
Abstract
The recovery of microbial community and activities is crucial to the remediation of contaminated soils. Distance-dependent variations of microbial community composition and metabolic characteristics in the rhizospheric soil of hyperaccumulator during phytoextraction are poorly understood. A 12-month phytoextraction experiment with Sedum alfredii in a Cd-contaminated soil was conducted. A pre-stratified rhizobox was used for separating sub-layer rhizospheric (0–2, 2–4, 4–6, 6–8, 8–10 mm from the root mat)/bulk soils. Soil microbial structure and function were analyzed by phospholipid fatty acid (PLFA) and MicroResp™ methods. The concentrations of total and specified PLFA biomarkers and the utilization rates for the 14 substrates (organic carbon) in the 0–2-mm sub-layer rhizospheric soil were significantly increased, as well as decreased with the increase in the distance from the root mat. Microbial structure measured by the ratios of different groups of PLFAs such as fungal/bacterial, monounsaturated/saturated, ratios of Gram-positive to Gram-negative (GP/GN) bacterial, and cyclopropyl/monoenoic precursors and 19:0 cyclo/18:1ω7c were significantly changed in the 0–2-mm soil. The PLFA contents and substrate utilization rates were negatively correlated with pH and total, acid-soluble, and reducible fractions of Cd, while positively correlated with labile carbon. The dynamics of microbial community were likely due to root exudates and Cd uptake by S. alfredii. This study revealed the stimulations and gradient changes of rhizosphere microbial community through phytoextraction, as reduced Cd concentration, pH, and increased labile carbons are due to the microbial community responses.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Large quantities of cadmium (Cd) have been introduced into soils by both anthropogenic and geogenic sources (He et al. 2015). From 2005 to 2013, the nationwide soil survey was conducted by the Environmental Protection Ministry of China. The survey results showed that 7% of the investigated soil samples have been contaminated by Cd according to the soil standards of China (Sun et al. 2016). Cd is potentially hazardous due to its high toxicity and lack of biodegradability (Robson et al. 2014; Liao et al. 2015), which can easily transfer to rice and become a serious threat to human health through food chain (Aghababaei et al. 2014; Sousa et al. 2014). Thus, remediation of the Cd-contaminated soils is of great urgency to ensure soil quality and food security as well as to human health.
Conventional physical and chemical methods of removing Cd from soils are costly and often have negative impacts on soil environment (Ali et al. 2013; Ghnaya et al. 2015). Phytoextraction, the utilization of plants to uptake heavy metal from the soils, is regarded as a cost-effective and green way for the remediation of Cd-contaminated soil (Davari et al. 2015; Muehe et al. 2015). In the past few years, there has been a growing interest on studying the effectiveness of metal phytoextraction. It must be noted that the ultimate goal of any soil remediation process should not only be the removal of metals but also the restoration of the soil quality (Hernandez-Allica et al. 2006). As to this, soil microbial characteristics are considered as the most ecologically relevant indicators of soil quality and have been frequently used to evaluate the ecological status of soil under metal pollution (Stefanowicz et al. 2010; Gómez-Sagasti et al. 2012; Burges et al. 2015). Considering the important role of soil microorganisms played in nutrient cycling or availability, the recovery of microbial communities is vital for the sustainability of a soil (Kohler et al. 2016). Consequently, assessing the improvements of soil microbial properties, such as the structure and function of microbial community, is often carried out after the implementation of phytomanagement technologies (Perez-de-Mora et al. 2006; Lu et al. 2015; Kohler et al. 2016). However, to our understanding, the mechanisms of plant rhizosphere regulating soil microbial community structure and function during soil restoration, such as phytoextraction processes, are not well understood. Previous studies have demonstrated that microbial community composition and function can be greatly influenced by soil chemical status such as pH, organic matter, or pollutants (Breulmann et al. 2012; Chodak et al. 2013; Gartzia-Bengoetxea et al. 2016). Relatively few researches have been taken to explore the relationships between microbiological properties and chemical properties in the rhizosphere of hyperaccumulator during phytoextraction. A better understanding of the factors affecting rhizosphere microbial processes of hyperaccumulators is vital for the potential use of phytoextraction and may give insights into prospective ways to accelerate the soil restoration process.
Rhizosphere was first defined as the volume of soil influenced by roots or their exudates, which is commonly separated by gently shaking the roots (He et al. 2005; Mandal et al. 2014). However, functionally, rhizosphere should not be limited to specific volumes but rather characterized by gradients of soil characteristics, from the root surface to the surrounding soils (Aviani et al. 2006). Distance-dependent changes of root activities might lead to the distance-dependent varieties of root-microbe-soil interrelations (He et al. 2009). Phytoextraction takes advantage of the stimulating effect of plants on microbes in the rhizosphere (Yergeau et al. 2014; Columbus and Macfie 2015; Cui et al. 2016). Although much work has been conducted to evaluate the rhizosphere characteristics of hyperaccumulators in phytoextraction, the question of how far these rhizosphere effects on microbial composition and function may extend in the rhizosphere of hyperaccumulators during phytoextraction has not been determined. In an attempt to study the gradients of rhizosphere soil, a pre-stratified rhizobox has been designed which soil in close to roots and then separated by millimeter increments can be get intact in our previous research (Yang et al. 2013a).
The ancient silver mining (ASM) ecotype of Sedum alfredii Hance natively growing in Zhejiang, China, has been conformed as a Cd hyperaccumulator (Yang et al. 2013a, 2014). Using the pre-stratified rhizobox, a 12-month phytoextraction experiment with S. alfredii in a Cd-contaminated soil was conducted. We hypothesized that soil microbial community structure and function of rhizospheric soils with different distances from the root mat might be significantly different due to phytoextraction process, and the main variability might be driven by differences in chemical characteristics such as organic C, pH, and available Cd concentrations. Thus, the objectives were to (1) study the effects of S. alfredii growth on soil Cd speciation (Community Bureau of Reference (BCR)’s sequential extraction procedure), pH, and labile organic C; (2) explore the gradient changes of microbial community structure and metabolic activity in the rhizosphere by phospholipid fatty acid (PLFA) and MicroResp™ analysis, and (3) determine the relationships between microbial community structure or substrate utilization rate and chemical properties (pH, labile carbon, and Cd concentrations). In addition, the potential of S. alfredii growth in the restoration of microbiological properties of metal-contaminated soils was evaluated.
Materials and methods
Soil and plant
The tested soil was collected from a paddy field at the depth of 0–15 cm. Roots or the visible debris were removed; then, the soil was air-dried, ground, and sieved through less than 1 mm. The basic characteristics of the studied soil were pH 5.9, organic matter content 2.90%, total nitrogen 0.18%, total phosphorus 0.11%, available nitrogen 95.3 mg kg−1, and available phosphorus 33.4 mg kg−1. The initial Cd concentration of the studied soil was about 0.8 mg kg−1. In order to get a heavily contaminated soil, the soil was incubated with 15.0 mg Cd kg−1 soil added as CdCl2 for 2 months. After the 2-month incubation, the soil has a final concentration of 15.8 mg Cd kg−1 soil. The ASM ecotype of S. alfredii was collected from an abandoned silver mining area in Quzhou. The results of our previous investigation showed that the Cd concentrations of the collecting site ranged from 5.7 to 84.4 mg kg−1 (Yang et al. 2014). The shoot cuttings were pre-cultured in nutrient solutions under greenhouse conditions.
Rhizobox experiment and sampling
A pre-stratified rhizobox was used in our study, and the details of which have been described previously (Yang et al. 2013a). The soils in the lower zone of the rhizobox were divided into six sub-layers 0–2, 2–4, 4–6, 6–8, 8–10, and >10 mm (bulk soil) from the root mat by the pre-stratified nylon net (Fig. S1, Electronic Supplementary Material).
