
Abstract
Purpose
Soil nitrogen (N) availability is a critical determinant of plantation productivity in subtropical Australia and is influenced by the soil microbial community. The size, structure and function of the soil microbial community can be impacted by land-use change and residue management. The objectives of this study were to examine the impact of land-use change from (1) native forest (NF) to first rotation (1R) hoop pine plantation and (2) 1R hoop pine plantation to second rotation (2R) hoop pine plantation on the soil microbial community. The impact of residue management on the soil microbial community was also investigated in the 2R forest, where soil microbial parameters were measured in tree rows (2R-T) and windrows (2R-W). In addition, relationships between soil microbial parameters and soil N parameters were investigated.
Materials and methods
Each of the four treatments (NF, 1R, 2R-T and 2R-W) had five 24-m2 replicate plots from which 15 soil cores were collected and bulked at three depths (0–10, 10–20, 20–30 cm). Microbial biomass carbon (MBC) and N (MBN) and soil respiration were measured on field moist soils. In addition, carbon (C) source utilisation patterns were assessed using the whole soil MicroResp™ technique (Campbell et al. 2003).
Results and discussion
Results indicate that the land-use change from NF to 1R hoop pine plantation significantly reduced MBC, respiration rate, soil total C and total N. Furthermore, the land-use change appeared to have a significant impact on the soil microbial community composition measured using MicroResp™ profiles. Land-use change from 1R to 2R hoop pine plantation resulted in a decline in total C and MBN and a shift in microbial community composition. When compared to the 2R-T soils, the 2R-W soils tended to have a greater microbial biomass and respiration rate. Residue management also influenced the microbial community composition measured in the MicroResp™ profiles.
Conclusions
Results indicate that land-use change had a significant impact on the soil microbial community, which was likely to be related to shifts in the quality and quantity of organic inputs associated with the change in land use. This may have significant implications for the long-term productivity of the soil resource. Further studies are required to confirm a difference in microbial community composition associated with residue management. In addition, long-term experiments in subtropical Australia are necessary to verify the results of this snapshot study and to improve our understanding of the impact of single-species plantation forestry and residue management on the soil microbial community, soil N dynamics and ultimately the long-term sustainability of the soil resource.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
The soil microbial community plays a central role in organic matter turnover and the cycling of almost all major plant nutrients, including nitrogen (N) (Smith and Paul 1990; Doran and Zeiss 2000; Xu et al. 2008, 2009). As such, it is a key factor influencing ecosystem functioning and the sustainability of the soil resource (Sparling 1997). Research has shown that soil microorganisms are sensitive to land use and management and can be used to indicate soil health (Sparling 1997; Chen et al. 2000; Gomez et al. 2000; Li et al. 2004; Zhang and Xu 2008). Shifts in the soil microbial community (population, activity and/or composition) associated with land-use change and management techniques may influence soil N pools and dynamics (He et al. 2005; Pan et al. 2008, 2009; He et al. 2009a, b; Xu et al. 2009).
In forest soils, the growth, activity and composition (i.e. diversity) of the soil microbial community are affected by abiotic and biotic factors including climate, tree species, quality and quantity of organic matter input, nutrient availability and physical disturbance (Priha et al. 1999; Leckie et al. 2004; Grayston and Prescott 2005; Hannam et al. 2006; He et al. 2006). These factors may in turn be influenced by land-use change (e.g. harvesting and change in stand composition) and silvicultural techniques (e.g. residue management) (Burton et al. 2007a, b; Chen and Xu 2008; He et al. 2008; Xu et al. 2009). There has been a substantial amount of research into the impact of tree species and stand composition, as well as harvesting and residue management, on the soil microbial community in temperate and boreal forests (Zhang et al. 2007a, b; Huang et al. 2008; Zhang et al. 2009); however, there is limited information available for subtropical forests, particularly in relation to a chronosequence of land-use change and management.
A number of soil microbiological parameters have been used to assess the impact of land-use change and management on the size, activity and composition or diversity of the soil microbial community (Paul and Clark 1996; Xu et al. 2009). Soil microbial biomass and respiration can be used as general indexes of the size and activity (i.e. CO2-C evolution or C turnover) of the soil microbial community (Paul and Clark 1996). Both parameters tend to be sensitive to land-use change and management and have traditionally been used as indicators of soil fertility, with decreases indicating a decline in soil quality or health (Elliott et al. 1996; Chen et al. 2000). In addition, the shift in soil microbial community composition has been used as an indicator of soil health (e.g. Yao et al. 2000). Common methods used to assess shifts in the microbial community as a result of land use and management include phospholipid fatty acid analysis, denaturing gradient gel electrophoresis and community level physiological profiles (CLPP) (Campbell et al. 2003; Bucher and Lanyon 2005; Grayston and Prescott 2005; Cookson et al. 2007). The CLPP techniques measure patterns of carbon (C) source utilisation to detect the shift in soil microbial community functional diversity.
In this study, measurements of soil microbial biomass, activity and community structure (CLPP) were used to test the hypothesis that the land-use change from a mixed-species native forest (NF) to a single-species first rotation (1R) hoop pine plantation, and subsequent second rotation (2R) hoop pine plantation and associated residue management practices, would have a significant impact on the soil microbial community. In addition, relationships between soil microbial parameters and available soil N parameters were investigated.
