo-Pacific and Mediterranean regions (BouDagher-Fadel and Price 2013) are investigated and described in the following parts. As the sections are Burdigalian in age, the coral-bearing limestones must be related to Band-e-Chaker Unit as it is confirmed by McCall et al. (1994).
Material and methods
Three sections at Tejek, Kermestan, and Irer areas (close to Khomeinishahr City) in western Makran were measured. As it is known, most of Miocene deposits in western Makran are shale, marl, and sandstone (e.g., Peterson and Rudzinskas 1982; McCall et al. 1994). However, the studied sections are Miocene carbonates that are well exposed and contain remarkably well-preserved zooxanthellate and azooxanthellate corals. Lithological traits of the studied sections were distinguished in the field area and more complement studies were accomplished in the university labs. Different coral assemblage growth fabrics were classified according to Insalaco (1998), and different coral morphotypes were recorded. More than 500 coral specimens were sampled, and significant abundances of corals in the depositional sequences through field observations were recorded in photographs. Coral assemblages were systematically identified to have more accurate paleoecological results. Lithological XRF analysis of Tejek section was prepared in Central Laboratory of University of Isfahan; it is worth noting that to achieve better paleoecological interpretations, up to 50 thin sections and polished slabs were prepared and studied precisely. Polished slabs and macrospecimens were photographed by Canon digital camera (SX200) and thin sections were photographed.
Studied sections
Totally three stratigraphic sections were investigated in Bashagard area, around Band-e-Chaker Syncline in western Makran (Figs. 1 and 2). Considering lithostratigraphic and tectonic characteristics of Makran area, the investigation was focused on the Miocene coral-bearing sequences of the Kermestan, Irer, and Tejek sections. The study sections are discussed below.

Profiles of the studied sections. 1 Tejek section, 2 Kermestan section, 3 Irer section
Kermestan
The Kermestan section is located at 26° 40′ 26.62″ N and 57° 56′ 38.99″ E near Kermestan Village in northeast flank of Band-e-Chaker Syncline (Figs. 1 and 2). The studied interval in the Kermestan area is situated normally between Dehirdan and shaly, siltstone, and sandstone layers of Band-e-Chaker units (Plate 1; Fig. 3). The contacts between the studied carbonate sequence and underlying Dehirdan Unit are conformable. Here, the marly limestone sequence is the lower part of Band-e-Chaker Unit and comprises 10 m thickness, accompanied with high frequency of platy, solitary, and head corals. According to Insalaco (1998), the section could be divided into two parts: the lower part begins with dominancy of platestone fabric and continues to upper part with presence of platestone–domestone fabric. In this stratigraphic section, 24 genera are recovered. Some corals such as Acanthastraea (Acanthastraea) echinata, Cyphastraea progoensis, Heliastraea (Athecastraea) boehmi, Larterophyllia cf. turriformis, Oulophyllia eocenica, Pavona folium, Thegioastraea roasendai, Goniopora globulosa, Caulastraea sp., Alveopora meridionalis, Fungophyllia verbeeki, Leptophyllon nishihirai, Porites sp., Stylophora reussiana, Astrocoenia bistellata are present, apart from foraminifera, red algae, bryozoans, gastropods, bivalves and serpulid worms. Among the mentioned acquired corals, based on McCall et al. (1994), A. (Acanthastraea) echinata is index of Burdigalian and P. folium is index fossil of Early Miocene of the Makran Basin.

Stratigraphic columns of studied sections in Tejek, Kermestan, and Irer; different coral assemblages are assigned for each part: Assemblage 1 domestone part of the Tejek section, Assemblage 2 platestone part of the Kermestan section, Assemblage 3 platestone–domestone part of the Kermestan section, Assemblage 4 platestone part of the Irer section, Assemblage 5 platestone–domestone part of the Irer section
Irer
The Irer section is located near Irer Village at 26° 40′ 44.87″ N and 57° 56′ 14.70″ E in northeast flank of Band-e-Chaker Syncline (Figs. 1 and 2). Stratigraphically, the Irer section as well as Kermestan section is situated between Dehirdan and shaly and siltstone layers of Band-e-Chaker Units. Coral marly limestone which lies at the base of Band-e-Chaker Unit is 12 m thick (Plate 1; Fig. 3). The contacts between these units are conformable. This stratigraphic section includes platy, solitary, and head corals. Based on Insalaco (1998), this sequence could be divided into lower and upper parts as the Kermestan section; the lower part begins with dominancy of platestone fabric and continues to upper part with presence of platestone–domestone fabric. In this stratigraphic section there exist 15 genera. Some corals like T. roasendai, Acanthastraea (Isophyllastraea) madeirensis, Caulastraea sp., C. progoensis, Heliastraea (Heliastraea) craterophora, O. eocenica, Scapophyllia cylindrica, Lithophyllon floriformis persica, Acanthastraea brevis, Acanthastraea sp., H. (Heliastraea) craterophora, Gonipora sp., Porites sp., Astrocoenia gerthi, Tarbellastraea reussiana, P. folium are present, likewise minor constituents such as foraminifera, red algae, bryozoans, gastropods, bivalves and serpulid worms. Among the acquired corals, A. gerthi and L. floriformis persica are indices of Burdigalian and P. folium is index fossil of Early Miocene of the Makran Basin (McCall et al. 1994).
Tejek
The Tejek section is located at 26° 34′ 37.36″ N, 57° and 56′ 5.70″ E near Tejek Village near south side of Band-e-Chaker Syncline (Figs. 1 and 2). The studied section in Tejek is influenced by tectonic activities, and the pure limestone (with CaO concentration of 53.96 % w/w) of Band-e-Chaker Unit is over-thrusted onto shale and sandstone of the Dehirdan Unit. Consequently, the contact between the limestone part and the Dehirdan Unit is a tectonic angular contact. About 30 m of coral-bearing carbonate sequence is exposed with good fossil preservation (Plate 1; Fig. 3). Stratigraphically, the Tejek section is not of interest for this study because the lower contact is a tectonized angular boundary, and the upper contact is not limited by any other younger units. However, we do consider the lithology, faunal content, and age and compare the results with other coral-bearing limestone sections in the area. On the basis of Insalaco (1998), this sequence is classified as domestone. Some examples of the obtained corals from this stratigraphic section are A. gerthi, Caulastraea sp., Cyathoseris infundibuliformis, Favites cerium, Heliastraea (Heliastraeopsis) curta, Acropora pachymorpha, L. nishihirai, Alveopora sp., Porites sp., Goniopora sp., Hydnophora solidior, Monticulastraea astraeoides, Cyphastraea sp., Monticulastraea provincialis, T. roasendai, Thegioastraea multisepta, A. brevis, and Heterogyra meandrinoides; they are totally 22 genera. Minor components in the section are foraminifera, red algae, bryozoans, gastropods, bivalves, and serpulid worms. From the obtained corals, A. gerthi and H. solidior are indices of Burdigalian in the Makran Basin (McCall et al. 1994).
It is worth pointing out that based on distribution of foraminifera in Indo-Pacific and Mediterranean regions (BouDagher-Fadel and Price 2013), the presence of Miogypsina globulina in Tejek, Kermestan, and Irer represents Burdigalian deposits.
