
Abstract
Bisphenol A (BPA), an endocrine-disrupting chemical (EDC), has strong potential for daily exposure to humans and animals due to its persistence and widespread in the environment, so its effects directly concern public health. Although invertebrates represent important components of aquatic ecosystems and are at significant risk of exposure, there is little information about the biological effects of EDCs in these organisms. Astacus leptodactylus used in this study is one of the most consumed and exported freshwater species in Europe. In this study, the 96-h effect of BPA on A. leptodactylus was examined using various biomarkers. The LC50 value of BPA was determined as 96.45 mg L−1. After 96 h of exposure to BPA, there were increases in superoxide dismutase (SOD), glutathione peroxidase (GPx), glutathione S-transferase (GST), alanine aminotransferase (ALT), aspartate aminotransferase (AST) and lactate dehydrogenase (LDH) activities and levels of malondialdehyde (MDA), and total oxidant status context (TOSC), and there were decreases in the activity of glutathione reductase (GR), carboxylesterase (CaE), acetylcholinesterase (AChE), Na+/K+ ATPase, Mg2+ ATPase, Ca2+ ATPase, and total ATPase and the total antioxidant context (TAC). From the results of this study, it can be concluded that BPA has significant toxic effects on A. leptodactylus based on the selected biochemical parameters of antioxidant, cholinergic, detoxification, and metabolic systems in crayfish even at low doses. Thus, it can be said that BPA can seriously threaten the aquatic ecosystem and public health.
Graphical abstract

Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Endocrine-disrupting chemicals cause anxiety all over the world due to their destructive effects on organisms following exposure. Bisphenol A (BPA), 2,2-bis (4-hydroxyphenyl) propane, is one of the most endocrine-disrupting chemicals that has spread to the environment as a result of industrial and manufacturing activities. BPA is widely used in the production of epoxy resins, polycarbonate plastics, detergents, and personal care product packages (Giulivo et al. 2016; Esteve et al. 2016). BPA acts as an endocrine-disrupting chemical and causes significant biochemical and physiological changes in organisms by disrupting the functions of tissues and organs, the structure of cells, and reproductive efficiency (Grasselli et al. 2010). Because BPAs have serious toxic and carcinogenic effects on organisms, their use has been banned in many countries and has been seriously controlled in others (Duan et al. 2014). Many waterways contain significant amounts of BPA, so microorganisms, algae, invertebrates, and fish are exposed to this chemical, and humans may also be exposed via the consumption of food and water contaminated with BPA (Tsai 2006; Mikolajewska et al. 2015). BPA has high solubility in water (120–300 mg L−1) (Staples et al. 1998) and its concentration in surface waters has been determined in the range of 0.0005–0.41 μg L−1 (Fromme et al. 2002). Since the median BPA concentration is very low in surface waters uncontaminated with BPA, it may not pose a risk to aquatic organisms, but in areas where BPA is discharged from various industrial processes, aquatic organisms may be significantly affected (Bhandari et al., 2015). BPA concentrations rise 10 times in areas susceptible to wastewater contamination (Hohenblum et al., 2004) and more than 200 times in areas affected by municipal landfills (Rudel et al., 1998). It is of concern that these concentrations are higher than ≤ 0.01 μg L−1 (0.04 nM), which has been found to cause adverse effects on aquatic wildlife and laboratory animals (Bhandari et al., 2015). While the mean lethal concentration (LC50) value of BPA for aquatic invertebrates is in the range of 960–2,700 μg L−1, this parameter is in the range of 6,800–17,900 μg L−1 for fish, so even the presence of BPA in water bodies can cause fatal toxic effects on aquatic organisms (Mathieu-Denoncourt et al. 2016). Although BPA has a short half-life in the aquatic environment (in the range of 2.5 to 4 days) (Dorn et al. 1987; Rogers et al. 2013), it may have negative effects on the nervous system, histological structure, biochemical variables, morphology, behavior, growth, and reproduction of fish (Kinch et al. 2015; Hayashi et al. 2015).
Studies have documented that some invertebrates are very sensitive to even low concentrations of BPA, so invertebrates are often used as bioindicators to investigate the toxic effects of endocrine-disrupting chemicals both in situ and in the laboratory (Oehlmann et al. 2006). Crayfish are one of the most suitable bioindicators for pollution-monitoring studies due to their feeding habits in water bodies, so their biological response to environmental pollution may reflect the health status of aquatic ecosystems (Hong et al. 2018). Astacus leptodactylus (Eschscholtz, 1823), also known as the Turkish crayfish, Galician crayfish, or marsh or pond crayfish, is one of the most common freshwater species with the greatest potential for use in Europe and is also consumed as food (Harlıoğlu 2004; Ahn et al. 2006). A. leptodactylus is Turkey’s only indigenous freshwater crayfish and is issued regularly each year (Barim-Oz 2018).
Antioxidant parameters are important defense mechanisms that protect organisms against the effects of reactive oxygen species created by environmental pro-oxidants (Tabrez and Ahmad 2009). Therefore, in ecotoxicological studies, antioxidant enzymes and oxidative stress biomarkers are most often used to determine the early warning of the biological endpoint of a particular pollutant (Pandey et al. 2003). Many studies have been conducted on the induction of oxidative stress by BPA in fish (Hulak et al. 2013; Chepelev et al. 2013; Ge et al. 2014; Kalb et al. 2016); hovewer, no studies have been found in the literature about the toxicity of BPA on A. leptodactylus.
This study was designed to determine the toxic effects of BPA on A. leptodactylus with some specific biochemical tests. The main aim was observing changes in enzyme activities responsible for detoxification (glutathione S-transferase (GST), carboxylesterase (CaE)), antioxidant activities (glutathione peroxidase (GPx), superoxide dismutase (SOD), glutathione reductase (GR), malondialdehyde (MDA), total antioxidant context (TAC), total oxidant status context (TOSC)), enzymes of the cholinergic system (acetylcholin-esterase (AChE)), metabolic enzymes (alanine-aminotransferase (ALT), aspartate-aminotransferase (AST), lactate-dehydrogenase (LDH)) in the hepatopancreas, and enzymes responsible for ion transfer in the cell (total ATPase, Na+/K+ ATPase, Mg2+ATPase, Ca2+ ATPase) in the gill and abdominal muscle of crayfish acutely (96 h) exposed to varying BPA concentrations.
Materials and methods
Experimental animals and test chemicals
The crayfish used in the study were obtained from the crayfish-growing ponds of Fırat University Faculty of Fisheries, Elazığ, Turkey. Adult crayfish weighing approximately 15 ± 5 g were used regardless of gender. BPA applications were made on crayfish placed in 40-L containers made of polypropylene material with dechlorinated tap water rested for a week. The water used in the experiment was treated according to ASTM guidelines (ASTM 2014). The average values for pH, conductivity, temperature, and dissolved oxygen of dechlorinated tap water were 7.62 (7.53–7.71), 23.40 S cm−1 (20.82–25.98), 22.6 °C (21.8–23.4), and 7.49 mg L−1 (7.20–7.78), respectively. During 96 h of application, crayfish were not fed and water in containers was replaced with fresh water with the same proportion of BPA every 24 h, so static regeneration was performed. The laboratory was at room temperature (25 ± 1 °C) and the lighting was provided by natural daylight (12-h light and 12-h dark). The ventilation of the containers was done with an air pump. The pH values in all containers were recorded daily. Water samples from the control and application group (temperature, percent dissolved oxygen, pH and conductivity) were measured at the beginning of the experiment (0 h) using a multiparameter device (YSI Pro Plus Multi Parameter). The average values for pH, conductivity, temperature, and dissolved oxygen were 7.55 (7.44–7.67), 23.10 S cm−1 (21.73–24.20), 24.4 °C (23.8–25.6), and 7.28 mg L−1 (7.00–7.60), respectively. Bisphenol A with purity ≥ 99% was purchased from Sigma-Aldrich Chemical (St. Louis, MO, USA). A 5,000-mg L−1 stock solution was prepared from the BPA solution prepared in dimethyl sulfoxide (DMSO). No adverse effects on biomarkers were seen in the preliminary experiment using the DMSO control (0.001% DMSO (v/v)).
Determination of 96 h LC50 value and experimental design
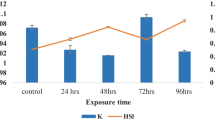
Concentrations of 0.25, 1, 4, 8, 16, 32, 64, 128, 512, and 2048 mg L−1 BPA were used from the stock solution. Individuals with immobilized bodies were considered dead, separated from those who were alive at 24, 48, 72, and 96 h, and the LC50 value was determined by Probit analysis of the Statistical Package for Social Sciences (SPSS) (Anderson 1982). Accordingly, the 96 h LC50 value of BPA was calculated as 96.45 mg L−1. Since there is no information in the literature about the maximum permissible limit of BPA for crayfish, three sub-lethal doses of this LC50 value (LC50/2, LC50/4, LC50/8) were applied to the animals.
Each container was designed to contain four animals and the study was performed in triplicate, so a total of 15 containers and 60 animals were used in five experimental groups as follows:
-
Control: No dose was administered to the animals.
-
Group LC50: 96.45 mg L−1 of BPA-treated animals.
-
Group LC50/2: 48.23 mg L−1 of BPA treated animals.
-
Group LC50/4: 24.11 mg L−1 of BPA treated animals.
-
Group LC50/8: 12.06 mg L−1 of BPA treated animals.
BPA was applied to the animals by mixing with the water in the containers. At the end of 96 h of treatment, crayfish were dissected and hepatopancreas, muscle, and gill tissues were removed.
Biochemical analyses
The dissection of crayfish was done under anesthesia on an ice bath (Leksrisawat 2010). Approximately 60 mg of samples were taken from the tissues. In a volume 4 times the total weight (w/v), homogenization was performed for 30 s at 2,000 rpm in a refrigerated homogenization buffer (0.1 M, pH 7.4 in potassium phosphate buffer (0.5 M KH2PO4, 0.5 M K2HPO4, 0.15 M KCl, 1 mM ethylenediaminetetraacetic acid [EDTA], 1 mM dithiothreitol [DTT]) using a glass-Teflon homogenizer (MTOPS MS3040D 2021) in an ice container. The homogenates were then transferred to clean microcentrifuge tubes after homogenization. The homogenates were centrifuged at 16,000 × g for 20 min at 4 °C (Hettich 460 R). The supernatant was collected after centrifugation and transferred to the clean microcentrifuge tubes, and enzyme activity in postmitochondrial fractions was examined. Total protein content and enzyme activity readings were performed in four replicates using a microplate reader (Thermo Varioscan Flash 2000). Without refreezing the assay samples, all of the assays were completed on the same day.