Four uniform S. alfredii seedlings were transplanted in one rhizobox. The experiment was carried out in a greenhouse, whose temperature and humidity were maintained at 20/25 °C night/day and 70%, respectively. Soil water content was adjusted to 70% of WHC by weighing the rhizobox and adding water from the bottom of rhizobox. The experiment was conducted with three replicates. After a 12-month growth, the samplings of plants and soils were carried out. The harvests involving the sequential removal of S. alfredii from the upper zone and sub-layer rhizospheric soils (from 0- to 10-mm distances of the root mat) and the bulk soil (>10 mm) were sampled. Then, one portion of the fresh soils was stored at 4 °C, and the MicroResp analysis was conducted within 1 week after sampling. One portion was freeze-dried for PLFA analysis. The other portions of soils were air-dried for analysis of chemical properties.
Soil chemical property analysis
Soil pH was measured in a 1:2.5 (w/v) soil solution after shaking for 1 h by a pH electrode. Soil labile organic carbon (C) or easily oxidizable carbon was measured according to Yuan (1963). Briefly, 0.5-g soil was oxidized by 66.7 mmol L−1 K2Cr2O7 at 130–140 °C. After the reaction, the excess dichromate was determined by titrating against 0.2 mol L−1 FeSO4. Soil Cd was fractionated with the modified BCR sequential extraction procedure (Pueyo et al. 2008). This procedure can be described as follows: step 1 (BI, exchangeable, and weak acid-soluble fraction): 1-g soil was extracted with 40 mL 0.11 mol L−1 acetic acid (v/w ratio = 40:1) by shaking in a shaker for 16 h; step 2 (BII, reducible fraction): the residue from the step 1 was extracted with 40 mL 0.5 mol L−1 hydroxylammonium chloride (v/w ratio = 40:1) by shaking in a shaker for 16 h; step 3 (BIII, oxidizable fraction): the residue from the step 2 was digested with H2O2 at 85 °C followed by extraction with 50 mL 1 mol L−1 ammonium acetate adjusted to pH 2.0 by shaking in a shaker for 16 h; step 4 (BIV, residual fraction): the residue from step 3 was digested with HNO3-HClO4-HF. The total Cd concentration was calculated as the sum of the previous four steps of Cd concentrations. Cd concentrations were determined by the atomic adsorption spectrophotometer (AAS).
PLFA analysis
The PLFAs were extracted, fractionated, and methylated as described by Wu et al. (2009). Briefly, the freeze-dried soils were extracted with a mixture of chloroform/methanol/citrate (1:2:0.8, v/v/v) buffer on a shaker for 2 h. Then, the obtained lipids were fractionated on silica-bonded phase columns (SPE-Si, Supelco, Poole, UK) into neutral lipids, glycolipids, and phospholipids. The phospholipid fractions were methylated by mild alkaline hydrolysis. The fatty acid methyl esters were identified by GC (Agilent N6890, USA) fitted with MIDI peak identification software (Version 4.5; MIDI Inc., Newark, DE). Methyl nonadecanoate fatty acid (19:0) was added as an internal standard. For each sample, the abundance of individual fatty acid methyl esters was expressed as nanomole PLFAs per gram soil. Individual PLFA markers were used to quantify the relative abundance of specific microbial groups. Bacterial biomass was represented by the PLFAs i15:0, a15:0, 15:0, i16:0, 16:1ω7c, 16:1ω9c, i17:0, a17:0, cy17:0, 17:0, 18:1ω7c, and cy19:0 (Frostegård and Bååth 1996). Biomarkers i15:0, a15:0, i16:0, i17:0, and a17:0 were used as markers for Gram-positive bacterial biomass (Zelles 1999). Gram-negative bacterial biomass was determined by using the following PLFAs: 16:1ω7c, 16:1ω9c, cy17:0, 18:1ω7c, and cy19:0 (Zogg et al. 1997). Fungal biomass was quantified as a sum of the PLFAs 18:2ω6,9c and 18:1ω9c (Myers et al. 2001). Arbuscular mycorrhizal (AM) fungal biomass was determined according to the PLFA 16:1ω5c (Olsson et al. 1995). Actinomycete biomass was represented by the PLFAs 16:0 (10Me), 17:0 (10Me), and 18:0 (10Me) (Turpeinen et al. 2004). Protozoan biomass was estimated on the basis of the PLFA 20:4ω6c (Myers et al. 2001).
Community-level physiological profile (MicroResp™)
Soil microbial community-level physiological profiles were determined by the MicroResp™ system (Macaulay Scientific Consulting Ltd., Aberdeen, UK). This approach uses a colorimetric detection to measure soil respiration in the presence of different carbon sources (Campbell et al. 2003). Fifteen carbon compounds such as carbohydrates (d-glucose, d-fructose, d-galactose, l-arabinose, and N-acetyl-glucosamine), amino acids (l-alanine, arginine, l-lysine, γ-amino butyric acid, and l-cysteine HCl), and carboxylic acids (citric acid, malic acid, oxalic acid, lactic acid, and protocatechuic acid) were used. The substrates used were selected on the basis of carbon sources that are ecologically relevant to soil especially those produced as root exudates (Campbell et al. 1997). Basal respiration rate was measured in wells with only water added. For each soil, three replicates were employed. The plate was read at 590 nm by using a microplate reader before and after 6 h of incubation at 25 °C. The absorbances were normalized for differences recorded before and after incubation and then converted to % CO2 by the calibration curve y = A + B/(1 + D × Ai), where A = −0.2265, B = −1.606, and D = −6.771. The % CO2 was converted to microgram CO2-C gram per hour using headspace volume, soil weight, and incubation time.
Statistical analysis
The data were expressed as mean (n = 3) with standard errors. One-way analysis of variance (ANOVA) was employed to determine the statistical differences of the values among soil samples from different distances of the root mat using SPSS 19.0 (SPSS Inc., USA). The principal component analysis (PCA) of soil PLFAs and MicroResp™ was performed using SPSS 19.0. Redundancy analysis (RDA) using Canoco for Windows 4.5 was used to determine (1) the relationships between soil microbial community structure and soil chemical properties (labile organic C, pH, and different forms of Cd concentrations); for this purpose, the concentrations of total and different groups (bacterial, Gram-positive bacterial, Gram-negative bacterial, actinomycete, fungal, AM fungal, and protozoa) of PLFAs and the microbial indicator ratios (fungal/bacterial, ratios of Gram-positive to Gram-negative (GP/GN) bacteria, cyclopropyl PLFAs to their monoenoic precursors, 19:0 cyclo/18:1ω7c, monounsaturated/saturated PLFAs) were used as the input data and (2) the relationships between soil substrate utilization rates and soil chemical properties (labile organic C, pH, and different forms of Cd concentrations). Correlation analysis between soil PLFA properties and substrate utilization rates and soil chemical properties using Pearson’s correlation coefficients by SPSS 19.0 were determined.
Results
Effects of S. alfredii growth on soil chemical properties
The pH value of 0–2-mm sub-layer rhizospheric soil was significantly (P < 0.05) decreased than those of 2–10-mm sub-layer rhizospheric and the bulk soils (Table 1). And the pH values of 2–4- and 4–6-mm sub-layer rhizospheric soils were also significantly (P < 0.05) decreased compared to those of 6–10-mm sub-layer rhizospheric and bulk soils.
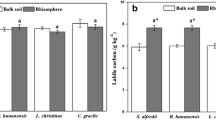
The labile organic C content of 0–2-mm sub-layer rhizospheric soil was significantly (P < 0.05) higher than those of 2–10-mm sub-layers and bulk soils (Table 1). The labile organic C contents of the 2–4-, 4–6-, and 6–8-mm sub-layer rhizospheric soils were also significantly (P < 0.05) increased compared to the bulk soil (Table 1).
The Cd concentrations of the total, BI, and BII fractions in the 0–2-mm sub-layer rhizospheric soil were significantly (P < 0.05) decreased compared to those of other sub-layers’ rhizospheric and bulk soils (Table 1). And Cd concentrations of the total, BI, and BII fractions in the 2–4- and 4–6-mm soils were significantly (P < 0.05) lower than those in the 6–8- and 8–10-mm sub-layers and the bulk soil (Table 1).
