
Abstract
The aim of this work was to characterize the structural and functional properties of hemp seed protein-derived acetylcholinesterase (AChE)-inhibitory enzymatic hydrolysates. Hemp seed protein isolate hydrolysis was performed using six different proteases (pepsin, papain, thermoase, flavourzyme, alcalase and pepsin + pancreatin) at different concentrations (1–4 %). The degree of hydrolysis was directly related to the amount of protease used but had no relationship with AChE-inhibitory activity. Amino acid composition results showed that the hemp seed protein hydrolysates (HPHs) had high levels of negatively charged amino acids (39.62–40.18 %) as well as arginine. The 1 % pepsin HPH was the most active AChE inhibitor with ~6 µg/mL IC50 value when compared to 8–11.6 µg/mL for the other HPHs. Mass spectrometry analysis showed that most of the peptides in all the hydrolysates were less than 1000 Da in size. However, the pepsin HPHs contained larger-sized peptides (244–1009 Da) than the papain HPHs (246–758 Da), which in turn was larger than the alcalase HPH (246–607 Da). The higher AChE-inhibitory effects of the pepsin HPHs may be due to increased synergistic effects from a wider peptide size range when compared to the papain and alcalase HPHs that had narrower ranges. The narrow peptide size range in the alcalase HPH confirms the higher efficiency of this protease in releasing small-sized peptides from food proteins.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The serine hydrolase, acetylcholinesterase (AChE), has long been known to play a very crucial physiological role in the central nervous system (CNS) through rapid acetylcholine (ACh) clearance from the synapse [1]. During neurotransmission, ACh is synthesized from choline and acetyl-CoA by the enzyme choline acetyltransferase. ACh is then released from the nerve into the synaptic cleft where it binds to receptors on the post-synaptic membrane thereby relaying the signal from the nerve to the brain [2]. AChE converts free ACh into the inactive metabolites, choline and acetate, thus promoting normal brain and muscle functions by preventing over-stimulation of the nervous system. The liberated choline is taken up again by the pre-synaptic nerve and combined with acetyl-CoA to synthetize ACh through the action of choline acetyltransferase [2]. Thus, normal functioning of AChE also ensures proper recycling of choline and acetyl-CoA such that an optimal level of ACh is maintained within the nervous system. As aging and dietary changes set in, less ACh is synthesized due to non-regeneration of used neurons and shortage of choline. Meanwhile, AChE continues to hydrolyze the small amounts of ACh; the net result is low ACh levels in the CNS. This leads to reduced functioning of nerve impulses and inadequate signal transmission to the brain, thus eventually contributing to memory impairment such as in Alzheimer’s disease (AD) and dementia [3].
AChE inhibition is an important research topic in the wide range of associated health implications in humans because of the ability to hydrolyze ACh at cholinergic synapses with higher catalytic efficiency than other known enzymes [4, 5]. AChE inhibitors are, therefore, employed to reduce the rate at which ACh is broken down, thereby increasing brain ACh concentration and combating the loss caused by the death of cholinergic neurons [6]. The approach involves the use of compounds that possess structural similarity to ACh such that they can fit into the enzyme active site, thereby reducing catalysis rate [2, 4]. As of today, the current available therapy for AD consists of the administration of synthetic AChE inhibitors due to their clinical efficacy in prolonging the half-life of ACh [1]. Such synthetic drugs (like tacrine, rivastigmine, galantamine, memantine and donepezil) are characterized by common negative side effects like nausea, vomiting, muscle cramps, decreased heart rate (bradycardia), decreased appetite and weight, as well as increased gastric acid production [4, 7, 8].
However, many approaches have been employed recently to address this critical problem of negative side effects, some of which include the use of natural polyphenolic plant products and their secondary metabolites [1, 7–9]. Zare-Zardini et al. [5] recently isolated a 31-mer peptide (Snakin-Z) from Ziziphus jujuba fruits, which inhibited in vitro AChE activity. However, to the best of our knowledge, there is scanty information on the potential use of naturally occurring bioactive peptides (hydrolysates) from plant proteins such as AChE inhibitors. Food protein hydrolysates contain peptides that consist of peptide bonds, which have structural similarity to the ester bond in ACh. Moreover, several peptides contain positively charged nitrogen atoms just like those found in ACh. Therefore, theoretically, small-sized peptides could simulate the structural conformation of ACh and enable peptide interaction with the AChE active site. Since peptides are not AChE substrates, such binding to the active site could exclude ACh, slow down enzyme catalysis and potentially enhance physiological levels of ACh during aging.