Environmental traits
Some paleoecological and environmental characteristics such as surface temperature, nutrient conditions, photic conditions, water depth, water energy, substrate conditions, and salinity are investigated and discussed. Based on Insalaco (1998) classification, different assemblages of corals are identified at the sections: platestone, platestone–domestone, and domestone (Plate 2). The platestone assemblages are present in the basal parts of Kermestan and Irer sections and mostly include agariciidae corals such as Pavona, Cyathoseris, and many other platy corals. The platestone–domestone assemblages including corals such as Caulastraea, Heliastraea, and Acanthastraea are laid on platestone fabric of Kermestan and Irer sections, and domestone fabric is present in the whole interval of Tejek section; most of them are dome-shaped poritidae–faviidae group like Goniopora, Porites, Tarbellastraea, Favites, Heliastraea, Thegioastraea, and Plesiastraea.

Coral fabrics in the studied sections. Domestone fabric in Tejek (1 and 2) and Kermestan (4); platestone–domestone fabric in Irer (5); and platestone fabric in Kermestan (3) and Irer (6)
Temperature
Temperature is one of the most significant factors for the life process and distribution of organisms (Flügel 2010). It controls CO2 solubility and CaCO3 saturation and thereby influences the amount of energy required to calcify and maintain skeletal carbonate (Mutti and Hallock 2003). Definitely, the distribution of zooxanthellate corals is controlled by conditions like sea surface temperature. Bosellini and Perrin (2008) suggest that although coral diversity patterns are related to a complex interplay between environmental factors and paleobiogeography, they are considerably controlled by climate variability and sea surface temperature. According to McCall et al. (1994), if the Makran coral fauna is compared with modem temperature patterns of generic richness of zooxanthellate corals in the Indo-Pacific described by Rosen (1984), it would indicate a mean minimum sea surface paleotemperature of at least 18 °C. Likewise, based on the method applied by Bosellini and Perrin (2008), considering the richness of zooxanthellate corals in the Makran Basin (McCall et al. 1994), sea surface temperature is inferred as 18–20 °C in Aquitanian–Burdigalian of Makran Basin. In this method, mean annual sea surface temperature is calculated using square root of zooxanthellate corals and an experimental presented curve (Bosellini and Perrin 2008) (Fig. 4). We also utilized this method to estimate the sea surface temperature in the three studied sections. It is obvious that the paleontological record is not as complete as the diversity at their lifetime (Bosellini and Perrin 2008) but at least indicates the least surface temperature. The diversities of symbiont-bearing corals genera in Tejek, Kermestan, and Irer (Table 3) are 22, 23, and 15, respectively. Therefore, based on the mentioned method, the least temperature range is from 19 to 20 °C in all sections (Fig. 4).

Main annual sea surfaces temperature curve (presented in Bosellini and Perrin 2008); the red circle shows the temperature range of the studied sections
This interpretation is confirmed by the presence of coralline red algae and larger hyaline foraminifera such as Amphistegina, Operculina, and Miogypsina co-occurring in the Tejek, Kermestan, and Irer sections (Plate 3) and indicates that carbonate successions are deposited in a tropical–subtropical environments (Brandano and Corda 2002). In sum, it seems that the studied carbonate corals are precipitated in a subtropical condition (Mutti and Hallock 2003; Flügel 2010; Wilson and Vecsei 2005).

Different minor constituents in the studied sections: 1 the presence of hyaline benthic foraminifera (Tejek section); 2 bioerosion traces (B.E) and red algae remains (R.A) (Irer section); 3, 6, 8 encrusting foraminifera (Acervulina sp.): Kermestan section (3) and Tejek section (8); 4 presence of red algae in Irer section; 5 large benthic foraminifera and an encrusting red algae on it (Kermestan section); 7 encrusting foraminifera and algae and red algae on an individual coral (Tejek section); 9 serpulid bioerosion trace in an Alveopora sp. (Tejek)
Nutrient conditions
Trophic conditions in marine environments are categorized as oligotrophic, mesotrophic, eutrophic, and hypertrophic in an increasing nutrient content (Mutti and Hallock 2003). Different components that could be used to infer nutrient conditions, such as zooxanthellate corals, foraminifera, corallinacea, bryozoans, and mollusk are present in western Makran sections (Tejek, Kermestan, Irer).
Zooxanthellate corals together with many larger foraminifera host symbiotic microalgae (Rasser et al. 2005). Generally, corals are supposed as light-dependant organisms that thrive in clear nutrient poor oligotrophic waters (e.g., Payros et al. 2010; Racey 2001), but different trophic conditions could be considered for zooxanthellate corals (Tomascik and Sander 1985; Hallock and Schlager 1986).
Lee (2006) concludes that the population density of the zooxanthellae is controlled by systematic nitrogen limitation within the host. When the level of external nitrogen increases (for example when conditions become fully eutrophic) (Hallock and Schlager 1986; Hallock 2001), the zooxanthellae outgrow their hosts and the host loses control over its symbiotic algae (Lee 2006) and symbiotic animals cannot compete with faster growing macroalgae (Birkeland 1987).
In the oligotrophic and slightly mesotrophic shallow waters of tropical to subtropical environments, the main carbonate producers are LBF, zooxanthellate corals, and coralline red algae (Langer et al. 1997; Hallock 2001; Halfar et al. 2004; Pomar et al. 2004; Brandano et al. 2009a, 2009b). In mesotrophic conditions, nutrient level is intermediate and light penetration is sufficient to support prolific calcareous algal production (Mutti and Hallock 2003); in this condition, bioeroding fauna is abundant and bioerosion rates are also promoted (Perrin 2002; Mutti and Hallock 2003; Hallock and Schlager 1986; Hallock 1988; James 1997; James et al. 1999).
To infer nutrient conditions some points are considered; filter-feeders are organisms which are controlled by nutrient input and consequently water transparency and availability to nutrients (Brandano 2001; Mateu-vicens 2007). Bivalves, sponges, bryozoans, some polychaetes (serpulid worms), and many other bioeroders are among filter-feeders (Sammarco and Michael 1990; Cosovic et al. 2004; Key et al. 2010). Therefore, boring hard substrate by bioeroders can provide an optimized condition for them in an environment with enhanced nutrient availability. As well, bioerosion and encrustation, abundance of bryozoans and decreasing LBF could be in virtue of higher nutrient availability (Morsilli et al. 2012). Additionally, common occurrence of encrusting biota suggests enhanced trophic levels that reach mesotrophic conditions, with competition for the substrate as the main limiting factor (Mutti and Hallock 2003). Also, water transparency and light penetration are affected by terrigenous discharge, nutrient input, dissolved organic matter, etc. (Morsilli et al. 2012). An increase of siliciclastic sediments can change water chemistry and enhance nutrients, as well as increased suspended matter in the water column results in reduction of light penetration (Flügel 2010).
The numbers of heterotrophic groups such as mollusks, bryozoans, serpulids, and relatively high abundance of encrusting bryozoans, red algae, and encrusting foraminifera (Acervulina sp., Miniacina sp.) in coral assemblage accompanied by enhanced bioerosion suggest the tendency of assemblage from oligotrophic conditions to mesotrophic conditions (Plate 3).
Tejek section is a coral-bearing sequence made up of pure limestone. Filter-feeders’ bio-eroding traces such as bivalves (Gastrochaenolites isp.) and serpulids (Trypanites isp.) are noticeably observed. As well, encrusting organisms including foraminifera (Acervulinids) and red algae are present in the section. Since Tejek section is made up of pure limestone, no evidence of siliciclastic input is seen. Considering the high dominancy of massive corals and domestone fabrics (higher calcification rate) in Tejek section, the presence of symbiont-bearing LBF and lithological features of the section (pure limestone deposits) that represent low turbidity and lack of siliciclastic input, oligotrophic condition is dominant, but accompaniment with bioeroding features such as boring traces created by serpulid worms and bivalves and also the presence of encrusting organisms like encrusting foraminifera and red algae, the oligotrophic to slightly mesotrophic conditions are considered for Tejek section. In very oligotrophic waters, physiologic function of corals may not be in the highest levels in different aspects; for example, corals growth may not be in its maximum condition (Dunn et al. 2012). Therefore, considering mixotrophy of corals (Hallock 1997), oligotrophic to slightly mesotrophic condition can provide an optimized condition for calcification and coral growth.