Effects of S. alfredii growth on soil microbial community structure (PLFA)
The number of individual PLFAs identified in sub-layer rhizospheric soils ranged from 41 to 60, while only 39 was detected in the bulk soil (Table S1, Electronic Supplementary Material). Compared to the bulk soil, some PLFAs were only found in the 0–2-mm sub-layer rhizospheric soils such as saturated fatty acids: 10:00, i11:0, 11:00, a13:0, a16:0, and i19:0; hydroxy group fatty acids: 11:0 3OH, 12:0 2OH, 14:0 2OH, and 16:0 2OH; and unsaturated fatty acids: 15:1ω6c, 16:1ω11c, 18:3ω6c, and 19:1ω9,11c. The concentrations of total microbes, total bacterial, Gram-positive and Gram-negative bacterial, actinomycete, fungal, AM fungal, and protozoan PLFAs in the 0–2-mm sub-layer rhizospheric soils were significantly (P < 0.05) enhanced compared to those in 2–10-mm rhizospheric and the bulk soils (Fig. 1). The actinomycete, AM fungal, and protozoan PLFAs in the 2–4-mm sub-layer rhizospheric soils were significantly (P < 0.05) higher than those in the bulk soil (Fig. 1).

Concentrations of total, bacterial, Gram-positive (GP) bacterial, Gram-negative (GN) bacterial, actinomycete, fungal, AM fungal, and protozoa PLFAs of the sub-layer rhizospheric (0–2, 2–4, 4–6, 6–8, 8–10 mm) and bulk (>10 mm) soils. Bars represent standard errors. Different letters on the error bars indicate significant differences at P < 0.05
The ratios of the taxonomic groupings were calculated (Fig. 2). The ratio of fungal-to-bacterial (F/B) PLFAs and monounsaturated to saturated (M/S) PLFAs were significantly (P < 0.05) increased in the 0–2-mm sub-layer rhizospheric soil than those in the 4–10-mm sub-layer rhizospheric and the bulk soils (Fig. 2). The F/B of 2–4-mm sub-layer rhizospheric soil was also significantly (P < 0.05) increased than that in the bulk soil. The GP/GN in the 0–2-mm sub-layer rhizospheric soil was significantly (P < 0.05) decreased than those in 6–10-mm sub-layer rhizospheric and the bulk soils (Fig. 2). Cyclopropyl PLFAs to their monoenoic precursors (cy/pre) and 19:0 cyclo/18:1ω7c in the 0–2-mm sub-layer rhizospheric soil were significantly (P < 0.05) lower than those in 4–10-mm sub-layer rhizospheric and the bulk soils. The cy/pre and 19:0 cyclo/18:1ω7c in the 2–4-mm sub-layer rhizospheric soil were significantly (P < 0.05) lower than those of bulk soil (Fig. 2).

Fungal/bacterial PLFAs (F/B), GP/GN bacterial PLFAs (GP/GN), cyclopropyl PLFAs to their monoenoic precursors (cy/pre), 19:0 cyclo/18:1ω7c, and monounsaturated/saturated PLFAs (M/S) of the sub-layer rhizospheric (0–2, 2–4, 4–6, 6–8, 8–10 mm) and bulk (>10 mm) soils. Bars represent standard errors. Different letters on the error bars indicate significant differences at P < 0.05
The effects of the phytoextraction process on PLFA composition were determined by PCA. As shown in Fig. 3, the first principal component accounted for 46.91%, and the second component for 16.4% of the total variation in PLFA profiles. The results of PCA suggested that the effects of plant growth resulted in distance dynamics of microbial community composition. The PCA results showed that the 0–2-mm sub-layer rhizospheric soil was separated clearly from the 2–10-mm sub-layer rhizospheric and the bulk soil by positive PC1 values (Fig. 3a). The 2–4-mm sub-layer rhizospheric soil was separated clearly from the 0–2- and 4–10-mm sub-layer rhizospheric soils and the bulk soil by positive PC2 values (Fig. 3a). In Fig. 3a, there appeared to be little separation between 4– 6-, 6–8-, and 8–10-mm sub-layer rhizospheric soils and the bulk soil. PCA of individual PLFAs (Fig. 3b) showed that most of the individual PLFAs laid to the right of the plot, such as the fungal biomarkers (18:2ω6,9c and 18:1ω9c), bacterial biomarkers (i16:0, i17:0,18:1ω7c, cy17:0, a17:0, etc.), Gram-negative bacterial biomarker (16:1ω7c, cy17:0), Gram-positive bacterial biomarker (i15:0, a15:0, i16:0, i17:0, a17:0), AM fungal biomarker (16:1ω5c), and some hydroxy group fatty acids (11:0 3OH, i11:0 3OH). These PLFA biomarkers all had highly positive loading scores for PC1. This indicated that these different groups of soil microorganisms were stimulated in 0–2-mm sub-layer rhizospheric soil due to the growth of S. alfredii. One of the actinobacteria PLFA biomarkers (10Me18:0) and some hydroxy group fatty acids (12:0 3OH, i15:0 3OH, and i13: 03OH) laid closely to the center of the plot. Only few fatty acids (14:0 3OH, 14:0 2OH, 9:0, 11:0 and i18:0) had negative loading values for PC1.

Principal component analysis (PCA) of PLFA patterns. a Results present each soil sample. The sub-layer rhizospheric soils from different distances of the root surface were represented by 0–2, 2–4, 4–6, 6–8, and 8–10. Bulk is the bulk soil (>10 mm). b The loadings of the individual PLFA for PC1 and PC2
Influence of S. alfredii growth on soil microbial metabolic function (MicroResp™)
As shown in Fig. 4, the highest induced respiration rate was observed after applications of carbohydrates such as d-glucose and d-fructose and so was the lowest as arginine added. The C substrate utilization rates for all the tested substrates (except arginine) of the 0–2-mm sub-layer rhizospheric soil were significantly (P < 0.05) enhanced than those of the bulk soil. The utilization rates for d-glucose, l-arabinose, and l-alanine in the 2–4-mm sub-layer rhizospheric soils were also significantly (P < 0.05) higher compared to those in the bulk soil (Fig. 4).

Substrate-induced respiration responses for 15 C sources and basal respiration in the sub-layer rhizospheric (0–2, 2–4, 4–6, 6–8, 8–10 mm) and bulk (>10 mm) soils. Bars represent standard errors (SE). Different letters on the error bars indicate significant differences at P < 0.05
The results of PCA based on the MicroResp™ data showed that phytoextraction also resulted in distance dynamics of microbial carbon utilization rates. As shown in Fig. 5, the first principal component accounted for 79.19% and the second component for 6.69% of the total variation in soil carbon utilization rate profiles. In Fig. 5a, the 0–2-mm sub-layer rhizospheric soil was separated clearly from the 2–10-mm sub-layer rhizospheric and bulk soils along the first component. The 2–4-, 4–6-, 6–8-, and 8–10-nm sub-layer rhizospheric soils and the bulk soil were not clearly separated. In the PCA of C individual carbon sources, all the 15 carbon sources (except arginine) and water have highly positive values for PC1 (Fig. 5b). This indicated that the carbon utilization rates for the carbon sources and water of the 0-2-mm sub-layer rhizospheric soil could be stimulated by S. alfredii growth.

Principal component analysis (PCA) of MicroResp carbon utilization profiles. a Results present each soil sample. The sub-layer rhizospheric soils from different distances of the root surface were represented by 0–2, 2–4, 4–6, 6–8, and 8–10. Bulk is the bulk soil (>10 mm). b The loadings of the individual 15 carbon sources and water for PC1 and PC2
Relationships between soil PLFAs or metabolic profiles and chemical properties
To examine the relationships between PLFAs and soil chemical properties, redundancy analysis (RDA) was performed (Fig. 6). Eigenvalues of RDA showed that axes 1 and 2 explained 82.6 and 0.6% of the variance of the PLFA data and relationships between PLFA-chemical properties, indicating that soil microbial PLFAs were strongly correlated with soil chemical parameters. Soil labile carbon; pH; and total, BI, and BII fractions of Cd were significant (P < 0.05) for explaining the variations of PLFAs. The total, bacteria, fungi, actinomycete, AM fungal, protozoa PLFAs, M/S, and F/B were significantly (P < 0.05) and negatively correlated with pH and total, BI, and BII fractions of Cd while significantly (P < 0.05) positively correlated with labile C (Table S2, Electronic Supplementary Material).