2 Materials and methods
2.1 Site description and sample collection
This study was conducted in Yarraman State Forest, southeast Queensland, Australia (26° 52′ S, 151° 51′ E). Details of the study area were provided by Chen et al. (2004). In brief, annual rainfall at this site ranges between 433 and 1,110 mm, with an average of 816 mm. The soil was a Typic Durustalf (Soil Survey Staff 1999), with a clayey texture (Chen et al. 2004). The NF site is classified as a mixed rainforest/scrub and is dominated by bunya pine (Araucaria bidwilli Hook.), yellowwood (Terminalia oblongata F. Muell. subsp. oblongata), crows ash (Pentaceras australis R.B.) and lignum-vitae (Premna lignum-vitae), with emergent hoop pine (Araucaria cunninghamii Aiton ex A. Cunn.). Experimental sites measuring 0.2 ha in area were located in adjacent NF, 1R hoop pine plantation (53 years old) and 2R hoop pine plantation (5 years old). Both the 1R and 2R hoop pine plantation sites were converted from native forest in 1952. The first rotation of hoop pine at the 2R site was clearfall harvested in 1999, and post harvest residues from the 1R plantation were formed into windrows approximately 6 m apart, using a D6 bulldozer with shear blade. The areas between windrows were then used as tree-planting rows for the 2R hoop pine plantation. Hence, the 2R plantation experimental area was divided into two treatments based on the residue management practices. These were (1) tree planting row (2R-T) and (2) windrow of harvest residues (2R-W). A buffer area of at least 100 m was left between experimental areas to avoid edge effects. Each of the four treatments had five 24-m2 replicate plots. Fifteen soil cores were randomly collected from each plot at three depths (0–10, 10–20 and 20–30 cm), using a 7.5-cm-diameter auger and bulked. All samples were transported to the laboratory where field moist soils were well mixed and passed through a 2-mm sieve and the fraction <2 mm was stored at 4°C until the analysis could be conducted. A subsample of each soil was air-dried at room temperature for analysis of soil basic physical and chemical characteristics, which are shown in Table 1. Soil total C and N were analysed using an isotope ratio mass spectrometer with a Eurovector Elemental Analyser (Isoprime-EuroEA 3000, Milan, Italy). Soluble organic N (SON) and mineral N were measured using the methods described in Burton et al. (2007a, b). Soil CEC, pH and bulk density were measured using the methods described by Rayment and Higginson (1992).
2.2 Microbial biomass C and N
Microbial biomass C (MBC) and microbial biomass N (MBN) were measured using the fumigation–extraction method described by Vance et al. (1987). In brief, fumigated and nonfumigated soils (10 g dry weight equivalent) were extracted with 40 ml of 0.5 M K2SO4 (soil/extractant ratio 1:4). Samples were shaken for 30 min, and filtered through a Whatman 42 filter paper and frozen until further analysis could be conducted. Soluble organic C and total N in the extracts of fumigated and nonfumigated samples were determined using a SHIMADZU TOC-VCPH/CPN analyser (fitted with TN unit). Microbial biomass C and N were calculated using a conversion factor for C (E c) of 2.64 (Vance et al. 1987) and for N (E n) of 2.22 (Brookes et al. 1985; Jenkinson 1988).
2.3 Soil respiration
Soil respiration was measured using the method described by Chen et al. (2000). Field moist subsamples (20 g dry weight equivalent) were placed in beakers and aerobically incubated at 22°C and at constant humidity in sealed 1-L glass jars. Carbon dioxide evolved from the soil was trapped in 0.1 M NaOH and measured after 24 h, 3 days, 7 days, 14 days, 21 days and 28 days by titration with 0.05 M HCl to the phenolphthalein end point after the addition of 1 M BaCl2. A number of controls (i.e. jars without soil) were subjected to the same conditions and used as blanks. The amount of carbon dioxide evolved was calculated from the difference in molarity between the NaOH from blanks and samples. The metabolic quotient (qCO2) was calculated as the ratio of respiration (µg CO2-C g–1 h–1) to MBC.
2.4 Community level physiological profiles
Carbon source utilisation patterns (often referred to as community level physiological profiles, or CLPP) were assessed using the whole soil MicroResp™ technique. Analysis was performed within 72 h of the completion of sample processing. The MicroResp™ colourimetric detection plates were prepared and profiles obtained according to Campbell et al. (2003). Briefly, soil samples, all of which were at >40% of the water holding capacity, were conditioned at 25°C in a humid environment for 2 days prior to analysis. Based on the work of Campbell et al. (1997, 2003), 15 ecologically relevant and easily dissolvable C substrates were selected (Table 2), and stock solutions were made from which 25-ml aliquots could be dispensed to deliver 30 mg C per gram of soil water to each deep well. MicroResp™ analysis was carried out in triplicate. The C solutions and water (to be used as basal respiration) were dispensed into a deep well plate before soil subsamples, each with a total volume of 300 μl, and were placed into each well of the deep well plate using the method described by Campbell et al. (2003). The deep well plate was then immediately sealed with a gasket and detection plate and incubated at 25°C for 6 h. In order to calculate colour development (C utilisation), the detection plate colour was measured as absorbance at 590 nm immediately before and after the 6-h incubation using a Biolog™ microplate reader (B62302A, Molecular Devices, CA, USA). The 6-h absorbance data were normalised for any differences in detection plates recorded prior to incubation. Basal respiration (for water) and substrate-induced respiration (SIR) (for individual C substrates) were calculated as CO2-C evolved according to Campbell et al. (2003). For each plate, the average amount of CO2-C that evolved per sample was calculated and used to normalise individual well responses before multivariate analysis was conducted.