Total protein analysis
Total protein analysis was done according to the Bradford method (Bradford 1976). Bovine serum albumin was used as the standard. The required total protein amounts (mg mL−1) to calculate enzyme activities and MDA levels were determined from 5 µL supernatant obtained after homogenization and centrifugation of each tissue of the animals used in the experiment, as mentioned above. Five microliters of sample and 250 µL Bradford solution were pipetted into a 96-well plate, and then incubated in the dark for 15 min. Total protein amounts were calculated by taking the absorbance value read at 595-nm wavelength. The protein amounts used for the determination of enzyme activity and MDA levels are shown in Tables 3 and 4.
Determination of activity and levels of enzymatic and non-enzymatic biomarkers
Activities of AChE, GST, CaE, GR, SOD, GPx, ALT, AST, and LDH, and levels of MDA, TAC, and TOSC were performed in hepatopancreas tissue. In gill and muscle tissues, adenosine triphosphatase activities (Na+/K+ ATPase, Mg2+ ATPase, Ca2+ATPase, total ATPase) were measured.
AChE activity
AChE activity was determined as the cholinergic system enzyme. In AChE activity, the method developed by Ellman et al. (1961) in a spectrophotometer was adapted to a microplate (Ozmen et al. 2008). As the substrate, acetylcholine iodide (ACTI) was used. Ten microliters of the supernatant obtained from the sample and 200 µL of a mixture of ACTI and DTNB prepared in Trizma buffer (pH 8) was pipetted to the microplate wells. Specific enzyme activity was calculated according to the absorbance value at 412 nm.
GST activity
GST and CaE activities were measured as the enzymes responsible for the detoxification. In determining the GST activity, the method created by Habig et al. (1974) was used, adapted to a microplate reader. CDNB (1-chloro-2,4-dinitrobenzene) and reduced glutathione were used as the substrate and cofactor, respectively. A mixture of 100 µL of GSH, 100 µL of phosphate buffer, 10 µL of CDNB, and 10 µL of supernatant was pipetted into microplate wells and an absorbance reading was performed at 344 nm.
CaE activity
To measure the CaE activity, the protocol of Santhoshkumar and Shivanandappa (1999) was modified to work with a microplate reader. PNPA (p-nitrophenol acetate) was used as the substrate. Two hundred fifty microliters of Trizma buffer (pH 7.4) with 5 µL of sample were placed in microplate wells and and incubated at 25 °C for 3 min. Five microliters of PNPA was pipetted onto this mixture and the absorbance value at 405 nm was measured.
GR activity
A version of the method developed by Cribb et al. (1989), modified to work in a microplate reader, was used to determine GR activity. Twenty microliters of GSSG was added to the reaction solution containing 150 µL of DTNB (5,5′-dithiobis (2-nitrobenzoic acid)), 20 µL of NADPH (nicotinamide adenine dinucleotide phosphate), and 20 µL of sample. Specific GR activity was calculated from the absorbance values at 405 nm obtained by reducing the amount of DTNB during GSH formation from GSSG.
SOD activity
SOD activity was determined according to the method of Sun et al. (1988). In this method, nitroblue tetrazolium, reduced from the xanthine/xanthine oxidase reaction, was an indicator of the substrate superoxide. The color change created by these superoxide radicals by interacting with nitroblue tetrazolium at 540 nm was recorded as absorbance value and enzyme activity was calculated.
GPx activity
GPx activity was determined by a method developed by Bell et al. (1985) that was adapted to a microplate reader. Hydrogen peroxide (H2O2) and sodium azide (NaN3) were used as the substrate and catalase inhibitor, respectively. GPx activity was calculated by measuring the absorbance value at 340 nm.
MDA level
The hepatopancreas MDA level was measured based on the relative production of the reactive substances of thiobarbituric acid (TBA) according to the method developed by Placer et al. (1966). In this method, the color change of MDA in TBA was determined at a wavelength of 532 nm and the level of MDA was calculated as nanomoles/gram of wet tissue weight.
ALT, AST, and LDH activities
To evaluate the metabolic toxicity, the activities of ALT, AST, and LDH were determined using commercial test kits (Biolabo 80,027, Biolabo 80,025 and Biolabo 92,111, respectively).
TAC and TOSC levels/calculation of the OSI value
TAC and TOSC assays and calcultions were performed by using commercial test kits by established Erel (Reel Assay Diagnostics) (Erel 2005). The principle of the TAC analysis is based on the conversion of the dark blue-green stable 2,2′-azino-di-3-ethylbenzthiazoline sulfonate (ABTS) radical to the colorless reduced ABTS form by the antioxidants in the sample. The amount of antioxidants in the sample correlates with the loss of this color. The assay is calibrated using a stable antioxidant standard solution, which is a vitamin E analog known as Trolox. TAC levels were calculated from absorbance values as a result of end-point measurement at 660 and expressed as micromoles of Trolox equivalents L−1. The principle of the TOSC test is that the oxidants present in the sample oxidize the Fe2+ ion-chelator complex to the Fe3+ ion. In an acidic medium, Fe3+ ion forms a colored complex with chromogen and the color intensity is determined spectrophotometrically. The color density is proportional to the total amount of oxidant molecules in the sample. The calibration for the determination was carried out with hydrogen peroxide. TOSC levels were calculated from absorbance values as a result of end-point measurement at 530 nm and expressed as mmol H2O2 equivalents L−1. The following formula was used to calculate the oxidative stress index (OSI) (arbitrary unit: AU) and expressed as a percentage (Erel 2005).
ATPase activities
The activity of adenosine triphosphatase (ATPase), the enzyme responsible for ion transfer in gill and muscle tissues, was measured according to the of method Atlı and Canlı (Atlı and Canlı 2011). For the measurement of Na+/K+ ATPase and Mg2+ ATPase enzyme activities, 70 µL of incubation mixture (pH 7.7) consisting of 1 mM ouabain, 40 mM Tris–HCl, 4 mM MgCl2, 20 mM KCl, and 100 mM NaCl was added to 10 µL of supernatant. For the determination of Ca2+ ATPase activity, a mixture (pH 7.7) of 70 µL of 1 mM CaCl2, 40 mM Tris–HCl, 4 mM MgCl2, and 1 mM EGTA was added to 10 µL of the supernatant. Samples were pipetted into a 96-well plate and incubated at 37 °C for 5 min. After the first incubation, 15 µL of 3 mM Na2ATP was added to the mixture to start the reaction, which was incubated for an additional 30 min at 37 °C. Then, the reaction was stopped by adding 40 µL of ice-cold dH2O to the mixture. In addition, to serve as an ATP blank, ouabain-free medium, supernatant, and dH2O were added to a 96-well plate. To serve as a sample blank, ouabain-free medium, Na2ATP, and dH2O were added to a 96-well plate. The non-enzymatic hydrolysis of ATP was corrected using these blanks. During the incubation of the final reaction mixture, the inorganic phosphate (Pi) formed by the enzymatic/non-enzymatic hydrolysis reaction of 3 mM ATP was measured spectrophotometrically according to the protocol established by Atkinson et al. (1973). Following the completion of the reaction, 170 µL of the base mixture consisting of 80 mM polyoxyethylene 10 lauryl ether and 16 mM ammonium molybdate was pipetted into 96-well plates. Then, the final reaction mixture was incubated for 10 min at 25–28 °C. After incubation, the inorganic phosphate (Pi) value was calculated from the absorbance value at 390 nm of the yellow compound formed by the main reagent (Atkinson et al. 1973). The activity of ATPase was measured in mol Pi mg protein−1 h−1.
Na+/K+ ATPase value was found by subtracting Mg2+ ATPase activity (mixture not containing ouabain) from the total ATPase activity (mixture containing ouabain). The difference between the enzyme activities obtained depending on the presence or absence of CaCl2 and MgCl2 was used to measure the Ca2+ ATPase and Mg2+ ATPase activities, respectively.
Measurement of the actual BPA concentrations in test waters
The actual concentrations of BPA in the test waters were measured with the liquid chromatography tandem mass spectrometry (Shimadzu Prominence LC-20AD/XR LCMS-8040) at Adıyaman University Central Research Laboratory. The standard of BPA with purity ≥ 99% was obtained from Sigma-Aldrich Chemical (St. Louis, MO, USA). The calibration curve was prepared in a standard concentration range of 50–500 µg L−1. The linearity (r2) for all analytes was > 0.999. The limit of detection (LOD) was 0.06 µg L−1 and the limit of quantification (LOQ) was 0.20 µg L−1. The recovery values were between 88.4 and 115.7%. The retention time of BPA was approximately 5 min. BPA was detected through the transitions 227.20 → 132.80 mass-to-charge ratios (m/z) and 227.20 → 212.20 m/z. The instrumental conditions of LCMSMS are shown in Table 1.
Data analysis
The data were statistically analyzed using the computer software program SPSS 22. The Shapiro–Wilk test (p < 0.05) was used to assess data normality. The Kruskal–Wallis test was used to compare biochemical data from different groups. To see whether there was a substantial difference between the classes, the Mann Whitney U test was used. p < 0.001 was used as the standard for statistical significance.