Biplot of redundancy analysis of the relationships between PLFA and environmental variables (pH, labile organic carbon, and different fractions of Cd)
As to the metabolic activity, the eigenvalues of RDA showed that RDA 1 and 2 explained 72.4 and 3.8% of the varieties of soil microbial carbon utilization rate profiles (Fig. 7). Soil labile carbon; pH; and total, BI, and BII fractions of Cd were significant (P < 0.05) for explaining the variation of MicroResp data. Substrate utilization rates for the carbon sources were significantly (P < 0.05) negatively correlated with pH and total, BI, and BII fractions of Cd (except arginine and protocatechuic acid) and significantly (P < 0.05) positively correlated with labile carbon (Table S3, Electronic Supplementary Material).

Biplot of redundancy analysis of the relationships between carbon source utilization profiles and environmental variables (pH, labile organic carbon, and different fractions of Cd)
Discussion
Effects of S. alfredii growth on soil chemical characteristics
Rhizosphere characteristics play an important role in metal accumulation of hyperaccumulators, particularly pH and available organic C (Kwonrae et al. 2010; Alford et al. 2010). A decrease in pH values was found in our study. Hyperaccumulators could enhance metal solubility via exudation of organic acids (Fitz and Wenzel 2002). As to Cd hyperaccumulation, the increase of Cd uptake because of the decrease of rhizosphere soil pH was reported (McGrath et al. 1997; Wei and Twardowska 2013). However, the controversy existed in the literature about the effects of hyperaccumulator growth on rhizosphere pH. For instance, Kwonrae et al. (2010) reported an increased pH in the rhizosphere of Indian mustard in the phytoremediation process. Labile organic C produced by degradation of roots or exudation of organic compounds may influence the solubility of metals due to formation of metal-C complexes (Otero et al. 2012). Similar to our study, the increasing of labile C in the rhizosphere of hyperaccumulators was reported by previous studies (Kim et al. 2010; Wei and Twardowska 2013). Thus, in this study, rhizosphere acidification and the increased labile organic C might be important factors influencing the Cd bioavailability in soil or uptake by S. alfredii.
The metals extracted by the BCR method of different steps represent different degrees of bioavailability (Anju and Banerjee 2010). In this procedure, the first step is considered as the water soluble, exchangeable, and associated with carbonate, the second as Mn/Fe oxide bound, the third as organic matter bound, and the fourth as metal in the mineral matrix (Sungur et al. 2014). In this study, after phytoextraction by S. alfredii, the Cd concentrations of first and second steps had decreased greatly. The metals extracted at the first step were considered as the most available forms (Chlopecka 1996). The decrease of these forms of Cd in the rhizosphere of hyperaccumulators was frequently reported, such as Thlaspi caerulescens (Puschenreiter et al. 2003; Dessureault-Rompre et al. 2010) and Sedum plumbizincicola (Liu et al. 2011; Li et al. 2014). In rhizospheric soil, the concentrations of metals rely on the balance between those released from soil and its uptake by plants. The results of our study indicated that the rate of Cd uptake by S. alfredii was higher than the activating rate of Cd from rhizospheric soil. The reducible fraction (Mn/Fe oxide bound) is reported to be not easily available for plant uptake (Wang et al. 2009; Chen et al. 2014). However, in this study, the reducible fraction of Cd was significantly reduced after phytoextraction. These results indicated that the exchangeable, water-soluble, carbonate, and Fe–Mn oxide-bound fraction of Cd were the main portions for the uptake by S. alfredii during phytoextraction.
Effects of S. alfredii growth on soil microbial community structure
The analysis of PLFAs provides quantitative data on soil microbial community composition. This approach has been widely used to determine the shifts in the structure of microbial community response to metal stress (Chodak et al. 2013; Mackie et al. 2015; Zornoza et al. 2016). The total PLFA concentration has been used as a measure of soil microbial biomass (Zelles 1999). In this study, the total and all specified groups of PLFA concentrations clearly increased in the sub-layer rhizospheric soils compared to the bulk soil. These were consistent with previous studies which showed that plant growth could significantly increase soil PLFAs (Hortal et al. 2013; Yoshitake et al. 2013; Qin et al. 2014). Compared to the unplanted soil, the presence of surfaces for soil microbial survival and the organic matters released by roots may lead to higher microbial populations in the planted soils (Cui et al. 2016). It has been frequently reported that root exudation stimulated the growth of microorganisms in the rhizosphere soil (Qin et al. 2014; Ridl et al. 2016). In our study, the extent of the rhizosphere effects of phytoextraction by S. alfredii on microbial community pattern could be shown. The PLFA concentrations decreased as the distances from the root surface increased and the 0–2- and 2–4-mm sub-layer rhizospheric soils were significantly influenced by S. alfredii growth. PCA also showed that 0–2- and 2–4-mm soils were separated from 4 to 10 mm and bulk soils. In our previous research, it was found that soil microbial biomass carbon was changed as the distances from the root mat varied (Yang et al. 2013a). Liu et al. (2012) reported that the microbial properties in the soils at 0–4-mm distance from the root mats were significantly different from the non-rhizosphere soil. Soil organic carbon sources excreted by roots were reported to be vital in determining the rhizosphere soil microbial compositions in vegetation process (Tscherko et al. 2004). However, the stimulation of soil microorganisms by roots is distance-dependent, owing to the gradual disappearance of root exudates due to radial dilution and microbial consumption. What is more, phytoextraction by S. alfredii significantly reduced the available and total Cd of the 0–2- and 2–4-mm soils, which indicated that the lower concentrations of Cd might also promote soil microbial growth. This is supported by negative relationships between soil PLFAs and Cd concentrations.
The ratios of different groups of microbial PLFAs have been used to detect soil microbial community shifts. And these ratios could indicate whether specific microbial groups are favored by the changes of soil conditions (Mitchell et al. 2015). The ratio of F/B or M/S has been used to indicate environmental change and function of the soil microbial communities (Hinojosa et al. 2005; Strickland and Rousk 2010). Excessive toxic metals in the soils were reported to exert a reduction of F/B and M/S (Pennanen et al. 1996; Hinojosa et al. 2005; Carrasco et al. 2010). The higher value of F/B or M/S is considered to be indicative for a more sustainable ecosystem and has positive effects on nutrient uptake and retention (Bossio and Scow 1998; Azarbad et al. 2016). In this study, the phytoextraction by S. alfredii significantly increased these ratios in the 0–2 (F/B and M/S) and 2–4 mm (F/B) sub-layer rhizospheric soils, indicating that phytoextraction alleviates Cd stress on microbial community. In addition, the higher F/B in rhizosphere might be related to the input of labile carbon, and the results of correlation analysis supported this. The increasing of organic carbon is usually followed by an increasing of fungi biomass. Zornoza et al. (2015) also reported a significant correlation between soil fungal biomass and organic carbon content.
The cy/pre, 19:0 cyclo/18:1ω7c, and GP/GN ratios of the PLFAs have been also commonly used as indicators of environmental stress to the microbial community, such as contamination, salt, or nutritional stress (Pennanen et al. 1996; Ying et al. 2013; Azarbad et al. 2016). The increase of cyclopropyl fatty acids (such as cy19:0) is an adaptation mechanism of microorganisms that is induced by metal toxicity (Frostegård et al. 1993). In our study, the significantly reduced cy/pre and 19:0 cyclo/18:1ω7c in the 0–2- and 2–4-mm soils indicate low Cd toxicity due to phytoextraction. This is supported by the positive relationships between these ratios and Cd concentrations (Fig. 6). These were in agreement with the finding by Carrasco et al. (2010) which showed that cyc/prec ratio was positively correlated to the soluble contents of heavy metals. Garcia-Sánchez et al. (2015) also reported that cy/pre ratio was increased under metal stress. In addition, high concentration of labile carbon from root exudates might also contribute to this. For example, Qin et al. (2014) showed that cy/pre was decreased in the rhizosphere soils of Cucurbita species compared to the unplanted soils. In our study, phytoextraction by S. alfredii decreased the ratio of GP/GN. This result suggested that the remediation process favored GN bacteria growth more than the GP bacteria. Gram-negative bacteria tend to grow well under substrate-rich conditions and adapt quickly to different environmental conditions (Chang et al. 2016). Garcia-Sánchez et al. (2015) found that the ratio of GP/GN was significantly increased in metal-contaminated soil, which indicated that GP/GN might be associated with metal stress. The results of RDA analysis in our study also support this. The low ratio of GP/GN was associated with the high content of organic C in the rhizospheric soils. The decrease of GP/GN in the rhizosphere due to phytoremediation was also reported in a previous study (Wei et al. 2014).