Kermestan and Irer sections are composed of marly limestone and the ratio of domestone/platestone and are different from Tejek section. In these sections, noticeable bioerosions traces created by filter-feeders’ activities such as bivalves (Gastrochaenolites isp.) and serpulid worms (Trypanites isp.) are observed. Encrusting organisms including foraminifera (Acervulinids) and red algae are also abundant (Plate 3), while LBF are less abundant. As already mentioned, lithologically, Kermestan and Irer sections are composed of marly limestone; accordingly, the presence of silts and detrital sediments is clearly seen; such condition could be observed through the sequences. So, it seems that water transparency in Kermestan and Irer sections is reduced. Therefore, based on Morsilli et al. (2012) and Flügel (2010), water transparency is concordant with enhanced nutrients availability. Totally, considering the presence of filter-feeders and bioeroders, normal presence of encrusters as an indicator of mesotrophic condition (Mutti and Hallock 2003), and tendency of red algae to be present in mesotrophic condition (Bassi 2005), it seems that the dominant nutrient condition in Kermestan and Irer had been mesotrophic. Considering the low ratio of domestone/platestone in Kermestan and Irer sections, one of the reasons that can lower calcification is enhanced nutrient availability (e.g., mesotrophic condition); such condition could result in lower regeneration, lower skeletal density, and eventually lower calcification (Hallock and Schlager 1986; Stambler et al. 1991; Loya et al. 2004; Fabricius 2005; Dunn et al. 2012).
Photic conditions and water depth
Photosynthesis-bearing symbionts are directly related to light condition; this condition causes dependency of some organisms to light. The zooxanthellate corals, LBF, and red algae are among the light-dependent organisms in Tejek, Kermestan, and Irer sections. The zooxanthellate corals as the main constituents of the studied sections are very sensitive to light conditions (Trench 1981; Cairns 1999). The light intensity and penetration at the sea floor is controlled by water transparency and water depth (Reiss and Hottinger 1984; Hallock 1987; Renema and Troelstra 2001; Beavington-Penney and Racey 2004; Novak et al. 2013). The light penetration is influenced by some factors (such as clastic sediments input, nutrients input, dissolved organic matters) which control water transparency (Morsilli et al. 2012).
On the basis of lithological analysis in the Kermestan, Irer, and Tejek sections, it seems that the water transparency has been different in the studied sections. In the Tejek section that is composed of pure limestone, high transparency condition has been prevailed. But, in Irer and Kermestan section which are composed of marly limestone to limestone and the presence of siltstone and some clastic deposits are obvious, the water transparency has not been high; such conditions have been prevailed through Irer and Kermestan sequences and affected the biotic characteristics.
Many authors (e.g., Hallock and Schlager 1986; Lee 1999; Pomar 2001; Flügel 2010) investigated on light classification of water column and used different terms such as euphotic, mesophotic, oligophotic, and disphotic/aphotic. in a decreasing way. Corals appear in variety of shapes; based on the situation of corals and light penetration in different depths, light level influences the shape of corals (Plate 4) that host zooxanthellae (Baker and Weber 1975; Graus and Macintyre 1976; Huston 1985; Barnes and Hughes 1988). The morphology of corals is influenced by light transparency. In reduced light, the shape of corals may change to maximize light catchment (Titlyanov et al. 1996). To optimize light catchment, corals become platy foliaceous (Plate 4) in higher depths and would be developed at and below 4 % of surface irradiation (Graus and Macintyre 1976; Barnes and Hughes 1988). In such conditions, according to Hallock and Schlager (1986), platy corals are under 4–20 % of surface light intensities and as a mesophotic zone may be distinguished between the euphotic and the oligophotic zones (Pomar 2001) (Fig. 5). Based on Hallock and Schlager (1986) in high light levels, head corals live in more than 20 % of surface light intensities and branching corals are present in more than 60 % of surface light intensities.

Some corals with different features: 1, 2, 3 platy corals with projections on them: Tejek section (1) and Kermestan section (2, 3); 4, 9 irregular growth form of individual corals: Irer section (4), (Kermestan) (9); 5 transverse view of a foliaceous coral (Irer section); 6 serpulid worm behind a platy coral (Irer section); 7 growth of a coral larva on upper surface of an individual coral (Tejek section); 8 growth of an individual coral on a head coral (Irer section); and 10 extensive bioerosion on an individual coral (Kermestan section); scale bars are 1 cm

Light classification, based on light intensity and penetration in seawater column (presented in Morsilli et al. 2012)
In Kermestan and Irer sections, platy foliaceous corals are abundant (Plate 4) in the lower parts, while this kind of corals could not be observed directly in the depositional sequence of Tejek, although some reworked platy corals may be seen in this area. In Kermestan, Irer, and Tejek sections, head corals are present in dome-shaped, massive, and hemispherical forms. It should be noted that branching corals are also present in very low amounts in Tejek section. Based on the distribution of platy corals, head corals, and branching corals, euphotic to slightly mesophotic condition has been prevailed in Tejek section, and mesophotic to euphotic condition has been dominant in Irer and Kermestan sections. Some carbonate constituents in carbonate successions are photo-independent biota that are not controlled by photo (Pomar 2001; Flügel 2010) such as bryozoans, bivalves, gastropods, crinoids, and majority of individual corals. Many authors have reported different deposition depths for different components (Table 4) that are also present in the studied sections.
Poritidae and faviidae corals are among the constituents of all the studied sections especially in domestone intervals. Accompanied with these hemispherical and dome-shaped corals are some LBF such as Amphistegina, Miogipsina, and Operculina. in all sections. Agraciidae corals and some other platy and foliaceous corals such as Acanthastraea are abundant in Kermestan and Irer sections. Furthermore, corallinacean red algae are another constituent of the studied sections.
Based on the Table 4, at the first glance, we observe that different depths are proposed for different groups. Therefore, transparency, light penetration and other factors should be considered. Transparency and light penetration are the most important factors in water depth definition. Transparency increases the light penetration, therefore, photic zones (euphotic, mesophotic, oligophotic) continue to deeper depths, while in less transparent water, the photic zones are limited to shallower parts (Fig. 6). In this extensive range from transparent water to turbid water, a defined depth may represent euphotic, mesophotic or oligophotic zone. Moreover, based on water transparency, a taxon (e.g., corallinacean red algae, Leptoseris) which is present in a defined photic zone can occur in various depths (Fig. 6).

Changes of different photic divisions in transparent and turbid water. The black pentagons show that all of the photic zones could be present in the same depth in shallower parts; the red stars (e.g., Leptoseris in mesophotic zone) and the pink drops (e.g., corallinacean red algae near the limit of light penetration) indicate that depending on water transparency, the same taxons could be present in different depths
Based on Hallock (1987, 1988), and considering trophic resources and reduction of illumination with water depth, Sanders and Baron-Szabo (2005) have presented a chart that could be used for defining the depth range in different conditions (Fig. 7). Accordingly, the depth ranges are plotted on the mentioned chart. Tejek section could be deposited in a depth range between 12 and 85 m, and Kermestan and Irer sections could be deposited in 5 to 62 m depth range.