The integrated biomarker response
Integrated biomarker response (IBR) was used to include all of the identified biochemical marker reactions into a single overall stress index to assess the risk potential of BPA. The IBR indexes were calculated according to the method defined by Arzate-Cárdenas and Martínez-Jerónimo (2012). The mean and standard deviation (SD) for each exposure and each biomarker were used to calculate the IBR index. Each response’s average value was normalized individually using the formula Y = (X − m)/SD, where Y is the biochemical marker’s standardized value, X is the average value of a biochemical marker for each treatment, and m is the average of the biochemical markers computed for the treatments. Depending on the biological impact, the Z values were computed as Z = Y (inhibition) or Z = − Y (activation). S =|min|+ Z, where S ≥ 0 and |min| represents the absolute value of the minimum of all application groups for each biochemical marker used to calculate the score (S). The average of the scores at each biochemical parameter level was calculated as (S1 × S2)/2 + (S2 × S3)/2 + … (Sn − 1 × Sn)/2 and used to calculate the IBR index, which was then normalized. The total number of biochemical markers was divided by the estimated values (Broeg and Lehtonen 2006; Kim et al. 2014a; Samanta et al. 2018a). A star plot was used to show the average scores. Individual biomarker scores were utilized to show the star plot in the traditional IBR technique. IBR indexes were calculated as follows:
Results and discussion
The actual concentrations of BPA in test waters
BPA concentrations in test waters measured by LCMSMS are shown in Table 2. Actual concentrations of BPA detected in water before application were approximately 15% higher than nominal concentrations. After 24 h of exposure, the actual BPA concentrations detected in water were approximately 6% lower than the nominal concentrations. BPA is a moderately hydrophobic compound that is highly soluble in water (300 g m−3) because it contains functional groups sensitive to hydrolysis (USEPA 2010). The loss of BPA at the end of the application may be due to conversion of BPA into its metabolites as a result of hydrolysis, biodegradation, and photolysis (Im and Löffler 2016) or absorption/matabolization of BPA by the crayfish (Iwano et al. 2018).
Toxicity assay
The LC50 value of BPA for A. leptodactylus was determined as 96.45 mg L−1. In other aquatic organisms, the 96 h LC50 value of BPA was reported by some researchers at the following concentrations: for Xiphophorus helleri, 0.018 mg L−1 (Kwak et al. 2001); for Oreochromis niloticus, 0.013 mg L−1 (Hamed and Abdel-Tawwab 2017); for Ctenopharyngodon idella, 6.32 mg L−1 (Faheem and Lone 2017); for zebrafish larvae, 8.04 mg L−1 (Chan and Chan 2012); and for midges, 2.7 mg L−1 (Gulley et al. 1996). According to the review by Mathieu-Denoncourt et al. (2016), the LC50 value of BPA ranges from 960 to 2,700 μg L−1 for aquatic invertebrates and 6,800 to 17,900 μg L−1 for fish. Compared to other aquatic invertebrates, A. leptodactylus showed higher tolerance to BPA, as supported by its high LC50 value. A similar situation was observed in a study for the marine rotifer Brachionus koreanus. The LC50 value determined for this species (20.924 mg/L) by Park et al. (2018) was found to be much higher than that of other aquatic invertebrates. The differences in the LC50 value reported for various aquatic organisms can be attributed to size, age, sex, species sensitivity, animal physiology, genetic background of the organisms (Tatarazako et al. 2004), the physicochemical properties of the water, dose, and the duration of exposure (Kontogiannatos et al. 2015).
Biochemical responses
Waterborne BPA can bind to estrogen receptors (ERs) in fish (Gibert et al. 2011), producing more estrogenic effects and arousing oxidative stress and affecting antioxidant defense system parameters in many aquatic organisms (Kankaya et al. 2015; Xu et al. 2015). In this study, significant changes were observed in all biochemical parameters analyzed (oxidative stress parameters, antioxidant enzymes, cholinergic system enzyme, metabolic enzymes, and ATPase activities) following short-term application of BPA to crayfish. The biochemical responses in the hepatopancreas are shown in Table 3. There have been studies on reproductive development, endocrine disruption, and immunotoxicity of BPA on aquatic organisms in the literature, but there are few reports on biochemical toxic effects in invertebrates. Therefore, we were not able to compare our findings with another study on the biochemical toxicity of BPA on A. leptodactylus. However, the biochemical toxic effects of BPA on other invertebrates depending on the exposure time and dose are discussed below according to the type of biochemical marker.
AChE is a serine protease responsible for the hydrolysis of neurotransmitter acetylcholine found in cholinergic synapses in the nervous system of animals (Orhan 2013; Hatami et al. 2019). All administered BPA concentrations significantly inhibited the AChE activity compared to the control. The lowest AChE activity was found as 1.40 ± 0.07 nmol min−1 mg protein−1 in LC50 dose with 72% decrease compared to the control. Inhibition of AChE enzyme in BPA-treated crayfish can lead to accumulation of acetylcholine (ACh) in the nervous system, overstimulation of cholinergic synapses, and impairment of neuromuscular attachment and vital activities of the central nervous system. Similar to our results, AChE inhibition was observed by various researchers as a result of BPA application in various fish species (Chen et al. 2017; Li et al. 2016; Hamed and Abdel-Tawwab 2017).
CaEs are cellular phase 1 metabolizing enzymes responsible for the hydrolysis of carboxyesters to the corresponding alcohol and carboxylic acid (Hatfield et al. 2016). In this study, CaE activity was inhibited without being dose-dependent. Inhibition at all concentrations was statistically significant. The highest inhibition compared to the control was observed at a concentration of LC50/8 with a rate of about 56%. The inhibition rate in the LC50 concentration was also around 56%. Inhibition of CaE suggests that BPA exposure may affect the normal detoxification functions in crayfish.
GST is a phase II detoxification enzyme which performs the conjugation function that converts phase I detoxification products into a more water-soluble form. In this way, it prevents reactive free radical accumulation and cellular damage (Chahine and O’Donnell 2011). GST is a detoxifying enzyme that mostly functions in the liver and prevents the toxic effects of various pollutants (Jain et al. 2010; Hamed 2015). In the present study, there was a non-dose-dependent increase in GST activity compared to control at all BPA concentrations. The highest increase was in the LC50/2 concentration as 387.24 ± 1.43 nmol min−1 mg protein−1 while this value was 171.87 ± 3.45 nmol min−1 mg protein−1 at control group. Similar results have been reported by many researchers (Wu et al. 2011a, b; Li et al. 2016; Kaya and Kaptaner 2016; Faheem and Lone 2017), i.e., that GST activity increased as a result of applying BPA in different concentrations in different fish species. GST is known to add the GSH group to xenobiotics or its metabolites, making them more soluble in water (Moorhouse and Casida 1992). Enhanced activity of GST may be thought to increase to detoxify BPA. If we want to evaluate this situation in invertebrates, we can give an example of a study by Park et al. (2018), which showed that the marine rotifer Brachionus koreanus given 1, 5, or 10 mg L−1 BPA for 24 h showed significantly increased GST activity in a dose-dependent manner. This shows that GST activity increases with short-term exposure to BPA. Moreover, GST activity was generally enhanced in marine invertebrate such as the bivalve mollusk (Mytilus gapploprovincialis), the freshwater snail (Bellamya purificata), or the water flea (Daphnia magna) exposed to BPA at various concentrations and durations (Wu et al. 2011a; Canesi et al. 2008; Jemec et al. 2012).
GR maintains reduced glutathione status by providing overall homeostatic oxido-reducing balance in the cell, and thus protects cells against reactive oxygen metabolites (Djordjevic et al. 2010). GR activity showed a significant dose-dependent decrease in all groups. In the LC50 and LC50/2 dose groups, significant inhibitions were observed at the rates of 64% and 59%, respectively. BPA is an endocrine disrupting agent and GR is inhibited due to oxidative stress occurring in organisms exposed to high concentrations of EDC. Glutathione reductase provides antioxidant protection to the cell by participating in the recycling of GSH from GSSG (Elia et al. 2003). With the effect of GR, the balance between GSH and GSSG inside the cell is maintained through the recycling of GSH (Schafer and Buettner 2001). Therefore, GSH, GSSG, and total glutathione contents are also reduced, so the glutathione-containing defense system remains weak against high doses of BPA exposure. Some researchers have reported a decrease in GSH content due to environmental oxidative stresses originating from xenobiotics (Verma and Srivastava 2003; Carvan et al. 2001; Velisek et al. 2011; Stara et al. 2014). Conversely, when Seoane et al. (2021) applied different concentrations of BPA (3.75, 7.5, 15, 30 mg L−1) to Asian clam Corbicula fluminea for 96 h, they observed significant dose-dependent increases in GR activity. In this study, although BPA was applied at the same time as ours, the doses applied were lower. This means that, at low doses of BPA exposure, GR activity first increases and is inhibited as the dose increases.
SOD is an important antioxidant enzyme that is involved in inhibiting the oxidant formation that occurs due to the proliferation of superoxide radicals in the cell (Zhang et al. 2004). In this study, SOD activity increased in a dose-dependent manner. Increases with LC50 and LC50/2, the highest concentrations, were statistically significant compared to the control. There was a 47% increase in SOD activity at the LC50 dose compared to the control. Increased SOD activity may be due to increased O2− production, which is considered to be the first defense mechanism against oxidative stress caused by xenobiotics. These results indicate that the antioxidant system is activated in response to BPA exposure (Clasen et al. 2014; Hemalatha et al. 2015).
GPx is an important antioxidant enzyme that plays a role in protecting organisms against the destructive effects of hydrogen peroxide (Orbea et al. 2002). In this study, GPx activity increased depending on the BPA concentration applied. The increase at the LC50/8 concentration was insignificant compared to the control, while the increase at the other concentrations was significant. GPx activity was highest in the LC50 group with a rate of approximately 70%. These results match with those of Wu et al. (2011a), who recorded significant increases in GPx activity in medaka (O. latipes) after treatment with BPA. These increases in GPx activity may be a function of the protective role of GPx against cell damage caused by oxidant production (El-Atti et al. 2019). In contrast, when Zhang et al. (2020) administered 225 μg L−1 BPA to red swamp crayfish Procambarus clarkii for 1 week, they observed significant increases in ROS levels and inhibition of SOD and GPx activities in the hepatopancreas. The level of biomarkers associated with oxidative stress first increases against xenobiotic exposure and then decreases due to the occurrence of oxidative damage (Rosenbaum et al. 2012). Therefore, we can attribute the inhibition of these enzymes in this study and their high activity in our study to our short-term application of BPA. Similarly, in the crab Charybdis japonica, SOD and GPx activities changed in response to 15 days of BPA administration at 0.125, 0.25, 0.50, and 1.00 mg L−1 in hemocytes, showing increases at the beginning of exposure and then decreases in a dose- and time-dependent manner (Peng et al. 2018).
