
Abstract
The foraminiferal limestones from the Middle Eocene Central Istrian region illustrate progressive deepening of depositional gradients. Shifting of Lower Cuisian to Upper Lutetian microfacies can be described in terms of a ramp model. The Orthophragminae-bearing parts of the foraminiferal limestones are interpreted in terms of larger foraminiferal faunal associations, planktonic foraminiferal relative abundance, limitations of algal endosymbionts, foraminiferal lamellar thickness and flattening of test shapes. Microfacies I contains the most diverse larger foraminiferal association with a predominance of large, thick nummulitids, assilinids, and asterocyclinids. Microfacies II is characterized by a gradual increase of Orthophragminae diversity and abundance. Nummulitids, equally abundant, are dominated by lenticular and subspherical specimens. The reduction in number of nummulitid specimens with characteristic biconical radiate morphologies, and relative abundance of flattened orthophragminids, characterizes Microfacies III. Scattered biodestructed orthophragminid tests and planktonic foraminifera constitute Microfacies IV, indicating the end of a long-lasting, shallow-marine Adriatic Carbonate Platform regime.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
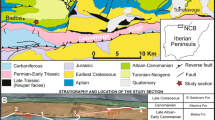
Eocene carbonate deposits are widespread in the central part of the Istrian Peninsula, Croatia and Slovenia (Fig. 1). The Eocene succession transgressively overlies Cretaceous sediments and comprises from the base upwards: brackish-lagoonal deposits of the Liburnian Formation (locally coals occur), platform limestones (known as foraminiferal limestones), Transitional Beds (“Marls with crabs” and “ Subbotina marls”), and basinal sediments (Flysch) (Stache 1889; Salopek 1954; Hamrla 1959, 1987; Šikić 1965; Bignot 1972; Drobne 1977, 1979, 2000; Drobne et al. 1979; Juračić 1980; Drobne and Pavlovec 1991; Drobne et al. 1991; Pavlovec et al. 1991; Ćosović and Drobne 1995, 1998; Ćosović 1996; Matičec et al. 1996; Tari Kovačić 1997; Drobne and Ćosović 1998). This succession provides an example of transition from platform to foreland basin and represents a sedimentary cycle during a general transgressive phase.

Simplified geological map of the Istrian Peninsula (after General geological map of former Yugoslavia, 1:500000), showing the locations of the sections discussed in this paper
This study deals with deposits accumulated during the latest stage of the Eocene carbonate platform. Its demise induced changes in biotic and abiotic environmental characteristics and caused an increase in abundance of orthophragminids and planktonic and smaller benthic foraminifera, whereas the shallow-water foraminifera such as alveolinids, orbitolitids and thick, so-called granulated nummulitids completely disappeared. The term Orthophragminae (Less 1987; Drooger 1993) is an informal denomination of a widely distributed group of larger, bilamellar-perforate foraminifera of Paleocene to Eocene age (i.e. Discocylinids, Asterocyclinids, Orbitoclypeids and Nemkovellas).
Stratigraphical sections (Pićan, Sveti Tom, Ragancini, Šterna, Slavec, and Osp), Late Cuisian—Late Lutetian (SBZ 12–SBZ 16, according to Serra-Kiel et al. 1998) in age, were measured to document vertical and lateral lithological and paleontological variations along the platform to basin transition.
The Eocene sediments have been studied in detail during the last years but less attention has been drawn to the transition from platform (shelf) to basin because (1) continuous shelf-to-basin transitions are not commonly exposed and (2) the lack of data regarding the identification of orthophragminids on a species level and their implementation into a paleoenvironmental reconstruction.
The objectives of this study are: (1) to refine previous stratigraphic and depositional models for the orthophragminid-bearing deposits and place them within a sequence stratigraphic framework; (2) to document the complex depositional processes that operated along this platform before its demise took place; (3) to document changes in sediment dispersal and depositional processes as the platform responded to increasing depositional gradients along the transition from outer platform (ramp) to basinal environment; and (4) to interpret the significance of orthophragminids as paleoenvironmental indicators. Paleogene sediments of the Central Istrian region provide an important, well-exposed analogue for stratigraphic interpretations of depositional processes on other, more poorly exposed or subsurface carbonate platform-to-basin transitions.
Geological setting
The Istrian region forms the northwestern part of the Eastern Adriatic Coast (Slovenia and Croatia). During the Middle Eocene, the Istrian Peninsula was part of the Adriatic Carbonate Platform (AdCP, HKA in Fig. 2). The studied sections are located along the southern and northern margin of the Central Istrian Paleogene Basin (Fig. 1). They range in age (Fig. 3) from Late Cuisian (Osp, Slavec), to Middle (Pićan, Ragancini and Sveti Tom) to Late Lutetian (Šterna).

Location of the Istrian Peninsula within the paleogeographic reconstruction at 46–40 Ma after Butterlin et al. (1993): A Apulia; UMB Umbria basin; MoB Molisa basin; HKA Dinaridic platform; Pl Pelagonian platform; PO Pindos zone; G Gavrovo zone; I Ionian basin; β Bauxite

Correlated schematic stratigraphic logs showing lithology, main fossils and Microfacies types (I–IV). The bold line at the upper parts of the sections represents the contact with overlying “ Subbotina marls” (Sm). The base of each of the sections starts at the contact with underlying foraminiferal limestones ( Flms). TST Transgressive System Tract, MFS Maximum Flooding Surface
The dynamic changes in depositional conditions during the Eocene are considered to reflect third-order depositional sequences described by Tucker (1993). Sedimentologic and paleoenvironmental conditions were conducive to producing some 120 to 200 m of foraminiferal limestones (Drobne and Pavlovec 1991) and at least 350 m of Flysch (Marinčić et al. 1996). The foraminiferal limestones, part of a catch-up system, are accumulated in intertidal to subtidal environments characterized by high biologic productivity. They are part of a Transgressive System Tract (TST). The transitional beds (“ Subbotina marls”) of deep subtidal origin are deposited above the TST and mark progressive deepening. They represent the maximum flooding event, and are characterized by a low rate of sedimentation, changes in microfossil assemblages and characteristic marl contents. The diachronous nature of the maximum flooding surface and associated condensed sections are illustrated in Fig. 3. This third-order sequence (Tucker 1993) is separated from the underlying Upper Cretaceous sediments by a pronounced unconformity associated with a major erosional hiatus.
Methods
The presented results are based on approximately 1,000 thin sections from samples collected in six stratigraphic sections (Figs. 1, 3). Thin section studies included visual determination of lithology; description of grain type, texture, and fossil content; and frequency estimation of bioclasts (Flügel 1982). According to the paleontologic content and sedimentologic features, four microfacies types have been identified. The orthophragminid test morphology was compared with that of nummulitids, assuming that similar morphology indicates the same environmental conditions (Hallock 1979; Hottinger 1983; Hallock and Glenn 1986), as indicated by habitats of present-day relatives of nummulitids (Hallock 1979; Reiss and Hottinger 1984; Hottinger 1997; Hohenegger et al. 2000; Langer and Hottinger 2000; Hohenegger and Yordanova 2001; Renema and Troelstra 2001).