Effects of S. alfredii growth on soil microbial metabolic functions
The ability to utilize variety of carbons reflects the activity of mineralization and functioning of a soil (Campbell et al. 2008; Yao et al. 2011; Ferris and Tuomisto 2015). The MicroResp™ developed by Campbell et al. (2003) is a method to study the carbon substrate utilization, which offers a rapid and sensitive method for the determination of differences in microbial community function (Creamer et al. 2016; Chapman et al. 2007; Burton et al. 2010). MicroResp™ has been frequently used to determine the effects of contaminants such as heavy metals on soil catabolic functions (Bérard et al. 2014; Kenarova and Boteva 2015). In our study, microbial catabolic activity measured by MicroResp™ was strengthened in the sub-layer rhizospheric soils due to the growth of S. alfredii, which was consistent with some previous studies. For example, Stefanowicz et al. (2012) showed that in metal-polluted soil, plants positively affected soil functional richness and apparently counteracted the toxic effects of metal contamination. Similarly, Epelde et al. (2010) reported that T. caerulescens growth increased the substrate-induced respiration rates in the Zn- and Cd-contaminated soil. These could be partly explained by the reduction of metal concentrations due to plant uptake. Numerous studies have indicated that excessive metals in soil are toxic to microorganisms and the ecological functions performed by them could be seriously inhibited (André et al. 2009; Kenarova et al. 2014). Metals are toxic to microorganisms by replacement of essential metals, damaging cell membrane and cellular function disruption (Epelde et al. 2015). Thus, lower catabolic functions of microorganisms in metal-contaminated soils were well documented (Boshoff et al. 2014; Klimek et al. 2016). In addition, carbon resources are considered as the driving force for microbial density and activity (Burton et al. 2010). Thus, the higher contents of organic carbon may also contribute to the increasing of metabolic activity of rhizosphere soil microorganisms. In this study, the highest level of substrate utilization rates was observed with carbohydrates and the lowest was arginine. The low substrate utilization rates for arginine were also reported by Campbell et al. (2008), since the utilization of arginine requires deaminase activity prior to catabolism. It has been argued that returning the functions to the contaminated soils is more important than returning microbial diversity per se (Setälä et al. 2005).
In this study, the results of pre-stratified rhizobox showed that soil microbial C utilization rates showed trend of decreasing as the distances from the root mat increased. And 0–2-mm soil was the main rhizosphere effects of S. alfredii on microbial metabolic activity. In our previous research, we found that soil microbial activities, as measured by enzyme activities, decreased as the distance from the root mat increased (Yang et al. 2013a). As shown by RDA analysis, labile C, pH, and Cd concentration explain most of the variations in metabolic activity. Thus, the distance-dependent variations of soil properties such as labile C, pH, and Cd concentration may result in the distance-dependent variations in microbial metabolic activity.
Correlations between soil PLFAs or metabolic functions and chemical properties
The pH is considered as one of the most important drivers for microbial community composition (Rousk et al. 2010; Zornoza et al. 2015, 2016). In this study, negative relationships between total or specific groups of PLFA concentrations and pH were found. This result is consistent with the result observed by Ge et al. (2013), which showed that total, bacterial, and fungal PLFAs were significantly and negatively related to pH. Zornoza et al. (2015) found that fungal growth decreased with increasing pH, despite the high availability of metals at low pH. However, the issue regarding the relationship between microbial and pH was controversial. For instance, Chodak et al. (2013) reported that some PLFA biomarkers of Gram-negative bacteria (16:1ω9, 18:1ω9t, cy17:0) were more abundant in soils with higher pH while others (βOH14:0, 18:1ω9t, cy17:0) were more abundant in lower pH. Rousk et al. (2010) showed that the bacteria biomass was positively correlated to soil pH while fungi were unaffected by pH. One possible explanation for the apparent disparity between different studies lies in the different soil environments. Soil microorganisms are usually carbon limited, and it has been reported that the contents of available carbon are vital for soil microbial community when nutrients are limited (Huang et al. 2015). In this study, during S. alfredii growth, the secreting of plant-derived C such as organic acids may benefit microbial population’s rapid growth in the rhizosphere soil. The positive relationship between labile C and PLFA concentrations supported this point. Similarly, a positive relationship between Gram-negative bacterial PLFAs, actinomycete, protozoan PLFAs, and soil labile C which are decomposed from shoot and root litter was reported by Breulmann et al. (2012). Mackie et al. (2015) also reported that soil bacterial PLFAs were positively correlated to extractable organic C. Heavy metals could severely restrain microbial growth, because of their known toxicity to microorganisms at high concentrations (Singh et al. 2014). The negative relationships between PLFAs and Cd concentrations found in our study were consistent with previous studies (Chodak et al. 2013; Yang et al. 2013b). The phytoextraction reduced available Cd, which alleviate the Cd toxicity and provide a better living condition for the soil microorganisms. Thus, the increased PLFAs of the sub-layer rhizospheric soils might be partly explained by decreasing of Cd concentrations caused by the uptake of S. alfredii.
It has been previously reported that plant cover is a key driver of soil microbial community activity (Breulmann et al. 2012). The change of soil microbial catabolic activities in revegetation was reported to be regulated by the dynamics of soil abiotic conditions (Yu et al. 2016). In our study, significant relationships were found among soil microbial carbon utilization rates and soil chemical properties (pH, total and available Cd, labile C) in the rhizosphere of S. alfredii. Soil pH is frequently reported to be a strong factor linked to soil microbial catabolic function in previous studies, such as Zhou et al. (2012) and Gartzia-Bengoetxea et al. (2016). Soil labile C comprises the most available sources for microorganisms, and many studies have revealed that soil microbial metabolic activities are strongly influenced by available carbon substrates (Burton et al. 2010; Tian et al. 2015). Similarly, in our study, it was found that microbial C utilization rates positively correlated with soil labile C content in the rhizosphere. This indicated that labile C secreted by S. alfredii might benefit soil microbiota and stimulate the activity of microbial community. The rhizospheric soils have higher levels of labile organic C, therefore supporting higher levels of microbial activity. Furthermore, negative correlations among soil microbial C utilization rates and Cd concentrations were found in our study, which indicated that the increasing of microbial activities might be also related to the decreasing of soil Cd concentration. High amounts of soil metals could damage proteins or disrupt cell membranes of microorganisms and thus negatively inhibit microbial activity (Leita et al. 1995). Yang et al. (2006) showed that both BI and BII Ni fractions were mostly responsible for shift of the soil microbial community-level physiological profiling. Kenarova et al. (2014) also reported that the carbon utilization rates of microbial communities were correlated significantly negatively with Cu, Zn, and Pb. In our study, the strong relationships between microbial metabolic profiles and total, B1, and B2 fractions of Cd addressed the important role of metal dynamic played during phytoextraction.