2.5 Statistical analysis
A split-plot factorial analysis of variance (ANOVA) was used to explore differences within biomass measurements based on the factors forest type and soil depth. Where significant differences were detected, pairwise comparisons were made using the Tukey adjustment for multiple range testing. One-way ANOVAs were used to evaluate the soil respiration and metabolic quotient data as well as MicroResp™ SIR data from individual C substrates. Least significant difference (P < 0.05) was used to separate treatment means when differences were significant. The assumptions of normality and equal variance were satisfied prior to this analysis being conducted in SAS version 9.1.3.
Patterns of C source utilisation among the forest types were examined by principal component analysis (PCA) and nonmetric multidimensional scaling (NMS) using Bray–Curtis distance measure. For the NMS analysis, the multiple response permutations procedure (MRPP) was used to determine whether groups were statistically different, and Bonferroni adjustment was used to ensure the overall error rate was 0.05. It should be noted that MRPP tests for differences among groups were based on both location and variation (Mielke and Berry 2001). Hence, significant differences found among the forest types may be due to either distance between the groups or variability within the groups. Cluster analysis was also performed using Bray–Curtis as the distance measure with graphical representations based on complete linkage for the hierarchical clustering. For the MicroResp™ data, substrates that did not induce respiration were removed for analysis. All multivariate analysis was carried out on normalised data using the Statistical package R version 2.4.0 (R Development Core Team 2008).
3 Results
3.1 Microbial biomass
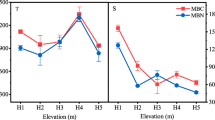
A significant interaction was found between forest type and soil depth for both MBC (P < 0.001) and MBN (P < 0.001). Hence, the extent to which forest type affected MBC and MBN varied with the soil depth. In the 0- to 10-cm layer, MBC ranged from 1,186 μg g–1 in the 2R-T soils to 2,156 μg g–1 in the NF soil, with values decreasing with soil depth (Table 3). The NF soil had significantly higher MBC than the 1R soil in the 0- to 10-cm layer, but there was no significant difference in MBC between 1R and 2R soils or between 2R-T and 2R-W soils at any depth.
The MBN ranged between 130 μg g–1 in the 2R-T soil and 231 μg g–1 in the NF soil, and also decreased with the soil depth (see Table 3). Although the NF soil tended to have higher MBN than the 1R soil in all depths, the differences were not significant. In the 0- to 10-cm layer, the 1R soil had significantly higher MBN than the 2R-T soil; however, values were similar at lower depths. There was no significant difference in MBN values between the 2R-T and 2R-W soils. The microbial C/N ratio (ratio of MBC to MBN) ranged between 7.3 and 9.4 in the 0- to 10-cm layer and tended to increase with the soil depth. There were generally no differences in the microbial C/N ratio among the forest types. The MBC constituted up to 2.5% of the soil total C, while MBN constituted up to 3.8% of the soil total N, neither were significantly affected by forest type (see Table 3).
Concentrations of MBC in the 0–10 cm were highly correlated with soil total C and N, hot water extractable organic C and N, hot KCl extractable organic C and N, and mineral N (Table 4). Concentrations of MBN in the 0- to 10-cm layer had similar relationships with total C and N, pools of soluble organic C (SOC), SON and mineral N, and soil C/N ratio as concentrations of MBC.
3.2 Soil respiration and metabolic quotients
The average soil respiration rate over the 28-day incubation ranged from 0.78 μg CO2-C g–1 h–1 in the 2R-T soil to 1.12 CO2-C g–1 h–1 in the NF soil (Fig. 1), while cumulative CO2-C production for the 28-day incubation period was between 530 μg CO2-C g–1 in the 2R-T soil and 755 μg CO2-C g–1 in the NF soil (Fig. 2). Both the average respiration rate and the cumulative CO2-C production were consistently higher in the NF soil compared to the 1R soil; however, no significant differences were found among the plantation soils (see Figs. 1 and 2). Metabolic quotients ranged from 0.52 in the NF soil to 0.74 in the 2R-T soil, and were not significantly different among the forest types. The average respiration rate was positively correlated with soil total C and N (P < 0.05), mineral N (P < 0.001), pools of soluble organic C extracted by hot water and hot KCl (P < 0.001) and soluble organic N extracted by hot water (P < 0.01) and hot KCl (P < 0.001), and negatively correlated with soil C/N ratio (P < 0.05) (see Table 4).