Measurements taken for each thin section include abundance estimations of orthophragminids and nummulitids as well as their test shapes. For this purpose, the orthophragminids were determined from axial or nearly axial sections. The planktonic ratio in foraminiferal associations, the Orthophragminid/Nummulitid ratio (O/N), the Discocyclinid/Asterocyclinid ratio (D/A), and the thickness (T)/diameter (D) ratio in orthophragminids and nummulitids (T/D), were counted in randomly selected areas of 1 cm2 of each thin-section using quantitative method: counting the total number (whole and broken specimens) of each fossil group.
Secondly, the composition of associated faunas was considered (presence of echinoids, molluscs, worm tubes, and bryozoans).
Thirdly, sedimentologic textures (cross-bedding) and further characteristics, such as the presence of glauconite (intraskeletal as foraminiferal chamber infillings or extraskeletal as isolated green to brown grains), burrowing, bioerosion, and incrustations have been considered qualitatively.
The material is housed in the Department of Geology and Paleontology, Faculty of Science in Zagreb, Croatia and in the I. Rakovec Paleontological Institute in Ljubljana, Slovenia.
Facies analysis of middle Eocene carbonate successions
Based on lithology, sedimentary characteristics, recognized “orthophragminid-species”, and associated benthic and planktonic foraminifera, there are four orthophragminid-microfacies in the foraminiferal limestones.
Microfacies I: Nummulitic limestones with orthophragminids
This microfacies is represented by fossiliferous packstones. Locally, the “ Nummulites bank” facies is developed as a consequence of winnowing processes (Šumbir and Pedrovica, in the vicinity of the Pićan section).
This microfacies is characterized by the most diverse foraminiferal fauna (Fig. 6 1; Table 1), comprised of orthophragminids (which constitute about 30% of total biogenic components), nummulitids (about 30%), sphaerogypsinids, rotaliids (heavy ornamented Neorotalia of the N. viennoti Greig group), amphisteginids, asterigerinids, orbitolitids, and alveolinids. Bryozoans and corallinaceans are found also. The family Nummulitidae is represented by assilinids, “operculinids”, and specimens of the genus Nummulites. In some places (“ Nummulites bank” or shoals) they account for 80 to 90% of all skeletal remains. Orthophragminids are identified as specimens of the following groups: Orbitoclypeus chudeaui Schlumberger, O. ramaroi (Samanta), Asterocyclina taramelli (Munier-Chalmas), A. stella (Gümbel), Discocyclina dispansa (Sowerby) and D. archiaci (Schlumberger) (Table 1). Orthophragminids are typically thickly lenticular to subsphaeroidal (T/D ratio between 0.45–0.6) in shape, though some flattened forms (T/D greater than five, always microsphaerical specimens) can be occasionally found. The robustness of shells, expressed by their thick walls and subglobular shape, is evident. The asterocyclinid/discocyclinid ratio shows a predominance of asterocyclinids. Juvenile individuals of larger foraminifera are abundant.

Triangle diagram summarizing where foraminiferal assemblages from three studied. Sections would generally plot according to nummulitids, rotaliids and orthophragminids contents in sediment. This diagram provides an indication of water depth, wave energy, and open marine affinities of the depositional environments of the three sections in the Middle Eocene. Deeper open marine conditions are indicated for faunas plotting toward the apex, assemblages from open platform to open shelf conditions are plot at the central part. The interpretation is based on Hallock and Glenn (1986) study
Two main growth morphologies of corallinaceans have been distinguished: thin laminar crusts (generally 0.5–1 mm thick) and small compact rhodoliths (1–3 cm in diameter, composed of several quite compact crusts). Locally, (Šterna section) algal shoals composed of red algae sands rich in alveolinids occur in which planktonic foraminifera are absent.
The main body of the unit is interbedded with nummulitid/orthophragminid wackestones.
Microfacies II: Discocyclinid-microfacies, lenticular to subsphaerical nummulitid and orthophragminid wackestone/packstone with smaller rotaliids
This microfacies is composed mainly of intensely bioturbated wackestones with local packestones. The diverse foraminiferal assemblage is dominated by orthophragminids (Fig. 6 2; Table 1). Other bioclasts include echinoids, bryozoans, corallinaceans, and planktonic and smaller benthic foraminifera. Locally, packestones are developed with tightly-packed flat discocyclinids, which are variably imbricated. The foraminifera are generally well preserved and do not show abrasion.
Nummulitids usually comprise less than 30% of the total biota in this microfacies, whereas free and fixed rotaliids make up 5–20%. As discocyclinid specimens outnumber (making up to 65%) all the foraminiferal specimens, this microfacies is named the discocyclinid-microfacies. The discocyclinid assemblage includes Dyscocyclina sella (D’Archiac), D. augustae Weijden, D. radians (D’Archiac), D. discus (Rütimeyer), D. trabayensis Neumann, D. dispansa and D. archiaci. Lenticular to ovate specimens (with T/D ratio between 0.4–0.45) of megalosphaeric forms of both orthophragminids and nummulitids predominate. The frequency of asterocyclinids and discocyclinids is more or less equal. The smaller benthic foraminiferal assemblage is relatively diversified, with a slight dominance of Gyroidinella, Asterigerina and Lenticulina specimens. Planktonic foraminifera begin to be present in this microfacies. The groundmass of sediments consists of very fine fragments of larger and smaller foraminifera, which occasionally contain subbotinids. The foraminiferal fauna is generally associated with fragments of corallinaceans, echinoids and crinoid stems. Glauconite grains (mainly foraminiferal chamber infillings) constitute a considerable part of the sediment particles (between 2.5–5%).
Microfacies III: Wackestones with flattened orthophragminids and planktonic foraminifera
The microfacies types in this group are mud-supported with planktonic foraminifera and a sparse, low diversity benthic assemblage dominated by flat Discocyclina and “ Operculina” (Fig. 6 3; Table 1). The sediments are locally burrowed, although horizons devoid of bioclasts and bioturbation are present.
The predominant bioclasts are larger benthic and planktonic foraminifera (e.g. subbotinids account for about 30% of total biomass). Among the larger foraminifera, the orthophragminids are represented by predominantly thin, strongly flattened shells (T/D ratio between 0.2–0.3) of Discocyclina ( D. sella, D. augustae, D. trabayensis, D. discus). The small, biconical, radiate nummulitids (very similar to those of the “ Globulus -Shales” in the Tremp basin, Pyrenean; Luterbacher 1970) with compressed test shapes occur along with remarkably elongated, thin-walled forms of “operculinids”. The larger foraminiferal population is composed mostly of adults, while juveniles are very rare (less than 5% of total biota). Smaller rotaliids seldom account for more than 5% of the total biota. The planktonic foraminiferal content is high; several species indicative for the Morozovella lehneri Zone of the Middle Eocene were determined in the Pićan (samples Pć 1/2239, Zajci 8/95) and Ragancini (sample R-L 5) sections. The planktonic foraminiferal fauna is taxonomically diverse (Table 1) in terms of species richness. Planktonic tests are filled with authigenetic glauconite.