Depth range of corals in different trophic conditions (adopted from Sanders and Baron-Szabo 2005). The curves are based on light intensities proposed by Hallock and Schlager (1986); <1 % minimal photosynthesis, 1–4 % little or no coral, 4–20 % platy corals, >20 % head corals, >60 % branching corals. The honeycomb filled area shows Kermestan and Irer sections depth range, and the break layered filled area shows Tejek section depth range
Water energy
Variable components of study sections and their morphological and textural traits are essential factors to define water energy level. Hydrodynamic energy also plays a role in the development of coral communities, and it could be estimated from the coral morphotypes (Tomás et al. 2008). Platy corals are characteristic of environments with low light penetration and/or reduced water energy (Schuster and Wielandt 1999; Esteban 1996; Riegl and Piller 2000; Bosellini et al. 2001, 2002). The domestone facies of Capo Testa (Italy) based on growth morphologies and sediment textures indicate relatively high hydrodynamic energy (Brandano et al. 2010). On the other hand, in the western Taurides of Turkey, the diversified colonies of massive, domal, hemispherical, and globuler forms with a lesser extent by branching colonies, dominated by Faviid and Heliastreid corals (Tarbellastraea, Heliastraea, Favites, Favia, Caulastraea, Aquitanastarea, Cladocora), Porites and Stylophora represent a marine environment with moderate to high water energy (Karabiyikoglu et al. 2005). Likewise, Yazdi et al. (2012) attributed the dome-shaped and massive corals like Tarbellastraea, Favites, Porites, Goniopora, and Siderastrarea. to high-moderate water energy.
Agraciidae such as Leptoseris and Cyathoseris and other kinds of platy corals are commonly distributed in the basal parts (indicating more depth) of coral-bearing sequences in Kermestan and Irer sections; therefore, platy corals accompanied with or without corallinacean red algae represent low energy water in the mentioned sections. Some of the platy corals implying low energy water are as follows: A. (Acanthastraea) echinata, L. floriformis persica, Larterophyllia cf. turriformis, Leptoseris sp., A. brevis, S. cylindrica, Boninastraea boninensis, C. infundibuliformis, A. (Isophyllastraea) madeirensis, Fasciatiphyllia, Hydnophyllia, O. eocenica, Acanthastraea (Isophyllastraea), and Hydnophyllia cerebriformis. These platy corals are situated in the platestone fabric.
Diversified massive corals such as Favites sp., T. reussiana, Porites sp., Astroria sp., T. roasendai, H. (Athecastraea) boehmi, Oulophyllia irradians, P. folium, Alveopora sp., C. progoensis, H. (Heliastraeopsis) curta, Phyllocoeniella archiaci, and Caulastraea are present in Tejek, Kermestan, and Irer sections. These dome-shaped massive corals are from domestone fabric of Tejek section and platestone–domestone fabric of Irer and Kermestan sections. Therefore, coral communities with hemispherical, massive, and domal colonies usually represent high to moderate energy (Brandano et al. 2010; Karabiyikoglu et al. 2005; Yazdi et al. 2012).
Substrate conditions
Substrate type plays an important role in controlling the distribution of biotic communities (Hallock and Schlager 1986; Pomar and Ward 1995, 1999, Pomar et al. 2004; Wilson and Vecsei 2005). The composition and type of carbonate substrates depend on depositional, biological, and diagenetic factors, and on the other hand, the substrate type affects feeding modes as well as the penetration and attachment (Flügel 2010). To investigate different characteristics of the study sections in this regard, some carbonate constituents, influenced by environmental conditions, were analyzed.
Scleractinian corals are the main constituents of the studied sections, and corallinacea, bivalves, bryozoan, foraminifera, serpulid worms are subordinate ones. Bioerosion is vastly distributed in the studied sections (Plates 3 and 4). The encrusting coralline algae are indicative of different substrate types in the studied areas; some of them are encrusted on sediment surfaces which show that they are developed over soft substrates (Bassi 2005) while they require low substrate mobility and hydrodynamic energy (Rasser and Piller 2004). Some coralline algae present in the studied sections are very well preserved and some of them (especially in Kermestan and Irer sections) are slightly broken, so according to Nebelsick and Bassi (2000), the well-preserved red algae should be indicative of quiet water environment with stable substrate and low sedimentation rate. It sounds that presence of some fractures in some red algae of the Kermestan and Irer sections are due to more instability of substrate and deposition of clastic sediments. Some other kinds of coralline algae prefer to encrust coral hard substrate (e.g., Nebelsick and Bassi 2000; Plates 3 and 4).
The presence of some common morphologies, related to soft substrates environment showing sedimentation and/or unstable substrate (Sanders and Baron-Szabo 2005), is as follows: The laminar and encrusting corals with projecting knobs (Kermestan and Irer sections; Plate 4) and some irregular individual corals (Kermestan and Irer sections; Plate 4) are some instances of soft substrate indicatives present in study areas. In some cases, free-living individual corals were situated on soft deposits developed on a flat Porites.
Corals are another and main hard substrate in study areas. They are influenced by boring (Flügel 2010), bioerosion, encrustation (Flügel 2010), and larva settling (Plate 4). Corallinacea, bryozoans, serpulid worms, bivalves, encrusting foraminifera (Acervulina, Miniacina), and gastropods affect and use the corals as hard substrate (Tejek, Kermestan, Irer; Plate 3).
Furthermore, Marl and siltstone sediments as clastic deposits were precipitated in Kermestan and Irer sections, while there were no clastic sediments in Tejek section; they could directly affect the substrate and corlas life. Considering the lithology characteristics, some evidence is observed in the studied fauna of Irer and Kermestan sections (mostly in upper parts) for precipitation. During the clastic sedimentation, some corals adopt themselves with the substrate level and some may be buried under sediments. As a result, stress bands created by pulsed sedimentation could be seen in some coral specimens of Kermestan and Irer sections, while such specimens could not be observed in Tejek section (Plate 5).

Stress bands (white arrows) created by pulsed sedimentation: 1 Goniopora sp. from Kermestan section, 2 individual coral from Kermestan section, and 3 Porites sp. from Irer section. scale bars are 1 cm
Salinity
According to presence of some cations such as Na, Mg, Ca, K, and Sr, salinity is categorized as freshwater, brackish, normal marine, and hypersaline conditions by Flügel (2010), while Mossadegh et al. (2009) divided it into 30–40 (normal salinity), 40–50, and ˃50 psu ranges. As it was previously mentioned, main constituents of three studied areas in western Makran are scleractinian corals, bryozoans, mollusks, and foraminifera (e.g., Miogypsinids, Operculina, Amphistegina, Acervulina, Miniacina). Logan and Cebulski (1970) and Kleypas et al. (1999) as well as Mossadegh et al. (2009) state that corals favorably develop in normal salinity waters in a narrow salinity ranges between 30 and 34 and 40 ‰. The presence of massive, domal, and hemispherical scleractinian corals such as Favites, Favia, and Caulastraea represents normal salinity (Karabiyikoglu et al. 2005). Moreover, occurrence of small and larger benthic foraminifera with hyaline wall indicates normal seawater conditions (Reiss and Hottinger 1984; Hallock and Glenn 1986; Geel 2000; Mossadegh et al. 2009; Mohammadi et al. 2011). Totally considering the distribution of corals, foraminifera, mollusks, etc., the coral-bearing deposits of Tejek, Kermestan, and Irer sections are precipitated in normal salinity conditions.