Food protein-derived hydrolysates have reduced the risk of imparting negative side effects in consumers and, therefore, could serve as suitable alternatives or complementary treatment to AD drugs. Previous works have demonstrated several in vitro [10, 11] and in vivo [12–14] anti-oxidant and antihypertensive effects of hemp seed protein hydrolysates (HPH). A previous work also showed the oxidative apoptosis protective effect of hemp seed peptides [15,] but so far there has been no report of AChE inhibition by food protein hydrolysates. Therefore, the aim of this study was to determine, for the first time, the in vitro AChE-inhibitory properties of several HPH generated from the action of different enzymes during proteolysis of hemp seed plant proteins. The amino acid composition, peptide size distribution and mass of potential peptides of the most active HPHs were determined.
Materials and Methods
Hemp Seed Products and Chemical Reagents
Defatted hemp seed protein meal (HPM) with 37 % protein content was a gift from Hemp Oil Canada (St. Agathe, MB, Canada). All the enzymes, pepsin (porcine gastric mucosa; E.C. 3.4.23.1; ≥250 U/mg), papain (papaya latex; E.C. 3.4.22.2), alcalase (Bacillus licheniformis; E.C. 3.4.21.62), flavourzyme (Aspergillus oryzae; ≥500 U/g), pancreatin (porcine pancreas; 647-014-00-9) and Electrophorus electricus (electric eel; 1256 U/mg protein) acetylcholinesterase Type VI-S, were purchased from Sigma-Aldrich (St. Louis, MO, USA). Thermoase (Aspergillus sp.; Protease A “Amano” 2 SD) was a gift from Amano Enzyme (Nishiki, Naka-ku, Nagoya, Japan). Dithio-bis-nitrobenzoic acid (DTNB) and acetylthiocholine were also purchased from Sigma-Aldrich. Other analytical-grade reagents were obtained from Fisher Scientific (Oakville, ON, Canada).
Preparation of Hemp Seed Protein Isolate (HPI)
HPI was produced from HPM according to the method of Girgih et al. [10] with slight modifications. HPM was dispersed in deionized water (1:20, w/v) and the dispersion was adjusted to pH 8.0 using 2 M NaOH to solubilize the proteins while stirring at 37 °C for 2 h; this was followed by centrifugation (7000g, 1 h, 4 °C). The precipitate was discarded and the supernatant filtered with cheese-cloth, adjusted to pH 5.0 with 2 M HCl to precipitate the proteins and then centrifuged (7000g, 1 h, 4 °C). The resultant precipitate was re-dispersed in deionized water, adjusted to pH 7.0 with 2 M NaOH and freeze-dried to obtain the HPI. Protein concentration of the HPI was determined using the modified Lowry method [16].
Preparation of Enzymatic Hemp Seed Protein Hydrolysates (HPHs)
Hydrolysis of the HPI was conducted using each of the following enzymes and reaction conditions: alcalase (50 °C, pH 8.0, 4 h); papain (65 °C, pH 6.0, 4 h), pepsin (37 °C, pH 2.0, 2 h), thermoase (50 °C, pH 8.0, 4 h), flavourzyme (50 °C, pH 8.0, 4 h) and pepsin + pancreatin (first with pepsin, 37 °C, pH 2.0, 2 h; then pancreatin at 37 °C, pH 7.5, 4 h) as previously described [17]. HPI (5 %, w/v, protein basis) was suspended in deionized water in a reaction vessel equipped with a stirrer, heated to the appropriate temperature and adjusted to the appropriate pH value prior to addition of the proteolytic enzymes. Each protease was added to the HPI slurry at five different enzyme-to-substrate protein ratios: 0.5:100, 1:100, 2:100, 3:100, and 4:100. During digestion, the reaction mixture pH was maintained constant by addition of 1 M NaOH. After digestion, the enzymes were inactivated by immersing reaction vessel in hot water bath at 95 °C for 10 min and allowed to cool. The undigested proteins were precipitated by centrifugation (8000g, 60 min at 4 °C) while the supernatant containing target peptides was freeze-dried as the HPH and stored at −20 °C until needed for further analysis. The protein contents of the freeze-dried HPHs were determined using the modified Lowry method [16].