Microfacies IV: Biodestructed orthophragminid wackestones
Biodestructed orthophragminid wackestones to mudstones form an up to 15 m thick unit between the Flysch and the Orthophragminae-bearing limestones. Sediments contain well-preserved crustaceans of Harpactocarcinus sp. (Salopek 1954; Drobne and Pavlovec 1991) and shark teeth. Bryozoans are common, and molluscs including oysters and gastropod molds; ahermatypic corals; and crinoid plates are also found. Detrital quartz is also present. Upward the unit passes into “Subbotina” marls (Transitional Beds: a condensed section rich in planktonic foraminifera).
In thin-sections this microfacies has a dark micritic matrix with fragments of bioclasts (Fig. 6 4). Teredinids, locally abundant (Šterna, Pićan), occur in small pieces of wood substrate that were presumably transported as driftwood and as bed load. Other bioclasts include echinoid fragments, planktonic foraminifera, orthophragminids and “operculinids”.
The wackestones—mudstones of this microfacies type often contain the lowest abundance and diversity of larger foraminifera. Larger foraminifera, nummulitids (“operculinids” and Nummulites specimens) and orthophragminids in particular, are the main recognizable components in this microfacies. Usually, the orthophragminids are very thin (T/D ratio less than 0.2) and specimens are sometimes broken. Rare tiny orthophragminid individuals occur ( D. discus, D. sella); they are small (the average size is by 20% reduced in comparison with those already described), with extremely thin walls, and a number of lateral layers, which indicate that the individuals are adults (more than 7 layers). Fragmentation of delicate Discocyclina tests, including regenerated ones (Fig. 4 1, marked), indicates that at least some of the fractured individuals were living in situ, their fracture being due to biotic processes (not transport breakages). Glauconite constitutes a considerable portion of the grains in the sediment. It occurs as foraminiferal chamber infillings (intraskeletal) and isolated grains (extraskeletal), together making up to about 10% of the sediment. Numerous smaller benthic specimens are dispersed in the matrix, which is partly composed of very small fragments of larger foraminifera.
Planktonics are generally abundant and diversified. The composition of the Pićan and Ragancini faunas are the same as those of Microfacies III. The assemblage from Šterna (samples Štr-1/0, 2/95, 21726) reveals a Middle Eocene age ( M. lehneri and ? Globigerapsis beckmanii Zones, P12 - ?P13). The nannofossils from Šterna (sample Štr-8/5) are poorly preserved and low in terms of species richness and abundances (Table 1); they indicate the same age.
Paleoenvironmental interpretation of microfacies
The distributional pattern of the microfacies types and their ecological gradient is based on four parameters: faunal association, planktonic ratio in foraminiferal associations, endosymbionts in the larger foraminifera, and lamellar thickness and flattening of larger foraminifera tests.
Faunal association (composition, changes, homeomorphs)
The faunal association was analysed with respect to microfacies distribution during the Middle Eocene. However, environments (protected shoals and lagoons) of deposition defined by predominance of alveolinids and orbitolitids are not considered in this study. The comparison of the studied Eocene succession with modern faunal associations (Reiss and Hottinger 1984; Piller 1994; Hohenegger et al. 2000; Hohenegger and Yordanova 2001; Renema and Troelstra 2001; Romero et al. 2002), Middle and Lower Miocene associations (Hottinger 1996; Hallock and Glenn 1986), and Eocene associations (Luterbacher 1970, 1984; Moody 1998) reveal faunal changes along an ecocline.
The grouping of foraminifera based on their living affinities provides an indication of water depth of the depositional environments of the foraminiferal associations. Following the Hallock and Glenn (1986) and Hallock (1999) model, three groups of foraminifera form the end members of a triangular diagram:
-
Group I: larger rotaliids with subspherical tests, with known shallow water affinities, i.e. large, thick nummulitids, sphaerogypsinids, neorotalids and thick amphisteginids (Fig. 7, 1–11).
Fig. 5 
Reconstructed block diagram of the Middle Eocene carbonate ramp showing the microfacies settings and ecological distribution of selected biota. This represents a wave-dominated carbonate ramp with minor storm activity
-
Group II: planktonic foraminifera, thin, flat nummulitids (mostly “ Operculina” sp.), flattened orthophragminids.
-
Group III: smaller rotaliids (free and fixed specimens, including lenticularids, cibicidids, and asterigerinids).

Shallow water, high energy conditions are indicated by microfaunas in the lower left corner, assemblages of restricted environments in the lower right, and deeper, open marine conditions in those of the top. This diagram (see Fig. 4) shows relatively large scatter of data from the Šterna section and good clustering of data from Pićan, whereas the Ragancini data show a bipartite clustering. Data suggest that the Pićan fauna was of deeper marine origin, whereas those from compared sections indicated different environments (transition from open platform and open shelf environments to toe of slope).

Microfacies types of the Middle Eocene Foraminiferal Limestone Formation, Istria (Croatia and Slovenia), all enlarged ×10, incident light. 1 Microfacies I: Nummulitic limestones, Nummulites sp., Discocyclina sp., Gyroidinella sp., Rotalia sp. and miliolids within micrite matrix. Thin-section, sample Pć 5/2229, Pićan section. 2 Microfacies II: Discocyclinid packstone, strongly dominated by Discocyclina ( D. archiaci (Schlumberger), D. augustae Weijden, D. dispansa (Sowerby)), while Asterocyclina sp. and Nummulites sp. occur, too. Thin-section, sample RL 3-2/95, Ragancini section. 3 Microfacies III: Fine-grained wackestone with planktonic foraminifera and discocyclinids D. sella (D’Archiac) loose into micrite matrix. Thin-section, sample Štr 2/1727, Šterna section. 4 Microfacies IV: Scarce discocyclinids and small nummulitids are dispersed within fine grained matrix along with planktonic foraminifera. Thin-section, sample Pć 9F, Pićan section
Changes in faunal associations reflect the differentiation of biotopes into several smaller areas, each controlled by bottom morphology and current patterns (which indirectly produced redistribution of nutrients, i.e. the Safaga Bay; Piller and Pervesler 1989).