MDA levels were significantly increased in a dose-dependent manner in all groups except for LC50/8. The highest level was at the LC50 dose with a rate of about 49%. An increased MDA content is associated with increased lipid peroxidation, which can lead to loss of cell integrity, i.e., increased cell permeability and DNA damage (Dominguez‐Rebolledo et al. 2010). Increasing MDA levels in this study may mean that oxidative stress had occurred as a result of BPA application (Chitra et al. 2003). In a study conducted by Abdel‐Tawwab and Hamed (2018), the MDA level increased significantly as a result of BPA application to Oreochromis niloticus for 6 weeks. The increase in the level of MDA can be attributed of a then-excessive production of ROS, which may be associated with the induction of oxidative stress after BPA exposure (Chitra et al. 2003).
ALT activity increased significantly compared to the control at all concentrations except for LC50/8. In terms of AST and LDH activities, a significant increase was observed at all BPA concentrations. These results coincide with a study by Eweda et al. (2020), who observed that hepatic ALT and AST values increased as a result of BPA administration to rats. Korkmaz et al. (2010) demonstrated that BPA treatment increased liver ALT, AST, and LDH activities and caused marked defects in liver morphology. The LDH and transaminases (AST and ALT) can be used for the evaluation of hepatotoxicity, and changes in AST and LDH activities may be considered an indicator of tissue damage under toxic substance stress (Rao 2006; Gholami-Seyedkolaei et al. 2013). Considering these views, in our results, it can be said that the high transaminase activity is due to the pathological effects of BPA on crayfish, and the increase in LDH activity is due to the fact that BPA reduces ATP production by blocking oxidative phosphorylation in mitochondria, thus creating hypoxic conditions.
Oxidative stress is the disruption of the balance between the oxidant and antioxidant systems as a result of the oxidant capacity exceeding the antioxidant capacity. TAC, TOSC, and OSI are parameters that reflect the general oxidative stress state of the organism (Aslan et al. 2007). TAC induced by hydroxyl radicals refers to the total antioxidative effect against free radical reactions (Karsen et al. 2012). TOSC reflects the total amount of oxidant molecules a chemical inducer creates in the organism. In our study, the TAC level was found to be significantly lower than the control in all BPA-applied groups. The TOSC level increased significantly due to the dose increase of BPA, except for the LC50/8 dose. It has been reported that BPA binds to oxygen radicals, converting them into various reactive metabolites that increase ROS production in the cell and inhibit antioxidant enzymes (Vahdati Hassani et al. 2018). It was observed that the TAC level in the colon, serum, and liver of BPA-treated mice was significantly lower than that in the control (Wang et al. 2019). OSI is an indicator showing the relationship between free radical–forming agents that cause oxidative stress in the organism and antioxidant defense systems against them (Sayed and Khalil 2016). In our study, as the BPA concentration applied increased, the OSI increased significantly compared to the control. Similar to our findings, in many animal studies in the literature, a number of BPA concentrations have been shown to significantly alter the total oxidative stress status of various tissues and organs (Hassan et al. 2012; Kalb et al. 2016; Moghaddam et al. 2015). In the freshwater bivalve Unio tumidus, 14 days of exposure to 200 ng L−1 BPA induced oxidative damage through excessive ROS production and DNA instability (Gnatyshyna et al., 2019).
ATPases are membrane-bound enzymes that play a role in cellular volume, osmotic regulation, pressure, and membrane permeability in fish (Sancho et al. 2003; Marshall and Grosell 2005). Fish gills play an important role in ion and acid–base homeostasis, gas exchange, filter feeding, and nitrogen excretion (Oğuz et al. 2018). Because the gill is the first organ to come into direct contact with water-soluble toxic substances, it is more sensitive to water pollutants and toxic injuries (Mirghaed et al. 2018). BPA is lipophilic, so it passes into biological bodies through breathing and nutrition then accumulates in fish through the gills and skin, so the gill is a potential target organ for BPA toxicity (Ismail et al. 2018). Na+/K+ ATPase, located on the gill membranes of fish, is responsible for the active transport of Na+ and K+ into and out of the cell; thus, the gill is the most suitable organ in which activity of Na+/K+ ATPase is observed (Li et al. 2011). Muscle tissue is the main consumable part of the fish and Ca2+ ATPase enzyme plays a role in ion metabolism in muscles (Saxena et al. 2000). For these reasons, we thought that the gill and muscle tissues were the most suitable tissues for evaluating ATPase activities. To our knowledge, this is the first demonstration of the toxic effects of BPA on ATPase activity in the crayfish. ATPase activity values in the gill and muscle tissues are shown in Table 4. In this study, all ATPase activities in both gill and muscle tissues were significantly inhibited. When all BPA concentrations were compared, the highest ATPase inhibitions were at the LC50 dose in both gill and muscle. Na+/K+ ATPase activity in the gill was inhibited at approximate rates of 26% and 24%, respectively, at the LC50 and LC50/2 doses. Activities of Mg2+ ATPase, Ca2+ ATPase, and total ATPase in the gill were inhibited in the rates of 46%, 39%, and 44%, respectively at the LC50 concentration. The inhibition rates of Na+/K+ ATPase, Mg2+ ATPase, Ca2+ ATPase, and total ATPase in the muscle were 36%, 48%, 43%, and 29%, respectively, at the LC50 dose. Na+/K+ ATPase is the most important ATPase in fish osmoregulation (Dogan et al. 2015).
Osmoregulator disruptions have been reported in fish exposed to EDC (McCormick et al. 2005). In this study, inhibition of Na+/K+ ATPase may mean that the transport of vital ions and nutrients is reduced due to the breakdown of cell membrane structure after BPA exposure (Begum 2011). Similar to our findings, Zhou et al. (2011) observed that activity of Na+/K+ ATPase decreased and oxidative damage occurred due to the disruption of cellular ionic homeostasis as a result of exposure of abalone embryos to BPA. In another study, Krishnapriya et al. (2017) observed that Na+/K+ ATPase activity decreased significantly in the gills of the carp Labeo rohita, after they applied BPA at sublethal concentrations. In the liver of Anabas testudineus, which was chronically exposed to BPA, Na+/K+ ATPase activity was inhibited, indicating decreased phosphorylation in the liver (Kumar and Sunny 2014). In this study, Ca2+ ATPase activity was significantly inhibited at all BPA concentrations. Since Ca2+ ATPase is an enzyme localized in the sarcoplasmic reticulum tubules that removes Ca2+ ions from the cytosol and lumen to balance Ca2+ levels in the cell, BPA application may have disrupted the intracellular ion balance by causing excessive accumulation of Ca2+ in the cell (Saxena et al. 2000). Similar to our results, Jalal et al. (2018) reported that low doses of BPA disrupted the structure of the calcium channels of cells by inhibiting Ca2+ ATPase, so Ca2+ homeostasis could not be achieved normally. Mg2+ ATPase is an enzyme that plays an important role in oxidative phosphorylation as well as transepithelial regulation of Mg2+ ions (Parvez et al. 2006). Since mitochondrial Mg2+ ATPase is an enzyme associated with energy metabolism, inhibition of this enzyme impairs oxidative phosphorylation (Dogan et al. 2015). Therefore, in our study, BPA may have disrupted the structure of mitochondrial membranes by inhibiting Mg2+ ATPase in gill and muscle tissues; thus, oxidative energy could not be converted into phosphate energy (Shwetha and Hosetti 2012). As can be seen from the inhibition rates, Na+/K+ ATPase inhibition was higher in muscle than in the gill, while the total ATPase rate was higher in the gill than in the muscle. Other inhibition rates were similar to each other. Comparing tissues in general, the results show that muscle tissue in crayfish is more sensitive to ATPase activity than gills. Supporting our findings, some researchers (Yoloğlu 2019; Uçkun and Öz 2020, 2021) have observed that ATPases are significantly altered in gill and muscle tissues as a result of pesticide exposure.
The IBR indices combining the responses of all biomarkers into a single overall stress index in the hepatopancreas, gill, and muscle are shown in Figs. 1, 2, and 3, respectively. IBR indices were computed using all biomarker data. There is no single biomarker response that can reveal the impact of a chemical on an organism without a doubt. To relate exposure to response and provide improved prediction tools for the evaluation of environmental exposure to pollutants, many biomarkers with various endpoints are required (Venturino et al. 2003). Using a battery of biomarkers and IBR, which is computed by integrating numerous biomarkers into a single value, is a more practical tool (Güngördü et al. 2016). Besides field studies, IBR has recently been used in laboratory research to determine the toxicological effects of xenobiotics on various species (Uçkun and Özmen 2021). The varying patterns of star plots in our IBR index figures reflect the integration profiles of multiple biochemical parameter responses in tissues (Kim and Jung 2016) exposed to different concentrations of BPA. Since the biomarkers analyzed in hepatopancreas were different from those in gill and muscle, different patterns of star plots were produced according to tissue type in the IBR index graphs. Hepatopancreatic IBR values with the LC50/4, LC50/2, and LC50 treatments were 2.42, 2.86, and 3.91 times higher than control, respectively, suggesting that LC50 and LC50/2 doses of BPA are more toxic to crayfish. This can also be understood from the fact that biomarker responses in the hepatopancreas were mostly induced as the applied BPA concentration increases, as can be seen in Table 4. The correlation between BPA concentration and IBR values shows that the hepatopancreas is significantly affected by BPA toxicity due to its detoxification function. In the gill and muscle tissues, as the concentration of BPA increased, the IBR values decreased compared to the control, and it was completely suppressed at the LC50 dose. In muscle, the LC50/8, LC50/4, LC50/2, and LC50 doses caused approximately 2.55-, 9.28-, 54.69-, and 100-fold decreases in the IBR values compared to the control, respectively. For the gills, these inhibition values were 8.91-, 21.18-, 22.95-, and 100-fold, respectively. Inhibitory effects in these tissues can be attributed to the increased toxic effects of BPA and inadequate individual biomarker responses (Kim and Jung 2016). Findings similar to ours regarding the IBR index were observed by Demirci et al. (2018) as a result of the application of various pesticides to Gammarus kischineffensis. Many researchers (Kim et al. 2013, 2014b; Kim and Jung 2016; Samanta et al. 2018a, b; Uçkun et al. 2021) have suggested that the IBR index is a useful tool for the integrative quantification of the molecular and biochemical biomarker responses in a short-term exposure to xenobiotics and a powerful tool to monitor aquatic pollution in situ. Kim et al. (2013) applied various concentrations of copper and benzo[a]pyrene to freshwater pale chub Zacco platypus and used the IBR index to compare their toxicity; they reported that IBR values correlated well with copper and benzo[a]pyrene concentrations. In another study, the same researchers reported that the integration of multilevel biomarker responses at the molecular, biochemical, and physiological levels was highly correlated with cadmium and benzo[k]fluoranthene concentrations (Kim et al. 2014a). Kim and Jung (2016) reported that several multilevel biomarker responses from IBR values in pale chub (Z. platypus) were significantly associated with metals accumulated in Z. platypus.