Amino Acid Composition Analysis
The amino acid profiles of HPI and HPHs were determined using the HPLC S4300 Amino Acid Analyzer (Sykam, Eresing, Bavaria, Germany) according to the method previously described [18] after samples were digested with 6 M HCl for 24 h. The cysteine and methionine contents were determined after performic acid oxidation [19] and the tryptophan content was determined after alkaline hydrolysis [20].
Determination of Degree of Hydrolysis (DH)
The percent DH of each HPH was determined according to the trinitrobenzene sulfonic acid (TNBS) method as previously described [21]. HPI was digested under vacuum with 6 M HCl for 24 h and the digest was used to determine total amino groups as l-leucine equivalent. The DH was calculated as the percentage ratio of the leucine equivalent of HPHs to that of HPI.
Analysis of Molecular Weight Distribution
Molecular weight (MW) distribution of HPHs was determined as previously described [17] using an AKTA FPLC system (GE Healthcare, Montreal, PQ, Canada) equipped with a Superdex Peptide 10/300 GL column (10 × 300 mm) and UV detector (λ = 214 nm). The column was calibrated with cytochrome C (12,384 Da), aprotinin (6512 Da), vitamin B12 (1855 Da), and glycine (75 Da) as the standard proteins/amino acid. Peptide sizes of the samples were estimated from a plot of log MW versus elution volume of the standard protein/amino acid.
Mass Spectrometry Analysis of Protein Hydrolysates
The mass spectrometry analysis of the protein hydrolysates was performed according to the previously described method [17] with slight modifications. Briefly, a 10-ng/µL aliquout of the HPH was dissolved in deionized water (containing 0.1 % formic acid as solvent A) and infused directly into the mass spectrometer (MS). The elution and MS scan was monitored with the Absciex QTRAP® 6500 MS System (Absciex, Foster City, CA, USA) coupled with electrospray ionization source using the following parameters: IonSpray voltage, 3.5 kV; temperature, 150 °C; ion source gas, 30; declustering potential, 100; entrance potential, 10; and flow rate, 30 µL/min for 3 min in the positive ion mode.
Acetylcholinesterase Inhibition Assays
AChE inhibition assay was performed as previously described [22] with slight modifications. Briefly, the reaction was carried out at room temperature using 0.1 M sodium phosphate buffer (pH 7.5) and the final assay mixture was 200 μL in a 96-well microplate (Costar, Corning, NY, USA). Aliquots of 20 µL HPHs were added to the microplate wells that contained 130 µL of the buffer. DTNB (20 µL) and acetylthiocholine (10 µL) were added to each well. Then, 20 µL of AChE (0.5 U/mL final assay concentration) was added to initiate the reaction and the mixture incubated for 15 min at room temperature. The absorbance and the reaction kinetics were read for 2 min at wavelength of 412 nm using a Synergy H4 microplate reader (Biotek Instruments, Winooski, VT, USA). In the uninhibited wells, 20 µL of the buffer was used to replace HPH. In the background wells, 20 µL of the buffer was used to replace AChE while 40 µL of buffer was used to replace both AChE and HPH in the blank wells. The concentration of sample that inhibited 50 % AChE activity (IC50) was calculated from a non-linear regression plot of percentage AChE activity versus HPH concentration.
Statistical Analysis
Triplicate determinations were used to obtain mean values and standard deviations. Statistical analysis was performed with SAS (Statistical Analysis Software 9.1) using one-way ANOVA. Duncan’s multiple-range test was carried out to compare the mean values for samples and significant differences taken at P < 0.05.