Comparisons between Eocene and Recent distributional patterns of larger foraminifera cannot be based on common or phylogenetically related taxa since orthophragminids and assilinids became extinct at the Eocene/Oligocene boundary. These genera were replaced during the mid-Miocene by those taxa which currently inhabit modern, shallow, tropical seas. Some of the described taxa are interpreted by taking into account the similarity of their morpho-functional characteristics with modern forms; others have modern equivalents whose habitat and environmental distribution is well known. Genera like Discocyclina, Orbitoclypeus, Asterocyclina can be regarded as homeomorphs to Cycloclypeus and Baculogypsinoides. The thin, flat tests of Cycloclypeus with an annular arrangement of chambers and a thicker test centre leading to biconvex forms are similar to Discocyclina and Orbitoclypeus. Most species of Cycloclypeus colonise the deepest environments, down to the lower limit of the photic zone, and they crawl on firm substrates (Reiss and Hottinger 1984; Hohenegger et al. 2000; Hohenegger and Yordanova 2001; Yordanova and Hohenegger 2002) but prefer attachment to macroids. Their plate-like forms show a high degree of downslope transport to the deepest zone (below 100 m in Central Pacific islands, ibid) as the result of high buoyancy. The orthophragminids at the lower limit of the photic zone are very flat, and their lateral chamberlets are particularly low in shape. The Baculogypsinoides individuals with 3–4 strong spines live on coral rubble and macroids, and are extremely rare on sandy bottoms in Okinawa (Hohenegger 2000; Hohenegger et al. 2000; Hohenegger and Yordanova 2001; Yordanova and Hohenegger 2002) and Spermonde Archipelago (Indonesia; Renema and Troelstra 2001). Baculogypsinoides spinosus Yabe and Hanzawa is adapted to live in low-light intensities, at maximum depths between 27 m in the outer shelf zone in Indonesia and 55 m in Okinawa. Test form, size, and spines are similar to the genus Asterocyclina (five well defined rays). According to the modern homeomorph asterocyclinids might have lived on firm substrate in high energy environments, along with asterigerinids and amphisteginids (less than 30 m; Hallock 1999). Smaller, globular nummulitid specimens are morphologically similar to Nummulites venosus (Fichtel and Moll), the living representative of the genus Nummulites. The thick-lenticular N. venosus inhabits sandy bottoms on the slope at moderate depths in the euphotic zone (40–80 m).
Planktonic ratio in foraminiferal associations
Planktonic foraminifera have great potential for paleoecological studies (Murray 1991). The Middle Eocene successions of Pićan, Ragancini, and Šterna (Table 1) contain planktonic foraminiferal associations, including species of typical low latitude, subtropical to warm temperate climate provinces (Spezzaferri 1995).
Muricate species (genera Acarinina, Truncorotaloides and Morozovella), and non-muricate Globigerinatheka are considered to have hosted algal symbionts and to have been dispersed within euphotic habitats. On the other hand, Subbotina and Turborotalia species occupied deeper water layers. According to Hemleben et al. (1989: 246), a noticeable presence of planktonic foraminifera reflects the reproductive depth where tests are produced, and consequently allows a minimum depth estimate. This corresponds to the lower limit of the photic zone, as indicated by the disappearance of both symbiont-bearing larger foraminifera and red algae.
The numbers of both genera and species indicate that water depths were predominantly shallow to intermediate at the time of test formation with only two deep-water genera (Table 1).
Algal endosymbionts in larger foraminifera
Symbiont-bearing foraminifera are restricted to the euphotic zone. Their presence of endosymbionts depends on the light conditions in the water column. Quality and quantity of light depends on water turbulence and depth.
Lateral chamberlets (Ferrandez-Canadell and Serra-Kiel 1992; Hottinger 1997, 2000) of all orbitoidal larger foraminifera are interpreted as symbiont greenhouses due to the presence of symbionts in the “lateral” chamberlets of the recent Baculogypsinoides (orthophragminid’s homeomorph). Interseptal lamellar piles, called light lenses, if combined with flattened test shapes, indicate a deep photic habitat (Ferrandez Canadell and Serra-Kiel 1992; Hottinger 1997, 2000).
The best argument for clear water environments in the Eocene carbonate ramp environments is the presence of fixed (sessile) foraminifera ( Gyroidinella, Carpenteria; Fig. 7 2, 10). Such a situation is characterized by a low nutrient content in the water column and a reduced water transparency due to the depth. This relationship supports the idea that the lower limit of the photic zone in the Paleogene Sea was placed much deeper than was expected. This hypothesis is supported by the global geodynamic reconstruction (Butterlin et al. 1993), where the studied area as a part of the AdCP was located at 35° N paleolatitude, allowing stronger insulation and deeper light-penetration than the Istrian area obtains nowadays. Water transparency controlled by the low planktonic density resulted in a transparent water column because the small number of planktonics allowed deeper penetration of the light that would be typical, placing the boundary of the lower photic zone deep in water column, and low sedimentation rate indicated by glauconite, microborings, and a reduced thickness of the sedimentary sequence
It is suggested (Hollaus and Hottinger 1998) that low temperatures do not hamper or suppress the photosynthesis of the symbionts but do suppress their growth and/or reproduction within the host cell because the lower water temperatures do not permit endosymbiotic algae to reproduce or to grow. The endosymbionts stay in the host cell and live and die without reproducing. Subsequently, the foraminifera will stay without endosymbiotic algae and also die. Symbiont-bearing foraminifera are thought to be restricted to a climatic belt bounded by the 15–20 °C winter isotherm of surface water (Murray 1991; Langer and Hottinger 2000).
Lamellar thickness and flattening of larger foraminiferal test shapes
Depth-dependence of larger foraminifera is mirrored by a combination of morphological traits influenced by light intensity and hydrodynamic force. This relationship could be used to estimate facies and depth in ancient warm seas using the concept of functional uniformitarianism (Hottinger 1997). In lamellar-perforate foraminifera, the thinning of the chamber walls results from thinning of each lamella documenting a decrease in the biomineralisation process with increased depth. Lamella thickness and flattening of shell shapes of larger foraminifera are products of water turbulence (Hallock 1979, 1999; Hallock and Glenn 1986; Hottinger 1977, 1997, 2000; Pecheux 1995).
To verify this pattern in orthophragminids we made the following measurements:
1.the thickness to diameter ratios (T/D, D = minimum to maximum diameter).
The orthophragminids from Microfacies I (mainly orbitoclypeids and asterocyclinids), by far the most spheroidal, have T/D ratios in excess of 0.45 (up to 0.6). These foraminifera occur with thick, lenticular assilinids and nummulitids. The discocyclinids, abundant in Microfacies II, appear to have T/D ratio of 0.4–0.45. Orthophragminids associated with planktonic foraminifera and “operculinids” have lower T/D ratios of about 0.2–0.3. There is an easily visible shape difference between individuals with T/D ratios of Indo-Pacific amphisteginids (Hallock 1985, 1999) related to depth preferences.
2. the shell thickness to number of equatorial chamberlets ratio (T/neqc).
The inflated and most robust forms, like specimens of Orbitoclypeus chudeaui which are the most common in Microfacies I, have ratios of about 0.09. On the other hand, flattened specimens like those of the Discocyclina sella group, common in Microfacies III, have ratios of 0.012–0.014. The orthophragminids from Microfacies II have ratios of about 0.03.
The shape trends evidenced by the two analysed ratios in studied orthophragminids exhibit a fairly straightforward depth zonation that is accompanied by fossil associations and sedimentologic characteristics. The shallowest dwelling orthophragminids are those with the highest T/D and T/neqc ratios, which occur in Microfacies I, while those from greater depths (Microfacies III) have lower ratios.
There is a relationship between size, longevity and fecundity (Hallock 1985). It is known that larger foraminifera living in fairly turbulent waters grow relatively large with a T/D ratio of 0.6 to 0.7. They produce up to ten times more offspring per reproduction than deeper dwelling species with a T/D ratio up to 0.3. The ratio between juvenile and adult specimens also reflects this relationship.