The IBR index of hepatopancreas

The IBR index of gill

The IBR index of muscle
Conclusion
While there is extensive information about the toxic effects of BPA on many fish species in the literature, its effect has not been studied in Astacus leptodactylus (determining the LC50 value and selected biomarkers for the first time in this organism), an economically important consumed organism in fresh waters, pointing to the originality of this study. The A. leptodactylus species used in this study are exported regularly every year, so the toxic effects of BPA, an EDC commonly found in aquatic environments on this species, have contributed greatly to the literature. In this study, it can be concluded that short-term application of BPA to crayfish significantly affects the activities of cholinergic system enzymes (AChE), detoxification enzymes (GST, CaE), metabolic enzymes (ALT, AST, LDH, ATPases), and oxidative stress/antioxydant markers (SOD, GPx, GR, MDA, TAC, TOSC). In conclusion, Astacus leptodactylus and selected biomarkers are useful indicators in the assessment of BPA toxicity on non-target organisms in aquatic environments.
Data availability
The data is available on request from the corresponding author.
Change history
23 December 2021
Insertion of space in equation.
References
Abdel-Tawwab M, Hamed HS (2018) Effect of bisphenol A toxicity on growth performance, biochemical variables, and oxidative stress biomarkers of Nile tilapia, Oreochromis niloticus (L.). J Appl Ichthyol 34:1117–1125. https://doi.org/10.1111/jai.13763
Ahn D, Kawai T, Kim S, Rho HS, Jung J, Kim WS, Lim BJ, Kim MS, Min GS (2006) Phylogeny of Northern Hemisphere freshwater crayfishes based on 16SrRNA gene analysis. Korean J Genetic 28:185–192
Anderson RL (1982) Toxicity of fenvalerate and permethrin to several nontarget aquatic invertebrates. Environ Entomol 11:1251–1257. https://doi.org/10.1093/ee/11.6.1251
Arzate-Cárdenas MA, Martínez-Jerónimo F (2012) Energy reserve modification in different age groups of Daphnia schoedleri (Anomopoda: Daphniidae) exposed to hexavalent chromium. Environ Toxicol Pharmacol 34:106–116. https://doi.org/10.1016/j.etap.2012.03.003
Aslan M, Sabuncu T, Kocyigit A, Celik H, Selek S (2007) Relationship between total oxidant status and severity of diabetic nephropathy in type 2 diabetic patients. Nutr Metab Cardiovasc Dis 17:734–740. https://doi.org/10.1016/j.numecd.2006.08.005
ASTM (American Society for Testing and Materials), E729–96 (2014) Standard guide for conducting acute toxicity tests on test materials with fishes, macroinvertebrates, and amphibians. In: Annual Book of ASTM Standards, Vol. 11.06. ASTM International, West Conshohocken, PA, United States, pp 1–22. https://doi.org/10.1520/E0729-96R14
Atkinson A, Gatemby AO, Lowe AG (1973) The determination of inorganic ortophosphate in biological systems. Biochim Biophys Acta 320:195–204. https://doi.org/10.1016/0304-4165(73)90178-5
Atlı G, Canlı M (2011) Essential metal (Cu, Zn) exposures alter the activity of ATPases in gill, kidney and muscle of tilapia Oreochromis niloticus. Ecotoxicology 20:1861–1869. https://doi.org/10.1007/s10646-011-0724-z
Barim-Oz O (2018) The effects on some non-enzymatic antioxidants and oxidative stress of Astacus leptodactylus (Esch., 1823) of starvation periods. Aquac Nutr 24:492–503. https://doi.org/10.1111/anu.12582
Begum G (2011) Organ-specific ATPase and phosphorylase enzyme activities in a food fish exposed to a carbamate insecticide and recovery response. Fish Physiol Biochem 37:61–69. https://doi.org/10.1007/s10695-010-9417-4
Bell JG, Cowey CB, Adro JW, Shanks AM (1985) Some effects of vitamine and selenium deprivation on tissue enzyme levels and indices of tissue peroxidation in rainbow trout (Salmo gairdneri). Br J Nutr 53:149–157. https://doi.org/10.1079/BJN19850019
Bhandari RK, Deem SL, Holliday DK, Jandegian CM, Kassotis CD, Nagel SC, Tillitt DE, Saal FSV, Rosenfeld CS (2015) Effects of the environmental estrogenic contaminants bisphenol A and 17α-ethinyl estradiol on sexual development and adult behaviors in aquatic wildlife species. Gen Comp Endocrinol 214:195–219. https://doi.org/10.1016/j.ygcen.2014.09.014
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254. https://doi.org/10.1016/0003-2697(76)90527-3
Broeg K, Lehtonen KK (2006) Indices for the assessment of environmental pollution of the Baltic Sea coasts: ıntegrated assessment of a multi-biomarker approach. Mar Pollut Bull 53:508–522. https://doi.org/10.1016/j.marpolbul.2006.02.004
Canesi L, Borghi C, Ciacci C, Fabbri R, Lorusso LC, Vergani L, Marcomini A, Poiana G (2008) Short-term effects of environmentally relevant concentrations of EDC mixtures on Mytilus galloprovincialis digestive gland. Aquat Toxicol 87(4):272–279. https://doi.org/10.1016/j.aquatox.2008.02.007
Carvan MJ, Sonntaq DM, Cmar CB, Cook RS, Curran MA, Miller GL (2001) Oxidative stress in zebrafish cells: potential utility of transgenic zebrafish as a deployable sentinel for hazard ranking. Sci Total Environ 274:183–196. https://doi.org/10.1016/S0048-9697(01)00742-2
Chahine S, O’Donnell MJ (2011) Interactions between detoxification mechanisms and excretion in malpighian tubules of Drosophila melanogaster. J Exp Biol 214:462–468. https://doi.org/10.1242/jeb.048884
Chan WK, Chan KM (2012) Disruption of the hypothalamicpituitary-thyroid axis in zebrafish embryo-larvae following waterborne exposure to BDE-47, TBBPA and BPA. Aquat Toxicol 108:106–111. https://doi.org/10.1016/j.aquatox.2011.10.013
Chen Q, Yin D, Jia Y, Schiwy S, Legradi J, Yang S, Hollert H (2017) Enhanced uptake of BPA in the presence of nanoplastics can lead to neurotoxic effects in adult zebrafish. Sci Total Environ 609:1312–1321. https://doi.org/10.1016/j.scitotenv.2017.07.144
Chepelev NL, Enikanolaiye MI, Chepelev LL, Almohaisen A, Chen Q, Scoggan KA, Coughlan MC, Cao XL, Jin X, Willmore WG (2013) Bisphenol A activates the Nrf1/2-antioxidant response element pathway in HEK 293 cells. Chem Res Toxicol 26:498–506. https://doi.org/10.1021/tx400036v
Chitra KC, Latchoumycandane C, Mathur PP (2003) Induction of oxidative stress by bisphenol A in the epididymal sperm of rats. Toxicology 185:119–127. https://doi.org/10.1016/S0300-483X(02)00597-8
Clasen B, Leitemperger J, Murussi C, Pretto A, Menezes C, Dalabona F, Marchezan E, Adaime MB, Zanella R, Loro VL (2014) Carbofuran promotes biochemical changes in carp exposed to rice field and laboratory conditions. Ecotoxicol Environ Saf 101:77–82. https://doi.org/10.1016/j.ecoenv.2013.12.012
Cribb AE, Leeder JS, Spielberg SP (1989) Use of a microplate reader in an assay of glutathione reductase using 5,5-dithiobis (2-nitrobenzoic acid). Anal Biochem 183:195–196. https://doi.org/10.1016/0003-2697(89)90188-7
Demirci Ö, Güven K, Asma D, Ögüt S, Uğurlu P (2018) Effects of endosulfan, thiamethoxam, and indoxacarb in combination with atrazine on multi-biomarkers in Gammarus kischineffensis. Ecotoxicol Environ Saf 147:749–758. https://doi.org/10.1016/j.ecoenv.2017.09.038
Djordjevic J, Djordjevic A, Adzic M, Niciforovic A, Radojcic MB (2010) Chronic stress differentially affects antioxidant enzymes and modifies the acute stress response in liver of wistar rats. Physiol Res 59:729–736. https://doi.org/10.33549/physiolres.931862
Dogan Z, Atli G, Canli M (2015) Effects of lead on ATPases in tissues of freshwater fish (Oreochromis niloticus) in differing calcium levels. Turk J Fish Aquat Sci 15:223–233. https://doi.org/10.4194/1303-2712-v15_2_04
Dominguez-Rebolledo ÁE, Fernández-Santos MR, Bisbal A, Ros-Santaella JL, Ramón M, Carmona M, Martínez-Pastor F, Garde JJ (2010) Improving the effect of incubation and oxidative stress on thawed spermatozoa from red deer by using different antioxidant treatments. Reprod Fertil Dev 22:856–870. https://doi.org/10.1071/RD09197
Dorn PB, Chou C, Gentempo JJ (1987) Degradation of bisphenol A in natural waters. Chemosphere 16:1501–1507. https://doi.org/10.1016/0045-6535(87)90090-7
Duan X, Li Y, Li X, Zhang D, Gao Y (2014) Alkylphenols in surface sediments of the Yellow Sea and East China Sea inner shelf: occurrence, distribution and fate. Chemosphere 107:265–273. https://doi.org/10.1016/j.chemosphere.2013.12.054
El-Atti MA, Desouky MMA, Mohamadien A, Said RM (2019) Effects of titanium dioxide nanoparticles on red swamp crayfish, Procambarus clarkii: bioaccumulation, oxidative stress and histopathological biomarkers. Egypt J Aquat Res 45:11–18. https://doi.org/10.1016/j.ejar.2019.01.001