Respiration rate (black bars) and metabolic quotient (grey bars) in the 0- to 10-cm soil layer of adjacent native forest (NF), 53-year-old first rotation hoop pine plantation (1R), 5-year-old second rotation tree row (2R-T) and second rotation windrow (2R-W) at the Yarraman site, subtropical Australia (standard errors shown by vertical bars)

Cumulative respiration rate in the 0- to 10-cm soil layer of adjacent native forest (NF), 53-year-old first rotation hoop pine plantation (1R), 5-year-old second rotation tree row (2R-T) and second rotation windrow (2R-W) at the Yarraman site, subtropical Australia (standard errors shown by vertical bars)
3.3 MicroResp™
All MicroResp™ results presented here are the means of the triplicates from each of the five replicates for the four treatments. The mean basal respiration (with no C source), measured from the wells containing water only, were 1.03, 0.99, 0.86 and 0.79 μg CO2-C g–1 h–1 in the 1R, NF, 2R-W and 2R-T soils, respectively. The only significant difference in the basal respiration was between the 1R soil and the 2R-T soil. Three of the fifteen C substrates, namely, γ-aminobutyric acid, 3,4-dihydroxybenzoic acid and l-cysteine hydrochloride, produced undetectable SIR in all soil samples. Table 2 displays mean SIR of the remaining 12 C substrates. The highest level of SIR in all forest types was observed with d-fructose, while the lowest detectable SIR was observed with l-lysine. The NF soil tended to have higher SIR than the 1R soil, although the difference was not always significant (see Table 2). In most cases, the 1R soil had similar SIR to the 2R soils; however, there were some instances (e.g. d-glucose, l-alanine, trehalose) where SIR was higher in the 1R soil than either or both of the 2R-T and 2R-W soils. No significant differences were found between the 2R-T and 2R-W soils (see Table 2).
PCA of the MicroResp™ data showed that PC 1 accounted for 71% of the variation while PC 2 accounted for a further 12% (a total of 83% for the first two PCs with 90% of the variation accounted for by the third PC). However, forest types were not separated into distinct groups based on the PCA (Fig. 3a). In contrast, NMS analysis of the MicroResp™ profiles revealed replicates of the different forest types tended to group together (Fig. 3b). Further analysis using MRPP revealed that the pattern of SIR in the NF soil was significantly different from the pattern produced for the 1R soil (δ = 0.0766, P = 0.0080). The pattern of SIR in the 1R soil was significantly different from that of the 2R-T soil (δ = 0.1246, P = 0.0060) and the 2R-W soil (δ = 0.1548, P = 0.0070). However, no significant difference was found between the patterns produced by the 2R-T and 2R-W soils (δ = 0.1985, P = 0.0170). These values are based on the P value obtained using Bonferroni adjustment (P = 0.0083).

Results of (a) PCA (PC1 accounted for 71% of the variation while PC2 accounted for a further 12% of the variation), (b) NMS and (c) cluster analysis (scale indicates Bray–Curtis distance with graphical representations based on complete linkage for the hierarchical clustering) of the normalized absorbance data of the 12 C-sources from the MicroResp™ profiles of the 0- to 10-cm soil layer of the adjacent native forest (NF) (numbers 16–20), 53-year-old first rotation hoop pine plantation (1R) (numbers 11–15), 5-year-old second rotation tree row (2R-T) (numbers 1–5) and second rotation windrow (2R-W) (numbers 6–10), at incubation time of 6 h
With the exception of one of the 2R-W replicates (sample number 6), cluster analysis of the MicroResp™ SIR profiles separated the NF, 1R, 2R-T and 2R-W replicates into distinguished clusters, linking together at a Bray–Curtis distance of approximately 0.10 (Fig. 3c). Cluster analysis of the MicroResp™ profiles revealed that the NF and 1R samples were most similar (linking together at a distance of 0.25) followed by 2R-W, which was linked with 1R and NF at a relative distance of 0.3, while 2R-T was most different from the other soils (see Fig. 3c).
4 Discussion
4.1 Soil microbial biomass and respiration
The mechanisms through which the land-use change from the NF to the hoop pine plantation may affect the soil microbial community are related to the change in tree species and the disturbance associated with logging of the NF as well as subsequent establishment of the 1R plantation and ensuing silvicultural techniques. Shifts in tree species may result in changes in the quality and quantity of both above-ground (litter) and below-ground (roots) organic matter input, as well as changes in microclimate (Priha and Smolander 1997; He et al. 2006). The NF in this study is composed of a mixture of tree species including both hardwood and conifer species, while the plantation is a single-species conifer forest. It has been suggested that due to the presence of a waxy surface layer and higher concentrations of recalcitrant compounds (e.g. phenolic compounds), conifer needles are more resistant to decomposition than leaf litter from hardwoods (Priha and Smolander. 1997; Li et al. 2004). Past studies have found a decline in microbial biomass associated with land-use change from native forest to plantations, as well as differences in soil respiration associated with different stand types (Waldrop et al. 2000; Priha et al. 2001; Chen et al. 2004). A comparison of beech (a northern hemisphere hardwood) and conifers (Scots pine and Norway spruce) found that MBN was significantly lower in the conifer stand compared to the beech stand (Zhong and Makeschin 2006). Similarly, Priha et al. (2001) found lower MBC and respiration under conifers (Scots pine and Norway spruce) compared to hardwoods (silver birch). However, Priha and Smolander (1997) compared hardwoods (silver birch) and conifers (Scots pine and Norway spruce) and found that stand type had no clear effect on MBC and MBN after 24 years.
In this study, concentrations of MBC and MBN (in all depths) and respiration rates in the NF and plantation sites were comparable to those reported for other plantation soils in southeast Queensland (Chen et al. 2002; Chen and Xu 2005). The land-use change from NF to 1R hoop pine plantation was associated with a significant reduction in the concentration of MBC in the 0- to 10-cm layer. Respiration rate, both on an hourly basis and over the 28-day incubation period, was also reduced as a result of the land-use change. Past research at this study site found that the NF soil had a lower alkyl C/O-alkyl ratio than the 1R soil (Chen et al. 2004). Also, analyses of litter and soil samples show that the NF had significantly lower C/N ratios (28 and 12, respectively) than the 1R forest (69 and 14, respectively). Together, these results indicate that the higher MBC and respiration in the NF compared to the 1R soil is associated with higher-quality organic matter input in the NF.