Interpretation
The paleogeographic reconstruction of the study area during Eocene times (Butterlin et al. 1993) and in particular its latitudinal position at about 35°N (Fig. 2) places the northwestern part of the AdCP at the northern fringe of the global desert belt, where continental runoff was probably low. Consequently, input of nutrients to the shallow-marine ecosystem was probably limited. The shallow carbonate platform was affected by relative sea-level changes corresponding to the sedimentary sequences of the Paleogene (Marjanac et al. 1998). The Lutetian of the AdCP is characterised by a gyre producing an isometric—oligotrophic condition all around its path (Fig. 2 in Hottinger 1996). The gyre regime resulted in high salinity (from 37.5 to more than 38‰) and a potential surface temperature of 20–25 °C (Barron and Peterson 1991). In the AdCP, oligotrophic conditions prevailed as suggested by a dominance of larger foraminifera.
Under such conditions the foraminiferal limestones rich in larger foraminifera (in particular, conical foraminifera, miliolids, alveolinids, orbitolitids, and nummulitids) were deposited. A constant rate of subsidence caused transgression and the establishment of a carbonate ramp recorded by deposition of nummulitid- to orthophragminid-bearing limestones in the study area. Carbonate production was initially able to keep pace with subsidence. However, an accelerating subsidence led to a reduction of carbonate production and the ramp drowned, as indicated by the vertical transition into hemipelagic marls (“ Subbotina Marls”) and turbiditic flysch. Fossil content, sedimentologic characteristics and geometric relationships between units containing different microfacies have been taken into consideration in order to establish the relationships of different carbonate platform settings. In the Cenozoic Tethyan realm, published data on ramps are scarce (Aigner 1983; Buxton and Pedley 1989; Accordi et al. 1998; Bassi 1998; Moody 1998; Nebelsick and Bassi 2000; Rasser 2000; Romero et al. 2002). Our hypothesis of a sedimentary model is governed by the following data: (1) available data from off-shore successions in the northern part of the Adriatic Sea (west of the studied area) reveal continuous basin sedimentation from Cretaceous to Oligocene (Premec Fuček et al. 1998); (2) published and obtained data on Istrian Eocene sediments show facies succession which extends from the protected zone (characterized by lagoonal facies) to the subtidal zone. The subtidal zone exhibits the largest environmental diversification, with a facies typical for sandy bottoms, vegetated bottoms (large distribution of epiphytic foraminifera in some levels; Drobne 1977, 1979), and muddy bottoms; (3) lack of reefal build-ups; (4) the geometry of Eocene sediments and facies distribution in Peri-Mediterranean realm (Tunisia: Moody 1998; France: Sinclair et al. 1998; Greece: Accordi et al. 1998; Italy: Bassi 1998); (5) facies associations and the paleogeographic position of this system compared to those of present-day shelf—lagoonal ramps sensu James et al. (1999); and (6) good stratigraphic correlation between Middle Adriatic off- and on-shore areas which suggest that platform facies (ranging from salt and brackish marshes of tidal flat to foreslope environments) are laterally extensive in the Dinaric direction (Tari Kovačić 1997).
Microfacies I
This microfacies (Fig. 6 1; Table 1, Fig. 5) represents the development of nummulite shoals or bars in a “proximal” middle ramp setting deposited near the fair weather wave base (FWWB). Currents were sufficient to concentrate the bioclasts but insufficient to cause significant abrasion or remove all of the mud matrix. Cross bedding indicates local high-energy conditions, probably above FWWB. The lack of abrasion of the foraminifera indicates autochthonous accumulations, winnowed in situ by oscillatory currents similar to those interpreted from Tertiary foraminiferal banks of Egypt (Aigner 1983). The interbedded wackestones represent the development of more protected intershoal areas, and the presence of alveolinid-nummulitid grainstones represents higher-energy restricted conditions. The relative abundance of juveniles indicates that foraminiferal composition reflects an assemblage preserved in life position.

Associated benthic foraminifera: 1 Sphaerogypsina carteri Silvestri, Asterigerina sp., Pićan section, sample Pć 5/2229, scale bar 500 μm. 2 Sphaeogypsina sp., Ragancini section, sample RL 5, scale bar 250 μm. 3 Sphaerogypsina globula Reuss, Pićan section, sample Pć 5/2229, scale bar 250 μm. 4, 7 Neorotalia vienotti Greig, axial sections, Šterna section, sample Štr 3/1727, scale bar 250 μm. 5 Neorotalia vienotti Greig, equatorial section, Šterna section, sample Štr 3/1727, scale bar 250 μm. 6 Neorotalia vienotti Greig, oblique section, Šterna section, sample Štr 3/1727, scale bar 250 μm. 8 Asterigerina sp., Šterna section, sample Štr 3-12/95, scale bar 250 μm. 9 Amphistegina sp., Pićan section, sample Pć 5/2229, scale bar 250 μm. 10 Gyroidinella magna Le Calvez, Pićan section, sample Pć 5/2229, scale bar 1 mm. 11 Assilina praespira (Douvillei), Sphaerogypsina sp., Amphistegina sp., Neorotalia sp., Ragancini section, sample RL 3-2/95, scale bar 1 mm
Microfacies II
This discocyclinid microfacies was deposited under low energy conditions, below FWWB and above storm wave base in the middle ramp setting, at a water depth estimated to have been between 30–60 m (middle neritic). Sediments may have been affected by bottom currents and storm processes (Fig. 5). The variation in the foraminiferal assemblage (abundance of discocyclinids and diversification of smaller rotaliids, Fig. 6 2) reflects the different water depths encountered in the ramp setting, with sediments containing Nummulites specimens reflecting shallower waters than those containing orthophragminids, mainly discocyclinids.
Microfacies III
The dominance of fine grained sediments and the lack of abraded detritus indicate a very low energy depositional environment, probably an outer ramp because of the presence of planktonic foraminifera (Fig. 5). The planktonic foraminiferal assemblage contains surface to deep-dwelling genera and species. This microfacies represents a paleoenvironment where orthophragminids lived “at their limits”. The predominance of adult orthophragminids may indicate that conditions were tolerable for their growth, but unfavourable for reproduction. Along with orthophragminids, “operculinids” and radiate, biconical nummulitids were found (Fig. 6 3). The presence of glauconite indicates deeper water and less oxygenated or reducing conditions with a low sedimentation rate (Odin and Matter 1981).
Microfacies IV
The large amount of fine grained sediments and the mode of preservation of the larger foraminifera indicate a low energy outer ramp setting (Fig. 5) with events of heavy erosion and floating transport from coastal areas. The glauconite grains occurring as infillings of chambers in larger and planktonic foraminifera support this interpretation (Odin and Matter 1981). Fracturing of delicate Discocyclina tests, including regenerated tests (Fig. 6 4), indicates that at least some of the specimens survived predator activities. The rare presence of quartz grains indicates sporadic terrigenous influx into the environment.