Results and Discussion
Degree of Hydrolysis and AChE Inhibition
Table 1 shows that the degree of hydrolysis (DH) was significantly (P < 0.05) affected by the type and level of proteases, which is consistent with the fact that these proteases have different specificities and will produce peptides at different rates. The flavourzyme-produced HPHs have the highest (P < 0.05) DH values, which can be attributed to the dual endoprotease and exopeptidase activities possessed by the enzyme. In contrast, thermoase and pepsin hydrolysates had the least DH values, which suggest that several peptide bonds within the hemp seed proteins were not accessible or were resistant to the catalytic activities of these two enzymes. As expected, higher DH values were obtained as the amount of protease used was increased, which indicates increased proteolysis. Protein content of the HPHs ranged from ~78.13–91.64 % (Table 1); therefore, AChE assays were performed on protein weight basis.
The data on inhibitory properties of the HPHs show that there was no direct relationship between the level of enzyme used and the ability of the protein hydrolysate to reduce AChE activity (Table 1). Thus, the results suggest that there is no direct relationship between peptide size (inversely related to DH) and AChE-inhibitory activity of the HPHs. It is more likely that AChE-inhibitory activity of the HPHs was determined mostly by the type and sequence of amino acids on the peptide chain. The results show that several of the HPHs have higher AChE-inhibitory activity than the 31 amino acid residue peptide (Snakin-Z) isolated from Ziziphus jujuba fruits [5]. At 10 µg/mL, the AChE activity inhibition by Snakin-Z was <20 % when compared to several of the HPHs with >20 % inhibition at same peptide concentration.
The higher AChE-inhibitory effects of some of the protein hydrolysates may be due to synergistic effects of several peptides when compared to the single Snakin-Z peptide. The highest (P < 0.05) AChE inhibition of 54 % was by the 1 % pepsin hydrolysate, which suggests the presence of peptides that bind strongly to AChE protein. The AChE-inhibitory activities of the HPHs which was up to 54 % for the 1 % pepsin hydrolysate is similar to the maximum value of 57 % that was reported by Hasnat et al. [6] for a Ganoderma lucidum aqueous polyphenolic extract at 2.00 mg/mL. Danis et al. [23] also reported about 56 % AChE inhibition by a methanolic extract of Rhus coriaria leaves but at 1.2 mg/mL. While it may not be totally correct to compare polyphenolic activity with that of a protein hydrolysate, it should be noted that the HPHs were tested at 120–200 times less than the natural plant extract concentrations. Therefore, on a weight basis, most of the HPHs have higher AChE-inhibitory activity than the G. lucidum and R. coriaria polyphenolic extracts.
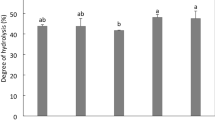
Based on the observed percentage AChE-inhibitory activities, the four HPHs with highest values were used for IC50 determination, which enables better comparison with literature values. The values obtained in this study (Fig. 1) for 1 % pepsin (5.95 ± 0.10 µg/mL), 3 % pepsin (8.04 ± 0.33 µg/mL), 3 % papain (8.97 ± 0.41 µg/mL) and 4 % alcalase (11.62 ± 0.32 µg/mL) are lower than those reported for different AChE-inhibitory phytochemical agents. For example, the following are typical IC50 values found in the literature: Pancratium illyricum L. alkaloid, 3.5 mg/mL [8]; GLBR, 1.01 mg/mL [6]; Snakin-Z peptide, 0.58 mg/mL [5]; Catharanthus roseus root alkaloids, 0.78 mg/mL [24]; R. coriaria leaves, 1.17 mg/mL [23]; and Amaryllidaceae alkaloids, 0.72 mg/mL [25]. The low IC50 results suggest that the HPHs are potentially excellent sources of highly potent peptides that can be used to inhibit AChE activity.