The hot water and hot KCl extractable SOC and SON pools are believed to represent labile fractions of soil organic C and N pools (Curtin and Wen 1999; Chen et al. 2004; Curtin et al. 2006; Chen and Xu 2008). The MBC, MBN and respiration were positively correlated with soluble organic C and N extracted by hot water and hot KCl, as well as soil total C and total N, and were negatively related to the soil C/N ratio (see Table 3). Higher percentages of total C and total N (see Table 1) and larger pools of labile SON and SOC in the NF soil compared to the 1R soil (Burton et al. 2007ab), together with the strong positive relationship, tend to suggest that the NF has greater quantity and quality of organic matter available for decomposition by the microbial community than the 1R forest. Smolander and Kitunen (2002) found that microbial biomass and activity were correlated with dissolved organic nitrogen, while Li et al. (2004) found that MBC and MBN were correlated with soil total C and total N. In this study, the negative relationship of microbial parameters with the soil C/N ratio suggests that N is limiting in the plantation soils, which have higher C/N ratios than the NF soil (see Table 1). Previous work on these soils showed that the NF soil had a significantly larger mineral N pool than the 1R soil (Burton et al. 2007a). The significant positive correlation of mineral N with MBC, MBN and respiration rate indicates that the larger soil microbial biomass and greater microbial activity in the NF soils are associated with the larger pools of mineral N in the NF soils. These results suggest that the change in microbial parameters associated with the change in land use may impact N transformation processes and mineral N availability which in turn may impact plantation productivity.
Plantation harvesting and the establishment of a subsequent hoop pine rotation may cause disturbance and compaction of the soil system as well as changes in the quantity and quality of organic matter and the microclimate, which may in turn affect the soil microbial community (Breland and Hansen 1996; Li et al. 2004). Research has revealed varying effects of harvesting on the soil microbial community, with researchers reporting no effect (e.g. Hannam et al. 2006), and decreases in biomass and respiration (e.g. Luizao et al. 1992; Pietikainen and Fritze 1995). Residue management may control the availability of organic matter and soil microclimate and therefore may also affect the soil microbial community (Chen and Xu 2005). A study in hoop pine plantations of subtropical Australia indicates that residue retention may decrease nutrient loss and increase soil C and N (Blumfield and Xu 2003; Mathers et al. 2003b). Furthermore, solid-state 13C NMR analysis of soils in hoop pine and eucalypt plantations of subtropical Australia revealed residue retention improved the quality of soil organic matter (Mathers et al. 2003a, b). A study undertaken in a 6-year-old slash pine plantation of subtropical Australia revealed that residue retention increased MBC and MBN, but had no significant effect on soil respiration and metabolic quotient (qCO2) (Chen and Xu 2005).
In this study, there were some indications that harvesting and residue management may influence the microbial community; however, the lack of statistical significance suggests that the impact is not highly significant 5 years into the 2R of the hoop pine plantation. Studies elsewhere have had varying results with Smaill et al. (2010), finding evidence that variation in microbial properties related to harvesting and residue management can persist for a considerable length of time. Alternatively, Hannam et al. (2006) found evidence to suggest that the soil microbial community may respond to harvesting immediately, but then return to preharvest levels within 4–5 years.
4.2 Community level physiological profiles
Community level physiological profiles, such as MicroResp™, produce results rapidly and have been used regularly in forest soil research (e.g. Li et al. 2004; Bucher and Lanyon 2005; Grayston and Prescott 2005; Huang et al. 2008) as a tool to indicate relative differences in community composition or “functional diversity”, based on differences in patterns of C substrate utilisation. It is important to note that CLPPs are only indicators of functional diversity or community composition based on the ability of the soil microbial community to utilise a range of C substrates. In this study, SIR for the MicroResp™ profiles was low when compared to a study by Campbell et al. (2003). However, similar to that study, the highest level of SIR in all forest types of this study was observed with d-fructose, while the lowest detectable SIR was observed with l-lysine. It is worth noting that although PCA is commonly used to analyse CLPP data, in this and other studies (e.g. Priha et al. 1999, 2001), treatments were not separated using PCA. Compared to PCA, which is based on measurements of variance in the data, NMS and cluster analysis are based on distance measures (Anderson 1984), and both were more successful in separating forest types than PCA.
As discussed previously, land use and management may not only affect the size and activity of the soil microbial community, but also influence its composition or functional diversity. This may result in a change in physiological capacity of the soil microbial community and may in turn affect organic matter decomposition and soil N cycling (Garland 1997; Waldrop et al. 2000; Larkin 2003; Carney and Matson 2006).