Elia AC, Galarini R, Taticchi MI, Dörr AJM, Mantilacci L (2003) Antioxidant responses and bioaccumulation in Ictalurus melas under mercury exposure. Ecotoxicol Environ Saf 55:162–167. https://doi.org/10.1016/S0147-6513(02)00123-9
Ellman GL, Courtney KD, Andres V, Featherstone RM (1961) A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem Pharmacol 7:88–95. https://doi.org/10.1016/0006-2952(61)90145-9
Erel O (2005) A new automated colorimetric method for measuring total oxidant status. Clin Biochem 38:1103–1111. https://doi.org/10.1016/j.clinbiochem.2005.08.008
Esteve C, Herrero L, Gómara B, Quintanilla-López JE (2016) Fast and simultaneous determination of endocrine disrupting compounds by ultra-high performance liquid chromatography-tandem mass spectrometry. Talanta 146:326–334. https://doi.org/10.1016/j.talanta.2015.08.064
Eweda SM, Newairy ASA, Abdou HM, Gaber AS (2020) Bisphenol A-induced oxidative damage in the hepatic and cardiac tissues of rats: the modulatory role of sesame lignans. Exp Ther Med 19:33–44. https://doi.org/10.3892/etm.2019.8193
Faheem M, Lone KP (2017) Oxidative stress and histopathologic biomarkers of exposure to bisphenol-A in the freshwater fish, Ctenopharyngodon idella. Braz J Pharm Sci 53:3–9. https://doi.org/10.1590/s2175-97902017000317003
Fromme H, Küchler T, Otto T, Pilz K, Müller J, Wenzel A (2002) Occurrence of phthalates and bisphenol A and F in the environment. Water Res 36:1429–1438. https://doi.org/10.1016/S0043-1354(01)00367-0
Ge LC, Chen ZJ, Liu H, Zhang KS, Su Q, Ma XY, Huang HB, Zhao ZD, Wang YY, Giesy JP, Du J, Wang HS (2014) Signaling related with biphasic effects of bisphenol A (BPA) on Sertoli cell proliferation: a comparative proteomic analysis. Biochim Biophys Acta 1840:2663–2673. https://doi.org/10.1016/j.bbagen.2014.05.018
Gholami-Seyedkolaei SJ, Mirvaghefi A, Farahmand H, Kosari AA (2013) Effect of a glyphosate-based herbicide in Cyprinus carpio: assessment of acetylcholinesterase activity, hematological responses and serum biochemical parameters. Ecotoxicol Environ Saf 98:135–141. https://doi.org/10.1016/j.ecoenv.2013.09.011
Gibert JM, Karch F, Schlötterer C (2011) Segregating variation in the polycomb group gene cramped alters the effect of temperature on multiple traits. PLoS Genet 7:1–12. https://doi.org/10.1371/journal.pgen.1001280
Giulivo M, Lopez de Alda M, Capri E, Barceló D (2016) Human exposure to endocrine disrupting compounds: their role in reproductive systems, metabolic syndrome and breast cancer. A Review Environ Res 151:251–264. https://doi.org/10.1016/j.envres.2016.07.011
Gnatyshyna L, Falfushynska H, Horyn O, Khoma V, Martinyuk V, Mishchuk O, Mishchuk N, Stoliar O (2019) Biochemical responses of freshwater mussel Unio tumidus to titanium oxide nanoparticles, bisphenol A, and their combination. Ecotoxicology 28(8):923–937. https://doi.org/10.1007/s10646-019-02090-6
Grasselli F, Baratta L, Baioni L, Bussolati S, Ramoni R, Grolli S, Basini G (2010) Bisphenol A disrupts granulosa cell function. Domest Anim Endocrinol 39:34–39. https://doi.org/10.1016/j.domaniend.2010.01.004
Gulley DD (1996) West Inc, Toxstat Version 3.5. Fish Physiology and Toxicology Laboratory. University of Wyoming, Laramie, WY, USA
Güngördü A, Uçkun M, Yoloğlu E (2016) Integrated assessment of biochemical markers in premetamorphic tadpoles of three amphibian species exposed to glyphosate-and methidathion-based pesticides in single and combination forms. Chemosphere 144:2024–2035. https://doi.org/10.1016/j.chemosphere.2015.10.125
Habig WH, Pabst MJ, Jakoby WB (1974) Glutathione S-transferases, the first enzymatic step in mercapturic acid formation. J Biol Chem 249:7130–7139. https://doi.org/10.1016/S0021-9258(19)42083-8
Hamed HS (2015) Impact of a short-term malathion exposure of Nile tilapia, (Oreochromis niloticus): the protective role of selenium. Int J Environ Monit Anal 3:30–37. https://doi.org/10.11648/j.ijema.s.2015030501.15
Hamed HS, Abdel-Tawwab M (2017) Ameliorative effect of propolis supplementation on alleviating bisphenol-A toxicity: growth performance, biochemical variables, and oxidative stress biomarkers of Nile tilapia, Oreochromis niloticus (L.) fingerlings. Comp Biochem Physiol C Toxicol Pharmacol 202:63–69. https://doi.org/10.1016/j.cbpc.2017.08.001
Harlıoğlu MM (2004) The present situation of freshwater crayfish, Astacus leptodactylus (Eschscholtz, 1823) in Turkey. Aquaculture 230:181–187. https://doi.org/10.1016/S0044-8486(03)00429-0
Hassan ZK, Elobeid MA, Virk P, Omer SA, ElAmin M, Daghestani MH, AlOlayan EM (2012) Bisphenol A induces hepatotoxicity through oxidative stress in rat model. Oxid Med Cell Longev 2012:1–6. https://doi.org/10.1155/2012/194829
Hatami M, Banaee M, Nematdoost Haghi B (2019) Sub-lethal toxicity of chlorpyrifos alone and in combination with polyethylene glycol to common carp (Cyprinus carpio). Chemosphere 219:981–988. https://doi.org/10.1016/j.chemosphere.2018.12.077
Hatfield MJ, Umans RA, Hyatt JL, Edwards CC, Wierdl M, Tsurkan L, Taylor MR, Potter PM (2016) Carboxylesterases: general detoxifying enzymes. Chem Biol Interact 259:327–331. https://doi.org/10.1016/j.cbi.2016.02.011
Hayashi L, Sheth M, Young A, Kruger M, Wayman GA, Coffin AB (2015) The effect of the aquatic contaminants bisphenol-A and PCB-95 on the zebrafish lateral line. Neurotoxicology 46:125–136. https://doi.org/10.1016/j.neuro.2014.12.010
Hemalatha D, Amala A, Rangasamy B, Nataraj B, Ramesh M (2015) Sublethal toxicity of quinalphos on oxidative stress and antioxidant responses in a freshwater fish Cyprinus carpio. Environ Toxicol 31:1399–1406. https://doi.org/10.1002/tox.22145
Hohenblum P, Gans O, Moche W, Scharf S, Lorbeer G (2004) Monitoring of selected estrogenic hormones and industrial chemicals in groundwaters and surface waters in Austria. Sci Total Environ 333(1–3):185–193. https://doi.org/10.1016/j.scitotenv.2004.05.009
Hong Y, Yang X, Huang Y, Yan G, Cheng Y (2018) Assessment of the oxidative and genotoxic effects of the glyphosate-based herbicide roundup on the freshwater shrimp, Macrobrachium nipponensis. Chemosphere 210:896–906. https://doi.org/10.1016/j.chemosphere.2018.07.069
Hulak M, Gazo I, Shaliutina A, Linhartova P (2013) In vitro effects of bisphenol A on the quality parameters, oxidative stress, DNA integrity and adenosine triphosphate content in sterlet (Acipenser ruthenus) spermatozoa. Comp Biochem Physiol C Toxicol Pharmacol 158:64–71. https://doi.org/10.1016/j.cbpc.2013.05.002
Im J, Löffler FE (2016) Fate of bisphenol A in terrestrial and aquatic environments. Enviro Sci Technol 50(16):8403–8416. https://doi.org/10.1021/acs.est.6b00877
Ismail NAH, Wee SY, Aris AZ (2018) Bisphenol A and alkylphenols concentrations in selected mariculture fish species from Pulau Kukup, Johor, Malaysia. Mar Pollut Bull 127:536–540. https://doi.org/10.1016/j.marpolbul.2017.12.043
Iwano H, Inoue H, Nishikawa M, Fujiki J, Yokota H (2018) Biotransformation of bisphenol a and its adverse effects on the next generation. In: Ahmed RG (ed) Endocrine Disruptors, 1st edn. IntechOpen, London, pp 63–77. https://doi.org/10.5772/intechopen.78275
Jain M, Ghanashyam C, Bhattacharjee A (2010) Comprehensive expression analysis suggests over lapping and specific roles of rice glutathione S-transferase genes during development and stress responses. BMC Genomics 11:1–17. https://doi.org/10.1186/1471-2164-11-73
Jalal N, Surendranath AR, Pathak JL, Yu S, Chung CY (2018) Bisphenol A (BPA) the mighty and the mutagenic. Toxicol Rep 5:76–84. https://doi.org/10.1016/j.toxrep.2017.12.013
Jemec A, Tišler T, Erjavec B, Pintar A (2012) Antioxidant responses and whole-organism changes in Daphnia magna acutely and chronically exposed to endocrine disruptor bisphenol A. Ecotoxicol Environ Saf 86:213–218. https://doi.org/10.1016/j.ecoenv.2012.09.016
Kalb AC, Kalb AL, Cardoso TF, Fernandes CG, Corcini CD, Varela Junior AS, Martínez PE (2016) Maternal transfer of bisphenol A during nursing causes sperm ımpairment in male offspring. Arch Environ Contam Toxicol 70:793–801. https://doi.org/10.1007/s00244-015-0199-7
Kankaya E, Kaptaner B, Doğan A, Celik I (2015) Toxicity of bisphenol a during the early life stages of Chalcalburnus tarichi (Pallas, 1811). Fresenius Environ Bull 24:977–985
Karsen H, Binici I, Sunnetcioğlu M, Baran AI, Ceylan MR, Selek S, Çelik H (2012) Association of paraoxonase activity and atherosclerosis in patients with chronic hepatitis. B Afr Health Sci 12:114–118. https://doi.org/10.4314/ahs.v12i2.6
Kaya O, Kaptaner B (2016) Antioxidant defense system parameters in isolated fish hepatocytes exposed to bisphenol A-Effect of vitamin C. Acta Biol Hung 67:225–235. https://doi.org/10.1556/018.67.2016.3.1