Inhibitory concentrations of the most active hemp seed protein hydrolysates that reduced acetylcholinesterase activity by 50 % (IC50). Bars with different letters have mean values that are significantly different (P < 0.05)
Analysis of Molecular Weight Distribution
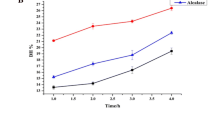
The MW of hydrolysate peptides is an important parameter in considering potential bioactivities. This is because of the general recognition that low molecular weight peptides have better chances of escaping structural degradation within the gastrointestinal tract and getting absorbed into blood circulation than larger peptides. Small-sized peptides are also more likely to fit into an enzyme active site and produce inhibitory effects than larger peptides. Therefore, one of the aims of protein hydrolysate production is to ensure the abundance of small-sized peptides in order to enhance potency against metabolic targets. The size exclusion chromatograms of the four most active in vitro AChE-inhibitory HPHs showed that the MW ranged from 300 to 9500 Da (Fig. 2). The 1 % pepsin HPH had four major peaks (A–D), but the intensity decreased when the concentration was increased to 3 % level, which is consistent with the increased DH (more extensive proteolysis) at the higher enzyme level.

Size-exclusion chromatograms of the most active acetylcholinesterase-inhibitory hemp seed protein hydrolysates
The 4 % alcalase hydrolysate showed lower levels of the 300–9500 Da peaks, which is consistent with having the highest DH among the four protein hydrolysates. Thus, alcalase seems to be more efficient than papain and pepsin in hydrolyzing the hemp seed proteins. However, protein hydrolysate activity is also highly dependent on the type of peptides produced and not just the hydrolysis efficiency. More so, the peaks produced by the proteases were different in nature and this could have been the basis for their in vitro AChE inhibitory activities. This is supported by the past study [17], which reported that the structure (size) and activity of peptides is largely dependent on its method of production (enzymes used). The results (<300 Da) obtained in this study for the alcalase HPH (even though at 4 % level) is in the range of those reported for canola protein hydrolysates (6.47–0.027 kDa) [26] and 2.6–0.14 kDa reported for rapeseed protein hydrolysates [17]. Alcalase produced a higher peak of smaller MW HPHs probably due to its broad specificity during protein digestion. Rapeseed [17] and canola [26] proteins have also been shown to be more susceptible to alcalase-induced proteolysis when compared to other proteases.
Amino Acid Composition and Peptide Mapping
Table 2 shows that the HPHs are composed of mostly negatively charged amino acids (NCAA), which was followed by hydrophobic amino acids (HAA), while the content of aromatic amino acids (AAA) was the least. The peripheral anionic site (PAS) is a unique surface area on the AChE protein that is believed to be an important binding site for inhibitors [4]. Within this binding site, positively-charged amino acids of inhibitory peptides can form a stable complex with the Trp286 indole ring of the AChE protein, which then prevents entry of substrates to the enzyme active site. Therefore, it is possible that the high arginine contents of the HPHs may have facilitated peptide–AChE interactions, which resulted in the observed inhibitions of enzyme activity.
The most active AChE-inhibitory hydrolysates viz: 1 and 3 % pepsin, 3 % papain and 4 % alcalase HPHs were subjected to mass spectrometry analysis to obtain several molecular species as shown in Fig. 3a–d. The mass spectra for 1 and 3 % pepsin HPHs indicate the presence of some ion masses (288, 318, 323, 491, 510, 518, 673, 690, 830, 848, 975 and 1009 Da) that are common to both samples. The results are consistent with the fact that proteolysis agent was the same for both 1 and 3 % pepsin HPHs. However, the intensities of some of the ions (323, 373, 491, 510, 518, and 673 Da) were higher in the 3 % pepsin HPH, which is consistent with a higher proteolysis level (DH) when compared to the 1 % pepsin HPH. Likewise, the MS scans obtained for 3 % papain and 4 % alcalase HPHs shared three similar ions (246, 288 and 359 Da), which indicate some similarity in catalytic activities of both enzymes. However, the 246, 288 and 359 Da ion intensities were stronger in the alcalase HPH, which reflects a higher catalytic rate than papain. Meanwhile, some of the species with higher molecular masses (for example, those with masses ≥800 Da) present in the 1 and 3 % pepsin HPHs spectra are seemingly diminished or absent from those of 3 % papain HPH and 4 % alcalase HPH. Overall, the alcalase HPH had the narrowest peptide size range, which may be attributed to higher protease efficiency when compared to papain. The wider peptide size range of the pepsin HPHs may be due to the higher selectivity of the protease action in combination with the shorter time period (2 h) used for catalysis when compared to 4 h for the papain and alcalase reactions.