Results of the MicroResp™ analysis revealed that although the two forest types had similar basal respiration, the NF soil tended to have a higher SIR than the 1R soil (see Fig. 3a). Further analysis of the patterns of substrate utilisation revealed separation of the NF and 1R soil into distinguished groups (NMS and cluster analysis). These results suggest that the microbial community in the NF soil has greater diversity and activity, and a different composition to the 1R soil. This result is supported by previous work at this study site in which greater microbial and fungal diversity, based on the culture-independent, DNA-fingerprinting method, was found in the NF soil compared to the 1R soil (He 2004; He et al. 2005). Litter, root and soil data from this study and previous studies at this site tend to suggest that the change in tree species/stand composition has reduced the quality and quantity of organic matter input, which may have contributed to the shift in soil microbial community composition and diversity (Burton et al. 2007a, b; Chen et al. 2004; Xu et al. 2008).
Harvesting of the 1R forest and conversion to 2R plantation and the ensuing residue management strategies may affect the soil microbial community (Breland and Hansen 1996; Li et al. 2004; Chen and Xu 2005; He et al. 2005). Comparison of the MicroResp™ profiles of the 1R and 2R soils indicates that there were some differences in the soil microbial community composition between the two forest types. However, the differences between the 1R and 2R soils were not as clearly defined as the differences between the NF and 1R soils. Previous studies at this site found changes in the quality and quantity of organic matter (e.g. root, litter and soil C/N ratios, and some pools of SON and SOC) associated with the conversion of 1R to 2R hoop pine plantation (Burton et al. 2007a, b). It is likely that these changes have contributed to the difference in microbial community composition or functional diversity between the 1R and 2R forests. Although the 2R-T and 2R-W appeared to be reasonably well separated in the cluster and NMS analysis of the MicroResp™ data, overall there was no significant difference in soil microbial community composition or diversity associated with residue management. This may be partly due to the large variability in the replicate MicroResp™ profiles within the 2R-W site.
5 Conclusions and recommendations
The results from this study indicate that the land-use change from the NF to the 1R hoop pine plantation is associated with reductions in soil microbial biomass and activity, and changes in the composition of the soil microbial community in the 0- to 10-cm soil layer. These changes are likely a consequence of reductions in the quantity and quality of organic matter inputs associated with the land-use change and may have significant implications for the long-term productivity of the soil resource.
While there is no evidence of changes in population size and respiration associated with the conversion of 1R to 2R hoop pine plantation, the Microresp™ data indicate that there are differences in the microbial community composition between the 1R and 2R soils. Residue management did not appear to have a significant influence on any of the microbial parameters, suggesting that the soil microbial community is resistant to this management technique. However, it is also possible that this result may be the consequence of the fact that samples are only representative of one sampling time, which occurred approximately 5 years after harvesting of the 1R plantation and establishment of the 2R plantation.
In addition, studies are required to confirm a difference in microbial community composition associated with residue management. As this is a snapshot study (i.e. results are based on a single round of sample collection), the long-term experiments with regular sampling throughout all seasons are required to verify the results of this study and to improve our understanding of the impact of land use and residue management on soil microbial community dynamics in subtropical Australia.
References
Anderson T (1984) An introduction to multivariate statistical analysis. Wiley, New York
Blumfield TJ, Xu ZH (2003) Impact of harvest residues on soil mineral nitrogen dynamics following clearfall harvesting of a hoop pine plantation in subtropical Australia. For Ecol Manage 179:55–67
Breland TA, Hansen S (1996) Nitrogen mineralization and microbial biomass as affected by soil compaction. Soil Biol Biochem 28:655–663
Brookes PC, Kragt JF, Powlson DS, Jenkinson DS (1985) Chloroform fumigation and the release of soil nitrogen: the effects of fumigations time and temperature. Soil Biol Biochem 17:831–835
Bucher A, Lanyon L (2005) Evaluating soil management with microbial community-level physiological profiles. Appl Soil Ecol 29:59–71
Burton JM, Chen CR, Xu ZH, Ghadiri H (2007a) Gross nitrogen transformations in adjacent native and plantation forests of subtropical Australia. Soil Biol Biochem 39:426–433
Burton JM, Chen CR, Xu ZH, Ghadiri H (2007b) Soluble organic nitrogen pools in adjacent native and plantation forests of subtropical Australia. Soil Biol Biochem 39:2723–2734
Campbell CD, Grayston SJ, Hirst DJ (1997) Use of rhizosphere carbon sources in sole carbon source tests to discriminate soil microbial communities. J Microbiol Meth 30:33–41
Campbell CD, Chapman SJ, Cameron CM, Davidson MS, Potts JM (2003) A rapid microtiter plate method to measure carbon dioxide evolved from carbon substrate amendments so as to determine the physiological profiles of soil microbial communities by using whole soil. Appl Environ Microbiol 69:3593–3599
Carney K, Matson P (2006) The influence of tropical plant diversity and composition on soil microbial communities. Microb Ecol 52:226–238
Chen CR, Xu ZH (2005) Soil carbon and nitrogen pools and microbial properties in a 6-year-old slash pine plantation of subtropical Australia: impacts of harvest residue management. For Ecol Manage 206:237–247
Chen CR, Xu ZH (2008) Analysis and behaviour of Soluble organic nitrogen in forest soils. J Soils Sediments 8:363–378
Chen C, Condron LM, Davis M, Sherlock RR (2000) Effects of afforestation on phosphorus dynamics and biological properties in a New Zealand grassland soil. Plant Soil 220:151–163
Chen CR, Xu ZH, Hughes JM (2002) Effects of nitrogen fertilization on soil nitrogen pools and microbial properties in a hoop pine (Araucaria cunninghamii) plantation in southeast Queensland, Australia. Biol Fertil Soils 36:276–283