Discussion and Conclusion
Makran Basin is formed in an active tectonic margin, influenced by Oman subduction (Dolati 2010; Burg et al. 2011). This Basin has experienced different tectonic histories through time. In the studied region, coral-bearing carbonate sequences are distributed around Band-e-Chaker Syncline, especially in its northeastern and southwestern flanks. Based on analysis of faunal content of the studied sections and the fauna presented in McCall et al. (1994)—as well as presence of M. globulina which is index of Burdigalian in Indo-Pacific and Mediterranean realms—the Tejek, Kermestan, and Irer sections are attributed to Burdigalian in age. As it is mentioned in previous parts, numerous environmental factors have affected formation and deposition of the coral-bearing sequences (Table 5). In the following, we will discuss and scrutinize the important environmental traits.
To define the sea surface temperature, some older references were cited; McCall et al. (1994) used the method of Rosen (1984) and calculated the temperature of 18 °C; based on zooxanthellate coral diversity in Makran Basin (McCall et al. 1994), Bosellini and Perrin (2008) calculated the Aquitanian–Burdigalian sea surface temperature of 18–20 °C. As well, the presence of coralline red algae and larger hyaline foraminifera (Brandano and Corda 2002) and paleolatitude of Makran Basin (McCall et al. 1994) are indicative of subtropical condition in the studied sections. To confirm the sea surface temperature, we used the diversity-based temperature curve, presented in Bosellini and Perrin (2008), and 19–20 °C were obtained for all three sections. The temperature ranges of Tejek, Irer, and Kermestan sections are almost the same.
Among the light-dependent organisms in the studied sections, zooxanthellate corals could be very good light intensity indicatives. The factors controlling the water transparency influence the light penetration (Morsilli et al. 2012). Lithological characteristics of Kermestan, Irer, and Tejek show that there are some differences between the water transparencies of the sections. Tejek section is composed of pure limestone, and Kermestan and Irer sections are composed of marly limestone. Based on various morphotypes of scleractinian corals (Hallock and Schlager 1986; Pomar 2001), euphotic to slightly mesophotic zone is prevailed in Tejek section and mesophotic to euphotic zones are present in Irer and Kermestan sections (Fig. 8). In the euphotic zone, head corals are present in all study sections, but in addition, slightly branching corals are as well present in Tejek section. In the mesophotic zone, the platy corals are dominant in Kermestan and Irer sections.

Trophic, photic, and energy conditions in studied sections
There is a general consent about the nutrient level required for coral growth (Dubinsky and Jokiel 1994), but there are still some debates (D’Angelo and Wiedenmann 2014), because nowadays corals with different trophic conditions exist in the world (Tomascik and Sander 1985; Hallock and Schlager 1986). Considering high abundance of massive corals in Tejek section, the presence of LBF and the lithology of the section (pure limestone, showing clear water), oligotrophic condition is prevailed, but accompaniment with bioeroding features implies the oligotrophic to slightly mesotrophic condition (Fig. 8). Kermestan and Irer sections are composed of marly limestone and their bioerosion ratio is more than Tejek section. Likewise, encrusting organism’s activities including foraminifera (Acervulina sp., Miniacina sp.) and red algae represents higher ratio than Tejek section. In this condition, LBF are reduced and the marly limestone deposits confirm the turbidity of the environment. The mesotrophic conditions are considered for Kermestan and Irer sections (Fig. 8). Different trophic levels could differently affect the carbonate deposition. In very oligotrophic nutrient condition, corals’ physiological performance may not be at their highest levels in all aspect; for instance, coral growth rates may not be at their maximum (Dunn et al. 2012; D’Angelo and Wiedenmann 2014). Dunn et al. (2012) believe that slightly increased nutrient levels may result in a better physiological performance of corals such as increased growth. Therefore, it seems that low nutrient levels (oligotrophic) to slightly increased nutrient levels (toward mesotrophic) could maximize the corals growth rate, such as what occurred in Tejek section; it may be an optimized condition for scleractinian zooxanthellate corals as mixotroph organisms.
Hydrodynamic energy plays a role in the development of coral communities and could be estimated from the coral morphologies (Tomás et al. 2008). As mentioned above, platy corals are characteristics of environments with low water energy (Schuster and Wielandt 1999; Esteban 1996; Riegl and Piller 2000; Bosellini et al. 2001, 2002). These kinds of corals including Agraciidae such as Leptoseris, Cyathoseris, and other kinds of platy corals are present in Kermestan and Irer sections. As another groups of corals, head corals represent moderate to higher water energy (Karabiyikoglu et al. 2005; Yazdi et al. 2012). Massive, hemispherical, and dome-shaped corals are present in all sections (Fig. 8). The water energy can control the CO2 content of water. The higher the water motion is, the higher the CO2 content released. Increasing the amount of CO2 dissolved in the water decreases the availability of carbonate (CO3 2−) ions and lowers the saturation of the major shell-forming carbonate minerals (Kleypas et al. 2006). The decreased carbonate ion concentration significantly reduces the ability of reef-building corals to produce their CaCO3 skeletons (Kleypas et al. 2006). So, higher water energy can release more CO2 from the water and enhance pH of water. In this condition, the calcification could be enhanced; this state is present in domestone parts of Tejek, Kermestan and Irer sections. Lower and platestone parts of Kermestan and Irer sections with platy corals indicating low water energy had to have more CO2 content and less pH and subsequently less rate of calcification.
As we know, the lithological features are different in the studied sections. Irer and Kermestan sections consist of marly limestone and Tejek section comprises of pure limestone. Marl and siltstone sediments as clastic deposits were precipitated in Kermestan and Irer sections, while there were no clastic sediments in Tejek section. Besides the lithology characteristics, some evidence could be observed in the biotic content of Irer and Kermestan sections. During clastic sedimentation, some corals adopt themselves with the substrate level. So, stress bands created by pulsed sedimentation could be seen in some coral specimens of Kermestan and Irer sections, while such specimens could not be observed in Tejek section.
Siliciclastic inputs can increase the acidity of water and subsequently may cause dissolution or may decrease carbonate precipitation. On the other hand, the presence of clastic sediments enhances water turbidity and decreases the light penetration. It causes less photosynthesis of symbionts and after that, the calcification rate would be decreased. Totally turbidity of water can (1) increase soft-substrate areas unfavorable to juvenile growth of corals, (2) increase the amount of metabolic energy needed for sediment rejection, (3) physically disturb and abrade coral polyps, (4) partially to completely kill corals by sediment veneering or burial, and (5) make corals more sensible to disease (Dodge et al. 1974; Loya 1976; Dodge and Vaisnys 1977; Dryer and Logan 1978; Lasker 1980; Rogers 1983; Hodgson 1990; Babcock and Davies 1991; Wittenberg and Hunte 1992; Stafford-Smith 1993; Riegl and Branch 1995; Wesseling et al. 1999; Yentsch et al. 2002, Sanders and Baron-Szabo 2005).
Salinity as another environmental factor could be estimated by presence of corals, foraminifera, etc., because they tolerate a defined limitation of salinity. According to the presence of massive, domal and hemispherical scleractinian corals (Karabiyikoglu et al. 2005) and occurrence of small and larger benthic foraminifera with hyaline wall (Reiss and Hottinger 1984; Hallock and Glenn 1986; Geel 2000; Mossadegh et al. 2009; Mohammadi et al. 2011), normal seawater conditions are estimated. In summary, considering the distribution of corals, foraminifera, mollusks, etc., the coral carbonates of Tejek, Kermestan, and Irer sections are precipitated in normal salinity conditions.