Electrospray ionization mass spectra of the most active acetylcholinesterase-inhibitory hemp seed protein hydrolysates: a 1 % pepsin; b 3 % pepsin; c 3 % papain; d 4 % alcalase
Conclusion
This study, to our knowledge, is the first to report the in vitro AChE-inhibitory activities of a food protein hydrolysate. The lack of a relationship between DH and AChE-inhibitory activities of the HPHs suggest that the peptide chain amino acid sequence was more important than the peptide chain length as an important structural feature for potency. The results showed the 1 % pepsin HPH as the most active, although it contained peptides that are also present in the 3 % pepsin HPH. Thus, the higher potency of 1 % pepsin HPH against AChE suggests that some of the initially formed active peptides were hydrolyzed when the enzyme concentration increased. Overall, the pepsin HPHs contained a wider range of peptide sizes, which may have produced higher levels of synergy and hence better AChE-inhibitory activities when compared to the narrower peptide size ranges in papain HPH and alcalase HPH. The IC50 values are lower than values reported for polyphenols and suggest high AChE-inhibitory potency of the peptides within the context of currently available inhibitory natural products. The dominance of low molecular peptides (<1 kDa) suggests that the HPHs contain potentially bioavailable components. Therefore, the HPHs may constitute important sources of natural AChE-inhibitory agents which may provide potential health benefits during metabolic disorders that involve the nervous system. Meanwhile, further work will be performed to determine actual amino acid sequences of the peptides present in the most active HPH.
References
Kumar S, Chowdhury S (2014) Kinetics of acetylcholinesterase inhibition by an aqueous extract of Cuminum cyminum seeds. Int J Appl Sci Biotech 2(1):64–68
Singh M, Kaur M, Kukreja H, Chugh R, Silakari O, Singh D (2013) Acetylcholinesterase inhibitors as Alzheimer therapy: from nerve toxins to neuroprotection. Eur J Med Chem 70:165–188
Willcox DC, Scapagnini G, Willcox BJ (2014) Healthy aging diets other than the Mediterranean: a focus on the Okinawan diet. Mech Ageing Dev 136–137:148–162
Saravanaraman P, Chinnadurai RK, Boopathy R (2014) A new role for the non-pathogenic non-synonymous single-nucleotide polymorphisms of acetylcholinesterase in the treatment of Alzheimer’s disease: a computational study. J Comp Biol 21(8):632–647
Zare-Zardini H, Tolueinia B, Hashemi A, Ebrahimi L, Fesahat F (2013) Antioxidant and cholinesterase inhibitory activity of a new peptide from Ziziphus jujuba fruits. Am J Alzheimer’s Dis Dem 28:702–709
Hasnat MA, Pervin M, Lim BO (2013) Acetylcholinesterase inhibition, in vitro and in vivo antioxidant activities of Ganoderma lucidum grown on germinated brown rice. Molecules 18:6663–6678
Ghribia L, Ghouilaa H, Omrib A, Besbesb M, Janneta HB (2014) Antioxidant and anti-acetylcholinesterase activities of extracts and secondary metabolites from Acacia cyanophylla. Asian Pac J Trop Biomed 4(S1):S417–S423
Iannello C, Pigni NB, Antognoni F, Poli F, Maxia A, de Andrade JP, Bastida J (2014) A potent acetylcholinesterase inhibitor from Pancratium illyricum L. Fitoter 92:163–167
Borowiec K, Szwajgier D, Targońsk Z, Demchuk OM, Cybulska J, Czernecki T, Malik A (2014) Cholinesterase inhibitors isolated from bilberry fruit. J Funct Foods 11:313–321
Girgih AT, Udenigwe CC, Aluko RE (2011) In vitro antioxidant properties of hempseed (Cannabis sativa L.) protein hydrolysate fractions. J Am Oil Chem Soc 88:381–389
Girgih AT, Udenigwe CC, Aluko RE (2013) Reverse-phase HPLC separation of hemp seed (Cannabis sativa L.) protein hydrolysate produced peptide fractions with enhanced antioxidant capacity. Plant Foods Hum Nutr 68:39–46