Chen CR, Xu ZH, Mathers NJ (2004) Soil carbon pools in adjacent natural and plantation forests of subtropical Australia. Soil Sci Soc Am J 68:282–291
Cookson WR, Osman M, Marschner P, Abaye DA, Clark I, Murphy DV, Stockdale EA, Watson CA (2007) Controls on soil nitrogen cycling and microbial community composition across land use and incubation temperature. Soil Biol Biochem 39:744–756
Curtin D, Wen G (1999) Organic matter fractions contributing to soil nitrogen mineralization potential. Soil Sci Soc Am J 63:410–415
Curtin D, Wright C, Beare M, McCallum F (2006) Hot water-extractable nitrogen as an indicator of soil nitrogen availability. Soil Sci Soc Am J 70:1512–1521
Doran JW, Zeiss MR (2000) Soil health and sustainability: managing the biotic component of soil quality. Appl Soil Ecol 15:3–11
Elliott L, Lynch J, Papendick R (1996) The microbial component of soil quality. In: Stotzky G, Bollag J (eds) Soil biochemistry. Marcel Dekker, Inc, New York, pp 1–20
Garland J (1997) Analysis and interpretation of community-level physiological profiles in microbial ecology. FEMS Microbial Ecol 24:289–300
Gomez E, Bisaro V, Conti M (2000) Potential C-source utilization patterns of bacterial communities as influenced by clearing and land use in a vertic soil of Argentina. Appl Soil Ecol 15:273–281
Grayston SJ, Prescott CE (2005) Microbial communities in forest floors under four tree species in coastal British Columbia. Soil Biol Biochem 37:1157–1167
Hannam KD, Quideau SA, Kishchuk BE (2006) Forest floor microbial communities in relation to stand composition and timber harvesting in northern Alberta. Soil Biol Biochem 38:2565–2575
He JZ (2004) Molecular biological studies of soil microbial communities under different management practices in forest ecosystems of Queensland. PhD Thesis, Griffith University, Brisbane, 161pp
He JZ, Xu ZH, Hughes J (2005) Analyses of soil fungal communities in adjacent natural forest and hoop pine plantation ecosystems of subtropical Australia using molecular approaches based on 18 S rRNA genes. FEMS Microbiol Lett 247:91–100
He JZ, Xu ZH, Hughes H (2006) Molecular bacterial diversity of a forest soil under different residue management regimes in subtropical Australia. FEMS Microbiol Ecol 55:38–47
He Y, Xu ZH, Chen CR, Burton J, Ma Q, Ge Y, Xu JM (2008) Light fraction and macroaggregate associated organic matters as early indicators for management-induced changes of soil in adjacent native and plantation forests of subtropical Australia. Geoderma 147:116–125
He JZ, Ge Y, Xu ZH, Chen CR (2009a) Linking soil bacterial diversity to ecosystem multifunctionality using backward-elimination boosted trees analysis. J Soils Sediments 9:547–554
He Y, Chen CR, Xu ZH, Williams D, Xu J (2009b) Assessing management impacts on soil organic matter quality in subtropical Australian forests using physical and chemical fractionation as well as 13C NMR spectroscopy. Soil Biol Biochem 41:640–650
Huang ZQ, Xu ZH, Chen CR (2008) Interactions among soil organic matter fractions, microbial communities and nitrogen transformations in two hardwood plantations of subtropical Australia: effect of mulch. Appl Soil Ecol 40:229–239
Jenkinson D (1988) Determination of microbial biomass carbon and nitrogen in soil. In: Wilson J (ed) Advances in nitrogen cycling in agricultural ecosystems, cab International, Wallingford, pp 368–385
Larkin RP (2003) Characterization of soil microbial communities under different potato cropping systems by microbial population dynamics, substrate utilization, and fatty acid profiles. Soil Biol Biochem 35:1451–1466
Leckie SE, Prescott CE, Grayston SJ, Neufeld JD, Mohn WW (2004) Characterisation of humus microbial communities in adjacent forest types that differ in nitrogen availability. Microb Ecol 48:29–40
Li Q, Lee Allen H, Wollum AG (2004) Microbial biomass and bacterial functional diversity in forest soils: effects of organic matter removal, compaction, and vegetation control. Soil Biol Biochem 36:571–579
Luizao RCC, Bonde TA, Rosswall T (1992) Seasonal variation of soil microbial biomass—the effects of clearfelling a tropical rainforest and establishment of pasture in the central Amazon. Soil Biol Biochem 24:805–813
Mathers NJ, Mendham DS, O’Connell AM, Grove TS, Xu Z, Saffigna PG (2003a) How does residue management impact soil organic matter composition and quality under Eucalyptus globulus plantations in southwestern Australia? For Ecol Manage 179:253–267
Mathers NJ, Xu ZH, Blumfield TJ, Berners-Price SJ, Saffigna PG (2003b) Composition and quality of harvest residues and soil organic matter under windrow residue management in young hoop pine plantations as revealed by solid-state 13C NMR spectroscopy. For Ecol Manage 175:467–488
Mielke P, Berry K (2001) Permutation methods: a distance function approach. Springer Series in Statistics. Springer
Pan KW, Xu ZH, Blumfield TJ, Tutua S, Lu MX (2008) In situ mineral 15N dynamics and fate of added 15NH4 + in hoop pine plantation of subtropical Australia. J Soil Sediments 8:398–405
Pan KW, Xu ZH, Blumfield TJ, Tutua S, Lu MX (2009) Application of (15NH4)2SO4 to study N dynamics in hoop pine plantation and adjacent native forest of subtropical Australia: the effects of injection depth and litter addition. J Soils Sediments 8:398–405
Paul E, Clark F (1996) Soil microbiology and biochemistry. Academic Press, San Diego, 340 pp
Pietikainen J, Fritze H (1995) Clear-cutting and prescribed burning in coniferous forest: comparison of effects on soil fungal and total microbial biomass, respiration activity and nitrification. Soil Biol Biochem 27:101–109
Priha O, Smolander A (1997) Microbial biomass and activity in soil and litter under Pinus sylvestris, Picea abies and Betula pendula at originally similar field afforestation sites. Biol Fertil Soils 24:45–51
Priha O, Grayston SJ, Pennanen T, Smolander A (1999) Microbial activities related to C and N cycling and microbial community structure in the rhizospheres of Pinus sylvestris, Picea abies and Betula pendula seedlings in an organic and mineral soil. FEMS Microbial Ecol 30:187–199
Priha O, Grayston SJ, Hiukka R, Pennanen T, Smolander A (2001) Microbial community structure and characteristics of the organic matter in soils under Pinus sylvestris, Picea abies and Betula pendula at tow forest sites. Biol Fertil Soils 33:17–24
R Development Core Team (2008) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, URL http://www.R-project.org.