Kim WK, Jung J (2016) In situ impact assessment of wastewater effluents by integrating multi-level biomarker responses in the pale chub (Zacco platypus). Ecotoxicol Environ Saf 128:246–251. https://doi.org/10.1016/j.ecoenv.2016.02.028
Kim WK, Lee SK, Choi K, Jung J (2013) Integrative assessment of biomarker responses in pale chub (Zacco platypus) exposed to copper and benzo [a] pyrene. Ecotoxicol Environ Saf 92:71–78. https://doi.org/10.1016/j.ecoenv.2013.02.010
Kim WK, Lee SK, Park JW, Choi K, Cargo J, Schlenk D, Jung J (2014a) Integration of multi-level biomarker responses to cadmium and benzo [k] fluoranthene in the pale chub (Zacco platypus). Ecotoxicol Environ Saf 110:21–128. https://doi.org/10.1016/j.ecoenv.2014.08.025
Kim WK, Park JW, Lim ES, Lee SK, Kim J, Kim S, Lee SW, Choi K, Jung J (2014b) Tissue-specific antioxidant responses in pale chub (Zacco platypus) exposed to copper and benzo [a] pyrene. Bull Environ Contam Toxicol 92(5):540–545. https://doi.org/10.1007/s00128-014-1213-x
Kinch CD, Ibhazehiebo K, Jeong JH, Habibi HR, Kurrasch DM (2015) Low-dose exposure to bisphenol A and replacement bisphenol S induces precocious hypothalamic neurogenesis in embryonic zebrafish. Proc Natl Acad Sci U S A 112:1475–1480. https://doi.org/10.1073/pnas.1417731112
Kontogiannatos D, Swevers L, Zakasis G, Kourti A (2015) The molecular and physiological impact of bisphenol A in Sesamia nonagrioides (Lepidoptera: Noctuidae). Ecotoxicology 24(2):356–367
Korkmaz A, Ahbab MA, Kolankaya D, Barlas N (2010) Influence of vitamin C on bisphenol A, nonylphenol and octylphenol induced oxidative damages in liver of male rats. Food Chem Toxicol 48:2865–2871. https://doi.org/10.1016/j.fct.2010.07.019
Krishnapriya K, Shobana G, Narmadha S, Ramesh M, Maruthappan V (2017) Sublethal concentration of bisphenol A induces hematological and biochemical responses in an Indian major carp Labeo rohita. Ecohydrol Hydrobiol 17:306–313. https://doi.org/10.1016/j.ecohyd.2017.06.003
Kumar KG, Sunny F (2014) Effect of bisphenol A on oxidative metabolism in a freshwater teleost, Anabas testudineus (Bloch). India J Aquat Biol Fish 2:122–128
Kwak H, Bae M, Lee M, Lee Y, Lee B, Kang K, Chae C, Sung H, Shin J, Kim J, Mar W, Sheen Y, Cho M (2001) Effects of nonylphenol, bisphenol A, and their mixture on the viviparous swordtail fish (Xiphophorus helleri). Environ Toxicol Chem 20:787–795. https://doi.org/10.1002/etc.5620200414
Lei B, Sun S, Xu J, Feng C, Yu Y, Xu G, Wu M, Peng W (2018) Low-concentration BPAF-and BPF-induced cell biological effects are mediated by ROS in MCF-7 breast cancer cells. Environ Sci Pollut Res 25(4):3200–3208. https://doi.org/10.1007/s11356-017-9709-7
Leksrisawat B, Cooper AS, Gilberts AB (2010) Muscle receptor organs in the crayfish abdomen: a student laboratory exercise in proprioception. J vis Exp 45:1–10. https://doi.org/10.3791/2323
Li D, Chen Q, Cao J, Chen H, Li L, Cedergreen N, Xie H, Xie L (2016) The chronic effects of lignin-derived bisphenol and bisphenol A in Japanese medaka Oryzias latipes. Aquat Toxicol 170:199–207. https://doi.org/10.1016/j.aquatox.2015.11.024
Li ZH, Li P, Randak T (2011) Evaluating the toxicity of environmental concentrations of waterborne chromium (VI) to a model teleost, Oncorhynchus mykiss: a comparative study of in vivo and in vitro. Comp Biochem Physiol C Toxicol Pharmacol 153:402–407. https://doi.org/10.1016/j.cbpc.2011.01.005
Marshall WS, Grosell M (2005) Ion transport, osmoregulation, and acid-base balance In The Physiology of Fishes (eds Evans DH, Claiborne J). CRC Press, Boca Raton, FL, pp 177–230
Mathieu-Denoncourt J, Wallace SJ, De Solla SR, Langlois VS (2016) Influence of lipophilicity on the toxicity of bisphenol a and phthalates to aquatic organisms. Bull Environ Contam Toxicol 97:4–10. https://doi.org/10.1007/s00128-016-1812-9
McCormick SD, O’Dea MF, Moeckel AM, Lerner DT, Björnsson BT (2005) Endocrine disruption of parr-smolt transformation and seawater tolerance of Atlantic salmon by 4-nonylphenol and 17 β-estradiol. Gen Comp Endocrinol 142:280–288. https://doi.org/10.1016/j.ygcen.2005.01.015
Mikolajewska K, Stragierowicz J, Gromadzinska J (2015) Bisphenol A application, sources of exposure and potential risks in infants, children and pregnant women. Int J Occup Med Environ Health 28:209–241. https://doi.org/10.13075/ijomeh.1896.00343
Mirghaed AT, Ghelichpour M, Mirzargar SS, Joshaghani H, Mousavi HE (2018) Toxic effects of indoxacarb on gill and kidney histopathology and biochemical indicators in common carp (Cyprinus carpio). Aquac Res 49:1616–1627. https://doi.org/10.1111/are.13617
Moghaddam HS, Samarghandian S, Farkhondeh T (2015) Effect of bisphenol A on blood glucose, lipid profile and oxidative stress indices in adult male mice. Toxicol Mech Methods 25:507–513. https://doi.org/10.3109/15376516.2015.1056395
Moorhouse KG, Casida JE (1992) Pesticides as activators of mouse liver microsomal glutathione-S-transferase. Pestic Biochem Physiol 44:83–90. https://doi.org/10.1016/0048-3575(92)90105-9
Oehlmann J, Schulte-Oehlmann U, Bachmann J, Oetken M, Lutz I, Kloas W, Ternes TA (2006) Bisphenol A induces superfeminization in the ramshorn snail Marisa cornuarietis (Gastropoda: Prosobranchia) at environmentally relevant concentrations. Environ Health Perspect 114:127–133. https://doi.org/10.1289/ehp.8065
Oğuz EK, Arihan O, Oğuz AR (2018) Oxidative and genotoxic effects of bisphenol A on primary gill cell culture of Lake Van Fish (Alburnus tarichi Güldenstädt, 1814). Chem Ecol 34:914–924. https://doi.org/10.1080/02757540.2018.1520846
Orbea A, Ortiz-Zarragoitia M, Solé M, Porte C, Cajaraville MP (2002) Antioxidant enzymes and peroxisome proliferation in relation to contaminant body burdens of PAHs and PCBs in bivalve molluscs, crabs and fish from the Urdaibai and Plentzia estuaries (Bay of Biscay). Aquat Toxicol 58:75–98. https://doi.org/10.1016/S0166-445X(01)00226-0
Orhan IE (2013) Nature: a substantial source of auspicious substances with acetylcholinesterase inhibitory action. Curr Neuropharmacol 11:379–387. https://doi.org/10.2174/1570159X11311040003
Ozmen M, Ayas Z, Güngördü A, Ekmekci GF, Yerli S (2008) Ecotoxicological assessment of water pollution in Sarıyar Dam Lake, Turkey. Ecotoxicol Environ Saf 70:163–173. https://doi.org/10.1016/j.ecoenv.2007.05.011
Pandey S, Parvez S, Sayeed I, Haque R, Bin-Hafeez B, Raisuddin S (2003) Biomarkers of oxidative stress: a comparative study of river Yamuna fish Wallago attu (Bl. and Schn.). Sci Total Environ 309:105–115. https://doi.org/10.1016/S0048-9697(03)00006-8
Park JC, Lee MC, Yoon DS, Han J, Kim M, Hwang UK, Jung JH, Lee JS (2018) Effects of bisphenol A and its analogs bisphenol F and S on life parameters, antioxidant system, and response of defensome in the marine rotifer Brachionus koreanus. Aquat Toxicol 199:21–29. https://doi.org/10.1016/j.aquatox.2018.03.024
Parvez S, Sayeed I, Raisuddin S (2006) Decreased gill ATPase activities in the freshwater fish Channa punctata (Bloch) exposed to a diluted paper mill effluent. Ecotoxicol Environ Saf 65:62–66. https://doi.org/10.1016/j.ecoenv.2005.07.010
Peng YQ, Wang MJ, Chen HG, Chen JH, Gao H, Huang HH (2018) Immunological responses in haemolymph and histologic changes in the hepatopancreas of Charybdis japonica (A. Milne-Edwards, 1861 (Decapoda: Brachyura: Portunidae) exposed to bisphenol. A J. Crustac Biol. 38(4):489–496. https://doi.org/10.1093/jcbiol/ruy046
Placer ZA, Cushman LL, Johnson BC (1966) Estimation of product of lipid peroxidation (malonyl dialdehyde) in biochemical systems. Anal Biochem 16:359–364. https://doi.org/10.1016/0003-2697(66)90167-9
Rao JV (2006) Biochemical alterations in euryhaline fish, Oreochromis mossambicus exposed to sub-lethal concentrations of an organophosphorus insecticide, monocrotophos. Chemosphere 65:1814–1820. https://doi.org/10.1016/j.chemosphere.2006.04.015
Rogers JA, Metz L, Yong VW (2013) Review: Endocrine disrupting chemicals and immune responses: a focus on bisphenol-A and its potential mechanisms. Mol Immunol 53:421–430. https://doi.org/10.1016/j.molimm.2012.09.013
Rosenbaum EA, Duboscq L, Soleño J, Montagna CM, Ferrari A, Venturino A (2012) Response of biomarkers in amphibian larvae to in situ exposures in a fruit-producing region in North Patagonia. Argentina Environ Toxicol Chem 31(10):2311–2317. https://doi.org/10.1002/etc.1950
Rudel RA, Melly SJ, Geno PW, Sun G, Brody JG (1998) Identification of alkylphenols and other estrogenic phenolic compounds in wastewater, septage, and groundwater on Cape Cod. Massachusetts Environ Sci Technol 32(7):861–869. https://doi.org/10.1021/es970723r