Girgih AT, Udenigwe CC, Li H, Adebiyi AP, Aluko RE (2011) Kinetics of enzyme inhibition and antihypertensive effects of hemp seed (Cannabis sativa L.) protein hydrolysates. J Am Oil Chem Soc 88:1767–1774
Girgih AT, He R, Malomo SA, Aluko RE (2014) Structural and functional characterization of hemp seed (Cannabis sativa L.) protein-derived antioxidant and antihypertensive peptides. J Funct Foods 6:384–394
Girgih AT, He R, Malomo SA, Alashi AM, Aluko RE (2014) Preventive and treatment effects of hemp seed (Cannabis sativa L.) meal protein hydrolysate against blood pressure in spontaneously hypertensive rats. Eur J Nutr 53:1237–1246
Lu RR, Qian P, Sun Z, Zhou XH, Chen TP, He JF, Zhang H, Wu J (2010) Hempseed protein derived antioxidative peptides: purification, identification and protection from hydrogen peroxide-induced apoptosis in PC12 cells. Food Chem 123:1210–1218
Markwell MAK, Haas SM, Bieber LL, Tolbert NE (1978) Modification of Lowry procedure to simplify protein determination in membrane and lipoprotein samples. Anal Biochem 87:206–210
He R, Malomo SA, Alashi A, Girgih AT, Ju X, Aluko RE (2013) Purification and hypotensive activity of rapeseed protein-derived renin and angiotensin converting enzyme inhibitory peptides. J Funct Foods 5:781–789
Bidlingmeyer BA, Cohen SA, Tarvin TL (1984) Rapid analysis of amino acids using pre-column derivatization. J Chrom B 336:93–104
Gehrke CW, Wall LL, Absheer JS, Kaiser FE, Zumwalt RW (1985) Sample preparation for chromatography of amino-acids: acid hydrolysis of proteins. J Assoc Off Anal Chem 68:811–821
Landry J, Delhaye S (1992) Simplified procedure for the determination of tryptophan of foods and feedstuffs from barytic hydrolysis. J Agric Food Chem 40:776–779
Adler-Nissen J (1979) Determination of the degree of hydrolysis of food protein hydrolysates by trinitrobenzenesulfonic acid. J Agric Food Chem 27:1256–1262
Ellman GL, Courtney KD, Andres V, Featherstone RM (1961) A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem Pharm 7:88–95
Danis O, Yuce-Dursun B, Çimen T, Demir S, Salan Ü, Yalçın G, Ogan A (2014) Evaluation of antioxidant, radical-Scavenging and acetylcholinesterase inhibitory activities of various culinary herbs cultivated in Southern Turkey. J Food Biochem 38:602–611
Pereira DM, Ferreres F, Oliveira JMA, Gaspar L, Faria J, Valentao P, Sottomayor M, Andrade PB (2010) Pharmacological effects of Catharanthus roseus root alkaloids in acetylcholinesterase inhibition and cholinergic neurotransmission. Phytomedicine 17:646–652
Lopez S, Bastida J, Viladomat F, Codina C (2002) Acetylcholinesterase inhibitory activity of some Amaryllidaceae alkaloids and Narcissus extracts. Life Sci 71:2521–2529
Alashi AM, Blanchard CL, Mailer RJ, Agboola SO, Mawson AJ, He R, Malomo SA, Girgih AT, Aluko RE (2014) Blood pressure lowering effects of Australian canola protein hydrolysates in spontaneously hypertensive rats. Food Res Int 55:281–287
Acknowledgments
This work was funded through a Natural Science and Engineering Research Council of Canada (NSERC) Discovery grant and a Manitoba Agri-Food Research and Development Initiative (ARDI) grant to Dr. RE Aluko. SA Malomo is a recipient of University of Manitoba Graduate Fellowship (UMGF) and Manitoba Graduate Scholarship (MGS) Awards for doctoral studies.
Author information
Authors and Affiliations
Corresponding author
About this article

Cite this article
Malomo, S.A., Aluko, R.E. In Vitro Acetylcholinesterase-Inhibitory Properties of Enzymatic Hemp Seed Protein Hydrolysates. J Am Oil Chem Soc 93, 411–420 (2016). https://doi.org/10.1007/s11746-015-2779-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11746-015-2779-0