Rayment G, Higginson F (1992) Australian laboratory handbook of soil and water chemical methods. Inkata Press, Melbourne
Smaill SJ, Clinton PW, Greenfield LG (2010) Legacies of organic matter removal: decreased microbial biomass nitrogen and net N mineralization in New Zealand Pinus radiata plantations. Biol Fertil Soils 46:309–316
Smith J, Paul E (1990) The significance of soil microbial biomass estimations. In: Stotzky G, Bollag J (eds) Soil biochemistry. Marcel Dekker, New York, pp 357–396
Smolander A, Kitunen V (2002) Soil microbial activities and characteristics of dissolved organic C and N in relation to tree species. Soil Biol Biochem 34:651–660
Soil Survey Staff (1999) Soil taxonomy a basic system of soil classification for making and interpreting soil surveys. Agriculture handbook, 436. USDA Soil Conservation Service, Washington
Sparling GP (1997) Soil microbial biomass, activity and nutrient cycling as indicators of soil health. In: Pankhurst CE, Doube BM, Gupta VVSR (eds) Biological indicators of soil health. CAB International, pp 97–119
Vance ED, Brookes PC, Jenkinson DS (1987) An extraction method for measuring soil microbial biomass C. Soil Biol Biochem 19:703–707
Waldrop MP, Balser TC, Firestone MK (2000) Linking microbial community composition to function in a tropical soil. Soil Biol Biochem 32:1837–1846
Xu ZH, Ward S, Chen CR, Blumfield T, Prasolova NV, Liu JX (2008) Soil carbon and nutrient pools, microbial properties and gross nitrogen transformations in adjacent natural forest and hoop pine plantations of subtropical Australia. J Soil Sediments 8:99–105
Xu ZH, Chen CR, He J, Liu J (2009) Trends and challenges in soil research 2009: linking global climate change to local long-term forest productivity. J Soils Sediments 9:83–88
Yao H, He Z, Wilson M, Campbell CD (2000) Microbial biomass and community structure in a sequence of soils with increasing fertility and changing land use. Microb Ecol 40:223–237
Zhang L, Xu ZH (2008) Assessing bacterial diversity in soil: a brief review. J Soils Sediments 8:379–388
Zhang L, Xu ZH, Patel BKC (2007a) Bacillus decisifrondis sp nov., isolated from soil underlying decaying leaf foliage. Int J Syst Evol Microbiol 57:974–978
Zhang L, Xu ZH, Patel BKC (2007b) Frondicola australicus gen. nov., sp nov., isolated from decaying leaf litter from a pine forest. Int J Syst Evol Microbiol 57:1177–1182
Zhang L, Xu ZH, Patel BKC (2009) Culture-dependent and culture-independent microbial investigation of pine litters and soil in subtropical Australia. J Soils Sediments 9:148–160
Zhong Z, Makeschin F (2006) Differences of soil microbial biomass and nitrogen transformation under two forest types in central Germany. Plant Soil 283:287–297
Acknowledgements
We acknowledge the Forestry Plantations Queensland and in particular Mr. Richard Jackson for allowing us access to the experimental site. We would like to thank Dr. Rui Yin, Mr. Yu Huang, Mr. Stephen Faggotter and Ms. Elizabeth Bridon for their assistance in soil sampling and processing. We also thank Mr. James McBroom for statistical advice. Finally, we would like to acknowledge the financial support provided by the CRC for Sustainable Production Forestry, Australian Rivers Institute and the Australian Research Council.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Hailong Wang
Rights and permissions
About this article
Cite this article
Burton, J., Chen, C., Xu, Z. et al. Soil microbial biomass, activity and community composition in adjacent native and plantation forests of subtropical Australia. J Soils Sediments 10, 1267–1277 (2010). https://doi.org/10.1007/s11368-010-0238-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11368-010-0238-y