Facies model for transition from the orthophragminid limestones to transitional beds
A depositional model showing the proposed morphology and facies architecture of the “Istrian Platform” is provided in Fig. 5. This model is largely comparable with those of other Eocene carbonate platforms (Bassi 1998; Moody 1998; Accordi et al. 1998; Sinclair et al. 1998), with depositional facies associations characteristic of a ramp setting, with an inner, middle and outer ramp. In the inner ramp (not subject of this paper) the most abundant lithofacies are medium to coarse-grained larger foraminifera—mollusc packstones (Drobne 1977, 1979, 2000; Drobne et al. 1979; Drobne and Pavlovec 1991; Drobne et al. 1991; Drobne and Ćosović 1998). The presence of larger foraminifera such as Coskinolina, Alveolina , Orbitolites , Nummulites and Assilina, place the depositional environment in the upper photic zone.
Middle to outer ramp settings (Microfacies I–IV) can be divided into three parts: medium to fine-grained foraminiferal packstones and floatstones dominated by assemblages of larger foraminifera, with common genera such as “ Operculina”, Nummulites , Discocyclina , Orbitoclypeus and Asterocyclina; fine-grained orthophragminidal wackestones and planktonic foraminiferal wackestones; to mudstones which are often bioturbated. The faunal association suggests that the depositional environment was situated in the mesophotic to aphotic zone. These provide a means of establishing a detailed subdivision of the ramp environments comparable with other published works on the environmental demands of these genera (Racey 1992; Ćosović and Drobne 1998; Hohenegger and Yordanova 2001; Romero et al. 2002). Bed-forms composed almost entirely of large, thin nummulitids and “operculinids” are interpreted, both in this paper and by Racey (1992), as being developed across the mid-ramp by periodic high energy para-autochthonous reworking and concentration of in situ foraminiferal faunas. Identification of larger foraminiferal assemblages provides not only a valuable tool for the reconstruction of paleoenvironments and relative sea-level curves through shallow-marine successions, but it also provides a means of determining the provenance of reworked, allochthonous units identified within outer ramp settings.
Microfacies I-IV represent deposition on a low-angle, relatively low energy carbonate ramp. Larger foraminifera dominated the ramp benthos. Sediments dominated by Nummulites, Amphistegina , Neorotalia specimens (Microfacies I, Fig. 6 1, Fig. 7 4–7, 9), molluscs and corallinaceans characterized the “proximal” middle ramp. Deposition occurred above FWWB, with oscillatory currents winnowing the sediments and removing some of the micrite, producing cross-bedding and imbricated bioclasts. The corallinacean (Microfacies I) occurrences suggest slightly deeper water, though still too shallow and light to support a foraminiferal assemblage. The corallinaceans partly bounded the sediment, baffling and preserving some micrite. The resulting algal shoals might have locally built up sufficient relief to produce protected back shoal areas, with deposition of nummulitids, amphisteginids, assilinids, and orbitoclypeids. The nummulite shoals were deposited in slightly deeper water, though still under the influence of wave action, producing imbrication of the foraminiferal tests, though the water energy was insufficient to winnow out the mud matrix. The middle ramp sediments deposited below FWWB are dominated by foraminiferal wackestones. They show a variation of the foraminiferal assemblage, which corresponds to increasing water depths. The middle ramp contains an assemblage dominated by robust orthophragminids and nummulitids, transitioning into a lower middle ramp assemblage dominated by flat discocyclinids and “operculinids”. The outer ramp was characterized by very low energy deposition of mudstones and marls. The lack of bioclasts and the appearance of planktonic foraminifera in a considerable amount indicate deep water (Table 1).
Facies architecture of the studied sediments from the Late Cuisian to Late Lutetian records facies migration during the transgressive phase in response to the interaction between tectonic subsidence, eustasy and nutrient conditions. During the transgressive phase, the middle ramp facies backstepped. This resulted from carbonate production in the aphotic zone by filter feeding organisms (bryozoans). The water depth independent biota were able to continue production even in conditions of progressive deepening (Transitional Beds).
Summary
During the Middle Eocene, the Istrian peninsula was part of the Adriatic Carbonate Platform, covered by a shallow oligotrophic sea. The Late Cuisian to Late Lutetian of the Central Istrian region marked by Orthophragminae-bearing limestones, culminated in the final demise of the long-lasting Adriatic Carbonate Platform setting.
The orthophragminal limestones are part of a Transgressive System Tract, and are grouped into four microfacies types according to: composition of foraminiferal associations, planktonic foraminiferal ratios, endosymbionts, and flattening of nummulitid and orthophragminid tests. The results of the present work confirm the utility of the Orthophragminae in reconstruction of the transition from shallow-marine to basin paleoenvironments.
-
1.
Microfacies analyses suggest a carbonate ramp model for the Lutetian. Alternation in the composition of foraminiferal associations may indicate rather stable, more or less oligotrophic, euphotic conditions until the end of sedimentation of Orthophragminae-limestones. The Orthophragminae-dominated microfacies imply environments generally below storm wave base in middle to outer ramp areas, where transport and redeposition of bioclastic sediment originating from shallower areas. They mark the time-transgressive drowning of parts of the Middle Eocene homoclinal ramp.
-
2.
There is regularity in succession of Microfacies types at studied sections in spite of age differences. This documents the overriding control of the larger benthic foraminiferal associations during the demise of the shallow marine carbonate regime.
-
3.
In the ecological gradient, the diversity of orthophragminids increases with progressive depth of deposition. This contrasts with the trend occurring at the lower limit of the photic zone, where only a few specialized larger foraminifera survive.
This can be explained by an independent second ecologic factor, eliminating the lowermost part of the depth ranges in the light gradient. This second factor could be a nutrient cline rising over the light limit. The so-called demise of the Adriatic Carbonate Platform is a signal of relative sea-level rise and a result of tectonic activities in the hinterland, altering the nutrient regime and/or coastal current pattern in the Middle Eocene Sea.
-
4.
Flattening of foraminiferal tests, reduction in the total number of asterocyclinds and orbitoclypeids, and an increase in diversity of discocyclinids occur with deepening of the environment. Flattening of the test and lamellar thickness may be a useful tool to determine Orthophragminae depth zonation.