Conclusions
Phytoextraction by S. alfredii significantly decreased the exchangeable, water-soluble, carbonate, and Fe–Mn oxide-bound forms of Cd in the 0–6-mm sub-layer rhizospheric soils compared to the bulk soil. The utilization rates for 14 carbon sources of soil microbial community were enhanced, as well as the phytoextraction increased the individual numbers and contents of total and specific groups of PLFAs and changed the structure of soil microbial communities in the sub-layer rhizospheric soils. The rhizosphere effects of S. alfredii growth on microbial community structure and metabolic functions were decreased as the distances from root surface increased, and the 0–2 mm was its mainly influence scope. The PLFA contents and substrate utilization rates were negatively correlated with pH and total, acid-soluble, and reducible fractions of Cd, while positively correlated with labile carbon. In conclusion, phytoextraction by S. alfredii changed the chemical properties and decreased available Cd contents of the rhizospheric soils. Microbial community structure was changed, and metabolic activities were enhanced. The distance dynamics of microbial community in the rhizospheric soil were related to the changes of pH, concentrations of available Cd, and labile carbon.
References
Aghababaei F, Raiesi F, Hosseinpur A (2014) The influence of earthworm and mycorrhizal co-inoculation on Cd speciation in a contaminated soil. Soil Biol Biochem 78:21–295
Alford ÉR, Pilon-Smits EA, Paschke MW (2010) Metallophytes—a view from the rhizosphere. Plant Soil 337:33–50
Ali H, Khan E, Sajad MA (2013) Phytoremediation of heavy metals—concepts and applications. Chemosphere 91:869–881
André A, Antunes SC, Gonçalves F, Pereira R (2009) Bait-lamina assay as a tool to assess the effects of metal contamination in the feeding activity of soil invertebrates within a uranium mine area. Environ Pollut 157:2368–2377
Anju M, Banerjee DK (2010) Comparison of two sequential extraction procedures for heavy metal partitioning in mine tailings. Chemosphere 78:1393–1402
Aviani I, Laor Y, Raviv M (2006) Limitations and potential of in situ rhizobox sampling for assessing microbial activity in fruit tree rhizosphere. Plant Soil 279:327–332
Azarbad H, van Straalen NM, Laskowski R, Nikiel K, Röling WF, Niklińska M (2016) Susceptibility to additional stressors in metal-tolerant soil microbial communities from two pollution gradients. Appl Soil Ecol 98:233–242
Bérard A, Mazzia C, Sappin-Didier V, Capowiez L, Capowiez Y (2014) Use of the MicroResp™ method to assess pollution-induced community tolerance in the context of metal soil contamination. Ecol Indic 40:27–33
Boshoff M, De Jonge M, Dardenne F, Blust R, Bervoets L (2014) The impact of metal pollution on soil faunal and microbial activity in two grassland ecosystems. Environ Res 134:169–180
Bossio DA, Scow KM (1998) Impacts of carbon and flooding on soil microbial communities: phospholipid fatty acid profiles and substrate utilization patterns. Microb Ecol 35:265–278
Breulmann M, Schulz E, Weibhuhn K, Buscot F (2012) Impact of the plant community composition on labile soil organic carbon, soil microbial activity and community structure in semi-natural grassland ecosystems of different productivity. Plant Soil 352:253–265
Burges A, Epelde L, Garbisu C (2015) Impact of repeated single-metal and multi-metal pollution events on soil quality. Chemosphere 120:8–15
Burton J, Chen C, Xu Z, Ghadir H (2010) Soil microbial biomass, activity and community composition in adjacent native and plantation forests of subtropical Australia. J Soil Sediments 10:1267–1277
Campbell CD, Grayston SJ, Hirst DJ (1997) Use of rhizosphere carbon sources in sole carbon source tests to discriminate soil microbial communities. J Microbiol Meth 30:33–41
Campbell CD, Chapman SJ, Cameron CM, Davidson MS, Potts JMA (2003) Rapid microtiter plate method to measure carbon dioxide evolved from carbon substrate amendments so as to determine the physiological profiles of soil microbial communities by using whole soil. Appl Environ Microb 69:3593–3599
Campbell CD, Cameron CM, Bastias BA, Chen CR, Cairney JWG (2008) Long term repeated burning in a wet scleophyll forest reduces fungal and bacterial biomass and responses to carbon substrates. Soil Biol Biochem 40:2246–2252
Carrasco L, Gattinger A, Fließbach A, Roldán A, Schloter M, Caravaca F (2010) Estimation by PLFA of microbial community structure associated with the rhizosphere of and growing in semiarid mine tailings. Microb Ecol 2:265–271
Chang EH, Chen TH, Tian G, Chiu CY (2016) The effect of altitudinal gradient on soil microbial community activity and structure in moso bamboo plantations. Appl Soil Ecol 98:213–220
Chapman SJ, Campbell CD, Artz RRE (2007) Assessing CLPPs using MicroResp: a comparison with Biolog and multi-SIR. J Soil Sediments 7:406–410
Chen Z, Ai Y, Fang C, Wang K, Li W, Liu S, Li C, Xiao J, Huang Z (2014) Distribution and phytoavailability of heavy metal chemical fractions in artificial soil on rock cut slopes alongside railways. J Hazard Mater 273:165–173
Chlopecka A (1996) Assessment of form of Cd, Zn and Pb in contaminated calcareous and gleyed soils in southwest Poland. Sci Total Environ 188:253–262
Chodak M, Gołębiewski M, Morawska-Płoskonka J, Kuduk K, Niklińska M (2013) Diversity of microorganisms from forest soils differently polluted with heavy metals. Appl Soil Ecol 64:7–14
Columbus MP, Macfie SM (2015) It takes an individual plant to raise a community: TRFLP analysis of the rhizosphere microbial community of two pairs of high- and low-metal-accumulating plants. Soil Biol Biochem 81:77–80
Creamer RE, Stone D, Berry P, Kuiper I (2016) Measuring respiration profiles of soil microbial communities across Europe using MicroResp™ method. Appl Soil Ecol 97:36–43
Cui H, Fan Y, Yang J, Xu L, Zhou J, Zhu Z (2016) In situ phytoextraction of copper and cadmium and its biological impacts in acidic soil. Chemosphere 161:233–241
Davari M, Homaee M, Rahnemaie R (2015) An analytical deterministic model for simultaneous phytoremediation of Ni and Cd from contaminated soils. Environ Sci Pollu Res 22:4609–4620
Dessureault-Rompre J, Luster J, Schulin R, Tercier-Waeber ML, Nowack B (2010) Decrease of labile Zn and Cd in the rhizosphere of hyperaccumulating Thlaspi caerulescens with time. Environ Pollut 158:1955–1962
Epelde L, Becerril JM, Kowalchuk GA, Deng Y, Zhou J, Garbisu C (2010) Impact of metal pollution and Thlaspi caerulescens growth on soil microbial communities. Appl Environ Microb 76:7843–7853
Epelde L, Lanzén A, Blanco F, Urich T, Garbisu C (2015) Adaptation of soil microbial community structure and function to chronic metal contamination at an abandoned Pb-Zn mine. FEMS Microbiol Ecol 91:1–11
Fitz WJ, Wenzel WW (2002) Arsenic transformation in the soil–rhizosphere–plant system, fundamentals and potential application of phytoremediation. J Biotechnol 99:259–278
Ferris H, Tuomisto H (2015) Unearthing the role of biological diversity in soil health. Soil Biol Biochem 85:101–109
Frostegård Å, Bååth E (1996) The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil. Biol Fert Soils 22:59–65
Frostegård Å, Tunlid A, Bååth E (1993) Phospholipid fatty acid composition, biomass and activity of microbial communities from two soil types experimentally exposed to different heavy metal. Appl Environ Microbiol 59:3605–3617
Garcia-Sánchez M, Garcia-Romera I, Cajthaml T, Tlustoš P, Száková J (2015) Changes in soil microbial community functionality and structure in a metal-polluted site: the effect of digestate and fly ash applications. J Environ Manag 162:63–73