References
Abbasi N (2000) Palaeoichnology, lithostratigraphy and sedimentary environments of Roksha and “Vaziri” units (Miocene) in Nikshahr-Ghasr Ghand areas (Makran). Dissertation, University of Shahid Beheshti
Aghanabati A (2004) Geology of Iran. Geological Survey of Iran, Tehran
Babcock R, Davies P (1991) Effects of sedimentation on settlement of Acropora millepora. Coral Reefs 9:205–208
Baker PA, Weber JN (1975) Coral growth rate: variation with depth. Earth Planet: Science Letter 27:57–61
Barnes RSK, Hughes RN (1988) An introduction to marine ecology. Blackwell, Oxford
Bassi D (2005) Larger foraminiferal and coralline algal facies in an Upper Eocene storm influenced, shallow water carbonate platform (Colli Berici, north-eastern Italy). Palaeogeogr Palaeoclimatol Palaeoecol 226:17–35
Beavington-Penney SJ, Racey A (2004) Ecology of extant nummulitids and other LBF: applications in palaeoenvironmental analysis. Earth-Sci Rev 67:219–265
Birkeland C (1987) Nutrient availability as a major determinant of differences among coastal hard-substratum communities in different regions of the tropics. In: Birkeland C (ed) Differences between Atlantic and Pacific tropical marine coastal systems: community structure, ecological processes, and productivity. UNESCO, Paris, pp. 45–90
Bosellini A, Ginsburg RN (1971) Form and internal structure of recent algal nodules (rhodolites) from Bermuda. J Geol 79:669–682
Bosellini FR, Perrin C (2008) Estimating Mediterranean Oligocene-Miocene seasurface temperatures: an approach based on coral taxonomic richness. Palaeogeogr Palaeoclimatol Palaeoecol 258:71–88
Bosellini FR, Russo A, Vescogni A (2001) Messinian reef-building assemblages of the Salento Peninsula (southern Italy): palaeobathymetric and palaeoclimatic significance. Palaeogeogr Palaeoclimatol Palaeoecol 175:7–26
Bosellini FR, Russo A, Vescogni A (2002) The Messinian reef complex of the Salento Peninsula (southern Italy): stratigraphy, facies and paleoenvironmental interpretation. Facies 47:91–112
Bosence DWJ (1983) The occurrence and ecology of recent rhodoliths. In: Peryt TM (ed) Coated grains. Springer, Berlin, Heidelberg, New York, pp. 225–242
BouDagher-Fadel MK, Price GD (2013) The phylogenetic and palaeogeographic evolution of the miogypsinid larger benthic foraminifera. J Geol Soc Lond 170(1):185–208
Bourrouilh-Le Jan FG, Hottinger L (1988) Occurrence of rhodolites in the tropical Pacific: a consequence of mid-Miocene paleo-oceanographic change. Sediment Geol 60:355–367
Brandano M (2001) Risposta fisica delle aree di piattaforma carbonatica agli eventi più significativi del Miocene nell’Apennino centrale. Sapienza University of Rome
Brandano M, Corda L (2002) Nutrients, sea level and tectonics: constrains for the facies architecture of a Miocene carbonate ramp in central Italy. Terra Nov. 14:257–262
Brandano M, Frezza V, Tomassetti L, Cuffaro M (2009a) Heterozoan carbonates in oligotrophic tropical waters: the Attard member of the lower coralline limestone formation (Upper Oligocene, Malta). Palaeogeogr Palaeoclimatol Palaeoecol 274:54–63
Brandano M, Frezza V, Tomassetti L, Pedley M, Matteucci R (2009b) Facies analysis and palaeoenvironmental interpretation of the Late Oligocene Attard Member (Lower Coralline Limestone Formation), Malta. Sedimentology 56:1138–1158
Brandano M, Tomassetti L, Bosellini F, Mazzucchi A (2010) Depositional model and paleodepth reconstruction of a coral-rich, mixed siliciclastic-carbonate system: the Burdigalian of Capo Testa (northern Sardinia, Italy). Facies 56:433–444
Burg JP, Bernoulli D, Dolati A, Muller C, Smit J, Spezzaferri S (2011) Stratigraphy and Structure of the Iranian Makran. In: AAPG International Conference and Exhibition, Milan, pp 23–26
Cairns SD (1999) Species richness of recent Scleractinia. Atoll Res Bull 459:1–46
Cosovic V, Drobne K, Moro A (2004) Paleoenvironmental model for Eocene foraminiferal limestones of the Adriatic carbonate platform (Istrian Peninsula). Facies 50(1):61–75
Crimes TP, McCall GJH (1995) A diverse ichnofauna from Eocene-Miocene rocks of the Makran range (S.E. Iran). Ichnos 3:231–258
D’Angelo C, Wiedenmann J (2014) Impacts of nutrient enrichment on coral reefs: new perspectives and implications for coastal management and reef survival. Curr Opin Environ Sustain 7:82–93
Dodd R, Stanton R (1990) Paleoecology: concepts and application. Wiley-Interscience Publication, Chichester
Dodge RE, Vaisnys JR (1977) Coral populations and growth patterns: responses to sedimentation and turbidity associated with dredging. J Mar Res 35:715–730
Dodge RE, Aller RC, Thomson J (1974) Coral growth related to resuspension of bottom sediments. Nature 247:574–577
Dolati A (2010) Stratigraphy, structural geology and low-temperature thermochronology across the Makran accretionary wedge in Iran. Dissertation, Swiss institute of technology
Dryer S, Logan A (1978) Holocene reefs and sediments of Castle Harbour, Bermuda. J Mar Res 36:399–425
Dubinsky Z, Jokiel PL (1994) Ratio of energy and nutrient fluxes regulates symbiosis between zooxanthellae and corals. Pac Sci 48:313–324
Dunn JG, Sammarco PW, LaFleur G Jr (2012) Effects of phosphate on growth and skeletal density in the scleractinian coral Acropora muricata: a controlled experimental approach. J Exp Mar Biol Ecol 411:34–44
Esteban M (1996) An overview of Miocene reefs from Mediterranean areas: general trend and facies models. In: Franseen EK, Esteban M, Ward WC, Rouchy J (eds) Models for carbonate stratigraphy from Miocene reef complexes of Mediterranean region. Concepts in sedimentology and paleontology, pp 3–53
Fabricius K (2005) Effects of terrestrial runoff on the ecology of coral reefs: review and synthesis. Mar Poll Bull 50:125–146
Falcon NL (1947) Raised beaches and terraces of the Iranian Makran coast. Geogr J 109-110:149–151
Farhoudi G, Karig DE (1977) Makran of Iran and Pakistan as an active arc system. Geology 5:664–668
Flügel E (2010) Microfacies of carbonate rocks, analysis, interpretation and application. Springer, Berlin
Fricke HW, Schumacher H (1983) The depth limits of Red Sea stony corals: an ecophysiological problem (a deep diving survey by submersible). Mar Ecol 4:163–194
Geel T (2000) Recognition of stratigraphic sequences in carbonate platform and slope deposits: empirical models based on microfacies analysis of Paleogene deposits in south-eastern Spain. Palaeogeogr Palaeoclimatol Palaeoecol 155:211–238
Graus RR, Macintyre IG (1976) Light control of growth form in colonial reef corals: computer simulation. Science 193:895–897
Halfar J, Godinez-Orta L, Mutti M, Valdez-Holguin J, Borges J (2004) Nutrient and temperature controls on modern carbonate production: an example from the Gulf of California, Mexico. Geology 32:213–216
Hallock P (1987) Fluctuations in the trophic resource continuum: a factor in global diversity cycles? Paleoceanography 2:457–471