References
Anonymous authors of Basic Geological Map of former Yugoslavia 1:500000, Federal Geological Institute (1970), Beograd
Accordi G, Carbone F, Pignatti J (1998) Depositional history of a Paleogene carbonate ramp (Western Cephalonia, Ionian Islands, Greece). Geol Romana n.s. 34:131–205
Aigner T (1983) Facies and origin of nummulitic buildups: an example from the Giza Pyramids Plateau (Middle Eocene, Egypt). Neues Jahrb Paläontol Abh 166:347–36
Barron EJ, Peterson WH (1991) The Cenozoic ocean circulation based on ocean General Circulation Model results. Palaeogeogr Palaeoclimatol Palaeoecol 83:1–28
Bassi D (1998) Coralline Algal Facies and their paleoenvironmentals in the Late Eocene of Northern Italy (Calcari di Nago, Trento). Facies 39:179–202
Bignot G (1972) Recherches stratigraphiques sur les calcaires du Cretace superieur et de l’Eocene d’Istrie et des regions voisines. Essai de revision du Liburnien. These Doct Etat, Trav Lab Micropaleont, 2. Univ Paris VI, Paris, 353 pp
Butterlin J, Vrielynck B, Guiraud R, Bignot G, Colchen M, Clermonte J, Andreieff P, Bellion Y, Benkhelil J, Cavelier C, Cornee JJ, Mercier de Lepinay B, Montenat C, Moreau C, Poisson A, Vila JM (1993) Lutetian palaeoenvironments (46 to 40 Ma). In: Dercourt J, Ricou LE, Vrielynck B (eds) Atlas Tethys palaeoenvironmental maps. BEICIP-FRANLAB, Rueil-Malmaison
Buxton M, Pedley H (1989) A standardized model for Tethyan Tertiary carbonate ramps. J Geol Soc 146/5:746–748
Ćosović V (1996) Discocyclinidal Foraminifera of Adriatic Carbonate Platform. Unpubl PhD Thesis, Univ Zagreb, 149 pp
Ćosović V, Drobne K (1995) Paleoecological significance of morphology of orthophragminids from the Istrian Peninsula (Croatia and Slovenia). Geobios 18:93–99
Ćosović V, Drobne K (1998) Lutetian Orthophragminae from the Istrian Peninsula (Adriatic Sea, Croatia and Slovenia). In: Drobne K, Hottinger L (eds) Paleogene shallow benthos of the Tethys. Dela—Opera SAZU 4 razr 34/2 Ljubljana, pp 173–181
Drobne K (1977) Alveolines Paleogenes de la Slovenie et de l’Istrie. Schweiz Paläontol Abh 99:1–132
Drobne K (1979) Paleogene and Eocene beds in Slovenia and Istria. 16th Europ Micropaleontol Colloq, pp 49–63
Drobne K (2000) The north-western part of the Adriatic Carbonate Platform from the K/T boundary to the Flysch Deposition during Paleocene—Eocene. Riassunti delle comunicazioni orali e dei poster. 80a Riunione Estiva, Trieste, 6–8 settembre 2000, pp 231–232
Drobne K, Ćosović V (1998) Repetitive order of appearance of larger Foraminifera and the ecological gradients preserved in Paleogene shallow benthic carbonate deposits. Tertiary to Recent larger Foraminifera. Their depositional environments and importance as petroleum reservoirs, conference and workshop: February 15–19th, 1998, Kingston Univ, 10 pp
Drobne K, Pavlovec R (1991) Paleocene and Eocene beds in Slovenia and Istria. Introduction to the Paleogene SW Slovenia and Istria. Field and guidebook IGCP Project 286 “Early Paleogene Benthos”, Second Meeting, pp 7–17
Drobne K, Pavlovec R, Ćosović V (1991) Section Ragancini-Lišani, Lower and Middle Lutetian. Introduction to the Paleogene SW Slovenia and Istria. Field and guidebook IGCP Project 286 “Early Paleogene Benthos”, Second Meeting, pp 83–85
Drobne K, Pavlovec R, Šikić L, Benić J (1979) Excursion F, Pićan, Istria—Cuisian, Lutetian. 16th Europ Micropaleontol Colloq, pp 177–184
Drooger WJ (1993) Radial Foraminifera, Morphometrics and Evolution. Verhandel Konink nederl Akad Wetten, Afd Natuurk E Reeks, Amsterdam, 241 pp
Ferrandez-Canadell C, Serra Kiel J (1992) Morphostructure and paleobiology of Discocyclina GÜMBEL, 1870. J Foramin Res 22/2:147–165
Flügel E (1982) Microfacies analysis of limestones, Springer, Berlin Heidelberg New York, 633 pp
Hallock P (1979) Trends in test shape with depth in large, symbiont-bearing Foraminifera. J Foramin Res 9:61–69
Hallock P (1985) Why are larger Foraminifera large? Paleobiology 11:195–208
Hallock P (1999) Symbiont-bearing Foraminifera. In: Sen Gupta BK (ed) Modern Foraminifera. Kluwer Academic Publishers, Dordrecht, pp 123–139
Hallock P, Glenn CE (1986) Larger Foraminifera: A tool for paleoenvironmental analysis of Cenozoic carbonate depositional facies. Palaios 1:55–64
Hamrla M (1959) O pogojih nastanka premogišč na Krasu. Geologija 5:180–264
Hamrla M (1987) Light reflectance of some Slovenian coals. Geologija 28/29:293–317
Hemleben C, Spindler M, Anderson OR (1989) Modern planktonic Foraminifera. Springer, Berlin Heidelberg New York, 363 pp
Hohenegger J (2000) Coenoclines of larger Foraminifera. Micropaleontol 46 (suppl 1):127–151
Hohenegger J, Yordanova E, Hatta A (2000) Remarks on West Pacific Nummulitidae (Foraminifera). J Foramin Res 30:3–28
Hohenegger J, Yordanova E (2001) Displacement of larger Foraminifera at the western slope of Motobu Peninsula (Okinawa, Japan). Palaios 16/1:53–72
Hollaus S, Hottinger L (1998) Temperature dependence of endosymbiontic relationships: evidence from the depth range of Mediterranean Amphistegina lessoni (Foraminiferida) truncated by the thermocline. Eclogae Geol Helv 90:591–597
Hottinger L (1977) Foraminifères operculinoformis. Mém Mus Nat Hist N Sér C Sc Terre 40. Muséum national d’Histoire naturelle Paris, 159 pp
Hottinger L (1983) Processes determining the distribution of larger Foraminifera in space and time. Utrecht Micropaleontol Bull 30:239–253
Hottinger L (1996) Sels nutritifs et bisedimentation. Mém Soc Géol France ns 169:99–107
Hottinger L (1997) Shallow benthic foraminiferal assemblages as signals for depth of their deposition and limitations. Bull Soc Geol Fr 168:491–505
Hottinger L (2000) Functional morphology of benthic foraminiferal shells, envelopes of cells beyond measure. Micropaleontol 46 (suppl 1):57–86
James NP, Collins LB, Bone Y, Hallock P (1999) Subtropical carbonates in a temperate realm: Modern sediments on the southwest Australia Shelf. J Sediment Res 69/6:1297–1321
Juračić M (1980) Depth of sedimentation of “Marls with Crabs” estimated from the ratio between planktonic and benthic Foraminifera. Geol Vjesnik 31 (1979):61–67
Langer M, Hottinger L (2000) Biogeography of selected “larger” foraminifera. Micropaleontol 46 (suppl 1):57–86
Less G (1987) Paleontology and stratigraphy of the European Orthophragminae. Geol Hungarica (Paleontologica) 51:373 pp