Gartzia-Bengoetxea N, Kandeler E, de Arano IM, Arias-González A (2016) Soil microbial functional activity is governed by a combination of tree species composition and soil properties in temperate forests. Appl Soil Ecol 100:57–64
Ge T, Chen X, Yuan H, Li B, Zhu H, Peng P, Li K, Jones DL, Wu J (2013) Microbial biomass, activity, and community structure in horticultural soils under conventional and organic management strategies. Eur J Soil Biol 58:122–128
Ghnaya T, Mnassri M, Ghabriche R, Wali M, Poschenrieder C, Lutts S, Abdelly C (2015) Nodulation by Sinorhizobium meliloti originated from a mining soil alleviates Cd toxicity and increases Cd-phytoextraction in Medicago sativa L. Front Plant Sci 6:863
Gómez-Sagasti MT, Alkorta I, Becerril JM, Epelde L, Anza M, Garbisu C (2012) Microbial monitoring of the recovery of soil quality during heavy metal phytoremediation. Water Air Soil Poll 223:3249–3262
He S, He Z, Yang X, Stoffella PJ, Baligar VC (2015) Soil biogeochemistry, plant physiology, and phytoremediation of cadmium-contaminated soils. Adv Agron 134:135–225
He Y, Xu J, Tang C, Wu Y (2005) Facilitation of pentachlorophenol degradation in the rhizosphere of ryegrass (Loliumperenne L.) Soil Biol Biochem 37:2017–2024
He Y, Xu J, Lv X, Ma Z, Wu J, Shi J (2009) Does the depletion of pentachlorophenol in root-soil interface follow a simple linear dependence on the distance to root surfaces. Soil Biol Biochem 41:1807–1813
Hernandez-Allica J, Becerril JM, Zarate O, Garbisu C (2006) Assessment of the efficiency of a metal phytoextraction process with biological indicators of soil health. Plant Soil 281:147–158
Hinojosa MB, Carreira JA, Garcıa-Ruız R, Dick RP (2005) Microbial response to heavy metal-polluted soils: community analysis from phospholipid-linked fatty acids and ester-linked fatty acids extracts. J Environ Qual 34:1789–1800
Hortal S, Bastida F, Armas C, Lozano YM, Moreno JL, García C, Pugnaire FI (2013) Soil microbial community under a nurse-plant species changes in composition, biomass and activity as the nurse grows. Soil Biol Biochem 64:139–146
Huang G, Cao YF, Wang B, Li Y (2015) Effects of nitrogen addition on soil microbes and their implications for soil Cemission in the Gurbantunggut Desert, center of the Eurasian continent. Sci Total Environ 515–516:215–224
Kenarova A, Boteva S (2015) Functional diversity of microorganisms in heavy metal-polluted soils. In: Heavy metal contamination of soils. Springer International Publishing, pp 245–257
Kenarova A, Radeva G, Traykov I, Boteva S (2014) Community level physiological profiles of bacterial communities inhabiting uranium mining impacted sites. Ecotox Environ Safe 100:226–232
Kim KR, Owens G, Kwon SL (2010) Influence of Indian mustard (Brassica juncea) on rhizosphere soil solution chemistry in long-term contaminated soils: a rhizobox study. J Environ Sci 22:98–105
Klimek B, Sitarz A, Choczyński M, Niklińska M (2016) The effects of heavy metals and total petroleum hydrocarbons on soil bacterial activity and functional diversity in the Upper Silesia Industrial Region (Poland). Water Air Soil Poll 227:1–9
Kohler J, Caravaca F, Azcon R, Díaz G, Roldan A (2016) Suitability of the microbial community composition and function in a semiarid mine soil for assessing phytomanagement practices based on mycorrhizal inoculation and amendment addition. J Environ Manag 169:236–246
Kwonrae K, Owens G, Naidu R, Soonlk K (2010) Influence of plant roots on rhizosphere soil solution composition of long-term contaminated soils. Geoderma 155:86–92
Leita L, De Nobili M, Muhlbachova G, Mondini C, Marchiol L, Zerbi G (1995) Bioavailability and effects of heavy metals on soil microbial biomass survival during laboratory incubation. Biol Fertil Soils 19:103–108
Li Z, Wu LH, Luo YM, Christie P (2014) Dynamics of plant metal uptake and metal changes in whole soil and soil particle fractions during repeated phytoextraction. Plant Soil 374:857–869
Liao QL, Liu C, Wu HY, Jin Y, Hua M, Zhu BW, Chen K, Huang L (2015) Association of soil cadmium contamination with ceramic industry: a case study in a Chinese town. Sci Total Environ 514:26–32
Liu D, Fang S, Tian Y, Dun X (2012) Variation in rhizosphere soil microbial index of tree species on seasonal flooding land: an in situ rhizobox approach. Appl Soil Eco 59:1–11
Liu L, Wu LH, Li N, Luo YM, Li SL, Li Z, Han CL, Jiang YG, Christie P (2011) Rhizosphere concentrations of zinc and cadmium in a metal contaminated soil after repeated phytoextraction by Sedum plumbizincicola. Int J Phytoremediat 13:750–764
Lu H, Li Z, Fu S, Méndez A, Gascó G, Paz-Ferreiro J (2015) Combining phytoextraction and biochar addition improves soil biochemical properties in a soil contaminated with Cd. Chemosphere 119:209–216
Mackie KA, Marhan S, Ditterich F, Schmidt HP, Kandeler E (2015) The effects of biochar and compost amendments on copper immobilization and soil microorganisms in a temperate vineyard. Agric Ecosyst Environ 201:58–69
Mandal A, Purakayastha TJ, Patra AK (2014) Phytoextraction of arsenic contaminated soil by Chinese brake fern (Pteris vittata): effect on soil microbiological activities. Biol Fert Soils 50:1247–1252
McGrath SP, Shen ZG, Zhao FJ (1997) Heavy metal uptake and chemical changes in the rhizosphere of Thlaspi caerulescens and Thlaspi ochroleucum grown in contaminated soils. Plant Soil 188:153–159
Mitchell PJ, Simpson AJ, Soong R, Simpson MJ (2015) Shifts in microbial community and water-extractable organic matter composition with biochar amendment in a temperate forest soil. Soil Biol Biochem 81:244–254
Muehe EM, Weigold P, Adaktylou IJ, Planer-Friedrich B, Kraemer U, Kappler A, Behrens S (2015) Rhizosphere microbial community composition affects cadmium and zinc uptake by the metal-hyperaccumulating plant Arabidopsis halleri. Appl Environ Microb 81:2173–2181
Myers RT, Zak DR, White DC, Peacock A (2001) Landscape level patterns of microbial community composition and substrate use in upland forest ecosystems. Soil Sci Soc Am J 65:359–367
Olsson PA, Baath E, Jakobsen I, Soderstrom B (1995) The use of phospholipid and neutral lipid fatty acids to estimate biomass of arbuscular mycorrhizal fungi in soil. Mycol Res 99:623–629
Otero XL, Álvarez E, Fernández-Sanjurjo MJ, Macías F (2012) Micronutrients and toxic trace metals in the bulk and rhizospheric soil of the spontaneous vegetation at an abandoned copper mine in Galicia (NW Spain). J Geochem Explor 112:84–92
Pennanen T, Frostegård Å, Fritze H, Bååth E (1996) Phospholipid fatty acids and heavy metal tolerance of soil microbial communities along two heavy metal-polluted gradients in coniferous forests. Appl Environ Microb 62:420–428
Perez-de-Mora A, Burgos P, Madejon E, Cabrera F, Jaeckel P, Schloter M (2006) Microbial community structure and function in a soil contaminated by heavy metals: effects of plant growth and different amendments. Soil Biol Biochem 38:327–341
Pueyo M, Mateu J, Rigol A, Vidal M, Lopez-Sanchez JF, Rauret G (2008) Use of the modified BCR three-step sequential extraction procedure for the study of trace element dynamics in contaminated soils. Environ Pollut 152:330–341
Puschenreiter M, Wieczorek S, Horak O, Wenzel WW (2003) Chemical changes in the rhizosphere of metal hyperaccumulator and excluder Thlaspi species. J Plant Nutr Soil Sci 166:579–584