Hallock P (1988) The role of nutrient availability in bioerosion: consequences to carbonate buildups. Palaeogeogr Palaeoclimatol Palaeoecol 63:275–291
Hallock P (1997) Reefs and reef limestones in earth history. In: Birkelund C (ed) Life and death of coral reefs. Chapman & Hall, New York, pp. 13–42
Hallock P (2001) Coral reefs, carbonate sediments, nutrients and global change. In: Stanley GD (ed) The history and sedimentology of ancient reef systems. Kluwer Academic Publishing/Plenum, New York, pp. 387–427
Hallock P, Glenn EC (1986) Larger foraminifera: a tool for paleoenvironmental analysis of Cenozoic carbonate depositional facies. PALAIOS 1:55–64
Hallock P, Schlager W (1986) Nutrient excess and the demise of coral reefs and carbonate platforms. PALAIOS 1:389–398
Hodgson G (1990) Tetracycline reduces sedimentation damage to corals. Mar Biol 104:493–496
Huston M (1985) Variation in coral growth rates with depth at Discovery Bay, Jamaica. Coral Reefs 4:19–25
Insalaco E (1998) The descriptive nomenclature and classification of growth fabrics in fossil scleractinian reefs. Sediment Geol 118:159–186
Iryu Y, Nakamori T, Matsuda S, Abe O (1995) Distribution of marine organisms and its geological significance in the modern reef complex of the Ryukyu Islands. Sediment Geol 99:243–258
Jacob KH, Quittmeyer RC (1979) The Makran region of Pakistan andiron: trench-arc system with active plate subduction. In: Earah A, DeJong K (eds) Geodynamics of Pakistan. Special Publication of Geological Survey of Pakistan, Quells, pp. 305–317
James NP (1997) The cool-water carbonate depositional realm. In: James NP, Clarke J (eds) Cool-Water Carbonates. SEPM Special Publication, pp 1–20
James NP, Collins LB, Bone Y, Hallock P (1999) Rottnest shelf to Ningaloo Reef cool-water to warm-water carbonate transition on the continental shelf of Western Australia. J Sediment Res 69:1297–1321
Karabiyikoglu M, Tuzcu S, Ciner A, Deynoux M, Orcen S, Hakyemez A (2005) Facies and environmental setting of the Miocene coral reefs in the late-orogenic fill of the Antalya Basin, western Taurides, Turkey: implications for tectonic control and sea-level changes. Sediment Geol 173:345–371
Key MM, Schumacher GA, Babcock LE, Frey RC, Heimbrock WP, Felton SH, Cooper DL, Gibson WB, Scheid DG, Schumacher SA (2010) Paleoecology of commensal epizoans fouling Flexicalymene (Trilobita) from the Upper Ordovician, Cincinnati arch region, USA. J Paleontol 84(6):1121–1134
Kleypas JA, Buddmeier RW, Archer D, Gattuso JP, Langdon C, Opdyke BN (1999) Geochemical consequences of increased atmospheric carbon dioxide on coral reefs. Science 284:118–120
Kleypas JA, Feely RA, Fabry VJ, Langdon C, Sabine CL, Robbins RR (2006) Impacts of ocean acidification on coral reefs and other marine calcifiers: a guide for future research, Report of a workshop held 18–20 April 2005. NSF, NOAA and the US Geological Survey, St Petersburg
Langer MR, Silk MT, Lipps JH (1997) Global ocean carbonate and carbon-dioxide production: the role of reef foraminifera. J Foramin Res 27:271–277
Lasker HR (1980) Sediment rejection by reef corals: the roles of behavior and morphology in Montastrea cavernosa (Linnaeus). J Exp Mar Biol Ecol 47:77–87
Lee RE (1999) Phycology. Cambridge University Press, Cambridge
Lee JJ (2006) Algal symbiosis in foraminifera. Symbiosis 42:63–75
Logan BW, Cebulski DE (1970) Sedimentary environments of Shark Bay, Western Australia. Memoir of the American Association of Petroleum Geologists 13:1–37
Loya Y (1976) Effects of water turbidity and sedimentation on the community structure of Puerto Rico corals. Bull Mar Sci 26:450–466
Loya Y, Lubinevsky H, Rosenfeld M, Kramarsky-Winter E (2004) Nutrient enrichment caused by in situ fish farms at Eilat, Red Sea is detrimental to coral reproduction. Mar Pollut Bull 49:344–353
Mateu-Vicens G (2007) Paleoecological and sedimentological analysis of Neogene carbonate platforms in the Western Mediterranean. Dissertation, Universitat de les Illes Balears, Palma de Mallorca, Spain
McCall GJH (1985) Explanatory Text of the Minab Quadrangle Map, scale 1:250,000: Geological Survey of Iran, Tehran
McCall GJH (1997) The geotectonic history of the Makran and adjacent area of southern Iran. J Asian Earth Sci 15(6):517–531
McCall GJH, Kidd RGW (1982) The Makran, Southeastem Iran: the anatomy of a convergent plate margin active from the Cretaceous to present. In: Leggett JK (ed) Trench-Forearc geology: sedimentation and tectonics on modern and ancient active plate margins. Blackwell Scientific Publications, Geological Society of London Special Publications, Oxford, pp. 387–397
McCall GJH, Rosen B, Darrell J (1994) Carbonate deposition in accretionary prism settings: early Miocene coral limestones and corals of the Makran mountain range in southern Iran. Facies 31:141–178
Mohammadi E, Safari A, Vaziri-Moghaddam H, Vaziri MR, Ghaedi M (2011) Microfacies analysis and paleoenviornmental interpretation of the Qom Formation, south of the Kashan, central Iran. Carbonate Evaporite 26:255–271
Morsilli M, Bosellini FR, Pomar L, Hallock P, Aurell M, Papazzoni CA (2012) Mesophotic coral buildups in a prodelta setting (late Eocene, southern Pyrenees, Spain): a mixed carbonate-siliciclastic system. Sedimentology 59:766–794
Mossadegh ZK, Haig DW, Allan T, Adabi MH, Sadeghi A (2009) Salinity changes during Late Oligocene to Early Miocene Asmari formation depositions, Zagros Mountains, Iran. Palaeogeogr Palaeoclimatol Palaeoecol 272:17–36
Mutti M, Hallock P (2003) Carbonate systems along nutrient and temperature gradients: some sedimentological and geochemical constraints. Int J Earth Sci 92:465–475
Nebelsick J, Bassi D (2000) Diversity, growth-forms and taphonomy: key factors controlling the fabric of coralline algal dominated shelf carbonates. In: Insalaco E, Skelton P, Palmer T (eds) Carbonate platform systems: components and interactions. Geological Society of London, Special volume, London, pp. 89–107
Niazi MH, Shimamura H, Matsu’ura M (1980) Microearthquakes and crustal structure off the Makran coast of Iran. Geophys Res Lett 7(5):297–300
Novak V, Santodomingo N, Rösler A, Martino ED, Braga JC, Taylor PD, Johnson KG, Renema W (2013) Environmental reconstruction of a late Burdigalian (Miocene) patch reef in deltaic deposits (East Kalimantan, Indonesia). Palaeogeogr Palaeoclimatol Palaeoecol 374:110–122
Payros A, Pujalte V, Tosquella J, Orue-Etxebarria X (2010) The Eocene storm-dominated foralgal ramp of the western Pyrenees (Urbasa–Andia Formation): an analogue of future shallow-marine carbonate systems? Sediment Geol 228:184–204
Perrin C (2002) Tertiary: the emergence of modern reef ecosystems. In: Kiessling W, Flügel E, Golonka J (eds) Phanerozoic reef patterns. SEPM, Special Publication, Tulsa, pp. 587–621
Perrin C, Bosence D, Rosen B (1995) Quantitative approaches to palaeozonation and palaeobathymetry of corals and coralline algae in Cenozoic reefs. In: Bosence DWJ, Allison PA (eds) Marine palaeoenvironmental analysis from fossils. Geol Soc, Special Publication, pp 181–229