Luterbacher HP (1970) Environmental distribution of Early Tertiary microfossils, Tremp Basin, Northeastern Spain. ESSO Production Research—European Laboratories, 46 pp
Luterbacher HP (1984) Paleoecology of foraminifera in the Paleogene of the Southern Pyrenees. Benthos’83, 2nd Int Symp Benthos Foraminifera, pp 389–392
Marinčić S, Šparica M, Tunis G, Uchman A (1996) The Eocene flysch deposits of the Istrian Peninsula in Croatia and Slovenia: regional, stratigraphic, sedimentological and ichnological analyses. Annales 9/96:139–156
Marjanac T, Babac D, Benić J, Ćosović V, Drobne K, Marjanac Lj, Pavlovec R, Velimirović Z (1998) Eocene carbonate sediments and sea-level changes on the NE part of Adriatic Carbonate Platform (Island of Hvar and Pelješac Peninsula, Croatia). In: Hottinger L, Drobne K (eds) Paleogene shallow benthos of the Tethys. Dela—Opera SAZU 4 razr 34/2, pp 243–254
Matičec D, Vlahović I, Velić I, Tišljar J (1996) Eocene limestones overlying Lower Cretaceous deposits of western Istria (Croatia): Did some parts of present Istria form land during the Cretaceous? Geol Croatica 49/1:117–127
Moody R (1998) Part 3: Background Information on Metlaoui Carbonates. In: Moody R, Brown A, Loucks R (eds) Tertiary to Recent larger Foraminifera: their depositional environments and importance as petroleum reservoirs. Field Trip Guide. Arco—BG plc—Carthago Oil—Chevron Kingston, pp 2–23
Murray JW (1991) Ecology and distribution of planktonic foraminifera. In: Lee JJ, Anderson RO (eds) Biology of Foraminifera. Academic Press, London, pp 255–285
Odin GS, Matter A (1981) De glaucodiarum origine. Sedimentology 28:611–641
Nebelsick JH, Bassi D (2000) Diversity, growth forms and taphonomy: key factors controlling the fabric of coralline algae dominated shelf carbonates. In: Insalaco E, Skelton PW, Palmer TJ (eds) Carbonate platform system: components and interactions. Geol Soc Lond 178:89–107
Pavlovec R, Drobne K, Ćosović V, Šikić L, Benić J (1991) The Pićan Profile, Middle and Upper Cuisian, Lower and Middle Lutetian. Introduction to the Paleogene SW Slovenia and Istria. Field and guidebook IGCP Project 286 “Early Paleogene Benthos”, Second Meeting, pp 73–82
Pecheux MJF (1995) Ecomorphology of recent larger foraminifer Operculina ammonoides. Geobios 28/5:529–566
Piller WE (1994) The Northern Bay of Safaga (Red Sea, Egypt): an actuopaleontological approach IV. Thin section analysis. Beitr Palaontol Ősterr 18:1–73
Piller WE, Pervesler P (1989) The Northern Bay of Safaga (Red Sea, Egypt): an actuopaleontological approach I. Topography and bottom facies. Beitr Palaontol Ősterr 15:103–147
Premec Fuček V, Babić Lj, Bajraktarević Z, Gušić I (1998) Planktonic foraminiferal biostratigraphy and paleoecology of the Middle to Upper Eocene succession in the north Adriatic Sea. In: Hottinger L, Drobne K (eds) Paleogene shallow benthos of the Tethys. Dela—Opera SAZU 4 razr 34/2, pp 255–271
Racey A (1992) The relative taxonomic value of morphological characters of the genus Nummulites (Foraminiferida). Micropaleontol 11(2):197–209
Rasser MW (2000) Coralline red algal limestones of the Late Eocene Alpine Foreland Basin in Upper Austria: component analysis, facies and paleoecology. Facies 42:59–92
Reiss Z, Hottinger L (1984) The Gulf of Aqaba. Ecological micropaleontology, Ecological Studies 50, Springer, Berlin Heidelberg New York, 354 pp
Renema W, Troelstra SR (2001) Larger Foraminifera distribution on a mesotrophic carbonate shelf in SW Sulawesi (Indonesia). Palaeogeogr Palaeoclimatol Palaeoecol 175:125–146
Romero J, Caus E, Rosell J (2002) A model for the palaeoenvironmental distribution of larger foraminifera based on late Middle Eocene deposits on the margin of the South Pyrenean basin (NE Spain). Palaeogeogr Palaeoclimatol Palaeoecol 179:43–56
Salopek M (1954) Beiträge zur Kenntnis des geologischen Baues der Becken von Labin und Pićan in Istrien. Prir istraž Jugosl akad znan umjet 26:5–58
Serra-Kiel J, Hottinger L, Caus E, Drobne K, Ferrandez C, Jauhri AK, Less G, Pavlovec R, Pignatti J, Samso JM, Schaub H, Sirel E, Strougo A, Tambareau Y, Tosquella J, Zakrevskaya E (1998) Larger foraminiferal biostratigraphy of the Tethyan Paleocene and Eocene. Bull Soc Géol Fr 169/2:281–299
Sinclair HD, Sayer ZR, Tucker ME (1998) Carbonate sedimentation during early foreland basin subsidence: the Eocene succession of the French Alps. In: Wright VP, Burchette TP (eds) Carbonate Ramps. Geol Soc Lond pp 205–227
Stache G (1889) Die Liburnische Stufe und deren Grenz-Horizonte. Abh kk Geol Reichsanst 13, 170 pp
Spezzaferri S (1995) Planktonic foraminiferal paleoclimatic implications across the Oligocene-Miocene transition in the oceanic record (Atlantic, Indian and South Pacific). Paleogeogr Palaeoclimatol Palaeoecol 114/1:43–74
Šikić D (1965) Geology of the paleogene deposits in Istria and Dalmatia. Unpubl PhD Thesis, Univ Zagreb, 149 pp
Tari Kovačić V (1997) The development of the Eocene platform carbonates from wells in the Middle Adriatic Off-Shore Area, Croatia. Geol Croatica 50/1:33–48
Tucker ME (1993) Carbonate diagenesis and sequence stratigraphy. In: Wright VP (ed) Sedimentology review /1, Blackwell, Oxford, pp 51–72
Yordanova EK, Hohenegger J (2002) Taphonomy of larger Foraminifera: relationship between living individuals and empty tests on flat reef slopes (Sesoko Island, Japan). Facies 46:169–204
Acknowledgement
This study was financed by Project 0119402 of Croatian Ministry of Science and Technology. This paper benefited from suggestions given by Lukas Hottinger, Vlasta Tari, Anna Fugagnoli and Davide Bassi and their insights are greatly appreciated. We wish to thank Josip Benić and Vlasta Premec Fuček for identification of nannofossils and planktonic foraminifera. We are very grateful to J. Kuss and M. Rasser for their critical reading of this paper. It was substantially improved by their suggestions. Thanks also go to the Editor-in-Chief, A. Freiwald, for valuable advices. We are also indebted to R. Košćal for making computer drawings. Carrie E. Schweitzer is gratefully acknowledged for English correction of the final text.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Ćosović, V., Drobne, K. & Moro, A. Paleoenvironmental model for Eocene foraminiferal limestones of the Adriatic carbonate platform (Istrian Peninsula). Facies 50, 61–75 (2004). https://doi.org/10.1007/s10347-004-0006-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10347-004-0006-9