Qin H, Brookes PC, Xu J (2014) Cucurbita spp. and Cucumis sativus enhance the dissipation of polychlorinated biphenyl congeners by stimulating soil microbial community development. Environ Pollut 184:306–312
Ridl J, Kolar M, Strejcek M, Strnad H, Stursa P, Paces J, Macek T, Uhlik O (2016) Plants rather than mineral fertilization shape microbial community structure and functional potential in legacy contaminated soil. Front Microbiol 7
Robson TC, Braungardt CB, Rieuwerts J, Worsfold P (2014) Cadmium contamination of agricultural soils and crops resulting from sphalerite weathering. Environ Pollut 184:283–289
Rousk J, Bååth E, Brookes PC, Lauber CL, Lozupone C, Caporaso JG, Knight R, Fierer N (2010) Soil bacterial and fungal communities across a pH gradient in an arable soil. The ISME J 4:1340–1351
Setälä H, Berg MP, Jones TH (2005) Trophic structure and functional redundancy in soil communities. In: Bardgett RD, Usher MB, Hopkins DW (eds) Biological diversity and function in soils. Cambridge University Press, Cambridge, pp 236–249
Singh BK, Quince C, Macdonald CA, Khachane A, Thomas N, Al-Soud WA, Sørensen SJ, He Z, White D, Sinclair A, Crooks B, Zhou J, Campbell CD (2014) Loss of microbial diversity in soils is coincident with reductions in some specialized functions. Environ Microbiol 16:2408–2420
Sousa NR, Ramos MA, Marques APGC, Castro PML (2014) A genotype dependent-response to cadmium contamination in soil is displayed by Pinuspinaster in symbiosis with different mycorrhizal fungi. Appl Soil Eco l76:7–13
Stefanowicz AM, Niklinska M, Kapusta P, Szarek-Łukaszewska G (2010) Pine forest and grassland differently influence the response of soil microbial communities to metal contamination. Sci Total Environ 408:6134–6141
Stefanowicz AM, Kapusta P, Szarek-Łukaszewska G, Grodzińska K, Niklińska M, Vogt RD (2012) Soil fertility and plant diversity enhance microbial performance in metal-polluted soils. Sci Total Environ 439:211–219
Strickland MS, Rousk J (2010) Considering fungal: bacterial dominance in soils—methods, controls, and ecosystem implications. Soil Biol Biochem 42:1385–1395
Sun Y, Xu Y, Xu Y, Wang L, Liang X, Li Y (2016) Reliability and stability of immobilization remediation of Cd polluted soils using sepiolite under pot and field trials. Environ Pollut 208:739–746
Sungur A, Soylak M, Yilmaz S, Özcan H (2014) Determination of heavy metals in sediments of the Ergene River by BCR sequential extraction method. Environ Earth Sci 72:3293–3305
Tian J, McCormack L, Wang J, Guo D, Wang Q, Zhang X, Yu G, Blagodatskaya E, Kuzyakov Y (2015) Linkages between the soil organic matter fractions and the microbial metabolic functional diversity within a broad-leaved Korean pine forest. Eur J Soil Biol 66:57–64
Tscherko D, Ute H, Marie-Claude M, Ellen K (2004) Shifts in rhizosphere microbial communities and enzyme activity of Poa alpina across an alpine chronosequence. Soil Biol Biochem 36:1685–1698
Turpeinen R, Kairesalo T, Häggblom MM (2004) Microbial community structure and activity in arsenic-, chromium- and copper- contaminated soils. FEMS Microbiol Ecol 47:39–50
Wang J, Zhang CB, Jin ZX (2009) The distribution and phytoavailability of heavy metal fractions in rhizosphere soils of Paulowniufortunei (seem) Hems near a Pb/Zn smelter in Guangdong, PR China. Geoderma 148:299–306
Wei J, Liu X, Zhang X, Chen X, Liu S, Chen L (2014) Rhizosphere effect of Scirpus triqueter on soil microbial structure during phytoremediation of diesel-contaminated wetland. Environ Technol 35:514–520
Wei S, Twardowska I (2013) Main rhizosphere characteristics of the Cd hyperaccumulator Rorippa globosa (Turcz.) Thell. Plant Soil 372(1–2):669–681
Wu Y, Ding N, Wang G, Xu J, Wu J, Brookes PC (2009) Effects of different soil weights, storage times and extraction methods on soil phospholipid fatty acid analyses. Geoderma 150:171–178
Yang WH, Hu H, Ru M, Ni WZ (2013a) Changes of microbial properties in (near–) rhizosphere soils after phytoextraction by Sedum alfredii H: a rhizobox approach with an artificial Cd-contaminated soil. Appl Soil Ecol 72:14–21
Yang L, Wang GP, Cheng Z, Liu Y, Shen ZG, Luo CL (2013b) Influence of the application of chelant EDDS on soil enzymatic activity and microbial community structure. J Hazard Mater 262:561–570
Yang WH, Li H, Zhang TX, Lin S, Ni WZ (2014) Classification and identification of metal-accumulating plant species by cluster analysis. Environ Sci Pollut Res 21:10626–10637
Yang Y, Campbell CD, Clark L, Cameron CM, Paterson E (2006) Microbial indicators of heavy metal contamination in urban and rural soils. Chemosphere 63:1942–1952
Yao HY, Campbell CD, Qiao XR (2011) Soil pH controls nitrification and carbon substrate utilization more than urea or charcoal in some highly acidic soils. Biol Fert Soils 47:515–522
Yergeau E, Sanschagrin S, Maynard C, St-Arnaud M, Greer CW (2014) Microbial expression profiles in the rhizosphere of willows depend on soil contamination. The ISME J 8:344–358
Ying JY, Zhang LM, Wei WX, He JZ (2013) Effects of land utilization patterns on soil microbial communities in an acid red soil based on DNA and PLFA analyses. J Soils Sediments 13:1223–1231
Yoshitake S, Fujiyoshi M, Watanabe K, Masuzawa T, Nakatsubo T, Koizumi H (2013) Successional changes in the soil microbial community along a vegetation development sequence in a subalpine volcanic desert on Mount Fuji, Japan. Plant Soil 364:261–272
Yu J, Unc A, Zhang X, Steinberger Y (2016) Responses of the soil microbial catabolic profile and diversity to vegetation rehabilitation in degraded semiarid grassland. Appl Soil Ecol 101:124–131
Yuan KN (1963) Studies on the organic-mineral complex in soil: the oxidation stability of humus from different organo-mineral complexes in soil. Acta Pedol Sin 11:286–293 (in Chinese)
Zelles L (1999) Fatty acid patterns of phospholipids and lipopolysaccharides in the characterisation of microbial communities in soil: a review. Biol Fert Soils 29:111–129
Zhou X, Wu H, Koetz E, Xu Z, Chen C (2012) Soil labile carbon and nitrogen pools and microbial metabolic diversity under winter crops in an arid environment. Appl Soil Ecol 53:49–55
Zogg GP, Zak DR, Ringelberg DB, MacDonald NW, Pregitzer KS, White DC (1997) Compositional and functional shifts in microbial communities due to soil warming. Soil Sci Soc Am J 61:475–481
Zornoza R, Acosta JA, Faz A, Bååth E (2016) Microbial growth and community structure in acid mine soils after addition of different amendments for soil reclamation. Geoderma 272:64–72
Zornoza R, Acosta JA, Martínez-Martínez S, Faz A, Bååth E (2015) Main factors controlling microbial community structure and function after reclamation of a tailing pond with aided phytostabilization. Geoderma 245:1–10
Acknowledgements
We sincerely acknowledge the supports from the National Natural Science Foundation of China (No. 41501345), the Natural Science Foundation of Fujian Province (No. 2015J01155), and the Foundation for Distinguished Young Scholars of Fujian Agriculture and Forestry University (No. XJQ201628).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Robert Duran
Electronic supplementary material
ESM 1
(DOC 185 kb)
Rights and permissions
About this article

Cite this article
Yang, W., Zhang, T., Lin, S. et al. Distance-dependent varieties of microbial community structure and metabolic functions in the rhizosphere of Sedum alfredii Hance during phytoextraction of a cadmium-contaminated soil. Environ Sci Pollut Res 24, 14234–14248 (2017). https://doi.org/10.1007/s11356-017-9007-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-017-9007-4