Peterson LW, Rudzinskas KK (1982) (Cartography): explanatory text of the Taherui quadrangle map 1:250000. Geological Survey of Iran, Tehran
Pomar L (2001) Ecological control of sedimentary accommodation: evolution from a carbonate ramp to rimmed shelf, Upper Miocene, Balearic Islands. Palaeogeogr Palaeoclimatol Palaeoecol 175:249–272
Pomar L, Ward WC (1995) Sea-level changes, carbonate production and platform architecture: the Llucmajor Platform, Mallorca, Spain. In: Haq B (ed) Sequence stratigraphy and depositional response to eustatic, tectonic and climatic forcing. Kluwer Academic, Dordrecht, pp. 87–112
Pomar L, Ward WC (1999) Reservoir-scale heterogeneity in depositional packages and diagenetic patterns on a reef-rimmed platform, Upper Miocene, Mallorca, Spain. American Association of Petroleum Geology Bulletin 83:1759–1773
Pomar L, Brandano M, Westphal H (2004) Environmental factors influencing skeletal-grain sediment associations: a critical review of Miocene examples from the western Mediterranean. Sedimentology 51:627–651
Racey A (2001) A review of Eocene nummulite accumulations: structure, formation and reservoir potential. J Petrol Geol 24:79–100
Rasser MW, Piller WE (2004) Crustose algal frameworks from the Eocene Alpine Foreland. Palaeogeogr Palaeoclimatol Palaeoecol 206:21–39
Rasser MW, Scheibner C, Mutti M (2005) A paleoenvironmental standard section for Early Ilerdian tropical carbonate factories (Corbieres, France; Pyrenees, Spain). Facies 51:217–232
Reiss Z, Hottinger L (1984) The Gulf of Aqaba: ecological micropaleontology. Springer, Berlin
Renema W, Troelstra SR (2001) Larger foraminifera distribution on a mesotrophic carbonate shelf in SW Sulawesi (Indonesia). Palaeogeogr Palaeoclimatol Palaeoecol 175:125–146
Riegl B, Branch GM (1995) Effects of sediment on the energy budgets of four scleractinian (Bourne 1900) and five alcyonaceans (Lamouroux 1816) corals. J Exp Mar Biol Ecol 186:259–275
Riegl B, Piller WE (1997) Distribution and environmental control of coral assemblages in northern Safaga Bay (Red Sea, Egypt). Facies 36:141–162
Riegl B, Piller WE (2000) Biostromal coral facies—a Miocene example from the Leitha limestone (Austria) and its actualistic interpretation. PALAIOS 15:399–413
Rogers CS (1983) Sublethal and lethal effects of sediments applied to common Caribbean reef corals in the field. Mar Poll Bull 14:378–382
Rosen BR (1984) Reef coral biogeography and climate through the late Cainozoic: just islands in the sun or a critical pattern of islands. In: Brenchley PJ (ed) fossils and Climate. Geol J, Special Issue, pp 201–262
Sammarco PW, Michael J (1990) Large-scale patterns in internal bioerosion of Porites: cross continental shelf trends on the Great Barrier Reef. Mar Ecol Prog Ser 59:145–156
Sanders D, Baron-Szabo RC (2005) Scleractinian assemblages under sediment input: their characteristics and relation to the nutrient input concept. Palaeogeogr Palaeoclimatol Palaeoecol 216:139–181
Schuster F, Wielandt U (1999) Oligocene and Early Miocene coral faunas from Iran: palaeoecology and palaeobiogeography. Int J Earth Sci 88:571–581
Sondhi VP (1947) The Makran earthquake, 28th November 1945: the birth of new islands. Indian Miner 1(3):146–154
Stafford-Smith MG (1993) Sediment-rejection efficiency of 22 species of Australian scleractinian corals. Mar Biol 115:229–243
Stambler N, Popper N, Dubinsky Z, Stimson J (1991) Effects of nutrient enrichment and water motion on the coral Pocillopora damicornis. Pac Sci 45:299–307
Steneck RS (1986) The ecology of coralline algal crusts convergent patterns and adaptative strategies. Annu Rev Ecol Syst 17:273–303
Testa V, Bosence DW (1999) Physical and biological controls on the formation of carbonate and siliciclastic bedforms on the north-east Brazilian shelf. Sedimentology 4:279–301
Titlyanov EA, Titlyanova TV, Leletkin VA, Tsukahara R, van Woesik R, Yamazato K (1996) Degradation and regulation of zooxanthellae density in hermatypic corals. Mar Ecol-Prog Ser 139:167–178
Tomás S, Löser H, Salas R (2008) Low-light and nutrient-rich coral assemblages in an Upper Aptian carbonate platform of the southern Maestrat Basin (Iberian Chain, eastern Spain). Cretac Res 29:509–534
Tomascik T, Sander F (1985) Effect of eutrophication on reef-building corals. I. Growth rate of the reef-building coral Montastrea annularis. Mar Biol 87:143–155
Trench RK (1981) Cellular and molecular interactions in symbioses between dinoflagellates and marine invertebrates. Pure Appl Chem 53:819–835
Veron JEN (2011) Corals: Biology, skeletal deposition, and reef-building. In: Hopley D (ed) Encyclopedia of modern coral reefs, structure, form and process. Springer, pp 275–281
Wesseling I, Uychiaoco AJ, Alino P, Aurin T, Vermaat JE (1999) Damage and recovery of four Philippine corals from short-term sediment burial. Mar Ecol Progr Ser 176:11–15
Wielandt-Schuster U, Schuster F, Harzhauser M, Mandic O, Kroh A, Rogl F, Reisinger J, Liebetrau V, Steininger FF, Piller WE (2004) Stratigraphy and paleoecology of Oligocene and Early Miocene sedimentary sequences of the Mesohellenic Basin (NW Greece). Cour Forsch Inst Senckenberg 248:1–55
Wilson MEJ, Vecsei A (2005) The apparent paradox of abundant foramol facies in low latitudes: their environmental significance and effect on platform development. Eatrh-Sci Rev 69:133–168
Wittenberg M, Hunte W (1992) Effects of eutrophication and sedimentation on juvenile corals. I. Abundance, mortality and community structure. Mar Biol 112:131–138
Yazdi M, Parvanenejad Shirazi M, Rahiminejad AH, Motavalipoor R (2012) Paleobathymetry and paleoecology of colonial corals from the Oligocene–early Miocene (?) Qom Formation (Dizlu area, central Iran). Carbonate Evaporite 27:395–405
Yentsch CS, Yentsch CM, Cullen JJ, Lapointe B, Phinney DA, Yentsch SW (2002) Sunlight and water transparency: cornerstornes in coral research. J Exp Mar Biol Ecol 268:171–183
Acknowledgments
Special thanks are due to Dr. Christian Chaix for providing helpful and constructive comments in confirming corals species identifications. Dr. Ali Bahrami is thanked for his field assistance and useful suggestions. Dr. Ebrahim Mohammadi improved the manuscript and is gratefully acknowledged. The first author is grateful to Dr. M. Allahkarampour Dill, Dr. Ali Ghabishavi, and Dr. and Dr. R. Shabafrooz for their useful discussions. Finally, we would like to thank the anonymous reviewer for insightful comments on the manuscript. This paper was financially supported by University of Isfahan.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Ghaedi, M., Johnson, K. & Yazdi, M. Paleoenvironmental conditions of Early Miocene corals, western Makran, Iran. Arab J Geosci 9, 686 (2016). https://doi.org/10.1007/s12517-016-2712-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12517-016-2712-3