Samanta P, Im H, Na J, Jung J (2018a) Ecological risk assessment of a contaminated stream using multi-level integrated biomarker response in Carassius auratus. Environ Pollut 233:429–438. https://doi.org/10.1016/j.envpol.2017.10.061
Samanta P, Im H, Yoo J, Lee H, Kim NY, Kim W, Hwang SJ, Kim WK, Jung J (2018b) Comparative assessment of the adverse outcome of wastewater effluents by integrating oxidative stress and histopathological alterations in endemic fish. J Hazard Mater 344:81–89. https://doi.org/10.1016/j.jhazmat.2017.10.016
Sancho E, Fernández-Vega C, Ferrando MD, Andreu-Moliner E (2003) Eel ATPase activity as biomarker of thiobencarb exposure. Ecotoxicol Environ Saf 56:434–441. https://doi.org/10.1016/S0147-6513(03)00077-0
Santhoshkumar P, Shivanandappa T (1999) In vitro sequestration of two organophosphorus homologs by the rat liver. Chem Biol Interact 119:277–282. https://doi.org/10.1016/S0009-2797(99)00037-X
Saxena TB, Zachariassen KE, Jorgensen L (2000) Effects of ethoxyquin on the blood composition of turbot, Scophthalmus maximus L. Comp Biochem Physiol C Pharmacol Toxicol Endocrinol 127:1–9. https://doi.org/10.1016/S0742-8413(00)00131-6
Sayed AEH, Abu Khalil NS (2016) Oxidative stress ınduction in monosex Nile tilapia (Oreochromis niloticus, Linnaeus, 1758): a field study on the side effects of methyltestosterone. J Aquac Res Dev 7:1–6. https://doi.org/10.4172/2155-9546.1000416
Schafer FQ, Buettner GR (2001) Redox environment ofthe cell as viewed through glutathione disulfide/glutathione couple. Free Radic Biol Med 30:1191–1212. https://doi.org/10.1016/S0891-5849(01)00480-4
Seoane M, Cid Á, Herrero C, Esperanza M (2021) Comparative acute toxicity of benzophenone derivatives and bisphenol analogues in the Asian clam Corbicula fluminea. Ecotoxicology 30(1):142–153. https://doi.org/10.1007/s10646-020-02299-w
Shwetha AD, Hosetti BB (2012) Effect of exposure to sublethal concentrations of zinc cyanide on tissue ATPase activity in the fresh water fish, Cirrhinus mrigala (HAM). Arch Biol Sci 64:257–263. https://doi.org/10.2298/ABS1201257D
Staples CA, Dorn PB, Klecka GM, O’Block ST, Harris LR (1998) A review of the environmental fate, effects, and exposures of bisphenol A. Chemosphere 36:2149–2173. https://doi.org/10.1016/S0045-6535(97)10133-3
Stara A, Kouba A, Velisek J (2014) Effect of chronic exposure to prometryne on oxidative stress and antioxidant response in red swamp crayfish (Procambarus clarkii). Biomed Res Int 2014:1–6. https://doi.org/10.1155/2014/680131
Sun Y, Oberley LW, Li Y (1988) A simple method for clinical assay of superoxide dismutase. Clin Chem 34:497–500. https://doi.org/10.1093/clinchem/34.3.497
Tabrez S, Ahmad M (2009) Effect of wastewater intake on antioxidant and marker enzymes of tissue damage in rat tissues: implications for the use of biochemical markers. Food Chem Toxicol 47:2465–2478. https://doi.org/10.1016/j.fct.2009.07.004
Tatarazako N, Ishibashi H, Teshima K, Kishi K, Arizono K (2004) Effects of triclosan on various aquatic organisms. Environ Sci 11(2):133–140
Tsai WT (2006) Human health risk on environmental exposure to bisphenol-A: a review. J Environ Sci Health C Environ Carcinog Ecotoxicol Rev 24:225–255. https://doi.org/10.1080/10590500600936482
Uçkun AA, Öz ÖB (2020) Acute exposure to the fungicide penconazole affects some biochemical parameters in the crayfish (Astacus leptodactylus Eschscholtz, 1823. Environ Sci Pollut Res 27:35626–35637. https://doi.org/10.1007/s11356-020-09595-2
Uçkun AA, Öz ÖB (2021) Evaluation of the acute toxic effect of azoxystrobin on non-target crayfish (Astacus leptodactylus Eschscholtz, 1823) by using oxidative stress enzymes, ATPases and cholinesterase as biomarkers. Drug Chem Toxicol 44(5):550–557. https://doi.org/10.1080/01480545.2020.1774604
Uçkun M, Özmen M (2021) Evaluating multiple biochemical markers in Xenopus laevis tadpoles exposed to the pesticides thiacloprid and trifloxystrobin in single and mixed forms. Environ Toxicol Chem 40(10):2846–2860. https://doi.org/10.1002/etc.5158
Uçkun M, Yoloğlu E, Uçkun AA, Öz ÖB (2021) Acute toxicity of ınsecticide thiamethoxam to crayfish (Astacus leptodactylus):alterations in oxidative stress markers. ATPases and cholinesterase. Acta Chim Sloven 68(3):521–531. https://doi.org/10.17344/acsi.2021.6823
USEPA, US Environmental Protection Agency (2010) Bisphenol A Action Plan (CASRN 80–05–7) [CA Index Name: Phenol,4,4'-(1-methylethylidene)bis-], Released March 29, 2010. Accessed at: https://www.epa.gov/sites/default/files/2015-09/documents/bpa_action_plan.pdf
Vahdati Hassani F, Abnous K, Mehri S, Jafarian A, Birner-Gruenberger R, Yazdian Robati R, Hosseinzadeh H (2018) Proteomics and phosphoproteomics analysis of liver in male rats exposed to bisphenol A: mechanism of hepatotoxicity and biomarker discovery. Food Chem Toxicol 112:26–38. https://doi.org/10.1016/j.fct.2017.12.021
Velisek J, Stara A, Kolarova J, Svobodova Z (2011) Biochemical, physiological and morfological responses in common carp (Cyprinus carpio L.) after long-term exposure to terbutryn intration. Pestic Biochem Physiol 100:305–313. https://doi.org/10.1016/j.pestbp.2011.05.004
Venturino A, Rosenbaum E, Caballero De Castro A, Anguiano OL, Gauna L, Fanovich De Schoreder T, Pechen De D’Angelo AM (2003) Biomarkers of effect in toads and frogs. Biomarkers 8:167–186. https://doi.org/10.1080/1354700031000120116
Verma RS, Srivastava N (2003) Effect of chlorpyrifos on thiobarbituric acid reactive substances, scavenging enzymes and glutathione in rat tissues. Indian J Biochem Biophys 40:423–428
Wang K, Zhao Z, Ji W (2019) Bisphenol A induces apoptosis, oxidative stress and inflammatory response in colon and liver of mice in a mitochondria-dependent manner. Biomed Pharmacother 117:1–8. https://doi.org/10.1016/j.biopha.2019.109182
Wu M, Xu H, Shen Y, Qiu W, Yang M (2011a) Oxidative stress in zebrafish embryos induced by short-term exposure to bisphenol a, nonylphenol, and their mixture. Environ Toxicol Chem 30:2335–2341. https://doi.org/10.1002/etc.634
Wu M, Xu H, Yang M, Xu G (2011b) Effects of chronic bisphenol A exposure on hepatic antioxidant parameters in medaka (Oryzias latipes). Toxicol Environ Chem 93:270–278. https://doi.org/10.1080/02772248.2010.530136
Xu EGB, Morton B, Lee JHW, Leung KMY (2015) Environmental fate and ecological risks of nonylphenols and bisphenol A in the Cape D’Aguilar Marine Reserve, Hong Kong. Mar Pollut Bull 91:128–138. https://doi.org/10.1016/j.marpolbul.2014.12.017
Yoloğlu E (2019) Assessment of Na+/K+-ATPase, Mg2+-ATPase, Ca2+-ATPase, and total ATPase activities in gills of freshwater mussels exposed to penconazole. Comm J Biol 3:88–92. https://doi.org/10.31594/commagene.632082
Zhang J, Shen H, Wang X, Wu J, Xue Y (2004) Effects of chronic exposure of 2,4-dichlorophenol on the antioxidant system in liver of freshwater fish Carassius auratus. Chemosphere 55:167–174. https://doi.org/10.1016/j.chemosphere.2003.10.048
Zhang Y, Mi K, Xue W, Wei W, Yang H (2020) Acute BPA exposure-induced oxidative stress, depressed immune genes expression and damage of hepatopancreas in red swamp crayfish Procambarus clarkii. Fish Shellfish Immunol 103:95–102. https://doi.org/10.1016/j.fsi.2020.04.032
Zhou J, Zhu XS, Cai ZH (2011) The impacts of bisphenol A (BPA) on abalone (Haliotis diversicolor supertexta) embryonic development. Chemosphere 82:443–450. https://doi.org/10.1016/j.chemosphere.2010.09.056
Author information
Authors and Affiliations
Contributions
Miraç Uçkun: resources, visualization, methodology, ınvestigation, data curation, and writing (reviewing and editing).
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This study did not use any kind of human participants or human data, which would require any kind of approval. The care and use of animals is conducted in accordance with all relevant international, state, and/or institutional guidelines. Both methods used in animal studies are compliant with the ethical principles of the organization or practice where the research was conducted.
Consent for publication
This study did not use any kind of individual data, such as videos or images.
Competing interests
The author declares no competing interests.
Additional information
Responsible Editor: Bruno Nunes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Uçkun, M. Assessing the toxic effects of bisphenol A in consumed crayfish Astacus leptodactylus using multi biochemical markers. Environ Sci Pollut Res 29, 25194–25208 (2022). https://doi.org/10.1007/s11356-021-17701-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-021-17701-1